Embed Size (px)
Citation preview

IntroductionSexual dimorphism is common in spiders and most dif-ferences between males and females are attributed either todifferences in reproductive roles or sexual selection.Differences in reproductive roles may account for size dif-ferences between males and females (e.g. Head 1995; Prenteret al. 1999; but see also Elgar 1991, Vollrath and Parker 1992,Moya-Laraño et al. 2002) and differences in trophic mor-phology, i.e. the sizes of chelicerae and venom glands(Walker and Rypstra 2001, 2002). In contrast, sexual selec-tion is thought to have caused comparatively longer legs inmales than females, particularly in species with sedentaryfemales (Gasnier et al. 2002; Framenau 2005), and it hasbeen argued that dimorphic colour patterns augment bodysize and condition both in male and female wolf spiders,resulting in increased mating success (e.g. Moya-Larañoet al. 2003). Sexual selection also explains the exaggerationof the visual courtship display of male wolf spiders throughmorphological and colour modifications of the pedipalps orsegments of the first leg (e.g. Barthel and Helversen 1990;Miller et al. 1998; Hebets and Uetz 2000). A most peculiardimorphic structure relates to the acoustic courtship displayof males of Hygrolycosa rubrofasciata Ohlert, 1865, Pardosa
sphagnicola (Dahl, 1908) and P. hyperborea (Thorell, 1872)(Kronestedt 1996 and references therein). Males of thesespecies display a ritualised drumming or tapping behaviourby flexing their abdomen onto the substrate to produce a dis-tinct sound or seismic signal. Knobby setae with a hirsutesurface on the ventral side of the abdomen in males arethought to improve the acoustic display of males in H. rubro-fasciata (e.g. Kronestedt 1996; Kotiaho et al. 1996; Mappeset al. 1996). Similar knobby setae have recently been foundin Passiena torbjoerni Lehtinen, 2005 (Lehtinen 2005), butthe courtship behaviour of this species is unknown and therole of the setae in acoustic signalling is untested.
The Australasian wolf spider fauna has received consider-able attention in recent years, with the revision of severalgenera, many of which are only known from theOriental/Pacific region (e.g. Vink 2001; Framenau and Vink2001; Framenau 2002, in press a, in press b). Three sub-families are present in Australia (Murphy et al. 2006;V. W. Framenau, unpublished data): the Venoniinae sensuLehtinen and Hippa (1979), including genera such asVenonia Thorell, 1894, Allotrochosina Roewer, 1960 andAnomalosa Roewer, 1960; an undescribed Australasian/Pacific subfamily represented by Artoria Thorell, 1877,
Invertebrate Systematics, 2006, 20, 185–202
10.1071/IS05036 1445-5226/06/020185© CSIRO 2006
Volker W. FramenauA,D and Jung-Sun YooA,B,C
ADepartment of Terrestrial Invertebrates, Western Australian Museum, Locked Bag 49, Welshpool DC, Western Australia 6986, Australia.
BSchool of Animal Biology, University of Western Australia, Crawley, Western Australia 6009, Australia.CResearch Institute for Natural Science, Dongguk University, Seoul 100-715, Korea.
DCorresponding author. Email: [email protected]
Abstract. The new Australian wolf spider genus Tuberculosa is revised, with T. harveyi, sp. nov. from the NorthernTerritory as type species. The genus includes a further three species from northern Queensland: T. austini, sp. nov.,T. hoggi (Framenau & Vink, 2001), comb. nov. and T. monteithi, sp. nov. The genus is defined by a unique sexualdimorphism: males carry modified tubercular setae on the ventral side of the third coxae, which are here comparedto the knobbed setae that can be found on the ventral surface of the abdomen in Hygrolycosa rubrofasciata Ohlert,1865 and Passiena torbjoerni Lehtinen, 2005 from two different lycosid subfamilies. Based on the structure of themale pedipalp, Tuberculosa belongs to the subfamily Lycosinae sensu Dondale (1986) with close affinities toVenatrix Roewer, 1960, because males of both genera have a tubercle on the outer edges of their fangs and thecymbium tip carries claw-like macrosetae. A cladistic analysis of all four Tuberculosa species, based on12 morphological characters and with Venatrix konei (Berland, 1924) as outgroup, revealed a topology withT. austini and T. harveyi as sister-species in the most derived clade (V. konei (T. monteithi (T. hoggi (T. harveyi,T. austini))). The distribution of Tuberculosa in the tropical north of Australia supports an origin of the genus, aswell as its putative sister-genus Venatrix, in the Palaearctic region, in contrast to all other Australian Lycosinae,which appear to be of Gondwanan origin.
Systematics of the new Australian wolf spider genus Tuberculosa(Araneae:Lycosidae)
www.publish.csiro.au/journals/is
CSIRO PUBLISHING

V. W. Framenau and J.-S. Yoo186 Invertebrate Systematics
Anoteropsis L. Koch, 1878, Tetralycosa Roewer, 1960,Diahogna Roewer, 1960 and several undescribed genera; andthe Lycosinae sensu Dondale (1986), represented by generasuch as Dingosa Roewer, 1955, Mainosa (Framenau, in pressa), Tasmanicosa Roewer, 1959, Venatrix Roewer, 1960 andVenator Hogg, 1900. The latter subfamily dominates theAustralian wolf spider fauna both in species richness andlocal abundance, but the generic relationships of manyAustralasian species remain unresolved. A recent molecularstudy suggested numerous wrong generic placements, in par-ticular of Australian species (Murphy et al. 2006). This studyalso showed that the Australian Lycosinae belong to two dif-ferent major clades. One of these is currently representedonly by the genus Venatrix, and morphological evidence sug-gests a phylogenetic relationship with the northern hemi-sphere genera Alopecosa Simon, 1885 and TrochosaC. L. Koch, 1847. The second, much larger Australasianclade includes all other Australian Lycosinae that were rep-resented in the analysis. The inclusion of the South AmericanPavocosa Roewer, 1960 in this clade suggests a Gondwananorigin of this group (Murphy et al. 2006).
We here report on a new lycosine genus with a veryunusual sexual dimorphism not previously reported in wolfspiders. The ventral side of the third coxae in males has afield of 9–15 modified tubercular setae (Fig. 1A). Thesesetae were first recorded for Venatrix hoggi Framenau &Vink, 2001. This species was initially included in the wolfspider genus Venatrix Roewer, 1960 owing to the presence ofa tubercle on the outer edge of the fangs in males and claw-like macrosetae on the tip of the cymbium (Framenau andVink 2001). However, the authors noted the unusual pedipalpconfiguration with a reduced terminal apophysis. Theyrefrained from erecting a new genus because of the lack ofmaterial (V. hoggi was then only known from two males)(Framenau and Vink 2001). Recent investigations of thelycosid collections from tropical northern Australia have
revealed three more species with the same conspicuous coxalsetae. These four species are revised here and a phylogenetichypothesis is presented based on a cladistic analysis of12 morphological characters, with Venatrix konei (Berland,1924) as an outgroup.
Material and methods
This revision forms part of a revision of Australian wolf spiders and isbased on the examination of all available type material of Australianlycosids and more than 20000 databased records of wolf spiders in allmajor museum collections in Australia.
Descriptions are based on specimens preserved in 70% ethanol.Internal female genitalia were prepared for examination by submersionin lactic acid at room temperature for 2–24 h. For clarity, the setae havebeen omitted from the illustrations of epigyna and male pedipalps.Scanning electron microscope (SEM) images were taken with a CarlZeiss Leo 1420VP (Oberkochen, Germany) after fixing the respectivespider parts in Karnovsky’s fixative (Sheehan and Hrapchack 1980) andosmic acid. The morphological nomenclature follows Framenau andVink (2001). All measurements are in millimetres (mm).
Phylogenetic analysis
We developed a phylogenetic hypothesis for the species of Tuberculosausing a cladistic analysis of morphological characters. We chose arepresentative of Venatrix with morphological affinities to Tuberculosaas an outgroup (Watrous and Wheeler 1981; Maddison et al. 1984, seealso Nixon and Carpenter 1993) since Venatrix is the putative sister-genus of Tuberculosa. The character matrix used in the cladistic analysiswas compiled in NEXUS Data Editor Version 0.5.0 (Page 2001). Allcharacters were binary and treated as unordered. As T. monteithi is onlyknown from males, female characters were excluded as missing data(‘?’). A heuristic search for maximum parsimony was performed withNONA version 2.0 (Goloboff 1993) within WinClada version 1.00.08(Nixon 2002) using the following commands: 1000 trees to hold (‘hold’)with 1000 replications (‘mult*N’) and 10 starting trees per replication(‘hold/’). Character optimisation was mapped onto the resulting topol-ogy for unambiguous characters only, and for fast (ACCTRAN) andslow (DELTRAN) optimisation. Twelve morphological characters, ofwhich three were somatic, six derived from male genitalia and threefrom female genitalia, remained parsimony informative for phylogeneticanalysis after eliminating autapomorphic characters a priori (Tables 1
Table 1. Characters and character states for the cladistic analysis of TuberculosaAll character states are considered unordered for the cladistic analysis in NONA
Somatic characters
(1) Carapace, median light band, shape: 0, strongly constricted in front of fovea; 1, slightly or not constricted in front of fovea.(2) Femora, dark annulations: 0, absent; 1, present.(3) Tibia I, number of retrolateral spines in males: 0, no spine; 1, one spine.
Genital characters, male(4) Palea, shape and sclerotisation: 0, apically bulging, not sclerotised; 1, apically flat, sclerotised.(5) Embolus, length: 0, approximately halfway along palea; 1, about as long as palea.(6) Embolus, concealment in ventral view: 0, exposed; 1, concealed.(7) Embolus, origin of sperm duct in ventral view: 0, central; 1, prolateral.(8) Terminal apophysis, texture: 0, not sclerotised; 1, sclerotised.(9) Terminal apophysis, shape: 0, lobe or tip; 1, concave, shape like a trough.
Genital characters, female(10) Epigyne, sclerotised lateral margins: 0, absent; 1, present.(11) Epigyne, angle of opening of anterior hoods: 0, ~45° to median line; 1, perpendicular to median line.(12) Copulatory duct: 0, narrow; 1, wide.

Invertebrate Systematics 187
and 2). Bremer ( = decay index) (Bremer 1988, 1994) and relativeBremer support (Goloboff 1993) were calculated using the followingcommands in NONA: ‘hold 1000 ; sub 1 ; find *; hold 2000 ; sub 3 ; find*; hold 4000 ; sub 5 ; find *; bsupport ; bsupport*;’.
Character coding was based on the specimens that were here usedfor species description, i.e. the holotype and paratypes in the cases ofTuberculosa austini, sp. nov., T. harveyi, sp. nov. and T. monteithi,sp. nov., and non-type material of T. hoggi (Framenau and Vink 2001).Outgroup comparison is based on a male (WAM T56058, fromSpecimen Hill Bushland Reserve, Bendigo Regional Park) and a female(WAM T56075, from Lake Eppalock) of Venatrix konei from Victoriacollected by the senior author. A detailed description of V. konei (asV. goyderi Hickman, 1944; new synonymy in Framenau (in press b)) canbe found in Framenau and Vink (2001).
Somatic characters
The four species of Tuberculosa do not vary noticeably in their somaticmorphology. The first character coded the presence and absence of aconstriction of the light median band in front of the fovea on the cara-pace (for example it is not constricted in T. harveyi, Fig. 2A). There wasalso some variation in the colouration of the femora, which showedsome dark annulations in two species, T. monteithei and V. konei (char-acter 2). Leg spination in wolf spiders varies considerably, evenbetween two opposite legs of a single specimen; however, we detectedconsistent patterns in the number of retrolateral spines on tibia I inmales (character 3).
Male genital characters
The palea region in Tuberculosa can either be bulging ventrally inits apical half, possibly a result of a lack of sclerotisation (Figs 2D, 4C,8C), or it may have a flat, sclerotised area that does not allow bulgingas indicated for T. hoggi (Fig. 6C) (character 4). The position and shapeof the embolus allowed the coding of two characters: character 5, therewas some variation in the length of the embolus in relation to the palea,i.e. it was either half as long as the palea (e.g. Figs 2D, 4C) or nearly aslong as the palea (e.g. Figs 6C, 8C); character 6, the embolus was eithertotally concealed by the tegulum and median apophysis in ventral view(e.g. Fig. 2E) or totally exposed (e.g. Fig. 6A). Character 6 related to theoverall shape of the bulb, in particular the relative size of the palea andtegulum. The sperm duct penetrates the palea dorsally and the positionwhere it becomes visible on the ventral side is central on the palea (e.g.T. hoggi, Fig. 6C) or prolateral (e.g. T. harveyi, Fig. 2D) (character 7).
The shape of the terminal apophysis was coded using two charac-ters: character 8, the terminal apophysis was either heavily sclerotised(T. harveyi (Figs 2D, E), T. austini (Figs 4A, C) and V. konei), or weaklysclerotised (T. monteithi, Figs 8A, C), or not sclerotised at all (T. hoggi,Figs 6A, C); character 9, the terminal apophysis was either a simple lobeor tip (Figs 6A, C, 8A, C) or concave to form the shape of a trough(Figs 2D, 4C).
Female genital characters
Tuberculosa harveyi and T. austini do not have a sclerotised lateralmargin bordering the epigyne atrium (Fig. 2G, 4D), in contrast toT. hoggi in which these margins reach halfway from the anterior cornerstowards the epigastric furrow (Fig. 6E) (character 10). In V. konei, thesemargins are very distinct along the whole length of the epigyne(e.g. Framenau and Vink 2001). The opening of the anterior hoodsnearly formed a right angle with a median line through the epigyne(Fig. 4D, 6D), or this angle was ~45° (Fig. 2G) (character 11). The inter-nal genitalia of Tuberculosa species are very similar, but there is somevariation in the width of the copulatory duct, which is narrow in T. hoggi(Fig. 6E) and V. konei, but much wider in the other species (Figs 2H, 4E)(character 12). The female of T. monteithi is not known and its futurediscovery may provide additional informative characters for the phylo-genetic analysis of Tuberculosa.
Abbreviations
Measurements (adult spiders, if not otherwise stated)
TL total lengthCL carapace length
CW carapace widthAL abdomen lengthAW abdomen width
Eyes
AE anteriorAME anterior medianALE anterior lateral
PE posteriorPME posterior medianPLE posterior lateral
Collections
NTMAG Museum and Art Gallery of the Northern Territory,Darwin
QM Queensland Museum, BrisbaneWAM Western Australian Museum, Perth
Key to the species of Tuberculosa
1. Light median band on carapace limited to anterior half, terminalapophysis forms straight sclerotised tip (Fig. 8C); (femaleunknown) . . . . . . . . . . . . . . . . . . . . . . T. monteithei, sp. nov.
Light median band on carapace reaches to posterior margin ofcarapace, terminal apophysis formed like a trough (Figs 2D,4C) or reduced to a lobe (Fig. 6C) (all known from females). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. Terminal apophysis reduced (Fig. 6C); sclerotised margin ofepigyne extends nearly halfway along the lateral border(Fig. 6D). . . . . . . . . . . . . T. hoggi (Framenau & Vink, 2001)
Terminal apophysis of male pedipalp formed like a trough(Figs 2D, 4C); female epigyne without sclerotised lateralmargins (Figs 2G, 4D) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3. Ventral apical edge of terminal apophysis of male pedipalpalmost straight (Fig. 2D); anterior hoods of epigyne almosttouch posterior margin medially (Fig. 2G) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. harveyi, sp. nov.
Ventral apical edge of terminal apophysis of male pedipalpcurved (Fig. 4C); median connection of anterior hood withconsiderable distance from posterior margin (Fig. 4D) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. austini, sp. nov.
Systematics of the new Australian wolf spider genus Tuberculosa
Table 2. Character matrix for cladistic analysisFor a description of characters and character states see Table 1
Taxa CharactersSomatic Male Female
genitalia genitalia
1 2 3 4 5 6 7 8 9 10 11 12V. konei 1 1 1 1 1 0 1 1 0 1 0 0T. austini 0 0 0 0 0 1 0 1 1 0 1 1T. harveyi 0 0 0 0 0 1 1 1 1 0 0 1T. hoggi 1 0 0 1 1 0 0 0 0 1 1 0T. monteithi 1 1 1 0 1 0 0 0 0 ? ? ?
V. W. Framenau and J.-S. Yoo188 Invertebrate Systematics
Systematics
Family LYCOSIDAE Sundevall
Subfamily LYCOSINAE Sundevall
Genus Tuberculosa, gen. nov.
Type species: Tuberculosa harveyi, sp. nov., here designated.
Diagnosis
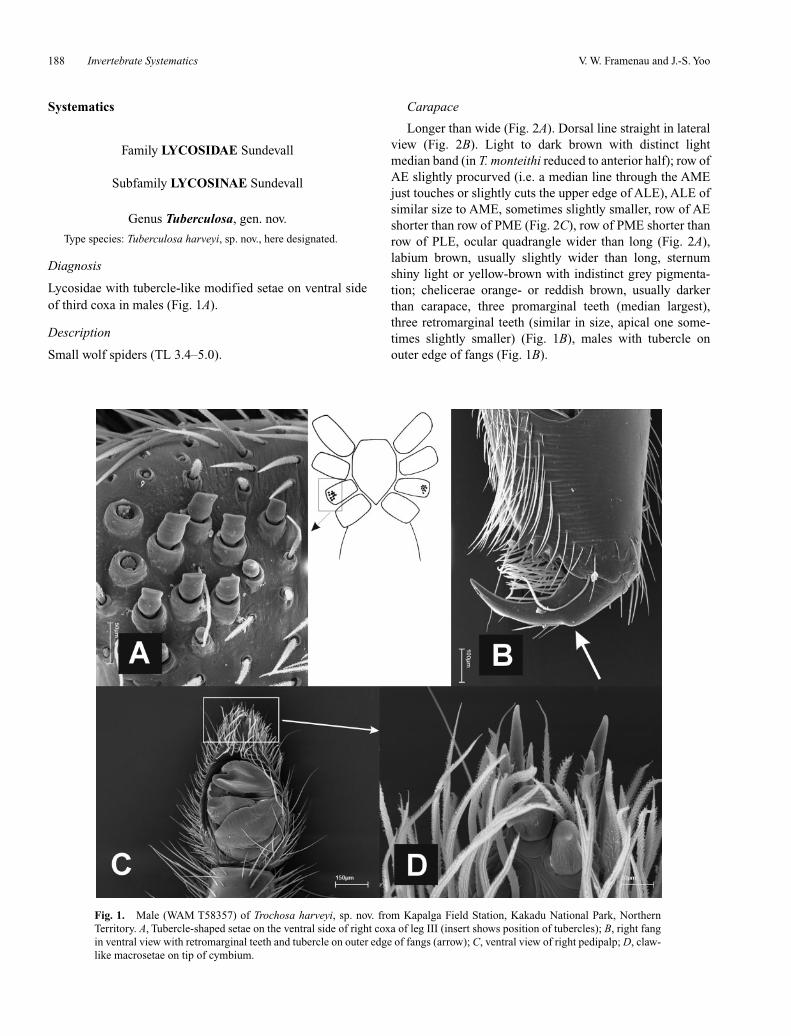
Lycosidae with tubercle-like modified setae on ventral sideof third coxa in males (Fig. 1A).
Description
Small wolf spiders (TL 3.4–5.0).
Carapace
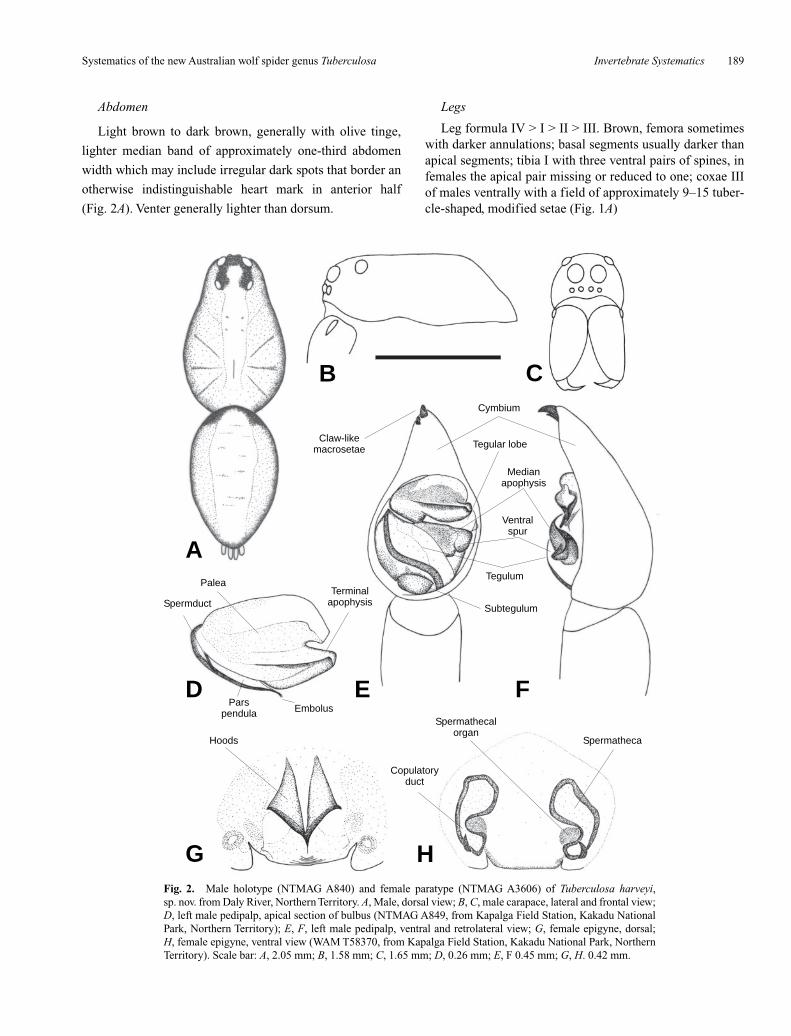
Longer than wide (Fig. 2A). Dorsal line straight in lateralview (Fig. 2B). Light to dark brown with distinct lightmedian band (in T. monteithi reduced to anterior half); row ofAE slightly procurved (i.e. a median line through the AMEjust touches or slightly cuts the upper edge of ALE), ALE ofsimilar size to AME, sometimes slightly smaller, row of AEshorter than row of PME (Fig. 2C), row of PME shorter thanrow of PLE, ocular quadrangle wider than long (Fig. 2A),labium brown, usually slightly wider than long, sternumshiny light or yellow-brown with indistinct grey pigmenta-tion; chelicerae orange- or reddish brown, usually darkerthan carapace, three promarginal teeth (median largest),three retromarginal teeth (similar in size, apical one some-times slightly smaller) (Fig. 1B), males with tubercle onouter edge of fangs (Fig. 1B).
Fig. 1. Male (WAM T58357) of Trochosa harveyi, sp. nov. from Kapalga Field Station, Kakadu National Park, NorthernTerritory. A, Tubercle-shaped setae on the ventral side of right coxa of leg III (insert shows position of tubercles); B, right fangin ventral view with retromarginal teeth and tubercle on outer edge of fangs (arrow); C, ventral view of right pedipalp; D, claw-like macrosetae on tip of cymbium.
Invertebrate Systematics 189
Abdomen
Light brown to dark brown, generally with olive tinge,
lighter median band of approximately one-third abdomen
width which may include irregular dark spots that border an
otherwise indistinguishable heart mark in anterior half
(Fig. 2A). Venter generally lighter than dorsum.
Legs
Leg formula IV > I > II > III. Brown, femora sometimeswith darker annulations; basal segments usually darker thanapical segments; tibia I with three ventral pairs of spines, infemales the apical pair missing or reduced to one; coxae IIIof males ventrally with a field of approximately 9–15 tuber-cle-shaped, modified setae (Fig. 1A)
Systematics of the new Australian wolf spider genus Tuberculosa
A
B C
D E F
G H
Cymbium
Medianapophysis
Terminalapophysis
Embolus
Tegulum
Subtegulum
Copulatoryduct
Spermatheca
Spermathecalorgan
Hoods
Palea
Spermduct
Claw-likemacrosetae Tegular lobe
Parspendula
Ventralspur
Fig. 2. Male holotype (NTMAG A840) and female paratype (NTMAG A3606) of Tuberculosa harveyi,sp. nov. from Daly River, Northern Territory. A, Male, dorsal view; B, C, male carapace, lateral and frontal view;D, left male pedipalp, apical section of bulbus (NTMAG A849, from Kapalga Field Station, Kakadu NationalPark, Northern Territory); E, F, left male pedipalp, ventral and retrolateral view; G, female epigyne, dorsal;H, female epigyne, ventral view (WAM T58370, from Kapalga Field Station, Kakadu National Park, NorthernTerritory). Scale bar: A, 2.05 mm; B, 1.58 mm; C, 1.65 mm; D, 0.26 mm; E, F 0.45 mm; G, H. 0.42 mm.

V. W. Framenau and J.-S. Yoo190 Invertebrate Systematics
Male pedipalp
Cymbium tip with two distinct claw-like setae (Fig. 1D),sometimes the smaller prolateral one missing; medianapophysis directed retrolaterally, ventral spur present; termi-nal apophysis of varying shape; embolus generally with ashort free tip, sometimes totally fused to palea; subtegulumoval and basal of tegulum.
Epigyne
Two distinct anterior hoods; median septum absent;spermathecae large and elongated, copulatory ducts shortand generally wide (except T. hoggi); spermathecal organspresent.
Remarks
Tuberculosa conforms to the delimitation of the subfamilyLycosinae because the male pedipalp has a transversemedian apophysis with a ventrally directed spur and asinuous channel on the dorsal side of the median apophysis(Dondale 1986). We reject current delimitations of tribes(Zyuzin 1993) or ‘groups’ (Dondale 1986) within theLycosinae based on the morphology of male and femalegenitalia, because recent molecular data suggest that genitaltraits do not necessarily reflect phylogenetic relationships inthis subfamily very well (Murphy et al. 2006). Consequently,we do not place Tuberculosa in any of these supragenericgroups.
Distribution
Tropical northern Australia (Northern Territory andQueensland).
Etymology
The generic name is derived from the tubercle-shaped mod-ified setae on the ventral side of the third coxae and thetubercle on the outer edge of the fangs in males, in combina-tion with -osa, a generic ending for lycosid genera. Thegender is considered feminine.
Tuberculosa harveyi, sp. nov.
(Figs 1A-D, 2A-H, 3)
Material examined
Holotype. �, Darwin [12°27′S, 130°52′E, Northern Territory,Australia], 17.xi.1975, CSIRO (NTMAG A840).
Paratypes. 2�, Daly River [13°18′S, 130°15′E, NorthernTerritory, Australia], 31.viii.1998, T. B. Churchill, CSIRO/PWCNT,quadrat K4, A2190 (NTMAG A850, A3606).
Other material examined. Australia: Northern Territory: 1�,3�, Daly River, 13°18′S, 130°15′E (NTMAG A852; A862, A864); 1�,East Point Reserve, 12°24′S, 130°49′E (NTMAG A837); 4� 4�,14 juv., Kapalga Field Research Station, Kakadu National Park,12°36′S, 132°20′E (NTMAG A849, A851, A863; WAM T58340,T58357, T58370–1); 1�, North Point, Kapalga, 12°25′S, 132°22′E
(QM S66769); 1�, West Alligator mouth, 12°10′S, 132°14′E(QM S66761).
Diagnosis
Most similar to T. austini. Ventral edge of trough-shapedterminal apophysis of male pedipalp more or less straight(not curved). Hood openings of female epigyne form anangle of ~45° with a median line through the epigyne (notperpendicular to this line) and median connection of thehoods much further away from the posterior margin of theepigyne.
Description
Male (based on holotype, NTMAG A840)
Carapace. Brown with distinct yellow-brown medianband that is widest behind the PLE and narrows continuouslyposteriad; indistinct dark radial pattern; brown setae, butwhite setae in median band; one long black bristle betweenAME, four long black bristles below AME.
Eyes. Row of AE < row of PME; row of AE slightlyprocurved.
Sternum. Shiny yellow-brown, brown setae of increas-ing length and density towards margins.
Labium. Brown; front end truncated and white.Chelicerae. Orange-brown, basally somewhat darker;
dark brown setae which are longer basally; three promarginalteeth, with the median largest; three retromarginal teeth, withthe apical smallest; tubercle on the outer edge of the fangs.
Pedipalp. Ventral spur of median apophysis situatedretrolaterally (Figs 2E, F); embolus sickle-shaped, reachinghalf way along palea (Figs 2D); pars pendula well developed;terminal apophysis shaped like a trough with straight ventraledge (Figs 2D, E).
Abdomen. Light olive-brown with light grey medianband and indistinct light heart mark in anterior half; brownsetae laterally, but white setae in median band, denser in theanterior heart mark. Venter: light yellow-brown; brown setae.Spinnerets: light yellow-brown.
Legs. Leg formula IV < I < II < III; brown, femora dis-tinctly darker; coxae III with 9 (right) and 12 (left) modified,tubercle-like setae on ventral side (Fig. 1A); spination of legI: femur: 3 dorsal, 1 apicoprolateral, 1 apicoretrolateral (onlyleft leg); tibia: 3 ventral pairs, 2 prolateral; metatarsus: 3ventral pairs, 1 prolateral, 1 retrolateral, 1 apicoventral, 1apicoprolateral, 1 apicoretrolateral.
Female (based on NTMAG A3606)
Carapace. As male but overall darker; two indistinctdarker spots in anterior half of median band; median bandslightly constricted in front of fovea; one long black bristlebetween AME, two long black bristles below AME.
Eyes, sternum and labium. As male.Chelicerae. Dark reddish-brown; setae and dentition as
male.

Invertebrate Systematics 191
Abdomen. Dorsally uniformly light olive-grey butmedially somewhat lighter; brown setae, silver-grey setaemedially; less dense brown macrosetae. Venter and spin-nerets: as male.
Epigyne, ventral view. Distinct anterior hoods, openingof these hoods form 45° angle with median line throughepigyne (Fig. 2G).
Epigyne, dorsal view. Spermathecae large, ovoid; widecopulatory ducts connect postero-retrolaterally (Fig. 2H).
Legs. Leg formula IV < I < II < III; dark brown; femorawith indistinct lighter longitudinal line dorsally; spination ofleg I: femur: 3 dorsal, 1 apicoprolateral; tibia: 2 ventral pairs,1 apicoventral on prolateral side; metatarsus: 3 ventral pairs,1 apicoventral.
Measurements
� holotype, NTMAG A840 (�, NTMAG A3606): TL 4.61(5.64), CL 2.35 (2.63), CW 1.50 (1.60). Eyes: AME 0.09(0.10), ALE 0.08 (0.10), PME 0.23 (0.24), PLE 0.18 (0.19).Row of eyes: AE 0.46 (0.54), PME 0.51 (0.58), PLE 0.60(0.71). Sternum (length/width) 1.03/0.75 (1.03/0.85).Labium (length/width) 0.28/0.32 (0.29/0.36). AL 1.88(2.82), AW 1.20 (1.69). Legs: Length of segments (femur +patella/tibia + metatarsus + tarsus = total length): Pedipalp0.75 + 0.75 + - + 0.59 = 2.09, I 1.41 + 1.77 + 1.13 + 0.85 =5.16, II 1.41 + 1.69 + 1.03 + 0.80 = 4.93, III 1.22 + 1.32 +1.22 + 0.75 = 4.51, IV 1.74 + 2.26 + 1.97 + 0.94 = 6.91(Pedipalp 0.85 + 0.85 + - + 0.56 = 2.26, I 1.60 + 1.88 + 1.13+ 0.85 = 5.46, II 1.50 + 1.60 + 1.03 + 0.75 = 4.88, III 1.41 +
1.41 + 1.22 + 0.66 = 4.70, IV 1.88 + 2.35 + 2.16 + 0.94 =7.33).
Variation
�(�) (range, mean ± s.d.): TL 4.42 – 4.79, 4.58 ± 0.16,n = 4; CL 2.35 – 2.63, 2.43 ± 0.12, n = 5; CW 1.50 – 1.69,1.54 ± 0.08; n = 5 (TL 4.51 – 5.64, 5.19 ± 0.48, n = 6; CL2.26 – 2.82, 2.48 ± 0.19, n = 6; CW 1.41– 1.79, 1.60 ± 0.15;n = 5). The number of specialised coxal tubercles in malesranges from 9 – 16 and can differ between legs of the samespecimen. Females appear to be somewhat darker than malesand indistinct lighter submarginal bands are present on thecarapace of some specimens.
Life history and habitat preferences
Males were found from September through to December andfemales in an extended period from August through toFebruary. Collection data suggest that this species inhabitsthe litter of open forests and possibly savannas.
Distribution
Northern parts of the Northern Territory (Fig. 3).
Etymology
This specific epithet is a patronym in honour of MarkHarvey from the Western Australian Museum for hisongoing support in revising the wolf spiders of Australia.
Systematics of the new Australian wolf spider genus Tuberculosa
Fig. 3. Records of Tuberculosa haveyi, sp. nov.
V. W. Framenau and J.-S. Yoo192 Invertebrate Systematics
Tuberculosa austini, sp. nov.
(Figs 4A-E, 5)
Material examined
Holotype. �, Davies Creek National Park [10°00′S, 145°34′E,Queensland, Australia], 29.x.1991 – 23.vii.1992, pitfall trap,P. Lawless, R. Raven, M. Shaw, NQ8 (QM S25137).
Paratype. �, data as holotype (QM S69318).Other material examined. Australia: Queensland: 3 �, Davies
Creek National Park, 10°00′S, 145°34′E (QM S69319, S25120); 1 �,Douglas Campus, James Cook University, Townsville, 19°15′S,146°49′E (QM S13050); 1 �, Forty Mile Scrub National Park, 18°05′S,144°51′E (QM S66760).
Diagnosis
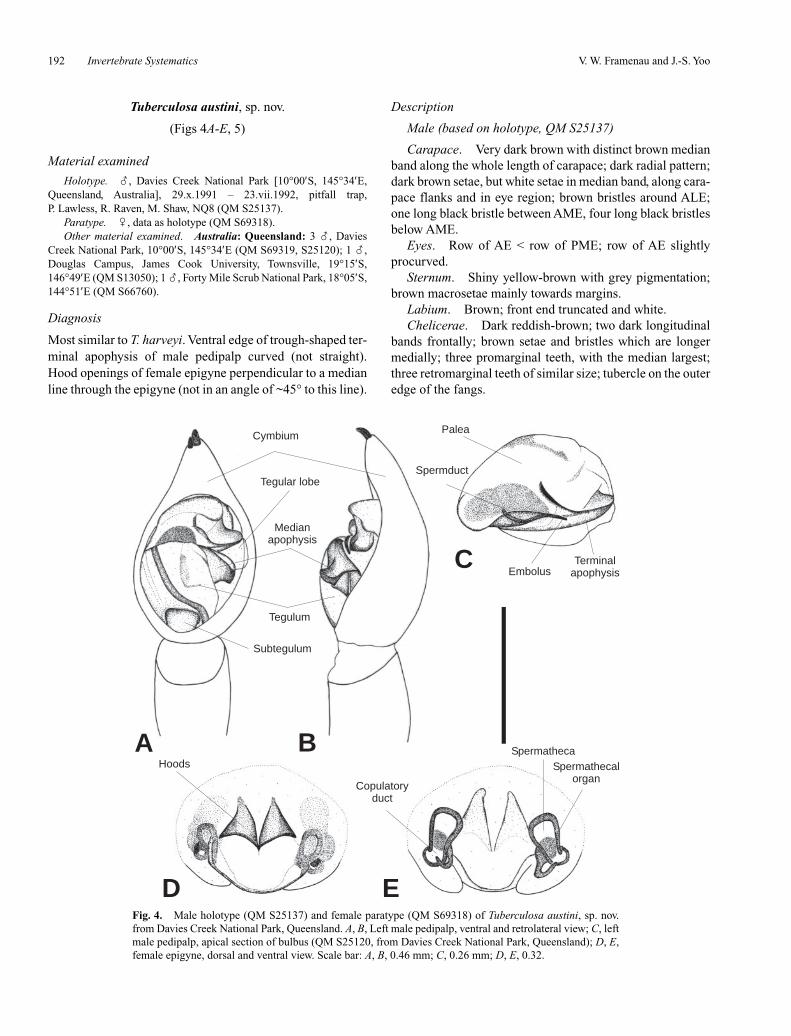
Most similar to T. harveyi. Ventral edge of trough-shaped ter-minal apophysis of male pedipalp curved (not straight).Hood openings of female epigyne perpendicular to a medianline through the epigyne (not in an angle of ~45° to this line).
Description
Male (based on holotype, QM S25137)
Carapace. Very dark brown with distinct brown medianband along the whole length of carapace; dark radial pattern;dark brown setae, but white setae in median band, along cara-pace flanks and in eye region; brown bristles around ALE;one long black bristle between AME, four long black bristlesbelow AME.
Eyes. Row of AE < row of PME; row of AE slightlyprocurved.
Sternum. Shiny yellow-brown with grey pigmentation;brown macrosetae mainly towards margins.
Labium. Brown; front end truncated and white.Chelicerae. Dark reddish-brown; two dark longitudinal
bands frontally; brown setae and bristles which are longermedially; three promarginal teeth, with the median largest;three retromarginal teeth of similar size; tubercle on the outeredge of the fangs.
C
A B
D E
Cymbium
Medianapophysis
TerminalapophysisEmbolus
Tegulum
Subtegulum
Copulatoryduct
SpermathecaSpermathecal
organHoods
Palea
SpermductTegular lobe
Fig. 4. Male holotype (QM S25137) and female paratype (QM S69318) of Tuberculosa austini, sp. nov.from Davies Creek National Park, Queensland. A, B, Left male pedipalp, ventral and retrolateral view; C, leftmale pedipalp, apical section of bulbus (QM S25120, from Davies Creek National Park, Queensland); D, E,female epigyne, dorsal and ventral view. Scale bar: A, B, 0.46 mm; C, 0.26 mm; D, E, 0.32.

Invertebrate Systematics 193
Pedipalp. Median apophysis pointing retrolaterallywith basally bent tip, ventral spur blunt (Figs 4A, B);embolus stout and tip pointing somewhat basally, reachinghalf way along palea (Fig. 4C); terminal apophysis shapedlike trough, ventral edge curved (Fig. 4A, C).
Abdomen. Dark olive-grey with yellow-brown medianband of approximately one third abdomen width; medianband with mottled grey pattern, in anterior half forming theindistinct borders of a lanceolate heart mark; brown setaeand macrosetae, but white setae in median band. Venter:olive-brown; brown setae. Spinnerets: light brown.
Legs. Leg formula IV < I < II < III; irregular dark greypigmentation; coxae III with 13 modified, tubercle-like setaeon ventral side; spination of leg I: femur: 3 dorsal, 1 apico-prolateral, 1 apicoretrolateral; tibia: 3 ventral pairs, 2 pro-lateral; metatarsus: 3 ventral pairs, 1 prolateral (2 on rightleg), 1 retrolateral, 1 apicoventral, 1 apicoprolateral, 1 api-coretrolateral.
Female (based on paratype, QM S69318)
Carapace, eyes, sternum and labium. As male, but moresetae on sternum.
Chelicerae. Dark reddish-brown; setae and dentition asmale.
Abdomen. Colour pattern similar but less distinct incomparison to male.
Epigyne, ventral view. Distinct anterior hoods, openingof hoods forms nearly 90° angle with a median line throughepigyne (Fig. 4D).
Epigyne, dorsal view. Oval spermathecae; copulatoryducts wide and short (Fig. 4E).
Legs. Leg formula IV < I < II < III; colour as male;spination of leg I: femur: 3 dorsal, 1 apicoprolateral; tibia:2 ventral pairs (of the apical third pair only one prolateral),1 prolateral; metatarsus: 3 ventral pairs, 1 apicoventral.
Measurements
� holotype, QM S25137 (� paratype, QM S69318): TL 4.61(4.89), CL 2.40 (2.30), CW 1.60 (1.41). Eyes: AME 0.09(0.09), ALE 0.09 (0.09), PME 0.24 (0.24), PLE 0.20 (0.20).Row of eyes: AE 0.44 (0.48), PME 0.56 (0.56), PLE 0.66(0.66). Sternum (length/width) 1.60/0.94 (1.03/0.85).Labium (length/width) 0.30/0.31 (0.31/0.38). AL 1.88(2.35), AW 1.32 (1.30). Legs: Lengths of segments (femur +patella/tibia + metatarsus + tarsus = total length): Pedipalp0.90 + 0.75 + - + 0.70 = 2.35, I 1.62 + 1.92 + 1.35 + 0.94 =5.83, II 1.50 + 1.82 + 1.32 + 0.87 = 5.51, III 1.45 + 1.50 +1.45 + 0.70 = 5.10, IV 1.99 + 2.39 + 2.26 + 0.70 = 5.10(Pedipalp 0.79 + 0.68 + - + 0.56 = 2.03, I 1.45 + 1.73 + 1.07+ 0.75 = 5.00, II 1.32 + 1.54 + 0.98 + 0.77 = 4.54, III 1.26 +1.37 + 1.13 + 0.60 = 4.36, IV 1.82 + 2.22 + 2.03 + 0.87 =6.94.
Variation
� (range, mean ± s.d.): TL 3.95 – 4.70, 4.18 ± 0.28; CL 2.16– 2.54, 2.34 ± 0.14; CW 1.41 – 1.65, 1.53 ± 0.08; n = 6. Thefemale of T. austini is only known from the paratype.
Systematics of the new Australian wolf spider genus Tuberculosa
Fig. 5. Records of Tuberculosa austini, sp. nov.
V. W. Framenau and J.-S. Yoo194 Invertebrate Systematics
Life history and habitat preferences
Tuberculosa austini has mainly been collected in Septemberand October. It has been found together with T. hoggi, whichsuggests that they share similar habitat requirements, dryeucalypt woodland.
Distribution
Eastern parts of central and northern Queensland (Fig. 5).
Etymology
The specific name is an patronym in honour of Andy Austinfrom the University of Adelaide for his ongoing support inrevising the wolf spiders of Australia.
Tuberculosa hoggi (Framenau & Vink, 2001), comb. nov.
(Figs 6A–E, 7)
Venatrix hoggi Framenau & Vink, 2001: 965–966, figs 46A–C, 47.
Material examined
Holotype. �, Helenvale, S of Cooktown [15°43′S, 145°14′E,Queensland, Australia], xi.1975, R.F. site 28, pitfall trap FN 601, coll.M. Gray (AM KS45407).
Paratype. �, data as holotype (AM KS 8151)..Other material examined. Australia: Queensland: 5�, 1 juv.,
Cape Cleveland Road, 19°21′23″S, 147°01′00″E (QM S19876,S22749, S40909, S69317); 4�, 2�, Davies Creek National Park,10°00′S, 145°34′E (QM S19798, S22707, SS25168, S66765); 7�, 6�,Eight Mile Creek, 18°41′00″S, 144°42′30″E (QM S21831, S21839,S21855, S21864, S21884, S22893, S22895, S22895, S35363, S69316);1�, nearby Emmett Creek, S of Townsville, 19°27′00″S, 147°02′40″E(QM S21959); 1�, 2�, nearby Running Creek, Innot Springs,17°18′45″S, 145°18′00″E (QM S24119, S66766); 1�, Sinclair Island,14°33′S, 144°54′E (QM S66762).
C
A B
D E
Cymbium
Medianapophysis
Terminalapophysis
Embolus
Tegulum
Subtegulum
Copulatoryduct
Spermatheca
Spermathecalorgan
Hoods
Palea
Spermduct
Tegular lobe
Lateral marginof epigyne
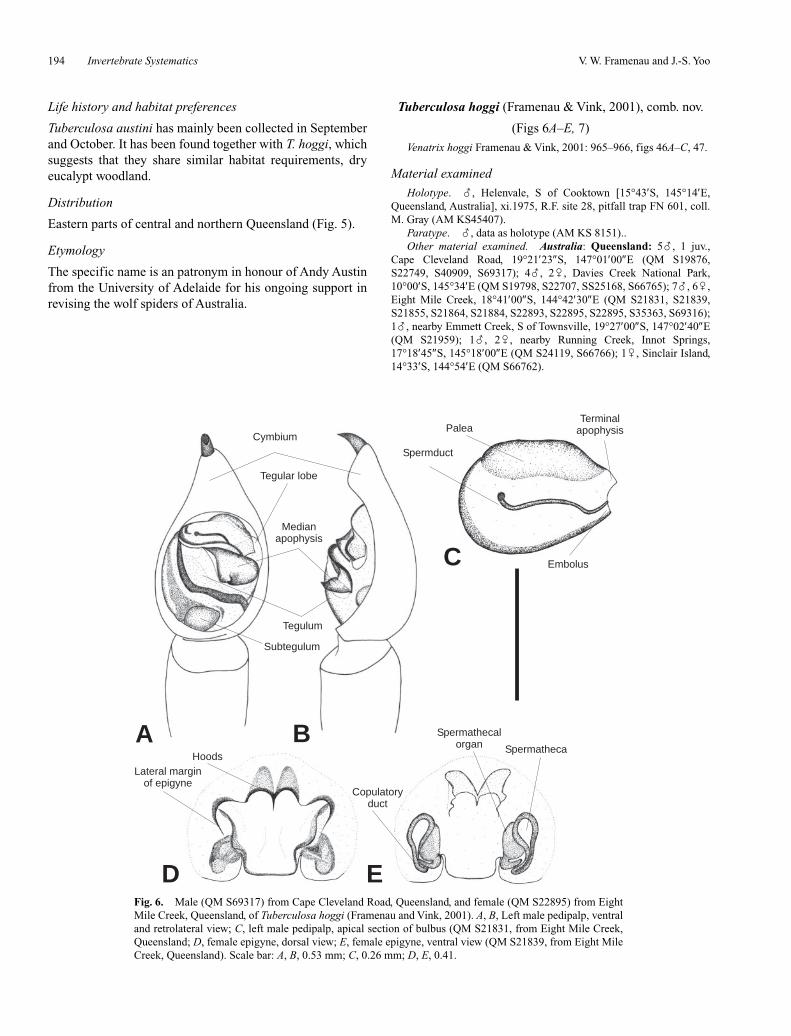
Fig. 6. Male (QM S69317) from Cape Cleveland Road, Queensland, and female (QM S22895) from EightMile Creek, Queensland, of Tuberculosa hoggi (Framenau and Vink, 2001). A, B, Left male pedipalp, ventraland retrolateral view; C, left male pedipalp, apical section of bulbus (QM S21831, from Eight Mile Creek,Queensland; D, female epigyne, dorsal view; E, female epigyne, ventral view (QM S21839, from Eight MileCreek, Queensland). Scale bar: A, B, 0.53 mm; C, 0.26 mm; D, E, 0.41.

Invertebrate Systematics 195
Diagnosis
Males with unique reduced, lobe-like terminal apophysis.Anterior sclerotised borders of female epigyne extending lat-erally past opening of hoods and curving around to nearlyhalf epigyne length (borders limited to the opening of theanterior hoods in all other Tuberculosa).
Description
Male (based on QM S69317)
Carapace. Dark brown with distinct light brownmedian band of slightly less than one third the carapacewidth; dark radial pattern; brown setae, but white setae inmedian and between PE; brown bristles around PME; onelong black bristle between AME, four long black bristlesbelow AME.
Eyes. Row of AE < row of PME; row of AE slightlyprocurved.
Sternum. Shiny yellow-brown with indistinct grey pig-mentation, brown setae of increasing length and densitytowards margins.
Labium. Brown; front end truncated and white.Chelicerae. Orange-brown; two dark longitudinal lines
frontally and grey pigmentation in between; dark brown bris-tles that are longer medially; three promarginal teeth, withthe median largest; three retromarginal teeth, with the apicalsmallest; tubercle on the outer edge of the fangs.
Pedipalp. Median apophysis pointing retrolaterally,ventral spur pointed (Figs 6A, B); embolus broad lamellarwithout distinct tip, terminal apophysis reduced (Fig. 6C).
Abdomen. Dark olive-grey with a lighter median bandthat has a mottled grey pattern forming the borders of anindistinct lanceolate heart mark in the anterior half; brownsetae, but white setae in median band. Venter: light olive-grey; light grey setae. Spinnerets: light olive-grey (asventer).
Legs. Leg formula IV < I < II < III; femora dark olive-brown with indistinct lighter longitudinal band medially;patellae, tibiae and metatarsi grey brown, tarsi brown; coxaeIII with 13 (right) and 14 (left) modified, tubercle-like setaeon ventral side; spination of leg I: femur: 3 dorsal, 1 apico-prolateral; tibia: 3 ventral pairs, 2 prolateral; metatarsus: 3ventral pairs, 1 prolateral, 1 retrolateral, 1 apicoventral, 1apicoprolateral, 1 apicoretrolateral.
Female (Based on QM S69316)
Carapace. As male but overall much lighter; distinctlight brown submarginal bands; setation as male with addi-tional white setae in submarginal bands.
Eyes, sternum and labium. As male.Chelicerae. Orange-brown with some grey pigmentation;
setae and dentition as male.Abdomen. Olive-grey; indistinct lanceolate heart mark
formed by indistinct dark mottled pattern; white setae.
Venter: light yellow-olive, brown setae and fewer macrose-tae. Spinnerets: yellow-brown.
Epigyne, ventral view. Distinct anterior hoods, theirsclerotised borders extend past hoods and reach around halfway along lateral borders of epigyne (Fig. 6D).
Epigyne, dorsal view. Oval spermathecae; copulatoryducts connecting posteriorly and narrow; spermathecalorgans large (Fig. 9E).
Legs. Leg formula IV < I < II < III; very light brown;femora with two distinct dark annulations; spination of leg I:femur: 2 dorsal, 1 apicoprolateral; tibia: 3 ventral pairs, 1prolateral; metatarsus: 3 ventral pairs, 1 apicoventral.
Measurements
�, QM S69317 (�, QM S69316): TL 4.37 (4.23), CL 2.21(2.35), CW 1.65 (1.60). Eyes: AME 0.09 (0.08), ALE 0.08(0.09), PME 0.25 (0.23), PLE 0.18 (0.19). Row of eyes: AE0.44 (0.47), PME 0.57 (0.56), PLE 0.68 (0.69). Sternum(length/width) 1.18/0.94 (1.08/0.85). Labium (length/width)0.29/0.33 (0.30/0.35). AL 1.79 (1.79), AW 1.13 (1.41). Legs:Length of segments (femur + patella/tibia + metatarsus +tarsus = total length): Pedipalp 0.90 + 0.83 + - + 0.71 = 2.44,I 1.60 + 1.99 + 1.24 + 0.88 = 5.71, II 1.47 + 1.79 + 1.15 +0.81 = 5.22, III 1.41 + 1.54 + 1.41 + 0.71 = 5.07, IV 2.01 +2.44 + 2.24 + 1.02 = 7.71 (Pedipalp 0.79 + 0.75 + - + 0.58 =2.12, I 1.50 + 1.82 + 1.03 + 0.75 = 5.10, II 1.35 + 1.62 + 1.00+ 0.71 = 4.68, III 1.30 + 1.32 + 1.11 + 0.64 = 4.37, IV 1.88+ 2.18 + 2.11 + 0.94 = 7.11.
Variation
�(�) (range, mean ± s.d.): TL 3.48 – 4.89, 4.15 ± 0.39; CL1.97 – 2.63, 2.33 ± 0.19; CW 1.32 – 1.79, 1.57 ± 0.15; n = 17(TL 3.95 – 4.89, 4.47 ± 0.32; CL 2.26 – 2.44, 2.34 ± 0.06;CW 1.50– 1.60, 1.56 ± 0.05; n = 7). Females appear to belighter and less distinctly coloured than males. Their sub-marginal light bands on the carapace can be broad and verydistinct but are sometimes dissolved in separate small spots.
Distribution
Eastern parts of north and central Queensland (Fig. 7).
Remarks
Tuberculosa hoggi was previously only known from twomales (Framenau and Vink 2001). The female is describedhere for the first time, and the variation of males is illustratedon a larger number of specimens.
Life history and habitat preferences
Tuberculosa hoggi is the most abundant of all Tuberculosa inmuseum collections. Collection data suggest a preference fordry eucalypt woodlands. Most specimens are from pitfalltraps which were exposed for too long to allow an interpre-tation of their phenology.
Systematics of the new Australian wolf spider genus Tuberculosa

V. W. Framenau and J.-S. Yoo196 Invertebrate Systematics
Tuberculosa monteithi, sp. nov.
(Figs 8A-C, 9)
Material examined
Holotype. �, Lockerbie [10°48′S, 142°27′E, Queensland,Australia], 16.-26.ix.1974, G.B. Monteith (QM S66764).
Paratype. �, data as holotype (QM S69320).
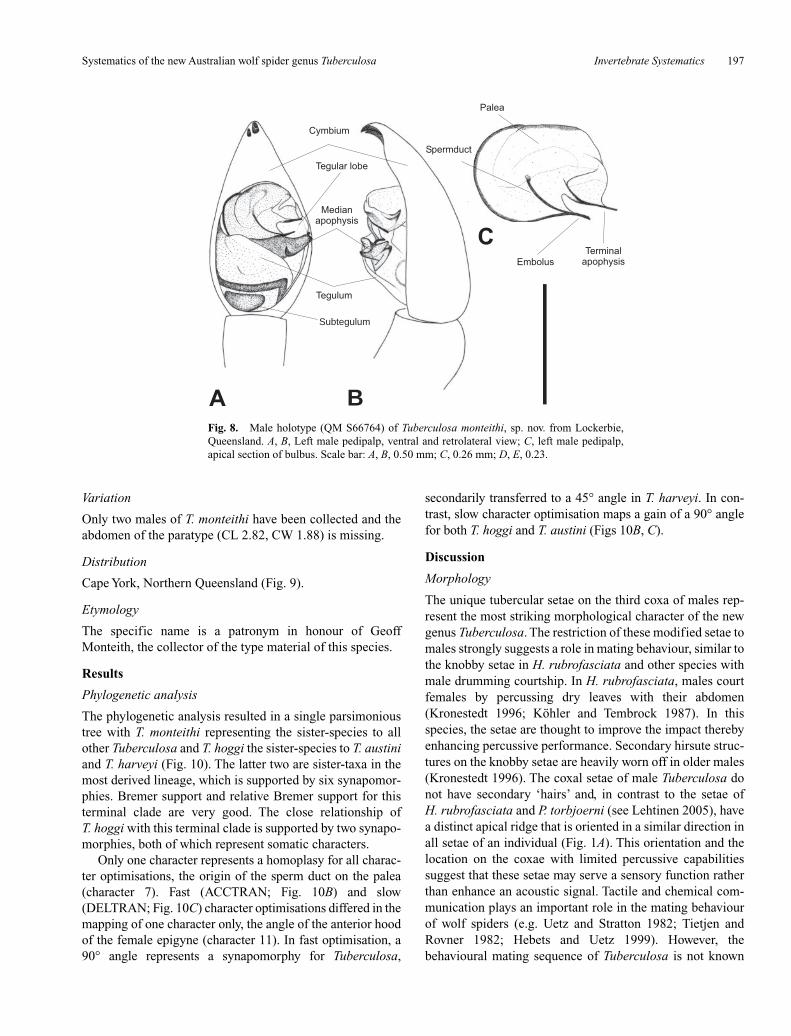
Diagnosis
Tegular apophysis of male pedipalp much slimmer than inany other Tuberculosa and terminal apophysis longer andnarrower; female unknown.
Description
Male (based on holotype, QM S66764)
Carapace. Dark brown with light-brown median bandthat is restricted to anterior half and approximately as wideas row of PLE; very indistinct light submarginal bands; darkradial pattern; head flanks somewhat lighter than carapace;few white setae, denser towards carapace margins; fewbrown bristles lateral of PME.
Eyes. Row of AE < row of PME; row of AE slightlyprocurved.
Sternum. Shiny yellow-brown with indistinct grey pig-mentation; brown macrosetae mainly near margins.
Labium. Brown; front end truncated and white.Chelicerae. Brown; few silver-white setae and some
brown bristles medially; three promarginal teeth, with the
median largest; three retromarginal teeth, with the apicalsmallest; tubercle on the outer edge of the fangs.
Pedipalp. Median apophysis pointing laterally, long andslim and reaching edge of cymbium (Figs 8A); terminalapophysis long and thin (Fig. 8C).
Abdomen. Brown; laterally with dark olive-greymottled pattern; few olive-grey spots in anterior half; whitesetae. Venter: brown with few olive-grey spots; white setaeand few brown macrosetae. Spinnerets: yellow-brown.
Legs. Leg formula IV < I < II < III; brown, femora withtwo dark annulations; coxae III with 14 (right) and 13 (left)modified, tubercle-like setae on ventral side; spination of legI: femur: 3 dorsal, 1 apicoprolateral; tibia: 3 ventral pairs,2 prolateral, 2 retrolateral; metatarsus: 3 ventral pairs, 1 pro-lateral, 1 retrolateral, 1 apicoventral, 1 apicoprolateral, 1apicoretrolateral.
Female
Unknown.
Measurements
� holotype, QM S66764: TL 4.70, CL 2.73, CW 1.88. Eyes:AME 0.08, ALE 0.08, PME 0.24, PLE 0.21. Row of eyes: AE0.50, PME 0.58, PLE 0.71. Sternum (length/width)1.32/0.94. Labium (length/width) 0.35/0.35. AL 1.88, AW1.32. Legs: Lengths of segments (femur + patella/tibia +metatarsus + tarsus = total length): Pedipalp 1.11 + 0.98 + -+ 0.77 = 2.86, I 1.79 + 2.20 + 1.41 + 1.03 = 6.43, II 1.65 +2.03 + 1.32 + 0.96 = 5.96, III 1.54 + 1.69 + 1.50 + 0.85 =5.58, IV 2.05 + 2.44 + 2.26 + 1.07 = 7.82.
Fig. 7. Records of Tuberculosa hoggi (Framenau and Vink, 2001).
Invertebrate Systematics 197
Variation
Only two males of T. monteithi have been collected and theabdomen of the paratype (CL 2.82, CW 1.88) is missing.
Distribution
Cape York, Northern Queensland (Fig. 9).
Etymology
The specific name is a patronym in honour of GeoffMonteith, the collector of the type material of this species.
Results
Phylogenetic analysis
The phylogenetic analysis resulted in a single parsimonioustree with T. monteithi representing the sister-species to allother Tuberculosa and T. hoggi the sister-species to T. austiniand T. harveyi (Fig. 10). The latter two are sister-taxa in themost derived lineage, which is supported by six synapomor-phies. Bremer support and relative Bremer support for thisterminal clade are very good. The close relationship ofT. hoggi with this terminal clade is supported by two synapo-morphies, both of which represent somatic characters.
Only one character represents a homoplasy for all charac-ter optimisations, the origin of the sperm duct on the palea(character 7). Fast (ACCTRAN; Fig. 10B) and slow(DELTRAN; Fig. 10C) character optimisations differed in themapping of one character only, the angle of the anterior hoodof the female epigyne (character 11). In fast optimisation, a90° angle represents a synapomorphy for Tuberculosa,
secondarily transferred to a 45° angle in T. harveyi. In con-trast, slow character optimisation maps a gain of a 90° anglefor both T. hoggi and T. austini (Figs 10B, C).
Discussion
Morphology
The unique tubercular setae on the third coxa of males rep-resent the most striking morphological character of the newgenus Tuberculosa. The restriction of these modified setae tomales strongly suggests a role in mating behaviour, similar tothe knobby setae in H. rubrofasciata and other species withmale drumming courtship. In H. rubrofasciata, males courtfemales by percussing dry leaves with their abdomen(Kronestedt 1996; Köhler and Tembrock 1987). In thisspecies, the setae are thought to improve the impact therebyenhancing percussive performance. Secondary hirsute struc-tures on the knobby setae are heavily worn off in older males(Kronestedt 1996). The coxal setae of male Tuberculosa donot have secondary ‘hairs’ and, in contrast to the setae ofH. rubrofasciata and P. torbjoerni (see Lehtinen 2005), havea distinct apical ridge that is oriented in a similar direction inall setae of an individual (Fig. 1A). This orientation and thelocation on the coxae with limited percussive capabilitiessuggest that these setae may serve a sensory function ratherthan enhance an acoustic signal. Tactile and chemical com-munication plays an important role in the mating behaviourof wolf spiders (e.g. Uetz and Stratton 1982; Tietjen andRovner 1982; Hebets and Uetz 1999). However, thebehavioural mating sequence of Tuberculosa is not known
Systematics of the new Australian wolf spider genus Tuberculosa
C
A B
Cymbium
Medianapophysis
TerminalapophysisEmbolus
Tegulum
Subtegulum
Palea
Spermduct
Tegular lobe
Fig. 8. Male holotype (QM S66764) of Tuberculosa monteithi, sp. nov. from Lockerbie,Queensland. A, B, Left male pedipalp, ventral and retrolateral view; C, left male pedipalp,apical section of bulbus. Scale bar: A, B, 0.50 mm; C, 0.26 mm; D, E, 0.23.

V. W. Framenau and J.-S. Yoo198 Invertebrate Systematics
and therefore an interpretation of the function of thetubercle-shaped setae remains speculative.
The distinct triangular anterior hoods are a commonfeature of females of Tuberculosa. Similar hoods are alsofound in V. konei and Tetralycosa arabanae, superficiallysupporting a relationship of Tuberculosa with both species.Although this holds true for V. konei as representative of theputative sister-genus Venatrix, male pedipalp morphologyand somatic characters contradict a close relationshipbetween Tuberculosa and Tetralycosa. The latter genusbelongs to a different subfamily altogether (Framenau et al.in press; Murphy et al. 2006). The convergent evolution of asimilar shape of anterior epigyne hoods is not unlikely takinginto account the function of these hoods during copulation.The ventral spur of the median apophysis of the male pedi-palp locks into this hood while the embolus is inserted at thebase of the epigyne (Fuhn and Niculescu-Burlacu 1971;Zyuzin 1993). The presence of similar structures in malesnecessitates matching structures in females even in unrelatedtaxa with an overall differing pedipalp arrangement.
Phylogeny
Owing to very uniform somatic morphology, the phylogenyof Tuberculosa is based on a limited number of characters. Inaddition, the female of T. monteithi is not known, potentiallyresulting in a further loss in resolution of the topology.However, the status of T. austini and T. harveyi as sister-species is well supported by several morphological charac-ters. Evident morphological synapomorphies include theconcave, trough-shaped terminal apophysis, unique within at
least Australian Lycosinae sensu Dondale (1986). This shapeis different to the reduced terminal apophysis in T. monteithiand T. hoggi, which appears to represent the ancestral statewithin Tuberculosa. In Venatrix, the putative sister-genus ofTuberculosa, the shape of the terminal apophysis is highlyvariable, most often sickle-shaped, but in contrast to thebasal species of Tuberculosa, generally sclerotised(Framenau and Vink 2001). The terminal apophysis repre-sents a particular problem in lycosid systematics and the des-ignation of homologous conditions on a higher systematicscale is contentious (see detailed discussion in Zyuzin 1993).In particular in genera with very derived pedipalp morphol-ogy such as Sosippus Simon, 1888, the phylogenetic inter-pretation of a variety of apophyses is difficult (Sierwald2000). This confusion is partly caused by differences in theposition and function of the terminal apophysis within theLycosidae (Zyuzin 1993). It also suffers from the intro-duction of a variety of terms for putatively homologousstructures, such as ‘conductor’ and ‘synembolus’. The intra-generic variation in the shape of the terminal apophysis, as inTuberculosa, suggests that this character is unsuitable forinferring higher phylogenetic relationships within theLycosidae.
Independent of character optimisation, the position of thesperm duct on the palea (character 7) is homoplasious in ouranalysis. Within Tuberculosa, the prolateral origin of thesperm duct in T. harveyi represents an unusual configuration,representing a reversal to the character state generally foundin other Lycosinae sensu Dondale (1986), includingVenatrix. Fast (Fig. 10B) and slow (Fig. 10C) character opti-
Fig. 9. Records of Tuberculosa monteithi, sp. nov.

Invertebrate Systematics 199
misations only differ in mapping the angle of the anteriorhoods of the female epigyne (character 11). However, thefemale of T. monteithi is not known, which makes an inter-pretation of the transformation of female character states dif-ficult. Alternative coding of this character for both possiblecharacters states in T. monteithi did not have an effect on thetopology presented here.
Monophyly of Venatrix
Venatrix is the best studied wolf spider genus in theAustralasian region, but a phylogenetic hypothesis for thisgenus does not exist (Framenau and Vink 2001; Framenau inpress b). The transfer of T. hoggi from Venatrix toTuberculosa could render the remaining species of Venatrixparaphyletic. It is beyond the scope of this study to present aphylogenetic hypothesis for both Venatrix and Tuberculosa
Systematics of the new Australian wolf spider genus Tuberculosa
4/58
Venatrix konei
Tuberculosa austini
Tuberculosa harveyi
Tuberculosa hoggi
Tuberculosa monteithi
7
1
12
1
10
0
9
1
6
1
5
0
1
1
3
0
2
0
Venatrix konei
Tuberculosa austini
Tuberculosa harveyi
Tuberculosa hoggi
Tuberculosa monteithi
8
1
7
1
4
1
11
0
7
1
4
1
12
1
10
0
9
1
8
1
6
1
5
0
1
1
3
0
2
0
11
1
Venatrix konei
Tuberculosa austini
Tuberculosa harveyi
Tuberculosa hoggi
Tuberculosa monteithi
8
1
7
1
4
1
11
17
1
11
1
4
1
12
1
10
0
9
1
8
1
6
1
5
0
1
1
3
0
2
0
A
B
C
1/50
Fig. 10. Phylogenetic reconstruction (tree length 17; consistency index (CI) = 0.70;retention index (RI) = 0.58) of Tuberculosa with varying character optimization. A,Unambiguous characters only (Bremer/relative Bremer (1–100) Supports shown in encirclednumbers); B, fast optimisation (ACCTRAN); C, slow optimisation (DELTRAN). Characters:black circles, syn- and autapomorphies; white circles, homoplasies. Character numbersappear above and character states below characters (see Table 1).

V. W. Framenau and J.-S. Yoo200 Invertebrate Systematics
to clarify the monophyly of Venatrix; however, there is amplemorphological support that the remaining species ofVenatrix, including V. konei, represent a monophyletic clade.Of particular importance are characters that are common toVenatrix and Tuberculosa but not present in other Lycosinaesensu Dondale (1986), since character states of the latter mayrepresent plesiomorphic conditions that cannot supportVenatrix monophyly. In that sense, the best support forVenatrix monophyly is provided by the dorsal edge of theclaw-like macrosetae at the tip of cymbium in males, whichis not present in other southern hemisphere Lycosinae. Thisedge is lobed or even serrated in all Venatrix, whereas it issmooth in Tuberculosa. The distinct ventral pattern with twolight lines (sometimes reduced to patches) on a dark surfaceis another character that unifies Venatrix. This pattern maynot be present in adult spiders, but can be seen in most juve-niles (e.g. V. lapidosa McKay, 1974; V. W. Framenau, per-sonal observation).
Biogeography
The putative sister-taxon to Tuberculosa, Venatrix, currentlyincludes 28 species (Framenau and Vink 2001; Framenau inpress b). Framenau and Vink (2001) realised close affinitiesof Venatrix to the northern hemisphere taxa Alopecosa andTrochosa. The type species of Alopecosa, A. fabrilis (Clerck,1757) has a tubercle on the outer edge of the fangs in males(e.g. Wiebes 1959:4, fig. 10), and ~10% of males of the typespecies of Trochosa, T. ruricola (De Geer, 1778) (Engelhardt1964). Curiously, no other representative of these two generaknown to us displays this tubercle. The tropical distributionof Tuberculosa supports the link of Venatrix with thenorthern hemisphere. Recently, Venatrix has also been foundon the Philippines and in Palau (Framenau in press b). Incontrast, all other lycosids of Australia appear to be ofGondwanan origin (Murphy et al. 2006).
It is possible that the distribution of Tuberculosa extendsbeyond the tropical northern parts of Australia into neigh-bouring countries, such as Indonesia and Papua New Guinea.Similar patterns of Oriental distribution with strongholdsnorth of Australia are prevalent for other wolf spider generasuch as Venonia and Zoica, although Venonia micarioides(L. Koch, 1877) spreads into the temperate southern regionsof Australia (V. W. Framenau and J.-S. Yoo, unpublisheddata).
Tuberculosa biogeography is at odds with the phylo-genetic history of the genus because the two phylo-genetically closest taxa T. harveyi and T. austini show a verydisjunct distribution. Tuberculosa austini is found in closeproximity to T. hoggi and T. monteithi in tropical northernand central Queensland, whereas T. harveyi is the onlyspecies from the Northern Territory. The geographical splitbetween the sister-taxa T. austini and T. harveyi coincideswith a biogeographic boundary also found in other groups,for example birds (line ‘D’, Cracraft 1982). A vicariance
event dividing Cape York and Arnhem Land, such as a tem-porary water body or areas of aridification, could serve as anexplanation. The Carpenteria Basin (today Gulf ofCarpenteria) was much wider and intruded further into theAustralian continent in the early Cretaceous (c. 110 millionyears ago) (Doutch 1972). Pleistocene (c. 2 million to 11000years ago) variations in sea level may have produced morerecent vicariance events also postulated for Pacific Islands(Yan and Kroenke 1993). Assuming an origin of Tuberculosain the Oriental region, an alternative hypothesis could beindependent immigrations of founder populations fromPapua New Guinea. There is evidence of several waves ofmigrations in other taxa, corresponding to openings andclosings of rainforest corridors between New Guinea andAustralia (Kikkawa et al. 1981; Monteith 1997). However,the interpretation of the biogeographic history of wolfspiders is hampered by the fact that a molecular clock datingspeciation events does not exist (Vink and Paterson 2003). Inaddition, the ability of wolf spiders to balloon in their larvalstages may account for disjunct distributions of closelyrelated species (e.g. Richter 1970; Greenstone 1982).
Acknowledgments
We are grateful to Mike Gray and Graham Milledge (AM),Robert Raven and Owen Seeman (QM) and Gavin Dally(NTMAG) for the loan of specimens for this study. We areparticularly grateful to Barbara Baehr (QM) and TraceyChurchill (NTMAG) and their respective families for theirhospitality during visits of the senior author to Brisbane andDarwin. Young-Bo Lee and An-Ja Ko (National Institute ofAgricultural Science and Technology, RDA, Suwon, Korea)took the SEM photographs. Melissa Thomas, Mark Harveyand Pekka Lehtinen gave helpful comments on earlier draftsof this manuscript. The Australian Biological ResourceStudy (ABRS) (to Mark Harvey, WAM, and Andy Austin,The University of Adelaide) provided funding for VWF. Thiswork was supported by the Korea Research FoundationGrant funded by the Korean Government (MOEHRD)(KRF-2005- 214-C00226).
References
Barthel, J., and Helversen, O. (1990). Pardosa wagleri (Hahn 1822) andPardosa saturatior Simon 1937, a pair of sibling species (Araneae,Lycosidae). Bulletin de la Société Europeenne d’Arachnologie 1,17–23.
Bremer, K. (1988). The limits of amino acid sequence data inangiosperm phylogenetic reconstruction. Evolution 42, 795–803.
Bremer, K. (1994). Branch support and tree stability. Cladistics 10,295–304. doi:10.1111/j.1096-0031.1994.tb00179.x
Cracraft, J. (1982). Geographic differentiation, cladistics, and vicari-ance biogeography: reconstructing the tempo and mode of evolu-tion. American Zoologist 22, 411–424.
Dondale, C. D. (1986). The subfamilies of wolf spiders (Araneae:Lycosidae). Actas X Congreso Internacional de Aracnología, Jaca,España 1, 327–332.

Invertebrate Systematics 201
Doutch, H. F. (1972). The palaeogeography of northern Australia andNew Guinea and its relevance to the Torres Strait area. In ‘Bridgeand Barrier: The Natural and Cultural History of Torres Strait’.(Ed. D. Walker.) pp. 1–10. (Reseach School of Pacific Studies,Department of Biogeography and Geomorphology, AustralianNational University: Canberra, Australia.)
Elgar, M. A. (1991). Sexual cannibalism, size dimorphism, andcourtship behaviour in orb-weaving spiders (Araneidae). Evolution45, 444–448.
Engelhardt, W. (1964). Die mitteleuropäischen Arten der GattungTrochosa C. L. Koch, 1848 (Araneae, Lycosidae). Morphologie,Chemotaxonomie, Biologie, Autökologie. Zeitschrift fürMorphologie und Ökologie der Tiere 54, 219–392.doi:10.1007/BF00390678
Framenau, V. W. (2002). Review of the wolf spider genus ArtoriaThorell (Araneae, Lycosidae). Invertebrate Systematics 16,209–235. doi:10.1071/IT01028
Framenau, V. W. (2005). Gender specific differences in activity andhome range reflect morphological dimorphism in wolf spiders(Araneae, Lycosidae). Journal of Arachnology 33, 334–346.
Framenau, V. W. (in press a). Mainosa, a new genus for the Australian‘Shuttlecock Wolf Spider’ (Araneae, Lycosidae). Journal ofArachnology.
Framenau, V. W. (in press b). The wolf spider genus Venatrix: newspecies, synonyms and generic transfers. Records of the WesternAustralian Museum.
Framenau, V. W., and Vink, C. J. (2001). Revision of the wolf spidergenus Venatrix Roewer (Araneae: Lycosidae). InvertebrateTaxonomy 15, 927–970. doi:10.1071/IT01008
Framenau, V. W., Gotch, T. B., and Austin, A. D. (In press). The wolfspiders of artesian springs in arid South Australia, with a revalida-tion of Tetralycosa (Araneae, Lycosidae). Journal of Arachnology.
Fuhn, I. E., and Niculescu-Burlacu, F. (1971). Lycosidae. FaunaRepublicii Socialiste România. Arachnida 5, 1–256.
Gasnier, T. R., de Azevedo, C. S., Torres-Sanchez, M. P., and Höfer, H.(2002). Adult size of eight hunting spider species in centralAmazonia: temporal variations and sexual dimorphism. Journal ofArachnology 30, 146–154.
Goloboff, P. (1993). ‘NONA, Version 2.0.’ (Instituto Miguel Lillo:Sierra Madre de Tucuman, Argentina.) Available online at:www.cladistics.com [verified March 2006].
Greenstone, M. H. (1982). Ballooning frequency and habitat pre-dictability in two wolf spider species (Lycosidae: Pardosa). TheFlorida Entomologist 65, 83–89.
Head, G. (1995). Selection on fecundity and variation in degree ofsexual size dimorphism among species (class: Araneae). Evolution49, 776–781.
Hebets, E. A., and Uetz, G. W. (1999). Female responses to isolatedsignals from multimodal male courtship display in the wolf spidergenus Schizocosa (Araneae: Lycosidae). Animal Behaviour 57,865–872. doi:10.1006/anbe.1998.1048
Hebets, E. A., and Uetz, G. W. (2000). Leg ornamentation and the effi-cacy of courtship display in four species of wolf spider (Araneae:Lycosidae). Behavioral Ecology and Sociobiology 47, 280–286.doi:10.1007/s002650050667
Kikkawa, J., Monteith, G. B., and Ingram, G. (1981). Cape YorkPeninsula: the major region of faunal interchange. In ‘EcologicalBiogeography in Australia’. (Ed. A. Keast.) pp. 1697–1742 (Junk:The Hague, The Netherlands.)
Köhler, D., and Tembrock, G. (1987). Akustische Signale bei derWolfsspinne Hygrolycosa rubrofasciata (Arachnida: Lycosidae).Zoologischer Anzeiger 219, 147–153.
Kotiaho, J., Alatalo, R. V., Mappes, J., and Parri, S. (1996). Sexualselection in a wolf spider: male drumming activity, body size, andviability. Evolution 50, 1977–1981.
Kronestedt, T. (1996). Vibratory communication in the wolf spiderHygrolycosa rubro-fasciata (Araneae, Lycosidae). Revue Suisse deZoologie hors série I , 341–354.
Lehtinen, P. T. (2005). Review of the Oriental wolf spider genusPassiena (Lycosidae, Pardosinae). Journal of Arachnology 33,398–407.
Lehtinen, P. T., and Hippa, H. (1979). Spiders of the Oriental-Australianregion. I. Lycosidae: Venoniinae and Zoicinae. Annales ZoologiciFennici 16, 1–22.
Maddison, W. P., Donague, M. J., and Maddison, D. R. (1984).Outgroup analysis and parsimony. Systematic Zoology 33, 83–103.
Mappes, J., Alatalo, R. V., Kotiaho, J., and Parri, S. (1996). Viabilitycosts of condition-dependent sexual male display in a drummingwolf spider. Proceedings of the Royal Society of London. Series B.Biological Sciences 263, 785–789.
Miller, G. L., Stratton, G. E., Miller, P. R., and Hebets, E. A. (1998).Geographic variation in male courtship behaviour and sexual isola-tion in wolf spiders of the genus Schizocosa. Animal Behaviour 56,937–951. doi:10.1006/anbe.1998.0851
Monteith, G. B. (1997). Revision of the Australian flat bugs of the sub-family Mezirinae (Insecta: Hemiptera: Aradidae). Memoirs of theQueensland Museum 41, 1–169.
Moya-Laraño, J., Halaj, J., and Wise, D. H. (2002). Climbing to reachfemales: Romeo should be small. Evolution 56, 420–425.
Moya-Laraño, J., Taylor, P. W., and Fernández-Montraveta, C. (2003).Body patterns as potential amplifiers of size and condition in a terri-torial spider. Biological Journal of the Linnean Society 78,355–364. doi:10.1046/j.1095-8312.2003.00148.x
Murphy, N. P., Framenau, V. W., Donellan, S. C., Harvey, M. S.,Park, Y.-C., and Austin, A. D. (2006). Phylogenetic reconstructionof the wolf spiders (Araneae, Lycosidae) using sequences from 12SrRNA, 28S rRNA and NADH1 genes: implications for classi-fication, biogeography and the evolution of web-building behavior.Molecular Phylogenetics and Evolution 38, 583–602.
Nixon, K. C. (2002). ‘WinClada, version 1.00.08.’ (Published bythe author: Ithaca, NY, USA.) Available online at:http://www.cladistics.com/ [verified March 2006].
Nixon, K. C., and Carpenter, J. M. (1993). On outgroups. Cladistics 9,413–426. doi:10.1111/j.1096-0031.1993.tb00234.x
Page, R. D. M. (2001). ‘NEXUS Data Editor for Windows, version0.5.0.’ (Published by the author: Glasgow, UK.) Available online at:http://taxonomy.zoology.gla.ac.uk/rod/NDE/nde.html [verifiedMarch 2006].
Prenter, J., Elwood, R. W., and Montgomery, W. I. (1999). Sexual sizedimorphism and reproductive investment by female spiders: a com-parative analysis. Evolution 53, 1987–1994.
Richter, C. J. J. (1970). Aerial dispersal in relation to habitat in eightwolf spider species (Pardosa, Araneae, Lycosidae). Oecologia 5,200–214. doi:10.1007/BF00344884
Sheehan, D. C., and Hrapchack, B. B. (1980). ‘Theory and Practice ofHistotechnology.’ 2nd edn. (Battelle Press: Columbus, OH, USA.)
Sierwald, P. (2000). Description of the male of Sosippus placidus, withnotes on the subfamily Sosippinae (Araneae, Lycosidae). Journal ofArachnology 28, 133–140.
Tietjen, W. J., and Rovner, J. S. (1982). Chemical communication inlycosids and other spiders. In ‘Spider Communication: Mechanismsand Ecological Significance’. (Eds P. N. Witt and J. S. Rovner.)pp. 249–279 (Princeton University Press: Princeton, NJ, USA.)
Uetz, G. W., and Stratton, G. E. (1982). Acoustic communication andreproductive isolation in spiders. In ‘Spider Communication:Mechanisms and Ecological Significance’. (Ed. P. N. Witt andJ. S. Rovner.) pp. 123–159 (Princeton University Press: Princeton,NJ, USA.)
Systematics of the new Australian wolf spider genus Tuberculosa

V. W. Framenau and J.-S. Yoo202 Invertebrate Systematics
Vink, C. J. (2001). A revision of the genus Allotrochosina Roewer(Lycosidae: Araneae). Invertebrate Taxonomy 15, 461–466.doi:10.1071/IT00022
Vink, C. J., and Paterson, A. M. (2003). Combined molecular andmorphological phylogenetic analyses of the New Zealand wolfspider genus Anoteropsis (Araneae: Lycosidae). MolecularPhylogenetics and Evolution 28, 576–587. doi:10.1016/S1055-7903(03)00219-7
Vollrath, F., and Parker, G. A. (1992). Sexual dimorphism and distortedsex ratios in spiders. Nature 360, 156–159. doi:10.1038/360156a0
Walker, S. E., and Rypstra, A. L. (2001). Sexual dimorphism in func-tional response and troph morphology in Rabidosa rabida (Araneae:Lycosidae). American Midland Naturalist 146, 161–170.
Walker, S. E., and Rypstra, A. L. (2002). Sexual dimorphism in trophicmorphology and feeding behavior of wolf spiders (Araneae:Lycosidae) as a result of differences in reproductive roles. CanadianJournal of Zoology 80, 679–688. doi:10.1139/z02-037
Watrous, L. E., and Wheeler, Q. D. (1981). The out-group comparisonmethod of character analysis. Systematic Zoology 30, 1–11.
Wiebes, J. T. (1959). The Lycosidae and Pisauridae (Araneae) of theNetherlands. Zoologische Verhandelingen 42, 1–79.
Yan, C. Y., and Kroenke, L. W. (1993). A plate tectonic reconstructionof the southwest Pacific, 0–100 Ma. Proceedings of the OceanDrilling Program, Scientific Results 130, 697–709.
Zyuzin, A. A. (1993). Studies on the wolf spiders (Araneae: Lycosidae).I. A new genus and species from Kazakhstan, with comments on theLycosinae. Memoirs of the Queensland Museum 33, 693–700.
Manuscript received 18 August 2005, revised and accepted 17 January2006.
http://www.publish.csiro.au/journals/is