Embed Size (px)
Citation preview

J. Cell Sci. 6, 87-107 (1970) 87
Printed in Great Britain
TEMPERATURE-DEPENDENT FUNCTIONAL
STRUCTURES IN THE POLYTENE
CHROMOSOMES OF PHASEOLUS, WITH
SPECIAL REFERENCE TO THE NUCLEOLUS
ORGANIZERS
W. NAGL*Max-Planck-Institut filr Pflanzengenetik, 6802 Ladenburg-Rosenhof, West Germany
SUMMARY
During the endopolyploidization of the suspensor cells of Phaseolus coccineus and P. vulgaris,polytene chromosomes and an important nucleolar apparatus develop within the nuclei. Thenucleolus-associated regions of the four nucleolus-organizing chromosomes and the regions ofthe secondary constrictions (intranucleolar chromatin threads) show a temperature-dependent,reversible variability in their structure. They split up and are dispersed at optimal temperatures,and they are compact and partly banded at too low or too high ones. Many nucleolar extrusions,and additional nucleoli which originate on distended elements of the heterochromatic parts ofdifferent giant chromosomes, occur at temperatures higher than 15 CC.
The polytene chromosomes of P. coccineus are able to form lateral loops projecting at rightangles to their axes, shortly after temperature elevation. This occurs by the uncoiling of thesuperficial chromomeres. Several loops synthesize droplets and, moreover, they formnucleolus-like bodies in a hitherto unknown manner. The duration of the lampbrush state isabout 3-4 h.
Cytochemical staining methods indicate the presence of DNA in all parts of the polytenechromosomes, including the intranucleolar secondary constriction and the lateral loops, and thepresence of RNA in all types of nucleoli and nucleolus-like bodies.
The structural modifications described are compared with the structure of salivary glandchromosomes of Diptera and lampbrush chromosomes in oocytes and spermatocytes. It issuggested that the structural changes are an expression of temperature-dependent changes ingene activity.
INTRODUCTION
Polytene chromosomes occur not only in animals, but also in Angiospermae(Tschermak-Woess, 1956, 1957; Hasitschka-Jenschke, 1959, 1962; Enzenberg, 1961;Nagl, 1962 a, b\ Erbrich, 1965). In some species of higher plants polytene chromosomes,which occur within the ovule, are found only occasionally (review by Tschermak-Woess, 1963), but in Rhinanthus and Phaseolus they are always present (Tschermak-Woess, 1957; Nagl, 1962a, b, 1969a). The giant chromosomes of Rhinanthus are,however, individually indistinguishable because of their granular, unbanded appear-ance, and the most favourable material for the study of plant polytene chromosomestherefore appears to be the suspensor of the legume Phaseolus. In this genus the highly
• Present address: Institute of Botany, The University of Vienna, 1030 Wien, Austria.

88 W. Nagl
endopolyploid basal cells of the suspensor contain 22 unpaired giant chromosomes,
which are individually distinguishable, at least in part, on the basis of their different
lengths, the different length ratios of their arms, and the specific distribution of eu-
and hetero-chromatic regions (Nagl, 19626, 1965, 1967). Moreover, different structural
modifications in the form of local swellings and lateral projections were found in the
species P. coccineus (Nagl, 1967), but the relationship of these to internal and external
factors remained unclear.
Temperature is an important factor in plant growth, and it influences the gross
morphology of the polytene chromosomes in Phaseolus vulgaris (Nagl, 1969a). In
Diptera, in addition to a similar effect of this kind (Bridges, 1936; Schultz, 1936;
Wolf, 1957; Bier, 1958, 1959), a change of the puffing pattern was observed following
temperature shocks (Berendes, van Breugel & Holt, 1965; Berendes, 1968). A study
was therefore made of the effect of growth temperature and of temperature shocks on
the morphology of the polytene chromosomes of Phaseolus. The results are presented
in this paper. Particular regard was given to the nucleolus-organizing chromosomes
and to the occurrence of additional nucleoli and nucleolus-like bodies, because these
give a morphological indication of the degree of nuclear activity.
MATERIALS AND METHODS
All studies and experiments were performed with phytotron-cultivated plants. The standardconditions were the following: a light-dark rhythm of 13:11 h, 6 5 % relative humidity, and atemperature of 27 °C in the light and 22 °C in the dark. In the experiments only the temperaturewas changed. One phytochamber was used throughout to maintain the standard conditions forcontrols. For each experiment 20 plants of Phaseolus vulgaris (variety 'Hild's Marona') and5 plants of Phaseolus coccineus (var. coccineus; seeds from the Botanical Garden of the Universityof Vienna) were used. Most of the studies were made with ovules of pods of a length between8 and 10 cm in P. vulgaris, and of 15 and 18 cm in P. coccineus. At this stage the cotyledons ofthe embryo occupy half of the embryo-sac. The nuclei examined for structural modifications ofthe polytene chromosomes showed a degree of polyploidy of 1024*, 2048*, or 4096.Y, estimatedfrom volume determinations (compare Nagl, 1962a, b) and by microphotometric measurementsof Feulgen-DNA (W. Nagl, unpublished).
The ovules were halved, and the suspensor—lying between the micropyle and the embryo—was taken out and fixed in ethanol/acetic acid (3/1) at 4 °C for 24 h. Other fixatives are un-favourable, as they do not allow squash preparations. The chromosomes are too large for paraffinsections to be useful. However, fixation in 10 % neutral formalin was carried out if proteins wereto be stained. The ethanol/acetic acid-fixed cells were treated with 45 % acetic acid at 22 °C for3—6 h, squashed and examined under a phase-contrast microscope. The quality of this prepara-tive technique was tested by comparison with living cells of lower endopolyploidy.
Cytochemical methods were used for the localization of DNA, proteins and RNA. DNA wasdetected using the Feulgen reaction; the hydrolysis was carried out with N HC1 (60 CC, 10 min)after pectinase treatment (compare Nagl, 1967), and sometimes with 10% perchloric acid(20 °C, 15 h); pectinase of Serva (Heidelberg), and />-rosaniline, free of acridine (Chroma-Ges.,Stuttgart) were used. Basic proteins (histones) of the chromosomes were stained according tothe method of Alfert & Geschwind (1953); for this, the formalin-fixed cells were hydrolysedwith trichloroacetic acid (5 %, 100 °C, 15 min), attached to the slides with Celloidin (1 %), andstained with fast green FCF (National Aniline Division, New York) at pH 8-2. The methylgreen-pyronin method of Brachet (1953) was used for the detection of RNA in nucleoli andnucleolus-like bodies (stains from G. Gurr Ltd, London); controls were treated with ribonu-clease (Serva, Heidelberg) for 3 h at pH 7 0 (3 mg/ml, 38 °C). Some cells were stained with fastgTeen at pH 50, and acid fuchsin at pH 7-0 for other proteins, including acid proteins. For

Polytene chromosomes in Phaseolus 89
studies of the gross morphology of the polytene chromosomes, the cells were hydrolysed withN HC1 at 55 CC for 15 min and stained with pyronin (Nagl, 1967), or hydrolysed for 30 min andstained with 005 % toluidine blue O (Merck A. G., Darmstadt) at pH 4-0 for 1 h.
Unless otherwise stated, drawings and photographs were made from unstained preparationsusing phase-contrast.
RESULTS
Development and gross morphology of the suspensor nuclei
During the development of the suspensor three stages are distinguishable: (a) theproliferation stage, which ends with a pro-embryo composed of diploid cells; (b) thedifferentiation stage, during which the suspensor grows endomitotically, and theembryo mitotically; and (c) the functional stage, during which the mature suspensorprobably nourishes the rapidly growing embryo, especially the cotyledons. When thecotyledons reach the chalazal end of the embryo-sac, the suspensor cells are crushedafter a short degeneration period.
The development of the polytene chromosomes was studied in the basal cells of thesuspensor during stage (b). The duration of this was found to be about one week, underthe standard conditions.
The diploid and low endopolyploid nuclei possess some small chromocentres (andendochromocentres) within a fine euchromatic background. Some of the heterochro-matic areas are associated with the spherical nucleolus, evidently being the nucleolusorganizers and the satellites (the diploid complement contains 4 nucleolus-organizingchromosomes). During the further endopolyploidization the endochromocentres en-large, and the euchromatic elements join to form granular parts of the future giantchromosomes (see Nagl, 19626), provided no mitosis interrupts the endopolyploidi-zation process (Nagl, 1969c). Finally, the whole chromatin of the nucleus is arrangedin 22 polytene chromosomes (Fig. 4). They have characteristic lengths, showing acharacteristic pattern of their eu- and hetero-chromatic portions (compare the idio-gram of Nagl, 1967). At optimal temperatures, the euchromatic regions of the chromo-somes have a granular appearance, but cooling of the plants results in the appearance of abanding pattern (Nagl, 1969a). The four polytene nucleolus-organizing chromosomesare the most easily identified ones (Nagl, 1965).
The nucleolus also enlarges significantly during nuclear growth. It persists duringendomitosis, which is of the common angiosperm-type (Nagl, 1965, 1967). As soon asthe polytene chromosomes become distinct, the hitherto spherical nucleolus elongates,and it becomes very irregularly shaped at higher degrees of endopolyploidy, with manylobes (Fig. 5). Within the nucleoli of all stages bright vacuoles can be seen. Chromaticmaterial, evidently forming the secondary constriction of the nucleolus-organizingchromosomes, is perceptible in some of the vacuoles. The total connexion between thenucleolus organizers and the satellites is commonly visible only after hydrolysis andstaining. It must be noted that in some cells the nucleolus divides into 2 or more parts.In this case, every nucleolar fragment is in connexion with one, or one pair, of chromo-somes, the larger pieces always with those of type I (see Nagl, 1965, 1967).

o,o W. Nagl
The effect of growth temperature on the nucleolus-associated regions of the polytevechromosomes
The structure of the nucleolus-associated regions of the polytene chromosomes, i.e.the nucleolus organizers and the satellites, was studied at three temperature levels:27722 °C, i5°/i2 CC, and i2°/8 °C (the first value denotes the light period, the secondthe dark period).
At 27722 °C, the nucleolus-associated regions split up into many threads of di-minishing thickness, running partly on the surface of the nucleolus, and penetratingpartly along vacuoles (Fig. 6). The satellites are shaped like a spider (Fig. 7), or theymay be fragmented into ramified or spherical patches, lying on the surface of thenucleolus (Fig. 5). If the temperature is lowered to 15 °C or 12 °C for 2 days or longer,the opened state of the nucleolus-associated regions is reduced, and the degree oframification is less than at higher temperatures. A significant change in the structureoccurs at i2°/8 °C. Under these conditions most of the nuclei have condensed nucleolusorganizers, and the satellites become compact and spherical (Figs. 8, 9). This structuralchange is reversible on returning the plants to one of the higher temperature levels forsome hours. It seems that the nucleolus organizers and satellites respond more quicklyto temperature alterations than the other parts of the chromosome complement, andthat the reaction to raising the temperature occurs more rapidly than does that tolowering it. Possibly only the intranucleolar chromatin is more sensitive to thoseinfluences.
The effect of temperature on the main nucleolus and the intranucleolar chromatin
The size and structure of the nucleolus, and the appearance of the intranucleolarchromatin (i.e. the secondary constrictions of the four nucleolus-organizing polytenechromosomes) were studied in plants kept for 2 days or longer at the 3 temperaturelevels noted above. In addition, temperature shocks of 35 °C were given for 4 h, and of45°-5O °C for 2 h.
The size of the main nucleoli increases with temperature, except at extreme levels.The nucleoli in plants cooled at i2°/8 °C for 2 days are unexpectedly large, occasionallyshowing extrusions along the chromosomes. It seems that material is squeezed out bythe condensing chromosomes and unites with the nucleolus. If the chilling time isextended beyond 10 days the nucleolus shrinks to a spherical structure of less than halfthe normal size. The nucleolar structure, characterized by a dense granular matrix andbright vacuoles at optimal temperatures, changes, with apparent loss of material, andat the end the nucleoli look like empty vacuoles. By a careful warming of the plants thestructure reverts to the original form. First, after some minutes, dark spots becomevisible within the nucleolus. Those spots and the nucleolus itself increase in size and,after some hours, the original picture is restored. On the other hand, there is a constantincrease of the nucleolar size between 12 °C and 27 °C. Under the standard conditions(27722 °C), the nucleoli produce many lobes and extrusions, and many sphericalparticles are extruded as free nucleoli into the karyoplasm. A temperature shock of35 °C for 4 h leads to the extrusion of numerous particles (Fig. 17). These extrusions

Polytene chromosomes in Phaseolus 91
again extrude material, lying as free bodies in the nucleus. Several vacuoles of the mainnucleolus join to form large central vacuoles. Finally, a temperature shock of 45 °C or50 °C for 2 h causes the disappearance of the granular cortex of the main nueleoli,
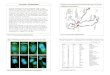
NucleolusVacuole
Satellite —
Secondaryconstriction
Centromere
Extrusion X°
3S°C
10//m
45 °C
Fig. 1. P. cocdneus and P. vulgaris, suspensor. Semi-diagrammatic figures of the nucleo-lus organizer, the secondary constriction, and the satellite of polytene chromosome I atdifferent temperatures. At 12 °C (day) and 8 °C (night), and following a temperatureshock of 45 °C, the chromosomal elements involved in the formation of the nucleolusare condensed; at 27°/22 °C, and following a shock of 35 °C, they are dispersed('puffed'). Note the different appearance of the secondary constriction at all tem-peratures. The right arm of the chromosome and the nucleolus are incompletelydrawn.
which become very translucent and small. The nueleoli again appear as emptyvacuoles. The effect of recooling was not investigated, but the nueleoli of plants whichwere allowed to grow at 2']°J22 °C for some days following a temperature shock showedthe same structure as those without a shock treatment.
The intranucleolar chromatin behaves in a similar manner to the nucleolus-

92 W. Nagl
associated regions of the polytene chromosomes. This chromatin, which constitutes thefibrils of the polytene secondary constrictions, is recognizable as a fibrillar connexionbetween the nucleolus organizers and the satellites following hydrolysis with N HC1,and it is visible in the form of threads within intranucleolar vacuoles in unhydrolysedpreparations. If the temperature is low (i2°/8 °C), these threads are thick and compact(Fig. 12). At higher temperatures (i5°/i2 °C and 27°/22 °C) the threads separate intoseveral thinner fibrils showing a fuzzy appearance (Fig. 13). Four hours after raisingthe temperature to 35 °C the intranucleolar chromatin has a disperse-chromomericappearance (Fig. 14). This may be the obvious expression of a puffed state of thischromatin. On the other hand, a temperature shock of 45 °C or 50 °C for 2 h inducesan entirely different structure. A marked condensation of the disperse chromatin to asingle thread per nucleolus-organizing chromosome occurs. These threads are mostlybanded (Fig. 15). High temperatures, therefore, change the structure of the nucleolus-related parts of the polytene chromosomes in a similar way as does low temperature.The structural changes described are reversible within a period of some hours; theyare summarized in Fig. 1.
In all experiments some plants were found which did not respond in the same way,exhibiting no significant structural change of the nucleolus-organizing chromosomes.The explanation of this requires further information about the relationship betweenchromosomal structure and function, which is not at present available.
A temperature-induced lampbrush state of the polytene chromosomes
In P. coccineus a conspicuous structural change of the polytene chromosomes oc-curred after a temperature elevation from i2°/8 °C to 22 °C, or from 22°/i5°C to300 C (and also at other temperature levels) in over half of some hundreds of cellsexamined. More rarely, a similar structural change was observed after the increase ofthe temperature correlated with the onset of day within the phytotrons. The suspensorcells were fixed at intervals of 10 min during the first hour after temperature elevation,and at every full hour subsequently up to 5 h.
During the first 20 min after temperature increase, all the polytene chromosomes ofthe temperature-sensitive cells develop narrow loops, at right angles to their axes, firstin distinct euchromatic regions (Fig. 2), later along the whole chromosome length, sothat the loops finally appear as a dense veil of processes radiating from the chromo-somes (Figs. 19, 21) and forming a dense fibrillar network within the nuclei (Fig. 18).The analysis of the loops is possible only after isolating the individual polytene chro-mosomes by squashing them out of the nuclei. In such preparations it can be seen thatthe chromosomes split up distally into fibrils with a simple structure; these fibrils areevidently oligotene fibres and single chromosomes of the polytene bundle. The lateralprojections show a duplex nature and are closed at their ends, i.e. they are real loops(Figs. 2, 22). The length of the loops varies from 2 to 25 /tm (their total length is doublethis), and their diameter varies from approximately 0-25 to 0-40 /tm. The duration ofthis temperature-induced lampbrush state amounts to 2 or 3 h; projections fromheterochromatic portions, not identified as loops, are recognizable for longer. Owingto the difficulty in distinguishing the individual giant chromosomes during the lamp-

Polytene chromosomes in Phaseolus 93
brush state, it has not so far been possible to distinguish between structures which are aconsequence of the rapid development of the loops, and those which may be an ex-pression of different activity in specific chromosomal loci. For the present it seemscertain that the loops start by uncoiling of the outer chromomeres of the polytenechromosomes, because the thickness of the chromosome axes corresponds to the num-ber of loops occurring. About 4 h after the onset of the temperature increase, most ofthe loops are retracted (Fig. 20).
10/<m
Fig. 2. Camera lucida drawing of the right arm of polytene chromosome I I I of P . coc-cineus in the early lampbrush state (10 min after temperature increase from 15° to30 °C); optical longitudinal-section.
Fig. 3. Schematic diagram to illustrate the development of a nucleolus-like body on anetwork formed by the loops of a suspensor polytene chromosome of P. coccineus(longitudinal section through the polytene chromosome; pc — diameter of the poly-tene chromosome, of which only the outer four sister chromosomes are drawn) : A, aloose band consisting of sister chromomeres ; B, the outermost chromomere uncoils toform a loop; c, the end of the loop forms a network and starts the synthesis of droplets ,while a second chromomere of the same band is uncoiling; D, the synthesized dropletsare joining to form a nucleolus-like body; E, a nucleolus-like body is finished; F , thenucleolus-like body (vacuolated) disengages completely from the network; and G, thealmost restored band.

94 W. Nagl
A morphologically conspicuous phenomenon indicating a high chromosomalactivity is the production of hundreds of droplets and particles by these lampbrushpolytene chromosomes, 20-40 min after their formation (Fig. 24). Moreover, the finalparts of some loops undergo a transformation to spherical networks, which becomeincreasingly loaded with droplets (Fig. 25). Evidently, the droplets join to form dense,nucleolus-like bodies, the magnitude of which is generally smaller than that of the' additional nucleoli' dealt with in the next section. They never produce extrusions, butdisengage themselves completely from the synthesizing loops. There remain behindonly the empty networks (Fig. 26). This behaviour of loops is shown schematically inFig. 3. The droplets and nucleolus-like bodies migrate to the nuclear boundary anddisappear in a short time.
The formation of loops is not inducible in every cell or plant, but in only about 60%.Evidently more factors are involved in the structural change of polytene chromosomesthan temperature alone.
Nucleolar extrusions and additional nucleoli
Nucleolar extrusions are spherical particles, ejected from the main and additionalnucleoli (Figs. 16, 17). The term 'additional nucleolus' is used in this paper fornucleolar bodies seeming to originate on heterochromatic elements of the polytenechromosomes at sites other than the nucleolus organizers (Figs. 10, 11). Both the ex-trusions and additional nucleoli are vacuolated like the main nucleolus. Chromatic dotsor filaments are visible within some of the vacuoles of additional nucleoli, indicatingthat the chromatin is not in the condensed state (Fig. 11). The number of nucleolarextrusions and also that of additional nucleoli increases from 1 to 3 in nuclei of cooledplants up to 20-40 at optimal temperatures.
The loci where additional nucleoli were found are as follows. At temperatures be-tween 150 and 35°C, one pair of polytene chromosomes in P. vulgaris always possessadditional nucleoli on their distal heterochromatic band. This pair of chromosomes isthe one which was found to be in the puffed state in most of the cells (Nagl, 19696).The possibility that these are a third pair of nucleolus-organizing chromosomes isimprobable, because their additional nucleoli unite with the main nucleolus very in-frequently (in less than 1 % of cells). Other polytene chromosomes of both species,among them also the nucleolus-organizing chromosomes, possess additional nucleolion the proximal heterochromatin (i.e. the region of the centromere), or on any band.The additional nucleolus was seen suspended on a heterochromatic filament origina-ting from a heterochromatic portion of the chromosome in several other cells (Fig. 10).
Staining properties of the polytene chromosomes and nucleoli
The Feulgen reaction indicates that all parts of the polytene chromosomes containDNA, as does the secondary constriction. This, the intranucleolar chromatin, is moreintensely stained the more it is condensed. The same is valid for staining of basicproteins with fast green at pH 8-2. It may be assumed, therefore, that the secondaryconstriction consists of a DNA-histone-complex, as do the other parts of the chromo-somes. The loops of the lampbrush state are not individually visible in Feulgen- or'

Polytene chromosomes in Phaseolus 95
fast green-stained preparations. In the former, the chromosomes possess a pinkfibrillar halo, indicating that the loops contain DNA. The individual loops are notrecognizable because the matrix is dissolved by hydrolysis. On the other hand, theformalin-fast green method preserves too much of the cytoplasm, thereby preventingdetailed analysis of the polytene chromosomes. An appearance comparable to that ofFeulgen-stained lampbrush polytene chromosomes, but somewhat more distinct, isgiven by staining the hydrolysed cells with toluidine blue (Fig. 21).
The RNA content of nucleoli was tested with methyl green-pyronin in combinationwith ribonuclease. This method does not preserve the chromosomal structure verywell, since swelling occurs. The polytene chromosomes stain violet, or, in severalpreparations, in a blue and violet pattern of banding. Following the treatment withribonuclease they stain green-blue. The main nucleoli and additional nucleoli of cellsnot treated with ribonuclease stain red, but the droplets and nucleolus-like bodies ofthe lampbrush stage stain only pink. If the cells are pretreated with ribonuclease, allthe nucleoli and nucleolus-like particles are preserved in form, but they appear to beunstained. It was generally noticed that the nucleolar extrusions and nucleolus-likebodies stain intensely with fast green at pH 5-0, and with acid fuchsin at pH 7-0. Thesereactions may indicate high contents of proteins, especially the acid ones. The mainnucleolus, on the contrary, stains more intensely with fast green at pH 8-2, which maysignify a high content of basic proteins.
Structural changes during the degeneration of the suspensor cells
All the structures described differ significantly from those seen at the onset of de-generation of the suspensor cells. When this occurs, the nucleoli dissolve, and thepolytene chromosomes disintegrate into longitudinal fibrils, which are evidently thesingle sister chromosomes which formed the polytene bundles (Fig. 23). Sometimesthey degenerate to pycnotic elements. Of course, the structural changes occurring indegenerating cells are in no way reversible.
DISCUSSION
The influence of temperature on the gross morphology of polytene chromosomes hasbeen known since the early studies of Diptera (Bridges, 1936; Schultz, 1936; Wolf,1957; Bier, 1958, 1959)- The chromosomes are more compact at low temperatures,and they are more diffuse at higher ones. Recently, a similar effect has been found in thepolytene chromosomes of the suspensor of P. vulgaris (Nagl, 1969 a). A more specificeffect was obtained with the aid of temperature shocks in Drosophila and Rhynchosciara:the induction of puffs, and the increased RNA synthesis in these puffs (Berendeset al. 1965; Pavan, 1965; Berendes, 1968). This gives evidence of temperature-dependent gene activity.
In the course of earlier investigations in Phaseolus an inexplicable structuralvariability of the giant chromosomes was noted (Nagl, 1965, 1967). The aim of thepresent study was to demonstrate that one of the factors influencing the detailedstructure of these plant polytene chromosomes may also be temperature. The nucleolus-

96 W. Nagl
organizing chromosomes have been found to be the most sensitive. At optimal tem-peratures, especially following a temperature shock of 35 °C, a 'puffing' of theintranucleolar and nucleolus-associated chromatin occurs, whereas this is extremelycondensed at too low or too high temperatures. Besides these reversible structuralmodifications, the size of the nucleolus changes with temperature: in general, it islargest at optimal temperature (compare Vogt-Kohne (1961) for Diptera), and in theseconditions the extrusion of numerous particles is recognizable. This extrusion pheno-menon has also been observed in other nuclei and interpreted as evidence of nuclearactivity (Gottschalk, 1951; Tschermak-Woess & Enzenberg-Kunz, 1965; Johnson &Jones, 1967; Kohlenbach, 1967). Since the injection of actinomycin D into the embryo-sac leads to a condensation of nucleolus-organizing polytene chromosomes and to adiminution of the nucleolar size similar to that produced by adverse temperatures(W. Nagl, unpublished observations), and since it is known that the synthetic activity ofcondensed chromatin is much less than that of diffuse chromatin, we conclude thatthere are temperature-dependent functional structures in the polytene chromosomes.
The formation of the nucleolus seems to be a result of the activity of the nucleolusorganizers (McClintock, 1934) and of the intranucleolar secondary constrictions(Heitz, 1931; Kaufmann, 1934; Kahn, 1962; Hsu, Brinkely & Arrighi, 1967). Theoccurrence of conspicuous chromatin structures within nucleoli has been reported alsoin other cells (Granboulan & Granboulan, 1964; Lettre, Siebs & Paweletz, 1966; LaCour, 1966), especially from salivary gland cells of Diptera (Heitz & Bauer, 1933;Kaufmann, 1938; Mechelke, 1953; Beermann, i960; Rodman, 1968); a single ex-ception was noted in Smittia, in which the chromatin of the nucleolus organizer is notdistinct from that in other bands (Jacob & Sirlin, 1964).
A specific' puffing' was hitherto found in only one pair of the polytene chromosomesof Phaseolus (Nagl, 19696). However, there are additional nucleoli at many chromo-somal sites other than the nucleolus organizers. These particles may indicate a syn-thesizing or storing function of several loci. Whether or not they are comparable withthe nucleolar bodies visible in other cells, and, particularly, in dipteran giant chromo-somes (see Beermann, i960; Swift, 1962; Jacob & Sirlin, 1963; Gabrusewycz-Garcia &Kleinfeld, 1966; Pelling & Beermann, 1966), cannot be decided at present. Like thelatter, they originate on heterochromatic parts of the polytene chromosomes, but evi-dently on loosened fibrils, because they possess threads or granules of intranucleolarchromatin (compare Pavan, 1965; Gabrusewycz-Garcia & Kleinfeld, 1966).
A very conspicuous functional structure is inducible within the greater part of thesuspensor cell nuclei of P. coccineus by a temperature increase of about 15 °C. Afterthis, the whole polytene complement assumes a lampbrush state for a period of 3-4 h.Generally, lampbrush chromosomes are meiotic chromosomes with projecting lateralloops, and probably occur in all oocytes (Gall, 1954; Callan, 1957, 1966; Kunz, 1967)and spermatocytes (Ris, 1945; Nebel & Hackett, 1961; Meyer, Hess & Beermann,1961). Like the puffs of polytene chromosomes, the loops are interpreted as an un-coiling of the chromosome fibres, which are in a condensed state within the chromo-meres (Callan, 1963; Gall, 1956). The hitherto described meiotic lampbrushchromosomes of plants have a fuzzy appearance, but loops were not identified (Grun,

Polytene chromosomes in Phaseolus 97
1958; Lu, 1967; Peveling, 1967). The polytene chromosomes of P. coccineus, on thecontrary, develop distinct loops, evidently by the uncoiling of the superficial chromo-meres. It seems that there is no essential difference between the chemical compositionof the loops in Phaseolus and in animals. The Feulgen reaction indicates DNA in theloops of the suspensor chromosomes and the presence of DNA has also been demon-strated for the loops in oocytes (Alfert, 1954; Callan & Macgregor, 1958). The loops inPhaseolus produce nucleolus-like bodies and droplets, believed to contain RNA on thebasis of the methyl green-pyronin method; RNA was also demonstrated in the matrixof animal loops (Gall, 1954; Gall & Callan, 1962; Macgregor & Callan, 1962). How-ever, more studies will be necessary to elucidate the function of this lampbrush state ofthe plant polytene chromosomes. For the present, it seems certain that the loops arenot an artefact, and they are distinct, for example, from the fuzzy appearance of dip-teran giant chromosomes following NaOH treatment (Painter, 1941). Apart from thefact that they have been seen after good fixation, the polytene chromosomes of Phaseolusdisintegrate into longitudinal fibrils during degeneration (Fig. 23) and following chemi-cal treatments (W. Nagl, unpublished).
The fact that the structures described are not inducible in all nuclei might be ex-plained by genetical differences between different plants, and by physiological anddevelopmental differences between single cells. Moreover, temperature may be one factoronly within a very complex regulation system. A high degree of variability of nuclearstructure without obvious cause is one of the unexplained phenomena in the study ofnuclei in general (Tschermak-Woess, 1963; Geitler, 1965). However, the presentfindings may indicate a possible direction for further research.
REFERENCESALFERT, M. (1954). Composition and structure of giant chromosomes. Int. Rev. Cytol. 3,
I3I-I75-ALFERT, M. & GESCHWIND, I. (1953). A selective staining method for the basic proteins of cell
nuclei. Proc. natn. Acad. Sci. U.S.A. 39, 991-999.BEERMANN, W. (i960). DerNukleolusalslebenswichtigerBestandteil des Zellkerns. Chromosoma
11, 263-296.BERENDES, H. D. (1968). Factors involved in the expression of gene activity in polytene chromo-
somes. Chromosoma 24, 418-437.BERENDES, H. D., BREUGEL, F. M. A. VAN & HOLT, T. K. H. (1965). Experimental puffs in
salivary gland chromosomes of Drosophila hydei. Chromosoma 16, 35-46.BIER, K. (1958). Beziehungen zwischen Wachstumsgeschwindigkeit, endometaphasischer
Kontraktion und der Bildung von Riesenchromosomen in den Nahrzellkernen von Calliphora.Z. Naturf. 13 b, 85-93.
BIER, K. (1959). Quantitative Untersuchungen iiber die Variabilitat der NShrzellkernstrukturund ihre Beeinflussung durch die Temperatur. Chromosoma 10, 619-653.
BRACHET, J. (1953). The use of basic dyes and ribonuclease for the cytochemical detectionof ribonucleic acid. Q. Jl microsc. Sci. 94, 1-10.
BRIDGES, C. B. (1936). Current methods for permanent carmine smears. Drosoph. Inf. Serv.6 > 3 i -
CALLAN, H. G. (1957). The lampbrush chromosomes of Sepia officinalis, Anilocra physodes, andScyllium catulus, and their structural relationship to the lampbrush chromosomes of Amphibia.Pubbl. Staz. zool. Napoli 29, 329-346.
CALLAN, H. G. (1963). The nature of lampbrush chromosomes. Int. Rev. Cytol. 15, 1-34.7 C E L 6

98 W. Nagl
CALLAN, H. G. (1966). Chromosomes and nucleoli of the axolotl, Ambystoma mexicanum.J. CellSci. i, 85-108.
CALLAN, H. G. & MACGREGOR, H. C. (1958). Action of deoxyribonuclease on lampbrushchromosomes. Nature, Lond. 181, 1479-1480.
ENZENBERG, U. (1961). Beitrage zur Karyologie des Endosperms. Ost. bot. Z. 108, 245-285.ERBRICH, P. (1965). Ober Endopolyploidie und Kernstrukturen in Endospermhaustorien. Ost.
bot. Z. 112, 197-262.GABRUSEWYCZ-GARCIA, N. & KLEINFELD, R. G. (1966). A study of the nucleolar material in
Sciara coprophila. J. Cell Biol. 29, 347-359.GALL, J. G. (1954). Lampbrush chromosomes from the oocyte nuclei of the newt._7. Morph. 94,
283-339-GALL, J. G. (1956). On the submicroscopic structure of chromosomes. Brookhaven Symp. Biol.
8, 17-32-GALL, J. G. & CALLAN, H. G. (1962). H3-uridine incorporation in lampbrush chromosomes.
Proc. natn. Acad. Sci. U.S.A. 48, 562-570.GEITLER, L. (1965). Riesenchromosomen bei Pflanzen. Forsch. Fortschr. 39, 295-298.GOTTSCHALK, W. (1951). Das Verhalten der Nukleolen wahrend der Meiosis von Solatium
lycopersicum. Chromosoma 4, 502-520.GRANBOULAN, N. & GRANBOULAN, P. (1964). Cytochimie ultrastructurale du nucl^ole. I. Mise
en evidence de chromatine a l'inte>ieur du nucteole. Expl Cell Res. 34, 71-87.GRUN, P. (1958). Plant lampbrush chromosomes. Expl Cell Res. 14, 619-621.HASITSCHKA-JENSCHKE, G. (1959). Vergleichende karyologische Untersuchungen an Antipoden.
Chromosoma 10, 229-267.HASITSCHKA-JENSCHKE, G. (1962). Notizen iiber endopolyploide Kerne im Bereich der Samen-
anlage von Angiospermen. Ost. bot. Z. 109, 125-137.HEITZ, E. (1931). Die Ursache der gesetzmStSigen Zahl, Lage, Form und GroCe pflanzlicher
Nukleolen. Planta 12, 775-844.HEITZ, E. & BAUER, H. (1933). Beweise fur die Chromosomennatur der Kernschleifen in den
Knauelkernen von Bibio hortulans L. Z. Zellforsch. mikrosk. Anat. 17, 67-82.Hsu, T. C , BRINKLEY, B. R. & ARRIGHI, F. E. (1967). The structure and behaviour of the
nucleolus organizers in mammalian cells. Chromosoma 23, 137—153.JACOB, J. & SIRI.IN, J. L. (1963). Electron microscope studies on salivary gland cells. I. The
nucleus of Bradysia mycorum F. (Sciaridae), with special reference to the nucleolus. J. CellBiol. 17, 153-165.
JACOB, J. & SIRLIN, J. L. (1964). Electron microscope studies on salivary gland cells. IV. Thenucleus of Smittia parthenogenetica (Chironomidae), with special reference to the nucleolusand the effects of actinomycin thereon. J. Ultrastruct. Res. 11, 315-328.
JOHNSON, J. M. & JONES, L. E. (1967). Behavior of nucleoli and contracting nucleolar vacuolesin tobacco cells growing in microculture. Am. J. Bot. 54, 189-198.
KAHN, J. (1962). The nucleolar organizer in the mitotic chromosome complement of Xenopuslaevis. Q. Jl microsc. Sci. 103, 407-409.
KAUFMANN, B. P. (1934). Somatic mitoses of Drosophila melanogaster. J. Morph. 56,125-155-
KAUFMANN, B. P. (1938). Nucleolus-organizing regions in salivary gland chromosomes ofDrosophila melanogaster. Z. Zellforsch. mikrosk. Anat. 28, 1-11.
KOHLENBACH, H. W. (1967). Nucleoli mit ' pulsierenden Vakuolen' bei kultivierten Rhoeo-Epidermiszellen. Z. PflPhysiol. 58, 187-190.
KUNZ, W. (1967). Lampenburstenchromosomen und multiple Nukleolen bei Orthopteren.Chromosoma 21, 446-462.
LA COUR, L. F. (1966). The internal structure of nucleoli. Chromosomes Today 1, 150-160.LETTRE, R., SIEBS, W. & PAWELETZ, N. (1966). Morphological observations on the nucleolus of
cells in tissue culture, with special regard to its composition. Natn. Cancer Inst. Monogr. 23,107-123.
Lu, B. C. (1967). The course of meiosis and behaviour during the ascus development of theAscomycete Gelasinospora calospora. Chromosoma 22, 210-226.
MACGREGOR, H. C. & CALLAN, H. G. (1962). The actions of enzymes on lampbrush chromo-somes. Q. Jl microsc. Sci. 103, 173-203.

Polytene chromosomes in Phaseolus 99
MCCLINTOCK, B. (1934). The relation of a particular chromosomal element to the developmentof the nucleoli in Zea mays. Z. Zdlforsch. mikrosk. Anat. 21, 294-328.
MECHELKE, F. (1953). Reversible Strukturmodifikationen der Speicheldrusenchromosomen vonAcricotopus lucidus. Chromosoma 5, 511-543.
MEYER, G. F., HESS, O. & BEERMANN, W. (1961). Phasenspezifische Funktionsstrukturen in denSpermatocytenkernen von Drosophila melanogaster und ihre Abhangigkeit vom Y-Chromo-som. Chromosoma 12, 676-716.
NAGL, W. (1962a). 4096-Ploidie und ' Riesenchromosomen' im Suspensor von Phaseoluscoccineus. Natunvissenscliaften 49, 261-262.
NAGL, W. (19626). Ober Endopolyploidie, Restitutionskernbildung und Kernstrukturen imSuspensor von Angiospermen und einer Gymnosperme. Ost. hot. Z. 109, 431-494.
NAGL, W. (1965). Die SAT-Riesenchromosomen der Kerne des Suspensors von Phaseoluscoccineus und ihr Verhalten wahrend der Endomitose. Chromosoma 16, 511-520.
NAGL, W. (1967). Die Riesenchromosomen von Phaseolus coccineus L.: Baueigenttimlichkeiten,Strukturmodifikationen, zusatzliche Nukleolen und Vergleich mit den mitotischen Chromo-somen. Ost. bot. Z. 114, 171-182.
NAGL, W. (1969a). Banded polytene chromosomes in the legume Pliaseohis vulgaris. Nature,Lond. 221, 70-71.
NAGL, W. (19696). Puffing of polytene chromosomes in a plant {Phaseolus vulgaris). Natunvis-senscliaften 56, 221-222.
NAGL, W. (1969c). Inhibition of polytene chromosome formation in Phaseolus by polyploidmitoses. Cytologia (in the Press).
NEBEL, B. R. & HACKETT, E. M. (1961). Lampbrush fine structure of vertebrate male chromo-somes in meiotic prophase. Natunuissenscliaften 48, 655.
PAINTER, T H . S. (1941). An experimental study of salivary gland chromosomes. Cold SpringHarb. Symp. quant. Biol. 9, 47—54.
PAVAN, C. (1965). Nucleic acid metabolism in polytene chromosomes and the problem of dif-ferentiation. Brookhaven Symp. Biol. 18, 222-241.
PELLING, C. & BEERMANN, W. (1966). Diversity and variation of the nucleolar organizing re-gions in Chironomids. Natn. Cancer Inst. Monogr. 23, 393-409.
PEVELING, E. (1967). Elektronenmikroskopische Untersuchungen an Zellkernstrukturenwahrend der Antherendifferenzierung. Ber. dt. bot. Ges. 8o, 303-304.
RlS, H. (1945). The structure of meiotic chromosomes in the grasshopper and its bearing on thenature of 'chromomeres' and 'lampbrush chromosomes'. Biol. Bull. mar. biol. Lab., WoodsHole 89, 242-257.
RODMAN, T. C. (1968). Intranucleolar DNA of polytene chromosomes. J. Cell Biol. 39, 113 A.SCHULTZ, K. (1936). Notes on methods for salivary chromosomes. Drosoph. Inf. Serv. 6, 35.SWIFT, H. (1962). Nucleic acids and cell morphology in Dipteran salivary glands. In The Mole-
cular Control of Cellular Activity (ed. J. M. Allen), pp. 73-125. New York: McGraw-Hill.TSCHERMAK-WOESS, E. (1956). Notizen uber die Riesenkerne und ' Riesenchromosomen' in den
Antipoden von Aconitum. Chromosoma 8, 114-134.TSCHERMAK-WOESS, E. (i957). Ober das regelmaBige Auftreten von 'Riesenchromosomen' im
Chalazahaustorium von Rliinanthus. Chromosoma 8, 523—544.TSCHERMAK-WOESS, E. (1963). Strukturtypen der Ruhekerne von Pflanzen und Tieren. Proto-
plasmatologia V/i . Wien and New York: Springer.TSCHERMAK-WOESS, E. & ENZENBERC-KUNZ, U. (1965). Die Struktur der hochendopolyploiden
Kerne im Endosperm von Zea mays, das auffallende Verhalten ihrer Nukleolen und ihrEndopolyploidiegrad. Planta 64, 149-169.
VOGT-KOHNE, L. (1961). Quantitative cytochemische Untersuchungen an Nukleolen ausSpeicheldrUsenkernen von Chironomus thummi. Chromosoma 12, 382-397.
WOLF, B. E. (1957). Temperaturabhangige Allozyklie des polyt&nen X-Chromosoms in denKernen der Somazellen von Phryne cincta. Chromosoma 8, 396-435.
(Received 13 January 1969—Revised 16 May 1969)
7-2

ioo W. Nagl
Fig. 4. P. coccineus, highly endopolyploid suspensor cell with polytene chromosomes.Note the size difference between this and the cells and nuclei of the integument andendosperm, which are respectively diploid and triploid. Original relationships of cellsaltered by the squash technique. Dark-field, x 160.Fig. 5. P. coccineus, vacuolated nucleolar lobes of a highly endopolyploid suspensor cellnucleus, at 27 °C. Arrows indicate a divided satellite. Phase contrast, x 1600.

Polytene chromosomes in Phaseolus i o r

io2 W. Nagl
Figs. 6-11. P. coccineus, suspensor. Phase contrast, x 1600.Fig. 6. Nucleolus organizer of polytene chromosome I, splitting up at 27°/22 °C.
Arrow indicates the centromere.Fig. 7. Satellite of polytene chromosome I, ramified. 27°/22 °C.Fig. 8. Nucleolus organizer of polytene chromosome I, condensed. i2°/8 °C. The
arrow indicates the centromere.Fig. 9. Satellite of polytene chromosome I, compact. i2°/8 °C.Fig. 10. Small additional nucleolus (arrow) originating on a heterochromatic filament
starting from a polytene chromosome. 27°/22 °C.Fig. 11. Additional nucleolus originating from the distal band of a polytene chromo-
some. 27°/22 °C. Note the chromatic areas within the vacuoles (arrows).

Polytene chromosomes in Phaseolus xsg

104 W. Nagl
Figs. 12-17. P- vulgaris, suspensor. Phase-contrast, x 1600.Fig. 12. Part of the condensed secondary constriction of a polytene chromosome
visible within an intranucleolar vacuole (arrow). i2°/8 °C.Fig. 13. Loosened threads of the secondary constriction with a fuzzy appearance,
visible within an intranucleolar vacuole. 27°/22 CC.Fig. 14. Gigantic vacuole of a nucleolar lobe (indicated by arrows). The 'puffed'
secondary constriction is visible as dispersed chromatin within the vacuole. Followinga temperature shock of 35 °C.
Fig. 15. Part of a condensed secondary constriction with a banded appearance. Fol-lowing a temperature shock of 50 CC.
Fig. 16. Additional, free nucleolus with an extrusion. 27°/22 °C.Fig. 17. Main nucleolus of a low endopolyploid cell, with many nucleolar extrusions.
Following a temperature shock of 35 °C.

Polytene chromosomes in Phaseolus

io6 W. Nagl
Figs. 18-26. P. coccineus, suspensor. Phase-contrast, except Fig. 21. x 1200, exceptFig. 22 which is x 240x3.
Fig. 18. Part of a nucleus with polytene chromosomes in the lampbrush state (arrows).Two hours after temperature increase from 150 to 30 °C.
Fig. 19. Lampbrush polytene chromosome, squashed out from the nucleus; 2 h aftertemperature increase from 15° to 30 °C.
Fig. 20. Late lampbrush state of a polytene chromosome in which most of the loopsare retracted; 4 h after temperature increase from 15° to 30 °C.
Fig. 21. Toluidine blue-stained polytene chromosome in the lampbrush state. Theloops appear as a veil around the chromosome (arrow). The cell was hydrolysed for30 min before staining.
Fig. 22. Higher enlargement of a single loop (arrow).
Fig. 23. Degenerating polytene chromosome, to demonstrate the differences from thelooped state. Note the longitudinal disintegration.
Fig. 24. Droplets, occurring above lampbrush polytene chromosomes.
Fig. 25. Filling up of networks formed by loops, to form nucleolus-like bodies(arrows).
Fig. 26. Empty networks (arrows), and a disengaged nucleolus-like body (doublearrow).

Polytene chromosomes in Phaseolus 107