Embed Size (px)
Citation preview

J. Cell Sri. 64, 255-264 (1983) 255Printed in Great Britain © The Company of Biologists Limited 1983
TOROIDAL BANDS IN POLYTENE CHROMOSOMESOF DROSOPHILA
VEIKKO SORSADepartment of Genetics, University of Helsinki, Finland
SUMMARY
Results obtained from the thin-section electron microscopy of salivary gland chromosomes ofDrosophila melanogaster mainly support the concept of cable-like organization of polytenechromosomes, with disk-like bands composed of parallel bundles of homologous chromomeres.Outward orientation of loop fibres may generally cause a toroidal bending in the chromomerebundles. Both longitudinal and transverse sections of polytene chromosomes indicate that the bandsmay contain toroidal subunits. Torus-shaped bands were only found in thin sections of the mostdistal and most proximal regions, as well as in certain heavy bands at the late-replicating regions ofpolytenized interphase chromosomes. This suggests that an incomplete duplication of chromomeresmay be a reason for torus formation, by preventing the separation of sister chromatids at the earliestphases of the polytenization process. The appearance of more numerous, but smaller, subunits inthin-sectioned faint bands is interpreted as a consequence of more complete segregation of sisterchromatids in those bands during polytenization.
INTRODUCTION
Recently, Mortin & Sedat (1982) reported results suggesting that most if not all thebands of polytene chromosomes of Drosophila melanogaster are torus-shaped. Ac-cording to Painter's (1934) review of the early history of studies of polytenechromosomes, the idea of a toroidal organization of the bands was presented byLeydig two years after Balbiani's first description of these striated ribbons in thesalivary gland cells of Chironomus. Several reports of both disk-like and torus-likeorganization of the bands can be found in the early literature of polytenechromosomes. Some investigators even tried to explain the banding pattern ofpolytene chromosomes as a helical type of organization, which was generally foundto be present in dividing chromosomes, both in mitosis and meiosis (cf. Painter, 1934;and reviews by Bauer, 1935; and Beermann, 1962).
The polyteny concept presented by Koltzoff (1934) also showed a preference fortoroidal organization of bands, while Bridges (1935) depicted the polytenechromosomes as cable-like structures with disk-like bands. Indisputable evidence forthe disk concept of bands was presented by Bauer (1935). He demonstrated clearlythat in cross-sections of fixed and stained polytene chromosomes, as well as in opticalsections of chromosomes in living salivary gland cells, the bands are evidently filledwith groups of parallel chromomeres. According to Bauer (1935) longitudinal sectionsalso provided no evidence to prove the existence of a hollow axis in the polytenizedinterphase chromosomes (cf. Beermann, 1962).
The hollow centre of the chromosomes, and particularly of the bands, if it does

256 V. Sorsa
exist, should be detectable in longitudinal thin sections cut along the long axis ofthe whole polytene chromosome. However, electron micrographs of serial sectionsalong the central axis of salivary gland chromosomes of D. melanogaster, used forthe electron microscopic (EM) revision of Bridges' maps, have not provided us withany evidence in favour of the torus concept as a general mode of organization of thebands (cf. Saura & V. Sorsa, 1979a,b,c,d; Saura, 1980; V. Sorsa & Saura, 1980a,b;V. Sorsa, Saura & Heino, 1983). Similarly, the earlier electron-microscopic observa-tions on polytene chromosomes, fixed and prepared for the EM by using severaldifferent methods, have also supported the disk-like organization of band chromatin(cf. e.g. Swift, 1962; Sorsa & Sorsa, 1967, 1968; M. Sorsa, 1969; Berendes, 1970;Lossinsky & Lefever, 1978; Semeshin, Zhimulev & Belyaeva, 1979; Mott, Burnett& Hill, 1980; ten Tusscher & Derksen, 1982; Zhimulev, Semeshin, Kulichkov &Belyaeva, 1982).
In the present report the possible existence of toroidal bands and their distributionin polytene chromosomes has been studied in a large collection of electronmicrographs taken from thin sections of both unsquashed and squashed salivary glandcells of D. melanogaster. On the basis of the results obtained some hypotheses arepresented for explaining the appearance and possible organization of toroidal bandsin polytene chromosomes.
MATERIALS AND METHODS
Salivary glands of third-instar larvae oiDrosophila melanogaster -were fixed within a few secondsof dissection, using procedures that allow exact comparison of the banding patterns with the revisedreference maps of Bridges (cf. Lindsley & Grell, 1968), also at the electron-microscopic level.Fixative used for this study were the following (v/v): (a) acetic acid/ethanol (1:3), (b) aceticacid/methanol (1:3), (c) formic acid/methanol (1:3), (d) formaldehyde (4%) in insect Ringer.Fixation was carried out in a refrigerator at +4°C and fixation time was approx. 1 h for the acidicfixatives and approx. 15 min for the formaldehyde-Ringer. Some of the acetoethanol-fixed glandswere embedded after dehydration and staining, without squashing. Some material was embeddedafter a light squashing. Most of the salivary glands were squashed on silicon-coated slides and
Fig. la-d. Electron micrographs of thin-sectioned salivary gland chromosomes of D.melanogaster. a. Partially cross-sectioned tip of X chromosome shows toroidal shape ofbands close to the telomere (t). Acetomethanol fixation and uranyl acetate staining, b.Toroidal group of chromatin fibres (arrow) at the chromocentre area close to thecentromere of chromosome 4. c. An axial section of the distal end of 2L chromosomeshowing strongly concave bands close to the tip (t) of chromosome arm. The cup-shapedbands obviously appear as closed toroids in certain types of cross-sections indicated by theparallel lines (cs). Nucleosome fibres (indicated by thin arrows) and some cross-sectionsand side-views of larger coils (thick arrows) can be recognized. Formaldehyde(4%)-Ringer fixation and uranyl acetate staining, d. Cross-section of the heavy doublet 3C 2-3at the proximal end of the inversion zeste+64b9 (stock obtained from M. M. Green) showsclosed toroids in both homologues (white arrows). The torus-shaped doublets 3C 2-3include some DNA from the rightmost end of originally triplicated white+R sublocusattached to the remnants of band 12B 9 at the proximal breakpoint of this inversion. Mostof the chromometric material of ring bands proposedly belongs to the late-replicatingregion demonstrated by Arcos-Teran (1972). Acetomethanol (1: 3) fixation, uranyl acetatestaining, cf. Fig. 3a. Bars, 0-1 ^m.
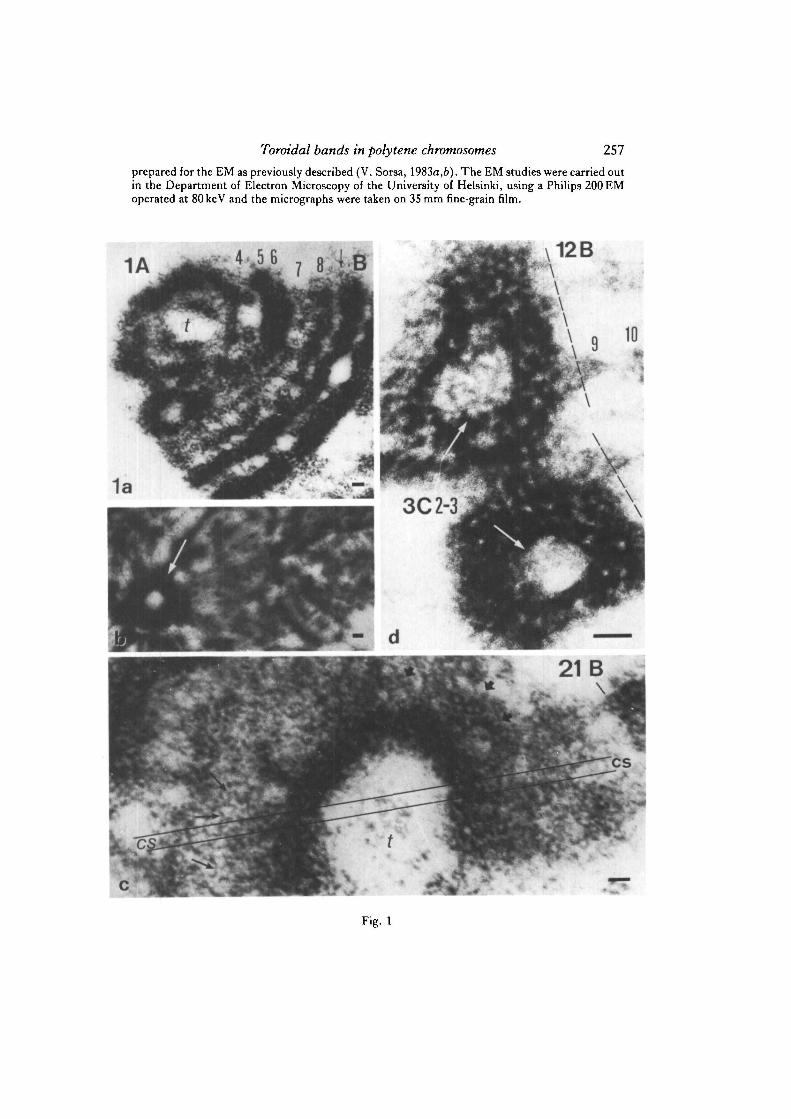
Toroidal bands in polytene chromosomes 257prepared for the EM as previously described (V. Sorsa, 1983a,b). The EM studies were carried outin the Department of Electron Microscopy of the University of Helsinki, using a Philips 200 EMoperated at 80keV and the micrographs were taken on 35 mm fine-grain film.
1A\12B
Fig. 1

258 V. Sorsa
RESULTS AND DISCUSSION
As has been shown at the light-microscopic level (Bridges, 1935), and also inelectron micrographs (Fig. la; 2d, e), most of the less-prominent bands are split intoparallel groups or bundles of homologous chromomeres (cf. also V. Sorsa & Saura,1980a,6; V. Sorsa, I982a,b). The cross-sections of heavy bands in unsquashedpolytene chromosomes (Fig. 2a, b) usually show several hollow centres, suggestingthat those bands are formed by several toroidal subunits. Thin sections of distal endsof chromosome arms show that many bands are strongly concave (Fig. lc), as demon-strated previously by Bauer (1936) (cf. also Beermann, 1962). The cup-shaped bandsmay appear as rings in certain cross-sections, as shown in Fig. lc. The actual shapeof the chromatin rings that appear in thin sections of the proximal ends of polytenechromosomes is difficult to demonstrate (Fig. lb). Occasionally, a torus-like organi-zation of chromatin can be also found in certain heavy bands in the middle ofchromosome arms (Fig. 2c). Clear evidence for the existence of torus-shaped bandscomes from the heavy doublet 3C 2—3 in the X chromosome (Fig. Id). In this frontallysectioned band at the proximal end of the inversion zeste+64b9 the asynapsed groupsof parallel chromatids, originating from the paired parental strands, seem to form twoclosed rings of chromatin. In the less-prominent bands the chromomeres often appearbent or as toroidal groups (Fig. 2d-i). In general, the results of the present study seemto suggest that a toroidal organization of chromatin appears in subunits of bands ratherthan in the entire bands of polytene chromosomes (cf. Sorsa & Sorsa, 1968).
According to the model proposed by Mortin & Sedat (1982), the axial fibres in boththe interbands and bands form a wide hollow cylinder in the polytene chromosome.The chromomeric loops are located mainly peripherally in this tube. The diameter ofthe axial tube would thus be essentially dependent on both the number and thethickness of individual axial fibres in the polytene chromosomes. Simple calculationsfor estimating the diameter of the axial cylinder, for example in a polytenechromosome of D. melanogaster, representing the polyteny degree of 512 strands perhomologue, suggest that the 'hollow axis' model may work only if: (a) the axial fibresare less than 5 nm thick, or (b) the degree of polyteny, at least in the axis, is lower than512 per homologue. For instance, if the interchromomeric fibres are supposed to beapprox. 30 nm thick helices of nucleosomes (Laird, Ashburner & Wilkinson, 1980),the diameter of a hollow axis formed by a single layer of 512 such fibres would be
Fig. 2a-i. Toroidal structures in the thin sections of unsquashed (a and b), slightlysquashed (c) and well-squashed (d-i) salivary gland chromosomes of D. melanogaster.a, b. A general view of a thin section of an unsquashed cell (a) and a higher magnificationof the cross-sectioned heavy band (b) showing four toroidal subunits in a band (whitearrows), c. An open-ended toroid in an unidentified heavy band. Acetoethanol fixation(1: 3), and uranyl acetate staining. Bars: a, 1 /mi; b, c, 0-2^m. d, e. A general view of anaxial thin section through the region 4D-9F of the X chromosome. The borders ofdivisions and subdivisions are marked by thin arrows according to the revised referencemap of X (Bridges, 1938). Circled areas (indicated by arrowheads f-i) are presented aslarger magnifications in f-i showing loop-like organization of chromomeres in manybands, cf. Fig. 3b—c. Bars: d—e, ljUm; f-i, 0-1 (Jin.
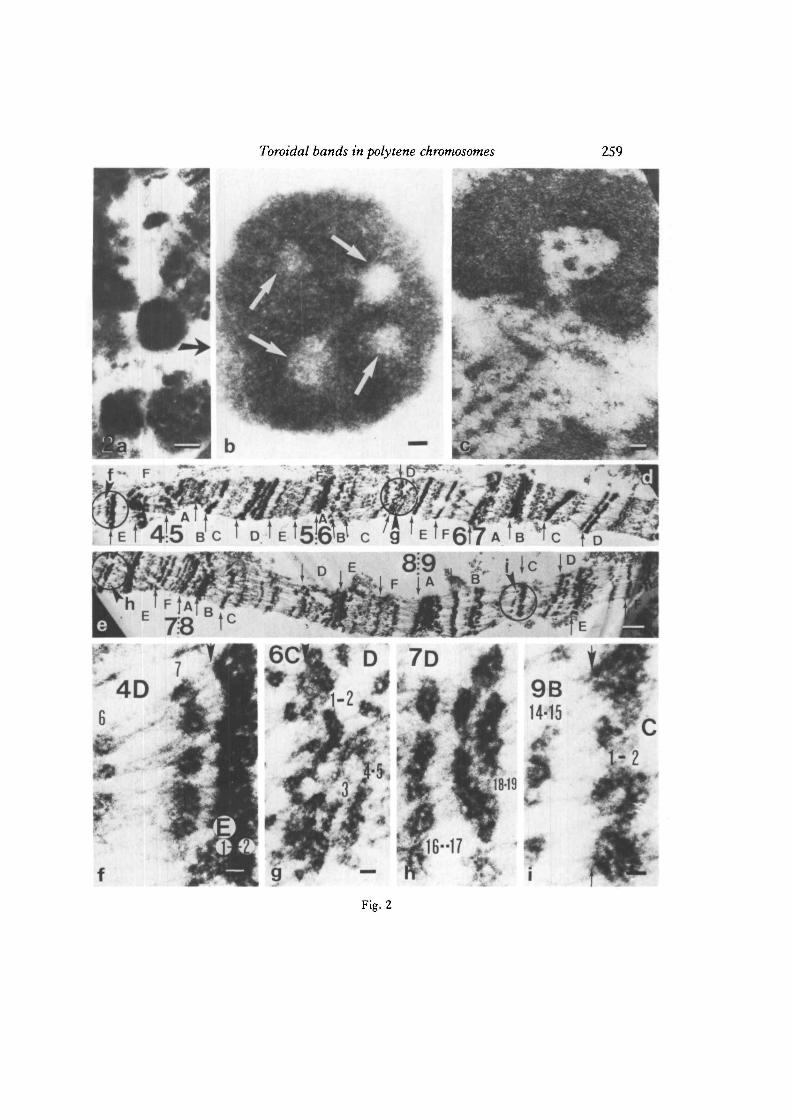
Toroidal bands in polytene chromosomes 259
Fig. 2
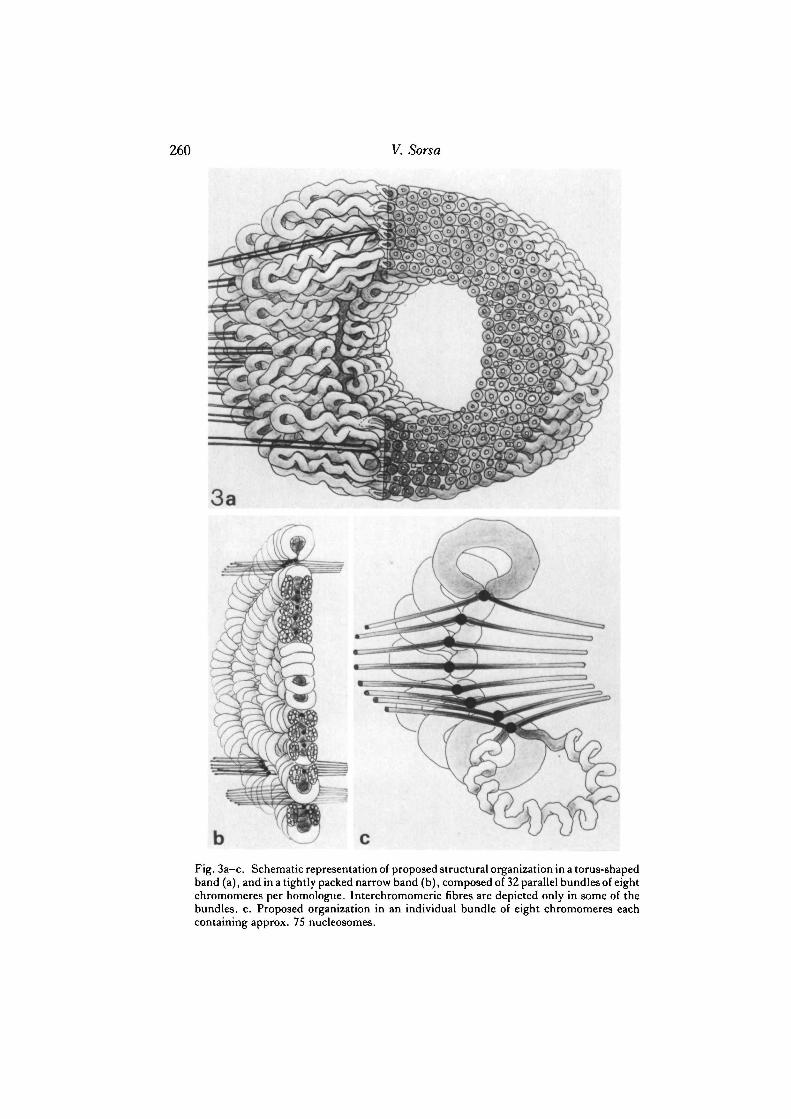
260 V. Sorsa
Fig. 3a—c. Schematic representation of proposed structural organization in a torus-shapedband (a), and in a tightly packed narrow band (b), composed of 32 parallel bundles of eightchromomeres per homologue. Interchromomeric fibres are depicted only in some of thebundles, c. Proposed organization in an individual bundle of eight chromomeres eachcontaining approx. 75 nucleosomes.

Toroidal bands in polytene chromosomes 261
roughly 5/im, even if the axial fibres are placed close together. In the polytenechromosomes flattened by squashing the axial tube alone should be even wider. Anillustration of highly polytenized salivary gland chromosomes of Chironomus, havinga degree of polyteny of approx. 8000-16000 (cf. Beermann, 1962), reveals quiteunreasonable results if the hollow axis model of Mortin & Sedat (1982) is adopted.
Conversely, if the hollow axis model is used for a rough estimation of the polytenydegree, e.g. in the torus-shaped doublet 3C 2-3 shown in Fig. Id, it gives the follow-ing results. In the cross-section of band 3C 2-3 the inner contour length of chromatinrings representing the homologues is approx. 500 nm. According to the structuralmodel proposed by Mortin & Sedat (1982), it corresponds to the axial tube formedby the layer of axial fibres in the polytene chromosomes. The contour length (500 nm)is equal to approx. 250 diameters of DNA double helix or to about 16 diameters of'solenoid-sized' (30 nm) chromatosome helices. According to the thin-section EM ofsquashed polytene chromosomes of Drosophila the average thickness of individualinterchromomeric fibres is about 5 nm (cf. V. Sorsa, 1982a,6). By using this averagediameter (5 nm) for the axial fibres the hollow axis model reveals a degree of polytenybetween 64 and 128 for the toroidal bands (3C 2—3) shown in Fig. Id.
The cross-sectional area of the ring bands 3C 2-3 (Fig. Id) is about 0-2/im2 perhomologue. This value can be used for a rough estimation of the degree of polytenyby comparing the average axial length of the doublet 3 C 2-3, which is approx .0-4 /xmin similarly prepared X chromosomes, with the axial length of the fraction of the sameband at the left end of the inversion, in (1) zeste+64b9 (stock obtained from M. M.Green). This fraction of band 3C 2 includes a complete white"1" locus (cf. V. Sorsa,Green & Beermann, 1973) attached to a small fraction of band 12B 9. Thus the bandcontains at least 15 X 103 bases of DNA in each parallel chromomere. The comparisonshows that the chromatin content of doublet 3C 2—3 is roughly 10 times the chromatincontent of the white"1" locus band in the inversion X. Accordingly, each individualchromomere of the doublet 3C 2-3 should contain at least 150 X 103 bases of DNA,which presumably is capable of forming approx. 750 nucleosome units. The cross-sectional area of such chromomeres is approx. 1700 nm2, if tightly filled with nucleo-some fibre, or approx. 2200 nm2, if filled with 30 nm thick chromatosome helicesformed of six nucleosomes per turn. Thus the maximum number of parallelchromomeres, i.e. the degree of polyteny, in the ring bands shown in Fig. Id is about118-91 per homologue. However, the densitometry of EM negatives representingthin-sectioned band chromatin indicates that the solid chromatin fibres constitute onlyabout 40 % of the total volume of thin sections (V. Sorsa, 19826). This suggests thatthe real chromomere content and, correspondingly, the number of parallelchromatids, if they are completely duplicated, is only about 47—36 per homologue inthe ring bands 3C 2-3 shown in Fig. Id. Arcos-Terdn (1972) has demonstrated thatthe region of the X chromosome including the doublet 3C 2-3 contains late-replicatingDNA, which may explain the estimated low degree of polyteny in these ring bands.It has been proposed that, due to retarded duplication of DNA the polytenizationcycles and segregation of sister chromatids have not been completed in thechromomeric loops of this doublet. 'Normal' polytenization of the distal fragment of

262 V. Sorsa
3C 2-3 containing the white"1" locus at the other end of the inversion indicates thatseveral potential replication origins may exist in the DNA of the doublet 3C 2-3, butreplication is prevented towards the weak point between 3C 2-3 and 3C 5-6. Theappearance of closed or open-ended toroids in the less highly polytenized regions atthe chromocentre and at the telomeres (Fig. la-c), as well as in the late-replicatingregion 3C 2-3 (Fig. Id) suggests that retarded replication of DNA may be a reasonfor the formation of torus-shaped bands. In other words, the existence of a singletoroidal group consisting of all the parallel chromomeres in a polytene chromosomeband indicates that the duplicated sister chromatids have not been able to separatefrom each other even after the earliest cycles of polytenization. Correspondingly, anappearance of four toroidal elements in a band (Fig. 2a, b) may be explained as aconsequence of incomplete segregation of duplicated sister chromomeres after thefour-stranded stage of polytenization etc. Evidently, the greater amount and morerepetitive nature of the DNA may also increase the compactness of those bands or ofsmaller toroidal groups.
Because cross-sections of polytene chromosomes showing an entire disk of parallelchromomeres composing a narrower band are extremely rare in electron micrographsand the identification of cross-sectioned regions is almost impossible until the thin-sectioning methods are refined, the number of parallel bundles and the organizationof fainter bands have been studied only in longitudinal sections. Thus it should beemphasized that only those fixation methods that allow the exact identification ofbands in thin-section EM have been used in this study. This methodological require-ment should not make it impossible to detect the hollow centres of bands if they existgenerally in polytenized chromosomes. The thickness of thin sections (=50— 80 nm)is evidently much lower than the expected diameter of the central holes in bands,which means that the possible central hole should be detectable in several serialsections along the chromosome. Analysis of electron micrographs does not support theexistence of a single 'hollow axis' in polytenized chromosomes. Although a centrallylocated hole can be found in certain prominent bands (e.g. in the subdivisions 5B, 6F,7A, 8C and 9EF in Fig. 2d-e), most probably the explanation is a local asynapsis ofhomologues. However, in certain tight bands, like 7D 1—2, the location of a distinctbreak in longitudinal thin section may be interpreted as the centre of a rather largetoroidal unit, comparable to the structures seen in cross-section in Fig. Id or Fig. 2b.
Excepting the late-replicating regions, the degree of polyteny, i.e. the total numberof parallel chromatids, seems to be roughly constant through long stretches of salivarygland chromosomes, which is also indicated by the rather uniform diameter of thechromosome (cf. Laird, Ashburner & Wilkinson, 1980). The remarkable variationfound in the organization of adjacent bands is seemingly caused by the differentialgrouping of parallel chromomeres. The highest countable number of parallel groupsis usually approx. 25 per thin section, which apparently corresponds to a total numberof approx. 250-500 parallel bundles per band in the whole chromosome, dependingon the degree of flattening of the chromosome region. Because this estimate is lowerthan the expected full polyteny degree (1024) it suggests that the sister chromomeresderived from the last or from the two latest cycles of polytenization have not separated.

Toroidal bands in polytene chromosomes 263
In many bands the chromomeres form even larger groups. In a large number ofBridges' bands the chromatin seems to be so tightly packed that counting of parallelsubunits is impossible.
The essential results of the present study may be summarized as follows. In thin-section electron microscopy of salivary gland chromosomes of D. melanogaster thetoroidal organization of whole bands was found only in the less-polytenized regions.The appearance of ring-shaped bands is proposed to be a consequence of incompleteor retarded duplication of chromomeric loops. This, together with a larger amountand greater degree of repetitiveness of DNA may prevent the segregation of sisterchromomeres and cause the grouping of chromatids even at the early phases ofpolytenization. In fainter bands composed of smaller chromomere units thechromatin consists of a number of separate groups, which is interpreted as aconsequence of easier segregation of duplicated chromatids during the early cycles ofthe polytenization process. Peripheral location of chromomeric loops may cause atoroidal bending also in smaller groups of parallel chromomeres (cf. Fig. 3).
The study was supported financially by the National Research Council of Sciences of Finland.I thank Dr Virpi Virrankoski-Castrodeza, Ms Anja O. Saura, Ph. Lie. and Ms Riikka Santalahti forskilful thin-sectioning of the chromosome material.
REFERENCES
ARCOS-TERAN, L. (1972). DNS-Replikation und die Natur der spat replizierenden Orte inX-Chromosom von Drosophila melanogaster. Chromosoma 37, 233-296.
BAUER, H. (1935). Der Aufbau der Chromosomen aus den Speicheldriisen von Chironomus Thum-mi Kiefer (Untersuchungen an der Riesenchromosomen der Dipteren. I.). Z. Zellforsch. nrikrosk.Anat. 23, 280-313.
BAUER, H. (1936). Structure and arrangement of salivary gland chromosomes in Drosophila species.Proc. natn. Acad. Set. U.SA. 22, 216-222.
BEERMANN, W. (1962). Riesenchromosomen. Protoplasmatologia, 6/D. Wien: Springer Verlag.BERENDES, H. D. (1970). Polytene chromosome structure at the submicroscopic level. I. A map of
region X, 1-4E of Drosophila melanogaster. Chromosoma 29, 118—130.BRIDGES, C. B. (1935). Salivary chromosome maps with a key to the banding of the chromosomes
of Drosophila melanogaster. J. Hered. 26, 60-64.BRIDGES, C. B. (1938). A revised map of the salivary gland X-chromosome of Drosophila
melanogaster. J. Hered. 29, 11-13.KOLTZOFF, N. K. (1934). The structure of the chromosomes in the salivary glands of Drosophila.
Science, N.Y. 80, 312-313.LAIRD, C. D., ASHBURNER, M. & WILKINSON, L. (1980). Relationship between relative dry mass
and the average band width in regions of polytene chromosomes of Drosophila. Chromosoma 76,175-189.
LINDSLEY, D. L. & GRELL, E. H. (1968). Genetic variations of Drosophila melanogaster, publ. 627.Washington, D.C.: Carnegie Instn of Washington.
LOSSINSKY, A. S. & LEFEVER, H. M. (1978). Ultrastructural banding observations in region1A-10F of the salivary gland X-chromosome of Drosophila melanogaster. Drosoph. Inf. Serv. 53,126-131.
MORTIN, L. I. & SEDAT, J. W. (1982). Structure of Drosophila polytene chromosomes, evidencefor a toroidal organization of the bands. J. Cell Sci. 57, 73-113.
MOTT, M. R., BURNETT, E. J. & HILL, R. J. (1980). Ultrastructure of polytene chromosomes ofDrosophila isolated by microdissection. J. Cell Sci. 45, 15-30.
PAINTER, T. S. (1934). Salivary gland chromosomes and the attack on the gene. J. Hered. 25,465-476.

264 V. Sorsa
SAURA, A. 0 . (1980). Electron microscopic analysis of the banding pattern in the salivary glandchromosomes of Drosophila melanogaster. Divisions 23 through 26 of 2L. Hereditas 93, 295-309.
SAURA, A. 0 . & SORSA, V. (1979a). Electron microscopic analysis of the banding pattern in thesalivary gland chromosomes of Drosophila melanogaster. Divisions 21 and 22 of 2L. Hereditas90, 39-49.
SAURA, A. O. & SORSA, V. (19796). Electron microscopic analysis of the banding pattern in thesalivary gland chromosomes of Drosophila melanogaster. Divisions 30 and 31 of 2L. Hereditas90, 257-267.
SAURA, A. O. & SORSA, V. (1979c). Electron microscopic analysis of the banding pattern in thesalivary gland chromosomes oiDmsophila melanogaster. Divisions 37, 38 and 39 of 2L. Hereditas91, 5-18.
SAURA, A. O. & SORSA, V. (1979<i). Electron microscopic analysis of the banding pattern in thesalivary gland chromosomes oiDmsophila melanogaster. Divisions 27, 28 and 29 of 2L. Hereditas91, 219-230.
SEMESHIN, V. F., ZHIMULEV, I. F. & BELYAEVA, E. S. (1979). Electron microscope autoradio-graphic study on transcriptional activity of Drosophila melanogaster polytene chromosomes.Chromosoma 73, 163-177.
SORSA, M. (1969). Infrastructure of the polytene chromosome in Drosophila melanogaster withspecial reference to electron microscopic mapping of chromosome 3R. Ann. Acad. Set. Fenn. ser.A, IVBiol. 151, 1-18.
SORSA, M. & SORSA, V. (1967). Electron microscopic observations on interband fibrils inDmsophila salivary chromosomes. Chromosoma 22, 32-41.
SORSA, M. & SORSA, V. (1968). Electron microscopic studies on band regions mDmsophila salivarychromosomes. Ann. Acad. Sci. Fenn. ser. A, IVBiol. 127, 1-8.
SORSA, V. (1982a). An attempt to estimate DNA content and distribution in the zeste-white regionof the X chromosome of Drosophila melanogaster. Biol. Zbl. 101, 8 1 - % .
SORSA, V. (19826). Volume of chromatin fibers in interbands and bands of polytene chromosomes.Hereditas 97', 103-113.
SORSA, V. (1983a). Interband fibrils of polytene chromosomes. Cell Differ. 12, 137-142.SORSA, V. (19836). Electron microscopic mapping and ultrastructure of Dmsophila polytene
chromosomes. In Insect Ultrastructure 2 (ed. H. Akai & R. C. King). New York: Plenum.SORSA, V., GREEN, M. M. & BEERMANN, W. (1973). Cytogenetic fine structure and chromosomal
localization of the white gene in Dmsophila melanogaster. Nature, new Biol. 245, 34—37.SORSA, V. & SAURA, A. O. (1980a). Electron microscopic analysis of the banding pattern in the
salivary gland chromosomes of Dmsophila melanogaster. Divisions 1 and 2 of X. Hereditas 92,73-83.
SORSA, V. & SAURA, A. O. (19806). Electron microscopic analysis of the banding pattern in thesalivary gland chromosomes oiDmsophila melanogaster. Divisions 3, 4 and 5 of X. Hereditas 92,341-351.
SORSA, V., SAURA, A. O. & HEINO, T. I. (1983). Electron microscopic analysis of the bandingpattern in the salivary gland chromosomes of Dmsophila melanogaster. Divisions 6 through 10of X. Hereditas 99, 181-200.
SWIFT, H. (1962). Nucleic acids and cell morphology in Dipteran salivary glands. In The MolecularContml of Cellular Activity (ed. J. M. Allen), pp. 73-125. New York: McGraw-Hill.
TEN TUSSCHER, B. & DERKSEN, J. (1982). The fourth chromosome of Chimnomus Malpighiantubules. Chromosoma 85, 643—658.
ZHIMULEV, I. F., SEMESHIN, V. F., KULICHKOV, V. A. & BELYAEVA, E. S. (1982). Intercalaryheterochromatin in Drosophila. I. Localization and general characteristics. Chromosoma 87,197-228.
{Received 26 April 1983-Accepted 8 June 1983)














![HSP70 overexpression may play a protective role in the ......ophila polytene chromosomes after thermal stress [23]. As a member of the HSP superfamily, HSP70 may act as a molecular](https://img.pdfslide.net/doc/110x75/60e7e367e6c0aa1dbe080064/hsp70-overexpression-may-play-a-protective-role-in-the-ophila-polytene-chromosomes.jpg)













