Embed Size (px)
Citation preview

1093
The Auk 117(4):1093–1115, 2000
CONSERVATION REPORT
THE AOU CONSERVATION COMMITTEE REVIEW OF THE BIOLOGY,STATUS, AND MANAGEMENT OF CAPE SABLE SEASIDE SPARROWS:
FINAL REPORT
JEFFREY R. WALTERS,1,8 STEVEN R. BEISSINGER,2 JOHN W. FITZPATRICK,3 RUSSELL
GREENBERG,4 JAMES D. NICHOLS,5 H. RONALD PULLIAM,6 AND DAVID W. WINKLER7
1Department of Biology, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061, USA;2Department of Environmental Science, Policy and Management, 151 Hilgard Hall 3110, University of California,
Berkeley, California 94720, USA;3Cornell Laboratory of Ornithology, 159 Sapsucker Woods Road, Ithaca, New York 14850, USA;
4Smithsonian Migratory Bird Center, National Zoological Park, Smithsonian Institution,Washington, D.C. 20008, USA;
5U.S. Geological Survey, Biological Resources Division, Patuxent Wildlife Research Center,Laurel, Maryland 20708, USA;
6Institute of Ecology, University of Georgia, Athens, Georgia 30605, USA; and7Department of Ecology and Evolutionary Biology, Cornell University, Ithaca, New York 14853, USA
THE CAPE SABLE SEASIDE SPARROW (Ammo-dramus maritimus mirabilis) was listed as anoriginal member of the federal list of endan-gered species in 1968. It is restricted to season-ally flooded prairies of extreme southern Flor-ida and is disjunct from all other conspecificbreeding populations (Kushlan et al 1982,McDonald 1988). Since the subspecies was de-scribed in 1919, its populations have been dis-covered and rediscovered, often only to dis-appear or decline to a handful of individuals(Werner and Woolfenden 1983, Kushlan andBass 1983). Although the sparrow historicallyis known from six distinct areas, at presentonly two of these areas support populationsnumbering in the hundreds or low thousandsof individuals.
Debates swirl around the status of these rem-nant populations. The main controversy en-compasses whether the sparrow, now largelyrestricted to Everglades National Park, is injeopardy of global extinction and if so, what ac-tions must be taken to prevent this from hap-pening. In November 1998, a panel of scientistswas assembled under the auspices of the Con-servation Committee of the American Orni-thologists’ Union (AOU) to evaluate the scien-tific evidence relevant to this controversy. ThePanel was charged with scrutinizing the evi-dence for the existence and probable causes ofglobal population decline in this subspecies,
8 E-mail: [email protected]
evaluating proposed management actions, andsuggesting further research necessary to man-age the remaining populations to maximizetheir chances of long-term persistence.
This document presents the conclusions ofthe Panel, which are based on our reading ofthe peer-reviewed and ‘‘gray’’ literature, inter-actions during a workshop held 9 to 11 Febru-ary 1999 at Florida International University inMiami with researchers investigating the spar-row’s biology, and site visits associated withthe workshop. In addition, researchers provid-ed the Panel with position papers summarizingtheir findings and conclusions prior to theworkshop and provided information in re-sponse to specific questions following theworkshop. Further information was obtainedthrough public comment on an initial draft ofthis report.
THE CAPE SABLE SEASIDE SPARROW IN
CONTEXT
The Cape Sable Seaside Sparrow is the onlyavian taxon entirely restricted to the Ever-glades ecosystem. Considerable evidence sug-gests that it is adapted to local conditions. TheCape Sable Seaside Sparrow was originally de-scribed as a distinct species, a status it officiallymaintained (contrary to recommendations ofGriscom 1944), along with the now extinctDusky Seaside Sparrow (A. m. nigrescens), until

1094 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
the 32nd supplement to the AOU check-list(AOU 1973).
The long-held view that the Cape Sable Sea-side Sparrow is a separate species was presum-ably based on its isolation, as well as its mor-phological and ecological distinctiveness. Eco-logically, the Cape Sable Seaside Sparrow joinsthe Dusky Seaside Sparrow as the only SeasideSparrows known to have occurred in fresh-water wetlands. The plumage is distinctivelymore olive above and streaked below com-pared with other Seaside Sparrows. The adap-tive significance of this coloration is unknown;one possibility is that it matches the back-ground of the greenish-gray periphyton (algae)layer that covers the soil in the marl prairiesthese birds inhabit. MacDonald (1988) ana-lyzed variation in all of the recognized SeasideSparrow subspecies and found the Cape SableSeaside Sparrow to be, along with the Texas (A.m. sennetti) and Dusky subspecies, significantlysmaller than other Seaside Sparrows. Robinsand Schnell (1971) determined that the CapeSable Seaside Sparrow was the most distinctiveof all Seaside Sparrow subspecies in terms ofoverall skeletal morphology, even when char-acters were standardized by sternum or hu-merus length. This suggests uniqueness in‘‘shape’’ as well as size. MacDonald (1988) an-alyzed sound spectrographs and concludedthat both Cape Sable and Dusky seaside spar-rows had songs that were distinctly more ‘‘in-sect-like’’ than songs of other Seaside Spar-rows.
As of this writing, no definitive analysis ex-ists of the relationship of Cape Sable SeasideSparrows to other Seaside Sparrows based onmolecular genetics (Avise and Nelson 1989, J.Avise pers. comm.). We note that the DuskySeaside Sparrow was distinct from other Sea-side Sparrows in many of the same ways as theCape Sable Seaside Sparrow, yet the Dusky wasnot genetically distinguishable from other At-lantic Coast populations based on mtDNAmarkers (Avise and Nelson 1989). Recent stud-ies of other sparrow taxa (e.g. Melospiza melodiaand M. georgiana) demonstrate substantial geo-graphically based, and presumably adaptive,morphological variation in the absence of geo-graphic structuring in mtDNA haplotypes(Zink and Dittmann 1993, Greenberg et al.1998). Therefore, little basis exists on which toargue against species-level recognition of a tax-
on that shows marked morphological diver-gence because of lack of differentiation inmtDNA (Zink and Kale 1995). In summary, un-equivocal morphological, behavioral, and eco-logical grounds exist for recognizing the CapeSable Seaside Sparrow as a unique subspeciesthat qualifies for protection under the Endan-gered Species Act.
As a subspecies that historically has occu-pied coastal marshes and inland prairies, theCape Sable Seaside Sparrow typifies the gen-eral threats to two particularly vulnerable com-ponents of the North American avifauna. First,as a taxon with a declining number of small, lo-cal populations in Spartina marshes, it joins alarge number of distinctive emberizid spar-rows associated with coastal wetlands, includ-ing other Seaside Sparrows (McDonald 1988)and subspecies of Song Sparrows (Melospizamelodia), Swamp Sparrows (M. georgiana), Salt-marsh Sharp-tailed Sparrows (Ammodramuscaudacutus), and Savannah Sparrows (Passercu-lus sandwichensis). Second, as a taxon restrictedto prairie habitat, it joins a large number ofsparrows and other birds associated with nat-ural grasslands that are declining substantially(Knopf 1995), including several other Ammo-dramus species such as Baird’s Sparrow (A. bair-dii), Henslow’s Sparrow (A. henslowii), andGrasshopper Sparrow (A. savannarum).
THE CURRENT CONTROVERSY
The current controversy centers on the scaleand implications of population declines and lo-cal extirpations that have been described forthis subspecies. Curnutt et al. (1998) and Nottet al. (1998) suggest that recent population de-clines are the direct result of water manage-ment practices that have altered the greater Ev-erglades ecosystem. Such an anthropogenic de-cline, if true, is all the more significant consid-ering the recent extinction of the Dusky SeasideSparrow, which was generally acknowledgedto have resulted from habitat mismanagement(Walters 1992).
Flooding and fire are the major explanationsfor the putative decline in Cape Sable SeasideSparrows (Curnutt et al. 1998, Nott et al. 1998).The prairie habitat that supports remainingpopulations is naturally prone to both. How-ever, researchers hypothesizing anthropogenicdecline note the increasingly restricted distri-

October 2000] 1095Cape Sable Seaside Sparrow Report
bution of the sparrow to a few habitat patches(Werner and Woolfenden 1983), along with hu-man-caused changes in flooding and fire re-gimes (Curnutt et al. 1998, Nott et al. 1998), asfactors that combine to threaten remainingpopulations. Specifically, emergency releases ofwater into the western portion of EvergladesNational Park appear to increase the frequencyand duration of flooding of the western popu-lation of sparrows during the breeding season.At the same time, structures restricting waterflow to the east create unnaturally frequentfire-prone conditions in the prairies inhabitedby northeastern populations (USDI 1998).
The potential for human influences on CapeSable Seaside Sparrow populations, particular-ly through catastrophic fire and the incursionof exotic trees (see Werner and Woolfenden1983), has been discussed since the subspecies’discovery. The potential problems of habitatfragmentation and increased frequency and ex-tent of catastrophic, dry-season fire were de-scribed by Werner and Woolfenden (1983).Kushlan et al. (1982) prepared a managementplan that, although not advocating any drasticmanagement actions, suggested that fire, flood-ing, and the encroachment of trees (particular-ly exotics) were threats in need of continuedmonitoring. Unnaturally frequent flooding andfire were cited by Post and Greenlaw (1994) asthreats to the prairie habitat of the Cape SableSeaside Sparrow.
Concern about the threats facing the spar-rows has heightened dramatically in recentyears, coinciding with an unusually wet periodin the Everglades system. Based on monitoringreinstated in 1992 of both habitat and popula-tions, an alarm was sounded about the effect ofthe high water years of 1993 to 1995 on thewestern Everglades (Orians et al. 1996). Re-cently, Curnutt al. (1998) and Nott et al. (1998)suggested that Cape Sable Seaside Sparrowsface imminent jeopardy of extinction owing toincreased risks from flooding and fire that re-sult from the shunting of water from east towest within the Everglades.
W. Post (pers. comm.) argues that the CapeSable Seaside Sparrow is not in imminent jeop-ardy. He suggests that local declines, if real, area natural consequence of the highly dynamicand vagile nature of the subspecies’ populationdynamics. As a subspecies adapted to habitatsthat are subjected to a highly unpredictable
disturbance regime, populations may shift lo-cations, disappearing from and reappearing inparticular habitat patches. Post further sug-gests that the primary native habitat of theCape Sable Seaside Sparrow, like other SeasideSparrows, was coastal Spartina marshes. In-deed, it was in such habitat that the species wasfirst discovered and subsequently extirpatedon Cape Sable. According to Post’s hypothesis,the disappearance of suitable Spartina habitatowing to both natural and anthropogenic caus-es has restricted the Cape Sable Seaside Spar-row to suboptimal inland prairie habitat. Asimilar argument was made by Post (pers.comm.) in relation to the decline of the DuskySeaside Sparrow.
The controversy surrounding the existenceand causes of a global decline in the Cape SableSeaside Sparrow is borne from the scientific un-certainty surrounding almost every facet of thebiology of this difficult-to-observe subspecies.The controversy has been nurtured further bythe possible implications that changes in habi-tat management have to the ecology of the Ev-erglades ecosystem and to the economic base ofpeople living in the vicinity.
Scientific uncertainty stems, in large part,from an incomplete historical record of thesubspecies’ distribution and abundance. Stim-son (1956) is credited with the first intensivesearch for populations away from the originalCape Sable and Ochopee sites. The first system-atic survey of the sparrow throughout itsknown range was conducted in 1981 (Kushlanand Bass 1983), although extensive surveysalso were conducted by Werner and Woolfen-den in the mid-1970s and by Kushlan and Bassfrom 1978 to 1980. Information on Cape SableSeaside Sparrows prior to the 1970s is anec-dotal and fragmentary, and no systematic sur-veys were conducted between 1981 and 1992.Extensive research on the population biology ofthe subspecies began in 1992, and many of theimportant details of these studies are not yetpublished.
Given the recent reports of population de-clines, how probable and imminent is the ex-tinction of the Cape Sable Seaside Sparrow un-der current management? This critical questionis difficult to answer definitively, because manyof the problems identified by researchers ascontributing to the decline eventually may bealleviated by long-term changes in water man-

1096 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
agement that already are planned. The emer-gency measures required to stabilize the pop-ulation in the interim, as well as the adequacyof the planned changes, are the management is-sues at hand.
SEASIDE SPARROW HABITAT AND WATER
MANAGEMENT IN THE EVERGLADES
Currently, the Cape Sable Seaside Sparrow isentirely restricted to marl prairies within andimmediately adjacent to Everglades NationalPark. Outside the park, much of this distinctiveprairie habitat has been converted to agricul-tural land, making proper management of rem-nant patches of prairie within the park espe-cially critical (DeAngelis et al. 1998). In thesubtropical climate of the Everglades, season-ality and year-to-year variation of rainfallgreatly affect ecological systems. Short-termand long-term cycles of water levels are drivenby seasonal variation in rainfall and by watermanagement regimes. Most rain occurs duringthe wet season of May through September(Thomas 1974, Duever et al. 1994). Less rainfalls in the other months, but the ‘‘dry season’’is rarely completely dry, because winter rainsperiodically occur when cold fronts movesouthward down the Florida peninsula. TheEverglades are less seasonal in rainfall, andpresumably in water levels, than are many oth-er lowland wetlands in the Neotropics (Beissin-ger and Gibbs 1993). Water levels and rainfallin South Florida also appear to follow longer-term drought/flood cycles with a periodicity offour to seven years (Thomas 1974, Beissinger1986).
Water management can have important in-fluences on the seasonality of water levels. Forexample, Beissinger (1986) showed that season-al fluctuations in Lake Okeechobee water levelsin the northern Everglades were small prior tothe completion of the dike around the lake in1930, but they increased greatly during themid-1900s when large volumes of water beganto be released from the lake via canals. Varia-tion in Okeechobee’s water levels has decreasedagain since the completion of the South FloridaWater Management Project.
Under natural conditions, water in the south-ern Everglades comes from local precipitationand from southward flows out of Lake Okee-chobee. Because of the slight tilt of the land,
and the lack of major topographic features, thiswater flows sheet-like across virtually the en-tire Everglades. Vegetation varies with subtlelocal differences in the annual period of flood-ing (hydroperiod), which are caused largely byminor microgeographic variation in topogra-phy and drainage. Marl prairie occurs withinthe zone intermediate between the permanent-ly flooded sloughs and the drier pine-dominat-ed high ground. Marl prairie is a relatively di-verse floristic association dominated by grass-es, sedges, and rushes growing on thin lime-stone soils that are seasonally flooded. Prairieoccurs where the hydroperiod is four to eightmonths long. Where hydroperiods are longer,taller marsh grasses and sedges dominate, andwhere hydroperiods are shorter, prairie per-sists only where fire eliminates woody plants.
A core issue is the effect of water manage-ment projects in recent decades on these prai-ries. Water flow has been under increasing hu-man management for nearly 100 years. Withurbanization of the greater Miami area and ag-ricultural development in areas to the northand east of Everglades National Park, the Unit-ed States Army Corps of Engineers began inthe 1950s to construct a series of structures andimpoundments that control water flowthroughout South Florida. Today, water flowinto Everglades National Park is controlled byfloodgates and levees in Water ConservationAreas 3A and 3B to the north. A series of flood-gates (S-12s) along the east-west oriented Tam-iami Trail at the north end of Shark RiverSlough allows managers to release water fromWater Conservation Area 3A (WCA 3A) south-ward into the western portion of the park (Fig.1). Water that historically would have flowedthrough WCA 3B into the eastern portion of thepark thereby may be diverted to the west. TheL-67 extension canal and levee prevent waterreleased from WCA 3A from flowing into theeastern portion of the park (Fig. 1). Diversionof so much water to the west has reduced theaverage hydroperiod of the marl prairie in thenortheastern portion of the park. Emergencyreleases of water from WCA 3A add to theflooding of areas west of Shark River Sloughduring years of high precipitation (Nott et al.1998).
One of the remaining sparrow populations(Population A) is affected by flooding of thesewestern areas, and another (Population D) is af-

October 2000] 1097Cape Sable Seaside Sparrow Report
FIG. 1. Location of Cape Sable Seaside Sparrow populations A to F in Everglades National Park in relationto Shark River Slough, Taylor Slough, the S-12 floodgates (S-12A to S12D), and the L-67 extension canal andlevee (L-67 Ext.). Major roads are shown for reference.
fected by additional releases of water into Tay-lor Slough in the southeastern portion of thepark (Fig. 1). Three populations (C, E, and F)occur in the prairies that experience unusuallyfrequent and prolonged dry conditions in thenortheastern portions of the park. The remain-ing population (B) occurs along the IngrahamHighway in the eastern part of the park.
The new water management strategyplanned for the Everglades calls for construc-tion of new structures that will restore morenatural patterns of water flow, especially in-creased flow into northeastern Shark RiverSlough and decreased flows west of Shark Riv-er Slough. Water managers have purposely pro-hibited significant flows into northeasternShark River Slough because such flows wouldincrease flooding of large areas of private landwithin the East Everglades (i.e. Everglades Na-tional Park Expansion Area) and would in-crease groundwater levels in another area ofprivate land (known as the ‘‘8.5 square mile’’area). The new plans call for purchase of these
private lands. In the interim, however, watercontinues to be routed west of Shark RiverSlough rather than into northeastern Shark Riv-er Slough. In wet years, water is either retainedin Water Conservation Area 3A or releasedwest of Shark River Slough and through a pointsource into Taylor Slough, resulting in floodingof those areas. Retention of water in WCA 3Aproduces abnormally high water levels that canprolong the flooding of tree islands (Guerra1996). This may have adverse effects on otherspecies of concern within the Everglades sys-tem, and on people. WCA 3A is inhabited bymembers of the Miccosukee tribe of NativeAmericans, and high water may result in flood-ing of culturally significant sites.
Below, we review and evaluate the scientificevidence bearing on population trends of CapeSable Seaside Sparrows and their significance,and provide recommendations for further re-search. We conclude by offering short-term andlong-term management recommendations basedon our findings and interpretations.

1098 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
RECENT POPULATION TRENDS
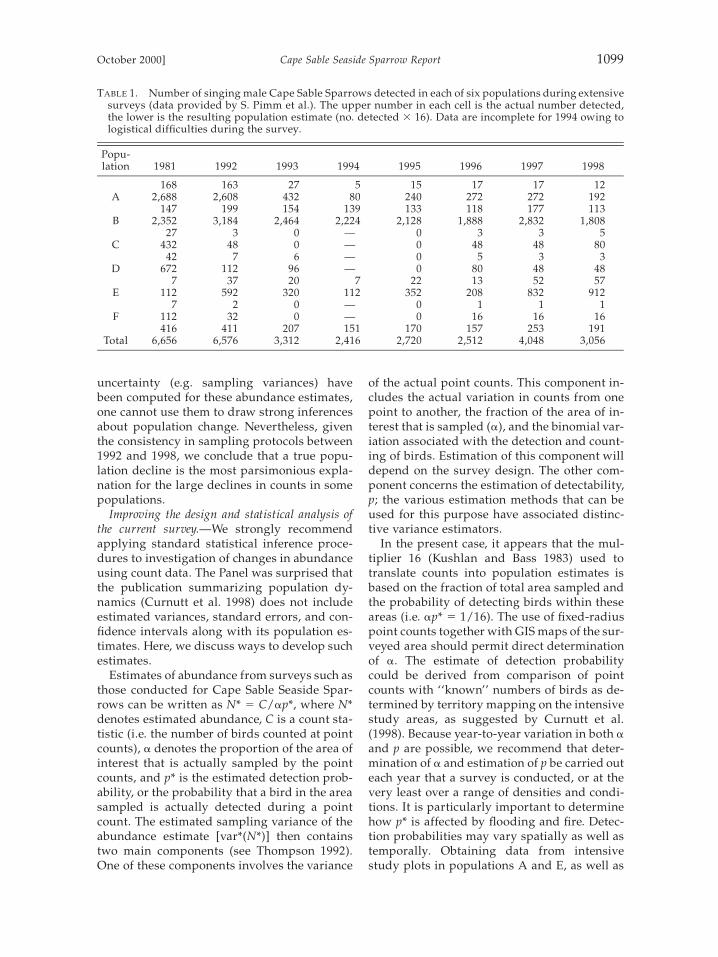
Published patterns.—As in most studies of avi-an populations, the number of adult individu-als forms the core statistic for inferences con-cerning population health. Since the 1970s,studies of Cape Sable Seaside Sparrows havefocused primarily on extensive counts of sing-ing males, conducted at 600 to 800 points thatare accessed primarily by helicopter and thatcover all of the known appropriate habitat forthe subspecies (Curnutt et al. 1998). Periodical-ly, sites not known to support sparrows alsohave been checked for the presence of singingbirds during the appropriate season. Censuspoints are distributed systematically at grid in-tersections with 1 km between points. Eachpoint is surveyed once during the morninghours (0630 to 0930) during the season of peakbreeding activity (mid-March through May).The census protocol was established by Kush-lan and Bass (1983) for the 1981 survey usingthe same helicopter-based approach followedby Werner and Woolfenden (1983) in the mid-1970s, and was used consistently for subse-quent surveys between 1992 and 1998. The onlychange in protocol was to shift from a nonran-dom order of visiting sites in 1981 and 1992 to1994, to a stratified random sampling order for1995 to present. The total population of terri-torial sparrows has been estimated by multi-plying the field counts by 16. This number wasoriginally based (Kushlan and Bass 1983) onthe need to multiply by two to account for fe-males and by eight to account for the area be-tween census points not covered in the survey.This estimation factor was found to be approx-imately correct when point counts were com-pared with actual mapped territories on inten-sive study plots (Curnutt et al. 1998).
Survey points are distributed across threedifferent areas from which Cape Sable SeasideSparrows have been reported in recent de-cades. The ‘‘Ingraham’’ population (PopulationB) was discovered in the 1970s along with theother small populations (C to F) in the easternEverglades. Since at least the 1950s, a largenumber of birds, originally referred to as thesouthern Big Cypress population, has beenknown to be scattered through the prairiesnorthwest of Shark River Slough (PopulationA). All of these areas, A to F (Fig. 1), werethought to contain substantial numbers of
birds in the mid- to late 1970s prior to the ini-tiation of the full population surveys describedabove. No evidence exists that more than ahandful of birds has occurred in recent decadesin either of the earliest known sites, the Ocho-pee prairies and the Spartina bakeri marshes ofCape Sable (Kushlan et al. 1982, USFWS 1998).These latter two populations are considered ex-tirpated.
Several estimates derived from counts ofsinging males are critical to management de-cisions: (1) the global breeding population (inthis case, number of territorial adults duringthe breeding season); (2) trends in the globalbreeding population; (3) spatial distribution ofindividuals, particularly as they are distribut-ed among the recognized populations (A to F);and (4) the pattern of change in the local pop-ulations. Some results from the surveys (Table1) are that (1) the estimated global breedingpopulation declined from approximately 6,500in 1981 and 1992 to approximately 3,000 (range2,416 to 4,048) from 1993 to 1998; (2) recentlythe only populations estimated to include morethan 200 adults are Population B (estimated at1,800 birds in 1998) and one of the three sub-populations of the northeast (E, estimated at900 birds in 1998); and (3) the other four pop-ulations are estimated to be much reducedfrom 1981. In the case of the western popula-tion (A), the estimated decline from more than2,500 to a few hundred individuals occurredafter the 1992 breeding season. Estimates forseveral of the populations (but not PopulationA) increased markedly between 1996 and 1998.
Declines in numbers of singing males count-ed were precipitous, and counts remained wellbelow initial (i.e. 1992) levels for a number ofyears after the decline. The Panel can proposeno credible confounding factors to explain asartifacts the large changes in numbers of sing-ing males detected in these standardized sur-veys. It is possible that systematic changes indetection probability occurred, brought aboutby changes in bird behavior as a consequenceof altered habitat condition. However, we con-sider this explanation to be substantially lessparsimonious than a genuine decline in num-ber of territorial males. Numbers of birds ob-served remained low even in years when con-ditions for detection were favorable (e.g. 1997),that is, when males would be expected to be ac-tive and conspicuous. Because no measures of
October 2000] 1099Cape Sable Seaside Sparrow Report
TABLE 1. Number of singing male Cape Sable Sparrows detected in each of six populations during extensivesurveys (data provided by S. Pimm et al.). The upper number in each cell is the actual number detected,the lower is the resulting population estimate (no. detected 3 16). Data are incomplete for 1994 owing tologistical difficulties during the survey.
Popu-lation 1981 1992 1993 1994 1995 1996 1997 1998
A168
2,688147
1632,608
199
27432154
580
139
15240133
17272118
17272177
12192113
B 2,35227
3,1843
2,4640
2,224—
2,1280
1,8883
2,8323
1,8085
C 43242
487
06
——
00
485
483
803
D 6727
11237
9620
—7
022
8013
4852
4857
E 1127
5922
3200
112—
3520
2081
8321
9121
F 112416
32411
0207
—151
0170
16157
16253
16191
Total 6,656 6,576 3,312 2,416 2,720 2,512 4,048 3,056
uncertainty (e.g. sampling variances) havebeen computed for these abundance estimates,one cannot use them to draw strong inferencesabout population change. Nevertheless, giventhe consistency in sampling protocols between1992 and 1998, we conclude that a true popu-lation decline is the most parsimonious expla-nation for the large declines in counts in somepopulations.
Improving the design and statistical analysis ofthe current survey.—We strongly recommendapplying standard statistical inference proce-dures to investigation of changes in abundanceusing count data. The Panel was surprised thatthe publication summarizing population dy-namics (Curnutt et al. 1998) does not includeestimated variances, standard errors, and con-fidence intervals along with its population es-timates. Here, we discuss ways to develop suchestimates.
Estimates of abundance from surveys such asthose conducted for Cape Sable Seaside Spar-rows can be written as N* 5 C/ap*, where N*denotes estimated abundance, C is a count sta-tistic (i.e. the number of birds counted at pointcounts), a denotes the proportion of the area ofinterest that is actually sampled by the pointcounts, and p* is the estimated detection prob-ability, or the probability that a bird in the areasampled is actually detected during a pointcount. The estimated sampling variance of theabundance estimate [var*(N*)] then containstwo main components (see Thompson 1992).One of these components involves the variance
of the actual point counts. This component in-cludes the actual variation in counts from onepoint to another, the fraction of the area of in-terest that is sampled (a), and the binomial var-iation associated with the detection and count-ing of birds. Estimation of this component willdepend on the survey design. The other com-ponent concerns the estimation of detectability,p; the various estimation methods that can beused for this purpose have associated distinc-tive variance estimators.
In the present case, it appears that the mul-tiplier 16 (Kushlan and Bass 1983) used totranslate counts into population estimates isbased on the fraction of total area sampled andthe probability of detecting birds within theseareas (i.e. ap* 5 1/16). The use of fixed-radiuspoint counts together with GIS maps of the sur-veyed area should permit direct determinationof a. The estimate of detection probabilitycould be derived from comparison of pointcounts with ‘‘known’’ numbers of birds as de-termined by territory mapping on the intensivestudy areas, as suggested by Curnutt et al.(1998). Because year-to-year variation in both aand p are possible, we recommend that deter-mination of a and estimation of p be carried outeach year that a survey is conducted, or at thevery least over a range of densities and condi-tions. It is particularly important to determinehow p* is affected by flooding and fire. Detec-tion probabilities may vary spatially as well astemporally. Obtaining data from intensivestudy plots in populations A and E, as well as

1100 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
existing plots in population B, is essential toevaluating the possibility of spatial variation inp*.
Estimation of var*(N*) involves two compo-nents as noted above. The first is based on thespatial or point-to-point variance in numbersof birds counted. Estimation of this componentfor the sparrow survey is not straightforwardbecause of the systematic sampling design, butseveral reasonable approaches are available(e.g. Cochran 1977, Thompson 1992). This com-ponent should not be based on an assumed un-derlying distribution (e.g. Poisson was as-sumed for discussion purposes in one of the re-view documents) but should be computed fromthe raw count data. The second component,var*(p*), will be based on the method used toestimate detection probability. If detectionprobability is estimated as a ratio of pointcounts to ‘‘known’’ numbers of birds (based onterritory mapping), then the variance of this es-timate can be obtained using standard expres-sions for the variance of a ratio estimator (e.g.Thompson 1992) that depend on details abouthow the data are aggregated for estimation.
In summary, although the count data ob-tained through reasonably consistent method-ology suggest substantial declines in sparrowabundance, the absence of estimates of preci-sion demands that care be exercised in inter-preting these data. Raw data obtained fromsurveys in 1981 and 1992 to 1997 can be usedto compute population estimates and relatedestimates of precision for each year of the sur-vey. We recommend that such estimation becarried out, because resulting estimates of pre-cision will be useful in providing an improvedability to assess the magnitude of historicalpopulation changes, as well as any changes inthe future.
General design recommendations for future sur-veys.—Besides computation of estimates of pre-cision based on existing data, the Panel sug-gests that several other changes be considered.Some involve the sampling of space. For ex-ample, controversy exists over potential spar-row habitat that was not sampled from 1992 to1997 (W. Post pers. comm.). Although no spe-cific areas of suitable but unsurveyed habitathave been identified, Post suggests that suchareas occur along the inland border of the man-grove zone in extreme western Everglades. Oneapproach to dealing with such a possibility
would involve stratification of a large area ofpotential habitat into adequate (i.e. likely tocontain some sparrows) and poor (unlikely tocontain sparrows) habitat strata. The ‘‘ade-quate habitat’’ stratum could be sampled athigh intensity (i.e. a would be high for this stra-tum), whereas the ‘‘poor habitat’’ stratumwould be sampled at a much lower intensity, re-flecting the low probability of birds inhabitingthis stratum.
Another consideration is the sampling de-sign itself. Historically, surveys have beenbased on a systematic design with point countsconducted at either 0.8 km or 1 km perpendic-ular distances in a checkerboard pattern. Sucha design was selected because information onlarge-scale bird distribution was considered tobe an important product of the survey (J. Kush-lan pers. comm.). If abundance estimation nowis the primary goal of the survey effort, how-ever, then a simple, random, or stratified ran-dom design might be more efficient and wouldpermit straightforward estimation of varianc-es. If continued collection of information aboutlarge-scale distribution remains a priority, thenthe current systematic design should be re-tained.
In addition to the sampling of space, it alsois possible to consider ways of estimating de-tection probability. If territory mapping is to becontinued in the future for reasons other thanestimation of p, and if it is sensible to regardbird numbers obtained from such exercises asapproaching true numbers, then it is reason-able to take advantage of these data for esti-mation of detection probability. Details of thiscalibration as described by Curnutt et al. (1998)are not entirely clear to us (e.g. it was not clearwhy the average of counts at four different sur-vey points was compared with the numbers ofbirds indicated by territory mapping on a sin-gle, intensive study plot). In any case, we rec-ommend that standard point counts be con-ducted each year within intensive study plotsin which actual bird abundance is documentedthrough territory mapping, so that direct esti-mation of detection probability is possible. Ifterritory mapping on intensive study plots is tobe discontinued, or it is not thought to providean accurate estimate of abundance, then othermethods for estimating detection probability,and hence bird abundance or density, may mer-it consideration. Distance sampling using var-

October 2000] 1101Cape Sable Seaside Sparrow Report
iable circular plots (Buckland et al. 1993) anddouble-observer sampling (original aerial sur-vey approach of Cook and Jacobson [1979]modified for point counts by Nichols et al.[2000]) both provide reasonable approaches toestimating the probability of detecting singlemales. (They do not, however, address theproblem of detecting birds that do not sing, seebelow.) We note that detection probability can-not be estimated well simply from replicatecounts (by the same or different persons) at thesame point(s).
We strongly recommend that estimation ofsampling variances accompany future abun-dance estimates, and that estimation of detec-tion probability be incorporated directly intothe survey design. Sparrow surveys clearly re-quire substantial expense and effort, and it is areasonable expectation that these efforts yieldinferences about variation in sparrow abun-dance over space and time. Such inferencesshould incorporate estimates of precision thatproperly account for possible spatial variationin counts, as well as detection probability andvariation associated with its estimation.
Beyond singing male surveys.—Three specificconcerns have been raised about reliance onsurveys of singing males (W. Post pers. comm.).First, singing is highly affected by both endog-enous and environmental factors, potentiallymaking detection probabilities low and vari-able. To partially address this concern, the Pan-el recommends that playback of song or othervocalizations, which is a standard method insingle-species studies, be incorporated into thesurvey work. This could be accomplished byadding a short playback session at the end ofeach standard point count. For example, appli-cation of playback to standard 10-min pointcounts has been shown to double the detectionprobability for forest birds such as Scarlet Tan-agers (Piranga olivacea; K. Rosenberg, J. Lowe,and A. Dhondt unpubl. data). Such a designwould allow for the continued gathering ofdata comparable to earlier parts of the study.We further recommend that playback be inves-tigated in pilot studies of variance in detectionprobability conducted in intensive study plots.
Second, the adult population at any one timemay include birds that do not attempt to breedbut nonetheless are alive and part of the futurebreeding population (i.e. floaters). The impor-tance of understanding the population dynam-
ics of floaters has been emphasized by Verner(1985) and others. In the Cape Sable SeasideSparrow, floaters may be more numerous inyears in which suitable habitat is reduced or atnon-peak periods of activity during the breed-ing season.
Third, the number of males attempting to at-tract mates may not indicate the number of fe-males actually engaged in reproductive activ-ity. In fact, singing rates actually may be higherfor unmated birds (Post 1974), resulting in tem-poral changes in detection probability associ-ated with changes in the proportion of birdsthat are paired. For most demographic models,the number of breeding females, not territorialmales, is the most relevant statistic.
The so-called floater question is amenable toinvestigation through color marking of individ-uals on intensive study plots. Flush-nettingmethods and radio tracking of individuals mayhelp determine the size of any nonbreedingcomponent of the population. If numbers offloaters can be estimated reasonably on the in-tensive study plots, then it would be possibleto include this component in the estimation ofdetection probability. The size of a floater com-ponent, and its variation in response to tem-poral changes in habitat quality, are importantto population modeling (see below), as well asto interpreting variation in the size of thebreeding population.
Reliance upon singing males rather than onfemales, although common in avian surveywork, is problematic. In the case of the Cape Sa-ble Seaside Sparrow, it may be possible to mon-itor the number of nesting females over largeareas to complement surveys of singing males.Females of at least one other subspecies of Sea-side Sparrow are known to give a ‘‘nest depar-ture’’ call (McDonald and Greenberg 1991), i.e.females utter a series of chip notes almost everytime they leave the nest from bouts of broodingor incubation. This call is given throughout theday and under a wide range of conditions. Nestdeparture often follows a predictable scheduleduring incubation, and determining the depar-ture schedules of females may enable the de-sign of a vocalization-based survey that pro-vides a more direct index of nesting activityand permits estimation of female detectionprobability and abundance. Effort should bemade to determine if female Cape Sable SeasideSparrows reliably display this behavior and

1102 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
whether this behavior could serve as the basisfor improved estimation of the breeding pop-ulation.
Conclusions about population trends.—Havingexplored ways to improve survey techniques,the Panel emphasizes that we view as parsi-monious and reasonable the conclusion thatPopulation A experienced a dramatic declineduring the 1990s, and that eastern PopulationsC, D, and F are smaller now than they were in1981. However, we strongly recommend thatpoint-count data be analyzed statistically usingan approach that accounts for relevant sourcesof variation (spatial variation, variable and un-known detection probabilities), because resultsof such analyses will provide a basis for stron-ger scientific inferences than can be made atpresent.
Future monitoring should be extensive andshould provide coverage of existing potentialhabitat. Given the potential for Cape Sable Sea-side Sparrows to shift breeding locations,broad geographic coverage is essential. Futuremonitoring also should deal explicitly with un-certainty in point-count data associated withspatial variation in bird density and variationin detection probability. The Panel envisions adouble-sampling approach in which intensivestudy plots are established at a subset of thepoints selected for sampling in the extensivesurvey. Work on these intensive plots would beused not only to estimate detection probability,but also to assess numbers of nonbreedingfloaters, to explore the incorporation of femalenest departure calls into the survey design, andpossibly to estimate vital rates using markedindividuals and nests. Intensive study plots al-ready have been established (S. Pimm pers.comm.), and some of the above data have beengathered from them. The Panel recommendsthat this effort be expanded and increased asoutlined above.
CAUSES OF POPULATION CHANGES
The Panel considered several proposed ex-planations for changes in the distribution andnumbers of Cape Sable Seaside Sparrows. Weaddress them here in sequence, beginning withthose we view as most likely to have affectedpopulation changes in the past 10 years.
Flooding.—A wealth of published informa-tion (e.g. Kushlan et al. 1982, Werner and Wool-
fenden 1983, Curnutt et al. 1998, Nott et al.1998) indicates that the Cape Sable SeasideSparrow is extremely sensitive to variations inthe nature and quality of its breeding habitat.It was clear to the Panel, both from the litera-ture and on the site visit, that relatively smallchanges in hydrology, especially water depthand hydroperiod, can produce critical changesin habitat quality that are visible even to the un-trained eye. A fundamental feature of the sub-species’ biology—nest placement—causes thissensitivity. Cape Sable Seaside Sparrows placetheir nests just above the bases of clumps ofwet-prairie vegetation (most often, now atleast, in tussocks of muhly grass [Muhlenbergiafilipes]; Werner 1975). Placement of a nest toolow in vegetation exposes the nest to the ever-present risk of flooding. If the nest is placed toohigh, the vegetation is too sparse to support thenest or to shield it from predators and the ele-ments.
Uncertainty exists over precisely how highabove ground (and especially standing water)the sparrows will build nests. Nott et al. (1998)cite a ‘‘10 cm rule’’ for maximum water depthover which the birds will initiate nesting, basedon absence of singing activity during surveyswhen water depth exceeded that level. In mod-eling population dynamics, Nott (1998:81) usesa 5-cm threshold of water depth for male sing-ing and nest building. Even the small sampleof nests analyzed by Dean and Morrison (1998)is sufficient to demonstrate that some nestingmay occur when average water depth exceedsthese thresholds. Thus, the Panel recognizessome risk in over-interpreting a hard criterionfor acceptable water levels when these are mea-sured over a grid of 0.5 to 1 km in scale. Becauseof variability in topography and vegetation, ap-parently some suitable nesting sites for spar-rows may remain in generally inundated hab-itat, and these sites can be essentially invisibleto large-scale measurements of water depth.
The Panel reached two conclusions about thisproblem. (1) The relation between water leveland nesting activity deserves further research,mainly to better define the correlation betweenwater levels measured at large scales and spar-row productivity. (2) A better understanding isnecessary to evaluate the effects of water levelduring the 1990s. The 1990s represent an un-usually rainy period in the Everglades, andduring this period managers frequently were

October 2000] 1103Cape Sable Seaside Sparrow Report
TABLE 2. Percent of habitat available for brood production for Cape Sable Seaside Sparrow population A.
1977 to 1992
One brood Two broods
1993 to 1996
One brood Two broods
n (years)MedianMeanSDRange
16917429
15–100
16364428
0–100
4161614
0–33
4578
0–18
concerned about high water levels. Data pro-vided by Nott et al. (1998) allow one to assessthe scale of water-level fluctuations during thisperiod and their likely effects on sparrow re-production. Although one might quibble aboutdetails and causes, it is clear from these datathat the magnitude and duration of floodingchanged substantially west of Shark RiverSlough, and that these changes likely had a ma-jor influence on Population A.
The case for massive disruption of sparrowbreeding because of flooding rests on severalimplications in Nott et al. (1998), especiallydata provided in their figure 2 and table 1. (Weassumed that the expressions ‘‘dry’’ and ‘‘wet’’used in reference to water levels describe levelsbelow or above, respectively, a hard criterionsuch as 5 cm or 10 cm, because these are else-where stated to be critical for sparrow breed-ing. We also assumed that the authors correctlyassessed such conditions as permitting or notpermitting breeding, respectively.) Nott et al.(1998: table 1) analyzed topographic variationin Area A by comparing water depths at 284census points in 1995 with water levels at mon-itoring station NP205 taken on the same date.Table 2 summarizes the percent of Area A thatthese authors deemed to be sufficiently dry fora sufficiently long period (see caveat above) topermit the production of one or two broods be-tween 1977 and 1996.
Persistently high water levels over the period1992 to 1996 were sufficient to have nearly pre-cluded successful breeding during both 1993and 1995. Even during 1994 and 1996, accept-able water depths were available for first clutch-es in less than one-third of the area (27 and33%, respectively), and almost no habitat re-mained at acceptable water levels to allow sec-ond broods. Over the 16-year period preceding1993, typically 70 to 90% of Area A was avail-able for first broods and 35 to 45% for secondbroods (Table 2). Comparable percentages from
1993 to 1996 were 15% and 4 to 7%. Even as-suming perfectly synchronous onset of breed-ing during these years, and allowing for mi-crosite variation that would have permittedlimited nesting when the area as a whole wasunsuitable, productivity of Population A from1993 to 1996 would have been only a small frac-tion of its average during the previous 15 years.
Deep water in Area A during the period ofinundation between 1993 and 1996 rendersmoot the question of precisely which habitat ac-ceptability rule one applies based on nestheights. During this period, water levels inmost of the area were well above the acceptablerange. Data in Lockwood et al. (1997) suggestthat even where sites are available for nesting,high water levels may result in elevated pre-dation and hence reduced nesting success. Ev-idence for this further effect of flooding is in-conclusive, however, in contrast to evidencethat reproduction is reduced when water levelsexceed typical nesting heights.
In short, we find the evidence convincing thatsuccessful breeding in Population A was sub-stantially reduced throughout the period from1993 to 1996 compared with earlier years, andreproduction may have been essentially nilduring at least two of these years (1993 and1995). For a small, sedentary songbird, such asignificant reduction in reproductive outputcan be expected to result in reduction of overallpopulation size.
Even before the manipulations by the Corpsof Engineers, the Everglades ecosystem expe-rienced substantial temporal variation in rain-fall and water levels (Blake 1980, Beissinger1986, Beissinger and Gibbs 1993, Deuver et al.1994). Is it likely that increased water levels,which appear to have caused cessation of nest-ing and concomitant decline in Population A,resulted simply from natural variation in cli-mate? Elevated water levels in the area of Pop-ulation A occurred during a series of uncom-

1104 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
monly wet years in South Florida, and no doubtthis contributed greatly to the problem.
Nott et al. (1998) attempted to isolate the ef-fects of water discharges into Everglades Na-tional Park through the S-12 structures by se-quentially removing through regression the ef-fects of base water level at the beginning of theseason and rainfall during the season. Residualwater depth after removing these two factorswas significantly correlated with dischargesfrom S-12A. Contributions of discharges fromthe remaining three S-12 structures were notexamined.
Although correlational analyses of this sortdo not yield strong inferences, the Panel viewsas reasonable Nott et al.’s (1998) conclusion thatthe concentrated releases of water from the S-12 structures from 1992 to 1995, above and be-yond existing water depth and seasonal rain-fall, directly led to the deep-water conditionswest of Shark River Slough. These in turn prob-ably caused habitat in the range of PopulationA to be unsuitable for breeding, and we con-clude that this likely played a major role in theapparent decline of Population A.
Good evidence also exists that extended hy-droperiods result in vegetation changes that re-duce habitat suitability for the sparrows, spe-cifically the conversion of prairies or marshesdominated by muhly to those dominated bysawgrass (Kushlan et al. 1982, Armentano et al.1995, Nott et al. 1998). This may have played arole in the decline of Population A, but it is bet-ter illustrated by changes in Population D in theTaylor Slough region. In fact, Kushlan et al.(1982) predicted that such a habitat conversion,and accompanying decline in sparrow num-bers, would occur in Population D owing to theconstruction of a pumping station immediatelyupstream in Taylor Slough. Nott et al. (1998)presented evidence that the increased hydro-periods resulting from releases from thepumping station have indeed resulted in thepredicted change in vegetation. Survey resultsprovide evidence of the predicted decline of thesparrow population (Table 1).
In some ways, effects of extended hydroper-iod on vegetation may be more important thaneffects on sparrow reproduction. That saw-grass-dominated prairie is unsuitable habitatfor sparrows is evident from Nott’s (1998) workon habitat use as a function of vegetative com-position generally, as well as information about
Populations A and D specifically (Nott et al.1998). Muhly prairie can withstand deep in-undation, and even whole years with little or nodry period, as long as these are interspersedwith years in which winter drying occurs(Kushlan et al. 1982). Because muhly prairiecan recover following a return to shorter hy-droperiods (Nott 1998), chronic alteration ofhydroperiod rather than occasional floodevents are the most problematic in bringingabout changes in vegetation. If water manage-ment produces long hydroperiods in Area Afrequently enough to alter its vegetation, as hasoccurred in Area D, then sparrow survival andreproductive rates will be moot because thehabitat will be unable to support successful re-production regardless of how many birdsmight be in the area.
Fire.—Compared with the role of water lev-els, the effects of fire on population sizesthrough the 1990s have been minor. We agree,however, that the potential capacity of fire to af-fect sparrow populations and habitats remainslarge. No catastrophic population declines inthe past decade can be directly ascribed to fire.Nevertheless, it appears likely that populationsin northeastern Everglades National Park, es-pecially Population F, may be prevented fromgrowing by the extremely high frequency of ar-son-caused fires during the dry season (Cur-nutt et al. 1998). The Ingraham fire burned partof the area occupied by population B in 1989.Survey data only three years later (1992) pro-vide evidence that this major fire did not havea catastrophic effect on Population B (Table 1).
The Panel concludes that in the short term,loss of populations to catastrophic fire is pos-sible but unlikely, even under current watermanagement practices. Risk of catastrophic firewill be greatly reduced by increased flows intonortheastern Shark River Slough, and the po-tential for catastrophe increases the longerthese larger flows are delayed. When fires oc-cur, as with the Ingraham fire, damage is likelyto affect a portion of a population rather thaneliminate all of it. However, fire does have po-tentially important effects on population dy-namics through altering habitat suitability.Curnutt et al. (1998) provide convincing evi-dence that fires are much more common in ar-eas at the eastern edge of Everglades NationalPark where the habitat frequently is dry, andwhere exposure to humans is especially great.

October 2000] 1105Cape Sable Seaside Sparrow Report
At issue is whether such effects played a sig-nificant role in the decline of the small north-eastern populations (C and F) and whether theymight threaten the larger one (E). Unfortunate-ly, the survey technique is not sensitive enoughto permit a direct approach to this question.
Disagreement exists about the effects of fireon sparrow populations. Abundant evidencepoints to a short-term negative effect of burn-ing on sparrow numbers (Werner 1975, Taylor1983, Werner and Woolfenden 1983, Curnutt etal. 1998). Burned habitat is avoided for up to ayear, and numbers then increase over the nextseveral years. There are conflicting reportsabout the duration of the effect and subsequentdeterioration of habitat in the continued ab-sence of fire. Werner (1975) suggested that useof otherwise suitable habitat declines to aban-donment after six years without fire. However,Taylor (1983) reported sparrows in sites thatwere unburned for more than a decade. Cur-nutt et al. (1998) pointed out that many birds inthe largest and most productive population (B)nested during the mid- to late 1990s in areasthat had not burned since the Ingraham fire in1989. Curnutt et al.’s (1998) cross-sectionalanalysis suggests that sparrow numbers in-crease for up to 10 years after fire. However,their analysis (see figure 5) was handicapped inthat it did not control for site effects. Many dif-ferences besides fire frequency and time sincelast fire exist among sites that were compared.Moreover, little question exists that at sometimescale, fire is necessary for continued oc-cupancy of a site by Cape Sable Seaside Spar-rows because it inhibits invasion by woodyplants (Craighead 1971). The aversion of spar-rows to woody vegetation in their nesting hab-itat, and resulting loss of habitat to invasion ofwoody vegetation, is well documented (Werner1975, Werner and Woolfenden 1983, Nott 1998).
Numerous factors complicate analysis of theeffects of fire frequency on habitat and sparrowpopulations. Areas with deeper soil appear torecover more rapidly from a fire (Taylor 1983,Curnutt et al. 1998) and to build up excess fuelsooner (A. Mayer unpubl. data). Fires set by hu-mans in the dry season often burn wider areasmore intensively than do lightning-caused firesduring the wet season.
The long-term role of fire should be investi-gated through a series of longitudinal studieswithin sites of known fire history having dif-
ferent hydrology and soil depths. Use of pre-scribed burning as an experimental treatmentlikely will be necessary to accomplish this. Re-cently established intensive study plots in pop-ulation E could be employed for this purpose.The Panel recommends a collaborative effortinvolving plant ecologists, ornithologists, andhabitat managers, and employing intentionalburns in the experimental design, to identifyhow soil, hydrology, and fire interact in affect-ing the dynamics of Cape Sable Seaside Spar-row populations.
Other possibilities.—One interpretation of thehistorical record of habitat distribution of theCape Sable Seaside Sparrow is that the currentplight of the sparrow results from its extirpa-tion from the now largely nonexistent Spartinabakeri habitats that once occurred along the Ev-erglades-marine interface at the turn of the cen-tury (W. Post pers. comm.). According to thishypothesis, the population might be larger andmore productive if the habitat that it once oc-cupied on Cape Sable was still available andsparrows still lived there. However, the recentproductivity of birds in good habitat (Lock-wood et al. 1997, Dean and Morrison 1998) isfully comparable to that of other populations ofthis species (e.g. Post et al. 1983) and appearsadequate to support a thriving population. Thedata required to determine how demographyin Spartina prairie compares with that in muhlyprairie do not (and probably never will) exist,but no direct evidence of any kind exists thatmuhly prairie represents suboptimal habitat.
All known threats to the Cape Sable SeasideSparrow involve alteration of habitat. The Panelencountered no evidence that the bird’s popu-lation is being affected by other biotic factors(e.g. unusual new predators, diseases, or com-petitors) or abiotic factors. The Panel explicitlyconsidered the possibility that Hurricane An-drew, which roared through most of the rangeof the sparrow in 1992, caused the decline, es-pecially in Population A. However, we findCornutt et al.’s (1998) arguments that Andrewwas not a primary factor in the decline of Pop-ulation A to be reasonable. Most important,Population A continued to decline for years af-ter Andrew, whereas Population B receivedonly slightly less extreme wind conditions thandid Population A, but exhibited no decline.
Conclusions.—The Panel concludes that pop-ulation declines of the Cape Sable Seaside Spar-

1106 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
row occurred largely as a result of reducedhabitat suitability. Habitat degradation owingto invasion by exotics and by woody vegeta-tion, which was of great concern in the early1980s (Kushlan et al. 1982), does not appear tothreaten areas where sparrows live currently.In contrast, the other threats to habitat qualityidentified in the 1980s, fire and flooding (Kush-lan et al. 1982), indeed appear to have wroughtthe damage it was feared they might. Declinesof Populations A and D likely can be attributedto extended hydroperiods that suppressed re-production and produced adverse changes invegetation. Populations C and F appear to bedepressed by reductions in habitat quality re-sulting from fire. Abnormally high fire fre-quency in these areas is the direct result of re-duced hydroperiods and proximity to humans.
RISK OF EXTINCTION
The current risk of extinction of the Cape Sa-ble Seaside Sparrow needs to be assessed care-fully because of recent declines of some of itspopulations, and because of the recent extinc-tion of its close relative, the Dusky SeasideSparrow. We will discuss extinction risk withrespect to current water management andplans for new water management described inthe Introduction. Because the new plan stipu-lates that some of the water currently held inWCA 3A will instead flow through WCA 3Binto northeastern Shark River Slough, we as-sume that Populations A and D will experienceextended hydroperiods less often under thenew plan than under current management. Wefurther assume that Populations C, E, and Fwill experience dry conditions less often.
Although the new water management plan isscheduled to be implemented in only five years,many obstacles to its implementation exist (e.g.land acquisition, construction of new struc-tures), and if the previous 50 years of Ever-glades history are any indication, delays arelikely. Therefore, risk of extinction in the inter-im cannot be ignored. The key element for thesparrows is that flow patterns produce hydro-periods similar to those that characterized themarl prairies prior to management of water byhumans. Flows west of Shark River Slough andinto Taylor Slough must be reduced, and flowsinto northeastern Shark River Slough in-creased, to accomplish this. The adequacy of
the new water management plan depends onthe extent to which the desired hydroperiodsare actually produced. In the interim, the bestalternative to reduce risk of extinction of thesparrow is to restore the required flow patternsto the maximum extent possible using existingstructures. Recognizing the reluctance of man-agers to adopt this alternative, we evaluate riskof extinction over the short term for two addi-tional scenarios.
The first scenario is ‘‘status quo’’ manage-ment. We consider the data collected during thelast two decades to reflect biological conse-quences of status quo management. The secondscenario is to retain water in WCA 3A in wetyears rather than allow emergency releasesthrough the S-12 structures into the area westof Shark River Slough. We assume in this sec-ond scenario that Population A would not besubjected to extended hydroperiods such asthose that occurred from 1993 to 1996, whereasin the first scenario these events will continueto occur when rainfall is sufficient. We furtherassume that Populations C, E, and F will con-tinue to experience dry conditions unusuallyfrequently in both short-term scenarios.
Here, we review aspects of demography rel-evant to extinction risk and then evaluate pre-vious efforts to model extinction risk. Finally,we address the following questions: (1) Will theplanned changes in flow patterns result in min-imal extinction risk for the Cape Sable SeasideSparrow? (2) What is the relative risk resultingfrom the two short-term management scenar-ios? Components of the second question in-clude the risk to Populations C, E, and F re-sulting from unusually frequent and pro-longed dry conditions, the risk to PopulationsA and D of continued releases of water intotheir habitat, and the risk to Population B of cat-astrophic fire.
Key features of demography.—To evaluate thepotential threat that factors such as floodingand fire represent to the continued existence ofthe Cape Sable Seaside Sparrow, we mustgauge the sparrow’s capacity to respond tothese threats. Many other species in the Ever-glades ecosystem (e.g. Snail Kites [Rostrhamussociabilis] Beissinger 1986, Takekawa and Beis-singer 1989; Wood Storks [Mycteria americana]Ogden 1996) have the capacity to move consid-erable distances in response to degradation ofhabitat. If Cape Sable Seaside Sparrows were

October 2000] 1107Cape Sable Seaside Sparrow Report
facultatively nomadic, they might be able tomove to better conditions when flooding or firedegrades their habitat, perhaps returningwhen conditions improve. Indeed, Post andGreenlaw (1994) refer to the Seaside Sparrowas a catastrophe-prone species with such abili-ties. Thus, it is especially important to examinedispersal behavior in evaluating extinctionrisk.
The history of the patchy and apparently dy-namic distribution of the Cape Sable SeasideSparrow (e.g. Kushlan et al. 1982) suggestssome ability to colonize newly available habitatas traditional areas become unsuitable. Infer-ences from historical distributions, however,must be weighed against the finding by Deanand Morrison (1998) that radio-tagged birds,even juveniles, show limited dispersal. ThePanel was impressed by this team’s abilities toaccount for and maintain radio-tracked birdsthrough many cycles of transmitter replace-ment and bird movements of up to 7 km (T.Dean unpubl. data). Telemetry data suggestthat when habitat remains in good conditionthe birds are quite sedentary, and that even dis-persing juveniles only rarely cross the kinds ofbarriers that separate different populations(Dean and Morrison 1998). Sample size for ju-venile birds remains small, however (n 5 11; T.Dean unpubl. data). Another piece of evidencethat adult sparrows have limited capacity forlarge-scale movements is that no compensatingincrease occurred in any of the other popula-tions after flooding of the habitat of PopulationA. The birds that disappeared from PopulationA did not appear to move elsewhere en masse.
Although no evidence of rapid shifts in dis-tribution through mass movements exists,gradual shifts could occur through populationextinction and recolonization events. Withoutbetter information on survival during floodingand fire episodes (see below), it is not possibleto determine precisely how prone the Cape Sa-ble Seaside Sparrow might be to local extinc-tion events. The historical extinctions at CapeSable and Ochopee involved changes in vege-tation that rendered habitat unsuitable (Kush-lan et al. 1982); hence, they have little bearingon assessing the vulnerability of sparrow pop-ulations to stochastic fluctuations in vital rates.The accuracy of population estimation is notknown, so it is not possible to determinewhether recent apparent extinction and recol-
onization events involving small populations(i.e. C, D, and F; see Table 1) were real or re-flected sampling error.
Telemetry data suggest that recolonizationability is limited in terms of distance and fre-quency. No movements between populationshave been observed, nor have movements beenobserved beyond barriers such as tree islandsand sloughs, although movements over contin-uous habitat of distances as large as those thatseparate Populations B to F have been docu-mented (Dean and Morrison 1998). The avail-able data suggest that emigration from goodhabitat probably is not frequent enough for oneto assume with any confidence that the variouspopulations are linked as a single metapopu-lation, in which populations are routinely ‘‘res-cued’’ from extinction by emigration from oth-er populations (Stacey and Taper 1992). How-ever, occasional movements that could result inrecolonization cannot be ruled out.
More problematic are movement patterns inresponse to adverse conditions, such as fires orfloods. Movement patterns are known tochange in response to such events in other Sea-side Sparrows (Post and Greenlaw 1994). Un-fortunately, no relevant data exist for the CapeSable Seaside Sparrow. Moreover, its distinctivehabitat and nonmigratory tendencies make iteasy to imagine that this population does notshare these habits with more northerly ones. Itmay be possible to evaluate the degree of iso-lation among the various Cape Sable SeasideSparrow populations by examining genetic re-lationships among them. If sparrows are to betrapped for other research purposes, thenblood, and perhaps tissue, should be taken topursue this possibility.
We further recommend that experimentalstudies be conducted in which movements ofindividuals following burning and flooding aremonitored using telemetry. In the case of fire,this could be incorporated into the longitudinalstudies using prescribed fire recommendedabove. In the case of floods, the study could becoordinated with planned water releases.These studies would allow researchers to mea-sure for the first time the movements of birdsfaced with degraded habitat. To date, all telem-etry work has been done in the relatively stableand suitable Ingraham population area. Thesestudies would also enable evaluation of varia-tion in detection probabilities (see above) and

1108 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
survival (see below) under adverse (but not un-common) conditions.
Limited dispersal and a dynamic spatial his-tory need not represent a paradox. Over longtime scales, rare long-distance movements ofjuveniles are capable of producing considerablerange dynamics. However, Cape Sable SeasideSparrows in the late 1990s appeared to live inan ecosystem with less total habitat, and per-haps less variety, than that in which the taxonprobably evolved. Marl prairies have been re-duced by development, chronic flooding, andinvasion of woody vegetation, and Spartinahabitat has been nearly eliminated by invasionof mangroves and changes in salinity (USDI1998, W. Post pers. comm.). Thus, whatever ca-pacity the species may have to shift its distri-bution to match changes in habitat suitability isnow constrained by limited habitat availability.The capacity for colonization of new habitatpatches probably exists, but the Panel sees noscientific rationale for gambling on a still un-proven potential for long-distance movementsas a responsible strategy for protecting thesparrow from population collapses in the fewusable habitat patches that remain.
How a species responds to adverse condi-tions depends on productivity and survivalrates within populations, as well as movementbetween populations. Species with high popu-lation growth rates potentially can recoverquickly from population declines and can ex-pand rapidly after recolonization. Nesting suc-cess of the Cape Sable Seaside Sparrow is low,but this is offset to some degree by the poten-tial for multiple brooding. Dr. Julie Lockwoodand collaborators have collected data on nest-ing success from intensive study plots (Lock-wood et al. 1997, J. Lockwood unpubl. data).Nests of this species are difficult to find and canbe logistically challenging to monitor, so sam-ple sizes are still small. Nevertheless, severalpatterns emerge. First, the Mayfield success es-timate for first nests that were initiated duringthe dry season in March was 43% (n 5 40), atypical value for a passerine. Second, nestingsuccess appeared to decline as the breedingseason advanced, a pattern seen in many avianspecies. Second broods that were initiated atthe end of the dry season in late April and Maywere less than half as likely to fledge young asfirst broods (16%, n 5 11). Nesting failure re-sulted predominantly from predation on eggs
or young, or flooding. A few individuals at-tempted third broods under highly favorableconditions (Lockwood et al. 1997). Fire andflooding can reduce the number of nesting at-tempts and, if severe enough, can terminate re-production completely. Fledging success aver-aged three young per successful nest under allconditions (Lockwood et al. 1997).
Data on survival of breeding males also areavailable from intensive study plots. Resight-ings of more than 100 birds banded from 1992to 1998 suggested a 50% survival rate (S. Pimmunpubl. data). However, this estimate was notcorrected for the probability of resighting. M.Nott (unpubl. data) recently applied Cormack-Jolly-Seber methods to estimate probabilities ofresighting and survivorship, obtaining an es-timate of 56% for survival of adult males fromthese same resighting data. This result shouldbe considered preliminary, because the analy-ses have not been published nor made availablefor examination.
Little else is known about survival rates ofCape Sable Seaside Sparrows. Data from juve-niles and females are insufficient to estimatesurvival rates accurately. Survival estimates forjuveniles represent a critical research need.Variation in survival is poorly described, evenfor adult males. Data from small samples sug-gest that survival of breeding males is substan-tially higher than 60% under favorable condi-tions (Werner and Woolfenden 1983), and sur-vival of nonbreeding males may be substan-tially lower than that of breeding males (S.Pimm unpubl. data). Annual variation and oth-er forms of heterogeneity in survival rates havenot been described. It is especially important toknow if survival is reduced by the same factorsthat reduce productivity, that is, flooding andfire. Studies of movements in response to fireand flooding recommended above also couldprovide valuable information on the effects ofthese events on survival. As data from inten-sive study plots accumulate, we also recom-mend that survival analyses examine differenc-es among individuals of different social status(i.e. breeders vs. nonbreeders) and among pop-ulations. Currently, sample sizes and data ap-pear insufficient to permit robust analyses ofthis type.
The effect that water management has on thecapacity for multiple brooding by Cape SableSeaside Sparrows is a critical issue. To further

October 2000] 1109Cape Sable Seaside Sparrow Report
TABLE 3. Estimates of lambda (geometric rate ofpopulation change) for Cape Sable Seaside Spar-rows based on various levels of annual survivaland numbers of nesting attempts per year. Popu-lations are stable when lambda 5 1 and declinewhen lambda , 1. Calculations assume age of firstbreeding at one year, three young raised per suc-cessful brood, an equal sex ratio at fledging, andthat third broods have the same success rate as sec-ond broods but are attempted by only one-third ofthe adults (Lockwood et al. 1997, Lockwood un-publ. data). Mayfield success of first and secondbroods was 43% and 16%, respectively.
No. nestingattempts/year
Survival rate
50% 55% 60%
0123
0.500.820.940.98
0.550.911.041.08
0.600.991.131.18
assess the significance of this issue for popu-lation behavior, we collated various parameterestimates to create a simple model of Cape Sa-ble Seaside Sparrow demography in the Ever-glades (Table 3). The objective of this exercisewas to evaluate the relationship between levelof multiple brooding and rate of populationchange (lambda). We developed a two-stage-class prebreeding population model with aone-year time step and (lacking data on juve-nile survival) no differences in survival be-tween juveniles and adults. Although there is alarge degree of variation in model outcomes,several conclusions emerged. First, lack of nest-ing has a dramatic effect on population behav-ior (as expected), but the opportunity to at-tempt a second brood may have a substantialeffect as well. Indeed, it is conceivable that theopportunity to attempt a second brood couldmake the difference between a declining and anincreasing population. Second, survival ratesrequired to achieve a stable population in theabsence of multiple brooding may be unreal-istically high. Even though we used juvenilesurvival rates that are probably too high, with-in the range of existing estimates of adult sur-vival, lamda only exceeds one if there are mul-tiple broods (Table 3). Recall, however, that un-der favorable conditions, adult survival occa-sionally might be higher than the values weused in the model.
If reproduction and survival are correlated(i.e. good years have high reproduction andhigh survival, and bad years suffer the reverse),
then sparrow populations are capable of a‘‘boom and bust’’ life history that characterizesother Everglades species (e.g. Beissinger 1995).Populations may suffer ‘‘bust’’ years when con-ditions are poor, productivity is negligible, andnumbers decline, but populations also may beable to increase rapidly when favorable condi-tions allow for pairs to raise more than onebrood.
The potential for considerable variability invital rates complicates evaluation of extinctionrisk. The range of conditions the sparrows reg-ularly encounter no doubt includes conditionsconducive to both population growth and pop-ulation decline. However, it is impossible to de-termine the relative frequency of periods ofgrowth and periods of decline without betterestimates of vital rates, and especially withouta better understanding of how these rates varyin response to environmental conditions.
Modeling extinction risk.—The likelihood thatpopulation fluctuations driven by flooding andfire events will result in extinction of sparrowpopulations in the absence of immigration hasbeen explored through population modeling.These attempts are preliminary and suffer frominsufficient data to estimate demographic pa-rameters and associated variation, as discussedabove. Nott (1998) developed a spatially explic-it, individual-based model (SIMSPAR) inwhich population dynamics are linked to hab-itat condition. SIMSPAR is data-demanding,but data available for modeling habitat occu-pancy and nesting density as a function of hab-itat condition appear to be sufficient to drawpreliminary conclusions. Habitat classificationis based on remote sensing data, and Nott(1998) described clear relationships betweenhabitat and numbers of breeding sparrows.Nott (1998) modeled availability of habitat clas-sified as suitable as a function of flooding andfire history and validated this component of themodel by demonstrating that birds have notbeen detected at survey locations classified asunsuitable. The availability of data on habitatand bird distribution makes this component ofSIMSPAR effective. The model appears capableof predicting habitat availability with reason-able accuracy.
In contrast to the habitat portion, parameter-ization of the demographic portion of the mod-el exceeds current knowledge of sparrow de-mography. Moreover, until more is known

1110 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
about the effects of habitat condition andbreeding status on movement and survival, itis not clear that modeling of sparrow demog-raphy needs to be either individual-based orspatially explicit. In the meantime, it might beuseful to simplify SIMSPAR’s demographicsubroutine. Finally, a complete sensitivity anal-ysis has not been performed, making it unclearhow much the model’s results are affected byvariation in some of the poorly known param-eters.
Nott’s (1998) model is a considerable achieve-ment in integrating habitat and population dy-namics and will be of general interest to mod-elers. However, its current application in theEverglades ecosystem should be limited toevaluating the comparative effects of differentmanagement scenarios and environmentalfluctuations on availability of suitable sparrowhabitat. It is premature to use this model togenerate specific probabilities of extinction(Beissinger and Westphal 1998). SIMSPAR alsomay be a useful tool for helping to set researchpriorities. A thorough sensitivity analysis canbe used to identify parameters that contributethe most to population behavior and therebyidentify where limited research funds shouldbe expended to improve the model’s predictivecapability.
The Panel concludes that, for now, the type ofsimple deterministic matrix model developedby S. Pimm (unpubl. data) is as appropriate forexamining population behavior as is more com-plex modeling, at least until more demographicdata are available. Pimm’s model has not beenwell documented, and several of the parametervalues employed require more rigorous exam-ination. To date, the model has been used pri-marily to explore the capacity for flooding andfire to negatively affect population behaviorthrough depression of survival and reproduc-tion (USDI 1998). The results suggest thatflooding poses a significant threat to Popula-tion A, and fire to the northeastern popula-tions, but not to Population B. Pimm’s modelhas not been used to explore how populationbehavior depends on values of demographicparameters, as we did in Table 3. This type ofmodel will be most useful in evaluating poten-tial effects of different management scenariosand alternative demographies, rather than at-tempting to estimate extinction risk. As addi-tional data required for more complex models
become available, it may be possible to employfirst a stochastic matrix model and eventuallySIMSPAR for these purposes.
Perhaps the strongest conclusion that onecan draw from modeling efforts to date is thatthey portray how flooding and fire, if they arefrequent enough and have strong enough ef-fects on reproduction and survival, might causemajor population declines. Furthermore, themagnitude of effects and frequencies requiredare realistic, at least for some portion of the do-main of possible mortality and reproductiverates. With this in mind, we now return to theissue of extinction risk under different watermanagement strategies.
Conclusions about extinction risk.—The capac-ity for recolonization is one factor determiningthe significance of the extinction of a particularpopulation. Recolonization of Population A ismost problematic because of its isolation fromthe other populations by distance and barriers,especially Shark River Slough. Populations B toF are in much closer proximity but are still sep-arated by barriers, albeit less-significant ones.Our first conclusion, then, is that extinction ofPopulation A may pose greater problems forfuture management than will extinction of theother small populations (C, D, and F).
Owing to the uncertainties about populationdynamics, it is difficult to evaluate the viabilityof particular sparrow populations individually,or the subspecies collectively. However, thecombination of empirical work and modelingresults makes a convincing case that continuedreleases of water into habitat occupied by Pop-ulation A pose a significant risk to that popu-lation’s continued existence. We conclude thatunder the current water management strategy,near-term extinction of Populations A and Dare real possibilities. We further believe that re-taining water in WCA 3A rather than releasingit west of Shark River Slough and into TaylorSlough in wet years will substantially reducethe risk of extinction of Populations A and D.
The case for risks posed by fire is less com-pelling. Population B remained large after themajor fire of 1989, and Pimm’s (unpubl. data)modeling supports the notion that populationfluctuations caused by fire likely are insuffi-cient to cause extinction of Population B. Thetemporary reduction in productivity and hab-itat suitability caused by fire has greater con-sequences for the smaller northeastern popu-

October 2000] 1111Cape Sable Seaside Sparrow Report
lations (C, E, and F) than for Population B. Inthese cases, extinction risk is a function of thenature and duration of interim water manage-ment. Population E may be large enough towithstand adverse effects of fire over the shortterm, even if the ‘‘short term’’ turns out to lastsomewhat longer than anticipated. We con-clude that extinction of Populations C and F isconceivable under both of the short-term sce-narios we evaluated, if interim water manage-ment persists substantially longer than antici-pated, but that recolonization of these popula-tions, should they go extinct, also appears to bea realistic possibility.
Thus, we perceive Populations A and D, andpossibly C and F, to be at risk under both of theshort-term scenarios. The risk of extinction ofthe total population obviously is increased bythe reduction of the number of large popula-tions from three to two. Establishment of a newlarge population to replace an existing oneseems to us a remote possibility, given the cur-rent lack of suitable unoccupied habitat. Gam-bling on being able to first create a large areaof new habitat and then successfully establishbirds there, when no candidate areas have beenidentified, is too risky a strategy to be seriouslyentertained. The best available means to reducethe risk of extinction of the Cape Sable SeasideSparrow is to retain and recover Population A.Population E should be monitored carefullywhile interim water management remains inplace, because the persistence of this popula-tion also is important to the future of the spar-row.
We further conclude that the new water man-agement plan will reduce extinction risk to thesparrow. The reductions in risk achieved willdepend on the extent to which alteration offlows west of Shark River Slough, in TaylorSlough, and in northeastern Shark River Sloughproduced by new structures and policies resultin hydroperiods that mimic historic ones. If thedistribution of water levels is as anticipated, therisks imposed by flooding and fire after imple-mentation should be lower than at any time inrecent history. A possible additional risk weperceive under the new water managementplan is loss of habitat to encroachment ofwoody vegetation. This means that the newmanagement regime must involve periodic fire.
MANAGEMENT RECOMMENDATIONS
Our main conclusions are that (1) extendedhydroperiods result in changes in vegetationand suppressed reproduction and thus repre-sent a serious threat to Cape Sable SeasideSparrows; (2) changes in water managementare required over the short term to preventsuch extended flooding; and (3) the new man-agement plan will alleviate this problem. Aprevious panel of scientists reached these sameconclusions (Orians et al. 1996). The manage-ment recommendations below follow fromthese conclusions.
Long-term management.—The amount of prai-rie habitat under protection within EvergladesNational Park appears to be sufficient to sup-port a self-sustaining population of Cape SableSeaside Sparrows. Indeed, the total populationsize attained at times within this habitat, esti-mated at 6,0001, may be larger than that ofmany other populations of the species in otherregions. The evidence is clear that some of theremaining habitat has been degraded in recentyears by frequent fires and extended hydroper-iods, but it is also clear that the habitat can re-cover once these effects are removed. The newwater management scheme promises to reducethese effects and to restore degraded habitat.From the perspective of Cape Sable SeasideSparrow management, we strongly endorsethis new scheme and urge that it be imple-mented with all possible speed. To maximizebenefits to the sparrow, this plan should be im-plemented in such a way that hydroperiods inthe prairies occupied by sparrows match his-toric ones as closely as possible. Once the newplan is implemented, monitoring and fire man-agement will be the predominant managementactivities. Habitat loss to encroachment bywoody vegetation must be prevented, perhapsthrough a prescribed burning program basedon improved information about optimum burn-ing intervals.
The natural population dynamics of the spar-row may be sufficient to fill the available suit-able habitat provided under the new watermanagement scheme. This process promisesnot only to preserve the remaining large pop-ulations (B and E), but also to recover the smallpopulations (A, C, D, and F) to higher levels.Recovery of these currently small populationsto historical levels is desirable for long-term

1112 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
sustainability of the Cape Sable Seaside Spar-row. If population growth does not occur as an-ticipated once the habitat has recovered, trans-location of individuals may be necessary.Translocation is appropriate only where unoc-cupied habitat has been restored to optimalconditions for sparrows. Currently, such habi-tat may exist within Population A and perhapsat Ochopee (Kushlan et al. 1982, Nott 1998). Ad-ditional unoccupied suitable habitat may ariseas a consequence of improvements in habitatconditions when the new management plan isimplemented.
Short-term management.—Retaining Popula-tion A until the new water management plan isimplemented at some future date is a majorconcern. Our recommendations about interimmanagement are based solely on the require-ments of Cape Sable Seaside Sparrows and ig-nore politics and the needs of other species in-habiting the Everglades. This approach befitsthe charge of the Panel. The best alternative isto reduce flows west of Shark River Slough andinto Taylor Slough, and increase flows intonortheastern Shark River Slough, to the extentpossible using existing structures. This alter-native would benefit Populations C, D, E, andF, as well as A. If this approach is not adopted,then releases of water west of Shark RiverSlough through the S-12 structures should beclosely regulated. In relatively dry years, re-leases could occur as in the past, but our rec-ommendation for wet years is that some waternormally released into Everglades NationalPark under existing policy instead be retainedin WCA 3A. Specifically, we recommend thatwater be managed to enable high productivityuntil Population A has recovered to at least1,000 breeding birds. A dry period of 50 to 60days beginning 15 March is the minimum re-quired to ensure reasonable productivity, anda period of 80 days is preferable. A dry periodof 50 to 60 days should allow most females tocomplete one brood, and a few to complete two,whereas an 80-day dry period should allowmost females to complete two broods (Nott etal. 1998).
In wet years, maintaining dry conditions inPopulation A will mean retaining water inWCA 3A rather than releasing it west of SharkRiver Slough. After Population A has recov-ered, S-12 releases could be allowed in wetyears. These should not occur in consecutive
years, or more often than about two years infive. It might even be possible to release waterone year in five prior to recovery of PopulationA without jeopardizing recovery. Obviously,such releases do not benefit the sparrow andwould have to be justified in terms of other con-siderations that outweigh harmful effects onPopulation A. Similarly, each year managerswill have to weigh potential increases in pro-ductivity of sparrows resulting from extendingthe dry period from 60 to 80 days (Table 3)against the consequences of retaining water inWCA 3A. The latter will be more severe in wet-ter years. Evaluating the biological consequenc-es of retaining water in WCA 3A is beyond thecharge of the Panel (see Armentano 1996 andreferences therein). We note that whatever ad-verse effects occur should be short-lived, be-cause the need to store water in WCA 3A willcease with adoption of the new long-term watermanagement scheme.
Releases of water into Taylor Slough shouldbe regulated similarly to releases west of SharkRiver Slough to avoid extirpation of PopulationD. We conclude that extinction of Population Ddoes not put the Cape Sable Seaside Sparrow inimminent danger of extinction, but managersmay prefer the actions necessary to retain thispopulation over those that will be required torestore it, should natural recolonization not oc-cur. If retaining Population D is treated as a pri-ority, then a recovery criterion based on habitatcondition should be developed for that popu-lation, and releases of water into Taylor Sloughshould be prevented until the criterion isachieved.
That fire might be too infrequent to preventinvasion of woody vegetation is unlikely in theshort term. Therefore, we recommend a policyof prevention and suppression of dry-seasonfires until the new management plan is in ef-fect. We do recommend, however, that pre-scribed burning during the wet season be acomponent of this policy. If managers elect notto increase flows into northeastern Shark RiverSlough prior to implementation of the newmanagement plan, then Populations C and Fwill remain at risk owing to adverse effects offire on habitat quality. Under these conditions,we do not recommend any effort to save themother than fire suppression, nor do we recom-mend translocation of individuals from otherpopulations to them. As long as abnormally

October 2000] 1113Cape Sable Seaside Sparrow Report
frequent dry conditions continue to prevail,translocation efforts will, in our opinion, be fu-tile. Efforts to restore Populations C and F,should they be extirpated, should be delayeduntil historic flow levels are restored to north-eastern Shark River Slough. However, more ag-gressive management of Population E shouldbe designed and implemented, should moni-toring indicate substantial declines in that pop-ulation.
Captive breeding.—Note that we do not in-clude captive breeding among our manage-ment recommendations. Captive breeding rep-resents a rescue operation (Snyder et al. 1996),and the Cape Sable Seaside Sparrow is not yetin need of rescue. The Panel views captivebreeding as risky, unnecessary, premature anddistracting at this time. The hope is that themanagement recommended will be sufficientto ensure that captive breeding will never berequired.
Recovery team.—Finally, we strongly recom-mend that a Federal recovery team be appoint-ed for the Cape Sable Seaside Sparrow to ad-vise local managers. The Everglades is not astatic system, and new challenges can be antic-ipated. A recovery team would serve as a valu-able advisory group as new issues arise.Among its members should be an avian popu-lation or conservation biologist, ornithologistswho have studied the Cape Sable Seaside Spar-row, a botanist specializing in wetlands, and ahydrologist. Such a group is needed to contin-ue the task of evaluating relevant scientific in-formation that we have attempted here, and toassist managers in maintaining compliancewith the species’ recovery plan (USFWS 1998).
ACKNOWLEDGMENTS
We thank the Science Coordination Team of theSouth Florida Ecosystem Restoration Task Force/Working Group for inviting us to conduct this reviewand for organizing and coordinating the review pro-cess. We especially thank Ronnie Best, who led theseefforts and acted as an effective liaison between thePanel and researchers. AOU President Frank Gillhelped arrange for the review and participated in allof its aspects. Researchers studying the Cape SableSeaside Sparrow went to considerable effort to pro-vide information in the form of unpublished data,position papers, presentations at the workshop, dis-cussions in the field, and comments on the draft re-port. We especially thank Sonny Bass, John Curnutt,Tylan Dean, Julie Lockwood, Philip Nott, Stuart
Pimm, and Wil Post for their efforts in this regard.Additional comments were provided by Keith Bild-stein, Donald DeAngelis, Dale Gawlik, Jon Greenlaw,Mark Kraus and Jerome Lorenz (National AudubonSociety Everglades Conservation Office and Taver-nier Science Center), Catherine Langtimm, SydneyMaddock (Biodiversity Legal Foundation), MaryAnn Poole, Bradford Sewell (Natural Resources De-fense Council), and Paul Wetzel.
LITERATURE CITED
AMERICAN ORNITHOLOGISTS’ UNION. 1973. Thirty-second supplement to the AOU Check-list ofNorth American Birds. Auk 90:411–419.
ARMENTANO, T. V. (Ed.). 1996. Ecological assessmentof the 1994–1995 high water conditions in thesouthern Everglades. United States Army Corpsof Engineers and Everglades National Park, Mi-ami, Florida.
ARMENTANO, T. V., R. F. DOREN, W. J. PLATT, AND T.MULLINS. 1995. Effects of Hurricane Andrew oncoastal and interior forests of southern Florida:Overview and synthesis. Journal of Coastal Re-search Special Issue No. 21:111–144.
AVISE, J. C., AND W. S. NELSON. 1989. Molecular ge-netic relationships of the extinct Dusky SeasideSparrow. Science 243:646–648.
BEISSINGER, S. R. 1986. Demography, environmentaluncertainty, and the evolution of mate desertionin the Snail Kite. Ecology 67:1445–1459.
BEISSINGER, S. R. 1995. Modeling extinction in peri-odic environments: Everglades water levels andSnail Kite population viability. Ecological Ap-plications 5:618–631.
BEISSINGER, S. R., AND J. P. GIBBS. 1993. Are variableenvironments stochastic? A review of methodsto quantify environmental predictability. Pages132–146 in Adaptation in stochastic environ-ments (J. Yoshimura and C. W. Clark, Eds.). Lec-ture Notes on Biomathematics, Springer-Verlag,Berlin.
BEISSINGER, S. R., AND M. I. WESTPHAL. 1998. On theuse of demographic models of population via-bility in endangered species management. Jour-nal of Wildlife Management 62:821–841.
BLAKE, N. M. 1980. Land into water–Water into land:A history of water management in Florida. Uni-versity Presses of Florida, Tallahassee.
BUCKLAND, S. T., D. R. ANDERSON, K. P. BURNHAM,AND J. L. LAAKE. 1993. Distance sampling. Chap-man and Hall, London.
COCHRAN. 1977. Sampling techniques, 3rd ed. JohnWiley and Sons, New York.
COOK, R. D., AND J. O. JACOBSON. 1979. A design forestimating visibility bias in aerial surveys. Bio-metrics 35:735–742.
CRAIGHEAD, F. C. 1971. Trees of South Florida, vol. 1.

1114 [Auk, Vol. 117Cape Sable Seaside Sparrow Report
University of Miami Press, Coral Gables, Flori-da.
CURNUTT, J. L., A. L. MAYER, T. M. BROOKS, L. MAN-NE, O. L. BASS, Jr., D. M. FLEMING, M. P. NOTT,AND S. L. PIMM. 1998. Population dynamics ofthe endangered Cape Sable Seaside Sparrow.Animal Conservation 1:11–21.
DEAN, T. F., AND J. L. MORRISON. 1998. Non-breedingseason ecology of the Cape Sable Seaside Spar-row (Ammodramus maritimus mirabilis): 1997–1998 field season final report. Unpublished re-port submitted to the United States Fish andWildlife Service.
DEANGELIS, D. L., L. J. GROSS, M. A. HUSTON, W.F.WOLFF, D. M. FLEMING, E. J. COMISKEY, AND S.M. SYLVESTER. 1998. Landscape modeling forEverglades ecosystem restoration. Ecosystems 1:64–75.
DUEVER, M. J., J. F. MEEDER, L. C. MEEDER, AND J. M.MCCOLLOM. 1994. The climate of South Floridaand its role in shaping the Everglades ecosys-tem. Pages 225–247 in Everglades: The ecosys-tem and its restoration (S. M. Davis and J. C. Og-den, Eds.). St. Lucie Press, Delray Beach, Florida.
GREENBERG, R., P. J. CORDERO, S. DROEGE, AND R. C.FLEISCHER. 1998. Morphological adaptation withno mitochondrial DNA differentiation in theCoastal Plain Swamp Sparrow. Auk 115:706–712.
GRISCOM, L. 1944. A second revision of the SeasideSparrows. Occasional Papers of the Museum ofZoology at Louisiana State University 19:313–328.
GUERRA, R. E. 1996. Impacts of the high water periodon 1994–95 on tree islands in Water Conserva-tion Areas. Pages 47–58 in Ecological assessmentof the 1994–1995 high water conditions in thesouthern Everglades (T. V. Armentano, Ed.).United States Army Corps of Engineers and Ev-erglades National Park, Miami, Florida.
KNOPF, F. L. 1995. Declining grassland birds. Pages296–298 in Our living resources: A report to thenation on the distribution, abundance, andhealth of U.S. plants, animals, and ecosystems(E. T. LaRoe, G. S. Farris, C.E. Puckett, P. D. Dor-an, and M. J. Mac, Eds.). United States NationalBiological Service, Washington, D.C.
KUSHLAN, J. A., AND O. L. BASS, Jr. 1983. Habitat useand the distribution of the Cape Sable Sparrow.Pages 139–146 in The Seaside Sparrow: Its biol-ogy and management (T. L. Quay, J. B. Funder-burg, Jr., D. S. Lee, E. F. Potter, and C. S. Robbins,Eds.). North Carolina Biological Survey Occa-sional Paper 1983–5.
KUSHLAN, J. A., O. L. BASS, Jr., L. L. LOOPE, W. B.,ROBERTSON, Jr., P. C. ROSENDAHL, AND D. L. TAY-LOR. 1982. Cape Sable Sparrow managementplan. South Florida Research Center Report M-660.
LOCKWOOD, J. L., K. H. FENN, J. L. CURNUTT, D. RO-SENTHALL, K. L. BALENT, AND A. L. MAYER. 1997.Life history of the endangered Cape Sable Sea-side Sparrow. Wilson Bulletin 109:720–731.
MCDONALD, M. V. 1988. Status survey of two FloridaSeaside Sparrows and taxonomic review of theSeaside Sparrow assemblage. Florida Coopera-tive Fish and Wildlife Research Unit TechnicalReport 32.
MCDONALD, M. V., AND R. GREENBERG. 1991. Nestdeparture calls in New World songbirds. Condor91:17–24.
NICHOLS, J. D., J. E. HINES, J. R. SAUER, F. W. FALLON,J. E. FALLON, AND P. J. HEGLUND. 2000. A double-observer approach for estimating detectionprobability and abundance from point counts.Auk 117:393–408.
NOTT, M. P. 1998. Effects of abiotic factors on popu-lation dynamics of the Cape Sable Seaside Spar-row and continental patterns of herpetologicalspecies richness: An appropriately scaled land-scape approach. Ph.D. dissertation, Universityof Tennessee, Knoxville.
NOTT, M. P., O. L. BASS, Jr., D. M. FLEMING, S. E. KIL-LEFFER, N. FRALEY, L. MANNE, J. L. CURNUTT, T.M. BROOKS, R. POWELL, AND S. L. PIMM. 1998.Water levels, rapid vegetational changes, and theendangered Cape Sable Seaside Sparrow. Ani-mal Conservation 1:23–32.
OGDEN, J. C. 1996. Wood Stork. In Rare and endan-gered biota of Florida (J. A. Rodgers, H. Kale IIIand H. T. Smith, Eds.). University Press of Flor-ida, Gainesville.
ORIANS, G. H., W. DUNSON, J. FITZPATRICK, D. GE-NEREUX, L. HARRIS, M. KRAUS, AND R. E. TURN-ER. 1996. Report of the panel to evaluate the eco-logical assessment of the 1994–1995 highwaterlevels in the southern Everglades. In Ecologicalassessment of the 1994–1995 high water condi-tions in the southern Everglades (T. V. Armen-tano, Ed.). United States Army Corps of Engi-neers and Everglades National Park, Miami,Florida.
POST, W. 1974. Functional analysis of space-relatedbehavior in the Seaside Sparrow. Ecology 55:564–575.
POST, W., AND J. S. GREENLAW. 1994. Seaside Sparrow(Ammodramus maritimus). In The birds of NorthAmerica, no. 127 (A. Poole and F. Gill, Eds.).Academy of Natural Sciences, Philadelphia, andAmerican Ornithologists’ Union, Washington,D.C.
POST, W., J. S. GREENLAW, T. L. MERRIAM, AND L. A.WOOD. 1983. Comparative ecology of northernand southern populations of the Seaside Spar-row. Pages 123–136 in The Seaside Sparrow: Itsbiology and management (T. L. Quay, J. B. Fun-derburg, Jr., D. S. Lee, E. F. Potter, and C. S. Rob-

October 2000] 1115Cape Sable Seaside Sparrow Report
bins, Eds.). North Carolina Biological SurveyOccasional Paper 1983–5.
ROBINS, J. D., AND G. D. SCHNELL. 1971. Skeletal anal-ysis of the Ammodramus-Ammospiza grasslandsparrow complex: A numerical taxonomic study.Auk 88:567–590.
SNYDER, N. F. R., S. R. DERRICKSON, S. R. BEISSINGER,J. W. WILEY, T. B. SMITH, W. D. TOONE, AND B.MILLER. 1996. Limitations of captive breeding inendangered species recovery. Conservation Bi-ology 10:338–348.
STACEY, P. B., AND M. TAPER. 1992. Environmentalvariation and the persistence of small popula-tions. Ecological Applications 2:18–29.
STIMSON, L. A. 1956. The Cape Sable Sparrow: Its for-mer and present distribution. Auk 73:489–502.
TAKEKAWA, J. E., AND S. R. BEISSINGER. 1989. Dis-persal, cyclic drought, and the conservation ofthe Snail Kite in Florida: Lessons in critical hab-itat. Conservation Biology 3:302–311.
TAYLOR, D. L. 1983. Management of the Cape SableSparrow. Pages 147–152 in The Seaside Sparrow:Its biology and management (T. L. Quay, J. B.Funderburg, Jr., D. S. Lee, E. F. Potter, and C. S.Robbins, Eds.). North Carolina Biological Sur-vey Occasional Paper 1983–5.
THOMAS, T. M. 1974. A detailed analysis of climato-logical and hydrological records of south Floridawith reference to man’s influence upon ecosys-tem evolution. Pages 82–122 in Environments ofsouth Florida, present and past (P. J. Gleason,Ed.). Memoir 2, Miami Geological Society, Mi-ami, Florida.
THOMPSON, S. K. 1992. Sampling. John Wiley andSons, New York.
UNITED STATES DEPARTMENT OF THE INTERIOR. 1998.Balancing on the brink. U.S. Department of theInterior, Vero Beach, Florida.
UNITED STATES FISH AND WILDLIFE SERVICE. 1998.Multi-species recovery plan for the threatenedand endangered species of South Florida. U.S.Fish and Wildlife Service, Atlanta.
VERNER, J. 1985. Assessment of counting techniques.Current Ornithology 2:247–302.
WALTERS, M. J. 1992. A shadow and a song: Thestruggle to save an endangered species. ChelseaGreen Publishing Company, Post Hills, Ver-mont.
WERNER, H. W. 1975. The biology of the Cape SableSeaside Sparrow. Project completion report pre-pared for the National Park Service, EvergladesNational Park, Florida.
WERNER, H. W., AND G. E. WOOLFENDEN. 1983. TheCape Sable Sparrow: Its habitats, habits and his-tory. Pages 55–75 in The Seaside Sparrow: Its bi-ology and management (T. L. Quay, J. B. Fun-derburg, Jr., D. S. Lee, E. F. Potter, and C. S. Rob-bins, Eds.). North Carolina Biological SurveyOccasional Paper 1983–5.
ZINK, R. M., AND D. L. DITTMANN. 1993. Gene flow,refugia, and the evolution of geographic varia-tion in the Song Sparrow (Melospiza melodia).Evolution 47:717–729.
ZINK, R. M., AND H. W. KALE. 1995. Conservation ge-netics of the Dusky Seaside Sparrow (Ammodra-mus maritimus nigrescens). Biological Conserva-tion 74:69–71.
Announcement
New Editor Selected.—Kimberly G. Smith of theUniversity of Arkansas in Fayetteville has been se-lected as the new Editor of The Auk. Please submit
manuscripts to The Auk, Department of Biological Sci-ences, University of Arkansas, Fayetteville, Arkansas72701, USA.





























