Embed Size (px)
Citation preview

Subnuclear Localization of FOS-LikeImmunoreactivity in the ParabrachialNucleus After Orofacial Nociceptive
Stimulation of the Awake Rat
OLA HERMANSON* AND ANDERS BLOMQVIST
Department of Cell Biology, Faculty of Health Sciences, University of Linkoping,S-581 85 Linkoping, Sweden
ABSTRACTWe used FOS-like immunohistochemistry to detect neuronal activity in the pontine
parabrachial nucleus after injection of formalin into the lower lip of the awake rat andcompared the labeling pattern with that seen after formalin injection into the hindpaw. Onehour after a formalin injection into the lip, many FOS-immunoreactive cells were seen in theparabrachial nucleus, preferentially on the side ipsilateral to the injection site. Detailedanatomical analysis revealed that FOS-immunoreactive neurons were localized predomi-nantly to three regions of the parabrachial nucleus: the external lateral, the external medial,and the Kolliker-Fuse subnuclei, with sparser labeling present in the dorsal and superiorlateral subnuclei and in the medial parabrachial nucleus. In contrast, a formalin injection intothe hindpaw resulted in dense FOS-labeling in the superior, dorsal, and central lateralsubnuclei, with sparse to moderate labeling in the Kolliker-Fuse nucleus, and sparse labelingin the external lateral and external medial subnuclei, as described previously (Hermansonand Blomqvist, J. Comp. Neurol., [1996] 368:45–56).
The distribution of FOS-labeled neurons after noxious orofacial stimulation correspondsto the termination pattern in the parabrachial nucleus of fibers that originate from neurons inthe marginal zone of the trigeminal dorsal horn and is different from that seen afternociceptive stimulation of other body parts. Considering the differences in efferent connec-tions of parabrachial subnuclei, the present findings imply that noxious information from theorofacial region to the parabrachial nucleus has other functional roles than noxiousinformation from the trunk and limbs. Such roles may include the integration of somatosen-sory and gustatory information, which has been suggested to be of importance for feedingbehavior. J. Comp. Neurol. 387:114–123, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: Kolliker-Fuse nucleus; somatosensory; pain; trigeminal dorsal horn; immediate
early gene
With the combined use of anatomical and electrophysi-ological techniques, Norgren and Leonard (1973) showedthat the pontine parabrachial nucleus (PB) receives tasteinformation from the nucleus of the solitary tract (NTS).Since that pioneering work, a large number of studies havedemonstrated that the PB is involved in special andgeneral visceral mechanisms (Aguero et al., 1993; Li andRowland, 1993; Chan and Sawchenko, 1994; Grigson et al.,1994; Lara et al., 1994; Swank and Bernstein, 1994).However, it has also been demonstrated that the PB is amajor target site for ascending projections from nocirespon-sive neurons in the spinal and trigeminal dorsal horn(Wiberg and Blomqvist, 1984; Cechetto et al., 1985; Hyldenet al., 1986; Wiberg et al., 1986; Blomqvist et al., 1989;
Hayashi and Tabata, 1989; Menetrey et al., 1989; Noguchiand Ruda, 1992; Light et al., 1993; Panneton et al., 1994;Slugg and Light, 1994; Wang et al., 1994; Bernard et al.,1995; Craig, 1995; Feil and Herbert, 1995). The spinal andtrigeminal fibers terminate largely in separate subnucleargroups of the PB. Hence, the spinal afferent fibers termi-
Grant sponsor: Swedish Medical Research Council; Grant number: 7879;Grant sponsor: Swedish Society for Medical Research.
*Correspondence to: Ola Hermanson, Department of Cell Biology, Fac-ulty of Health Sciences, University of Linkoping, S-581 85 Linkoping,Sweden. E-mail: [email protected]
Received 31 January 1997; Revised 6 May 1997; Accepted 21 May 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 387:114–123 (1997)
r 1997 WILEY-LISS, INC.

nate most densely in the dorsal lateral (PBdl) and centrallateral (PBcl) subnuclei (for nomenclature, see Fulwilerand Saper, 1984), whereas the trigeminal fibers terminateprimarily in the Kolliker-Fuse nucleus (K-F) and in theexternal lateral (PBel) and external medial (PBem) subnu-clei (Cechetto et al., 1985; Panneton et al., 1994; Slugg andLight, 1994; Feil and Herbert, 1995). In addition, thespinal input to the PB is predominantly contralateral, incontrast to the trigeminal input, which is mainly ipsilat-eral (Cechetto et al., 1985; Panneton et al., 1994; Slugg andLight, 1994; Feil and Herbert, 1995). Because the PBsubnuclei display distinct efferent connections, which dif-fer also in laterality (Fulwiler and Saper, 1984; Cechettoand Saper, 1987; Krukoff et al., 1993), the different termi-nation patterns of the spinal and trigeminal input implythat the information from different body regions may playdifferent functional roles.
Consistent with the presence of a nociceptive input tothe PB, noxious-evoked activity has been demonstrated inelectrophysiological studies of PB neurons (Hayashi andTabata, 1989; Bernard and Besson, 1990), but these stud-ies have not revealed differences in the response propertiesof PB neurons that would relate to the anatomical differ-ences in the organization of the afferent input from variousbody parts. However, because the electrophysiologicalstudies were performed on anesthetized animals andbecause PB neurons are involved in autonomic regulation,the cardiovascular and respiratory effects of anesthesiamight have influenced the results (Clement et al., 1996;Krukoff et al., 1992; Takayama et al., 1994). These limita-tions do not apply to the demonstration of synaptic activa-tion of neurons in unanesthetized animals in which theprotooncogene protein FOS is detected by immunohisto-chemistry (Hunt et al., 1987). By use of this technique, wedemonstrated, in a previous study, the subnuclear distribu-tion of PB neurons that were activated following noxiousstimulation of the neck, tail, and hindlimbs in unanesthe-tized rats (Hermanson and Blomqvist, 1996b). This distri-bution clearly differed from that seen in electrophysiologi-cal studies on anesthetized animals (Bernard and Besson,1990) and was largely consistent with the terminationpattern of the spinal afferent fibers. Thus, to furtheranalyze the relation between the ascending afferent inputand the distribution of noxious-evoked activity in the PB,we have, in the present study, used FOS-immunohisto-chemistry to compare the distribution of FOS-expressingneurons following noxious stimulation of the orofacialregion and the hindpaw, respectively.
MATERIALS AND METHODS
Male Sprague-Dawley rats (250-350 g; B&K Universal,Stockholm, Sweden) were used. All experimental proce-dures were approved by the Animal Care and Use Commit-tee at the University of Linkoping.
One rat at a time was placed in a separate cage in a dark,quiet room. After 3 hours of habituation, the rat received ashort ether anesthesia (60–90 seconds), during which30–50 µl of 1.85% paraformaldehyde in 0.9% NaCl (26–30°C) was injected into the lower lip on one side. The ratswere then allowed to survive 30 minutes (n52), 60–70minutes (n56), or 180 minutes (n52). Six other ratsreceived an injection of 50–100 µl of 1.85% paraformalde-hyde in 0.9% NaCl (26–30°C) into one hindpaw and werekilled 60–70 minutes after the injection. Previous analysishas shown that the number of FOS-labeled neurons in thePB following hindpaw stimulation was largest after thissurvival period (Hermanson and Blomqvist, 1996b).
At the end of the survival period, the animals were givenan overdose of sodium pentobarbital (120 mg/kg injectedinto the liver) and perfused transcardially within 3 min-utes with 150 ml 0.9% NaCl at room temperature, followedby 500 ml 4% paraformaldehyde in 0.1 M phosphate buffer(pH 7.4) at 4°C. The brain stem and spinal cord wereremoved, stored in a 20% sucrose-fixative solution for 20hours, and cut transversely at 40 µm on a freezingmicrotome. Every second pontine and every fourth medul-lary section was incubated overnight at room temperatureunder constant agitation in a solution of a rabbit anti-FOSantibody (Santa Cruz, Santa Cruz, CA, sc-04, 0.20 µg/ml;raised against a peptide of amino acids 3–16 of the FOSprotein; this antibody does not cross-react with any knownFOS-related antigen) and 0.5% bovine serum albumin(BSA) in 0.1 M phosphate buffer (pH 7.4). Bound primaryantibody was visualized with secondary antibody (swine-anti-rabbit, 1:100; Dakopatts, Copenhagen, Denmark; 1hour, room temperature) and peroxidase anti-peroxidase(PAP) complex (PAP-rabbit, 1:150; Dakopatts; 1 hour, roomtemperature), followed by 0.02% 3,3’-diaminobenzidinetetrahydrochloride, 0.01% hydrogen peroxide, and 2.5%nickel ammonium sulphate in 0.1 M sodium acetate buffer(pH 6.0) for 6–8 minutes. Adjacent sections were stainedwith thionin.
The sections were analysed in the light microscope at aprimary magnification of 34. Camera lucida drawings ofthe distribution of FOS-labeled neurons were made fromevery processed section (n 5 12–16) through the lateral PBin the animals that were killed 60-70 minutes after theformalin injection into the lower lip (n56; see Results). Asimilar analysis was made in animals killed 60–70 min-utes after a formalin injection into one hindpaw (n56). Theilluminations of the microscopic field and the drawingpaper were adjusted so that only profiles that displayed alabeling intensity above background level were seen, andonly such profiles were counted. Background level wasdetermined by examining areas not expected to expressFOS after nociceptive stimulation, such as the centralnucleus of the inferior colliculus. The boundaries of the PBsubnuclei were identified in adjacent thionin sections. Thenomenclature followed Fulwiler and Saper (1984) andHerbert et al. (1990). The dorsal lateral subnucleus wasconsidered to extend approximately 50–100 µm morerostrally than originally described by Fulwiler and Saper(1984). The extreme lateral subnucleus was not delineated
Abbreviations
me5 mesencephalic trigeminal tractK-F Kolliker-Fuse nucleusLC lateral crescent areaPB parabrachial nucleusPBcl, CL central lateral parabrachial subnucleusPBdl, DL dorsal lateral parabrachial subnucleusPBel, EL external lateral parabrachial subnucleusPBem, EM external medial parabrachial subnucleusPBil, IL internal lateral parabrachial subnucleusPBm, M medial parabrachial nucleusPBsl, SL superior lateral parabrachial subnucleusPBvl, VL ventral lateral parabrachial subnucleusscp superior cerebellar peduncle
FOS IN PB AFTER OROFACIAL NOXIOUS STIMULATION 115

due to its close proximity to the lateral lemniscus (seeHermanson and Blomqvist, 1996b).
Control procedures have been described in detail previ-ously (Hermanson and Blomqvist, 1996b). Thus, ratstreated with ether alone or subjected to sound stimulationdisplayed little FOS in the PB. After food deprivation, amoderate number of FOS-labeled neurons were seen pri-marily in the PBel (see Discussion). Additional controls inthe present series of experiments included injection ofsaline (0.9%, 30–50 µl, 26–30°C) or a local anesthetic,bupivacain (Marcaint, 2.5%, 30–50 µl, 26–30°C; Astra,Sodertalje, Sweden), into the lip (n 5 4 and 2, respec-tively), followed by 60 minutes survival.
RESULTS
Orofacial stimulation
Formalin injection into the lower lip evoked FOS-likeimmunoreactivity in the nuclei of neurons as previouslydescribed (Sambucetti and Curran, 1986). A large numberof neurons were seen in a cluster in the dorsal part of thetrigeminal dorsal horn (Fig. 1), being consistent with thesomatotopic organization of the primary afferent input(Marfurt, 1981; Jacquin et al., 1983; Pfaller and Arvidsson,1988) and with previous demonstrations of FOS in thetrigeminal dorsal horn after nociceptive orofacial stimula-tion (Strassman and Vos, 1993; Coimbra and Coimbra,1994; Kato et al., 1994). Many labeled neurons were alsodetected in the nucleus of the solitary tract (NTS), prefer-entially in its rostrolateral part. In addition, a smallnumber of labeled neurons were seen in lamina III–V ofthe medullary dorsal horn and in the ventrolateral me-dulla.
Many FOS-immunoreactive neurons were present in thePB (Figs. 2–5). In all subnuclei, the largest number ofneurons were seen in rats that had survived 60–70 min-utes after the formalin injection. Therefore, the distribu-tion and number of FOS-labeled neurons in the PB wereanalyzed in detail in these rats.
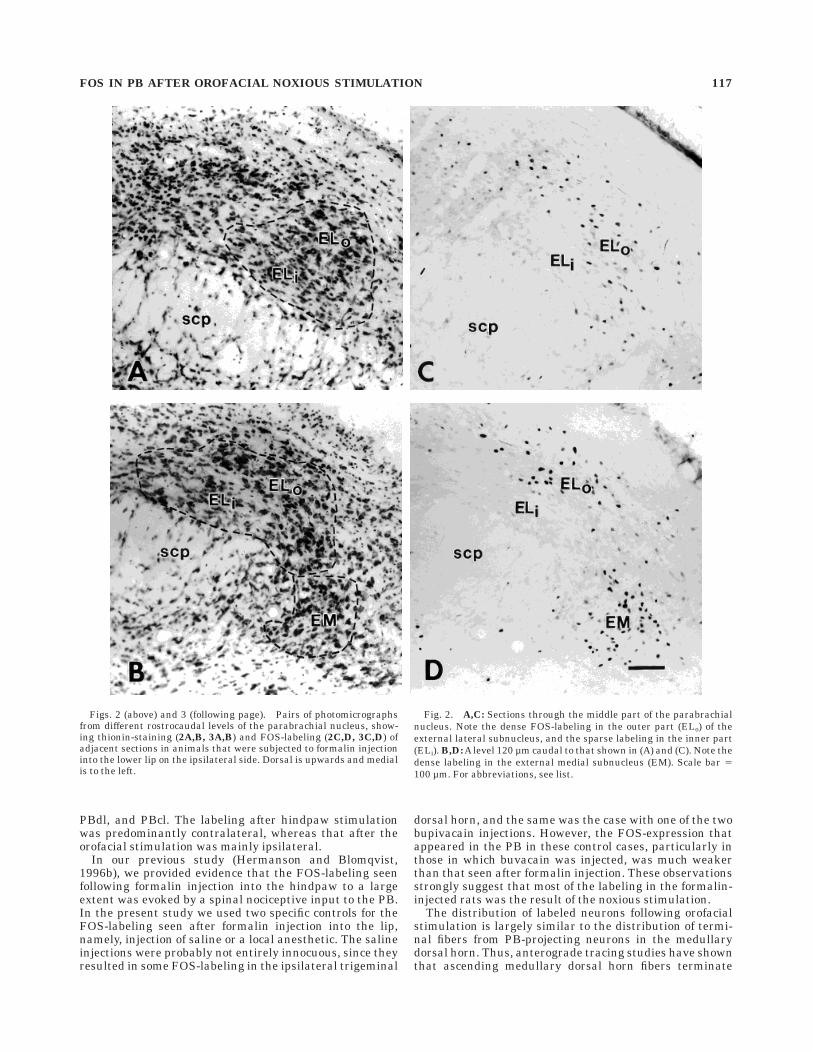
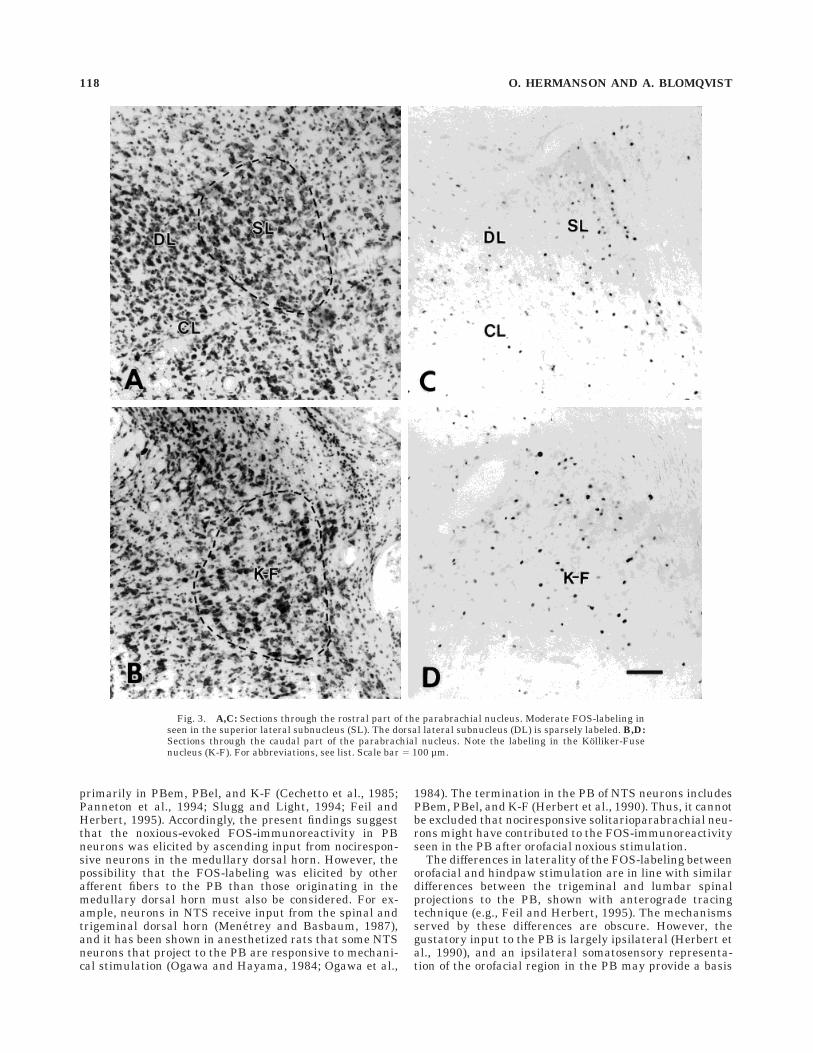
The labeling in the PB was predominantly localized tothe side ipsilateral to the injected lip, but many FOS-labeled neurons were found also on the contralateral side,and the subnuclear distribution of labeled neurons was thesame on both sides. Dense labeling was seen in threesubnuclei: PBem, PBel and K-F (Figs. 2–5). In PBem,FOS-labeled neurons were present throughout the entiresubnucleus (Fig. 2). In PBel, most of the labeling was seenin the outer part of this subnucleus (Fig. 2); this labelingextended also into adjacent parts of PBcl and the lateralcrescent area (PBlc; see Figs. 2,4). K-F was labeled through-out its extent (Fig. 3), but most labeled neurons werelocalized to its rostral levels, with the densest labelingseen 100–200 µm caudal to the level of separation betweenthe inferior colliculus and pons (Figs. 3,4).
In addition to the labeling in PBem, PBel, and K-F,moderate numbers of labeled neurons were seen in thesuperior lateral subnucleus (PBsl; Figs. 3–5), PBdl (Figs.4, 5), and in the medial PB (PBm). A small number ofFOS-labeled neurons were found in the ventral (PBvl) andinternal (PBil) lateral subnuclei, and in the medial parts ofPBcl (Fig. 4). At the very rostral levels of the PB, diffuselydistributed FOS-labeled neurons were seen; this labeling
extended in the rostrodorsomedial direction towards theventrolateral periaqueductal gray matter.
Injection of saline into the lip yielded a faint labeling ofneurons within the terminal zone of the mandibular nervein the dorsal part of the trigeminal dorsal horn (Marfurt,1981; Jacquin et al., 1983; Pfaller and Arvidsson, 1988).The labeling pattern in the PB was similar to that seenafter formalin injection, but the number of FOS-labeledneurons was much sparser, and the immunoreactivity inthe labeled neurons was generally much weaker. Injectionof bupivacain into the lip yielded in one of the two cases afaint labeling of neurons in the trigeminal dorsal horn,whereas in the other case no trigeminal dorsal hornlabeling was seen. Both these cases resulted in sparselabeling in the PB, preferentially located to the outer partof PBel and K-F. In these cases, as well as in the salinecontrols, a group of rather faintly labeled neurons was alsoseen in the caudal part of PBcl.
Hindpaw stimulation
Formalin injection into one hindpaw resulted in thesame labeling pattern as described previously (Herman-son and Blomqvist, 1996b). Thus, large numbers of labeledneurons were seen in PBsl, PBdl, and PBcl (Fig. 4), withthe majority of labeled neurons localized contralateral tothe formalin-injected side. Small to moderate numbers oflabeled neurons were also seen in K-F on both sides,whereas only few labeled neurons were found in PBel andPBem (Hermanson and Blomqvist, 1996b). Single FOS-immunoreactive neurons were only occasionally detectedin PBil and PBvl (Fig. 4).
DISCUSSION
This study shows that nociceptive stimulation of theorofacial region of the awake rat evokes a labeling patternthat is different from that seen after nociceptive stimula-tion of the hindpaw (present study; Hermanson and Blom-qvist, 1996b) as well as of the tail and neck (Hermansonand Blomqvist, 1996b). Thus, formalin injection into thelower lip resulted in a large number of labeled neurons inPBel, K-F, and PBem. In contrast, formalin injection intothe hindpaw resulted in labeling preferentially in PBsl,
Fig. 1. Photomicrograph showing FOS-labeled neurons in thedorsomedial part of the medullary dorsal horn following formalininjection into the lower lip. Dorsal is upwards and medial to the left.Scale bar 5 200 µm.
116 O. HERMANSON AND A. BLOMQVIST
PBdl, and PBcl. The labeling after hindpaw stimulationwas predominantly contralateral, whereas that after theorofacial stimulation was mainly ipsilateral.
In our previous study (Hermanson and Blomqvist,1996b), we provided evidence that the FOS-labeling seenfollowing formalin injection into the hindpaw to a largeextent was evoked by a spinal nociceptive input to the PB.In the present study we used two specific controls for theFOS-labeling seen after formalin injection into the lip,namely, injection of saline or a local anesthetic. The salineinjections were probably not entirely innocuous, since theyresulted in some FOS-labeling in the ipsilateral trigeminal
dorsal horn, and the same was the case with one of the twobupivacain injections. However, the FOS-expression thatappeared in the PB in these control cases, particularly inthose in which buvacain was injected, was much weakerthan that seen after formalin injection. These observationsstrongly suggest that most of the labeling in the formalin-injected rats was the result of the noxious stimulation.
The distribution of labeled neurons following orofacialstimulation is largely similar to the distribution of termi-nal fibers from PB-projecting neurons in the medullarydorsal horn. Thus, anterograde tracing studies have shownthat ascending medullary dorsal horn fibers terminate
Figs. 2 (above) and 3 (following page). Pairs of photomicrographsfrom different rostrocaudal levels of the parabrachial nucleus, show-ing thionin-staining (2A,B, 3A,B) and FOS-labeling (2C,D, 3C,D) ofadjacent sections in animals that were subjected to formalin injectioninto the lower lip on the ipsilateral side. Dorsal is upwards and medialis to the left.
Fig. 2. A,C: Sections through the middle part of the parabrachialnucleus. Note the dense FOS-labeling in the outer part (ELo) of theexternal lateral subnucleus, and the sparse labeling in the inner part(ELi). B,D: A level 120 µm caudal to that shown in (A) and (C). Note thedense labeling in the external medial subnucleus (EM). Scale bar 5100 µm. For abbreviations, see list.
FOS IN PB AFTER OROFACIAL NOXIOUS STIMULATION 117
primarily in PBem, PBel, and K-F (Cechetto et al., 1985;Panneton et al., 1994; Slugg and Light, 1994; Feil andHerbert, 1995). Accordingly, the present findings suggestthat the noxious-evoked FOS-immunoreactivity in PBneurons was elicited by ascending input from nocirespon-sive neurons in the medullary dorsal horn. However, thepossibility that the FOS-labeling was elicited by otherafferent fibers to the PB than those originating in themedullary dorsal horn must also be considered. For ex-ample, neurons in NTS receive input from the spinal andtrigeminal dorsal horn (Menetrey and Basbaum, 1987),and it has been shown in anesthetized rats that some NTSneurons that project to the PB are responsive to mechani-cal stimulation (Ogawa and Hayama, 1984; Ogawa et al.,
1984). The termination in the PB of NTS neurons includesPBem, PBel, and K-F (Herbert et al., 1990). Thus, it cannotbe excluded that nociresponsive solitarioparabrachial neu-rons might have contributed to the FOS-immunoreactivityseen in the PB after orofacial noxious stimulation.
The differences in laterality of the FOS-labeling betweenorofacial and hindpaw stimulation are in line with similardifferences between the trigeminal and lumbar spinalprojections to the PB, shown with anterograde tracingtechnique (e.g., Feil and Herbert, 1995). The mechanismsserved by these differences are obscure. However, thegustatory input to the PB is largely ipsilateral (Herbert etal., 1990), and an ipsilateral somatosensory representa-tion of the orofacial region in the PB may provide a basis
Fig. 3. A,C: Sections through the rostral part of the parabrachial nucleus. Moderate FOS-labeling inseen in the superior lateral subnucleus (SL). The dorsal lateral subnucleus (DL) is sparsely labeled. B,D:Sections through the caudal part of the parabrachial nucleus. Note the labeling in the Kolliker-Fusenucleus (K-F). For abbreviations, see list. Scale bar 5 100 µm.
118 O. HERMANSON AND A. BLOMQVIST
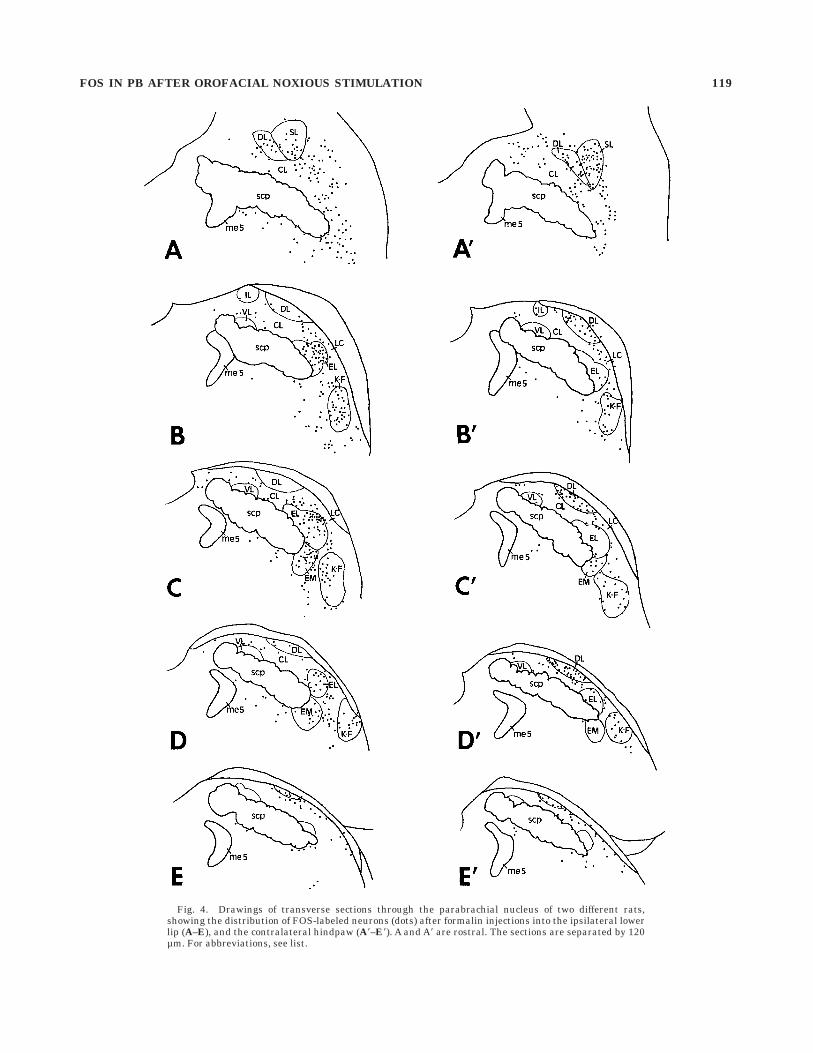
Fig. 4. Drawings of transverse sections through the parabrachial nucleus of two different rats,showing the distribution of FOS-labeled neurons (dots) after formalin injections into the ipsilateral lowerlip (A–E), and the contralateral hindpaw (A8–E8). A and A8 are rostral. The sections are separated by 120µm. For abbreviations, see list.
FOS IN PB AFTER OROFACIAL NOXIOUS STIMULATION 119
for the integration of these modalities in this brain stemregion. Interestingly, the thalamocortical representationof orofacial sensitivity has also a prominent ipsilateralcomponent in some species, although not in rats, matchingthe predominant ipsilateral representation of gustatoryinformation (for references, see Jones, 1985).
The present study is the first demonstration of noxious-evoked activity in PBel neurons of awake animals. TheFOS-immunoreactive neurons were preferentially foundin the outer part of PBel. This finding is consistent withprevious observations showing that fibers from neurons inthe trigeminal dorsal horn terminate in the outer part ofPBel (Cechetto et al., 1985) and support electrophysiologi-cal observations that PBel neurons are activated by nox-ious stimuli (Hayashi and Tabata, 1989; Bernard andBesson, 1990). It should be noted that in the study by Feiland Herbert (1995) tracer injection into a region of thespinal trigeminal nucleus, located slightly lateral to theFOS-labeled region in the present study, resulted in denseanterograde labeling preferentially in the ventral portionof PBel and only sparse labeling in the other parts of PBel.The discrepancy between the FOS labeling pattern inPBel, as shown by the present study, and the anterograde
labeling pattern found by Feil and Herbert (1995) mayreflect differences in the projection to PBel from differentparts of the spinal trigeminal nucleus.
Detailed anatomical studies have shown that PBelneurons project most densely to the lateral part of thecentral nucleus of the amygdala (CNA) (Bernard et al.,1991; Bernard et al., 1993; Krukoff et al., 1993), andnociceptive responses have been demonstrated in CNA(Bernard et al., 1990). Since trigeminoparabrachial fiberssynaptically contact dendrites of parabrachioamygdalianneurons (Ma and Peschanski, 1988), nociceptive orofacialstimulation may influence CNA neurons via PBel.
However, in several studies it has been suggested thatPBel, in addition to transmitting information from thetrigeminal region, may be a relay for noxious informationfrom the spinal cord to the amygdala (Ma and Peschanski,1988; Bernard et al., 1989; Bernard and Besson, 1990;Bernard et al., 1990; Bernard et al., 1994). The anatomicalevidence for this idea are meager; in the study by Ma andPeschanski (1988), synaptic contacts between trigemino-parabrachial fibers and PB neurons that projected to theamygdala were demonstrated, but no evidence for theexistence for a similar pathway from the spinal cord to theamygdala has yet been unequivocally demonstrated (seealso Ma et al., 1989). In consistence with the very sparsespinal input to PBel (Feil and Herbert, 1995), we foundthat nociceptive stimulation of the neck, tail, or hindlimbsfailed to evoke significant FOS-like immunoreactivity inPBel neurons (present study; Hermanson and Blomqvist,1996b; see also Lanteri-Minet et al., 1994; Bellavance andBeitz, 1996).
It has been suggested that the discrepancy between thelack of substantial spinal afferent projection to PBel andthe electrophysiological reports on nociceptive activationof PBel neurons from various body sites could be explainedby the presence of nociceptive spinal afferent input to PBeldendrites located within the termination field of the spinalfibers (Feil and Herbert, 1995; Herbert and Bellintani-Guardia, 1995; Saper, 1995). The possibility that thenociceptive input to PBel is not transmitted directly byspinoparabrachial fibers, but through a relay in the brain-stem must also be considered; thus, both NTS and theventrolateral medulla project densely to PBel (Herbert etal., 1990), and both regions receive a strong spinal input(Blomqvist and Berkley, 1992; Craig, 1995). As discussedpreviously (Hermanson and Blomqvist, 1996b), it is un-likely that these inputs, if present, synaptically activatePBel neurons, since PBel only displays sparse FOS-expression following nociceptive stimulation of other re-gions than the face. However, the possibility that thetrigeminal and spinal inputs activates different transcrip-tion mechanisms must be considered, particularly sincethe trigeminal target regions in the PB express otherneuropeptides than do the spinal target regions (Blom-qvist et al., 1994; Hermanson et al., 1995; Hermanson andBlomqvist, 1996a). So far, work along this line has notprovided evidence for a spinal activation of PBel. Thus, ina recent study detecting immunoreactivity for noxious-evoked phosphorylated cAMP-responsive element bindingprotein (CREB) in the PB, dense labeling was seen inmany nociresponsive PB subnuclei, but not in PBel (Her-manson and Blomqvist, 1996a). Other explanations for theelectrophysiological data involves the possibility that theanesthesia used in the electrophysiological studies re-duced the threshold for excitation of PBel neurons, so that
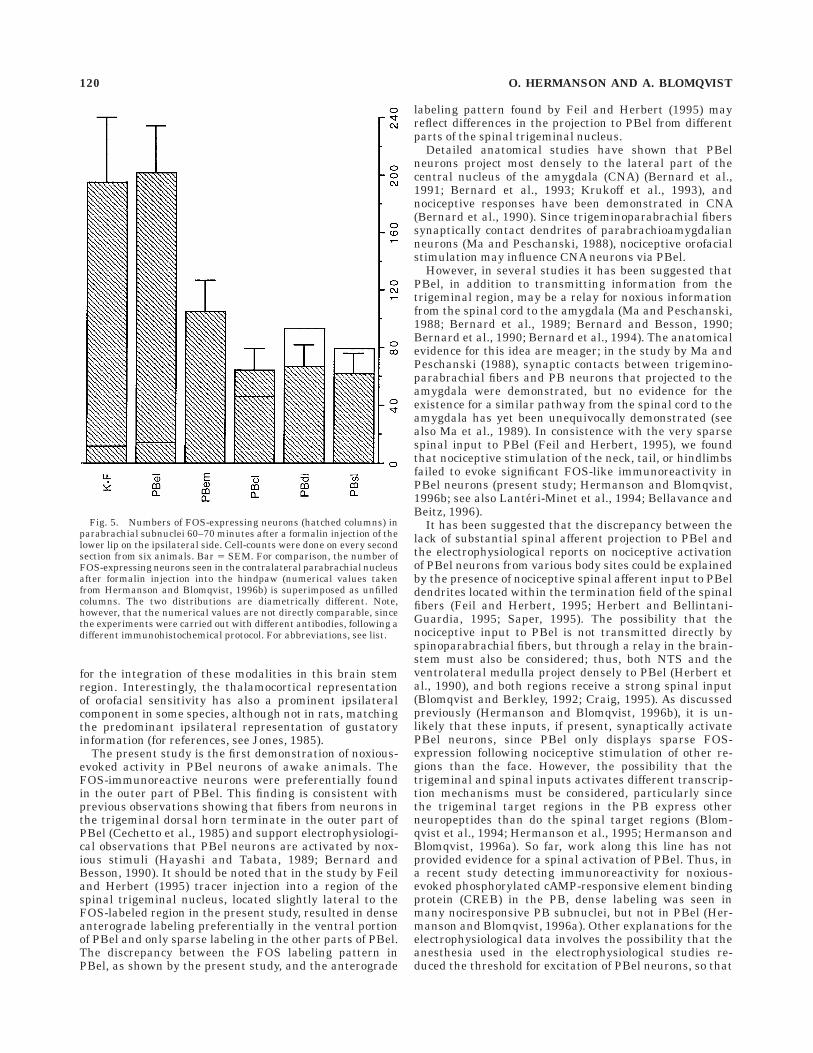
Fig. 5. Numbers of FOS-expressing neurons (hatched columns) inparabrachial subnuclei 60–70 minutes after a formalin injection of thelower lip on the ipsilateral side. Cell-counts were done on every secondsection from six animals. Bar 5 SEM. For comparison, the number ofFOS-expressing neurons seen in the contralateral parabrachial nucleusafter formalin injection into the hindpaw (numerical values takenfrom Hermanson and Blomqvist, 1996b) is superimposed as unfilledcolumns. The two distributions are diametrically different. Note,however, that the numerical values are not directly comparable, sincethe experiments were carried out with different antibodies, following adifferent immunohistochemical protocol. For abbreviations, see list.
120 O. HERMANSON AND A. BLOMQVIST

a putative spinal input to peripheral dendrites of PBelneurons, which in the awake animal is subthreshold, inthe anesthetized animal synaptically activated the neurons.
Neurons in PBel may play a role in gustatory mecha-nisms; for example, PBel and its efferent target the CNAhave been implicated in mechanisms of gustatory memory,such as taste aversion (Yamamoto et al., 1994; Yasoshimaet al., 1995), and lesions of the lateral PB abolishes such abehavior (Reilly et al., 1993; Grigson et al., 1994). It hasalso been suggested that the projection from the rostraldorsal part of the spinal trigeminal nucleus (i.e., thetermination zone of lingual nerve) to the PB represents amechanism for convergence of tongue somatic sensoryafferents with lingual gustatory information (Cechetto etal., 1985). Interestingly, neurons in the region of PBel havebeen shown to respond to both taste and oral mechanicalstimulation (Ogawa et al., 1982), and may accordinglyprovide the amygdala with such information. Indeed, ithas long been understood that orofacial sensory stimula-tion reaches the amygdala. As first recognized by Kluverand Bucy (1937, 1939), lesions of the amygdalian regionelicit a remarkable tendency to put all types of objects intothe mouth, a behavior believed to be due to impairedcapacity for crossmodal associations (Murray and Mish-kin, 1985).
In addition to oral information, general visceral informa-tion activates PBel neurons. For example, the anorecticagent dexfenfluramine has been shown to evoke FOS inneurons located in the outer part of PBel, resulting in aFOS-labeling pattern that is strikingly similar to that seenin the present study (Li and Rowland, 1993). Thus, activa-tion of PBel neurons may be the result of various stimulithat are associated with food avoidance or aversion.
In addition to the projection to the amygdala, PBelneurons have been shown to project to several limbicforebrain regions, such as the bed nucleus of the striaterminalis and the substantia innominata (Fulwiler andSaper, 1984; Moga et al., 1990; Alden et al., 1994) andpossibly also to brain stem structures, such as the periaq-ueductal gray matter (see Chamberlin and Saper, 1992).PBel contains a large number of neurons that expresspreproenkephalin mRNA (Blomqvist et al., 1994). This isof interest since FOS has been shown to regulate enkepha-lin transcription in some neuronal types (Sonnenberg etal., 1989), and since FOS is elicited in PBel by analgeticcompounds (Krukoff et al., 1992; Clement et al., 1996).Future studies may identify the detailed projections andneuropeptidergic contents of the orofacial-activated, FOS-labeled neurons in PBel.
FOS was consistently detected in the region just ventro-lateral to the superior cerebellar peduncle, including PBem.Neurons in this region are targeted by trigeminal fibers,and they have been shown to project to the thalamus,specifically to the parvicellular part of the ventropostero-medial nucleus (VPMpc) and the parvicellular part of theventroposterolateral nucleus (VPLpc) (Cechetto and Sa-per, 1987). VPMpc has long been implicated in gustatorymechanisms, whereas VPLpc has been shown to be in-volved in visceral mechanosensory mechanisms (Cechettoand Saper, 1987). It has been suggested (Xing et al., 1995)that VPLpc may be the rat equivalent to the nociceptive-and cold-specific ventromedial posterior nucleus (VMpo),which has been defined by immunohistochemical andelectrophysiological characterization in primates (Craig etal., 1994). The thalamic projection from the spinal and
trigeminal dorsal horn is sparse in rats compared to thatin cats, monkeys, and humans (Burstein et al., 1990; Iwataet al., 1992; Farnebo, Hamlin, and Blomqvist, unpublishedobservations). However, the present study suggests apossible alternative route for peripheral nociceptive stimu-lation to reach the posterior thalamus (cf., Floyd et al.,1996), and it is therefore of interest to study if thenoxious-induced FOS in the region of PBem is located inthalamic-projecting neurons.
Several studies have demonstrated that K-F is a majortarget site for the ascending trigeminal input (Cechetto etal., 1985; Panneton et al., 1994; Slugg and Light, 1994; Feiland Herbert, 1995), being in line with the present findingsthat K-F neurons are activated by noxious orofacial stimu-lation. Neurons in K-F have long been shown to beinvolved in respiration and antinociception (Bertrand andHugelin, 1971; Girardot et al., 1987; Young et al., 1992;Chamberlin and Saper, 1994). The analgetic effect of K-Fstimulation has been ascribed to the noradrenergic A7 cellgroup, which is partly localized to K-F (Westlund et al.,1983; Clark and Proudfit, 1991). However, we recentlydemonstrated that many K-F neurons are enkephalinergic(Blomqvist et al., 1994), that the enkephalinergic neurons,which make up the largest population, are separate fromthe noradrenergic neurons (Hermanson and Blomqvist,1995), and that the expression of preproenkephalin mRNAis increased in K-F following peripheral mechanical noci-ceptive stimulation (Blomqvist et al., 1994). These findingssupport previous electrophysiological observations show-ing non-noradrenergic, opioid-mediated spinal inhibitionfollowing electrical stimulation in the K-F region (Zhaoand Duggan, 1988; Zhuo et al., 1993). Enkephalinergicneurons in K-F project to the rostral ventrolateral medullaand the spinal cord (Blomqvist et al., 1994). Since FOSmay influence enkephalin transcription (Sonnenberg etal., 1989), FOS in K-F after orofacial noxious stimulationmay activate a descending opioidergic projection. How-ever, enkephalin transcription has in several recent stud-ies been suggested to be influenced by CREB rather thanby FOS (Borsook et al., 1994; Hyman et al., 1994; Konradiet al., 1995; Hermanson and Blomqvist, 1996a), suggest-ing that FOS in K-F may regulate other transcriptionalevents.
The FOS-labeling in PBsl and PBdl was sparser afterorofacial stimulation than after hindpaw stimulation. Thisobservation is consistent with the differences in termina-tion density in these subnuclei of fibers from the trigemi-nal and spinal dorsal horn, respectively (Cechetto et al.,1985; Slugg and Light, 1994; Feil and Herbert, 1995). PBslin the muskrat receives a contralateral input from trigemi-nothalamic tract neurons (Panneton et al., 1994), but inthe present study on the rat, the densest FOS-labeling inPBsl was found on the ipsilateral side, being in line withthe ipsilateral predominance of the trigeminal input toPBsl in this species.
In summary, the findings of this study show that manyneurons in the PB, particularly in PBel, PBem, and K-F,are activated by orofacial nociceptive stimulation. Thepattern of FOS labeling seems to reflect the terminationpattern in the PB of ascending medullary dorsal hornfibers. Thus, the topographical differences in the spinaland trigeminal terminations in the PB correspond todifferences in the distribution of nociceptive-responsiveneurons. This implies that noxious information from differ-
FOS IN PB AFTER OROFACIAL NOXIOUS STIMULATION 121

ent body parts may give rise to distinct responses byactivation of the different PB subnuclei.
LITERATURE CITED
Alden, M., J.-M. Besson, and J.-F. Bernard (1994) Organization of theefferent projections from the pontine parabrachial area to the bednucleus of the stria terminalis and neighboring regions: A PHA-L studyin the rat. J. Comp. Neurol. 341:298–314.
Aguero, A., M. Arnedo, M. Gallo, and A. Puerto (1993) The functionalrelevance of the lateral parabrachial nucleus in lithium chloride-induced aversion learning. Pharmacol. Biochem. Behav. 45:973–978.
Bellavance, L.L., and A.J. Beitz (1996) Altered c-fos expression in theparabrachial nucleus in a rodent model of CFA-induced peripheralinflammation. J. Comp. Neurol. 366:431–447.
Bernard, J.F., and J.M. Besson (1990) The spino(trigemino)pontoamygda-loid pathway: Electrophysiological evidence for an involvement in painprocesses. J. Neurophysiol. 63:473–490.
Bernard, J.F., M. Peschanski, and J.M. Besson (1989) A possible spino(tri-gemino)-ponto-amygdaloid pathway for pain. Neurosci. Lett. 100:83–88.
Bernard, J.F., G.F. Huang, and J.M. Besson (1990) Effect of noxioussomesthetic stimulation on the activity of neurons of the nucleuscentralis of the amygdala. Brain Res. 523:347–350.
Bernard, J.F., J. Carroue, and J.M. Besson (1991) Efferent projections fromthe external parabrachial area to the forebrain: A Phaseolus vulgarisleucoagglutinin study in the rat. Neurosci. Lett. 122:257–260.
Bernard, J.F., M. Alden, and J.-M. Besson (1993) The organization of theefferent projections from the pontine parabrachial area to the amygda-loid complex: A Phaseolus vulgaris leucoagglutinin (PHA-L) study inthe rat. J. Comp. Neurol. 329:201–229.
Bernard, J.F., G.F. Huang, and J.M. Besson (1994) The parabrachial area:Electrophysiological evidence for an involvement in visceral nociceptiveprocesses. J. Neurophysiol. 71:1646–1660.
Bernard, J.F., R. Dallel, P. Raboisson, L. Villanueva, and D. Le Bars (1995)Organization of the efferent projections from the spinal cervical enlarge-ment to the parabrachial area and periaqueductal gray: A PHA-L studyin the rat. J. Comp. Neurol. 353:480–505.
Bertrand, F., and A. Hugelin (1971) Respiratory synchronizing function ofnucleus parabrachialis medialis: Pneumotaxic mechanisms. J. Neuro-physiol. 34:189–207.
Blomqvist,A., and K.J. Berkley (1992)Are-examination of the spino-reticulo-diencephalic pathway in the cat. Brain Res. 579:17–31.
Blomqvist, A., W. Ma, and K.J. Berkley (1989) Spinal input to theparabrachial nucleus in the cat. Brain Res. 480:29–36.
Blomqvist, A., O. Hermanson, H. Ericson, and D. Larhammar (1994)Activation of a bulbospinal opioidergic projection by pain stimuli in theawake rat. NeuroReport 5:461–464.
Borsook, D., C. Konradi, O. Falkowski, M. Comb, and S.E. Hyman (1994)Molecular mechanisms of stress-induced proenkephalin gene regula-tion: CREB interacts with the proenkephalin gene in the mousehypothalamus and is phosphorylated in response to hyperosmolarstress. Mol. Endocrinol. 8:240–248.
Burstein, R., R.J. Dado, and G.J. Giesler Jr. (1990) The cells of origin of thespinothalamic tract of the rat: A quantitative reexamination. Brain Res.511:329–337.
Cechetto, D.F., and C.B. Saper (1987) Evidence for a viscerotopic sensoryrepresentation in the cortex and thalamus in the rat. J. Comp. Neurol.262:27–45.
Cechetto, D.F., D.G. Standaert, and C.B. Saper (1985) Spinal and trigemi-nal dorsal horn projections to the parabrachial nucleus in the rat. J.Comp. Neurol. 240:153–160.
Chamberlin, N.L., and C.B. Saper (1992) Topographic organization ofcardiovascular responses to electrical and glutamate microstimulationof the parabrachial nucleus in the rat. J. Comp. Neurol. 326:245–262.
Chamberlin, N.L., and C.B. Saper (1994) Topographic organization ofrespiratory neurons to glutamate microstimulation of the parabrachialnucleus in the rat. J. Neurosci. 14:6500–6510.
Chan, R.K.W., and P.E. Sawchenko (1994) Spatially and temporally differ-entiated patterns of c-fos expression in brainstem catecholaminergiccell groups induced by cardiovascular challenges in the rat. J. Comp.Neurol. 348:433–460.
Clark, F.M., and H.K. Proudfit (1991) The projection of noradrenergicneurons in the A7 catecholamine cell group to the spinal cord in the rat
demonstrated by anterograde tracing combined with immunocytochem-istry. Brain Res. 547:279–288.
Clement, C.I., K.A. Keay, B.K. Owler, and R. Bandler (1996) Commonpatterns of increased and decreased Fos expression in midbrain andpons evoked by noxious deep somatic and noxious visceral manipula-tions in the rat. J. Comp. Neurol. 366:495–515.
Coimbra, F., and A. Coimbra (1994) Dental noxious input reaches thesubnucleus caudalis of the trigeminal complex in the rat, as shown byc-fos expression upon thermal or mechanical stimulation. Neurosci.Lett. 173:201–204.
Craig, A.D. (1995) Distribution of brainstem projections from spinal laminaI neurons in the cat and the monkey. J. Comp. Neurol. 361:225–248.
Craig, A.D., M.C. Bushnell, E.-T. Zhang, and A. Blomqvist (1994) Athalamic nucleus specific for pain and temperature sensation. Nature372:770–773.
Feil, K., and H. Herbert (1995) Topographic organization of spinal andtrigeminal somatosensory pathways to the rat parabrachial and Kolliker-Fuse nuclei. J. Comp. Neurol. 353:506–528.
Floyd, N.S., K.A. Keay, and R. Bandler (1996) A calbindin immunoreactivedeep pain recipient thalamic nucleus in the rat. NeuroReport 7:622–626.
Fulwiler, C.E., and C.B. Saper (1984) Subnuclear organization of theefferent connections of the parabrachial nucleus in the rat. Brain Res.Rev. 7:229–259.
Girardot, M.-N., T.J. Brennan, M.E. Martindale, and R.D. Foreman (1987)Effects of stimulating the subcoeruleus-parabrachial region on thenon-noxious and noxious responses of T1-T5 spinothalamic tract neu-rons in the primate. Brain Res. 409:19–30.
Grigson, P.S., A.C. Spector, and R. Norgren (1994) Lesions of the pontineparabrachial nuclei eliminate successive negative contrast effects inrats. Behav. Neurosci. 108:714–723.
Hayashi, H., and T. Tabata (1989) Physiological properties of sensorytrigeminal neurons projecting to mesencephalic parabrachial area inthe cat. J. Neurophysiol. 61:1153–1160.
Herbert, H., M.M. Moga, and C.B. Saper (1990) Connections of theparabrachial nucleus with the nucleus of the solitary tract and themedullary reticular formation in the rat. J. Comp. Neurol. 293:540–580.
Herbert, H., and B. Bellintani-Guardia (1995) Morphology and dendriticdomains of neurons in the lateral parabrachial nucleus of the rat. J.Comp. Neurol. 354:377–394.
Hermanson, O., andA. Blomqvist (1995) Enkephalinergic and catecholamin-ergic neurons constitute separate populations in the rat Kolliker-Fuse/A7 region. Neurosci. Lett. 190:57–60.
Hermanson, O., and A. Blomqvist (1996a) Phosphorylated CREB in theparabrachial nucleus: Relation to preproenkephalin mRNA expression.Soc. Neurosci. Abstr. 22:116.
Hermanson, O., and A. Blomqvist (1996b) Subnuclear localization ofFOS-like immunoreactivity in the rat parabrachial nucleus after noci-ceptive stimulation. J. Comp. Neurol. 368:45–56.
Hermanson, O., M. Hallbeck, D. Larhammar, and A. Blomqvist (1995)Preprocholecystokinin mRNA expressing neurons in the rat parabra-chial nucleus. Soc. Neurosci. Abstr. 21:893.
Hunt, S.P., A. Pini, and G. Evan (1987) Induction of c-fos-like protein inspinal cord neurons following sensory stimulation. Nature 328:632–634.
Hylden, J.L.K., H. Hayashi, R. Dubner, and G.J. Bennett (1986) Physiologyand morphology of the lamina I spinomesencephalic projection. J.Comp. Neurol. 247:505–515.
Hyman, S.E., C. Konradi, L. Kobierski, R. Cole, P. Senatus, and D. Green(1994) Pharmacologic regulation of striatal proenkephalin gene expres-sion via transcription factor CREB. Prog. Clin. Biol. Res. 390:155–171.
Iwata, K., D.R. Kenshalo Jr., R. Dubner, and R.L. Nahin (1992) Dience-phalic projections from the superficial and deep laminae of the medul-lary dorsal horn in the rat. J. Comp. Neurol. 321:404–420.
Jacquin, M.F., K. Semba, M.D. Egger, and R.W. Rhoades (1983) Organiza-tion of HRP-labeled trigeminal mandibular primary afferent neurons inthe rat. J. Comp. Neurol. 215:397–420.
Jones, E.G. (1985) The Thalamus. New York: Plenum Press, 935 pp.Kato, J., S. Wakisaka, M.J. Tabata, Y. Sasaki, and K. Kurisu (1994)
Induction of Fos protein in the rat trigeminal nucleus complex duringan experimental tooth movement. Arch. Oral Biol. 39:723–726.
122 O. HERMANSON AND A. BLOMQVIST

Kluver, H., and P.C. Bucy (1937) ‘‘Psychic blindness’’ and other symptomsfollowing bilateral temporal lobectomy in Rhesus monkeys. Am. J.Physiol. 119:352–353.
Kluver, H., and P.C. Bucy (1939) Preliminary analysis of functions of thetemporal lobes in monkeys. Arch. Neurol. Psychiatry 42:979–1000.
Konradi, C., R.L. Cole, D. Green, P. Senatus, J.-C. Leveque, A.E. Pollack,S.J. Grossbard, and S.E. Hyman (1995) Analysis of the proenkephalinsecond messenger-inducible enhancer in rat striatal cultures. J. Neuro-chem.. 65:1007–1015.
Krukoff, T.L., T.L. Morton, K.H. Harris, and J.H. Jhamandas (1992)Expression of c-fos protein in rat brain elicited by electrical stimulationof the pontine parabrachial nucleus. J. Neurosci. 12:3582–3590.
Krukoff, T.L., K.H. Harris, and J.H. Jhamandas (1993) Efferent projectionsfrom the parabrachial nucleus demonstrated with the anterogradetracer Phaseolus vulgaris Leucoagglutinin. Brain Res. Bull. 30:163–172.
Lanteri-Minet, M., J. Weil-Fugazza, J. de Pommery, and D. Menetrey(1994) Hindbrain structures involved in pain processing as revaled bythe expression of c-FOS and other immediate early gene proteins.Neuroscience 58:287–298.
Lara, J.P., M.J. Parkes, L. Silva-Carvhalo, P. Izzo, M.S. Dawid-Milner, andK.M. Spyer (1994) Cardiovascular and respiratory effects of stimulationof cell bodies of the parabrachial nuclei in the anaesthetized rat. J.Physiol. (Lond.) 477:321–329.
Li, B.-H., and N.E. Rowland (1993) Dexfenfluramine induces Fos-likeimmunoreactivity in discrete brain regions in rats. Brain Res. Bull.31:43–48.
Light, A.R., M.J. Sedivec, E.J. Casale, and S.L. Jones (1993) Physiologicaland morphological characteristics of spinal neurons projecting to theparabrachial region of the cat. Somatosens. Mot. Res. 10:309–325.
Ma, W., and M. Peschanski (1988) Spinal and trigeminal projections to theparabrachial nucleus in the rat: Electron-microscopic evidence of aspino-ponto-amygdalian somatosensory pathway. Somatosens. Res.5:247–257.
Ma, W., A. Blomqvist, and K.J. Berkley (1989) Spino-diencephalic relaysthrough the parabrachial nucleus in the cat. Brain Res. 480:37–50.
Marfurt, C.F. (1981) The central projections of trigeminal primary afferentneurons in the cat as determined by the transganglionic transport ofhorseradish peroxidase. J. Comp. Neurol. 203:785–798.
Menetrey, D., and A.I. Basbaum (1987) Spinal and trigeminal projections tothe nucleus of the solitary tract: A possible substrate for somatovisceraland viscerovisceral reflex activation. J. Comp. Neurol. 255:439–450.
Menetrey, D., A. Gannon, J.D. Levine, and A.I. Basbaum (1989) Expressionof c-fos protein in interneurons and projection neurons of the rat spinalcord in response to noxious somatic, articular, and visceral stimulation.J. Comp. Neurol. 285:177–195.
Moga, M.M., H. Herbert, K.M. Hurley, Y.Yasui, T.S. Gray, and C.B. Saper(1990) Organization of cortical, basal forebrain, and hypothalamicafferents to the parabrachial nucleus in the rat. J. Comp. Neurol.295:624–661.
Murray, E.A., and M. Mishkin (1985) Amygdalectomy impairs crossmodalassociation in monkeys. Science 228:604–606.
Noguchi, K., and M.A. Ruda (1992) Gene regulation in an ascendingnociceptive pathway: Inflammation-induced increase in preprotachyki-nin mRNA in rat lamina I spinal projection neurons. J. Neurosci.12:2563–2572.
Norgren, R., and C.M. Leonard (1973) Ascending central gustatory path-ways. J. Comp. Neurol. 150:217–238.
Ogawa, H., and T. Hayama (1984) Receptive fields of solitario-parabrachialrelay neurons responsive to natural stimulation of the oral cavity inrats. Exp. Brain. Res. 54:359–366.
Ogawa, H., T. Hayama, and S. Ito (1982) Convergence of input from tongueand palate to the parabrachial nucleus neurons of rats. Neurosci. Lett.28:9–14.
Ogawa, H., T. Imoto, and T. Hayama (1984) Responsiveness of solitario-parabrachial relay neurons to taste and mechanical stimulation appliedto the oral cavity in rats. Exp. Brain. Res. 54:349–358.
Panneton, W.M., S.N. Johnson, and N.D. Christensen (1994) Trigeminalprojections to the peribrachial region in the muskrat. Neuroscience58:605–625.
Pfaller, K., and J. Arvidsson (1988) Central distribution of trigeminal andupper cervical primary afferents in the rat studied by anterogradetransport of horseradish peroxidase conjugated to wheat germ aggluti-nin. J. Comp. Neurol. 268:91–108.
Reilly, S., P.S. Grigson, and R. Norgren (1993) Parabrachial nucleus lesionsand conditioned taste aversion: Evidence supporting an associativedeficit. Behav. Neurosci. 107:1005–1017.
Sambucetti, L.C., and T. Curran (1986) The Fos protein complex isassociated with DNA in isolated nuclei and binds to DNA cellulose.Science 234:1417–1419.
Saper, C.B. (1995) The spinoparabrachial pathway: Shedding new light onan old path. J. Comp. Neurol. 353:477–479.
Slugg, R.M., and A.R. Light (1994) Spinal cord and trigeminal projections tothe pontine parabrachial region in the rat as demonstrated withPhaseolus vulgaris leucoagglutinin. J. Comp. Neurol. 339:49–61.
Sonnenberg, J.L., F.J. Rauscher III, J.I. Morgan, and T. Curran (1989)Regulation of proenkephalin by Fos and Jun. Science 246:1622–1625.
Strassman, A.M., and B.P. Vos (1993) Somatotopic and laminar organiza-tion of Fos-like immunoreactivity in the medullary and upper cervicaldorsal horn induced by noxious facial stimulation in the rat. J. Comp.Neurol. 331:495–516.
Swank, M.W., and I.L. Bernstein (1994) c-FOS induction in response to aconditioned stimulus after single trial taste aversion learning. BrainRes. 636:202–208.
Takayama, K., T. Suzuki, and M. Miura (1994) The comparison of effects ofvarious anesthetics on expression of FOS protein in the rat brain.Neurosci. Lett. 176:59–62.
Wang, L.G., H.M. Li, and J.S. Li (1994) Formalin induced FOS-likeimmunoreactive neurons in the trigeminal spinal caudal subnucleusproject to contralateral parabrachial nucleus in the rat. Brain Res.649:62–70.
Westlund, K.N., R.M. Bowker, M.G. Ziegler, and J.D. Coulter (1983)Noradrenergic projections to the spinal cord of the rat. Brain Res.263:15–31.
Wiberg, M., and A. Blomqvist (1984) The spinomesencephalic tract in thecat: Its cells of origin and termination pattern as demonstrated by theintraaxonal transport method. Brain Res. 291:1–18.
Wiberg, M., J. Westman, and A. Blomqvist (1986) The projection to themesencephalon from the sensory trigeminal nuclei: An anatomicalstudy in the cat. Brain Res. 399:51–68.
Xing, G., L. Zhang, L. Zhang, T. Heynen, T. Yoshikawa, M. Smith, S. Weiss,and S. Detera-Wadleigh (1995) Rat PPAR delta contains a CGG tripletand is prominently expressed in the thalamic nuclei. Biochem. Biophys.Res. Comm. 217:1015–1025.
Yamamoto, T., T. Shimura, N. Sakai, and N. Ozaki (1994) Representation ofhedonics and quality of taste stimuli in the parabrachial nucleus of therat. Physiol. Behav. 56:1197–1202.
Yasoshima Y., T. Shimura, and T. Yamamoto (1995) Single unit responses ofthe amygdala after conditioned taste aversion in conscious rats. Neu-roReport 6:2424–2428.
Young, R.F., V. Tronnier, and P.C. Rinaldi (1992) Chronic stimulation of theKolliker-Fuse nucleus region for relief of intractable pain in humans. J.Neurosurg. 76:979–985.
Zhao, Z.-Q., and A.W. Duggan (1988) Idazoxan blocks the action ofnoradrenaline but not spinal inhibition from electrical stimulation ofthe locus coeruleus and nucleus Kolliker-Fuse of the cat. Neuroscience25:997–1005.
Zhuo, H., S.J. Fung, and C.D. Barnes (1993) Opioid action on spinal cordreflexes due to dorsolateral pontine tegmentum stimulation. Neurophar-macology 32:621–631.
FOS IN PB AFTER OROFACIAL NOXIOUS STIMULATION 123





























