-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
1/6
The Active Sites of Fructose
6-Phosphate,2-kinase:Fructose-2,6-bisphosphatase from Rat
Testis
ROLES OF ASP-128, THR-52, THR-130, ASN-73, AND TYR-197*
(Received for publication, August 16, 1996, and in revised form,
December 12, 1996)
Kosaku Uyeda, Xiao-Li Wang, Hiroyuki Mizuguchi, Yang Li, Cu
Nguyen,and Charles A. Hasemann
From the Department of Veterans Affairs Medical Center, Research
Service, Dallas, Texas 75216 and the Departments ofBiochemistry and
Internal Medicine, University of Texas Southwestern Medical Center
at Dallas, Dallas, Texas 75235
To investigate the role in catalysis and/or substratebinding of
the Walker motif residues of rat testis fruc-tose
6-phosphate,2-kinase:fructose-2,6-bisphosphatase(Fru
6-P,2-kinase:Fru-2,6-Pase), we have constructedand characterized
mutant enzymes of Asp-128, Thr-52,Asn-73, Thr-130, and Tyr-197.
Replacement of Asp-128 by Ala, Asn, and Ser resulted in a small
decrease in Vmaxand a significant increase in Km values for both
sub-
strates. These mutants exhibited similar pH activityprofiles as
that of the wild type enzyme. Mutation ofThr-52 to Ala resulted in
an enzyme with an infinitelyhigh Km for both substrates and an
800-fold decreasedVmax. Substitution of Asn-73 with Ala or Asp
caused a100- and 600-fold increase, respectively in KFru 6-P
withonly a small increase in KATP and small changes in
Vmax.Mutation of Thr-130 caused small changes in the
kineticproperties. Replacement of Tyr-197 with Ser resulted inan
enzyme with severely decreased binding of Fru 6-Pwith 3-fold
decreased Vmax. A fluorescent analog of
ATP,2(3)-O-(N-methylanthraniloyl)ATP (mant-ATP) servedas a
substrate with Km 0.64 M, and Vmax 25 milli-units/mg and was a
competitive inhibitor with respect to ATP. When mant-ATP bound to
the enzyme, fluores-
cence intensity at 440 nm increased. mant-ATP bindingof the wild
type and the mutant enzymes were comparedusing the fluorometric
method. The Kd values of theT52A and D128N enzymes were infinitely
high and couldnot be measured, while those of the other mutant
en-zymes increased slightly. These results provide evi-dence that
those amino acids are involved in substratebinding, and they are
consistent with the crystallo-graphic data. The results also
suggest that Asp-128 doesnot serve as a nucleophile in catalysis,
and since thereare no other potential nucleophiles in the active
site, wehypothesize that the Fru 6-P,2-kinase reaction is medi-ated
via a transition state stabilization mechanism.
Fru 2,6-P21
is the most potent activator of phosphofructoki-
nase (PFK), and its synthesis and degradation are catalyzed
by
a bifunctional enzyme, Fru 6-P,2-kinase:Fru-2,6-bisphos-
phatase (Fru 6-P ATP i Fru 2,6-P2 ADP and Fru 2,6-P23 Fru 6-P
Pi). Several isozymic forms of the enzyme from
mammalian tissues have been characterized (1). They are all
homodimers with Mr ranging from 108,000 to 120,000. The
primary structures of these enzymes revealed that the
catalytic
domains are highly conserved, and the kinase and the phos-
phatase domains reside in the N-terminal half and the C-terminal
half, respectively (27).
The reaction catalyzed by Fru 6-P,2-kinase follows ternary
complex formation (8) with direct transfer of the -phosphate
of
ATP to the 2-OH of-D-Fru 6-P (9). Little is known about the
amino acid residues involved in substrate binding and
cataly-
sis. The Fru 6-P binding sites of Fru 6-P,2-kinase have been
studied by chemical modification and site-directed mutagene-
sis. The result of affinity labeling experiments revealed
that
Cys-107 and Cys-196 of rat liver Fru
6-P,2-kinase:Fru-2,6-Pase
and Cys-105 of the rat heart enzyme appear to be near or at
the
Fru 6-P binding site (10, 11). Site-directed mutagenesis of
the
rat testis enzyme demonstrated that Arg-102 (Arg-105 of the
liver enzyme) is essential for Fru 6-P binding (12).
Similarly,
the importance of this Arg residue in the liver and
muscleenzymes was shown by Rider et al. (13) and Kurland et al.
(14)
by mutagenesis of the Arg residue to Ala, which resulted in
a
200-fold increase in KFru 6-P. Arg-195 and Gly-48,
respectively,
were shown to be essential for Fru 6-P and ATP binding in
the
kinase reaction by site-directed mutagenesis in the liver
isozyme (15). More recently, Vertommen et al. (16) showed
that
site-directed mutagenesis of Lys-54 and Thr-55 of the liver
isozyme (which correspond to Lys-51 and Thr-52 of the testis
enzyme) resulted in a 5000-fold decrease in the kinase
activity.
Since mutation of Thr-55 to Cys resulted in loss of the
kinase
activity but the mutated enzyme still binds mant-ATP and
ATP, they suggested that the Thr residue may be involved in
catalysis.
The Fru 6-P,2-kinase reaction is very similar to that cata-
lyzed by PFK (Fru 6-P ATP 3 fructose 1,6-bisphosphate ADP). The
crystal structure of Escherichia coli PFK has been
solved (17). Based on the structures of complexes with Fru
6-P
and imidoadenosine 5-triphosphate (AMPPNP), Hellinga and
Evans (18) concluded that Asp-127 acts as a base, which in-
creases nucleophilicity by abstracting a H from the 1-OH of
Fru 6-P. In support of this idea is the observation that
muta-
* This work was supported by a grant from the Department of
Vet-erans Affairs, Grant DK16194 from the National Institutes of
Health(to K. U.), and a grant from the Welch Foundation (to C. A.
H.). Thecosts of publication of this article were defrayed in part
by the paymentof page charges. This article must therefore be
hereby marked adver-tisement in accordance with 18 U.S.C. Section
1734 solely to indicatethis fact.
To whom correspondence should be addressed: Dept. of
VeteransAffairs Medical Center, 4500 S. Lancaster Rd., Dallas, TX
75216. Tel.:214-372-7028; Fax: 214-372-9534.
1 The abbreviations used are: Fru-2,6-P2, fructose
2,6-bisphosphate;PFK, phosphofructokinase; Fru 6-P, fructose
6-phosphate; Fru-2,6-Pase, fructose-2,6-bisphosphatase; mant-ATP,
2(3)-O-(N-methylan-thraniloyl)-ATP; mant-ADP,
2(3)-O-(N-methylanthraniloyl)-ADP;
AMPPNP, 5-adenylylimidodiphosphate; WT, wild type; RT2K,
fructose6-phosphate,2-kinase:fructose-2,6-bisphosphatase; NMP
kinase, nucle-oside monophosphate kinase; HPLC, high pressure
liquid chromatog-raphy; MES, 4-morpholineethanesulfonic acid; E
t, total enzyme
concentration.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 272, No. 12, Issue of
March 21, pp. 78677872, 1997Printed in U.S.A.
This paper is available on line at
http://www-jbc.stanford.edu/jbc/ 7867
-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
2/6
tion of Asp-127 to Ser decreases the enzyme activity by
18,000-
fold compared with WT PFK (18). In the event that the mech-
anism of the Fru 6-P,2-kinase reaction is similar to that of
PFK, as has been widely presumed, then a nucleophile should
be identifiable that would activate the 2-OH of Fru 6-P for
attack on the ATP -phosphate.
Recently, we have crystallized (19) and solved the three-
dimensional structure of rat testis Fru
6-P,2-kinase:Fru-2,6-
Pase (RT2K) complexed with Mg2-ATP (20). These results
demonstrated that the Fru 6-P,2-kinase domain of the enzyme
does not resemble the structure of PFK as had been presumed
(21), but rather this domain is structurally similar to the
family
of nucleoside monophosphate kinases (NMP kinases), includ-
ing adenylate kinase (22) and uridylate kinase (23). The
Mg2-
ATP binding regions of the NMP kinases and the Fru 6-P,2-
kinase domain are remarkably similar, consisting of
classical
Walker-A (GXXGXGKT) and -B (ZZZD; where Z represents a
hydrophobic amino acid) motifs (24), which comprise a phos-
phate binding loop and a Mg2-coordinating aspartate residue,
respectively. In the Fru 6-P,2-kinase domain, the conserved
Lys and Thr residues of the Walker-A motif are Lys-51 and
Thr-52, while Asp-128 is the conserved residue of the
Walker-B
motif. No Fru 6-P was observed in the crystal structure, but
the
Fru 6-P binding site can be accurately predicted both by
ho-mology with the NMP binding sites of the NMP kinases and by
the location of residues previously identified to affect Fru
6-P
binding (1214). By modeling Fru 6-P in this presumed sub-
strate binding site, we have been able to search for a
potential
nucleophile for the activation of the 2-OH of Fru 6-P. With
the
exception of Asp-128, there is no Asp or Glu in the active
site
that might fulfill this role. We have identified Asn-73,
Thr-130,
and Tyr-197 as the only amino acids in the vicinity of the
binding site that might potentially act as weak nucleophiles
and/or assist in substrate binding.
To investigate the role in catalysis and/or substrate
binding
of the conserved Walker motif residues and the weak nucleo-
philes in the substrate binding site, we have prepared
mutant
enzymes at Asp-128, Thr-52, Asn-73, Thr-130, and Tyr-197.The
kinetic properties and the nucleotide binding of these
mutant enzymes have been determined and are compared with
the WT enzyme.
EXPERIMENTAL PROCEDURES
Rabbit muscle PFK was prepared as described (25). The cDNA
en-coding RT2K was prepared as described (7). Restriction enzymes,
T4DNA ligase, and T4 polynucleotide kinase were purchased from
New
England BioLabs (Beverly, MA). The Muta-Gene M13 in vitro
mutagen-esis kit was purchased from Bio-Rad. The pT77 RNA
polymerase/promoter plasmid (26) was a gift of Dr. Stan Tabor
(Harvard MedicalSchool). The Sequenase version 2 sequencing kit was
purchased fromU.S. Biochemical Corp. mant-ATP and mant-ADP were
synthesized bythe method of Hiratsuka (27) and purified by HPLC on
a Partisil 10SAX column (86 250 mm; Whatmann, Hillsboro, OR),
eluting with 0.6M ammonium phosphate (pH 4.0) (28). The product was
an equilibriummixture of 70% 3 isomer and 30% 2 isomer (28). All
other chemicalswere reagent grade and obtained from commercial
sources.
Site-directed MutagenesisPlasmid RT2K/pT77 containing the
RT2K gene cloned in a pT77 vector (7) was digested with XbaI
andHindIII, and the isolated 1.7-kilobase pair fragment was ligated
intothe XbaI-HindIII site of M13mp18 (M13/RT2K). The ligation
mixturewas used to transform E. coli JM109 competent cells. The
phage har-boring M13/RT2K was harvested and transfected into E.
coli CJ236(dutung). The purified recombinant M13/RT2K phage was
used toprepare uracil-containing single-stranded template.
Synthetic oligonu-cleotide primers used for constructing various
mutants are shown inTable I. The oligonucleotide-directed in vitro
mutagenesis was per-formed as described by Kunkel (29) using the
Muta-Gene M13 in vitromutagenesis kit. The double mutants
designated as T52A/D128A and
T52S/D128A were constructed as follows. The pT77 vector
containingD128A mutant of RT2K/pT77 DNA was digested with EcoRI,
isolated,
and introduced into EcoRI-digested RT2K/pT77 containing the
muta-
tion of T52A. The synthesized double-stranded DNA was used to
trans-form E. coli MV1190 competent cells. Mutant derivatives were
identi-
fied by DNA sequencing (30), and the DNAs were digested with
NdeIand HindIII. The DNA fragments containing the mutated RT2K
geneswere subcloned into the NdeI-HindIII sites of RT2K/pT77 and
ex-pressed in E. coli as before (7). The WT and mutant Fru
6-P,2-kinase:
Fru-2,6-Pase enzymes were purified as described previously (31).
How-ever, some of the mutant enzymes required slight modification
of thisprocedure, which are described under Results.
Assay Method for Fru 6-P,2-kinaseThis assay was based on
thedetermination of Fru 2,6-P2 and is the same as described
previously (32)with slight modification. The reaction mixture in a
final volume of 50 lcontained 100 mM Tris/HCl (pH 7.5), 0.1 mM
EDTA, 10 mM MgCl2, 2 mM
ATP, and 2 mM Fru 6-P. The mixture was incubated at 30 C for 10
min. At the end of the reaction, 0.1 N NaOH (50 l) was added, and
themixture was heated for 90 s at 80 C. Suitable aliquots were
assayed forFru 2,6-P2 as described by Uyeda et al. (33). One unit
of enzyme activity
is defined as the amount of enzyme that catalyzes the formation
of 1mol of Fru 2,6-P2 /min under these conditions.
Assay Method for Fru-2,6-PaseThis assay measures the formationof
Fru 6-P fluorometrically coupled to NADPH formation and
wasdescribed previously (31). The reaction mixture (in a final
volume of 1.0ml) contained 100 mM Tris/HCl (pH 7.5), 0.2 mM EDTA,
100 M NADP,0.4 unit of Glu 6-P dehydrogenase, 1 unit of
phosphoglucose isomerase,and varying amounts of Fru 2,6-P2. The
enzyme was desalted by columncentrifugation (34) in 15 mM Tris
sulfate (pH 7.5), 0.5 mM EDTA, and 5mM dithiothreitol. The reaction
was initiated with the addition of en-zyme and followed at 25 C by
measuring the NADPH formation at
452-nm emission and excitation at 350 nm using an
Aminco-BowmanSeries 2 luminescence spectrometer (SLM Aminco/Urbana,
IL).
Fluorescence MeasurementsFluorescence spectra were determinedat
25 C with an SLM Aminco Bowman Series 2
spectrofluorometer.Excitation and emission slits were set to 4 nm.
Spectra were correctedfor the buffer background but not for the
instrument response func-tions. Binding of mant-ATP to enzymes was
measured at 25 C in areaction mixture in a final volume of 0.2 ml,
containing 50 m M Trisphosphate (pH 7.5), 0.1 mM EDTA, 5 mM
dithiothreitol, and 1 M Fru6-P,2-kinase:Fru-2,6-Pase. Suitable
aliquots (1 l) of 10 M mant-ATPwere added and corrected for
dilution. Fluorescence intensity due to
free mant-ATP was measured in the absence of enzyme and used
forcorrection. The excitation and emission were 280 nm (or 350 nm)
and450 nm, respectively. The advantage of using 280-nm excitation
wasthat thefluorescence of mant-ATP was much lower than that at 340
nm.
Separation and Determination of mant-ADPmant-ADP and mant-ATP
were separated on a Partisil 10 SAX column (4.6 250 mm) usinga
Dionex HPLC (Dionex/Sunnyvale, CA) and a Ratio-2
fluorometer(Optical Technology Devices, Elmsford, NY) equipped with
a flow cell.mant-ADP was eluted from the column with a linear
gradient of 0.250.5 M ammonium phosphate (pH 4.0), containing 25%
ethanol with aflow rate of 1 ml/min. Under these conditions
mant-ADP was usually
eluted after 7.5 min, while mant-ATP was eluted with 0.5 M
ammoniumphosphate (after 15 min). To determine the concentration of
mant-ADP,
a standard curve of mant-ADP was generated by the HPLC
chromatog-
TABLE IOligonucleotides used for mutagenesis
These nucleotides are complementary to the DNA of RT2K at 182202
(T52A and T52S); 246267 (N73A and N73D); 410430 (D128AD128N); 419
436 (T130A); 416 436 (T130ST130V); 616 634 (Y197Fand Y197S).
Oligonucleotide Sequence
T52A 5-AGA AAT GTA GGC CTT GCC CCT-3T52S 5-AGA AAT GTA GGA CTT
GCC CCT-3
N73A 5- TA C TG A CC C AC G GC G AA T TC C C- 3N73D 5- TA C TG A
CC C AC G TC G AA T TC C C- 3
D128A 5-ATT GGT AGC AGC AAA AAC CGC-3D128S 5-ATT GGT AGC ACT AAA
AAC CGC-3D128K 5-ATT GGT AGC CTT AAA AAC CGC-3D128E 5-ATT GGT AGC
CTC AAA AAC CGC-3D128N 5-ATT GGT AGC ATT AAA AAC CGC-3
T130A 5-GGT GGT ATT GGC AGC ATC-3T130S 5-GGT GGT ATT GGA AGC ATC
AAA-3T130V 5-GGT GGT ATT GAC AGC ATC AAA-3
Y197F 5-GTT TTC AAA GCA TTC AAT-3Y197S 5-GTT TTC AGA GCA TTC
AAT-3
Active Site Residues of Fru 6-P,2-kinase7868
-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
3/6
raphy of varying concentrations (010 pmol) of mant-ADP and
by
calculating the concentration from the peak heights of the
mant-ADPfluorescence peaks. Samples (10 l) containing mant-ADP were
chro-matographed under the identical conditions, and the
concentrations
were calculated using the standard curve.Other
MethodsSDS-polyacrylamide slab gel electrophoresis was
performed with the Phast System (Pharmacia Biotech Inc.).
Proteinconcentration was determined by the Bradford method (35)
using bo-
vine serum albumin as a standard.
RESULTS
Expression and Purification of Various MutantsThe mu-
tated enzymes described here were purified using the same
procedure as that for the WT enzyme (31). However, D128N
and D128S mutant enzymes were eluted from the Yellow-3
column (fractionation step 4) with 20 mM ATP, while T52A and
Y197S were eluted from the same column with 0.2 or 0.3 M
potassium phosphate included in buffer A (Tris/P, 50 M, pH
7.5, 0.1 M EDTA; 0.1 M EGTA, 5% glycerol, 2 mM dithiothre-
itol, 1% polyethyleneglycol (M 300)). All of the enzymes
were
purified to apparent homogeneity as judged by SDS-polyacryl-
amide gel electrophoresis.
Steady State Fru 6-P,2-kinase KineticsTable II summa-
rizes the kinetic parameters of the WT RT2K and various
mutant enzymes.
T52A and T52SThr-52 to Ala altered the Km for both Fru
6-P and ATP to infinite value, i.e. not saturable, and the
Vmaxwas decreased to 11000 that of the WT enzyme. However,
Thr-52
to Ser increased the Km and Vmax values about 2-fold.
Similar
changes in the kinetic parameters were observed with T55C
and T55S mutants of rat liver Fru 6-P,2-kinase (16). Thus,
the
side chain hydroxyl group appears to be essential for binding
ofboth substrates and also for the catalysis. As depicted in Fig.
1,
the crystal structure showed that the side chain OH of Thr
is
hydrogen-bonded to Mg2, which is chelated to the PO4 of
ATP (or 2-phosphate of Fru 2,6-P2; Ref. 20). Thus, this
inter-
action may stabilize the pentacoordinated transition state.
Ap-
parently, the extra methyl group present in Thr does not
sig-
nificantly interfere with this interaction.
Asp-128 MutantsMutation of Asp-128 to Ala, Asn, and Ser
resulted in an 824-fold decrease in Vmax and a 30250-fold
increase and 33200-fold increase in KATP and KFru 6-P
values,
respectively. The changes in the kinetic properties of the
Ala
mutant were similar to those reported with the liver isozyme
(13). Asp-128 to Glu increased Vmax 2-fold, and Km values of
KATP and KFru 6-P increased 13- and 210-fold, respectively.
Surprisingly, the Asp to Lys mutation increased the Km
values
comparable with the other mutants, but the Vmax was de-
creased by 1100. Since the carboxyl group of Asp is chelated
to
Mg2 (Fig. 1), one would expect the negative charge to be
essential in the substrate binding and stabilization of the
tran-
sition state. However, it appears that while the anionic
inter-
action with Mg2 may not be essential for ATP binding, it is
required for catalysis.
T52A/D128A and T52S/D128ABoth of these double mu-
tant enzymes showed extremely high Km values for both sub-
strates as did the single mutant, T52A. The T52A/D128A mu-
tant displayed only trace kinase activity, but surprisingly,
the
T52S/D128A mutant retained the same Fru 6-P,2-kinase activ-
ity as the WT enzyme although half of that of the single
mutant
T52S. These results may suggest a more important role for
Thr-52 than for Asp-128 in efficient catalysis.Thr-130
MutantsRelatively small changes in the kinetic
properties, especially the Vmax values, were observed with
Thr-
130 to Ala or Ser mutation. Mutation to Val increased KFru
6-P350-fold without affecting the Vmax. Thr-130 is located near
the
furanose ring of Fru 6-P and may not have strong interaction
except to place the sugar moiety in proper orientation for
effi-
cient catalysis.
Tyr-197 MutantsMutation of Tyr to Phe increased KATPand KFru 6-P
by 6- and 70-fold, respectively, without change in
Vmax. However, mutation to Ser produced a large increase in
KFru 6-P
and a 19-fold increase in KATP. The Vmax decreased to13
that of the WT enzyme. These results suggest that Tyr-197
provides a hydrophobic pocket for Fru 6-P.
Asn-73 MutantsThe N73A mutant showed a 130-fold in-crease in
KFru 6-P and a 4-fold increase in KATP, but the Vmax value was only
slightly decreased (50% of WT). The N73D
mutant increased KFru 6-P to 600-fold and decreased Vmax to
9%
of WT without affecting KATP. The Vmax/KFru 6-PEt value of
the
N73D mutant was 0.02% of WT, which was 121 that of N73A.
Asn-73 appears to be essential for Fru 6-P binding, probably
through hydrogen bonding, but not essential for ATP binding
or
catalysis. This interpretation is consistent with the
crystal
structure (Fig. 1), which shows that Asn-73 is located near
the
furanose ring of Fru 6-P and may orient the Fru 6-P to a
proper
position for catalysis through hydrogen bonding with the
sugar
moiety. Introduction of a negative charge at Asn-73 (N73D)
may affect the interaction of Thr-52 and Asp-128, which are
essential for catalysis with Fru 6-P, and causes a large
decrease
TABLE IIKinetic constants of rat testis Fru 6-P,2-kinase:
Fru-2,6-Pase and the mutant enzymes
The results were averages of three determinations, and the
standard errors were 15%. The range of substrate concentrations
used for thedetermination of K
mvalues varied from 0.1 to 5 Km values.
EnzymeFru 6-P,2-kinase Fru-2,6-Pase
KATP KFru 6-P Vmax/Et V/KATP Et V/KFru 6-PEt KFru 2,6-P2 Vmax/Et
V/KFru2,6P2Et
mM mM s1 M1 s1 M1 s1 M s1 M1 s1
WT 0.1 0.04 0.073 730 1800 0.04 0.013 330,000T52A 20 30 0.0001
0.08 0.036 450,000
T52S 0.19 0.11 0.15 790 1400 0.05 0.016 320,000D128A 2.9 5.0
0.0037 1.3 0.74 0.16 0.017 110,000D128N 25 1.3 0.0031 0.12 2.4 0.03
0.012 400,000D128S 7.5 8.0 0.0092 1.2 1.2 0.08 0.017 210,000D128K
2.2 4.5 0.00061 0.28 0.14 0.06 0.0079 130,000D128E 1.3 8.4 0.14 110
17 0.07 0.013 190,000T52A/D128A 20 30 0.0092 0.16 0.0092
58,000T52S/D128A 20 30 0.073 0.09 0.046 510,000T130A 0.16 0.28
0.070 440 250 0.06 0.027 450,000T130S 0.03 0.20 0.046 1500 230 0.07
0.044 630,000T130V 1.4 14 0.092 66 6.6 0.07 0.039 560,000N73A 0.4
5.3 0.037 93 7.0 0.12 0.028 230,000N73D 0.09 24 0.0082 91 0.34 0.04
0.011 280,000
Y197F 0.6 2.8 0.083 140 30 0.14 0.017 120,000 Y197S 1.9 30 0.023
12 0.13 0.031 240,000
Active Site Residues of Fru 6-P,2-kinase 7869
-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
4/6
in Vmax/KFru 6-PEt.
Other Asp and Glu MutantsWe have altered all of the
highly conserved aspartates among the various isozymes to
Asn and some of the conserved glutamates to Gln includingAsp-94,
-112, -160, -189, -205, -209, and -231 and Glu-97, -120,
-135, -155, and -195. These mutant enzymes were purified to
homogeneity, and their kinetic parameters were determined.
These mutations did not cause significant changes in the ki-
netic properties of the kinase activity compared with the WT
enzyme.
Fru-2,6-Pase Activities of the Mutant EnzymesIn general,
all of these mutant enzymes showed only small changes (24-
fold increase) in KFru 2,6-P2 and Vmax values of
Fru-2,6-Pase.
mant-ATP as a SubstrateThe binding of mant-ATP, a flu-
orescent derivative of ATP, to various mutant enzymes and
the
WT enzymes was compared. mant-ATP serves as a substrate
for the WT Fru 6-P,2-kinase, and the apparent Kmant-ATP and
Vmax
were 0.64 M and 25 milliunits/mg, respectively (data not
shown). It is surprising that the Km value for mant-ATP is
156-fold smaller than that for ATP, since subsequent
examina-
tion of the crystal structure reveals that there is space
avail-
able to accommodate the methylanthraniloyl substituent with
the potential for favorable packing interactions. To
determine
if mant-ATP binds at the same catalytic site as ATP, Fru
6-P,2-kinase activity was measured at varying ATP or Fru 6-P
concentrations in the presence of constant concentration of
2
M mant-ATP. The kinase activity with mant-ATP alone as a
substrate in this reaction mixture (containing ATP as well)
was
determined by measuring the formation of mant-ADP. The
mant-ADP and mant-ATP in the reaction mixture were first
separated by HPLC chromatography and quantitated fluoro-
metrically as described under Experimental Procedures. The
results demonstrated that mant-ATP was a competitive inhib-itor
with respect to ATP but noncompetitive with respect to Fru
6-P. The KiATP and Ki
Fru 6-P values estimated from the plots
were 0.82 M and 2.8 M, respectively.
Binding of mant-ATP to WT Fru 6-P,2-kinase:Fru-2,6-Pase
was determined fluorometrically. When mant-ATP (2 M) was
bound to the enzyme, the fluorescence maximum decreased
from approximately 450 to 440 nm when excited at 350 nm, and
the fluorescence intensity increased compared with free
mant-
ATP (Fig. 2, a versus c). The mant-ATP binding to the enzyme
required Mg2 (Fig. 2, b versus c), since there was no change
in
the fluorescence in the absence of Mg2. The addition of 1 mM
ATP decreased the fluorescence at 440 nm, and the resulting
spectrum was identical to that of free mant-ATP (Fig. 2d),
suggesting that ATP displaced the enzyme-bound mant-ATP
completely. The same qualitative results were obtained when
mant-ATP was excited at 280 nm, but a much larger difference
in the fluorescent increase of bound mant-ATP was observed
compared with 350-nm excitation.
This increase in the fluorescence intensity was used to
study
the binding of mant-ATP to the WT and some of the mutant
enzymes, and the dissociation constants calculated from the
double reciprocal plots are summarized in Table III. The
dis-sociation constants for mant-ATP of the WT enzyme (0.80 M)
were comparable with the Kmant-ATP (0.64 M) and considerably
lower than KATP (100 M) and confirmed the tighter binding
than ATP. T52A and D128N did not bind mant-ATP, but other
mutant enzymes bound the ATP derivative as well as the WT
enzyme. Thus, the differences in the mant-ATP bindings to
these enzymes were similar to the differences in the KATP values
(Table I), although the former values were nearly 2
orders of magnitude lower than the latter values (Table II).
These differences suggest that the binding of mant-ATP by
the
enzyme was determined partly by hydrophobic interaction of
the mant- group.
These results, such as the large differences in the binding
constants of mant-ATP versus ATP, were in agreement with
FIG. 2. Emission spectra of mant-ATP bound to Fru 6-P,2-ki-
nase:Fru-2,6-Pase. The reaction mixture contained, in a final
volumeof 0.2 ml, 50 mM Tris phosphate (pH 7.5), 0.1 mM EDTA, 5 mM
dithio-threitol with 2 M mant-ATP (a), 1 M dimer (0.1 mg/ml) the
rat testisenzyme, and 2 M mant-ATP (b), 1 M enzyme, 2 M mant-ATP,
and 5mM MgCl2 (c); and 1 M enzyme, 2 M mant-ATP, 5 mM MgCl2, and
1mM ATP (d). The emission spectra were scanned at 25 C from 380
to560 nm with the excitation wavelength at 350 nm using an
SLMspectrofluorometer Series 2.
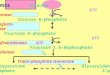
FIG. 1. Stereo diagram of the sub-strate binding region of the
Fru6-P,2-kinase domain of Fru 6-P,2-ki-nase:Fru-2,6-Pase. The side
chains mu-tated in this study are drawn as ball andstick models, as
is the Mg2-ATP. Thehydrogen bond interactions between the
ATP, Mg2, Thr-52, and Asp-128 are
drawn as solid white lines. Fru 6-P hasbeen modeled into the
proposed Fru 6-Pbinding site based on 1) the coincidence ofan empty
cavity in the crystal structurewith the substrate binding site of
the ho-mologous NMP kinases and 2) the conver-gence of residues
shown to affect the KFru6-P in this region. The proximity of
Tyr-197, Thr-130, and Asn-73 to the ribosering of Fru 6-P is
apparent.
Active Site Residues of Fru 6-P,2-kinase7870
-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
5/6
those reported recently with the rat liver Fru 6-P,2-kinase
and
its derivative containing six His residues (H6) at the C
termi-
nus (16). However, there were several important differences.
The rat liver enzyme (H6) contains high and low affinity
bind-
ing sites for Mg-mant-ATP and mant-ATP with Kd values of
108 M and 105 M, respectively. Moreover, mant-ATP binding
to the low affinity site is unaffected by mutation of Thr-55
and
Lys-54. We found that the rat testis enzyme had one binding
site for Mg-mant-ATP with the Kd values ranging from 107 to
106 M, and as shown in Fig. 2, its binding requires Mg 2.
Themutant enzymes showed different Kd values, which were con-
sistent with the changes in the KATP values. These
differences
may be due to the differences between these isozymes, which
may be a reflection of differences in the regulatory domains
since their catalytic domains are very similar.
pH Activity ProfilesIf Asp-128 is the nucleophile involved
in catalysis, mutation of this residue may show an altered
pH
activity profile. To investigate this possibility, the Fru
6-P,2-
kinase activity of the WT, D128A, and D128N enzymes at pH
values between 6 and 9.5 were determined. The results shown
in Fig. 3 indicated that the WT enzyme showed the pH opti-
mum between 7 and 8.5, and the apparent pK values for the
ascending and the descending rims were 6.6 and 9, respec-
tively. The pH profiles of the mutant enzymes were similar
tothat of the WT enzyme. The results suggested that Asp-128
does not serve as a base in the catalysis, confirming the
muta-
tion results. These pH activity curves are completely
different
from that of rat muscle bifunctional enzyme in which the ki-
nase activity increases linearly from 6 to 10, and the
optimum
is pH 10 (13). Furthermore, mutation of Asp-128 to Ala of
the
muscle enzymes results in loss of the pH optimum at 10 and
becomes independent of pH. Since the active sites of all of
these
isozymes are highly conserved, it is unlikely that these
differ-
ences are due to tissue-specific isozymic differences.
DISCUSSION
All kinase reactions or phosphoryl transfer reactions
require
a base (nucleophile) for catalysis, positive charges to
neutral-
ize/stabilize the negative charges of the ATP phosphates,
andresidues to position the substrates and stabilize the
transition
state. The determination of the three-dimensional structure
has demonstrated that the active site of the Fru
6-P,2-kinase
domain of the Fru 6-P,2-kinase:Fru 2,6-P2 appears to satisfy
these requirements with the exception of a catalytic nucleo-
phile. It was the goal of this study to probe by mutagenesis
the
residues in the active site for their potential role as the
appar-
ently missing nucleophile. The effect of mutagenesis of the
residues located at or near the active site presented herein
is
consistent with the three-dimensional model determined by
crystallography as shown in Fig. 1. Thr-52 and Asp-128 are
hydrogen-bonded/salt-bridged to the Mg2 ion of the Mg2-
ATP complex. Accordingly, all but the most conservative mu-
tations of these residues have a very significant impact on
the
KATP. Thr-52 is also close enough to the -phosphate of the
ATP
to be directly hydrogen-bonded to it, and this may explain
the
most extreme effect on KATP of the T52A mutation. The pro-
found effect on Vmax of the T52A mutation and the comparably
modest effects of the Asp-128 mutations were not predicted.
We
would have expected the charge interaction of Asp-128 with
the
Mg2 ion to dominate the coordination of the Mg2-ATP, while
our results indicate that Thr-52 is the more important Mg2
ligand. The Mg2 ion may play a role in catalysis by
stabilizing
the negative charge involved in transition state formation.
Thr-52 may have an additional role in the ATP binding. Ac-
cording to the crystal structure of the bifunctional enzyme,
the
kinase active site is nearly identical to the GTP binding site
of
p21ras, and Thr-52 in the former is situated at the position
analogous to Ser-17 in the latter (36, 37). Muegge et al.
(38)
proposed that the interaction of Ser-17 with the GTP -phos-
phate maintains a conformation of the P-loop main chain at-
oms, which optimize the orientation of the main chain
dipoles
for interaction with the GTP phosphates. Thus, a component
of
the decreased catalysis observed in our T52A mutants might
be
due to a loss of an essential main chain conformation in the
P-loop, which promotes optimal electrostatic interactions
with
the ATP. Further mutagenesis and crystallography of mutant
enzymes or transition state mimics will be necessary to
resolve
the structural basis for this observation.
It should be noted that the observed changes in the
kineticproperties of the mutated enzymes were not due to global
structural changes for the following reasons. 1) All of
these
enzymes were purified to homogeneity with the same proce-
dure as that used for the WT enzyme. The only exception was
that some of the mutants bound to an affinity column (Yel-
low-3) more tightly than the WT enzyme, requiring slightly
higher ATP concentration and/or higher phosphate for
elution,
although they showed increased Km values for the
substrate(s).
2) Intrinsic tryptophan fluorescence spectra of Asp-128 and
Thr-52 mutants were identical to that of the WT enzyme (data
not shown).
The effect on the KFru 6-P of the mutants, which would ap-
parently only affect Mg2-ATP binding, can best be explained
by assuming that ligand binding in the Fru 6-P,2-kinase
active
TABLE IIIDissociation constants of mant-ATP
The Kd
values were the average S.E. of three determinations. T52Aand
D128N did not bind mant-ATP. The binding was determined
flu-orometrically as described under Experimental Procedures.
Enzyme Kdmant-ATP
M
WT 0.80 0.05T130A 0.90 0.10
R136L 1.9 0.10T52S 1.6 0.10T52AD128N
Y197F 1.5 0.10
FIG. 3. Fru 6-P,2-kinase pH activity profiles of WT, D128A,
andD128N Fru 6-P,2-kinase:Fru-2,6-Pase. Fru 6-P,2-kinase activity
wasdetermined as described under Experimental Procedures, except
thebuffer mixture consisted of 0.1 M MES, 0.051 M
N-ethylmorpholine, and0.051 M diethanolamine at varying pH values,
and 0.1 and 20 m M Fru
6-P for the WT and the neutral enzymes, respectively, were used.
Thereaction was initiated with 10 mMATPand 20m M MgCl2 forthe
mutantenzymes and with 1 mM ATP and 20 mM MgCl2 for the WT
enzymes.
Active Site Residues of Fru 6-P,2-kinase 7871
-
8/14/2019 The Active Sites of Fructose 6-Phosphate,2-Kinase
6/6
site is an ordered process in which ATP binds to the enzyme
first followed by Fru 6-P. Earlier kinetic studies (8)
suggested
that the addition of the substrates to liver Fru 6-P,2-kinase
is
random, but it is possible that under these assay conditions
the
ordered substrate addition was favored. Thus, mutations af-
fecting the ability to bind ATP will concomitantly affect Fru
6-P
binding, while the inverse is not necessarily true.
While the crystal structure of the Fru 6-P,2-kinase:Fru-2,6-
Pase did not include Fru 6-P bound at its active site, the
homology of this enzyme with the NMP-kinases and the loca-
tion of several residues known to affect KFru 6-P have
allowed
for a confident prediction of the Fru 6-P binding site. As
shown
in Fig. 1, there are several residues in the vicinity of the
Fru
6-P binding site of the enzyme that are potential (albeit
weak)
nucleophiles, including Thr-130, Tyr-197, and Asn-73. While
mutations of each of these residues (or Thr-52 and Asp-128)
has
an impact on KFru 6-P (or KATP), their low impact on
Vmax(1150-fold reductions) would indicate that none of them act
as
a nucleophile for activating substrate (compared with the
13,000-fold reduction observed for the D127A mutation of the
PFK nucleophile (39)). Again, turning to the structure in Fig.
1,
this is reasonable, since their relation to a modeled Fru
6-P
substrate places them in positions to bind the ribose moiety
(Asn-73, Thr-130, Tyr-197) or the Mg2
-ATP (Thr-52, Asp-128)and not in the immediate vicinity of the
gap between the
-phosphate and the Fru 6-P 2-OH, where an activating nu-
cleophile would be expected to reside. In addition, we have
mutated all of the highly conserved Asp and Glu residues in
the
Fru 6-P,2-kinase domain, but none of these mutant enzymes
were affected significantly.
The evidence presented here supports our contention that
the catalytic mechanism of the Fru 6-P,2-kinase reaction may
not involve a strong nucleophile but instead is mediated via
transition state stabilization. This may be the reason for
the
extremely low value for kcat of 0.07 s1 of Fru 6-P,2-kinase.
This is in contradiction to the widely held belief that the
Fru
6-P,2-kinase reaction would be PFK-like (21). This is not
surprising in light of the structural differences between the
Fru6-P,2-kinase domain and the PFK structures and the unantic-
ipated similarity between the Fru 6-P,2-kinase domain and
the
NMP-kinase and G-protein structures. These latter enzymes
are believed to catalyze their reactions by a transition
state
stabilization mechanism.
AcknowledgmentsWe thank Drs. Paul A. Srere and Paul F. Cookfor
critical reading of this paper.
REFERENCES
1. Uyeda, K. (1991) in CRC Review (Kuby, S. A., ed) Vol. 2, pp.
445456, CRCPress, Boston, MA
2. Rider, M. H., Marchand, M. J., Hue, L., and Rousseau, G. G.
(1987) FEBS Lett.
2243, 3173213. Algaier, J., and Uyeda, K. (1988) Biochem.
Biophys. Res. Commun. 153,
3283334. Lively, M. O., El-Maghrabi, M. R., Pilkis, J., DAngelo,
G., Colosia, A. D.,
Ciavola, J.-A., Fraser, B. A., and Pilkis, S. J. (1988) J. Biol.
Chem. 263,839849
5. Crepin, T. M., Darville, M. I., Michel, A., Hue, L., and
Rousseau, G. G. (1989)Biochem. J. 264, 151160
6. Sakata, J., and Uyeda, K. (1990) Proc. Natl. Acad. Sci. U. S.
A. 87, 495149557. Sakata, J., Abe, Y., and Uyeda, K. (1991) J.
Biol. Chem. 266, 1576415770
8. Kitajima, S., Sakakibara, R., and Uyeda, K. (1984) J. Biol.
Chem. 259,
689669039. Kountz, P. D., Freeman, S., Cook, A. G., El-Maghrabi,
M. R., Knowles, J. R.,
and Pilkis, S. J. (1988) J. Biol. Chem. 263, 160691607210.
Sakakibara, R., Kitajima, S., Hartman, F. C., and Uyeda, K. (1984)
J. Biol.
Chem. 259, 140231402811. Kitamura, K., Uyeda, K., Hartman, F.
C., Kangawa, K., and Matsuo, H. (1989)
J. Biol. Chem. 264, 6344634812. Tsujikawa, T., Watanabe, F., and
Uyeda, K. (1995) Biochemistry 34,
63896393
13. Rider, M. H., Crepin, K. M., deChoedt, M., Bertraud, L.,
Vertommen, D., andHue, L. (1995) Biochem. J. 309, 341346
14. Kurland, I., Chapman, B., Lee, Y-H., and Pilkis, S. J.
(1995) Biochem. Biophys.Res. Commun. 213, 663672
15. Li, L., Lin, K., Kurland, I. J., Correia, J. J., and Pilkis,
S. J. (1992) J. Biol.Chem. 267, 43864393
16. Vertommen, D., Bestrand, L., Sontag, B., DiPietro, A.,
Louckx, M. P., Vidal, H.,
Hue, L., and Rider, M. H. (1996) J. Biol. Chem. 271,
178751788017. Shirakihara, Y., and Evans, P. R. (1988) J. Mol.
Biol. 204, 973994
18. Hellinga, H. W., and Evans, P. R. (1987) Nature 327,
437439
19. Istvan, E. S., Hasemann, C. A., Kurumbail, R. G., Uyeda, K.,
and Deisenhofer,J. (1995) Protein Sci. 4, 24392441
20. Hasemann, C. A., Istvan, E. S., Uyeda, K., and Deisenhofer,
J. (1996)Structure4, 10171029
21. Bazan, J. F., Fletterick, R. J., and Pilkis, S. J. (1989)
Proc. Natl. Acad. Sci.U. S. A. 86, 96429646
22. Muller, C. W., and Schulz, G. E. (1992) J. Mol. Biol. 224,
15917723. Muller-Dieckmann, H-J., and Schulz, G. E. ( 1995) J. Mol.
Biol. 246, 522530
24. Walker, J. E., Saraste, M., Runswicks, M. J., and Gay, N. J.
(1982) EMBO J.1, 945951
25. Uyeda, K., Miyatake, A., Luby, L. J., and Richards, E. G.
(1978) J. Biol. Chem.253, 83198327
26. Tabor, S., and Richardson, C. (1985) Proc. Natl. Acad. Sci.
U. S. A. 82,
1074107827. Hiratsuka, T. (1983) Biochim. Biophys. Acta 742,
496508
28. Hazlett, T. L., Moore, K. J. M., Lowe, P. N., Jameson, D.
M., and Eccleston, J.F. (1993) Biochemistry 32, 1357513583
29. Kunkel, T. A. (1985) Proc. Natl. Acad. Sci. U. S. A. 82,
48849230. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc.
Natl.Acad. Sci.U. S. A.
74, 5463546731. Tominaga, N.,Minami, Y.,Sakakibara, R.,and
Uyeda,K. (1993)J. Biol.Chem.268, 1595115957
32. Furuya, E., and Uyeda, K. (1981) J. Biol. Chem. 256,
7109711233. Uyeda, K., Furuya, E., and Luby, L. J. (1981) J. Biol.
Chem. 256, 83948399
34. Penefsky, H. S. (1977) J. Biol. Chem. 252, 2891289935.
Bradford, M. M. (1976) Anal. Biochem. 72, 248254
36. DeVos, A. M., Tory, L., Milburn, M., Matias, P. M.,
Jancarik, J., Noguchi, S.,Nishimura, K., Miura, K., Ohtsuka, E.,
and Kim, S. H. (1988) Science 239,888893
37. Pai, E. F., Kabsch, W., Krengel, U., Holmes, K., John, J.,
and Wittinghofer, A.(1989) Nature 341, 209214
38. Muegge, I., Schweins, T., Langen, R., and Warshel, A. (1996)
Structure 4,475489
39. Berger, S. A., and Evans, P. R. (1992) Biochemistry 31,
92379242
Active Site Residues of Fru 6-P,2-kinase7872