Embed Size (px)
Citation preview

Eur. J. Biochem. 79, 419-432 (1977)
Synthesis by Avian-Myeloblastosis-Virus RN A-D ependent DNA P ol ymer ase of Discrete Reverse Transcripts of Bacteriophage RNA Polyadenylated in vitro
Rent DEVOS, John VAN EMMELO, Paul CELEN, Elie GILLIS, and Walter FIERS
Laboratorium voor Moleculaire Biologie and Laboratorium voor Fysiologische Scheikunde, Rijksuniversiteit Gent
(Received January 3, 1977)
After polyadenylation in vitro, several viral RNAs become excellent templates for the synthesis of complementary DNA using RNA-dependent DNA polymerase from avian myeloblastosis virus, in the presence of oligo(dT) as a primer. If dTTP was the only deoxynucleoside triphosphate present, the reverse transcriptase catalyzed a ‘slippage’ reaction on the poly(A) tail, especially if the dTTP concentration exceeds 1 pM. In the case of MS2 RNA-poly(A), this poly(dT) synthesis could be specifically inhibited, but not completely abolished, by addition of dGTP. Lowering the dTTP con- centration also decreased the average size of the poly(dT) products formed. In the presence of all four dNTPs, but using a low dTTP concentration to avoid slippage of the primer, discrete partial transcripts could be synthesized, complementary to the 3’-terminal region of the viral RNA. Although the average size of the products increases at higher temperatures of incubation, the pattern of the labeled transcripts, as revealed by gel electrophoresis, was mainly dependent on the nature of the deoxynucleoside triphosphate present in limiting concentration. These results indicate that it is not the secondary structure of the template but its primary sequence that is the major determinant of the discrete partial transcripts. Restricted synthesis, using a combination of only three or less dNTPs, results in very small cDNA products. The successive incorporation of the different dNTPs added to the primer, using MS2 RNA-poly(A) or Qp RNA-poly(A) as a template, was in complete agreement with the known 3’-terminal sequence. Under the conditions used and with MS2 RNA-poly(A) or poly(A) as template, a ribosyl primer could not be substituted for the deoxyribosyl primer. Similarly, with MS2 RNA-oligo(C) only (dG),, was an efficient primer for reverse transcription. Ribonucleoside triphosphates were very poor substrates.
The DNA polymerase associated with oncorna viruses has been extensively used in vitro to synthesize cDNAs complementary to eukaryotic_messenger RNAs (e.g. [1,2]). Although most of the ‘variables of the reaction conditions have been optimized, many un- certainties still remain regarding the transcription of natural RNA templates into long representative DNA copies under various conditions [3]. It has often been observed that transcription predominantly leads to products shorter than the RNA template, both in the
endogenous reaction (i.e. incubation in vitro of dis- rupted oncorna viruses with dNTPs) and in the exo- genous reaction using purified reverse transcriptase [4]. Nevertheless, sequence representation in the product can be high especially in the endogenous reaction. The size of the cDNA can be increased by using a higher concentration of the dNTPs in the reaction mixture [5]. The secondary structure of the RNA template and/or the RNase H associated with the DNA polymerase, however, could also play a major role in determining the size of the product. Another characteristic feature of the reverse trans- criptase appears to be the high error-frequency during
This paper is XXXVI of a series; paper xxxV was by Min J0u3 Van Montagu & Fiers [Biochem. Biophys. Res. Commun. 73, 1083-1093 (197h)l ,-- . -,>. ~ ~. -..-
Abbreoiutions. AMV, avian myeloblastosis virus; (dT),, and DNA polymerization i61. (dG),, , decathymidylic acid and decadeoxyguanylic acid, respec- tively, each carrying a 5’-phosphate group; cDNA, complementary
nuclease H.
Some important problems regarding the reverse transcription concern the reaction mechanism of the
DNA; dNTP, deoxynucleoside triphosphate; RNase H; ribo- RNA-directed DNA synthesis and the fidelity and Enzymes, ATP : RNA adenylyltransferase (EC 2.7.7.19); AMV efficiency Of copying a given sequence Of an
RNA-dependent DNA polymerase (EC 2.7.7.7); RNase H (EC 3.1.4.34).
RNA template. Moreover, for some purposes it may be important to start polymerisation from a unique

420 Discrete Reverse Transcription Products of MS 2 RNA-poly(A)
site, in order to obtain products which all have an identical 5' initial sequence. This is readily achieved with oncorna virus genomes, which contain an ap- propriately positioned tRNA primer, but poses special problems in the case of a homopolymer primer as elongation by slippage rather than actual copying can readily occur.
Recently we developed a method to add a poly(A) tail to the 3' end of bacteriophage RNA, using Escherichia coli ATP:RNA adenylyltransferase [7,8]. It is now well established that poly(A)-terminating RNAs can be used as a template for RNA-dependent DNA polymerase. Since the complete primary struc- ture of MS2 RNA is known and good secondary structure models for this molecule exist [9], we have here a reliable model template, MS2 RNA-poly(A), to study some of the problems mentioned above. This report deals with the use of polyadenylated bacterio- phage RNA for studying some aspects of the reverse transcriptase reaction, and with the conditions for obtaining discrete cDNA products. The use of these procedures for sequencing the 3'-terminal regions of RNAs which cannot easily be labeled in uiuo will be documented in a forthcoming publication.
MATERIALS AND METHODS
Materials
All 3H- and I4C-labeled nucleotides were purchased from the Radiochemical Centre, Amersham. [ E - ~ ~ P ] dNTPs either were obtained from New England Nuclear, Boston, or from the Radiochemical Centre, Amersham. Unlabeled nucleotides were from Sigma or Boehringer; (dT),, , (dG),, , oligo(1) and (U), were purchased from P. L. Biochemicals Inc. (Milwaukee, Wis.) and poly(U) and poly(A) from Boehringer. MS2 RNA and QP RNA were prepared by sodium dodecylsulphate/phenol extraction of purified virus. ATP :RNA adenylyltransferase was purified from E. coli B/r as described previously [I 01. AMV-containing chicken plasma prepared by Dr J. W. Beard was a kind gift from Dr G. Marbaix and Dr G. Huez. AMV DNA polymerase was purified from the plasma ac- cording to Kacian and Spiegelman [l l] . Further pre- parations of purified AMV reverse transcriptase were generously supplied by Dr M. A. Chirigos (National Cancer Institute, Bethesda, Maryland) and Dr J. W. Beard (Life Sciences Inc., Gulfport, Florida).
Polyadenylation of Viral R N A
MS2 RNA and QP RNA were polyadenylated by means of E. coli ATP :RNA adenylyltransferase, and further purified by zonal centrifugation [7,8]. Reverse transcription of MS2 RNA-poly(A) purified by oligo (dT)-cellulose chromatography was not inhibited by
addition of a five-fold excess of unmodified MS2 RNA (results not shown). Therefore the material in the gradient peak fraction (which still contains unmodi- fied RNA) was used directly for the AMV reverse transcriptase reactions, without further purification.
RNA-Dependent D N A Polymerase Assay
The standard reaction mixture for synthesis of cDNA contained the following: 50 mM Tris-HC1 (pH 8.3), 20 mM KCl, 4 mM dithiothreitol, 10 mM MgC1, , 20 pg/ml actinomycin D, 50 pg/ml polyadeny- lated RNA (unless indicated otherwise), 10 - 20 pg/ml (dT),, , unlabeled dNTPs at 100 pM (unless indicated otherwise), labeled dNTPs as indicated and 20 units/ml AMV reverse transcriptase (unless indicated other- wise). The RNA solution generally contributed 5 - 10% sucrose to the reaction mixture. The template was preincubated for 15 min at 37 "C with the (dT),, in 0.1 M NaCl before addition to the reaction mixture. Then enzyme was added and the incubation was continued at 37 "C (unless indicated otherwise). At appropriate times samples were withdrawn formeasure- ment of the incorporation by trichloroacetic acid precipitation. Large-scale reactions were stopped by addition of EDTA to a final concentration of 20 mM; the mixture was extracted with phenol/chloroform/ isoamyl alcohol containing 1 % sodium dodecylsul- phate and passed through a Sephadex G-50 column (0.5 x 20 cm) equilibrated with 10 mM Tris-HC1 (pH 7.9,O.l M NaCl, 1 mM EDTA, at 4 "C. The excluded material was then ethanol-precipitated for 30 min at -70 "C after addition of 100 pg carrier tRNA. The material was dissolved in Tris/borate/EDTA electro- phoresis buffer containing 7 M urea [12] and boiled for 3 min prior to electrophoresis. When the reaction mixture was only 10 p1, EDTA, 40 pg tRNA and solid urea were added, and the mixture was boiled for 3 min.
Polyacrylamide Gel Electrophoresis
A slab gel apparatus was used as described before [13]. The cDNA products were examined in poly- acrylamide gels, containing Tris/borate/EDTA buffer (0.09 M Tris/borate, 2.5 mM EDTA, pH 8.3) in 7 M urea. The buffer in the reservoirs was Tris/borate/ EDTA without urea. Before loading, sucrose to a final concentration of 6% and a set of tracking dyes (xylene cyanol FF, trypane red, bromephenol blue, eosine and fluoresceine) were added to the samples.
RESULTS
RNA-Dependent D N A Synthesis on Polyadenylated MS2 R N A
Bacteriophage MS2 RNA to which a 3'-terminal poly(A) tail has been added [7], functions as an efficient

R. Devos. J. van Emmelo, P. Celen, E. Gillis, and W. Fiers 42 1
bottom
A to+ Omin
2min A l 6
" 0 10 15 2 0 0 10 15 20 0 30 60
Fraction number Incubation time (min)
Fig. 1. Effect of length of the poly iA) tail, added lo MS2 RNA. on subsequent synthesis o f c D N A by AMV reoerse trunscriptase. (A) 40 pg of MS2 RNA was polyadenylated for 0, 2 ,4 and 6 min at 37 "C, with ATP: RNA adenylyltransfelase using ['4C]ATP (15.6 Cijmol), and puri- fied by zonal centrifugation as described before [7]. Absorbance at 254 nm (-); acid-precipitable radioactivity (- - - -). (Fractions are numbered from top to bottom). (B) 0.2 pg of the RNA in the peak fractions of each gradient was incubated at 37 "C with 0.4 unit of AMV RNA-dependent DNA polymerase in a 0.02-ml reaction mixture with [3H]dGTP (38 pM, 13 Cijmmol) as a labeled precursor (cf. Materials & Methods). After appropriate times 4-pl samples were withdrawn for acid-precipitable radioactivity measurements
Table 1. Effect of a po ly (A) tail and an oligo(dT) primer on the synthesis of DNA, complementary to MS2 RNA, by A M V reverse transcriptase 0.1-ml reaction mixtures containing 1 unit AMV reverse transcriptase and 3 pg MS2 RNA or MS2 RNA-poly(A) were incubated for 60 min at 37 "C with or without 300 pmol (dT),,. [3H]dGTP was used at 240 pM (1 Ci/mmol), the other dNTPs at 100 pM
Template Primer dGMP incorporated (acid-precipitable material)
pmol MS2 RNA (dT),, 0.27 MS2 RNA - 0.28
MS2 RNA-poly(A) - 0.50 MS2 RNA-pOly(A) (dT),, 72
template for the AMV reverse transcriptase. Fig. 1(B) shows the activity exhibited by the AMV enzyme with MS2 RNA, polyadenylated for different times with ATP :RNA adenylyltransferase. The RNA was first purified by zonal centrifugation (Fig. 1A) and each MS2 RNA peak fraction was used for synthesis of cDNA. Our previous results indicated that in- creased incubation time with transferase results in longer poly(A) tails while the fraction of adenylated molecules remains almost constant [7]; 4 min of in- cubation with the ATP transferase seems to ensure an optimal length of poly(A) added (60- 100 AMP residues) for the reverse transcription (Fig. 1B). Lower incorporation is obtained with the 6-min transferase product; possibly the reverse transcriptase becomes
more engaged in poly(dT) synthesis on the longer poly(A) tail (vide infra) . The reverse transcription is totally dependent on the presence of oligo(dT) as a primer (Table 1). Unmodified MS2 RNA either with or without primer added was not significantly trans- cribed. Actinomycin D (tested between 0 and 40 pg/ml) did not affect the total incorporation (data not shown).
Fig. 2 demonstrates that if one of the dNTPs is present in a lower concentration, the initial rate as well as the total incorporation is considerably di- minished. Lowering the dCTP or the dATP concentra- tion to 1 pM resulted in a two-fold decrease of total incorporation. Thus it is important to use all four dNTPs at high concentration (> 50 pM) in order to obtain optimal yield of cDNA.
'Slippage' of AMV Reverse Transcriptase on the Poly(A) Tail of MS2 RNA-poly(A)
In the presence of [3H]dTTP as the only substrate the AMV DNA polymerase can synthesize poly(dT) using MS2 RNA-poly(A) as a template (Fig. 3A). On the other hand, when the same reaction was carried out in the presence of [a-32P]dGTP, no significant in- corporation of dGMP could be observed (Fig. 3B), while the rate of the [3H]dTMP incorporation was approximately three times slower than in the absence of dGTP. Fig. 3C shows that when dATP and dCTP were also present in the reaction, both [3H]dTMP and [32P]dGMP were incorporated, but the rate of dTMP incorporation was comparable to that observed with dGTP and dTTP alone. These results show that

422 Discrete Reverse Transcription Products of MS2 RNA-poly(A)
Incubation time (min)
Fig. 2. Effect of deoxynucleoside triphosphate concentration on the rate of synthesis of DNA complementary to M S 2 RNA-poly(A) by A M V reuerse transcriptase. 0.02-ml reaction mixtures were used and the dNTP concentration was varied as indicated. 2-pl aliquots were withdrawn at indicated times, acid-precipitated and counted. (A) Variation of dTTP and dCTP; [3H]dGTP (38 pM, 13 Ci/mmol). (B) Variation of dATP and dGTP; [3H]dCTP (32 pM, 23 Ci/mmol)
A B C 8 - - 4 2
2 I
a -
N 0%
0 30 60 90 120 30 60 90 120 30 60 90 120 Incubation t ime (rnin)
Fig. 3. Transcription of M S 2 RNA-poly(A) by AMP’ polymerase in the presence of dTTP, dTTP plus dGTP, and all four deoxynurleoside triphosphates. 0.1-ml reaction mixtures (Materials & Methods) containing 1 pg MS2 RNA-poly(A) and 0.1 unit of AMV DNA polymerase were incubated at 37 “C in the presence of (A) 30 pM [3H]dTTP (4.3 Ci/mmol), (B) 30 pM [3H]dTTP (as in A) plus 30 pM [cc-~’P]~GTP (2 Ci/mmol), (C) 30 pM [3H]dTTP and 30 pM [w3’P]dGTP (as in B) plus dATP and dCTP, 100 pM each. At the indicated times 10-p1 samples were withdrawn for measurement of acid-precipitable material
[u-~’P]~GTP inhibits dTMP incorporation. That this inhibition was not due specifically to the presence of an (w3’P)-labeled dNTP is demonstrated by the ex- periment shown in Fig. 4, where the reactions with dTTP alone, dTTP plus dATP or dTTP plus dCTP resulted in a three times higher incorporation of dTMP compared to the reaction with dTTP plus dGTP. Undoubtedly, the AMV reverse transcriptase carries out a ‘slippage’ reaction on the poly(A) tail of MS2 RNA-poly(A), as has also been reported for globin mRNA [14]. This ‘slippage’ can only be slowed down by a second dNTP if the latter is dGTP, because this supplies the first three nucleotides complementary to the 3‘ end of MS2 RNA and in this way anchors the primer [15]. The rate and total amount of synthesis of
poly(dT) by reverse transcriptase using poly(A). (dT),, as template-primer cannot be lowered by adding dGTP to the reaction (results not shown). Furthermore the dTMP incorporation with poly(A) as a template continued linearly, whereas with MS2 RNA-poly(A), poly(dT) synthesis leveled off (Fig. 3A). The addition of rGTP did not lower the uncontrolled poly(dT)- synthesizing activity on MS2 RNA-poly(A) (Fig. 4B). Obviously it is much more difficult for the enzyme to incorporate a ribonucleotide at a complementary site than to await ‘slippage’ of the poly(dT) product in order to incorporate a new dTMP residue.
We found that all commercial primer preparations tested did contain significant amounts of oligo(dT) series with lengths smaller and larger than ten (Fig. 5).

R. Devos, J. van Emmelo, P. Celen, E. Gillis, and W. Fiers 423
I A
30 60 90 Incubation time (min)
Fig. 4. Transcription of M S 2 RNA-poly(A) by AMV reverse transcriptase in the presence of two nucleoside triphosphates. The reaction mix- tures (50 pl), as in the legend to Fig. 3 , were incubated at 37 “C and 10-p1 aliquots were withdrawn at the indicated times. (A) 30 p M [3H]dTTP (4 Ci/mmol) either alone (A-A), or in the presence of 100 pM dATP (0- O), or 100 pM dCTP (0 -0). or 100 pM dGTP (M- m). (B) 30 pM [3H]dTTP (4 Ci/mmol) either alone (0-0) or in the presence of 60 pM GTP (A-A), or 120 pM GTP (o-o), or 100 pM dGTP
This was demonstrated by 5’-labeling using [y-32P]ATP and TCinduced polynucleotide kinase and analysis on polyacrylamide gel. In the further experiments [5’-32P] oligo(dT) which mainly consisted of (dT),, was used as a reference. The absolute length of these products was determined by ‘wandering spot’ analysis (Dr J. Merregaert, unpublished). Another batch of oligo (dT) which mainly contained (dT),, was used for the actual elongation reactions. When contamination with other homologues was too high, however, primers with defined length were prepared by electrophoresis on a 24% polyacrylamide gel and elution.
The length of the poly(dT) product obtained with poly(A). (dT),, as a template decreases by lowering the concentration of dTTP in the reaction mixture (Fig. 5). If dTTP was used at a concentration of 0.1 pM, elongation of the primer by only one dTMP could be observed. That this single product was indeed formed by addition (and not by terminal exchange) of one dTMP residue has been confirmed using purified (dT),, primer (not shown). When the sub- strate was present at a concentration of 1 pM, limited extension of the primer was observed, while at 30 pM large poly(dT) molecules were synthesized. Similarly, using MS2 RNA-poly(A).(dT),, as a template and dTTP at a concentration of 30 pM as a substrate large poly(dT) products were formed (Fig. 6A), while little or almost no elongation of (dT),, took place at 1 pM and 0.1 pM dTTP respectively (Fig. 6D,G). On the other hand, when dGTP was also present, total dTMP incorporation diminished at a dTTP concentration of 30 pM (Fig. 6B), while at lower dTTP concentration very small discrete products were formed (Fig. 6E, H). In conclusion, in order to reduce the ‘slippage’ of the AMV reverse transcriptase on the poly(A) tail of
MS2 RNA-poly(A) and hence to obtain homogene- ously initiated products, it is important to decrease the dTTP concentration. Lowering of the incubation temperature also resulted in the formation of smaller oligo(dT) products (data not shown).
Discontinuous Trunscription of Polyadenyluted RNA
Fig. 6C, F, I shows the size distribution of the reverse transcripts obtained from MS2 RNA-poly(A) in the presence of different concentrations of [ C ( - ~ ~ P ] dTTP, the other three dNTPs being kept at 100 pM. At a dTTP concentration of 30 pM (Fig. 6C) a hetero- geneous size distribution of cDNA is observed, with some discrete bands of more highly labeled material. Since the four dNTPs were present at a moderately high concentration, one could indeed expect that rather long cDNA molecules would be formed [l]. Moreover, this cDNA mainly arises from the extension of large poly(dT) stretches and hence the 5‘ end of these products is heterogeneous. This conclusion is based on pyrimidine tract analysis of products made in the presence of 5’-labeled (dT),, primer (not shown). On the other hand, a dTTP concentration of 1 pM (Fig. 6F) predominantly leads to synthesis of products with a defined length, and even smaller, discrete bands are observed at a dTTP concentration of 0.1 pM (Fig. 61). We believe that at low dTTP concentrations ‘slippage’ of the primer is prevented and only those molecules which are hybridized adjacent to the natural 3‘ end of MS2 RNA have a chance to become elongat- ed. This leads to a homogeneous 5’ end of the product, starting with the original (dT),, primer. But the low dTTP concentration also creates an unfavorable con- dition for transcription and results in discontinuous

424 Discrete Reverse Transcription Products of MS 2 RNA-poly(A)
Fig. 5. Autoradiogram of P’P]poly(dT) product synthesized using poly(A) as a template in the presence of different concentrations of [ u - ~ ’ P ] ~ T T P . 10-p1 reaction mixtures were made up, containing 1 Fg poly(A), 30 pmol (dT),,, 0.2 unit AMV DNA polymerase and [E-~’P]~TTP (70 Ci/mmol). After 60 min at 25 “C the reaction mixtures were made 7 M in urea and analysed by electrophoresis on a 24% polyacrylamide gel in 7 M urea. Slot 1: commercial [5‘-32P](dT),-,, preparation; slot 2: [32P]cDNA synthesized in the presence of [ U - ~ ~ P J ~ T T P at 0.1 pM; slot 3: [u-~’P]~TTP at 1 pM; slot 4: [cz-~’P]~TTP at 30 pM. (Note that the primer used for reactions 2, 3 and 4 was homogeneous (dT),, and different from the labeled reference sample in slot 1 . )
synthesis by the AMV reverse transcriptase. In further experiments we have chosen a dTTP concentration of 1 pM for discontinuous synthesis of cDNA, because it allows homogeneous initiation of heteropolymer synthesis (dTTP < 30 pM) and produces discrete bands
Fig. 6. Autoradiogram of f ’P]cDNA synthesized by AMV DNA polymerase with MS2 RNA-poly(A) and different concentrations of [ci 32P]dTTP. 10-p1 reaction mixtures as in the legend to Fig. 5, but with 0.4 pg MS2 RNA-poly(A) as template, were incubated for 60 min at 37 “C and electrophoresed in a 10-24% discontinuous polyacrylamide gel containing 7 M urea. (A) cDNA synthesized in the presence of [ce3’P]dTTP at 30 pM (100 Cijmmol). (B) As for (A) but with dGTP at 100 pM. (C) As for (B) but with also dATP and dCTP each at 100 pM. (D), (E) and (F) As for (A), (B) and (C) respectively, but with [G~-~’P]~TTP at 1 pM. (G), (H) and (I) As for (A), (B) and (C) respectively, but with [E-~’P]~TTP at 0.1 pM
of appropriate length (dTTP > 0.1 pM). Furthermore, we could ascertain the precise elongation of the primer molecules, which are hybridized adjacent to the original 3’ end of MS2 RNA by preincubation in a reaction mixture in which dGTP is the only substrate present. This predominantly leads to the addition of three dGMP residues to the primer (vide infiu), at which point further elongation would require dTTP.
Fig. 7 shows typical electrophoretic patterns of the DNA transcripts using MS2 RNA-poly(A) as a template, in the presence of all four dNTPs, with dTTP kept at 1 pM. Discrete fragments ranging from approximately 50 to 300 nucleotides in length are pre- sent. The labeling pattern of the smaller transcripts also differs according to the [E-~’P]~NTP used, an

R. Devos, J. van Emmelo, P. Celen, E. Gillis, and W. Fiers 425
Fig. 7 . Discontinuous synthesis of cDNA with polyadenylated MS2 RNA by AMV reverse transcriptase. 0.1-ml reaction mixtures (Materials and Methods) containing 2 units AMV DNA polymerase, 300 pmol (dT),, and 5 pg MS2 RNA-poly(A), were incubated for 60 min at 25 “C, in the presence of three unlabeled dNTPs at 100 pM each (except dTTP at 1 pM) and one [w3’P]dNTP at 7.5 pM (except [ U - ~ ~ P I ~ T T P at 1 pM). The mixtures were preincubated for 30 min at 25 “C with dGTP as the only dNTP present. After synthesis, the cDNA was recovered by phenolization followed by gel filtration and precipitation with two volumes of ethanol. After centrifugation the material was redissolved in 20 pl electrophoresis buffer containing 7 M urea, and electrophoresed in a 10% poly- acrylamide gel. (A) Slot 1 : [5‘-32P](dT),, reference; slot 2: 32P- labeled Simian virus 40 Hind restriction fragment K (210 nucleo- tides in length); slot 3: [32P]cDNA synthesized with MS2 RNA- poly(A) in the presence of [ E - ~ ~ P ] ~ A T P (90 Ci/mmol); slot 4: as slot 3, but with [M-~’P]~CTP (90 Ci/mmol) as the labeled dNTP; slot 5: as slot 4, but with [M-~’P]~CTP (15 Ci/mmol). The approxi- mate length of the indicated components has been confirmed by pyrimidine tract analysis. (B) Slot 1: as slot 3 in (A), but with [w3’P]dTTP (100 Ci/mmol) as the labeled dNTP; slot 2: as slot 3 in (A), but with [ G I - ~ ~ P J ~ G T P (100 Ci/mmol) as the labeled dNTP
Fig. 8. Discontinuous synthesis of cDNA with MS2 RNA-poly(A) and QB RNA-poly(A) by A M V reverse transcriptase. [3ZP]~DNA complementary to MS2 RNA-poly(A) and QB RNA-poly(A) was synthesized in 25-p1 reaction mixture and electrophoresed as described in the legend to Fig. 7. Labeled dNTPs at 7.5 pM and unlabeled dNTPs at 100 pM (except dTTP at 1 pM) were used. Slot 1 : [32P]cDNA products of MS2 RNA-poly(A), in the presence of [ L Y - ~ ~ P I ~ A T P (1 00 Ci/mmol); slot 2: as slot 1 , but with [w3’P]dCTP (100 Ci/mmol) as the labeled dNTP; slot 3 : [3ZP]cDNA products of QB RNA-poly(A) in the presence of [ u - ~ ~ P I ~ A T P (100 Ci/mmol); slot 4: as slot 3, but with [ cx -~~PI~CTP (100 Ci/mmol) as the labeled dNTP
effect which is probably enhanced by their being pre- sent at low concentration (7.5 pM), while the con- centration of the other unlabeled dNTPs (except dTTP) was 100 pM each. The approximate length of the transcripts was determined by analysis of the pyrimi- dine tracts. These results were in full agreement with the expected distribution from the known nucleotide sequence of the 3‘ end [16]. Qfi RNA-poly(A), used as a template under similar conditions, also yields many discrete products in the same size class as obtained

426
Fig. 9. Effect of the incubation temperature on the pattern of the discrete reverse transcripts synthesized by A MV DNA polymerase using MS2 RNA-poly(A) as a template. 0.8 pg MS2 RNA-poly(A), purified by (dT)-cellulose chromatography, was preincubated for 20 rnin at 25 "C with dGTP at 100 pM in a 40-pl reaction mixture, containing 1 unit AMV reverse transcriptase. [ G I - ~ ~ P ] ~ T T P (100 Ci/ mmol, final concentration 1 pM), together with dATP and dCTP (final concentration 100 pM each) were then added (final volume 50 ~ 1 ) and the incubation was continued for 30 min at the indicated temperature. After phenolization and gel filtration, the cDNA was electrophoresed as described in the legend of Fig. 7
with MS2 RNA-poly(A) (Fig. 8). Obviously, the pattern of bands obtained depends on the template RNA.
We further examined the mechanism by which these incomplete, discrete reverse transcripts originate on the MS2 RNA-poly(A) template. If regions of extensive secondary structure are responsible for the discontinuous synthesis, one would expect that at
Discrete Reverse Transcription Products of MS2 RNA-poly(A)
1 5 OIo 'XP.
Fig. 10. Effect of the nature of the limiting deoxynucleoside triphos- phate on the discontinuous transcription of MS2 RNA-poly(A) by AMV reverse transcriptase. Four 10-pl reaction mixtures containing 0.5 pg MS2 RNA-poly(A) and 0.2 unit of AMV reverse transcriptase were preincubated for 30 rnin at 25 "C in the presence of [cx-~'P]~GTP (80 Ci/mmol) at 1 pM. The incubation was then continued for 60 rnin at 25 "C without further addition of dNTPs (e), or by adding dGTP to a final concentration of 500 pM, together with either dTTP (b), dCTP (c) or dATP (d) at 0.1 pM and two other dNTPs at 100 pM each. After addition of solid urea to make 7 M, the mixtures were heated for 3 rnin at 100 "C and loaded directly on a discontinuous 15-24% polyacrylamide gel. (a) [5'-32P](dT)+ll reference
higher temperatures some hairpins would become destabilized resulting in the disappearance of cor- responding bands in the transcription pattern. The autoradiographies illustrated in Fig. 9, however, show that although the average size of the reverse trans- cripts increases at higher temperatures, no discrete transcripts of intermediate size disappear. Hence these experiments do not allow one to correlate the discrete
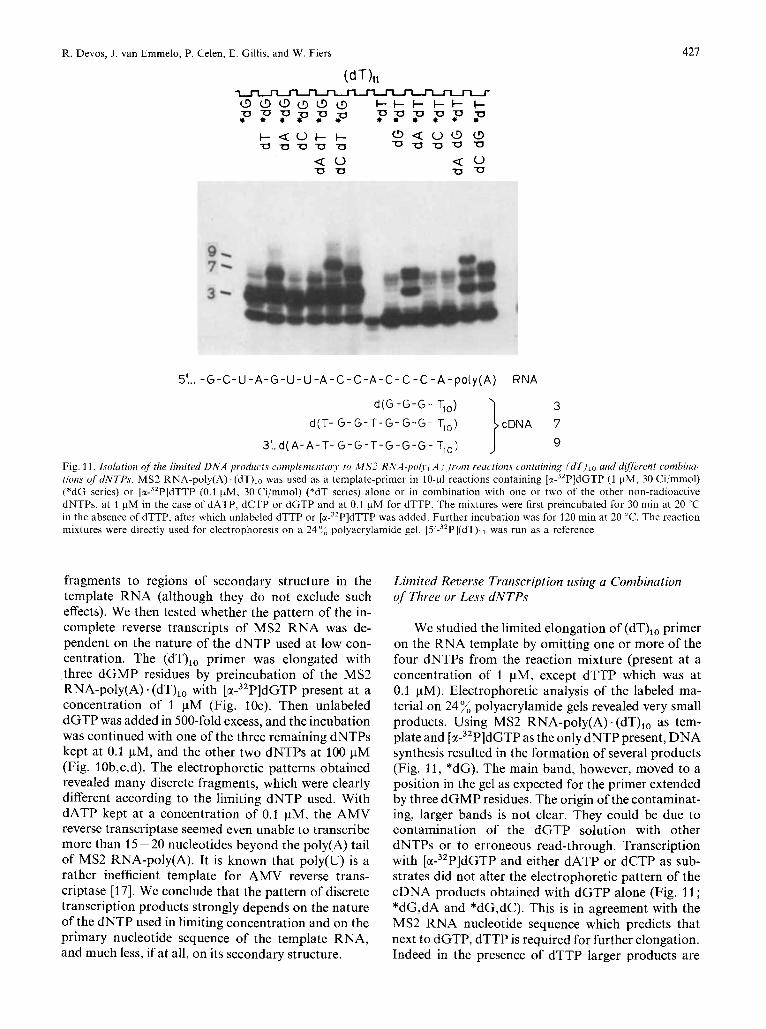
R. Devos, J . van Emmelo, P. Celen, E. Gillis, and W. Fiers 421
5‘ ... - G - C - U - A - G - U - U - A - C - C - A - C - C - C - A - p o l y ( A ) RNA
3 cDNA 7 I 9
d ( G - G - G - T,o) d (T- G - G - T - G - G - G - T,o)
31. d( A-A-T- G - G - T - G - G - G - T,o)
Fig. 11. I,wlution of the limited DNA products complenic~ntur~~ to MS2 RNA-Iio!,.( A J ,froin reuctions containing (dT)lo and dgferent combina- tions 0j’dNTP.r. MS2 RNA-poly(A) (dT)lo was used as a template-primer in 10-pl reactions containing [E-~’P]~GTP (1 pM, 30 Ci/mmol) (*dG series) or [E-~’P]~TTP (0.1 pM, 30 Ci/mmol) (*dT series) alone or in combination with one or two of the other non-radioactive dNTPs, at 1 pM in the case of dATP, dCTP or dGTP and at 0.1 pM for dTTP. The mixtures were first preincubated for 30 min at 20 “C in the absence of dTTP, after which unlabeled dTTP or [E-~’P]~TTP was added. Further incubation was for 120 min at 20 “C. The reaction mixtures were directly used for electrophoresis on a 2404 polyacrylamide gel. [5’-32P](dT)ll was run as a reference
fragments to regions of secondary structure in the template RNA (although they do not exclude such effects). We then tested whether the pattern of the in- complete reverse transcripts of MS2 RNA was de- pendent on the nature of the dNTP used at low con- centration. The (dT),, primer was elongated with three dGMP residues by preincubation of the MS2 RNA-poly(A) . (dT),, with [M-~’P]~GTP present at a concentration of 1 pM (Fig. 10e). Then unlabeled dGTP was added in 500-fold excess, and the incubation was continued with one of the three remaining dNTPs kept at 0.1 pM, and the other two dNTPs at 100 pM (Fig. 10b,c,d). The electrophoretic patterns obtained revealed many discrete fragments, which were clearly different according to the limiting dNTP used. With dATP kept at a concentration of 0.1 pM, the AMV reverse transcriptase seemed even unable to transcribe more than 15 - 20 nucleotides beyond the poly(A) tail of MS2 RNA-poly(A). It is known that poly(U) is a rather inefficient template for AMV reverse trans- criptase [17]. We conclude that the pattern of discrete transcription products strongly depends on the nature of the dNTP used in limiting concentration and on the primary nucleotide sequence of the template RNA, and much less, if at all, on its secondary structure.
Limited Reverse Transcription using a Combination of Three or Less dNTPs
We studied the limited elongation of (dT),, primer on the RNA template by omitting one or more of the four dNTPs from the reaction mixture (present at a concentration of 1 pM, except dTTP which was at 0.1 pM). Electrophoretic analysis of the labeled ma- terial on 24 % polyacrylamide gels revealed very small products. Using MS2 RNA-poly(A) . (dT),, as tem- plate and [w3’P]dGTP as the only dNTP present, DNA synthesis resulted in the formation of several products (Fig. 11, *dG). The main band, however, moved to a position in the gel as expected for the primer extended by three dGMP residues. The origin of the contaminat- ing, larger bands is not clear. They could be due to contamination of the dGTP solution with other dNTPs or to erroneous read-through. Transcription with [M-~’P]~GTP and either dATP or dCTP as sub- strates did not alter the electrophoretic pattern of the cDNA products obtained with dGTP alone (Fig. 3 1 ; *dG,dA and *dG,dC). This is in agreement with the MS2 RNA nucleotide sequence which predicts that next to dGTP, dTTP is required for further elongation. Indeed in the presence of dTTP larger products are

428 Discrete Reverse Transcription Products of MS 2 RNA-poly(A)
0 U
I- -0
5!..-G-A-U-C-U-G-C-U-U-U-G-C-C-C-U-C-U-C-U-C-C-U-C-C-C-(A)-pO~y(Al R N A 3
cDNA 14 i 23
d(G- G-G-T,,) d(G- G- G- A-G- A-G- A-G- G-A-G-G-G-T,,)
31 .... d(A-G-A-C-G-A-A-A-C-G- G- G- A-G- A-G- A-G- G-A-G-G-G-To)
Fig. 12. Limiled cDNA products c o n z ~ ~ ~ e n r r r i l a ~ } ~ to Qfl RA’A-~J(I!,.( A J s~nthrsized by A M V r n ‘ e r , ~ e truriscriptusr , f h n reactions contuining u combination of dNTPs. 10-pl reaction mixtures containing QB RNA-poly(A) . (dTjlo and either [X-~~PI~CTTP (*dG series) or [x-~’P]~TTP (*dT series) alone, or in combination with one or two of the other unlabeled dNTPs were used (concentrations as indicated in the legend to Fig. 11). The mixtures were incubated for 60 min at 20 “C and subjected to analysis on a 24n4 polyacrylamide gel as in Fig. 11. Dye markers: CFF = xylene cyanol, TR = trypane red, EOS = eosine. Reference: [5’-32P](dT)+~~.
formed. The main transcript corresponds to an elongation of the primer molecule with seven nucleo- tides (Fig. 11, *dG,dT). Next, dATP was required for further elongation of the primer by two nucleotides (Fig. 11 ; *dG,dT,dA). Nevertheless, the four-nucleo- tide elongation product is still present in considerable amount. This intermediate can perhaps be explained by the hairpin structure at the 3‘ end of MS2 RNA [16]. This hairpin may form an obstacle for the reverse transcriptase thus slowing down its further trans- cription under these conditions. Alternatively, it may be that the enzyme has considerable difficulty in in-
corporating two succeeding nucleotides present in low concentration (in this case dGTP).
When [ X - ~ ~ P I ~ T T P is present as the only substrate at a concentration of 0.1 pM, the main reaction is the addition of a single residue, thus forming (dT),, . This reaction can occur with any (dT),, primer hybri- dized along the poly(A) tail of the MS2 RNA template and is similar to the reaction observed with poly(A) (cf. above and Fig. 5). The primer (dT),, which is hybridized just adjacent to the heteropolymer end of MS2 RNA, however, can only be elongated if dGTP is also present (*dT,dG); we have verified that in this
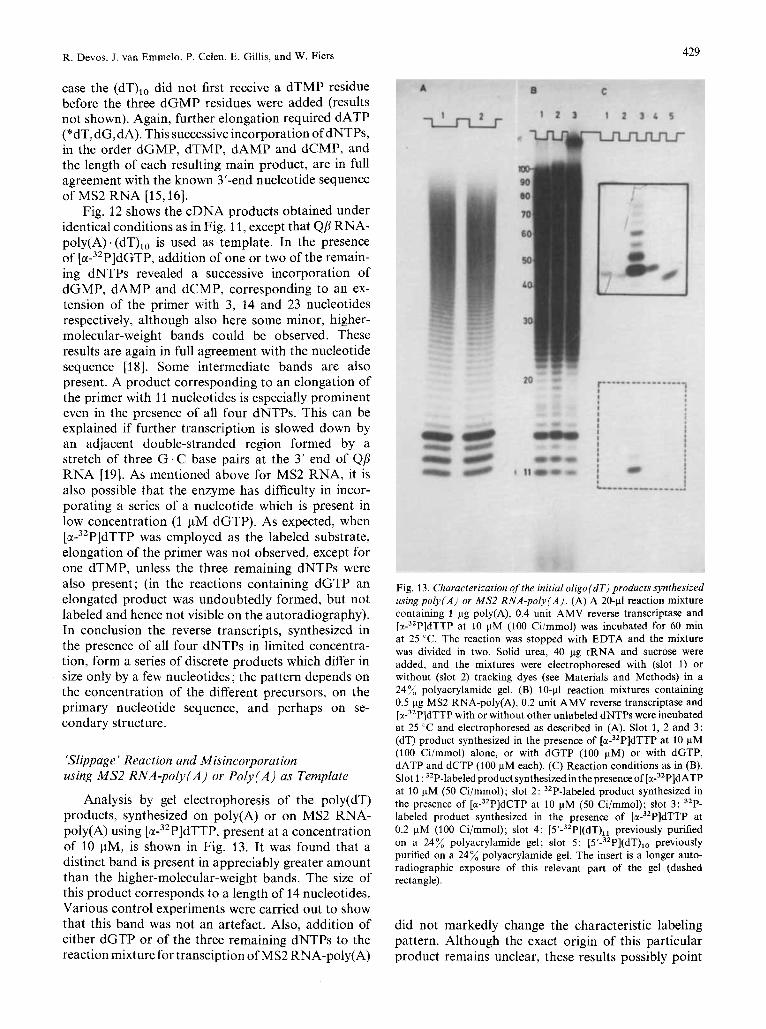
R. Devos, J. van Emmelo, P. Celen, E. Gillis, and W. Fiers 429
case the (dT),, did not first receive a dTMP residue before the three dGMP residues were added (results not shown). Again, further elongation required dATP (*dT, dG,dA). This successive incorporation of dNTPs, in the order dGMP, dTMP, dAMP and dCMP, and the length of each resulting main product, are in full agreement with the known 3’-end nucleotide sequence of MS2 RNA [15,16].
Fig. 12 shows the cDNA products obtained under identical conditions as in Fig. 11, except that Qp RNA- poly(A). (dT),, is used as template. In the presence of [LX-~~PI~GTP, addition of one or two of the remain- ing dNTPs revealed a successive incorporation of dGMP, dAMP and dCMP, corresponding to an ex- tension of the primer with 3, 14 and 23 nucleotides respectively, although also here some minor, higher- molecular-weight bands could be observed. These results are again in full agreement with the nucleotide sequence [18]. Some intermediate bands are also present. A product corresponding to an elongation of the primer with 11 nucleotides is especially prominent even in the presence of all four dNTPs. This can be explained if further transcription is slowed down by an adjacent double-stranded region formed by a stretch of three G . C base pairs at the 3‘ end of Qp RNA [19]. As mentioned above for MS2 RNA, it is also possible that the enzyme has difficulty in incor- porating a series of a nucleotide which is present in low concentration (1 pM dGTP). As expected, when [LX-~’P]~TTP was employed as the labeled substrate, elongation of the primer was not observed, except for one dTMP, unless the three remaining dNTPs were also present; (in the reactions containing dGTP an elongated product was undoubtedly formed, but not labeled and hence not visible on the autoradiography). In conclusion the reverse transcripts, synthesized in the presence of all four dNTPs in limited concentra- tion, form a series of discrete products which differ in size only by a few nucleotides; the pattern depends on the concentration of the different precursors, on the primary nucleotide sequence, and perhaps on se- condary structure.
‘Slippage’ Reaction and Misincorporation using MS2 RNA-poly(A) or Poly(A) as Template
Analysis by gel electrophoresis of the poly(dT) products, synthesized on poly(A) or on MS2 RNA- poly(A) using [ M - ~ ~ P I ~ T T P , present at a concentration of 10 pM, is shown in Fig. 13. It was found that a distinct band is present in appreciably greater amount than the higher-molecular-weight bands. The size of this product corresponds to a length of 14 nucleotides. Various control experiments were carried out to show that this band was not an artefact. Also, addition of either dGTP or of the three remaining dNTPs to the reaction mixture for transciption of MS2 RNA-poly(A)
Fig. 13. Characterization of the initial oligo(dT) products synthesized usingpoly(A) or MS2 RNA-poly(A). (A) A 20-p1 reaction mixture containing 1 pg poly(A), 0.4 unit AMV reverse transcriptase and [z-~’P]~TTP at 10 pM (100 Ci/mmol) was incubated for 60 min at 25 “C. The reaction was stopped with EDTA and the mixture was divided in two. Solid urea, 40 pg tRNA and sucrose were added, and the mixtures were electrophoresed with (slot 1) or without (slot 2) tracking dyes (see Materials and Methods) in a 24% polyacrylamide gel. (B) 10-p1 reaction mixtures containing 0.5 pg MS2 RNA-poly(A), 0.2 unit AMV reverse transcriptase and [w3’P]dTTP with or without other unlabeled dNTPs were incubated at 25 “C and electrophoresed as described in (A). Slot 1, 2 and 3: (dT) product synthesized in the presence of [ G ( - ~ ~ P ] ~ T T P at 10 pM (100 Ci/mmol) alone, or with dGTP (100 pM) or with dGTP, dATP and dCTP (100 pM each). (C) Reaction conditions as in (B). Slot 1 : 32P-labeled product synthesized in the presence of [w3’P]dATP at 10 pM (50 Ci/mmol); slot 2: 32P-labeled product synthesized in the presence of [a-32P]dCTP at 10 FM (50 Ci/mmol); slot 3 : 32P- labeled product synthesized in the presence of [w3’P]dTTP at 0.2 pM (100 Ci/mmol); slot 4: [5’-32P](dT),l previously purified on a 24% polyacrylamide gel; slot 5: [5’-32P](dT)l, previously purified on a 24% polyacrylamide gel. The insert is a longer auto- radiographic exposure of this relevant part of the gel (dashed rectangle).
did not markedly change the characteristic labeling pattern. Although the exact origin of this particular product remains unclear, these results possibly point

430 Discrete Reverse Transcription Products of MS 2 RNA-poly(A)
Table 2. Eflect of ( A ) primer and ( B ) nucleoside triphosphate on the reverse transcription of poly(A) and MS2 RNA-poly(A) (A) 1 pg poly(A) or 1 pg MS2 RNA-poly(A) samples were prein- cubated in 0.5 M KCI for 15 min at 37 "C with either 1 pg (dT),,, 1 pg (U), or 1 pg poly(U) (pretreated for 1 h at 37 "C with alkaline phosphatase and purified on a Sephadex G-50 column). The template- primer complex was then added to a 5O-pl reaction mixture, con- taining [3H]dTTP at 30 pM (4 Ci/mmol) as the only deoxynucleoside triphosphate present, 0.2 unit AMV reverse transcriptase and either 8 mM MgCI, or 0.5 mM MnCI, as indicated. After 60-min incubation at 37 "C acid-precipitable radioactivity was measured. (B) 1 pg MS2 RNA-poly(A) was preincubated for 15 min at 37 "C with 1 pg (dT),,, and then added to a reaction mixture (final volume 30 pl) containing either [3H]dCTP at 65 pM (23 Ci/mmol) in the presence of 8 mM MgCI,, or [3H]CTP at 88 pM (17 Ci/mmol) in the presence of 8 mM MgCI, or 2 mM MnCI,, and dATP, dGTP and dTTP at 100 pM each. The reaction was started by addition of 0.2 unit AMV DNA polymerase and incubated for 60 min at 37 "C
(A) dTMP incorporated with Primer Template
8 mM MgCI, 0.5 mM MnCI,
pmol/O.l ml
(B) Nucleoside triphosphate dCMP or CMP
incorporated
[3H]dCTP, 8 mM MgC1, [3H]CTP, 8 mM MgC1, [3H]CTP, 2 mM MnCl,
pmol/O.l ml
135 5.9 5.2
towards a special characteristic of the initiation reaction by the AMV reverse transcriptase, starting from poly(A). oligo(dT). The use of [ u - ~ ~ P I ~ T T P at 0.2 pM leads almost exclusively to the formation of a single product (Fig. 13C), while with [x-~'P]~ATP or [ u - ~ ~ P I ~ C T P a labeled product is also formed al- though in much smaller amounts than with dTTP. That the AMV DNA polymerase can elongate the primer with a non-complementary nucleotide on MS2 RNA-poly(A) has been confirmed by snake venom degradation of the 32P-labeled product (not shown). This result may be due to misincorporation of the type found by Battula et al. [6]. However, it is not known yet, whether the reverse transcriptase does in- corporate a non-complementary nucleotide in the MS2 cDNA copy when all four deoxynucleotide triphosphates are present.
Use of a Ribosyl Primer and Ribonucleoside Triphosphates
The natural primer in oncorna viruses is a tRNA [20] ; therefore one would expect that oligoribonucleo- tides can also be used as primers for the synthesis of discrete products. This was tested by using poly(A) as a template with either (U), or poly(U) (treated with alkaline phosphatase) as a primer, but no [3H]dTMP incorporation could be detected. Similarly, using MS2 RNA-poly(A), only a little elongation of the (U), or poly(U) primer was observed with or without MnC1, present in the reaction (Table 2A). In order to further investigate whether oligo(1) would serve as a primer for AMV reverse transcriptase, an oligo(C) tail was added to the 3' end of MS2 RNA. MS2 RNA-oligo(C) was synthesized using ATP : RNA adenylyltransferase' , under somewhat different conditions as used for adenylation (pH 8.4, 8 mM MgCl,, 7.5 mM MnCl, and 4 mM dithiothreitol). The cytidylyltransfer activity thus obtained was about one third of that found pre- viously for adenylyltransfer [7]. Fig. 14(A) shows the purification of MS2 RNA-oligo(C) on a sucrose gradient. Using (dG),, as a primer, it is evident from Fig. 14(B) that MS2 RNA-oligo(C) is an efficient template for reverse transcription. On the other hand, using oligo(1) as a primer, no significant synthesis on MS2 RNA-oligo(C) could be detected. MS2 RNA- oligo(C) can be used as a template for QP replicase, which in the presence of rGTP synthesizes an oligo(G) complementary to the 3' oligo(C) tail; the resulting complex, however, could not be used as a template- primer for the AMV reverse transcriptase (data not shown). These results on the absence of priming ability of oligoribonucleotides are somewhat unex- pected in view of the fact that the natural primer is a tRNA molecule. This could perhaps be due to loss of a factor or to conversion of the p form of the enzyme to the CI form, or to a different conformation of the template-primer complex.
We also tested whether ribonucleoside triphos- phates could be incorporated by the AMV reverse transcriptase using MS2 RNA-poly(A) . (dT),, as a template. The results shown in Table 2(B) indicate that, even in the presence of MnCl, , maximal [3H]CMP incorporation is only one thirtieth of the incorporation observed with [3H]dCTP.
DISCUSSION
The addition of a poly(A) tail to an RNA offers the possibility of transcribing this RNA with RNA- dependent DNA polymerase, using oligo(dT) as a primer. Since it has been shown that the AMV reverse
In view of this broader specificity, the enzyme may perhaps more properly be named NTP : RNA nucleotidylyltransferase.

R. Devos, J. van Emmelo, P. Celen, E. Gillis, and W. Fiers 431
t OP bottom
Fraction number Incubation time (rnin)
Fig. 14. Purijicution of M S 2 RNA-oligoiCJ by zonul centrifugation und reverse trunscriptase template ej’ciency using (dG), , as a primer. (A) A I-ml reaction mixture containing 50 mM Tris-HC1 (pH 8.4), 8 mM MgCl,, 7.5 mM MnCl,, 4 mM dithiothreitol, 0.5 mM EDTA, 0.15 M NaC1, 0.3 mM [3H]CTP (50 Ci/mol), 50 pg MS2 RNA and 20 pg ATP: RNA adenylyltransferase was incubated for 15 min at 37 “C. The reaction was stopped by addition of 10 p1 20% sodium dodecylsulphate and 10 mg solid EDTA, and the solution was further treated with a phenol/chloroform/isoamylalcohol mixture as described in Materials and Methods. The RNA was recovered by ethanol precipitation and analysed on a 10-30% sucrose gradient. AZ54 (-); acid-precipitable radioactivity (- ---). (B) 1 pg MS2 RNA-oligo(C) from the absorbance peak fraction of the sucrose gradient was preincubated for 35 min at 37 “C in the presence of either (a) 300 pmol (dG),, or (b) 1 pg oligo(1). Subsequently, the incubation was continued in a 2 0 4 reaction mixture (cf. Materials and Methods), containing 0.2 unit AMV reverse transcriptase, 10 pM [a-jZP]dTTP (75 Cijmmol) and 100 pM of unlabeled dNTPs. After different times 2 4 samples were withdrawn for acid-precipitable radioactivity measurements
transcriptase can generate incomplete, discrete trans- cripts, new methods may be devised on this basis for the nucleotide sequence determination of the 3‘ end of an RNA. Many studies have been devoted to an optimalization of the experimental conditions for the reverse transcriptase reaction using synthetic oligo- ribonucleotides and polyribonucleotides as primer- template combinations [21]. Also procedures for ob- taining complete copies of natural mRNAs have been much improved [ l , 31. AMV reverse transcriptase, however, ‘slips’ during transcription of poly(A) [17] (and this paper) and it may be important to eliminate this side reaction when transcribing natural mRNAs.
Our results indicate that natural RNA molecules polyadenylated in vitro using ATP :RNA adenylyl- transferase, become efficient templates for reverse transcription. However, as observed for globin mRNA, poly(dT) products were formed, prior to the extension of the primer beyond the poly(A) tail. Addition of only the first deoxynucleoside triphosphate comple- mentary to the 3‘ end of the RNA, together with dTTP, reduces this poly(dT)-synthesizing activity, and seems to depress slippage. By reducing the dTTP con- centration we could further eliminate synthesis of poly(dT) on the poly(A) tail. Presumably when dTTP is present in low concentration, the enzyme pre- ferentially starts elongation of those primer molecules, which are hybridized adjacent to the original 3’ end of the RNA. This contributes to the homogeneity of the product. On the other hand, lowering the dTTP
concentration generates discrete, partial reverse trans- cripts. This could also be accomplished by lowering the concentration of any other deoxynucleoside tri- phosphate other than dTTP. Since the incomplete transcripts occur as discrete size classes, rather than as a continuous spectrum, it is of interest to look where exactly these products terminate on the RNA template. Efstratiadis et al. [ l ] proposed that incomplete trans- cripts may be generated by the existence of regions with extensive secondary structure in the RNA tem- plate. We did observe that by increasing the incubation temperature larger transcripts were obtained. How- ever, the specific banding pattern corresponding to the size of the discrete products remained qualitatively the same. This means that none of the bands could be correlated with the specific destabilization of certain structural features upon increasing the temperature. On the other hand we did find that the nature of the limiting deoxynucleoside triphosphate profoundly af- fects the qualitative pattern of the incomplete trans- cripts. Therefore particular nucleotide sequences must play an important role in blocking the reverse trans- criptase at specific positions. For example, since poly(U) and poly(G) were transcribed at very low rates by AMV reverse transcriptase [17], it is possible that oligo(U) or oligo(G) stretches in the template are responsible for slowing down transcription, when dATP or dCTP respectively are present in limiting amounts. This is consistent with our observation that lowering the concentration of dATP or dCTP in the

432 R. Devos, J. van Emmelo, P. Celen, E. Gillis, and W. Fiers: Discrete Reverse Transcription Products of MS2 RNA-poly(A)
reaction was more effective for decreasing the total activity of the enzyme, than lowering the concentration of dTTP or dGTP respectively.
It has been shown that when the AMV reverse transcriptase reaction is run in the presence of three or less triphosphates, synthesis proceeds up to a posi- tion where the missing triphosphate should have been incorporated. Polyadenylated MS2 or QB RNAs and a combination of two or three deoxynucleoside tri- phosphates, present in limiting amounts, leads to several discrete products, differing only by a few residues in length. The main bands completely cor- respond to what one would expect on the basis of the primary structure. However, even under these con- ditions intermediate bands are also present and these can either be explained by blocking of the AMV poly- merase due to a double-stranded region with high stability in the template, or by the difficulty which the enzyme experiences when it has to incorporate several residues in a row of a precursor present in limiting concentration. (The latter hypothesis would involve at least two precursor binding sites on the enzyme, a catalytic site and an entry site; reversible binding at the catalytic site would only lead to incorporation when the entry site is also occupied by a matching precursor.)
The series of discrete products with a homogeneous 5’ end and increasing size offers obvious advantages for the nucleotide sequence determination of the 3’ end of any RNA. This application will be further docu- mented in a forthcoming paper.
We thank Dr R. Contreras and Dr M. Ysebaert for preparing [5’-”P](dT),,, Dr J. Merregaert and Mr F. Molemans for pyrimi- dine tract analysis, and Dr J. Beard and Dr M. Chirigos for a generous supply of AMV reverse transcriptase (Office of Program and Logistics, Viral Cancer Program, Viral Oncology, Division of Cancer Cause and Prevention, Bethesda, Maryland, 20014) This research program was supported by a grant from the Belgian Fonds voor Kollektief Fundamenteel Onderzoek (1 0300).
REFERENCES
1.
2.
3. 4.
5.
6.
7.
8.
9.
10. 11.
12.
13.
14.
15. 16.
17.
18.
19.
20.
21.
Efstratiadis, A., Maniatis, T., Kafatos, F. C., Jeffrey, A. &
Monahan, J., Harris, S., Woo, S. & O’Malley, B. (1976) Bio-
Molling, K. (1976) Science (Wash. D . C . ) 264, 111-112. Haseltine, W. A,, Kleid, D. G., Panet, A., Rothenberg, E. &
Imaizumi, T., Diggelmann, H. & Schemer, K. (1975) Proc.
Battula, N., Dube, D. K. & Loeb, L. A. (1975) J . Biol. Chem.
Devos, R., Gillis, E. & Fiers, W. (1976) Eur. J . Biochem. 62,
Devos, R., Van Emmelo, J., Seurinck-Opsomer, C., Gillis, E.
Vournakis, J. W. (1975) Cell, 4, 367-378.
chemistry, 16, 223 -233.
Baltimore, D. (1976) J . Mol. Biol. 106, 109-131.
Nut1 Acad. Sci. U.S.A. 70, 1122-1126.
250, 8404 - 8408.
401 -410.
& Fiers, W. (1976) Biochirn. Biophys. Acta, 447, 319- 321.
Fiers, W., Contreras, R.; Duerinck, F., Haegeman, G., Iseren- tant, D., Merregaert, J., Min Jou, W., Molemans, F., Raey- maekers, A., Van den Berghe, A., Volckaert, G. & Ysebaert, M. (1976) Nature (Lond.) 260, 500-507.
Sippel, A. E. (1973) Eur. J . Biochem. 37, 31 -40. Kacian, D. L. & Spiegelman, S. (1974) Method Enzymol. 29,
Maniatis, T . , Jeffrey, A. & van de Sande, H. (1975) Bioche- mistry, 14, 3787 - 3794.
De Wachter, R. & Fiers, W. (1971) Methods Enzymol. 21, 167-178.
Falvey, A. K., Kantor, J. A., Robert-Guroff, M. G., Picciano, D. J., Weiss, G. B., Vavich, J. M. & Anderson, W. F. (1974) J . Biol. Chem. 249, 7049-7056.
De Wachter, R. & Fiers, W. (1967) J . Mol. B i d . 30, 507-527. Vandenberghe, A., Min Jou, W. & Fiers, W. (1975) Proc. Natl
Acad. Sci. U.S .A. 72, 2559 -- 2562. Falvey, A. K., Weiss, G. B., Krueger, L. J., Kantor, J.A. &
Anderson, W. F. (1976) Nucleic Acid Res. 3, 79-88. Weissmann, C., Billeter, M. A,, Goodman, H. M., Hindley, J.
& Weber, H. (1973) Annu. Rev. Biochem. 42, 303 - 328. Flavell, R. A,, Sabo, D. L. O., Bandle, E. F. & Weissmann, C.
(1975) Proc. Nut1 Acad. Sci. U.S.A. 72, 367-371. Dahlberg, J. E., Sawyer, R. C., Taylor, J. M., Faras, A. J., Le-
vinson, W. E., Goodman, H. M. & Bishop, J. M. (1974) J . Virol. 13, 1126-1133.
Marcus, S . L. & Modak, J. (1976) Nucleic Acid Res. 3 , 1473- 1486.
150- 173.
R. Devos, J . van Emmelo, and W. Fiers, Laboratorium voor Moleculaire Biologie, Fakulteit der Wetenschappen, Rijksuniversiteit te Gent, K. Lod Ledeganckstraat 35, B-9000 Gent, Belgium
P. Celen and E. Gillis, Laboratorium voor Fysiologische Scheikunde, Fakulteit voor Geneeskunde, Rijksuniversiteit te Gent, K. Lod Ledeganckstraat 35, B-9000 Gent, Belgium


![RT-PCR [Uyumluluk Modu] - turkpath.org.tr · Doğal revers transkriptaz örnekleri: HIV-1 (human immunodeficiency virus) M-MLV (Moloney murine leukemia virus) AMV ( avian myeloblastosis](https://img.pdfslide.net/doc/110x75/5ccf047188c993fb7c8df0ea/rt-pcr-uyumluluk-modu-dogal-revers-transkriptaz-oernekleri-hiv-1-human.jpg)