Embed Size (px)
Citation preview

الجوهىريت الجزائريت الديوقراطيت الشعبيت Ministry of High Education and Scientific Research
N° d’ordre : 91/DS/2018
N° de série : 04/BA/2018
Thesis submitted for the degree of Doctorate in Sciences
Option: Animal physiology
Topic:
Presented by: AKLIL Badiaa
Examination board:
President: N. BAAZIZ Prof. University Frères Mentouri Constantine
Supervisor: S. ZERIZER Prof. University Frères Mentouri Constantine
Examiners: K. BOUBEKRI MC. University Frères Mentouri Constantine
S. DAHAMNA Prof. University of Sétif
S. KHENNOUF Prof. University of Sétif
C. ABDENNOUR Prof. University of Annaba
2017 / 2018
The effect of Argania spinosa on plasma
Homocysteine, Lipids, Antioxidant enzymes and
Aortas Sections in Methionine induced
Hyperhomocysteinemia in mice
University of des frères Mentouri Constantine
Faculty of life and Natural Sciences
Department of Animal Biology
جاهعت اإلخىة هنتىري قسنطينت
لىم الطبيعت و الحياة ـــليت عـــك
ىاى ــيــحــىلىجيا الـيـسن بــــق

Dedication
This thesis is dedicated to the memory of my father Omar, who would have been
happy to see me at this step.
To my mother Fatma, for her constant, unconditional love and support.
To my husband Ali, who has been a constant source of support and
encouragement during all the hard periods of research and life.
To my children: Haithem, Nouha and Assil.
To all my family, the symbol of love and giving.
And to all my friends who encourage and support me.

Acknowledgements
First, I am deeply grateful to the Almighty Allah who helped me to
complete this thesis.
I wish to express my deepest gratitude to my supervisor Prof. S. Zerizer
for introducing me to the interesting field of science and for providing me with
the opportunity to carry out this study, and for her invaluable advice, patience
and inspiring guidance throughout this work.
I am grateful to Prof. Z. Kabouche for providing me the plant material.
I would like to thank all the jury members, Prof. N. BAAZIZ
(president of the jury), Dr. K. BOUBEKRI (examiner of the doctorate thesis),
Prof. S. DAHAMNA (examiner of the doctorate thesis), Prof. S. KHENNOUF
(examiner of the doctorate thesis), and Prof. C. ABDENNOUR (examiner of the
doctorate thesis), for their interest in my work.
I wish to express my sincere thanks to Dr. Guy D’hallewin for accepting
me in his laboratory and giving me the opportunity to work with his exceptional
laboratory team; my sincere thanks also go to Pr. G. Orru to help me conduct
laboratory tests related to research.
I wish to thank Dr. G. Piacherri for her help and assistance.
I would like to thank everyone who provided any assistance even with a
single word and especially my friends.

Table of contents
LIST OF ABBREVIATIONS
LIST OF FIGURES
LIST OF TABLES
INTRODUCTION…………………………………………………………………………… 01
LITERATURE REVIEW
1- Homocysteine ………………………………………………………………...................
03
1-1 Structure and Forms of homocysteine………………………………………………. 03
1-2 Biosynthesis and Metabolism of homocysteine………………………………........... 03
1-2 Regulation of Metabolism…………………………………………………………….
05
2- Hyperhomocysteinemia …………………………………………………………………. 06
2-1 Definition…………………………………………………………………………….. 06
2-2 Causes of Hyperhomocystenemia …………………………………………………… 06
2-2-1 Genetic Defects …………………………………………………………………. 06
a- Cysthathionine beta synthase deficiency……………………………………….. 07
b- N-5, 10-Methylene tetrahydrofolate reductase deficiency……………………… 07
2-2-2 Vitamin Deficiencies …………………………………………………………….. 07
2-2-3 Other Causes of Hyperhomocysteinemia ………………………………………
08
3- Pathogenicity of Hyperhomocysteinemia ………………………………………………
08
3-1 Hyperhomocysteinemia and cardiovascular disease……………………………….. 08
3-1-1 Endothelial dysfunction………………………………………………………… 08
3-1-1-1Homocysteine-Induced Oxidative Stress Condition………………………… 10
a- Uncoupling of NO synthase ………………………………………………… 11
b- Accumulation of Asymmetric Dimethylarginine…………………………… 12
3-1-1-2Homocysteine induced protein modification and endoplasmic reticulum
stress …………………………………………………………………… …..
13
3-1-1-3 Homocysteine induced inflammatory/prothrombotic conditions…………. 13
3-1-2 Atherosclerosis……………………………………………………………………. 15
3-2 Hyperhomocysteinemia and dyslipedimia……………………………………………… 17
3-3 Hyperhomocysteinemia and hepatic disease…………………………………………… 18
3-4 Hyperhomocysteinemia and carcinogenesis …………………………………………… 18

Table of contents
4- Therapy of hyperhomocysteinemia ……………………………………………………
19
5- Argania spinosa………………………………………………………………………………….
19
5-1 Description ……………………………………………………………………………
19
5-2 Botanical classification……………………………………………………………….. 20
5-1 Therapeutic properties………………………………………………………………….
21
6- Bacterial Biofilm and cardiovascular diseases…………………………………................. 22
6-1 Definition of Biofilm………………………………………………………………… 22
6-2 Stages of Biofilm formation………………………………………………………….
22
6-3 Biofilm and cardiovascular infections ……………………………………………….. 23
6-4 Bacterial strains ……………………………………………………………………. 25
6-4-1 Streptococcus mutans…………………………………………………………………… 25
6-4-2 Streptococcus intermedius and Streptococcus anginosus …………………………. 25
6-4-1 Staphylococcus haemolyticus ………………………………………………………….. 26
6-4-3 Streptococcus uberis……………………………………………………………………..
26
MATERIALS AND METHODS
1- Biological plant…………………………………………………………………………….
27
2- Animals………………………………………………………………………………….. 27
2-1 Experimental animals…………………………………………………………….. …… 27
2-2 Experimental treatments………………………………………………………………… 28
2-3- Blood and tissue sampling………………………………………………………………
28
3- Methods…………………………………………………………………………….......... 30
3-1Chemical products……………………………………………………………………… 30
3-2 Equipments…………………………………………………………………………….. 30
3-3 Biochemical analysis………………………………………………………………….. 30
3-3-1 Plasma Hcy determination…………………………………………………………..
30
3-3-2 Lipids determination…………………………………………………………………
31

Table of contents
3-3-3 Determination of Aspartate Aminotransferase and Alanine aminotransferase
activities………………………………………………………………………………
33
3-4 Determination of oxidative stress parameters …………………………………................. 34
3-4-1 Tissue homogenate preparation………………………………………………………. 34
3-4-2 Glutathione assay……………………………………………………………………… 34
3-4-2 Catalase activity……………………………………………………………….............. 36
3-5 Histological analysis……………………………………………………………..............
37
4- Biofilm formation and quantification ……………………………………………………. 38
4-1 Strains and culture conditions……………………………………………………….. 38
4-2 Crystal Violet Biofilm formation screening assay……………………………………..
38
5- Statistical analysis…………………………………………………………………………..
40
RESULTS AND DISCUSSION
Chapter 1: Effect of treatment on body weight, Hcy levels, lipid profile, liver enzyme
activities and antioxidants markers in mice
1- Body weight …………………………………………………………………………..
41
2- Effect of treatment on lipid profile in mice……………………………………................ 41
2-1 Triglycerides……………………………………………………………………………. 41
2-2 Total cholesterol ……………………………………………………………………….. 42
2-3 HDL-c……………………..…………………………………………………………… 43
2-4 LDL-c…………… .……………………………………………………………………..
3- Effect of treatments on Homocysteine levels ……………………………………………..
44
45
4- Effect of treatment on liver enzyme activity……………………………………………… 46
4-1 Aspartate Aminotransferase………………………………………………………….. 46
4-2 Alanine aminotransferase ……………………………………………………………..
47
5- Effect of treatment on antioxidants markers ……………………………………………… 48
5-1 Reduced Gluthatione …………………………………………………………………. 48
5-2 Catalase activity……………………………………………………………………….
49

Table of contents
Chapter 2: Effect of treatment on histology of aorta heart and liver
1- Histological study of the heart…………………………………………………..............
51
2- Histological study of the aorta…………………………………………………………..
51
3- Histological study of the liver…………………………………………………...............
51
Chapter 3: Anti-biofilm formation of Argan oil “In vitro” Study
DISCUSSION……………………………………………………………………………………..
68
CONCLUSION AND PERSPECTIVES……………………………………………………….
85
REFERENCES…………………………………………………………………………………..
87
APPENDICES
PAPER
ملخص بالعربيةال

List of Abbreviations
.HRO2-: Hydroperoxyl
•NO: Nitric Oxide
•NO2: Nitrogen Dioxide
•O2-: Superoxide
•OH: Hydroxyl
•RO2: Peroxyl
5, 10-MTHF: 5, 10-Methylene Tetrahydrofolate
5, 10MTHFR: 5, 10- Methylene Tetrahydrofolate Reductase
5-MTHF: 5-Methylene Tetrahydrofolate
ADMA: Protein Asymmetric Dimethyl Arginine
AECA: Anti-Endothelial Cell Antibodies
ALT: Alanine Aminotransferase
Ang II: Angiotensin II
ANOVA: One-way Analysis Of Variance
anti-oxLDL: Anti-Oxidized LDL Antibodies
APLA: Anti-Phospholipid Antibodies
apoB: Apolipoprotein B
apoE : Apolipoprotein E
AST: Aspartate Aminotransferase
BH4: Tetrahydrobiopterin
BSA: Bovine Serum Albumin
CβS: Cystathionine β-Synthase
CoNS : Coagulase-negative staphylococci
Cth−/−: Cystathionine Deficient Mice
CVD: Cardiovascular Diseases
CYP7A1: Cholesterol 7A-Hydroxylase
DHF: Dihydrofolate
DNA: Deoxyribonucleic Acid
DTNB: Dithiobis-2-Ditrobenzoic Dcid
EDHF: Endothelium-Derived Hyperpolarizing Factor
EDTA: Tris Ethylene Di-amine Tetra Acetic acid
eNOS: Endothelial Nitric Oxide Synthase
EPS: Exopolysaccharides

List of Abbreviations
ER: Endoplasmic Reticulum
ET-1: Endothelin-1
GCT: γ-cystathionase
GSH: Hepatic Reduced Glutathione
GST: Glutathione S-Transferase
H2O2: Hydrogen Peroxide
Hb: Hepatocellular Ballooning
Hcy: Homocysteine
HHcy: Hyperhomocysteinemia
HNO2: Nitrous oxide
HOCl: Hydrochlorous Acid
Hsp90: Heat Shock Protein 90
HTL: Homocysteine Thiolactone
ICAM-1: Intercellular Adhesion Molecule-1
IE: Infective Endocarditis
LCAT: Lecithin-Cholesterol Acyltransferase
LDL-c: Low Density Lipoprotein
LPS: Lipopolysaccharides
MAT I/III: Methionine Adenosyl Transferases I and III
MCP-1: Monocyte Chemoattractant Protein 1
MS: Methionine Synthase
MTHFR: Methylene Tetrahydro Folate Reductase
NAD: Nicotinamide Adenine Dinucleotide
NADH: Nicotinamide Adenine Dinucleotide
NF-κB : Nuclear Factor-kappa B
NO: Nitric Oxide
NOS: Nitric oxide Synthase
O2·−: Superoxide
ONOO−: Peroxynitrite
oxLDL: Oxidized Low Density Lipoprotein
PC: Phosphatidylcholine
PE: Phosphatidyl Ethanolamine
PEMT: Phosphatidyl Ethanolamine Methyl Transferase

List of Abbreviations
PGI2: Prostaglandin I 2
PLP: Pyridoxal 5-Phosphate
PON1: Paraoxonase 1
QS: Quorum Sensing
RNS: Reactive Nitrogen Species
RONOO: Alkyl Peroxynitrates
ROS: Reactive Oxygen Species
SAG: Streptococcus Anginosus Group
SAH: S-Adenosyl –L-homocysteine
SAM: S-Adenosyl Methionine
SMCs: Smooth Muscle Cells
SPSS: Statistical Package for Social Science
SREBP-1: sterol regulatory element-binding protein
TBS: Tris-Buffered Saline
TG: Triglycerides
THF: tetrahydrofolate
TNF-α : Tumor Necrosis Factor-α
UPR: Unfolded Protein Response
VCAM-1: Vascular Cell Adhesion Molecule-1
VEC: Vascular Endothelial Cells
VLDL: Very Low Density Lipoproteins
VSMC: Vascular Smooth Muscle Cells
XDH : Xanthine Dehydrogenase
XO : Xanthine Oxidase
XOR : Xanthine Oxido Reductase

List of Figures
Figure 01. Homocysteine metabolism………………………………………………….
04
Figure 02. Potential mechanisms of homocysteine-induced endothelial dysfunction.. 09
Figure 03. Major endogenous sources of reactive oxygen species (ROS) and reactive
nitrogen species (RNS) in the cardiomyocyte…………………………………………..
10
Figure 04. Central role of endothelial NO synthase (eNOS) uncoupling in the
pathogenesis of endothelial dysfunction…………………………………………………
12
Figure 05. Regulatory circuits in inflammation and endothelial dysfunction………….
14
Figure 06. Hyperhomocysteinemia and etiopathogenesis of atherosclerosis …………..
16
Figure 07. Possible interactions between hyperhomocystenemia and hyperlipidemia in
cell pathology……………………………………………………………………………
17
Figure 08. Kernel and tegument of Argania spinosa L………………………………..
20
Figure 09. Distribution area of the Argan tree in Algeria ……………………………
21
Figure 10. Various stages of biofilm formation and development……………………
23
Figure 11. Blood and tissue sampling…………………………………………………. 29
Figure 12. Effect of L-methionine intake on mice weight during 21 days……………
41
Figure 13. Interaction of L-methionine and A. spinosa seeds on the triglycerides in
mice during 21 days of treatment……………………………………………………….
42
Figure 14. Interaction of L-methionine and A. spinosa seeds on the T-CHO in mice
during 21 days of treatment……………………………………………………………..
43
Figure15. Interaction of L-methionine and A. spinosa seeds on the HDL-c in mice
during 21 days of treatment. ……………………………………………………………
44
Figure 16. Interaction of L-methionine and A.spinosa seeds on the LDL-c in mice
during 21 days of treatment………………………………………………………………
45
Figure 17. Interaction of L-methionine and A. spinosa seeds on the homocysteine
levels in mice during 21 days of treatment…………………………………………….
46
Figure 18. Interaction of L-methionine and A. spinosa seeds on the AST in mice
during 21 days of treatment……………………………………………………………...
47
Figure 19. Interaction of L-methionine and A.spinosa seeds on the ALT in mice during
21 days of treatment…………………………………………………………………….
48

List of Figures
Figure 20. Interaction of L-methionine and A.spinosa seeds on the reduced glutathione
in mice during 21 days of treatment……………………………………………………..
49
Figure 21. Interaction of L-methionine and A.spinosa seeds on the catalase activity in
mice during 21 days of treatment……………………………………………………….
50
Figure 22. Histological sections of heart tissue in experimental groups (F, M, MP and
P)………………………………………………………………………………………..
53
Figure 23. Histological sections of the arch aorta in experimental groups (F, M, MP
and P)…………………………………………………………………………………….
55
Figure 24. Histological sections of the abdominal aorta in experimental groups (F, M,
MP and P)……………………………………………………………………….............
57
Figure 25. Histological sections of the iliac aorta in experimental groups (F, M, MP
and P)……………………………………………………………………….....................
59
Figure 26. Histological sections of liver tissue in experimental groups (F, M, MP and
P)………………………………………………………………………………………….
61
Figure 27. Inhibitory effect of Argan oil on S. intermedius biofilm formation………….
64
Figure 28. Inhibitory effect of Argan oil on S. haemolyticus biofilm formation……….
64
Figure 29. Inhibitory effect of Argan oil on S. mutans biofilm formation……………..
65
Figure 30. Inhibitory effect of Argan oil on S. anginosus biofilm formation………….
65
Figure 31. Inhibitory effect of Argan oil on S. uberis biofilm formation………………
66
Figure 32. Effect of the first dilution (100 g/mL) concentration of Argan oil on
elimination of biofilms………………………………………………………………….
67

List of Tables
Table 01: Structures and forms of Hcy and related amino acids …………………….......
03
Table 02: Composition of diet for 1 kg of food taken by the mice during 21 days
(ONAB) ………………………………………………………………………………….
27
Table 03: Treatment of mice…………………………………………………………...
28
Table 04: Concentrations and amounts of reagents needed for the dosage of catalase
activity. …………………………………………………………………………………….
36
Table 05: Reduction percent of biofilm for test bacteria treated with different
concentrations of Argan oil. ………………………………………………………………
66

Introduction

Introduction
1
Homocysteine "hypothesis of arteriosclerosis" was first proposed by McCully in 1969,
when he observed premature atherothrombosis of the peripheral, coronary, and cerebral
vasculature in children with homocystinuria, an in born error in methionine metabolism
(Rasmussen and Moller, 2000).
Homocysteine (Hcy) a type of amino acid that is naturally found in blood plasma is
not harmful at normal levels, but when its levels are too high, health problems can result. If
unhealthy levels of Hcy increase in the blood, the delicate lining of an artery (endothelium)
can be damaged. Also, Hcy can both initiate and potentiate atherosclerosis (Saleh, 2015).
Therefore, is considered as an emerging cardiovascular risk factor (Athyros et al., 2010).
Homocysteine induced injury to the arterial wall is one of the factors that can initiate
the process of atherosclerosis, leading to endothelial dysfunction and eventually to heart
attacks and strokes (Gallai et al., 2001; Papatheodorou and Weiss, 2007).
Oxidative stress induced by Hcy is reflected by a decrease in serum total anti-oxidant
capacity. The oxidative stress resulting from elevated serum Hcy can oxidize membrane lipids
and proteins and stimulate the activation of Nuclear Factor-kappa B (NFκB), and
consequently increase the expression of inflammatory factors in vivo. Hcy can be converted to
a highly reactive thiolactone which is able to react with proteins forming- NH-CO-adducts,
thus affecting body proteins and enzymes (Ramakrishnan et al., 2006).
The studies suggest that certain chronic infections increase the risk for cardiovascular
disease and that such infections may be considered novel and potentially modifiable risk
factors (Epstein et al., 1999). Specific pathogens along with their potential contribution by
direct or indirect mechanisms to atherosclerosis pathogenesis have been recently reviewed
(Rosenfeld and Campbell, 2011).
Biofilm is an aggregate of microorganisms in which cells are adhere to each other to a
surface. The adherent cells are embedded within a self-produced matrix of extracellular
polymeric substance (Gupta, 2015).
Biofilms can cause chronic infections and are associated with a number of chronic
disease states including cystic fibrosis, infectious endocarditis, and chronic wounds (Singh et
al., 2000; James et al., 2008). In this context, the development of bacterial resistance to
presently available antibiotics has necessitated the need to search for new antibacterial agents.

Introduction
2
Plants as a source of medicinal compounds have continued to play a dominant role in
the maintenance of human health since ancient times. According to the World Health
Organization plant extracts or their active constituents are used as folk medicine in traditional
therapies of 80% of the world’s population. Over 50% of all modern clinical drugs are of
natural product origin (Kirbag et al., 2009).
This study was designed to investigate the beneficial effects of Algerian plant Argania
spinosa (using the powdered seeds and oil) belongs to the family Sapotaceae. The therapeutic
benefits of A. spinosa have been claimed by previous studies which have confirmed that A.
spinosa have several biological effects including: antiproliferative (Bennani et al., 2006;
Drissi et al., 2006; Samane et al., 2006; Bennani et al., 2009), Hypolipidemic,
hypocholesterolemic ( Berrougui et al., 2003), antiatherogenic (Berrougui et al., 2004; Cherki
et al., 2005; Cherki et al., 2006), antiradical (Drissi et al., 2004; Amzal et al., 2008) and anti-
inflammatory activities (Alaoui et al., 1998).
The main objectives of this thesis are:
Induce hyperhomocysteinemia by administration of high L-methionine dose, in an in vivo
animal;
Examine the effect of L-methionine on the weight.
Examine the effect of L-methionine on some biochemical parameters such as plasma
Hcy, triglycerides (TG), Total cholesterol (T-CHO), low density lipoprotein (LDL-c),
high density lipoprotein (HDL-c), ALT, AST, reduced glutathione (GSH), and catalase
activity.
Examine the effect of L-methionine on different sections of aorta, heart and liver.
Evaluate the effect of the powdered seeds of A. spinosa seeds on hyperhocysteinemia and
other biochemical paramaters.
Evaluate the effect of the powdered seeds of A. spinosa on the structure disorders of
aorta, heart and liver induced by high L-methionine intake; and
Assess the anti-biofilm activity of Argan oil against 5 bacterial strains, which can induce
cardiovascular problems.

Literature
Review

Literature Review
3
1- Homocysteine
Homocysteine (Hcy) is a natural sulfur-containing amino acid produced in the
metabolism of the essential amino acid methionine, which is derived from dietary protein
(Narmatha et al., 2015). Normally human Hcy levels range from 4 to 12.3 μmol/l (Elhawary
et al., 2013). The levels of Hcy increase with aging, and are typically higher in men than
women (Nygard et al., 1995; Refsum et al., 2006). However, elevated plasma Hcy
concentrations have important implications for human health and disease (Jing et al., 2014).
1-1 Structure and forms of Homocysteine
Homocysteine is present in different forms (Ganguly and Alam, 2015): around 1%
circulates as free thiol, 70–80% remains disulphide-bound to plasma proteins, mainly albumin
and 20–30% combines with itself to form the dimer Hcy or with other thiols (Hankey and
Eikelboom, 1999).
Table 01: Structures and forms of Hcy and related amino acids (Miller, 2013).
1-2 Biosynthesis and metabolism of Homocysteine
The single source of Hcy in humans is dietary methionine (Miller, 2013). Methionine
is converted into S-adenosylmethionine (SAM), which then loses a methyl moiety and
becomes S-adenosyl-homocysteine (SAH), which finally hydrolyzes into Hcy and adenosine
(Tchantchou, 2006).
Homocysteine is metabolized via two pathways (Figure 01). The first one is
remethylation, where Hcy is reconverted into methionine (Tchantchou, 2006). In this
pathway, Hcy reacquires a methyl group in a reaction catalyzed by the zinc-dependent

Literature Review
4
enzyme, Methionine Synthase (MS), with methyl tetrahydrofolate serving as the methyl
donor and vitamin B12 serving as a cofactor. This reaction occurs in all mammalian cells.
Alternatively Hcy can be remethylated in a folate and vitamin B12-independent reaction using
betaine as the methyl donor and catalyzed by Betaine-Homocysteine-Methyltransferase. This
reaction occurs primarily in the liver and to a lesser extent in the kidney and possibly in the
brain (Miller, 2013).
The second pathway is transsulfuration, where Hcy is converted into cystathionine to
form cysteine by cystathionine-ß-synthase (CBS), with vitamin B6 as a co-factor
(Tchantchou, 2006; Plazar and Jurdana, 2010). Cystathionine is then cleaved to form α-
ketobutyrate and cysteine in a second PLP-dependent reaction catalyzed by cystathionase.
Further metabolism of cysteine leads to the formation of Glutathione or inorganic sulfate
(Miller, 2013).
Figure 01. Homocysteine metabolism (Škovierová et al., 2016).

Literature Review
5
1-3 Regulation of Metabolism
Perturbations in methyl group metabolism and Hcy balance have emerged over the
past few decades as having defining roles in a number of pathological conditions. Numerous
nutritional, hormonal, and genetic factors that are characterized by elevations in circulating
homocysteine concentrations are also associated with specific pathological conditions,
including cancer development, autoimmune diseases, vascular dysfunction, and
neurodegenerative disease (Schalinske and Anne Smazal, 2012).
Because Hcy has many metabolic routes for its production and utilization, a number of
key proteins involved in these processes factor heavily in the regulation of Hcy balance. When there is an excess of methionine, Hcy is metabolized via the pathway of trans-
sulfurylation, producing cystathionine and cysteine in turn. Conversely, under conditions of
methionine deficiency, Hcy is remethylated into methionine. Hcy is remethylated in the liver
via betaine-homocysteine-methyltransferase; however, in most tissues, Hcy is remethylated
into methionine by methionine synthase (MS), which uses vitamin B12 as a co-factor and 5-
Methylene Tetrahydrofolate (5-MTHF) as a substrate (Marinou et al., 2005).
An additional level on Hcy metabolism is exerted by oxidative stress, which reduces
methionine synthase activity. This may occur by oxidative inactivation of the vitamin B12
cofactor or by the oxidation of cysteine residues that are important for zinc binding. By
inhibiting methionine synthase, oxidative stress tends to divert Hcy toward cystathionine
synthesis away from methionine synthesis. This serves to increase the synthesis of
glutathione, a product of Hcy metabolism through the transsulfuration pathway and an
important intracellular antioxidant (Miller, 2013).
In addition, plasma Hcy levels are affected by menopause (Hak et al., 2000), diabetes
(Wijekoon and Brosnan, 2007), and thyroid disorders (Saleh, 2015). These observations
suggest that hormones, including estrogen, insulin, thyroxine, and thyroid stimulating
hormone, may directly or indirectly affect Hcy metabolism. The mechanisms by which these
hormones affect Hcy metabolism are poorly understood (Miller, 2013).

Literature Review
6
2- Hyperhomocysteinemia
2-1 Definition
In 1969, McCully reported two patients with homocystinuria who presented with
premature atherosclerosis at the ages of 2 months and 8 years, respectively. Since then,
increasing evidence suggests that even modest elevations in plasma homocysteine called
hyperhomocystenemia( HHcy), may act as an independent risk factor for atherosclerosis in
the general population (Guthikonda and Haynes, 2006). Hyperhomocystenemia is a metabolic
systemic disorder with defects in sulphur-containing amino acid (methionine and cysteine)
metabolism leading to abnormally higher amounts of Hcy (Veeranki and Tyagi, 2013).
Several types of HHcy are classified in relation to the Hcy
concentration: moderate (16–30 μM), intermediate (31–100 μM), and severe (higher than 100
μM) (Liu et al., 2007).
Hcy occurs in human blood plasma in several forms, including the most reactive one,
homocysteine thiolactone (HTL) – a cyclic thioester, which represents less than <1 % of total
plasma Hcy. The increase in extracellular Hcy is toxic to cells and tissues and it has the
potential to initiate a broad array of vascular complications (Domagała et al., 2006;
Jakubowski, 2008).
2-2 Causes of Hyperhomocysteinemia
Elevations in Hcy concentration may be triggered by a diverse group of stimuli
internal and external to the body. Diet, genetics, medications, lifestyle, and systemic illnesses
may all, separately or in combination, result in HHcy (Refsum et al., 2004).
2-2-1 Genetic Defects
Inherited deficiencies of enzymes in the methionine-Hcy pathway produce HHcy (Guthikonda and Haynes, 2006), which are observed in individuals with homozygous genetic
defects affecting cystathionine β-synthase (CβS), N-5,10-Methylene Tetrahydrofolate
reductase (MTHFR), or any of several enzymes responsible for the conversion of vitamin
B12 to its methionine synthase-associated cofactor form (Miller, 2013).

Literature Review
7
These autosomal recessive genetic disorders, collectively termed homocystinuria
because Hcy accumulates in the urine as well as the blood, are associated with severe
premature vascular disease, including thrombosis and atherosclerosis ,mental retardation,
dislocation of the eyelens and skeletal malformations (Miller, 2013).
a- Cysthathionine beta synthase deficiency
Cysthathionine beta synthase (CβS) is responsible for the irreversible degradation of
homocysteine. Without CβS, the entire transsulfuration pathway of the methionine cycle is
shut off (Brandon, 2009).
Mutations in the gene coding for the enzyme (CβS) lead to classical homocystinuria with
severe HHcy and/or homocystinuria (Gaustadnes et al., 1998), which is an autosomal
recessive disease characterized by a severely elevated plasma Hcy and Hcy excretion in the
urine (Liselotte, 2003).
b- N-5,10-Methylene Tetrahydrofolate reductase deficiency
N-5,10-Methylene Tetrahydrofolate reductase (MTHFR) is a folate cycle enzyme that
generates the methyl donor, 5-methyltetrahydrofolate, that is used for Hcy remethylation by
MS (Lentz, 2005). The most common one that is detected worldwide and has a high
incidence in different populations, is single nucleotide polymorphisms of N-5,10-methylene
tetrahydrofolate reductase which has been associated with mild (13–24 μM) and moderate
(25–60 μM) HHcy (Curro et al., 2014). The most common enzyme defect associated with
moderately raised total Hcy is a point mutation (C-to-T substitution at nucleotide 677) in the
coding region of the gene for MTHFR, which is associated with a thermo labile MTHFR
variant that has about half-normal activity (Hankey and Eikelboom, 1999).
2-2-2 Vitamin Deficiencies
Hyperhomocystenemia can also arise from nutritional deficiencies of folate, vitamin
B6, and vitamin B12 (Curro et al., 2014) which are essential cofactors in Hcy-methionine
metabolism. Therefore, low vitamin B availability (B6, B12 and folic acid) leads to impaired
remethylation of Hcy to methionine and thus to Hcy accumulation (Mangge et al., 2014).
However, the nature of HHcy caused by vitamin B6 deficiency differs from that caused by
folate and vitamin B12 deficiencies. In vitamin B6 deficiency, fasting blood levels of Hcy are

Literature Review
8
usually not elevated or only slightly elevated. However, after a protein meal or after
consumption of an oral methionine load does plasma Hcy become abnormally elevated in
vitamin B6-deficient patients. In contrast, plasma Hcy levels tend to be elevated regardless of
prandial state in patients with folate or vitamin B12 deficiency (Miller, 2013).
2-2-3 Other Causes of Hyperhomocysteinemia
Several diseases such as renal and thyroid dysfunction, cancer, psoriasis, and diabetes
as well as various drugs, alcohol, tobacco, coffee, older age and menopause, are believed to
be associated with moderately elevated Hcy concentrations (Faeh et al., 2006).
A rise in serum creatinine also leads to a rise in fasting total homocysteine (Hankey and
Eikelboom, 1999). Other causes of HHcy include leukemia (Refsum et al., 1991), sickle cell
anemia (Houston et al., 1997), polycythemia vera, and idiopathic thrombocytosis (Gisslinger
et al., 1999).
3- Pathogenicity of Hyperhomocysteinemia
3-1 Hyperhomocysteinemia and cardiovascular disease
Cardiovascular diseases (CVD) comprise a class of diseases that involve heart and
systemic blood vessels. A large number of epidemiological studies have demonstrated that
mild HHcy is a prevalent risk factor for stroke, cardiovascular disease, and venous
thromboembolism (Den Heijer et al., 2005).
3-1-1 Endothelial dysfunction
Endothelium is composed by a single layer of endothelial cell, which lines the interior
surface of vascular lumen, between blood and vascular smooth muscle cells (VSMC) of all
kinds of blood vessels and the whole circulatory system (Lai and Kan, 2015). It plays an
important role in many physiological functions, including the control of blood cell trafficking,
vasomotor tone, vessel permeability, and hemostatic balance (Aird, 2007).
Endothelial cells produce a wide variety of substances in response to various physical
and chemical stimuli, including vasodilator substances such as nitric oxide (NO), prostacyclin
(PGI2), and endothelium-derived hyperpolarizing factor (EDHF), and vasoconstrictor
substances such as endothelin-1 (ET-1), angiotensin II (Ang II), thromboxane A2 (TXA2) or
free radicals (Aird, 2004).

Literature Review
9
Endothelial dysfunction can be described as an imbalance between vasodilator and
vasoconstrictor produced by the endothelium, and it has been regarded as the core systemic
pathological status in the process of atherosclerosis and cardiovascular disease (Lai and Kan,
2015) (Figure 02).
Three mechanisms have been suggested explaining HHcy-could leads to impaired
Endothelial-dependent dilatation:
1- Oxidative stress conditions: the disruptive uncoupling of NO synthase activity,
quenching of NO, and enzymatic inhibition;
2- Endoplasmic reticulum stress with eventual endothelial cell apoptosis;
3- Chronic inflammation/prothrombotic conditions.
.
Figure 02. Potential mechanisms of homocysteine-induced endothelial dysfunction
(Lai and Kan, 2015)

Literature Review
10
3-1-1-1 Homocysteine induced oxidative stress condition
Oxidative stress is defined in general as an imbalance between elevated ROS/RNS
production and removal by the endogenous antioxidant system (José, 2014). The reactive
oxygen species (ROS) include free radicals such as superoxide (•O2-), hydroxyl (•OH),
peroxyl (•RO2), hydroperoxyl (•HRO2-), as well as non radical species such as hydrogen
peroxide (H2O2).
The reactive nitrogen species (RNS) include free radicals like nitric oxide (•NO) and
nitrogen dioxide (•NO2), as well as non radicals such as peroxynitrite (ONOO-), nitrousoxide
(HNO2) and alkyl peroxynitrates (RONOO). Of these reactive molecules, •O2-, •NO and
ONOO- are the most widely studied species and play important roles in the cardiovascular
complications (Jeanette et al., 2005).
Endogenous sources of ROS can be subdivided into mitochondrial and cytosolic,
including NADPH oxidases (Nox), uncoupled NO synthases (NOSs), and xanthine oxidase
(XO). Noting that in cardiomyocytes, mitochondria are the major source of ROS, which are
generated as byproducts of electron flow through the electron transport chain (ETC),
predominantly at complexes I and III. (Figure 03).
Figure 03. Major endogenous sources of reactive oxygen species (ROS) and reactive nitro-
gen species (RNS) in the cardiomyocyte (José, 2014)

Literature Review
11
a- Uncoupling of NO synthase
Nitric oxide (NO) has been shown to be a major modulator of vascular homeostasis and to
have a vasoprotective effect against atherosclerosis (Moncada et al., 1991). It inhibits platelet
aggregation, leukocyte migration, and adhesion to endothelium, and it also attenuates vascular
muscle cell proliferation and migration, which collectively promote atherosclerosis (Knowles
et al., 2000).
Nitric oxide is majorly synthesized by the endothelial isoform of NO synthase (eNOS) in
response to the vasodilation stimulus. Endothelial NO diffuses across to VSMC where it
activates cytosolic guanylylcyclase, increases cyclic GMP production, and leads to vascular
smooth muscle relaxation (Deanfield et al., 2005).
Therefore, the loss of endothelial-mediated vasodilatory ability that characterized by the
tipping of the vascular balance toward an abnormally constrictive, inflammatory and
prothrombombic state is considered to be one of the earliest manifestations of cardiovascular
damage ( lai and kan, 2015 ), and is considered to be pivotal in the initiation and progression
of atherosclerosis (Ross , 1999) .
In endothelial cells, eNOS is inactive when it is bonded with caveolin 1 (cav-1). When it
becomes active, eNOS disassociates from cav-1 and binds with calmodulin (CAM) and heat
shock protein 90 (Hsp90) and together with phosphorylation of serine sites lead to the
vasodilataion (Kietadisorn et al., 2012) (figure 04).
Tetrahydrobiopterin (BH4), an essential cofactor of eNOS, is necessary for optimal eNOS
activity (Kietadisorn et al., 2012). The reduction in BH4 availability, followed by the
uncoupling of eNOS, is the significant mark in Hcy-mediated oxidative stress (Dhillon et al.,
2003; Rochette et al., 2013). This is an exact feature of endothelial dysfunction that directly
precedes the appearance of atherosclerosis (He et al., 2010).

Literature Review
12
Figure 04. Central role of endothelial NO synthase (eNOS) uncoupling in the pathogenesis
of endothelial dysfunction (Kietadisorn et al., 2012)
Homocysteine also induces NADPH oxidase activity (NOX). There are multiple
isoforms of NADPH NOX, with endothelial cell mostly exhibiting the isoform NOX2. Hcy
increased endothelial cells NADPH oxidase expression in a time- and dose-dependent manner
(Tyagi et al., 2005). The up-regulated NADPH oxidase likely represents an initiating source
of oxidative stress in endothelial cells that triggers other dormant ROS producers in HHcy (lai
and kan, 2015 ).
Homocysteine contains a highly reactive sulfhydryl (-SH) group. The sulfhydryl group
readily self-oxidizes to form disulfide linkage with other free thiols, along with the generation
of superoxide radicals as a byproduct (McDowell and Lang , 2000). In addition, the self-
oxidation of Hcy to Hcy and Hcy-thiolactone generates (ROS) and further contributes to the
vascular toxicity of homocysteinemia (Andersson, 1995).
b- Accumulation of Asymmetric Dimethylarginine
Recently, the protein asymmetric dimethylarginine (ADMA), an endogenous eNOS
inhibitor, has garnered interest as a potential biomarker for endothelial dysfunction (Zhang et

Literature Review
13
al., 2012). Hcy is also known to decrease NO production by increasing the ADMA (Eren et
al., 2014). Elevated plasma ADMA is an emerging cardiovascular risk factor that is prevalent
in patients with hypercholesterolemia, hyperhomocysteinemia, diabetes mellitus, and
hypertension (Boger, 2003). In addition to inhibiting production of NO, ADMA may also
promote the uncoupling of eNOS, directly contributing to increased oxidative stress
(Vallance, 2001; Boger, 2003). Elevated plasma ADMA correlated directly with impairment
of endothelium-dependent relaxation of the carotid artery in monkeys with
hyperhomocysteinemia caused by a diet enriched with methionine and deficient in folate
(Boger, 2000). Elevated plasma ADMA was also reported to correlate with impaired
endothelial function in a rat model of HHcy (Fu et al., 2005) and in human subjects with
acute HHcy induced by oral methionine loading ( Stuhlinger et al., 2003).
3-1-1-2 Homocysteine induced protein modification and endoplasmic
reticulum stress
The Endoplasmic Reticulum (ER) plays a pivotal role in proper assisted protein
folding and post-translational modifications of proteins for appropriate function, membrane
targeting and secretion. Any process that interferes with ER function results in unfolded
protein response (UPR) and ER stress (Veeranki and Tyagi, 2013).
Elevated Hcy levels lead to ER stress and induce protein modification through an
alternative mechanism mediated by the cyclic thioester form of Hcy (homocysteine
thiolactone). Homocysteine thiolactone is formed by methionyl-tRNA synthetase as an error-
editing reaction when homocysteine becomes mis-incorporated into methionyl-tRNA in place
of methionine (Santulli and Iaccarino, 2013).
When intracellular concentrations of Hcy become elevated, Hcy can participate in
disulfide exchange reactions with ER proteins, leading to the misfolding of newly synthesized
secretory and membrane proteins such as thrombomodulin (Lentz and Sadler, 1993). The
cellular consequences of this ER stress, include dysregulation of lipid metabolism, activation
of inflammatory pathways, and impaired insulin signaling. ER stress can also lead to
apoptotic cell death (Kaufman, 2002).
3-1-1-3 Homocysteine induced inflammatory/prothrombotic conditions
Hyperhomocystenemia enhanced vascular inflammation (Durand et al., 1997).

Literature Review
14
During inflammation (Figure 05), Tumor Necrosis factor-α (TNF-α) exerts its effects
on the endothelium through its receptor, TNFR. Binding of TNFR by TNF-α leads to
diminished eNOS protein expression via suppression of promoter activity and destabilization
of its mRNA. TNFR suppresses eNOS activity by preventing the degradation of its
endogenous inhibitor, ADMA. TNFR signaling also induces the transcription factor NF-κB
leading to enhanced expression of intercellular adhesion molecules: intercellular adhesion
molecule-1 (ICAM-1); vascular cell adhesion molecule-1 (VCAM-1), TNF-α and NADPH-
oxidase-1(Nox1). NF-κB induction is also mediated by oxidized low density lipoprotein
(oxLDL), ROS and binding of various autoantibodies (AECA: anti-endothelial cell
antibodies; APLA: antiphospholipid antibodies; anti-oxLDL: anti-oxidized LDL antibodies).
eNOS uncoupling, mediated in part by ROS, is associated with reduced NO production and
enhanced generation of ROS. eNOS activity is also suppressed by oxLDL (Steyers and
Miller, 2014).
Figure 05. Regulatory circuits in inflammation and endothelial dysfunction
(Steyers and Miller, 2014)

Literature Review
15
HHcy also induces a prothrombotic condition (Dayal et al., 2006) including enhanced
platelet activation, enhanced coagulation (Undas et al., 2005), and attenuated fibrinolysis,
resulting from posttranslational modification of fibrinogen by homocysteinylation (Sauls et
al., 2006).
3-1-2 Atherosclerosis
The mechanism of atherosclerotic disease in HHcy is directly related to vascular
endothelial cell damage which leads to vascular endothelial dysfunction, and enhanced
oxidative stress (Malinowska et al., 2012; Yilmaz, 2012).
Atherosclerosis initiates from disrupted endothelium which allows circulating
apolipoprotein B (apoB) containing lipoproteins to penetrate and accumulate in
subendothelium where they further undergo chemical modification. Modified lipoproteins,
particularly, oxidized low-density lipoprotein (LDLs), promote the proinflammatory
phenotype of endothelial cells for increased vascular cell adhesion protein 1 (VCAM1) and
intercellular adhesion molecule 1 (ICAM1) expression and proinflammatory cytokine
production, all of which attract circulating white blood cells homing to the lesion site (Estruch
et al., 2013; Milstone et al., 2015). Following infiltration into the lesion site, monocytes,
dendritic cells and T lymphocytes uptake fat and cholesterol to become foam cells that
aggravate the inflammation cascade site (Haka et al., 2015; Cochain and Zernecke 2015)
(Figure 06).
Elevated plasma Hcy has been considered as an independent risk factor for
atherosclerotic vascular disease (Bautista et al., 2002; Cui et al., 2008).
Indeed, increased oxidative stress, alterations of lipid metabolism and induction of
thrombosis have been suggested to be pathogenic links which are present between HHcy and
atherosclerosis (Eren et al., 2014).

Literature Review
16
Figure 06. Hyperhomocysteinemia and etiopathogenesis of atherosclerosis
(Sainani et al., 2008)
Homocysteine-induced histopathologic findings include thickness of the intima layer,
disruption of the elastic lamina, smooth muscle hypertrophy, platelet aggregation, and the
white thrombus formation (McCully, 1969; Tsai et al., 1994). These changes may be mediated
by several pathophysiologic mechanisms: upregulation of the monocyte chemoattractant
protein-1 and interleukin-8 expression and secretion, and subsequent leukocyte recruitment ]
(Poddar et al.,2001); binding of thiolactone with low density lipoprotein (LDL) cholesterol to
form aggregates, which are phagocytosed by macrophages in tunica intima that in turn enrich
the atherosclerotic plaques with lipid (McCully, 1996) smooth muscle cell proliferation and
increased collagen production (Majors et al ., 2002 ); attenuation of endothelial tissue
plasminogen activator binding sites; increased blood viscosity; protein C inhibition; factors
VIIa and V activation; increased fibrinopeptide A and prothrombin fragments 1 and 2;
decreased endothelial antithrombotic activity (Nappo et al., 1999); increased oxidative stress
(Mansoor et al., 1995 ) and platelet aggregation (McCully and Carvalho, 1987).
The study of (Lentz, 2005) demonstrate that HHcy was shown to accelerate the
development of atherosclerosis in susceptible models such as the apolipoprotein E (apoE)-
deficient mouse. (Hofmann et al., 2001) reported that apoE deficient mice fed a
hyperhomocysteinemic diet for 8 weeks developed atherosclerotic lesions in the aortic sinus
that were of greater size and complexity than those seen in apoE-deficient mice fed normal
chow. The vascular lesions in the hyperhomocysteinemic apoE-deficient mice contained high

Literature Review
17
levels of inflammatory mediators such as the leukocyte adhesion molecule, VCAM-1
(Hofmann et al., 2001).
3-2 Hyperhomocysteinemia and dyslipedimia
Dyslipidemia, as a risk factor of CVD, is manifested by elevation or attenuation of
plasma concentration of lipoproteins. Generally, it is defined as the total cholesterol, LDL,
triglycerides, apo B or Lp (a) levels above the 90th percentile or HDL and apo A levels below
the 10th percentile of the general population (Dobsn et al., 1996).
Both Hcy and lipids are toxic in vascular cells and hepatocytes which could indicate
interactions between the two pathways. Possible mechanism might be that the intake of
saturated fatty acids can lead to increased Hcy by increasing the production of
phosphatidylcholine (PC) from phosphatidylethanolamine (PE) via the phosphatidyl
ethanolamine methyltransferase (PEMT) pathway (Berstad et al., 2007).
Phosphatidyl ethanolamine methyltransferase consumes three SAM molecules for
transforming (PE) to (PC). The reaction produces three SAH molecules that are hydrolyzed to
Hcy via SAH-hydrolase (Figure 07). Another possible explanation could be that a diet rich in
fatty acids might contains more methionine, the precursor of Hcy (Obeid and Herrmann,
2009).
Figure 07. Possible interactions between hyperhomocysteinemia and hyperlipidemia in cell
pathology (Obeid and Herrmann, 2009)

Literature Review
18
Studies have reported that oxidative stress and inhibition of NO release were induced
by Hcy, which also promoted a lower expression of paraoxonase 1 (PON1) and enhanced the
production of ROS and a lower activity of PONs, in patients with HHcy. Hcy-induced ROS
downregulates the expression of HDL-associated PON1, which accelerates the development
of atherosclerosis (Maron and Michel, 2012; Eren et al., 2013). Paraoxonase 1 is a HDL-
associated enzyme esterase which appears to contribute to the anti-oxidant and anti-
atherosclerotic capabilities of HDL (Parra et al., 2007).
3-3 Hyperhomocysteinemia and hepatic disease
The liver is central for the synthesis and metabolism of Hcy and related thiols, given
that the majority of dietary methionine is metabolized in this organ (Mato and Lu, 2005). The
Changes of Hcy metabolism were reported during liver damage associated to alterations of
lipid metabolism (Werstuck et al., 2001; Obeid and Herrmann, 2009).
Hyperhomocystenemia is also implicated in hepatic disorders, such as alcoholic liver disease
(Roblin et al., 2007), cirrhosis (Bosy- Westphal et al., 2001), steatosis and fibrosis (Adinolfi
et al., 2005; Ventura et al., 2005). This correlation is pertinent, as far as the liver is central in
Hcy metabolism (Brosnan et al., 2004).
3-4 Hyperhomocysteinemia and carcinogenesis
For the past several years, a link has been established between certain cancers and
elevated plasma Hcy. Increased plasma Hcy concentration is a risk factor for cancer and even
as a novel tumor marker (Plazar and Jurdana, 2010). Folate depletion promotes the
development of cancer, particularly colorectal cancer, whereas high doses of folic acid
enhance the growth of cancer cells. Folate, Vitamin B12, and Vitamin B6 have a number of
biologic roles that make them potentially important in cancer (Qureshi et al., 2016).
Defective metabolism of Hcy in carcinogenesis is well documented, but the
pathophysiology is not fully understood (Fassbender et al., 1999; Sun et al., 2002). Malignant
cells are characterized by high a growth rate, and the methionine requirement increases in
these cells due to increased protein synthesis and transmethylation reactions. Normal cells
meet their methionine requirement by synthesizing it from homocysteine. In contrast,

Literature Review
19
methionine-dependent malignant cells in organs such as the lung, kidney, breast, colon and
bladder cannot convert Hcy to methionine, which results in Hcy accumulation (Cellarier et al.,
2003).
4- Therapy of Hyperhomocysteinemia
The primary goal of treatment is to lower blood levels of Hcy to normal. Treatment for
HHcy involves the use of vitamins, such as folic acid, Vitamin B12, and pyridoxine. Folic
acid and vitamins predominantly act under fasting condition and pyridoxine acts after meals.
Pyridoxine reduces Hcy levels by 22%. Folic acid alone reduces Hcy level by 22% and
Vitamin B12 by 11%. When both administered together, it causes a reduction of 38.5%
(Lehmann et al., 2003).
5- Argania spinosa
The herbs have been the basis for many medicinal therapies. Among these herbs: Argania
spinosa.
5-1 Description
Argania spinosa is a tropical tree that belongs to the Sapotaceae family (Chaussod et al.,
2005). This plant is endemic in southwestern Algeria and Morocco (Msanda et al., 2005).
Because of its ability to survive to arid and semi arid regions (Naggar and Mhirit, 2006), It
protects soil from desertification and erosion (Alados and El Aich, 2008).
In addition to these important ecological aspects. The Argan tree is exploited essentially
for its fruits. The endosperm seed of the fruit constitutes a good potential source of edible oil
for human consumption and is endowed with important medicinal properties (Charrouf and
Guillaume, 1999). The leaves of this tree are also used as "hanging forage" for cattle (goats
and sheep) and this forage is complemented by the energetic leftovers obtained after the oil
preparation (Charrouf and Guillaume, 1999). Indeed, Argan oil is rich in essential
polyunsaturated fatty acids. It is a source of oleic acid (47.7%) and linoleic acid (29.3)
(Rahmani, 2005) and it is rich in minor and noble compounds like tocopherols, polyphenols,
sterols, carotenoids, xanthophyls, squalen (Khallouki et al., 2005), and saponins (Guillaume
and Charrouf, 2005).

Literature Review
20
5-2 Botanical classification
Figure 08. Kernel and tegument of Argania spinosa L (Ould safi, 2014)
Kingdom Plantae
Sub-division Angiospermae
Class Dicotylédonae
Sub-class Asteridae
Order Ericales
Family Sapotaceae
Genus Argania
Species Argania spinosa
Tegument
Kernel

Literature Review
21
Figure 09. Distribution area of the Argan tree in Algeria (Kechairi, 2009)
5-3 Therapeutic properties
The main traditional use of Argan oil is by far for nutritional purposes. Natives either
directly eat the oil on toasts, generally for breakfast, or use it for frying.
As cosmetic, the oil is traditionally indicated to cure all kind of pimples on the skin and
more particularly juvenile acne and chicken pox pustules. It is also recommended to reduce
dry skin problems and slow down the appearance of wrinkles. It is also used in rhumatology.
For these indications, the oil is used as a skin lotion and applied on the area to be cured.
(Charrouf and Guillaume, 1999).
In addition, Many scientific studies have reported that the oil has many pharmacological
effects, such as antioxidant (El Baabli et al., 2010), antiproliferative (Bennani et al., 2007),
cardioprotective (Charrouf et al., 2007) and hypolipemiant activities (Drissi et al., 2004).

Literature Review
22
6- Bacterial Biofilm and cardiovascular diseases
6-1 Definition of Biofilm
A biofilm is a structured community of bacterial cells enclosed in a self-produced
matrix that adheres to inert or living surfaces, including tissues, industrial surfaces, and
artificial devices, such as intrauterine contraceptive devices, implants and prosthetic medical
devices, catheters, dental materials, cardiac valves, and contact lenses. Biofilms form when
bacterial colonizers adhere to surfaces in aqueous environments and excrete a slimy, glue-like
substance composed of exopolysaccharides (EPS). The EPS can consist of cellulose,
alginates, poly-N-acetylglucosamine, extracellular teichoic acid, various proteins, lipids, and
extracellular RNA or DNA (Sun et al., 2013).
One of the most important characteristic of biofilms is their increased tolerance to
antimicrobial agents (Wimpenny et al., 2000). Bacteria within a biofilm are several orders of
magnitude more resistant to antibiotics, compared with planktonic bacteria (Rabin et al.,
2015)
Biofilms contain channels that allow water, nutrients and oxygen circulation (De Beer
et al., 1994). However, during biofilm formation a gradient of available substances is
established, making the outer layers becoming aerobic and metabolically active, while the
inner ones become anaerobic, nutrient deficient and slowed down growth (Werner et al.,
2004; Costerton et al., 2005; Bjarnsholt et al., 2013).
6-2 Stages of Biofilm formation
The process of biofilm formation is complex, but generally recognised as consisting of
five stages (Palmer and White, 1997) (Figure 10):
Development of a surface conditioning film;
Movement of microorganisms into close proximity with the surface;
Adhesion (reversible and irreversible adhesion of microbes to the conditioned
surface);
Growth and division of the organisms with the colonisation of the surface,
microcolony formation and biofilm formation; phenotype and genotype changes and
Biofilm cell detachment/dispersal each of these processes will be considered in turn.

Literature Review
23
Figure 10. Various stages of biofilm formation and development (Gupta et al., 2016)
6-3 Biofilm and cardiovascular infections
The human body contains a large number of bacteria but their localization in healthy
individuals is normally restricted to certain body areas such as the skin, the mucosae of buccal
and nasal cavities, vagina and the gastrointestinal tract (Costello et al., 2009; Ma et al., 2012).
and endothelium (Kokare et al. 2009).
The biofilm infection strategy is one of attaching to host tissue surfaces, producing
aging of host cells in the area of infection, and providing nutrition to the biofilm constituents
through inflammatory pathways which lead to the production of plasma exudate that can be
used by the community as a nutrient source (Hall-Stoodley et al., 2004; Hall‐Stoodley and
Stoodley, 2009). Biofilms can cause chronic infections and are associated with a number of
chronic disease states including cystic fibrosis, infectious endocarditis, and chronic wounds
(Singh et al., 2000; James et al., 2008).
Infections of the cardiovascular system, including those involving prostheses and
devices, are a globally recurring problem. Vascular infections are often life-threatening,
spread easily, and costly to treat. Furthermore, infection is a common problem affecting the
success of biomedical implants, such as vascular stents (Habash and Reid, 1999). Bacteria can

Literature Review
24
be introduced through surgical interventions, travel through the bloodstream and infect the
endothelial cells lining the blood vessels. Cardiovascular disease has also been linked to
microbial infection (Lowy, 1998), with attachment of bacterial pathogens to endothelium or
extracellular matrix being an initial step in the process (Beachey, 1981).
Atherosclerotic plaque contains bacteria and other microorganisms (Fabricant et al.,
1978; Ott et al., 2006). However, early efforts to determine the clinical significance of the
presence of these microorganisms in plaque proved inconclusive, mainly due to the failure of
poorly designed antibiotic trials (Rosenfeld and Campbell, 2011). Many epidemiological
studies have established positive associations between cardiovascular disease risk factors,
morbidity, mortality, and markers of infection. Specific pathogens along with their potential
contribution by direct or indirect mechanisms to atherosclerosis pathogenesis have been
reviewed (Rosenfeld and Campbell, 2011).
First, the most common source of microorganisms within atherosclerotic plaques most
closely correlate with the oral microbiome, rather than bacteria from any other niche, such as
gut, skin, or sinus (Hayashi et al., 2010; Jain and Douglas, 2014). Secondly, the arrangement
of the microorganisms within the plaques is heterogeneous (Wolcott et al., 2012), in that the
samples followed a pattern of regions of high microbial density directly adjacent to an area
which was almost void of microbial DNA. Finally, it has been shown that the regions of high
microbial density were polymicrobial. These features suggested microorganisms at the
arterial wall are in biofilm mode of growth (Dalton et al., 2011; Wolcott et al., 2012).
The general hypothesis that chronic infections can contribute to the development of
atherosclerosis has come from: direct effects of infectious agents on cellular components of
the vessel wall; increased expression of cytokines, chemokines; and cellular adhesion
molecules, resulting in local endothelial dysfunction and immune responses targeted at self-
proteins located in the vessel wall due to molecular mimicry (Epstein et al., 1999).

Literature Review
25
6-4 Bacterial strains
Clinical isolated bacteria used in the study are:
6-4-1 Streptococcus mutans
Streptococcus mutans, a Gram-positive facultative anaerobic bacterium, is generally
known to be a pathogen of dental caries (Hamada and Slade, 1980), and its surface protein
antigens have been investigated to clarify their role as virulence factors. It is known to be
associated with bacteremia and infective endocarditis (IE). It is also of interest that S. mutans
was shown to possess these two, indicating that the bacterium is a possible candidate for
inclusion in the group of bacterial species involved with atheromatous plaque formation
properties (Kuramitsu et al., 2001 ; Chia et al., 2004; Nakano et al., 2005).
The role of S. mutans in atherogenesis has been investigated. Although these bacteria are
capable of invading endothelial cells and stimulating the production of inflammatory markers,
in addition to being detected at a high frequency in these lesions (Nakano et al., 2006; Nakano
et al., 2009).
Several in vitro studies have shown that S. mutans has the ability to adhere to collagen
type 1 (Nomura et al., 2012), induce platelet aggregation (Matsumoto-Nakano et al., 2009),
invade human endothelial cells, and induce increased production of interleukin (IL) 1, IL-6,
monocyte chemoattractant protein 1 (MCP-1) and foamy macrophages, which are strongly
associated with the pathogenesis of atherosclerosis (Nagata et al., 2011). Studies using animal
models observed that an infection with the invasive strain of S. mutans OMZ175 accelerates
the development of atherosclerotic plaques and increases the inflammatory response in an
ApoE-null mouse when compared to the control without S. mutans infection (Kesavalu et al.,
2012). These results suggest that invasive strains of S. mutans may be related to vascular
disease in humans, possibly contributing to the progression of atherosclerotic lesions.
6-4-2 Streptococcus intermedius and Streptococcus anginosus
Streptococcus intermedius and Streptococcus anginosus are two members of the
Streptococcus anginosus group (SAG), also known as the "Streptococcus milleri" group, one
of five groups collectively known as viridans group streptococci, consists of the species S.
intermedius, S. anginosus, and S. constellatus (Whiley and Beighton, 1991) . A variety of
clinical diseases have been associated with infection with the different members of the SAG
(Claridge et al., 2001).

Literature Review
26
Streptococcus intermedius has a tendency to cause abscess formation commonly found in
the liver and brain, but is rarely the etiologic agent in infective endocarditis (Whiley et al.,
1992; Rashid et al., 2007). Previous studies on infective endocarditis caused by SAG have
relied on phenotypic methods for identification. (Sussman et al., 1986) studied 36 patients
with viridans streptococcal endocarditis, and identification at the species level was determined
by using biochemical tests. Four of the cases were found to be S. intermedius.
Recently, (Cooper and Gotoff, 2016) reported a case of purulent pericarditis with associated
subdiaphragmatic and hepatic collections due to S. anginosus.
6-4-3 Staphylococcus haemolyticus
Staphylococcus haemolyticus plays an important role in hospital-acquired opportunistic
infections related to implanted medical devices (Mack et al., 1996; Mehta et al., 1997; Mack
et al., 2006). Furthermore, S. haemolyticus has the highest level of antimicrobial resistance
among all CoNS (Coagulase-negative staphylococci) (Froggatt et al., 1989; Chiew et al.,
2007).
The study of Ott et al., (2006) reported that S .haemolyticus among bacterial agents could
have secondarily colonized atheromatous lesions and could act as an additional factor
accelerating disease progression.
6-4-4 Streptococcus uberis
Streptococcus uberis is an environmental Gram-positive bacterium belonging to the
Streptococcaceae family. It is responsible for a high percentage of mastitis in dairy cattle and
it is rarely associated with human infections (Zadoks, 2007). Only a few case reports have
described human infections with this microorganism, which is widely difficult to identify.
In vitro experiments demonstrated that S. uberis can readily develop penicillin
resistance, and microbial analysis of bacterial population in treated milk showed that S. uberis
can grow even in cold storage (Haenni et al., 2010; Rasolofo et al., 2010). It has been
hypothesized that the stability of this pathogen under various environmental conditions and
the expression of virulence factors (Haenni et al., 2010), may expand the pathogenicity of
this bacterium from cattle to humans as described for Streptococcus agalactiae, a pathogen
that is associated with both animal and human diseases ( Zadoks et al., 2011).
Recently, Gulen and his collabators (2013) identified S. uberis from urine samples of
seven of 148 patients by phenotypic methods.

Materials and
Methods

Materials and methods
27
1- Biological plant
In this study, Argania spinosa seeds were collected from Tindouf which is located in
South west of Algeria, on December 2011 (L.O.S.T As. 12. 11).
The powdered seeds of A. spinosa seeds were obtained by cutting the fruits into pieces
to obtain seeds, and then the seeds were subjected to size reduction to a coarse powder using a
mechanical grinder.
2- Animals
2-1- Experimental animals
The experiments were performed on 28 adult male abino Mus Musculus mice (2.5- 3
month old), weighing (30– 35g), given from central pharmacy, Algeria.
The animals were separated and housed 7 per plastic cages covered with a stainless
wire netting, a layer of sawdust is placed at the bottom of each cage, and it is regularly renewed
every two days. They were maintained under standard laboratory conditions of humidity,
temperature and light.
Animals were fed with normal commercial pellet diet (LA RATION, Bouzeriaa, Algeria) and
water ad libitum. The animals were acclimatized to laboratory conditions for one week prior
to experiment.
Table 02: Composition of diet for 1 kg of food taken by the mice during 21 days (ONAB):
Composition Amount in g / kg Percentage %
Corn 620 62
Soja 260 26
Phosphate 16 1,6
Limestone 9 0,9
Cellulose 10 1
Minerals 10 1
Vitamins 10 1

Materials and methods
28
2-2- Experimental treatments
Animals were divided into four groups of similar mean body weights and fed for 21
days with control and experimental diet:
The control group (F) was fed with white bread (0.50 mg/mice);
The second group (M) was fed with L-methionine (500 mg/kg/day);
The third group (MP) was fed with L-methionine (500 mg/kg/day) and
treated with A. spinosa powdered seeds (150mg/kg/day), While;
The positive control group (P) was treated with the powdered seeds of A.
spinosa (150 mg/kg/day) only.
The methionine and plant extract were given in white bread (0.50mg/mice), and animals
were allowed free access to food and water.
During all the treatment period (21 days), body weight of the mice was measured daily
at the same time.
Each dose of methionine or plant is incorporated into a flour ball (0.50 mg) and then
administered to the mouse orally.
Table 03: Treatment of mice (n=7, for 21 days).
2-3- Blood and tissue sampling
At the end of the experiments, blood samples were collected after fasting, from the retro
orbital vein into EDTA tubes by using glass capillaries. They were centrifuged immediately,
and plasma was frozen under -20°C until assay time.
Experimental
group
Substance
administered
Daily dose
F
Flour 0,5 mg/mice
M
Flour +L-Methionine 500 mg/kg/mice
MP
Flour+ L-methinine +
plante
500 mg/kg+150mg/kg/mice
P
Flour +Plante 150 mg/kg/mice

Materials and methods
29
After the blood samples collection, the animals were sacrificed. Then, organs used for
histological analysis (Aorta, Heart and Liver) were quickly removed, rinsed with saline solution
(0.9%), and fixed in formalin 10%. The techniques used in this research study are summarized
in (figure 11).
Figure 11. Blood and tissue sampling.
28 adult male Abino Mus
Musculus mice
The positive
control group (P) d’engraissement
(15 semaines)
Blood sample
Histological investigations of aorta, heart and liver
The third group
(MP)
iment
d’engraissement
(15 semaines)
8 lapins nourris
à l’aliment
d’engraissement
The second group
(M) d’engraissement
(15 semaines)
The control group
(F)
d’engraissement
(15 semaines)
Tissue sample
Blood sample Blood sample Blood sample
Tissue sample Tissue sample Tissue sample
Dosage of biochemical
parameters: Hcy, TG,
CHO, HDL-c , LDL-c,
ALT, AST
FSH, LH, Testostérone,
Oestradiol
Determination of
oxidative stress
parameters : GSH and
catalase activity
Dosage of biochemical
parameters: Hcy, TG,
CHO, HDL-c , LDL-c,
ALT, AST
FSH, LH, Testostérone,
Oestradiol
Determination of
oxidative stress
parameters: GSH and
catalase activity
Dosage of biochemical
parameters: Hcy, TG,
CHO, HDL-c , LDL-c,
ALT, AST
FSH, LH, Testostérone,
Oestradiol
Determination of
oxidative stress
parameters : GSH and
catalase activity
Dosage of biochemical
parameters: Hcy, TG,
CHO, HDL-c , LDL-c,
ALT, AST
FSH, LH, Testostérone,
Oestradiol
Determination of
oxidative stress
parameters : GSH and
catalase activity

Materials and methods
30
3- Methods
3-1Chemical products
Chemical products used in our study are:
L-methionine, chloroform, NaCL 0.9%, formalin 10%, dithiobis-2-nitrobenzoic acid (DTNB),
sulfo-salicylic acid (0.01M), Bovine Serum Albumin (BSA), orthophosphoric acid (85%), Tris
Ethylene Di-amine Tetra Acetic acid (EDTA, 0.02M), different concentrations of ethanol
(50%, 70%, 95% and 96%), HCl, NaOH, NaCl, butanol, xylene, paraffin and glycerin.
3-2 Equipments
Precision weighing balances (readability 0.01g) to determine the weight of the mice,
Precision Weighing Balances (readability 0.0001g) to determine the quantity of methionine,
Heating magnetic stirrer, pH meter, Centrifuge, Spectrophotometer , Oven, Microtome and
Photo microscope connected to computer.
3-3 Biochemical analysis
Plasma Hcy and lipids status determination were performed in the medical laboratory of
IBN SINA, Constantine.
3-3-1 Plasma Hcy determination
Homocysteine levels were measured by competitive solid phase chemiluminescance
immunoassay (IMMULITE).
Homocysteine involved a preliminary manual sample pretreatment step. Hcy in the
plasma sample is released from its binding proteins and converted to SAH by an off-line 30
minute incubation at 37°C in the presence of SAH hydrolase and Dithiothreitol. The treatment
sample and alkaline phosphate –labeled-anti-SAH antibody are simultaneously introduced into
a test unit containing an SAH coated polystyrene bead. During 30 minutes of incubation, the
converted SAH from the sample completes with the immobilized SAH for binding the alkaline
phosphatase labeled-anti SAH antibody conjugated. Unbound enzyme conjugated is removed
by centrifugal wash. The substrate is added and the procedure continues as described for the
typical immunoassays.

Materials and methods
31
3-3-2 Lipids determination
Total cholesterol, HDL-c, LDL-c and triglycerides concentrations were assessed using
colorimetric automatic procedures (Auto-analyzer type Integra 400).
Clinical significant of total cholesterol
Cholesterol is an unsaturated alcohol of the steroid family of compounds and found in blood,
bile, and brain tissue. It is synthesized in many types of tissues, but particularly in the liver and
intestinal wall. It serves as a precursor to bile acids, adrenal and gonadal steroid hormones and
vitamin D (Cox and Garcia-Palmieri, 1990).
Epidemiological studies have shown a positive relationship between total cholesterol
concentrations and mortality from coronary heart disease (Obeid and Herrmenn, 2009).
The series of reactions involved in the assay system are as follows:
Cholesterol oxidase
Cholesterol esters + H2O Cholesterol + fatty acids
Cholesterol Oxidase
Cholesterol + O2 Cholest-4-ene-3-one + H2O2
Peroxidase
2 H2O2 + amino-4-antipyrine + phenol Quinoneimine dye + 4 H2O
The intensity of the color produced is directly proportional to cholesterol concentration. It is
determined by measuring the increase in absorbance at 500 – 550 nm.
The concentration of cholesterol was calculated by using the following formulae:
Absorbance of Sample
Cholesterol Concentration= (Cholesterol standard)*200 mg/dl
(mmol/L) Absorbance of Standard
Clinical significant of triglyceride
Triglyceride (TG) is water insoluble lipids, synthesized in the intestinal mucosa by the
esterification of glycerol and free fatty acids. They represent a concentration source of
metabolic energy.

Materials and methods
32
Triglyceride are transported in the blood as core constituents of all lipoproteins, but the
greatest concentration of these molecule is carried in the TG-rich chylomicrom and very low
density lipoproteins (VLDL) (Rifai et al., 2001).
The triglycerides are determined after enzymatic hydrolysis with lipases. The indicator is a
quinoneimine formed from hydrogen peroxide, 4-aminophenazone and 4-chlorophenol under
the catalytic influence of peroxidase (Young, 2001).
Lipoprotein Lipase (LP)
Triglycerides (TG) Glycerol+ fatty acids
Glycerol Kinase (GK)
Glycerol +ATP Glycerol-3-phosphate (G3P) + ADP
Glycerol Phosphate Oxidase (GPO)
G3P + O2 Dihydroxyacetone phosphate (DAP) + 2 H2O2
Peroxidase (POD)
2 H2O2+ 4-AAP + 4-Chlorophenol Quinoneimine dye + 4 H2O
The concentration of triglycerides was calculated by using the following formulae:
Absorbance of Sample
TG concentration = * 200 (Standard concentration)
Absorbance of Standard
= mg/ml *0.0114 mmol/L.
Clinical significant of HDL-c
High density lipoprotein cholesterol (HDL-c) also known as "good" cholesterol, molecules
consisting of cholesterol and protein that carry cholesterol from cells back to the liver (Obeid
and Herrmenn, 2009).
HDL-c was determined with enzymatic procedure after lipoproteins were precipitate by
phosphotungstate in the presence of magnesium ions. After centrifugation, the HDL cholesterol

Materials and methods
33
in the supernatant is determined by the same technique as the total enzymatic cholesterol, and
the calculation as shown below:
Absorbance of Sample
HDL-c Concentration = (Standard concentration)*200 mg/dL
Absorbance of Standard
Clinical significant of low density lipoprotein cholesterol
Low density lipoprotein cholesterol (LDL-c) particle carry cholesterol from the cell back to
the tissue. LDL-c is known as bad cholesterol because high levels are thought to increase the
risk of heart disease.
LDL-c concentration was obtained by direct calculation according to Friedwald formula:
LDL = total cholesterol - HDL - triglycerides / 5
When the level of TG is greater than 3.4 g / l (3.75 mmol / L), LDL cholesterol cannot be
calculated by this formula, it should be assayed by a direct enzymatic method.
3-3-3 Determination of Aspartate Aminotransferase and Alanine
aminotransferase activities
Aspartate Aminotransferase (AST) and Alanine aminotransferase (ALT) values were
assessed using colorimetric automatic procedures (Auto-analyzer type Integra 400).
Aspartate Aminotransferase
Aspartate Aminotransferase is a cellular enzyme present in many tissues such as heart,
skeletal muscles, kidney, brain, liver, pancreas or erythrocytes. It exists in two isoforms,
cytoplasmic and mitochondrial. The determination of AST activity in serum is used mainly to
assess the liver damage.

Materials and methods
34
Principle:
Aspartate Aminotransferase catalyzes the transfer of an amino group from glutamic acid
to oxaloacetic acid with the formation of α-ketoglutarate and L-aspartate. In the Alera assay,
the reaction mixture contains an excess of malate dehydrogenase. In the presence of reduced
nicotinamide adenine dinucleotide (NADH), the malate dehydrogenase converts oxaloacetic
acid to malic acid with the oxidation of NADH to nicotinamide adenine dinucleotide (NAD).
NADH absorbs strongly at 340 nm, whereas NAD does not. Therefore the rate of conversion of
NADH to NAD can be determined by monitoring the decrease in absorbance bichromatically at
340-647 nm.
Alanine aminotransferase
Alanine aminotransferase is a cytoplasmic enzyme. It is primarily localized in
hepatocytes. It is released into the blood during the cell damage. The determination of ALT
activity in serum is used mainly to assess the liver damage.
Principle:
Alanine aminotransferase in serum converts the L-alanine and α-keto-glutarate in the
reaction to L-glutamate and pyruvate. The pyruvate that is formed reacts with reduced
nicotinamide adenine dinucleotide (NADH) in the presence of lactate dehydrogenase to form
lactic acid and oxidized nicotinamide adenine dinucleotide (NAD). The rate of conversion of
the reduced cofactor to the cofactor can be determined by monitoring the decrease in
absorbance bichromatically at 378 nm- 505 nm.
3-4 Determination of oxidative stress parameters
3-4-1 Tissue homogenate preparation
0,5g of the liver was homogenized in 2ml of TBS (Tris 50 mM, NaCl 150 mM, pH 7.4).
The homogenates were centrifuged at 9000 g for15 min at 4˚C. And the supernatant was used
for determination of reduced glutathione, the catalase activities and protein concentrations.
3-4-2 Glutathione assay
Glutathione (GSH) is a water-soluble tripeptide (γ-glutamyl-cysteinylglycine) produced
naturally by the liver. Due to the thiol function of cysteine, glutathione is an important
compound in maintaining the redox balance of the cell. It maintains in the proper redox state

Materials and methods
35
OD * 1*1.525
GSH (nmol/mg of protein) =
13100*0.8*0.5.mg protein
the thiol groups of soluble and structural proteins, and participates in the detoxification of
hydroxyperoxides. In addition to detoxification, GSH plays a role in other cellular reactions,
including, the glyoxalase system, reduction of ribonucleotides to deoxyribonucleotides,
regulation of protein and gene expression via thiol disulfide exchange reactions (Townsend et
al., 2003).
The concentrations of the GSH are proportioned by the method of Weckbecker and Cory as
shown below, The spectrophotometric reader assay method for GSH involves oxidation of
GSH by the sulfhydryl reagent 5,5′-dithio-bis2-nitrobenzoic acid (DTNB) to form the yellow
derivative 5′-thio-2-nitrobenzoic acid (TNB),which is measurable at 412 nm.
Liver homogenate sample (0.8ml) was deproteinized with (0.2ml) of 5-sulfosalicylic acid
solution (0.25%) and was allowed stand on ice for 10 min. Following centrifugation at 1000
tours/mn) during 5minutes to remove the precipitated protein. (0.5 ml) of supernatant was
mixed with 1 ml Tris/EDTA buffer (pH 9.6) and (0.025 ml) of DTNB-reagent (0.01M 5,5'-
dithiobis-2-nitrobenzoic acid) and left at room temperature for 5 min. Then the absorption was
measured at 412 nm using a spectrophotometer by comparing to the blank reaction.
Glutathione concentration was obtained by direct calculation of the following formulae:
OD: optical density
1: total volume of solutions in the deproteinisation (0.8ml homogenate+ 0.2ml 5-
sulfosalicylic acid).
1.525: total volume of the solutions used in the assay of GSH (0.5ml supernatant+ 1 ml
Tris/EDTA+ 0.025 ml DTNB).
13100: absorbance coefficient at Groupment—SH to 412nm.
0.8: volume of homogenat sample.
0.5: volume of supernatant.

Materials and methods
36
Protein determination
Protein concentration was measured by the method of Bradford (1976), using bovine serum
albumin as standard. The procedure is based on the formation of a blue complex between the
comaissie bruillant blue G-250 dye, and proteins in solution. The amount of absorption is
proportional to the protein present.
Liver homogenate sample 0.1ml was mixed with 5ml Bradford reagent and was allowed
stand for 5min. Then the absorbance was measured at 595 nm using a spectrophotometer by
comparing to the blank reaction.
The protein concentration of a test sample is determined by comparison to that of a standard
series of bovine serum albumin to reproducibly exhibit a linear absorbance profile in this assay
(Figure 01 annex).
3-4-3 Catalase activity
Catalase is a common enzyme found in nearly all living organisms exposed to oxygen. It
catalyzes the decomposition of hydrogen peroxide (H2O2) to water and oxygen. Catalase is a
tetramer of four polypeptide, It contains four porphyrin heme (iron) groups that allow the
enzyme to react with the hydrogen peroxide. It was estimated in the liver homogenate in a UV
spectrophotometer as described by Aebi (1984). The specific activity of catalase has been
expressed as mmol of H2O2 consumed/min/ mg protein. The difference in absorbance at 240
nm per unit time is a measure of catalase activity.
The reaction is believed to occur in two stages:
Catalase-(Fe III) + H2O2 [Catalase-H 2O-(Fe V)] + H2O
[Catalase-H 2O-(Fe V)]+ H 2O2 Catalase-(Fe III) + H2O + O2
2 H2O2 2 H2O + O2
Table 04: the concentrations and amounts of reagents needed for the dosage of catalase
activity.
Sq: supernatant quantity
Reagents Sample (μl) blank (μl)
Phosphate buffer (100Mm, PH7.5) 790 800
H2O2 (500Mm) 200 200
Sq (1 to 1.5 mg prt/ml) 10 0

Materials and methods
37
The activity of catalase was estimated by the decrease of absorbance at 240 nm for 1 min
(15 and 60 seconds ) as a consequence of H2O2 consumption.
Catalase activity was obtained by direct calculation of the following formulae:
Catalase activity (mmol H2O2/min/ mg prot) =
ΔDO
ε ×L × χ × Fd
ε : extinction coefficient (= 0.043 mM-1.cm -1).
L : The length of the cuvette used (1 cm).
χ : protein quantity mg/ml.
Fd : 0.02 (dilution factor of the H2O2 in the buffer).
λ : 240 nm.
3-5 Histological analysis
After the blood samples collection, the animals were sacrificed, and samples for light
microscopic investigations were obtained from aorta, heart and liver. For histological
investigations the aorta was divided into 4 sections (arch, thoracic, abdominal, and iliac).
The samples were rinsed of all adherent tissues with saline solution (0.9%).
Then, they are kept in small containers filled with diluted formol 10%. Liver, heart and the
different parts of the aorta were included in Bouin solution for 5 min (until colored), and the
deshydration was performed through a series of ethanol solution (50 %, 70 %, 96 %), each step
was placed for approximately 30 min (3×30 min=1h 30min).
The tissues were then kept in small containers filled with butanol for 3 days. After that they
were cleared in Xylene for 10 min with two exchanges.
For the next step, the organs were immersed in paraffin at 60 °C for 1h and 30 min at
three exchanges. The sectioning was performed with a microtome (Leica RM 2135, laboratory
EL-YASSEMINE, ANNABA).
Paraffin slices, 5μm thick were stained following the heamatoxylin eosin staining
protocol (Appendix).

Materials and methods
38
4- Biofilm formation and quantification
4-1 Strains and culture conditions
The following strains were used in this study:
Staphylococcus haemoliticus clinical isolate NC1 ;
Streptococcus intermedius DSMZ 20573 (German Collections of
Microorganisms and Cell Cultures);
Streptococcus anginosus, clinical isolate NC10 ;
Streptococcus mutans CIP103220 (Collection Institut Pasteur) and
Streptococcus uberis human clinical isolate NC6.
These strains were identified by 16S rRNA gene sequencing, using the Sanger capillary
sequencing procedure with ABI 310 apparatus (Applied Byosystem). The bacteria prior the
use were stored at – 80°C in a tube contained the proper medium broth (Microbiol, UTA,
Cagliari, Italy) with 20% glycerol.
4-2 Crystal Violet Biofilm formation screening assay
Preparation of inoculum:
The bacteria (Streptococcus spp.) were grown in anaerobic Schaedler agar (Microbiol,
UTA, Cagliari, Italy) at 37°C for 24 hours with a CO2 concentration of 5% and in Muller-
Hinton agar (Microbiol, UTA, Cagliari, Italy) for aerobic bacteria (S. haemolyticus).
After the incubation, 5 ml of SH Broth was inoculated with the following bacterial
species until a final concentration of 106cell/ml
for each bacterial species using a
spectrophotometer at 620 nm (DMS100s, Varian, NH, USA).
Protocol for quantification of biofilm on microplate
For an in vitro biofilm evaluation we used the protocol described by (Merritt et al.,
2005).
Briefly, each antibacterial combination was suspended in a 96 well microplate, where
100 L of SH Broth and 100 L of the Argan oil were added in the first well. From the first
well, 12 serial dilutions were done (from 100 to 0,04 g/ml). Next, 100 L of microbial

Materials and methods
39
suspension was added into each well. However, positive control wells contained SH Broth and
microbial suspension, and negative control wells contained SH Broth and Argan oil only.
The experiment was performed in duplicate.
Then, The plate was agitated on microplate shaker and incubated for 37°C for 48h
aerobically (S. haemolyticus), or in 5% CO2 (Streptococcus spp.) to permit biofilm formation.
After this, the plate samples were gently washed three times with Phosphate-buffered
saline (PBSGibco®) to eliminate planktonic cells and the adhering cells in the biofilm were
stained with 100 µl of 0.1 % v/v of crystal violet solution (Microbial, Uta , Cagliari) for 10
minutes at 25°C. After three washes with PBS solution, as described previously, 200 µl of 30%
v/v acetic acid were added in every well to solubilize the dye from the bacterial biomass. The
biofilm amount was measured with a plate reader spectrophotometer (SLT-Spectra II, SLT
Instruments, Germany) at 620 nm.
In these conditions, the minimum biofilm inhibition concentration (MBIC) is defined as
the lowest concentration of an antimicrobial required to inhibit the formation of novel biofilms,
and it was determined by observing a reduction of 90% in color intensity at 620 nm ABS, as
compared to positive controls in the microplate wells.
The absorbance of biofilm formation was calculated using the ratio between the values
of wells with and without Argan oil.
The percentage of biofilm inhibition was calculated by the formula:
C= mean OD of positive control,
B = mean OD of negative control,
T= mean OD of test wells.

Materials and methods
40
5- Statistical analysis
Values obtained were expressed as mean ± SEM and subjected to statistical analysis
using one-way analysis of variance (ANOVA), followed by Tukey’s post-hoc test. The
programme used for the analysis was SPSS (version 20). P < 0.05 values were considered as
significant different.
The comparison is considered, according to the probability (P) as:
No significant if P >0.05.
Significant (*) if P <0.05.
Highly significant (**) if P <0.01.
Very highly significant (***) if P <0.001.

Results and
Discussion

Chapter 1
Effect of L-methionine and A. spinosa
powdered seeds on body weight, Hcy levels,
lipid profile, liver enzyme activities and
antioxidants markers in mice

Results and Discussion
41
1- Body weight
In the present study, we examined the effects of of L-methionine on the weight of mice.
No significant difference was found in body weight among the four groups (F), (M),
(MP) and (P) during the experimental period, indicating that (500mg/ Kg) of L-Methionine
or/and (150 mg/ Kg) of the powdered seeds of A. spinosa supplementations do not affected
significantly mice growth P> 0.05 (Figure 13).
Figure 12. Effect of L-methionine intake on mice weight during 21 days.
F: Control group received flour at 0.5mg/mice; M: group received methionine at dose 500mg/kg; MP: group
received methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group received
Argania spinosa powdered seeds at dose 150mg/kg.
2- Effect of treatments on lipid profile in mice
2-1 Triglycerides
As shown in the (Figure 13), the concentrations of the TG were in: the (F) group (0.61
±0.05 g/L), (M) group (0.81 ± 0.06 g/L), (MP) group (0.57 ± 0.05 g/L) and (P) group (0.55±
0.04
15,00
20,00
25,00
30,00
35,00
40,00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
wei
gh
t (g
)
F
M
PM
P
Days

Results and Discussion
42
g/L) .Our data indicated that there is a high significant difference between groups (P =
0.002).
The Tukey test revealed significant difference between the groups (F and M) P= 0.029,
and a high significant difference between the groups (M and MP) (P=0.006) and the groups
(M and P) (P= 0.003).
Figure 13. Interaction of L-methionine and A. spinosa seeds on the triglycerides in mice
during 21 days of treatment.
Values are shown as mean ±SEM (n = 5); *p<0.05 and **p<0.01 .
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
2-2 Total cholesterol
The concentrations of the T-CHO were in: the (F) group (1.09 ±0.05 g/L), (M) group
(1.29 ± 0.08 g/L), (MP) group (1.04 ± 0.04 g/L) and (P) group (0.95± 0.11 g/L). Our results
shown that there is a significant difference among the four groups (P = 0.017).
The Tukey test revealed significant difference between the groups (M and P) P= 0.012.
(Figure 14)
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
F M MP P
Tri
gly
ceri
des
(g
/L)
* ** **

Results and Discussion
43
Figure 14. Interaction of L-methionine and A. spinosa seeds on the T-CHO in mice during 21
days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
2-3 HDL-c
The present data showed that there is a significant difference in the concentrations of the
HDL-c among the four groups: (F) group (0.88 ±0.02 g/L), (M) group (0.70± 0.07 g/L), (MP)
group (0.84 ± 0.03 g/L) and (P) group (0.72± 0.03 g/L) (P = 0.017).
The Tukey test revealed significant difference between the groups (F and M) (P= 0.033).
(Figure 15)
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
F M MP P
Tota
l Ch
ole
ste
rol (
g/L)
*

Results and Discussion
44
Figure15. Interaction of L-methionine and A. spinosa seeds on the HDL-c in mice during 21
days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L- methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150 mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
2-4 LDL-c
The Figure 16 demonstrated that the concentrations of the LDL-c were in: the (F)
group (0.09 ±0.02 g/L), (M) group (0.42 ± 0.12 g/L), (MP) group (0.08 ± 0.02 g/L), (P) group
(0.06± 0.6 g/L). Our results showed that there is a high significant difference among the four
groups (P = 0.002).
The Tukey test revealed a high significant difference between the (F and M) P= 0.008,
the groups (M and MP) P=0.005 and the groups (M and P) P= 0.003.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
F M MP P
HD
L-c
(g/L
)
*

Results and Discussion
45
Figure 16. Interaction of L-methionine and A. spinosa seeds on the LDL-c in mice during 21
days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05 and **p<0.01.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
3- Effect of treatments on Homocysteine levels
After 21 days of experiments, results showed that there is a very significant difference
(p=0.007) between 4 groups : plasma t-Hcy of the (M) group achieved higher levels, and the
average was 10.04±0.83 μmol/l, significantly higher than that of the control group F
(6,84±0.51 μmol/l) and control positive group (P) (7.1±0.88 μmol/l). However, in the
combined treatments of A. spinosa crude extract with L-methionine, the Hcy concentration
was (7.26±0.46 μmol/l), showed a significant decline in plasma t-Hcy, demonstrating that
A.spinosa crude extract appears to be effective in preventing the increase of t-Hcy levels.
The Tukey test revealed a high significant difference between the groups (F and M) (P=
0.01), the groups (M and P) (P= 0.01) and a significant difference between the groups (M and
MP) (P=0.028). (Figure 17)
0
0,1
0,2
0,3
0,4
0,5
0,6
F M MP P
LD
L-c
(g
/L)
** ** **

Results and Discussion
46
Figure 17. Interaction of L-methionine and A. spinosa seeds on the Homocysteine levels in
mice during 21 days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05 and **p<0.01.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
4- Effect of treatment on liver enzymes activities
4-1 Aspartate Aminotransferase
The concentrations of the AST were in: the (F) group (129 ±65 UI/L), (M) group
(386.14 ± 53.13 UI/L), (MP) group (308.37 ± 55.5 UI/L) and (P) group (197.18± 35.72 UI/L).
Our data shown that there is a very high significant difference among the four groups (P =
0.001).
0
2
4
6
8
10
12
F M MP P
Hcy
(µ
mo
lL)
** * **

Results and Discussion
47
The Tukey test revealed a very high significant difference between the groups (F and
M) (P= 0.001), and a significant difference between the groups (F and MP) (P=0.02) and a
high significant difference between the groups (M and P) (P= 0.01). (Figure 18)
Figure 18. Interaction of L-methionine and A. spinosa seeds on the AST in mice during 21
days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05, **p<0.01 and ***p<0.001.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
4-2 Alanine aminotransferase
The concentrations of the ALT showed that there is a difference among the four groups
but not significantly (P> 0.05): in the (F) group (38.89 ±4.01 UI/L), (M) group (49.04.14 ±
5.31 UI/L), (MP) group (45.14 ± 15.14 UI/L) and (P) group (35.29± 10.34 UI/L).
ALT levels were increased in (M) group and decreased in (MP) group but not significantly
compared to the other groups (Figure 19).
0
50
100
150
200
250
300
350
400
450
500
F M MP P
AS
T (
UI/
L)
*** ** *

Results and Discussion
48
Figure 19. Interaction of L-methionine and A.spinosa seeds on the ALT in mice during 21
days of treatment.
Values are shown as mean ±SEM (n = 7).
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
5- Effect of treatment on antioxidants markers
5-1 Reduced Gluthatione
The concentrations of the reduced glutathione (GSH) were in: the (F) group (8.03±0.55
nmol/mg protein), (M) group (4.48±0.55 nmol/mg protein), (MP) group (6.1±0.5 nmol/mg
protein) and (P) group (7.36±1.28 nmol/mg protein). Our data showed that there is a high
significant difference among the four groups (P = 0.01).
The Tukey test revealed a high significant difference between the groups (F and M) (P=
0.01) and a significant difference between the groups (M and P) (P=0.048) (Figure 20).
0
10
20
30
40
50
60
F M MP P
AL
T (
UI/
L)

Results and Discussion
49
Figure 20. Interaction of L-methionine and A.spinosa seeds on the reduced glutathione
in mice during 21 days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05 and **p<0.01.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
5-2 Catalase activity
After 3 weeks of treatment, our results showed that there is a significant difference
(p=0.02) among the four 4 groups: Hepatic catalase activity was in: the (F) group (61.37
±6.39 mmol/mg protein), (M) group (45.82±5.83 mmol/mg protein), (MP) group (62.26±3.32
mmol/mg protein) and (P) group (52.16±3.19 mmol/mg protein).
The Tukey test revealed a significant difference between the groups (F and M) (P=
0.03) and the groups (M and MP) (P= 0.02) (Figure 21).
0,00
1,00
2,00
3,00
4,00
5,00
6,00
7,00
8,00
9,00
10,00
F M MP P
GSH
(n
mo
l/m
g p
rote
in)
**
*

Results and Discussion
50
Figure 21. Interaction of L-methionine and A.spinosa seeds on the catalase activity in mice
during 21 days of treatment.
Values are shown as mean ±SEM (n = 7); *p<0.05.
F: Control group received flour at 0.5mg/mice; M: group received L-methionine at dose 500mg/kg; MP: group
received L-methionine at dose 500mg/kg + Argania spinosa powdered seeds at dose 150mg/kg; P: group
received Argania spinosa powdered seeds at dose 150mg/kg.
0
10
20
30
40
50
60
70
80
F M MP P
Ca
tala
se a
ctiv
ity
(mm
ol/
mg
pro
tein
)
* *

Chapter 2
Effect of L-methionine and A. spinosa
powdered seeds on histology of aorta, heart
and liver

Results and Discussion
51
The administration of L-methionine (500mg/Kg/animal) during 3 weeks appears to alter
the aorta, heart and liver structures compared to the control group.
1- Histological study of the heart
Changes observed by light microscopy were more apparent in the sections of (M)
group than in (F) group-stained, paraffin-embedded sections. These changes consisted of
presence of lysis, and architectural changes of cardiomyocytes (Figure22: (B) and (C). There
were no apparent differences in severity of lesions among samples obtained from group (MP)
and (P) (Figure22: (D) and (E)).
2- Histological study of the aorta
In the group (M), the aortic intima showed degeneration and desquamation of
endothelial cells with fenestration, it is observed in the media lysis, formation of foam cells
laden with small lipid droplets and oval nuclei, disappearance of elastics fibers especially in
the arch aorta. (Figure 23: (B) and (C); Figure 24: (B); Figure 25: (B), (C), (D) and (E)).
However, in the control group (F), the aortic sections have intact endothelium and
spindle shaped mediocytes nuclei. Also we have observed intact aorta in the group (MP)
treated with L-methionine and A. spinosa (Figure 23: (D); Figure 24: (C); Figure 25: (F)) and
in the group (P) treated with A. spinosa only (Figure 23: (E); Figure 24: (D); Figure 25: (G)).
Noting that all these microscopic changes were observed in the histological sections of
(arch, abdominal and iliac) aorta. For the thoracic aorta sections in experimental groups (F,
M, MP and P), any change was observed.
3- Histological study of the liver
The microscopic analysis of the liver of the control group (F), showed the
parenchymal cells which are hepatocytes. These polygonal cells are joined to one another
anastomosing plates, with borders that face either the sinusoid or adjacent hepatocytes (Figure
26: (A)).
For the group (M), our data showed various pathological alterations in liver of mice
induced by the oral L-methionine administration. These alterations were marked by

Results and Discussion
52
destruction of membrane cells, hepatocellular ballooning, hypertrophy of some hepatocyte
nuclei, cytoplasmic vacuolization, and macrovesicular hepatic steatosis with a large lipid
vacuole filling the hepatocyte cytoplasm and around the blood vessels. (Figure 26: (B), (C),
(D), (E) and (F)).
The liver of (P) group showed a normal structure (Figure 26: (H). Thus, in
combination group (MP), were A. spinosa powdered seeds were administered with 500
mg/Kg of L-methionine showed reparative changes (Figure 26: (G)).

Results and Discussion
35
Figure 22. Histological sections of heart tissue in experimental groups (F, M, MP and P)
X 100 X400
X 100 X 100 X 100

Results and Discussion
54
(A). Histological sections of the cardiac muscle. Control’s Hematoxylin eosin staining (x100).
The heart histology of the group (F) was intact, with the presence of cardiac muscle fibers and
their nuclei.
(B) and (C). Histological sections of the cardiac muscle. 21 days of oral L-methionine
application (500 mg/kg/day). Hematoxylin eosin staining (Bx400, Cx100).
We observed in group (M), lysis and architectural changes of cardiomyocytes.
(D). Histological section of the cardiac muscle. 21 days of oral L-methionine (500 mg/kg/day)
+ A. spinosa powdered seeds (150mg/kg/day) application. Hematoxylin eosin staining (x100).
In the histological sections of the cardiac muscle of group (PM), no lysis was observed.
(E). Histological section of the cardiac muscle. Positive Control’s Hematoxylin eosin
staining (x100).
No change was observed.
ACC. Architectural Changes of Cardiomyocytes, CMF. Cardiac Muscle Fibers, MCN. Muscle Cell Nuclei,
L. Lysis.

Results and Discussion
55
Figure 23. Histological sections of the arch aorta in experimental groups (F, M, MP and P)
X 100
X 100
X 400 X 100 X 100

Results and Discussion
56
(A). Histological sections of the arch aorta. Control’s Hematoxylin eosin staining (x100).
The endothelium of the arch aorta of the group (F) was intact.
(B) and (C). Histological sections of the arch aorta. 21 days of oral methionine application
(500 mg/kg/day). Hematoxylin eosin staining (B x 100, C x400).
We observed in group (M), in the media the presence of lipids droplets, and the disappearance
of elastics fibers. We noted also appearance of the oval nuclei.
(D). Histological section of the arch aorta. 21 days of oral methionine (500 mg/kg/day) +
A.spinosa powdered seeds (150mg/kg/day) application. Hematoxylin eosin staining (x100).
In the histological sections of the arch aorta of group (PM), no change was observed.
(E). Histological section of the arch aorta. Positive Control’s Hematoxylin eosin staining
(x100).
No change was observed.
FC. Foam Cells, FN. Fibroblast Nuclei, IEND. Intact Endothelium, LD. Lipid Droplets, Lu. Lumen, ON.
Oval Nuclei, SN. Spindle Nuclei.

Results and Discussion
57
Figure 24. Histological sections of the abdominal aorta in experimental groups (F, M, MP and P)
X 100 X 100 X 100 X 100

Results and Discussion
58
(A). Histological sections of the abdominal aorta. Control’s Hematoxylin eosin staining
(x100).
This photograph showed the intact endothelium of the abdominal aorta of the group (F).
(B). Histological section of the abdominal aorta. 21 days of oral methionine application (500
mg/kg/day). Hematoxylin eosin staining (x100).
The aortic intima of group (M), showed endolysis.
(C). Histological section of the abdominal aorta. 21 days of oral methionine (500
mg/kg/day) + A. spinosa powdered seeds (150mg/kg/day) application. Hematoxylin eosin
staining (x100).
In the histological section of the abdominal aorta of group (PM), no lysis was observed.
(D). Histological section of the abdominal aorta. Positive Control’s Hematoxylin eosin
staining (x100).
In the positive control group (P), the aortic sections have intact endothelium and spindle
shaped mediocytes nuclei.
END. Endolysis, FN. Fibroblast Nuclei, IEND. Intact Endothelium , LU. Lumen, SN. Spindle Nuclei.

Results and Discussion
Figure 25. Histological sections of the iliac aorta in experimental groups (F, M, MP and P)
59
X 100 X 100 X 400
X 400 X 400 X 100 X 100

Results and Discussion
60
(A). Histological sections of the iliac aorta. Control’s Hematoxylin eosin staining (x100).
The histology of the iliac aorta of the group (F) was intact.
(B), (C), (D) and (E). Histological section of the iliac aorta. 21 days of oral L-methionine
application (500 mg/kg/day). Hematoxylin eosin staining (B x 100; D, E, Cx400).
Changes observed by light microscopy in the group (M), were degeneration and desquamation
of endothelial cells, we also observed in the media lysis, fenestration, formation of foam cells,
lipids droplets and oval nuclei.
(F). Histological section of the iliac aorta. 21 days of oral L-methionine (500 mg/kg/day) +
A. spinosa powdered seeds (150mg/kg/day) application. Hematoxylin eosin staining (x100).
In the histological section of the iliac aorta of group (PM), no change was observed.
(G). Histological section of the iliac aorta. Positive Control’s Hematoxylin eosin staining
(x100).
No change was observed.
D. Desquamation, END. Endolysis, F. Fenestration, FC. Foam Cells, FN. Fibroblast Nuclei, IEND. Intact
Endothelium , LU. Lumen, ON. Oval Nuclei, SN. Spindle Nuclei.

Results and Discussion
61
Figure 26. Histological sections of liver tissue in experimental groups (F, M, MP and P)
X 100 X 100 X 400 X 400
X 400 X 400 X 100 X 100

Results and Discussion
62
(A). Histological section of the liver. Control’s Hematoxylin eosin staining (x100).
The liver of control group showed a normal structure.
(B), (C), (D) (E), and (F). Histological sections of the liver. 21 days of oral L-methionine
application (500 mg/kg/day). Hematoxylin eosin staining ((B) x(100); (C), (D) (E), and (F)
x400).
We observed in the liver of the group (M), destruction of membrane cells, hepatocellular
ballooning, hypertrophy of some hepatocyte nuclei, cytoplasmic vacuolization, and
macrovesicular hepatic steatosis with a large lipid vacuole filling the hepatocyte cytoplasm
and around the blood vessels.
(G). Histological section of the liver. 21 days of oral L-methionine (500 mg/kg/day) +
A.spinosa powdered seeds (150mg/kg/day) application, Hematoxylin eosin staining (x100).
The histological section of the liver of group (PM), showed reparative changes.
(H). Histological section of the liver. Positive Control’s Hematoxylin eosin staining (x100).
Any change was observed in this histological section.
CV: Centro Lobular Vein, Ds. Destruction of membrane cells, H: Hepatocyte, S: Sinusoid. Hb. Hepatocellular
Ballooning, Hy. Hypertrophy of Hepatocyte Nuclei, ICF. Inflammatory Cell Foci, S. Sinusoid, V. Vacuole.

Chapter 3
Anti-biofilm formation of Argan oil
“In vitro” Study

Results and Discussion
63
Results of the biofilm assay are shown in (Figures 27, 28, 29,30, 31).
These results showed that there was a trend of increasing inhibition of S. intermedius,
St haemolyticus, S. mutans, S. anginosus and S. uberis biofilm formation as the Argan oil was
getting more concentrated.
As known that the minimum biofilm inhibition concentration (MBIC) is defined as
the lowest concentration of an antimicrobial required to inhibit the formation of novel
biofilms, and it was determined by observing a reduction of 90% in color intensity at 620 nm,
as compared to positive controls in the microplate wells.
Argan oil had a MBIC value of 100 g/mL when tested against S. intermedius, St
haemolyticus, S. mutans, and S. anginosus. Of all the 12 concentartions that were investigated
for their effects on growth, the first concentration (first dilution) was the most effective in the
inhibition of growth of these mentioned bacteria (Table 05, Figure 32), and produced a
percent reduction of 93.81 %, 100 %, 100 %, 100 %, 78.78 % for. S. intermedius, St
haemolyticus, S. mutans, S. anginosus and S. uberis respectively. However, the lesser
concentrations did not significantly inhibit biofilm formation.
The acceptable concentration of Argan oil (100 µg/mL) indicated the highest
antibacterial effect on biofilms of S. intermedius, St haemolyticus, S. mutans, S. anginosus S.
but not S. uberis (78.78 % lower than 90%).
Our study leads to the conclusion that Argan oil is able to efficiently reduce biofilms
formation of (Streptococcus mutans, Streptococcus anginosus, Streptococcus intermedius,
and staphylococcus haemolyticus) strains.

Results and Discussion
64
Figure 27. Inhibitory effect of Argan oil on S. intermedius biofilm formation.
Figure 28. Inhibitory effect of Argan oil on S. haemolyticus biofilm formation.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
100 50 25 12,5 6,25 3,12 1,56 0,78 0,39 0,19 0,09 0,048
Bio
film
fo
rma
tio
n a
t 6
20
nm
Concentrations of dilutions (µg/mL)
S intermedius +
argan oil
S intermedius
0,0
0,5
1,0
1,5
2,0
2,5
3,0
100 50 25 12,5 6,25 3,12 1,56 0,78 0,39 0,19 0,09 0,048
Bio
film
fo
rma
tio
n a
t 6
20
nm
concentrations of dilutions (µg/mL)
S. haemolyticus +
argan oil
S.haemolyticus

Results and Discussion
65
Figure 29. Inhibitory effect of Argan oil on S. mutans biofilm formation.
Figure 30. Inhibitory effect of Argan oil on S. anginosus biofilm formation.
0,00
0,50
1,00
1,50
2,00
2,50
3,00
3,50
100 50 25 12,5 6,25 3,12 1,56 0,78 0,39 0,19 0,09 0,048
Bio
film
fo
rmat
ion
at
62
0 n
m
concentrations of dilutions (µg/mL)
S.mutans + argan
oil
S. mutans
0,0
0,5
1,0
1,5
2,0
2,5
100 50 25 12,5 6,25 3,12 1,56 0,78 0,39 0,19 0,09
Bio
film
fo
rma
tio
n a
t 6
20
nm
concentrations of dilutions (µg/mL)
S. anginosus+ argan oil
S.anginosus
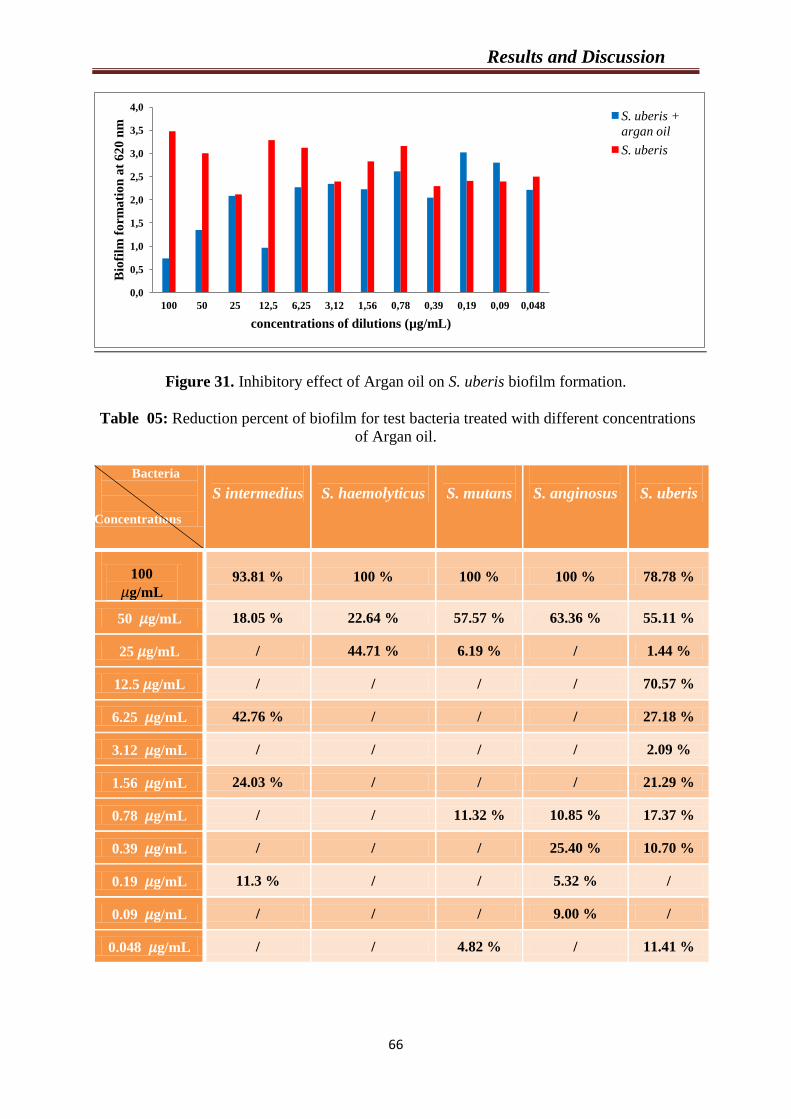
Results and Discussion
66
Figure 31. Inhibitory effect of Argan oil on S. uberis biofilm formation.
Table 05: Reduction percent of biofilm for test bacteria treated with different concentrations
of Argan oil.
Bacteria
Concentrations
S intermedius
S. haemolyticus
S. mutans
S. anginosus
S. uberis
100
g/mL
93.81 % 100 % 100 % 100 % 78.78 %
50 g/mL 18.05 % 22.64 % 57.57 % 63.36 % 55.11 %
25 g/mL / 44.71 % 6.19 % / 1.44 %
12.5 g/mL / / / / 70.57 %
6.25 g/mL 42.76 % / / / 27.18 %
3.12 g/mL / / / / 2.09 %
1.56 g/mL 24.03 % / / / 21.29 %
0.78 g/mL / / 11.32 % 10.85 % 17.37 %
0.39 g/mL / / / 25.40 % 10.70 %
0.19 g/mL 11.3 % / / 5.32 % /
0.09 g/mL / / / 9.00 % /
0.048 g/mL / / 4.82 % / 11.41 %
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
100 50 25 12,5 6,25 3,12 1,56 0,78 0,39 0,19 0,09 0,048
Bio
film
fo
rma
tio
n a
t 6
20
nm
concentrations of dilutions (µg/mL)
S. uberis +
argan oil
S. uberis

Results and Discussion
67
Figure 32. Effect of the first dilution (100 g/mL) concentration of Argan oil on
biofilm inhibition.
0%
20%
40%
60%
80%
100%
93,81% 100% 100% 100%
78,78% R
edu
ctio
n p
erce
nt%

Results and Discussion
68
DISSCUSION
This research investigated the relationship between dietary intake of L-methionine and
total plasma Hcy concentration (t-Hcy). This (t-Hcy) is a biomarker of the homocysteine-
methionine cycle and dysfunction in this cycle can lead to several cardiovascular diseases. In
other hand, we evaluate the protective and the preventive effect of the powdered seeds of A.
spinosa against HHcy, hyperlipidemia, oxidant status and damages in the aorta, heart and
liver induced by high methionine intake in mice. Also, this study investigated the antibacterial
effect of Argan oil on biofilm formation.
1- Body weight
The first experiment evaluates the effect of L-methionine on the weight. The results
showed that there is no significant difference in weight of mice among the four groups, which
means that the body weight of mice was not affected by L-methionine intake.
Our results are in agreement with (Lamda, 2000) who indicated a stable weight despite
the applied treatment (L-methionine and cholesterol). Another work of (Zarrouk et al., 2010)
who reported that a similar evolution of weight between animals subjected to methionine and
the control group during the first month of the experiment (administration of 70 mg/kg during
6 months in an athero-sensitive model (Psammomys obesus)). However, the study of (Zerizer,
2006) reported an increased in the weight of rats treated with 200 mg/kg of L-methionine for
21 days. Also, the study of (Boudebouz, 2013; Sakhri, 2014) showed an increase in the weight
of mice treated with 400 mg/kg of L-methionine for 3 weeks. However, the work of
(Benmebarek, 2013) indicated a significant decrease in the weight of mice treated with
200mg/kg of L-methionine for the same period. Given the conflicting results, we cannot
conclude that there is a relationship between HHcy and the weight of mice.
2- Effect of treatment on Hcy levels, lipid profile, liver enzyme activities
and antioxidants markers in mice
The current study has shown that high L-methionine diet in mice clearly caused a
significant increase in plasma t-Hcy levels, T-CHO, TG , AST activities, and significant
decrease in HDL-c, reduced glutathione, and catalase activity in liver.

Results and Discussion
69
Hyperhomocysteinemia results from deficiency of one or more vitamin cofactors,
genetic conditions as reduced activity of enzymes involved in Hcy metabolism, such as CβS
deficiency, deficient activity of methionine adenosyl transferases I and III (MAT I/III), or
non-genetic conditions included liver disease, or high dietary methionine intake (Mudd, 2011;
Hoffman, 2011).
Methionine feeding has often been used to elevate serum and tissue Hcy levels to
study the pathogenesis of HHcys-related metabolic disturbances (Yalçınkaya et al., 2009).
In the present study, only the higher L-methionine dose (500mg/kg) for 21 days of the
experiment, was sufficient to induce HHcy in mice notable by a highly significant increase of
plasma Hcy compared to the normal group; suggesting a detrimental effect of excess dietary
methionine, which may have a practical significance in humans consuming large amounts of
foods high in methionine such as animal proteins (Bhandari et al., 2011).
Acquired results are similar to several studies which have shown that methionine
enriched-diet induced a significant increase of plasma t-Hcy (Mudd, 2011; Viggianoa et al.,
2012; Yamada et al., 2012; Kirac et al., 2013).
This potential effect of high L-methionine diet could possibly be due to up-regulation
in the enzymes that metabolize Hcy, for example cystathione β-synthase. It is clear that tissue
concentration of Hcy is maintained at low levels by regulating production and efficient
removal of this thiol (Stipanuk, 2004).
When methionine is in excess, Hcy is directed to the transulfuration pathway. In this
way, Hcy is converted to cystathionine and then cysteine by the pyridoxal 5-phosphate, the
physiological active coenzyme form of vitamin (B6) dependent enzyme. Cysteine is the rate-
limiting component in the synthesis of glutathione.
Studies have shown that high Hcy concentration could cause oxidative damage to
cells, and cysteine may act as a pro-oxidant agent causing the overproduction of free radicals
and hydrogen peroxide, and may further lead to gene mutation and subsequent development
of cancer (Hogg, 1999; Stipanuk, 2004) and cardiovascular disease.

Results and Discussion
70
Actually, Hcy can be harmful to cells because it evokes oxidative stress through the
production of ROS, binds to nitric oxide (NO) and produces homocysteinylated proteins, or
led to the accumulation of its precursor, S-adenosyl homocysteine, a potent inhibitor of
biological transmethylations (Kamal and Thanaa, 2009).
In this context, the study of (Heydrick et al., 2004) indicated that high Hcy
concentrations increased formation of ROS and lipid oxidation in endothelial cells but not
vascular smooth muscle cells. Indeed, Hcy induced expression of iNOS and decreased eNOS
expression, which led to a decreased NO bioavailability (Kamal and Thanaa, 2009).
Also, other studies have demonstrated the involvement of Hcy actions linked to
oxidative stress (Streck et al., 2003; Matté et al., 2009). It was previously shown that acute
Hcy administration decreased TRAP,Na+, K+-ATPase and catalase (CAT) activities by 20%,
60% and15%, respectively in hippocampus of rats (Wyse et al., 2002). In addition, it was
recently shown that chronic Hcy administration increased DNA damage and disrupted
antioxidant defenses (enzymatic and non-enzymatic) in parietal cortex and blood plasma
(Matté et al., 2009).
Meanwhile, we found that the diet supplemented with the powdered seeds of A.
spinosa was effective in prevention against HHcy in mice exposed to a L-methionine enriched
diet, indicating that this medicinal plant has the potential to reduce t-Hcy levels in vivo. Some
evidences showed that lowering Hcy by nutrition interventions might offer preventive or
therapeutic benefits against cardio- and cerebrovascular diseases, although controversy still
exists (Spence, 2007; Abraham and Cho, 2010). Several dietary factors, including folate,
vitamins B6 and B12, and betaine have been demonstrated to be effective in decreasing
plasma Hcy level (Lin et al., 2006; Spence, 2007).
Our results are in agreement with (Hamelet et al., 2007; Yalcınkaya et al., 2009 ;
Meng et al., 2013) who reported that quercetin, catechin and taurine supplementation are
effectives in attenuating the increase of the serum Hcy level as induced by a Methionine
enriched diet in rats and mice respectively.
The protective effect of Argan oil is probably due to its high contents of antioxidants,
particularly polyphenols, tocopherols and sterols, which are known as powerful antioxidants
(Masella et al., 2001).

Results and Discussion
71
As found in the present investigation, mice fed with high Methionine enriched diet
have shown an increase in serum T-CHO, LDL-C, and TG concentrations; however, HDL-C
values were lower.
An association between hyperlipidemia and HHCY has been suggested. Results
obtained are in agreement with the previous experimental studies of (Lee et al., 2002; Obeid
and Herrmenn, 2009; Tselmin et al., 2013; Boudebouz, 2013, Sakhri, 2014). However, it’s
differed from studies of (Zerizer, 2006 and Zerizer et al., 2008) who observed no relationship
between HHcy and lipid status.
Both Hcy and lipids are toxic in vascular cells and hepatocytes, which could indicate
interactions between the two pathways (Obeid and Herrmann, 2009).
Low HDL-c in combination with raised triglyceride levels is considered an
atherogenic lipid profile (Obeid and Herrmann, 2009). In addition, LDL-c elevation in
hyperhomocystenemic rats may be attributed to the reduction in the number of LDL receptor
or reduced LDL binding to its receptor in rats. Changes in LDL- receptor contribute to the
elevation in serum cholesterol levels induced by methionine (Bhandari et al., 2011).
Also, the low methyl group availability decreases the synthesis of
phosphatidylcholine and the secretion of VLDL, causing triglyceride accumulation in the liver
(Li and Vance, 2008; Bravo et al., 2011). It has been reported that severe HHcy, caused by
CβS deficiency, leads to disturbances in the regulation of lipid metabolism and fat
accumulation in the liver (Gaull Get al., 1974; Namekata et al., 2004).
An inverse association between Hcy and lipoproteins, especially HDL, has been well
described in humans (Moat et al., 1999; Poloni et al., 2012) and various animal models of
HHcy (Poloni et al., 2015). It has been reported that HHcy alters intracellular lipid
metabolism (Werstuck et al., 2001) and may be associated with hepatic fat accumulation
(Carvalho et al., 2013). It seems that hypomethylation associated with HHcy is responsible for
lipid accumulation in tissues. HHcy and hypercholesterolemia are linked to the development
of atherothrombotic diseases (Obeid and Herrmann, 2009). However, many studies have
indicated that cholesterol homeostasis is a major mechanism for suppressing cardiovascular
disease (Koyama et al., 2015).

Results and Discussion
72
HDL is involved in several biological processes that counteract inflammation and
oxidative stress, by beneficially influencing, such as pancreatic beta-cell function, endothelial
vasoreactivity, endothelial apoptosis, restorative processes and monocyte activation as well as
adhesion molecules expression, thus being highly vasculoprotective (Chapman et al., 2011).
The effect of Hcy on HDL is probably related to inhibiting enzymes or molecules
participating in HDL-particle assembly (Obeid and Herrmann, 2009). Therefore, increased
risk for atherosclerosis and dysfunctional HDL particles in HHcy subjects might be related to
low activity of Paraoxonase-1 PON1 (Holven et al., 2008). PON1 is a serum HDL-associated
phosphotriesterase secreted mainly by the liver and shows a Hcy thiolactonase activity
(Domagala et al., 2006) thus protecting from atherosclerosis (Bhattacharyya et al., 2008).
In addition, HDL-c are antiatherogenic lipoproteins implicated in the protection of
LDL against oxidation (Mackness et al., 1993). Low HDL-c attributed to its central function
in the reverse of CHO transport, a process whereby excess cell CHO is taken up and
processed by HDL particles for further delivery to the liver for metabolism (Martinez et al.,
2004). Also the study of (Tselmin et al., 2013) suggests that the decrease in HDL is due to an
inhibition synthesis of Apo AI, the major apolipoprotein of HDL by Hcy.
On the other hand, A. spinosa powdered seeds administration caused a significant
reduction of TG, t-CHO and LDL-c showing the beneficial effect of this plant in the treatment
of the hyperlipidemia. This protection related to the decrease level of Hcy and therefore the
suppression of their cytotoxic effects on different organs. These corrections related to the
antioxidants and anti-inflammatory components of A. spinosa seeds.
Recent experimental study has shown that Argan oil blunted the increases of T-Cho,
LDL-C and TG concentrations in a high-fat diet (Sour et al., 2015).
Virgin Argan oil induces a lowering of the triglycerides level in men (Derouiche et al.,
2005) and of plasma LDL-c in healthy subjects (Drissi et al., 2004; Cherki et al., 2005).
Similar studies have demonstrated that the phenolic extract of Argan oil inhibits
human LDL-c oxidation and increases the cholesterol efflux from human T-helper precursor-1
macrophages (Berrougui et al., 2006). (Drissi et al., 2004) demonstrated that regular
consumption of virgin Argan oil induces a lowering of LDL-c and has antioxidant properties.

Results and Discussion
73
The decrease in cholesterol concentration could be due to a low intestinal absorption of
cholesterol because of the activity of saponins in Argan oil (Berrougui et al., 2003).
The impact of Argan oil consumption on oxidative stress plasma markers and HDL
paraoxonase 1 (PON1) activity has been evaluated. After three weeks of daily Argan oil
consumption (25 mL/day), plasma PON1 activity, antioxidant vitamins, and LDL
susceptibility to oxidation were measured. A significant increase in PON1 activity and
benficial effects on plasma lipid peroxide, conjugated dienes, and vitamin E concentration
were observed (Cherki et al., 2005).
Epidemiological studies have shown that consumption of food and beverages rich in
phenols can reduce the risk of heart disease by slowing the progression of atherosclerosis
principally by protecting LDL from oxidation (Duffy and Vita, 2003).
Argan oil contains a high amount of Vitamin E but also a non-negligible proportion of
phenolic compounds. The study of (Berrougui et al., 2003; Berrougui et al., 2004) have
previously demonstrated that chronic ingestion of crude Argan oil not only reduces plasma
cholesterol and LDL levels in rats fed with hypercholesterolemic diet , but also improve
endothelial function and prevents high blood pressure.
These effects are principally related to the richness of this oil in oleic and linoleic
acids and tocopherol.
The antioxidant activity of polyphenolics is principally defined by the presence of
orthodihydroxy substituents, which stabilize radicals and chelate metals. The antioxidant
effect of phenolic acids and their esters depends on the number of hydroxyl groups in the
molecule. This antioxidant compound protects against LDL-oxidation (Chen et al., 2004).
Concerning the anti-oxidative parameters, results shown that the content of GSH and
catalase activity in liver tissue was significantly decreased in response to the oral methionine
administration. Our findings are supported by the study of (Sakhri, 2014) who reported a
decrease in GSH levels of mice treated with 400 mg/kg of L-methionine for 21 days. Recently
(Meng et al, 2013) also found that the content of GSH in serum or liver tissue was decreased
in response to quercetin administration in rats fed a Methionine enriched diet.
It was reported that lower level of glutathione, the major intracellular antioxidant, is
accompanied by decreased level of S-adenosylmethionine (SAM), methyl donor in reactions

Results and Discussion
74
catalyzed methyltransferase and the main metabolic regulator of Hcy synthesis (Li and Vance,
2008; Bravo et al., 2011).
Glutathione is a key buffer of intercellular oxidative reduction reaction, and its
dependent antioxidant enzymes include glutathione S-transferase (GST) and glutathione
peroxidase (GPx), which play a fundamental role in cellular defense against reactive free
radical and other oxidant species (Stipanuk et al., 2006). Organic peroxides can also be
reduced by GPx and GSH S-transferase. Catalase can also reduce hydrogen peroxide but it is
present only in peroxisome. This makes GSH particularly important in the mitochondria in
defending against both physiologically and pathologically generated oxidative stress
(Fernández-Checa et al., 1997; Garcia-Ruiz and Fernández-Checa, 2006). There is
accumulating data that reduced GSH levels occur in many human diseases and they contribute
to worsening of the condition (Ballatori et al., 2009). While, oxidative injury plays a dominant
role in GSH depletion in many of these disorders, some are causally related to reduced
expression of GSH synthetic enzymes (Lu, 2009).
More, hyperaccumulation of methionine sulfoxide in the liver may induce more
serious (oxidative) hepatotoxicity in Cth−/− mice, whose levels of several antioxidative
cysteine metabolites, including GSH, and taurine/hypotaurine, were all downregulated
(Yamada et al., 2012). Hyperhomocysteinemia leads to increased oxidative stress via the
generation of ROS which weaken intracellular antioxidation defense systems or elicit
intracellular redox controlled inflammation responses (Welch et al., 1998).
In this study, we also verified the effect of Hcy on catalase, since this enzyme has the
higher activity among the hepatic antioxidant enzymes (Polavarapu et al., 1998; Kasdallah-
Grissa et al., 2007).
A significant inhibition of hepatic catalase activity was observed in
hyperhomocysteinemic mice. In agreement with our data, other studies suggest a negative
correlation between plasma Hcy levels and catalase activity in liver of rats, pointing a
significant reduction of hepatic antioxidant defenses (Woo et al., 2006; Chanson et al., 2007;
Matté et al., 2009).
In addition, it has been shown that Hcy can directly act on catalase and inhibit the
breakdown of H2O2 by conversion of the enzyme into the inactive form (Milton, 2008). Loss

Results and Discussion
75
of catalase activity is associated with increased susceptibility to oxidative stress (Góth et al.,
2004; Ho et al., 2004). The mechanism of Hcy inhibition of catalase is shared with a number
of inhibitors including 3-amino-1:2:4:- triazole (Margoliash and Novogrodsky , 1958; Putnam
et al., 2000) and amyloid-ß ( Milton, 1999).
Indeed, we demonstrated that A. spinosa powdered seeds when given in combination
with high L-methionine diet increased not significantly liver GSH level and significantly
catalase activity, indicating its beneficial effect in prevention against oxidative stress in vivo.
In accordance with our results, (Necib et al., 2013) demonstrated that Argan oil treatment
augments the GSH against mercuric chloride induced oxidative stress in experimental rats.
The elevated level of GSH protects cellular proteins against oxidation through glutathione
redox cycle and directly detoxifies reactive species (Ketterer, 1998). Also, our results showed
an increase of catalase activity in (MP) group. These results are in agreement with those of
(Benajiba et al., 2002) who showed that the activities of cytosolic CAT were significantly
higher in Wistar rats treated with Argan oil in comparison with untreated rats.
Argan oil is rich en polyphenols. These compounds have been found to modulate
expression and activity of catalase and eNOS in several tissues, increases catalase activity in
guinea pig cardiac tissue (Floreani et al., 2003), or rat liver (Kasdallah-Grissa et al., 2007),
while red wine polyphenols extract increases phosphorylation of eNOS in porcine coronary
arteries (Madeira et al., 2009) and in aorta and carotid artery of Zucker Fatty rat, a model of
obesity, with an increase of NO bioavailability (Agouni et al., 2009).
For liver enzyme activities, data have shown a significant increase of AST in (M)
group but not ALT values. We believe that the absence of significant differences in ALT
levels in our study could be due to the short period of treatment with Hcy (21 days only).
Liver injury could be identified by serum markers, such as (ALT) and (AST), which
are increased as a result of hepatic necrosis (Ozer et al., 2008). However, increased activity of
AST indicated cardiac disorders (Killip and Payne, 1960).
(Yalcınkaya et al., 2009) have shown that a high methionine diet supplemented for 6
months caused HHcy, and increased serum ALT and AST levels in rats. The increment of the

Results and Discussion
76
activities of AST and ALT in serum may be mainly due to the leakage of these enzymes from
the liver cytosol into the blood stream (Navarro et al., 1993).
In this context, Hcy has been related to hepatotoxic conditions in numerous reports,
which have showed a positive correlation between HHcy and plasma aminotransferases
activities in clinical and experimental studies (Huang et al., 2001 ; Frelut et al., 2006; Woo et
al., 2006) associated with ROS production and hepatic lipid peroxidation (Huang et al., 2001 ;
Woo et al., 2006).
Previously, we demonstrated that the administration of A.spinosa powdered seeds
increased tissue antioxidant capacity and significantly protected the liver against liver injury
in mice. Simultaneously, the serum activity of AST was significantly decreased, indicating
that a toxic action was displayed with excessive intake of A. spinosa extract crude.
3- Effect of treatment on histology of aorta heart and liver
Histological analysis showed that HHcy induced by the high methionine intake
prompted an angiotoxic activity on the aorta (arch, abdominal and iliac), cardiac and liver
tissue damages. This was observed through the loss and degeneration of endothelium,
formation of foam cells in the different sections of the aorta, alteration of the cardiac muscle,
and liver macrovesicular steatosis.
In our experimental situation, is due to elevated Hcy levels, which perturbs lipid
profile, and decreases tissue level of reduced GSH, and catalase activity, the well known bio-
markers of oxidative stress.
The obtained data in the current investigation were in agreement with the studies of
(Boudebouz, 2013; Sakhri 2014). They reported that the high dose of methionine perturbed
the structures of aorta, heart and liver.
It is belived that HHcy leads to endothelial cell damage, reduction in the flexibility of
vessels, and alters the process of homeostasis (Baszczuk and Kopczynski, 2014).
Homocysteine-induced injury to the arterial wall is one of the factors that can initiate
the process of atherosclerosis, leading to endothelial dysfunction and eventually to heart
attacks and strokes (Gallai et al., 2001; Papatheodorou and Weiss, 2007). Evidence from

Results and Discussion
77
animal models of HHcy suggest that endothelial dysfunction is largely due to oxidative stress
and decreased bio- availability of NO (Pacher, 2007), NO may protect against the onset of
vascular diseases (Cooke and Dzau, 1997).
Homocysteine promoted oxidative stress through production of reactive oxygen
species (ROS). ROS disrupts endothelial cell integrity, which in turn, can cause endothelial
cell damage predispose affected vessels to the subsequent development of atherosclerosis
(Kanani et al., 1999).
Recently, the study of (Chen et al., 2015) demonstrated that HHcy promotes
atherosclerosis progress. This may be associated with decreased levels of endothelial or aortic
protein S-nitrosylation which plays an important role in the regulation of cardiovascular
functions in nitric oxide (NO) Pathway.
Another possible mechanisms by which Hcy must be contributing to atherogenesis
and thrombosis include increased smooth muscle cell proliferation, cytotoxicity, increased
oxidative stress, stimulation of low density lipoprotein oxidation, induction of endothelial
dysfunction, enhanced coagulability and platelet activation (Willoughby et al., 2002; Luo et
al., 2006).
In addition, in the present study an increase in atherogenic index was found,
consequently of the increase of cholesterol and decrease of HDL-c. This atherogenic index
indicates the disposition of foam cell or fatty infiltration or lipids in heart, coronaries, aorta
liver and kidneys. The higher the atherogenic index, the higher is the risk of the above organs
for damage (Mehta et al., 2003).
Studies in several animal species, including rabbits, baboons, and rats, have
demonstrated desquamation of endothelial cells, fragmentation of the internal elastic lamina,
disruption of elastic fibers, and focal areas of smooth muscle hyperplasia (Rolland et al.,
1995). (Zulli and Hare, 2009) has been reported that the combination of high methionine and
cholesterol increased the alterations of the arterial wall structures and the thickness of the
aortic wall in animal models.
However, in methionine-treated animals, it was shown an aortic angiotoxic action
following to an increase of plasma Hcy levels. (Raghuveer et al., 2001) have reported that,

Results and Discussion
78
acute elevations in plasma Hcy after methionine loading causes vessel endothelial dysfunction
and this could be reversed by administration of vitamin E in humans.
It has been documented that Hcy can interact with different plasma and cellular
proteins and by forming mixed disulfide conjugates, alters the physicochemical properties of
the affected proteins. This has been also proposed as a potential mechanism for Hcy induced
cellular dysfunction (Barbato et al., 2007).
Accumulating evidence indicates that oxidative stress was the major mechanism of
vascular injury caused by Hcy. The active free sulfhydryl contained in Hcy is prone to self-
oxidation under catalysis of the copper found in blood, forming homocysteine–homocysteine
mixed bisulfide. In this course, many series of oxygen reactions with associated toxic effects
could be produced, including the formation of hydrogen peroxide (H2O2), superoxide anion,
and the hydroxyl radical. This induced membrane lipid peroxidation, directly resulted in
chemical injury of vascular endothelial cells (VEC) and impaired endothelial function
(Nygard et al.,1997). Then, by carboxyl methylation, Hcy decreased activity of membranes.
Accordingly, Hcy inhibited the cell cycle of VEC, decreased the regeneration capacity of the
endothelium and induced endothelial dysfunction (Lee and Wang, 1999). Therefore, the
supplementation with antioxidants like the crude extract of A.spinosa to reduce some of the
prooxidant activity of Hcy is beneficial to alleviate endothelial injury
In addition to early cardiovascular events, patients with homocysteinuria develop
hepatic steatosis or ‘‘fatty liver” which is characterized by enlarged, multinucleated
hepatocytes containing microvesicular lipid droplets (Mudd et al., 1995), which can progress
into hepatocellular carcinoma (Dara and Kaplowitz, 2011). It has been reported that Hcy
elicits hepatic damage in experimental models by three mechanisms: oxidative stress,
endoplasmic reticulum stress, and activation of proinflammatory factors (Ji and Kaplowitz ,
2004; Robert et al., 2005). (Woo and Siow, 2008) showed that HHcy, induced by high-
methionine diet for 4 weeks, elevated the expression and the protein synthesis of monocyte
chemoattractant protein-1 (MCP-1) in plasma and in liver tissue homogenate, due to
hepatocyte production, suggesting that Hcy may contribute to chronic inflammation in this
organ studied hyperhomocysteinemic CβS-deficient mice and demonstrated foci of
perilobular mononuclear inflammatory infiltrate around the vessels.

Results and Discussion
79
Also, (Ji and Kaplowitz, 2004; Matté et al., 2009) showed that HHcy increased
mediators of inflammation, such as nuclear factor kappa B NFκB, interleukin (IL)-1b, IL-6
and IL-8 in liver.
We also observed the disruption of the elastic lamina and disorganization of elastic
fibers. It has been reported that the disappearance of the aortic elastic lamina might be due to
production of elastase, a serine protease, by increased number of SMCs induced by Hcy
(Jourdheuil-Rahmani et al., 1997; Zulli et al., 1998).
In addition, A.spinosa powdered seeds treatment was found to decrease highly and
significantly serum AST activities and to ameliorate histopathologic changes in the liver,
heart due to the high-fed methionine diet, and prevents the endothelial alteration as shown by
the morphological data. The study of (kumar et al., 2010) suggested that an increase intake of
antioxidants appeared to be protective in cardiovascular diseases. Also, epidemiological
studies have shown that consumption of food and beverages rich in phenols can reduce the
risk of heart disease by slowing the progression of atherosclerosis principally by protecting
LDL from oxidation (Duffy and Vita, 2003).
The work of Zerizer & Naimi (2004), and Benmebarek et al., (2013) proved that , the
administration of L-methionine (200 mg/kg) to rats and mice respectively during 21 days,
could damage the aorta and heart tissue and the treatment of these animals with vitamins B9,
B12 , B6 and Stachys mialhesi extract respectively corrected these alterations.
Another work of (Zerizer et al., 2008) established that higher level of Hcy could
stimulate the angiogenesis on the aorta of rats, and the treatment with the extracts of
medicinal plants Stachys mialhesi and Chrysanthemum Macrocarpum could inhibit the
angiogenesis.
4- Anti- biofilm effect of Argan oil
Bacterial infectious diseases represent an important cause of morbidity and mortality
worldwide. Therefore, the development of new antimicrobial agents for the treatment of
bacterial infections is of increasing interest.
Normally, bacterial infectious diseases could be assumed as being a “biofilm type of
infection” (Draelos, 2010), and recent research has shown that generally these bacteria

Results and Discussion
80
structured in a sessile form are more resistant to various antimicrobial treatments (Olsen,
2015).
One of the most important characteristic of biofilms is their increased tolerance to
antimicrobial agents (Wimpenny et al., 2000). The presence of exopolysaccharides, which
accounts for a majority of the dry mass of the biofilm, is known to act as a mechanical barrier,
thereby diminishing the effectiveness of various antibiotics (Stewart, 1996). Moreover, the
negatively charged polymers present on the surface of the biofilm matrix are known to
interact with positively charged antibiotics (such as aminoglycoside), thereby
limiting/slowing the penetration of these drugs (Nichols et al., 1988).
Additionally, altered environmental conditions may trigger differential oxygen
concentration, such as the surface of the biofilms may show higher oxygen levels, whereas a
scarcity of oxygen may be observed toward the core of the biofilms, thereby making the core
less accessible to the antibiotics (De Beer et al., 1994; Yang et al., 2008).
Biofilms exhibit regions of oxygen limitation and nutrient deficiency that, in contrast
to planktonic cultures, are non-uniformly distributed (Xu et al., 1998). Bacteria located
within starvation zones may be metabolically dormant and thus it has been suggested that
growth-dependent changes in cellular processes may contribute to reduced sensitivity to many
classes of antimicrobial agents (Borriello et al., 2004 ; Field et al., 2005).
Many bacteria are known to regulate their cooperative activities and physiological
processes through a mechanism called quorum sensing (QS), in which bacterial cells
communicate with each other by releasing, sensing and responding to small diffusible signal
molecules. A quorum sensing mechanism, including symbiosis, formation of spore or fruiting
bodies, bacteriocin production, genetic competence, programmed cell death, virulence and
biofilm formation (Li and Tian, 2012).
Many naturally occurring compounds found in plants, herbs, and spices have been
shown to possess antimicrobial functions and serve as a source of antimicrobial agents against
pathogens (Deans and Ritchie 1987; Kumar et al., 2006).

Results and Discussion
81
Thus, the antibacterial agents derived from the natural source (plants) may serve as an
effective alternative, due to the presence of secondary metabolites, which are known to enjoy
selectional advantages against the resistance organisms (Butler and Buss, 2006).
In this study, the anti-biofilm formation of the Algerian Argan oil was assessed against
5 species of bacteria, that belong to Gram positive (Streptococcus mutans, Streptococcus
anginosus, Streptococcus intermedius, Streptococcus uberis and Staphylococcus
haemolyticus) strains.
The results obtained with the broth microdilution test showed for the first time, that
there was a trend of increasing inhibition of S. intermedius, St haemolyticus, S. mutans and S.
anginosus biofilm formation as the Argan oil was getting more concentrated.
The first one (100 µg/ml) of Argan oil was effective to remove S. uberis biofilm but
not to kill the cells. However, the highest effect of this oil was on S. intermedius, St
haemolyticus, S. mutans, S. anginosus biofilms with reduction percent greater than 90%.
The inhibitory activity produced by the Argan oil could be due to
the presence of individual phytochemicals, which are active against this strains of bacteria or
could be as a result of the synergistic effect of two or more phytochemicals that are
contained in the oil (Da Silva et al., 2016). The Argan oil was found to display antibiofilm
properties, probably by the richness of natural phenols such as: caffeic acid, vanillic acid,
tyrosol, epicatechin …. (Charrouf and Guillaume, 2007).
Argan oil has been confirmed to contain phytochemicals such as steroids,
tannins, flavonoids and saponins (Charrouf and Guillaume, 2007). And these phytochemicals
have been demonstrated to associate with bacterial proteins and inhibit microbial adhesion,
enzymes, cell envelop and transport proteins (Samy et al., 2010; Upadhyay et al., 2014). It
is, therefore, likely that the Argan oil inhibited biofilm formation through some of these
mechanisms. Bacterial adhesion is important during biofilm formation (Rabin et al., 2015)
and agents that disrupt bacterial adhesion to surfaces have the potential to act as anti-biofilm
agents.
In this context, the study of (Stojkovic et al., 2013) demonstrated a good antioxidant
and antimicrobial activity of caffeic acid. Also, a number of polyphenols (hydroxycinnamic

Results and Discussion
82
acid, rutin, epicatechin) have been found to block quorum sensing in Chromobacterium
violaceum (Nazzaro et al., 2013).
The microbial biofilm inhibition can be managed either by preventing the attachment
of the organism to the surface or by breaking the structure of the biofilm
if they formed (Gupta et al., 2016).
Another explanation that Argan oil might have contained antibacterial phytochemicals
that stimulate a stress response in the biofilm. Several studies have demonstrated that when
microbes are stressed, such as under antibiotic treatment, genes associated with biofilm
formation are stimulated and the bacteria convert to the biofilm phenotype (Ackart et al.,
2013).
Recently, Lotfi et al., (2015) revealed that Argan oil (extracted from the kernels of the
Argan tree of Bechar, Algeria ) has shown a positive effect on resistant planktonic bacteria
studied in particular staphylococcus aurus and staphylococcus white.
According to our finding, spices such as garlic (Bjarnsholt et al., 2005), ginger,
cinnamon (Niu et al., 2006) , clove (Khan et al., 2009), cumin (Packiavathy et al., 2012) and
turmeric (Packiavathy et al., 2014) have been found to display QS-mediated biofilm
inhibitory properties.
The findings suggest that curcumin derived from C. longa significantly inhibits the
formation of biofilm by reducing exopolysaccharide as well as alginate production, probably
by interfering with the signal molecules of the QS system. Studies have also shown that
curcumin enhances the susceptibility of the organism to conventional antibiotic (Packiavathy
et al., 2014).
Zingerone, one of the main chemical constituents of ginger (Zingiber officinale),
displays the ability to inhibit as well as eradicate biofilms formed by P. aeruginosa. In a study
conducted with ginger extract, pretreatment with the extract effectively reduced the
production of EPS (Kim and Park, 2013). Zingerone also increased the susceptibility of the
organism to conventional antibiotics, when used in combination with the standard antibiotics
such as ciprofloxacin (Kumar et al., 2013).

Results and Discussion
83
Garlic (Allium sativum) was found to block QS mediated biofilm formation and
virulence development by inhibition of LuxR-type receptor (Prigent-Combaret et al., 1999)
Essential oils are aromatic oily liquids from plant materials and are well known for
their antibacterial properties (Burt, 2004). A variety of researches have shown that the
biofilm could be removed effectively by essential oils for example essential oils of Piper
bredemeyeri, Piper brachypodom, Piper bogotence, Gaultheria procumbens L., Achillea
millefolium (yarrow), Syzygium aromaticum (clove), Coriandrum sativum (coriander),
Cinnamomum verum (cinnamon), thyme and Origanum vulgare (oregano) showed inhibitory
effects on biofilm formation at sublethal concentrations (Khan et al., 2009; Musthafa et
al.,2010; Olivero et al., 2011; Jadhav et al., 2013)
Traditional Chinese medicinal herbs have also been screened for antibiofilm properties
against microbes like C. violaceum and P. aeruginosa (Koh and Tham, 2011).
Also, Alcoholic extracts of some medicinal plants of Egypt were investigated for anti-
QS activity against C. violaceum (Zaki et al., 2013). Findings also reveal information about
the effectiveness of some plants (leaves of Adhatoda vasica Nees, Bauhinia purpurea L.,
Lantana camara L., Myoporum laetum G. Forst., the fruits of Piper longum L., and aerial
parts of Taraxacum officinale) against the QS system (Zaki et al., 2013).
The microorganisms used in this study are generally known to be a pathogen of dental
caries. In this context, infections, including those caused by oral bacteria, are more likely
involved in CVD progression than previously thought. Many studies have established a
plausible link between oral bacteria and atherosclerosis, which is extremely complex and it’s
highly likely that more than one mechanism is involved (Leishman et al., 2010).
Endocarditis is the interaction between the surfaces of the
endothelium and bacteria. Though the early association is weak, but with the advent of any
wound, the microbes turn opportunistic and form a strong biofilm-aided association which can
damage heart valves (Kokare et al., 2009). The organisms can primarily enter into the blood
stream through oropharynx, genitourinary tract and gastrointestinal tract. Generally, the
adherence of microbes to the intact endothelium is very poor, but in case of a wounded or
damaged epithelium, a condition namely nonbacterial thrombotic endocarditis develops

Results and Discussion
84
(Kokare et al., 2009). In this condition, the red blood cells, platelets and fibrin accumulate at
the site of injury. Endothelial cells secrete fibronectin which have the ability to bind to
collagen, fibrin, human cell as well as bacteria. Microbes like Staphylococcus and
Streptococcus sp. have fibronectin receptors which can form biofilms on the site of injury as
well as damage the tissue of the valves (Kokare et al., 2009). Therefore, an invasion of
endothelial cells by oral bacteria may lead to changes in the proinflammatory and
proatherogenic properties of endothelial cells as well as programmed cell death, all of which
are indicative of endothelial dysfunction (Leishman et al., 2010).
Through the results obtained, Argan oil can be used in the manufacturing of the herbal
toothpaste, which prevents cardiovascular disease indirectly.

Conclusion
and
Perspectives

Conclusion and perspectives
85
The purpose of this study was to induce hyperhomocysteinemia by administration of
high dose of L-methionine (500mg/kg) during 21 days in an in vivo animal; therefore evaluate
the protective and preventive effect of the powdered seeds of A.spinosa against the metabolic
and structural disorders induced in L-methionine treated mice. On the other hand, investigated
the antibiofilm effect of Argan oil againt 5 species of bacteria. These bacteria can lodge on
heart valves and cause infection of the endocardium.
The current study has shown that high L-methionine diet clearly caused some metabolic
disorders manifested by:
Hyperhomocysteinemia.
Hyperlipedemia with an increase in the total lipid rate: the total cholesterol and
triglycerides.
Dyslipoproteinemia with a decrease in HDL-c and an increase LDL-c.
An increase in the atherogenic index, consequently of the increase of cholesterol and
decrease of HDL-c.
Anti-oxidative disorder by the depletion of the reduced glutathione and the catalase
activity.
Increase of liver enzyme especially AST.
In addition, histology analyses showed cellular damages of aorta, heart and liver
characterized by:
Loss and degeneration of endothelium.
Formation of foam cells in the different sections of the aorta.
Formation of lipids droplets in aorta and liver.
Disappearance of elastics fibers especially in the arch aorta sections.
Alteration of the cardiac muscle.
Destruction of membrane cells, hepatocellular ballooning, hypertrophy of
some hepatocyte nuclei, cytoplasmic vacuolization, and macrovesicular
hepatic steatosis.

Conclusion and perspectives
86
Meanwhile, A diet enriched with (150mg/kg/day) of Algerian Argania spinosa
seeds was effective in preventing the increase of these metabolic disorders, endothelial
alteration, and the heart and liver damages induced by methionine enriched diet in mice.
Also, Considering the results obtained from the in vitro study, we can conclude that
the Algerian Argan oil was able to efficiently reduce biofilms formation of (Streptococcus
mutans, Streptococcus anginosus, Streptococcus intermedius and staphylococcus
haemolyticus) strains, when oil was getting more concentrated.
This preventive protection of Argania spinosa (seeds and oil ) might be related to its
content of many antioxidants and anti inflammatory compounds such as tocopherols (vitamin
E), phenols (caffeic acid, oleuropein, vanillic acid, tyrosol, catechol, resorcinol,
epicatechin and catechin), carotenes, squalene, and fatty acids, (80% unsaturated fatty acids).
Based on the findings of this study our future work and perspectives can evaluate
many topics:
Purification and determination of the bioactive molecules presented in the
powdered seeds of Argania spinosa.
Determination of antioxidant enzyme superoxide dismutase and glutathione-s-
Transferase.
Study the gene expression of antioxidant enzymes.
Determination of pro-inflammatory cytokines in mice administered with high
dose of L-Methionine and treated with Argania spinosa.
Isolate the specific antibacterial principles in Argan oil.
Determine the activity of the plant extract on other types of bacteria as
staphylococcus aurus, in addition to the synergistic activity of this medicinal
plant with antibiotics.

References

References
87
Abraham J. M., Cho L. (2010). The homocysteine hypothesis: still relevant to the
prevention and treatment of cardiovascular disease?. Cleveland Clin J Med. 77: 911– 8.
Ackart D., Hascal-Dove L., Ceres S. M., Kirk N., Podell B., Maled C. (2013).
Expression of antimicrobial tolerance by attached communities of Mycobacterium
tuberculosis. Pathog Dis. 70: 359–69.
Adinolfi L. E., Ingrosso D., Cesaro G., Cimmino A., D’Anto M., Capasso R., ZappiaV.,
Ruggiero G. (2005). Hyperhomocysteinemia and the MTHFR C677T polymorphism promote
steatosis and fibrosis in chronic hepatitis C patients. Hepatology. 41: 995–1003.
Aebi H. (1984). Catalase in vitro. Methods in enzymology. 105: 121-126.
Agouni A., Lagrue-Lak-Hal A. H., Mostefai H. A., Tesse A., Mulder P., Rouet P., et al. (2009). Red wine polyphenols prevent metabolic and cardiovascular alterations associated
with obesity in Zucker fatty rats (Fa/Fa). PLoS ONE. 5: 5557.
Aird W.C. (2004). Endothelium as an organ system. Crit Care Med. 32(5):271–279.
Aird W. C. (2007). Phenotypic heterogeneity of the endothelium. Circulation research. 100:
174-190.
Alados C. L., El Aich A. (2008). Stress assessment of argan (Argania spinosa (L.) Skeels) in
response to land uses across an aridity gradient: translational asymmetry and branch fractal
dimension. J arid environments. 72: 338-349.
Alaoui K., Lagorce J. F., Cherrah Y., Hassar M., Amarouch H., Roquebert J. (1998).
Analgesic and anti-inflammatory activity of saponins of Argania spinosa. Ann Pharm Fr. 5:
220-8.
Amzal H., Alaoui K., Tok S., Errachidi A., Charof R., Cherrah Y., Benjouad A. (2008).
Protective effect of saponins from Argania spinosa against free radical-induced oxidative.
haemolysis Fitoterapia . 79(5): 337- 44.
Andersson A., Lindgren A., Hultberg B. (1995). Effect of thiol oxidation and thiol export
from erythrocytes on determination of redox status of homocysteine and other thiols in plasma
from healthy subjects and patients with cerebral infarction. Clin Chem. 41: 361 – 366.
Athyros G. Tziomalos K. , Karagiannis A ., Mikhailidis D. P. (2010). Homocysteine: An
Emerging Cardiovascular Risk Factor that Never Really Made it. Open Clin Chemist J. 3:19-
24.
Ballatori N., Krance S. M., Notenboom S., Shi S., Tieu K., Hammond C. L. (2009).
Glutathione dysregulation and the etiology and progression of human diseases. Biol Chem.
390: 191–214.
Barbato J. C., Catanescu O., Murray K., DiBello P. M., Jacobsen D. W. (2007). Targeting
of metallothionein by L-homocysteine: a novel mechanism for disruption of zinc and redox
homeostasis. Arterioscler Thromb Vasc Biol. 27: 49–54.

References
88
Baszczuk A., Kopczynski Z. (2014). Hyperhomocystenemia in patients with cardiovascular
disease. Postepy Hig Med Dosw. 2: 579-89.
Bautista L. E., Arenas I. A., Penuela A., Martinez L. X. (2002). Total plasma homo-
cysteine level and risk of cardiovascular disease: a meta-analysis of prospective cohort
studies. J Clin Epidemiol. 55: 882– 887.
Beachey E. H. (1981). Bacterial adherence: adhesion-receptor interactions mediating the
attachment of bacterial to mucosal surfaces. J Infect Dis. 143:325-345.
Benajiba N., Morel S., De Leiris J., Boucher F., Charrouf Z., Mokhtar N. (2002). The
effect of argan oil on heart function during ischemia and reperfusion. Therapie. 57: 246-52.
Benmebarek A., Zerizer S., Laggoune S. et Kabouche Z. (2013). Effect of Stachys
milhesi de Noé on the inflammation induced by hyperhomocysteinemia in cardiovascular
diseases. Der Pharmacer Letter 5 : 212- 223.
Bennani H., Drissi A., Giton F., Kheuang L., Fiet J., Adlouni A. (2006). Antiproliferative
effect of polyphenols and sterols of virgin argan oil on human prostate cancer cell lines.
Cancer Detect Prev . 31: 64- 69.
Bennani H., Drissi A., Giton F., Kheuang L., Fiet J., Adlouni A. (2007). Antiproliferative
effect of polyphenols and sterols of virgin argan oil on human prostate cancer cell
lines. Cancer detection and prevention. 31: 64-69.
Bennani H., et al. (2009). Impact of argan oil on prostate cancer antiproliferative effect:
Study of polyphenols. Rev Franco Lab. 416: 23-26.
Berrougui H., Alvarez S., Perez-Guerrero C., Ettaib A., Hmamouchi M., Marhuenda E.,
Herrera M.D. (2004). Argan (Argania spinosa) oil lowers blood pressure and improves
endothelial dysfunction in spontaneously hypertensive rats. Br J Nutr . 92: 921- 929.
Berrougui H., Cloutier M., Isabelle M., Khalil A. (2006). Phenolic-extract from argan oil
(Argania spinosa L.) inhibits human low-density lipoprotein (LDL) oxidation and enhances
cholesterol efflux from human THP-1 macrophages. Atherosclerosis. 184: 389–96.
Berrougui H., Ettaib A., Herrera Gonzalez M. D., Alvarez de Sotomayor M., Bennani-
Kabchi N., Hmamouchi M. (2003). Hypolipidemic and hypocholesterolemic effect of argan
oil (Argania spinosa L.) in Meriones shawi rats .J Ethnopharmacology. 89: 15–18.
Berstad P., Konstantinova S.V., Refsum H., Nurk E., Vollset S.E., Tell G.S., Ueland
P.M., Drevon C.A., Ursin G. (2007) . Dietary fat and plasma total homocysteine
concentrations in 2 adult age groups: the Hordaland Homocysteine Study. Am J Clin Nutr.
85: 1598–1605.
Bhandari U., Pathan R. A., Kuma V., Khanna N. (2011). Ameliorative role of
atorvastatin on methionine- induced hyperhomocystenemia and hematological changes in
albino rats. Indian J Experiment Bio. 49: 132-139.

References
89
Bhattacharyya T., Nicholls S.J., Topol E.J., Zhang R., Yang X., Schmitt D., Fu X., Shao
M., Brennan D.M., Ellis S.G., Brennan M.L., Allayee H., Lusis A.J., Hazen S.L. (2008).
Relationship of paraoxonase 1 (PON1) gene polymorphisms and functional activity with
systemic oxidative stress and cardiovascular risk. JAMA . 299: 1265–1276.
Bjarnsholt T., Ciofu O., Molin S., Givskov M., Hoiby N. (2013). Applying insights from
biofilm biology to drug development - can a new approach be developed?. Nat Rev Drug
Discov. 12: 791-808.
Bjarnsholt T., Jensen P., Rasmussen T., Christophersen L., Calum H., Hentzer M., et
al. (2005). Garlic blocks quorum sensing and promotes rapid clearing of pulmonary
Pseudomonas aeruginosa infections. Microbiology. 151: 3873-80.
Boger R. H. (2003).The emerging role of asymmetric dimethylarginine as a novel
cardiovascular risk factor. Cardiovasc Res. 59: 824–33.
Boger R. H., Bode-Boger S. M., Sydow K., Heistad D. D., Lentz S. R. ( 2000). Plasma
concentration of asymmetric dimethylarginine, an endogenous inhibitor of nitric oxide
synthase, is elevated in monkeys with hyperhomocyst( e)inemia or hypercholesterolemia.
Arterioscl Thromb Vasc Biol. 20: 1557–64.
Borriello G., Werner E., Roe F., Kim A. M., Ehrlich G. D., Stewart P.S. (2004). Oxygen
limitation contributes to antibiotic tolerance of Pseudomonas aeruginosa in biofilms.
Antimicrob Agents Chemother .48: 2659–64.
Bosy-Westphal A., Petersen S., Hinrichsen H., Czech N., Muller J. M.( 2001). Increased
plasma homocysteine in liver cirrhosis. Hepatol Res. 20: 28–38.
Boudebouz S. (2013). Effet comparatif de l’extrait butanolique de la plante Stachys mialhesi
avec les vitamines B9 et B12 sur quelques paramètres biochimiques et la structure de l’aorte
chez des souris induites par une hyperhomocysteinemia. Thesis of magister. University of
Constantine 1.
Brandon B. (2009). Changes in serum homocysteine levels after roux en Y gastric bypass
surgery in severe obesity. Doctorat Thesis, university of Winston-Salem, North Carolina.
Bravo E., Palleschi S., Aspichueta P., Buqué X., Rossi B., Cano A., et al. (2011). High fat
dietinduced non-alcoholic fatty liver disease in rats is associated with hyperhomocysteinemia
caused by down regulation of the transsulphuration pathway. Lipids Health Dis.10:60.
Brosnan J. T., Jacobs R.L., Stead L. M., Brosnan M. E. (2004). Methylation demand: a
key determinant of homocysteine metabolism. Acta Biochim Pol. 51: 405–413.
Burt S. A., Ojo-Fakunle V.T.A., Woertman J., Veldhuizen E. J. A. (2014). The natural
antimicrobial carvacrol inhibits quorum sensing in Chromobacterium violaceum and reduces
bacterial biofilmformation at sub-lethal concentrations. PLoS One. 9: e93414.
Burt S. (2004). Essential oils: their antibacterial properties and potential applications in
foods: a review. Int J Food Microbiol. 94: 223- 53.

References
90
Butler M., Buss A. (2006).Natural products- the future scaffolds for novel antibiotics?.
Biochem Pharmacol. 12: 919-29.
Carvalho S. C., Cartaxo M.T., Siqueira M. D., Siqueira E.R., Gomes A.V., Silva K. A.,
Bezerra L.C. (2013). Plasmatic higher levels of homocysteine in Non-alcoholic fatty liver
disease (NAFLD). J Nutrition. 12: 37.
Cellarier E., Durando X., Vasson M. P., Farges M. C., Demiden A., Maurizis J. C.,
Madelmont J. C., Chollet P. (2003). Methionine dependency and cancer treatment. Cancer
Treat Rev. 29: 489- 499.
Chanson A., Rock E., Martin J. F., Liotard A., Brachet P. (2007). Preferential response of
glutathione-related enzymes to folate-dependent changes in the redox state of rat liver. Eur J
Nutr. 46: 204–212.
Chapman M. J., Ginsberg H. N., Amarenco P., Andreotti F., Borén J., Catapano A. L.,
Descamps O. S., Fisher E. (2011). Triglyceride-rich lipoproteins and high-density
lipoprotein cholesterol in patients at high risk of cardiovascular disease: evidence and
guidance for management. Eur Heart J. 32: 1345-1361.
Charrouf Z., Guillaume D. (2007). "Phenols and Polyphenols from Argania
spinosa".American J Food Tech. 2: 679.
Charrouf Z., Benomar M., Guillaume D. (2007). Huile d’argan et système cardiovasculaire,
une revue. Coeur Vaisseaux. 4 : 50-55.
Charrouf Z., Guillaume D. (1999). Ethnoeconomical and phytochemical study of Argania
spinosa (L) Skeels. J Ethnopharmacol. 67:7–14.
Chaussod R., Adlouni A., Christon R. (2005). L’arganier et l’huile d’argane au Maroc: vers
la mutation d’un système agroforestier traditionnel : enjeux et contribution de la recherche.
Cah Agric. 14: 351-6.
Chen C. Y., Milbury P. E., Kwak H. K., et al. (2004). Avenanthramides and phenolic acids
from oats are bioavailable and act synergistically with Vitamin C to enhance hamster and
human LDL resistance to oxidation. J Nutr. 134:1459–66.
Chen Y., Liu R., Zhang G., Yu Q., Jia M., Zheng C., Wang Y., Xu C., Zhang Y. (2015).
Hypercysteinemia promotes atherosclerosis by reducing protein S-nitrosylation. Biomedicine
Pharmacotherapy. 70: 253–259.
Cherki M., Berrougui H., Drissi A., Adlouni A., Khalil A. (2006). Argan oil: which
benefits on cardiovascular diseaese?. Pharmacol Res. 54: 1-5.
Cherki M., Derouiche A., Drissi A., Bamou Y., El Messal M., Idrissi Oudghiri A. (2005).
Consumption of argan oil may have an antiatherogenic effect by improving paraoxonase
activities and antioxidant status: Intervention study in healthy men. Nutr Metab Cardiovasc
Dis. 15: 352-360.
Chia J. S., Lin Y. L., Lien H.T., Chen J. Y. (2004). Platelet aggregation induced by
serotype polysaccharides from Streptococcus mutans. Infect Immun. 72: 2605-17.

References
91
Chiew Y. F., Charles M., Johnstone M. C., Thompson K. M., Parnell K. D., Penno. E. C. (2007). Detection of vancomycin heteroresistant Staphylococcus haemolyticus and
vancomycin intermediate resistant Staphylococcus epidermidis by means of vancomycin
screening agar. Pathology. 39: 375–377.
Claridge J.E., Attorri S., Musher D.M., Hebert J., Dunbar S. (2001). Streptococcus
intermedius, Streptococcus constellatus, and Streptococcus anginosus ("Streptococcus milleri
group") are of different clinical importance and are not equally associated with abscess. Clin
Infect Dis. 32:1511-1515.
Cochain C., Zernecke A. (2015). Macrophages and immune cells in atherosclerosis: Recent
advances and novel nconcepts. Basic Res Cardiol. 110: 34.
Cooke J.P., Dzau V. J. (1997). Nitric oxide synthase: role in the genesis of vascular disease.
Annu Rev Med . 48: 489–509.
Cooper J. D., Gotoff R. A. (2016). Streptococcus anginosus purulent pericarditis with
contiguous hepatic and subdiaphragmatic fluid collections: First Reported Case. Infect dis clin
pract. 24: 37-39.
Costello E. K., Lauber C. L., Hamady M., Fierer N., Gordon J. I., Knight R. (2009).
Bacterial community variation in human body habitats across space and time. Science.
326:1694e7.
Costerton J. W., Montanaro L., Arciola C. R. (2005). Biofilm in implant infections: its
production and regulation. Int J Artif Organs. 28: 1062-8.
Cox R. A., Garcia-Palmieri M. R. (1990). Cholesterol, triglycerides, and associated
lipoproteins. Clinical Methods: The History, Physical, and Laboratory Examinations. 3rd
edition.
Cui R., Moriyama Y., Koike K. A., Date C., Kikuchi S., Tamakoshi A., Iso H. (2008).
Serum total homocysteine concentrations and risk of mortality from stroke and coronary heart
disease in Japanese: the JACC study. Atherosclerosis. 198: 412–418.
Curro M., Gugliandolo A., Gangemi C., Risitano R., Ientile R., Caccamo D. (2014).
Toxic effects of mildly elevated homocysteine concentrations in neuronal-like cells.
Neurochem Res. 39: 1485–95.
Da Silva A. P. S. A., da Silva L. C. N., da Fonseca C. S. M., de Araújo J. M., dos Santos
Correia M. T., da Silva Cavalcanti M., de Menezes Lima V. L. (2016). Antimicrobial
Activity and Phytochemical Analysis of Organic Extracts from Cleome spinosa. Frontiers in
microbiology, 7: 963.
Dalton T., Dowd S. E., Wolcott R. D. (2011). An in vivo polymicrobial biofilm wound
infection model to study interspecies interactions. PloS One. 6: 11-e27317.

References
92
Dara L., Kaplowitz C. Ji .N. (2011). The contribution of endoplasmic reticulum stress to
liver diseases. Hepatology. 53: 1752–1763.
Dayal S., Wilson K. M., Leo L., Arning E., Bottiglieri T., Lentz S. R. (2006). Enhanced
susceptibility to arterial thrombosis in a murine model of hyperhomocysteinemia. Blood. 108:
2237– 2243.
De Beer D., Stoodley P., Roe F., Lewandowski Z. (1994). Effects of biofilm structure on
oxygen distribution and mass transport. Biotechnol Bioeng. 43: 1131-8.
Deanfield J., Donald A., Ferri C., Giannattasio C., Halcox J., Halligan S., Lerman A.,
Mancia G., Oliver J.J., Pessina A. (2005). Working Group on Endothelin and Endothelial
Factors of the European Society of Hypertension: Endothelial function and dysfunction. J
Hypertens. 23: 7–17.
Deans G., Ritchie G. (1987). Antibacterial properties of plant essential oils. Int J Food
Microbio. 5:165–180.
Den Heijer M., Lewington S., Clarke R. (2005). Homocysteine, MTHFR and risk of
venous thrombosis: a meta-analysis of published epidemiological studies. J Thromb Haemost.
3: 292–9.
Derouiche A., Cherki M., Drissi A., Bamou Y., El Messal M., Idrissi-Oudghiri A. (2005).
Nutritional intervention study with Argan oil in man: effects on lipids and apolipoproteins.
Ann Nutr Metab. 49:196–201.
Dhillon B., Badiwala M.V., Maitland A., Rao V., Li S.H., Verma S. (2003).
Tetrahydrobiopterin attenuates homocysteine induced endothelial dysfunction. Mol Cell
Biochem. 247: 223-27.
Dobsn A., Filipiak B., Kuulasmaa K. (1996). Relations of changes in coronary disease rates
and changes in risk factor levels: methodological issues and a practical example. Am J
Epidemiol. 143: 1025–34.
Domagala T .B., Lacinski M ., Trzeciak W. H., Mackness B., Mackness M. I.,
Jakubowski H. (2006). The correlation of homocysteine-thiolactonase activity of the
paraoxonase (PON1) protein with coronary heart disease status. Cell Mol Biol . 52: 4–10.
Draelos Z. D. (2010) .The biofilm: skin health and beauty in a contaminated world. J Cosmet
Dermatol. 9: 167-168.
Drissi A., Bennani H., Giton F., Charrouf Z., Fiet J., Adlouni A. (2006). Tocopherols and
saponins derived from Argania spinosa exert an antiproliferative effect on human prostate
cancer. Cancer Invest. 24: 588-592.
Drissi A., Girona J., Cherki M., Godas G., Derouiche A., El Messal M., Saile R., Kettani
A., Sola R., Masana L., Adlouni A. (2004). Evidence of hypolipemiant and antioxidant
properties of Argan oil derived from the Argan tree (Argania spinosa). Clin Nutr. 23:1159-
1166.

References
93
Duffy S.J., Vita J. A. (2003). Effects of phenolics on vascular endothelial function. Curr
Opin Lipidol. 14: 21–7.
Durand P., Lussier-Cacan S., Blache D. (1997). Acute methionine load-induced
hyperhomocysteinemia enhances platelet aggregation, thromboxane biosynthesis, and
macrophage-derived tissue factor activity in rats. Faseb J. 11: 1157–1168.
El Babili F., Bouajila J., Fouraste I., Valentin A., Mauret S., Moulis C. (2010). Chemical
study, antimalarial and antioxidant activities, and cytotoxicity to human breast cancer cells
(MCF7) of Argania spinosa. Phytomedicine. 17: 157-160.
Elhawary N. A., Hewedi D., Arab A., Teama S., Shaibah H., Tayeb M. T., Bogari N.
(2013). The MTHFR 677T allele may influence the severity and biochemical risk factors of
Alzheimer’s disease in an Egyptian population. Disease markers. 35: 439-446.
Epstein S.E., Zhou Y. F., Zhu J. (1999). Infection and atherosclerosis emerging mechanistic
paradigms. Circulation. 100: e20-8.
Eren E., Ellidag H .Y., Aydin O., Yılmaz N. (2014). Homocysteine, Paraoxonase-1 and
Vascular Endothelial Dysfunction: Omnibus viis Romam Pervenitur. J Clin Diagn Res. 8:
CE01-CE04.
Eren E., Yilmaz N., Aydin O. (2013). Functionally Defective High-Density Lipoprotein and
Paraoxonase: A Couple for Endothelial Dysfunction in Atherosclerosis. Cholesterol. 2013:
792090.
Estruch M., Bancells C., Beloki L., Sanchez-Quesada J. L., Ordonez-Llanos J., Benitez
S. (2013). CD14 and TLR4 mediate cytokine release promoted by electronegative LDL in
monocytes. Atherosclerosis. 229: 356–362.
Fabricant C., Fabricant J., Litrenta M., Minick C. (1978). Virus-induced atherosclerosis.
J Exp Med. 148: 1, 335–340.
Faeh D., Chiolero A., Paccaud F. (2006). Homocysteine as a risk factor for cardiovascular
disease: should we (still) worry about it?. Swiss Med Wkly.136:745–56.
Fassbender K., Orell M., Hennerici M. (1999). Plasma homocysteine concentrations in
cerebrovascular disease. Stroke. 30: 2224-38.
Fernández-Checa J., Kaplowitz N., Garcia-Ruiz C., Colell A., Miranda M., Mai M.,
Ardite E., Morales A. (1997). GSH transport inmitochondria: defense against TNF-induced
oxidative stress and alcohol-induced defect. American J Physiology-Gastrointestinal and
Liver Physiology . 273:G7-G17.
Field T. R., White A., Elborn J. S., Tunney M. M. (2005). Effect of oxygen limitation on
the in vitro antimicrobial susceptibility of clinical isolates of Pseudomonas aeruginosa grown
planktonically and as biofilms. Eur J Clin Microbiol Infect Dis. 24:677–87.
Floreani M., Napoli E., Quintieri L., Palatini P. (2003). Oral administration of
transresveratrol to guinea pigs increases cardiac DT-diaphorase and catalase activities, and
protects isolated atria from menadione toxicity. Life Sci. 24: 2741–50.

References
94
Frelut M. L., Emery-Fillon N., Guilland J. C., Dao H. H., de Courcy G. P. (2006).
Alanine amino transferase concentrations are linked to folate intakes and
methylenetetrahydrofolate reductase polymorphism in obese adolescent girls. J pediatric
gastroenterology and nutrition. 43: 234-239.
Froggatt J. W., Johnston J. L., Galetto D. W., Archer G. L. (1989). Antimicrobial
resistance in nosocomial isolates of Staphylococcus haemolyticus. Antimicrob. Agents
Chemother. 33:460–466.
Fu Y-F, Xiong Y, Guo Z. (2005). A reduction of endogenous asymmetric dimethylarginine
contributes to the effect of captopril on endothelial dysfunction induced by homocysteine in
rats. Eur J Pharmacol. 508: 175.
Galkina E., Ley K. (2009). Immune and inflammatory mechanisms of atherosclerosis. Annu
Rev Immunol. 27: 165-197.
Gallai V., Caso V., et al. (2001). Mild hyperhomocysteinemia: a possible risk factor for
cervical artery dissection. Stroke. 32: 714–718.
Ganguly P ., Alam S. F. ( 2015) . Role of homocysteine in the development of cardiovascular
disease. Nutr J. 14:6.
Garcia-Ruiz C., Fernández-Checa J.C. (2006). Mitochondrial glutathione: hepatocellular
survival–death switch. Journal of gastroenterology and hepatology. 21(s3).
Gaull G., Starman J.A., Schaffner N. (1974). Homocystinuria due to cystathionine synthase
deficiency: enzymatic and ultrastructural studies. The Journal of pediatrics. 8 : 381-390.
Gaustadnes M., Kluijtmans L.A., Jensen O.K., et al. (1998). Detection of a novel deletion
in the cystathionine beta-synthase (CβS) gene using an improved genomic DNA based
method. FEBS Lett. 431:175-179.
Gisslinger H., Rodeghiero F., Ruggeri M., Heis-Vahidi Fard N., Mannhalter C.,
Papagiannopoulos M. (1999). Homocysteine levelsin polycythaemia vera and essential
thrombocythaemia. Br J Haematol. 105: 551–5.
Góth L., Rass P., Páy A. (2004). Catalase enzyme mutations and their association with
diseases. Mol Diagn. 8: 141-9.
Guillaume D., Charrouf Z. (2005). Saponines et métabolites secondaires de l'arganier
(Argania spinosa). Cahiers Agricultures. 14: 509-516.
Gulen D., Kaya A. D., Aydin M., Tanriverdi Y. (2013). Urinary tract infections caused by
Streptococcus uberis: A pathogen of bovine mastitis report of seven cases. Afri J Microbiol
Res. 7: 3908–12.
Gupta A. (2015). Biofilm Quantification and Comparative Analysis of MIC (Minimum
Inhibitory Concentration) & MBIC (Minimum Biofilm Inhibitory Concentration) Value for
Different Antibiotics against E. coli. Int J Curr Microbiol App Sci. 4: 198-224.

References
95
Gupta P., Sarkar S., Das B., Bhattacharjee S., Tribedi P. (2016). Biofilm, pathogenesis
and prevention—a journey to break the wall: a review. Archives of microbiology. 198: 1-15.
Guthikonda S. M. D., Haynes W. G. (2006). Homocysteine: Role and Implications in
Atherosclerosis. Current Atherosclerosis Reports. 8: 100–106.
Habash M., Reid G. (1999). Microbial biofilms: their development and significance for
medical device-related infections. J Clin Pharmacol. 39: 887-899.
Haenni M., Galofaro L., Ythier M., Giddey M., Majcherczyk P., Moreillon P., et al. (2010). Penicillin-binding protein gene alterations in Streptococcus uberis isolates presenting
decreased susceptibility to penicillin. Antimicrob Agents Chemother. 54:1140–5.
Hak A. E., Polderman K.H., Westendorp I. C., Jakobs C., Hofman A., Witteman J.
C., Stehouwer C. D. (2000). Increased plasma homocysteine after menopause.
Atherosclerosis. 149: 163-8.
Haka A. S., Singh R. K., Grosheva I., Hoffner H., Capetillo-Zarate E., Chin H. F.,
Anandasabapathy N., Maxfield F.R. (2015). Monocyte-Derived Dendritic Cells
Upregulate Extracellular Catabolism of Aggregated Low-Density Lipoprotein on Maturation,
Leading to Foam Cell Formation. Arterioscler Thromb Vasc Biol. 35: 2092–2103.
Hall-Stoodley L., Costerton J.W., Stoodley P. (2004). Bacterial biofilms: from the natural
environment to infectious diseases. Nat Rev Microbiol . 2: 95–108.
Hall‐Stoodley L., Stoodley P. (2009). Evolving concepts in biofilm infections. Cell
Microbiol. 11: 1034–1043.
Hamada S., Slade H. D. (1980). Biology, immunology, and cariogenicity of Streptococcus
mutans. Microbiol Rev. 44: 331-84.
Hamelet J., Demuth K., Dairou J., Ledru A., Paul J.L., Dupret J.M., et al. (2007).
Effects of catechin on homocysteine metabolism in hyperhomocysteinemic mice. Biochem
Biophys Res Commun. 355: 221–7.
Hankey G. J., Eikelboom J. W. (1999). Homocysteine and vascular disease. Lancet. 354:
407–13.
Hayashi C., Gudino C.V., Gibson F.C., Genco C. A. (2010). Review: pathogen-induced
inflammation at sites distant from oral infection: bacterial persistence and induction of cell-
specific innate immune inflammatory pathways. Mol Oral Microbiol. 25: 305–316.
He L., Zeng H., Li F., Feng J., Liu S., Liu J. (2010). Homocysteine impairs coronary artery
endothelial function by inhibiting tetrahydrobiopterin in patients with hyperhomocysteinemia.
Am J Physiol Endocrinol Metab. 299: 1061-5.
Ho Y. S., Xiong Y., Ma W., Spector A., Ho D. S. (2004). Mice lacking catalase develop
normally but show differential sensitivity to oxidant tissue injury. J Biol Chem. 279: 32804-
12.

References
96
Hoffman M. (2011). Hypothesis: hyperhomocysteinemia is an indicator of oxidant stress.
Med Hypotheses 77:1088–1093.
Hofmann M. A., Lalla E., Lu Y., Gleason M. R., Wolf B. M., Tanji N., Ferran L. J. Jr.,
Kohl B., Rao V., Kisiel W., Stern D. M., Schmidt A.M. (2001). Hyperhomocysteinemia
enhances vascular inflammation and accelerates atherosclerosis in a murine model. J Clin
Invest. 107: 675–83.
Hogg N. (1999). The effect of cysteine on the auto-oxidation of homocysteine. Free Radic
Biol Med . 27: 28- 33.
Holven K. B., Aukrust P., Retterstol K., Otterdal K., Bjerkeli V., Ose L., Nenseter M. S.,
Halvorsen B. (2008). The antiatherogenic function of HDL is impaired in
hyperhomocysteinemic subjects. J Nutrition . 138: 2070–2075.
Houston P. E., Rana S., Sekhsaria S., Perlin E., Kim K. S., Castro O.L.( 1997).
Homocysteine in sickle cell disease: relationship to stroke. Am J Med. 103:192–6.
Huang R.F., HsuY.C., Lin H.L., Yang F.L. (2001). Folate d.epletion and elevated plasma
homocysteine promote oxidative stress in rat livers. J Nutr. 131: 33–38.
Jadhav S., Shah R., Bhave M., Ea P. (2013). Inhibitory activity of yarrow essential oil on
Listeria planktonic cells and biofilms. Food Control. 29:125-30.
Jain R., Douglas R. (2014). When and how should we treat biofilms in chronic sinusitis? .
Curr Opin Otolaryngol Head Neck Surg. 22: 16–21.
Jakubowski H. (2008).The pathophysiological hypothesis of homocysteine thiolactone-
mediated vascular disease. J Physiol Pharmacol. 59:155–67.
James G.A., Swogger E., Wolcott R. (2008). Biofilms in chronic wounds. Wound Repair
Regen. 16:37–44.
Jeanette S.J., Alex K.H., David J.R., Adviye E. (2005). Oxidative stress and the use of
antioxidants in diabete: linking basic science to clinical practice .Cardiovascular
Diabetology. 4:5
Ji C., Kaplowitz N. (2004). Hyperhomocysteinemia, endoplasmic reticulum stress, and
alcoholic liver injury. World J Gastroenterol. 10: 1699–1708.
Jing M., Rech L., Wu Y., Goltz D.,Taylor C. G., House J. D. (2014). Effects of zinc
deficiency and zinc supplementation on homocysteine levels and related enzyme expression
in rats. J Trace Elem Med Biol. 30: 77-82.
José M. G. (2014). Oxidative Stress and Cell Death in Cardiovascular Disease: A Post-
Genomic Appraisal. Post-Genomic Cardiology, second edition. Chapter 14. 978-0-12-
404599-6.
Jourdheuil-Rahmani D., Rolland P.H., Rosset E., Branchereau A., Garcon D. (1997).
Homocysteine induces synthesis of a serine elastase in arterial smooth muscle cells from
multi-organ donors. Cardiovasc Res. 34: 597–602.

References
97
Kamal A. A., Thanaa M. A. (2009). Oxidative markers, nitric oxide and homocysteine
alteration in hypercholesterolimic rats: role of atorvastatine and cinnamon. Int J Clin Exp
Med. 2: 254- 265.
Kanani P. M., Sinkey C. A., Browning R. L., Allaman M., Knapp H. R., Haynes W. G. (1999). Role of oxidant stress in endothelial dysfunction produced by experimental
hyperhomocysteinemia in humans. Circulation. 100: 1161–8.
Kasdallah-Grissa A., Mornagui B., Aouani E., Hammami M., El May M., Gharbi N.,
Kamoun A., El-Fazaa S. (2007). Resveratrol, a red wine polyphenol, attenuates ethanol-
induced oxidative stress in rat liver. Life Sci. 80: 1033–1039.
Kaufman R. J. (2002). Orchestrating the unfolded protein response in health and disease. J
Clin Invest. 110: 1389–98.
Kechairi R. (2009). Contribution à l'étude écologique de l'arganier Argania spinosa (L)
skeels. dans la région de Tindouf (Algérie) (Doctoral dissertation).
Kesavalu L, Lucas AR, Verma RK, et al. (2012). Increased atherogenesis during
Streptococcus mutans infection in ApoE-null mice. J Dent Res. 9: 255–60.
Ketterer B. (1998). Glutathione-stransferase and prevention of cellular free radical damage.
Free radical Research. 28: 647-658.
Kevin L. S., Anne L. S. (2012). Homocysteine Imbalance: a Pathological Metabolic Marker :
a Pathological Metabolic Marker. American Society for Nutrition. 3: 755–762.
Khan M., Zahin M., Hasan S., Husain F., Ahmad I. (2009). Inhibition of quorum sensing
regulated bacterial functions by plant essential oils with special reference to clove oil. Lett
Appl Microbiol. 49: 354- 9.
Khallouki F., Spiegelhalder B., Bartsch H., Owen R. W. (2005). Secondary metabolites of
the argan tree (Morocco) may have disease prevention properties. African J Biotechnology. 4:
381-388.
Kietadisorn R., Juni R. P., Moens A.L. (2012). Tackling endothelial dysfunction by
modulating NOS uncoupling: new insights into its pathogenesis and therapeutic possibilities
Am J Physiol Endocrinol Metab. 302: E481–E495.
Killip T., Ann Payne M. (1960). High serum transaminase activity in heart disease:
circulatory failure and hepatic necrosis. Circulation. 21: 646–660.
Kim H., Park H. (2013). Ginger extract inhibits biofilm formation by Pseudomonas
aeruginosa PA14. Plos One. 8: E76106.
Kirac D., Negis Y., Ozer N. K. (2013). Vitamin E attenuates homocysteine and cholesterol
induced damage in rat aorta. Cardiovascular Pathology. 22: 465–472.
Kirbag S., Zengin F., Kursat M. (2009). Antimicrobial Activities of Extracts of some
Plants. Pakistan J Botany . 41: 2067-2070.

References
98
Knowles J. W., Reddick R. L., Jennette J. C. (2000). Enhanced atherosclerosis and kidney
dysfunction in eNOS(-/-) Apoe(-/-) mice are ameliorated by enalapril treatment. J Clin Invest
.105: 451-458.
Koh K., Tham F. (2011). Screening of traditional chinese medicinal plants for quorum-
sensing inhibitors activity. J Microbiol Immunol Infect. 44:144-8.
Kokare C.R., Chakraborty S., Khopade A.N., Mahadik K.R. (2009)
Biofilm: importance and applications. Indian J Biotechnol
8:159–168.
Koyama T., Matsui D., Kuriyama N., Ozaki E., Tanaka K., Oze I., Hamajima N., Wakai
K., Okada R ., Arisawa K., Mikami H., et al. (2015). Genetic variants of SLC17A1 are
associated with cholesterol homeostasis and hyperhomocysteinaemia in Japanese men. Scie
Rep. 5: 15888.
Kumar L., Chhibber S., Harjai K. (2013). Zingerone inhibit biofilm formation and improve
antibiofilm efficacy of ciprofloxacin against Pseudomonas aeruginosa PAO1. Fitoterapia. 90:
73- 8.
Kumar S .V. Neelam P. H ., Rajani. M. (2006). Search for antibacterial and antifungal
agents from selected Indian medicinal plants. J Ethnopharmaco. 107: 182–188.
kumar S.V., Saritha G., Fareedullah M. d. (2010).role of antioxidants and oxidative stress
in cardiovascular diseases. An Biol Res. 1:158-173.
Kuramitsu H. K., Qi M., Kang I. C., Chen W. (2001). Role for periodontal bacteria in
cardiovascular diseases. Ann Periodontol. 6: 41-7.
Heydrick S. J., Weiss N., Thomas S. R., Cap A. P., Pimentel D. R., Loscalzo J., Keaney
J. F. (2004). L-Homocysteine and L-homocystine stereospecifically induce endothelial nitric
oxide synthase-dependent lipid peroxidation in endothelial cells. Free Radical Biology and
Medicine. 36: 632-640.
Lai W. K. C., Kan M. Y. (2015). Homocysteine-Induced Endothelial Dysfunction. Ann
Nutr Metab. 67: 1–12.
Lamda S. (2000). Effets d’interaction des vitamines B6, B9 et B12, chez les rats
normotendus et hypertendus soumis a un regime riche en methionine associee ou non au
cholesterol. Thèse de Magister en Biologie et physiologie cellulaire animale. Université
Constantine 1: 1-119.
Lee M. E., Wang H. (1999). Homocysteine and hypomethylation. A novel link to vascular
disease. Trends Cardiovasc Med. 9: 49–54.
Lee Y. J., Kang S. W., Yang J. I., Choi Y. M., Sheen D., Lee E. B., Choi S. W., Song Y.
W. (2002). Coagulation parameters and plasma total homocysteine levels in Behcet’s disease.
Thrombosis Research. 106: 19-24.

References
99
Lehmann M., Regland B., Blennow K., Gottfries C. G. (2003). Vitamin B12- B6-folate
treatment improves blood-brain barrier function in patients with hyperhomocysteinaemia and
mild cognitive impairment. Dement Geriatr Cogn Disord. 16: 145-50.
Leishman S.J., Doand H.L., Ford P.J. (2010).Cardiovascular disease and the role of oral
bacteria. J Oral Microbiol . 2: 5781.
Lentez S. R. (2005). Mechanisms of homocysteine-induced atherothrombosis . J Thromb
Haemost. 3: 1646–54.
Lentz S.R., Sadler J. E. (1993). Homocysteine inhibits von Willebrand factor processing
and secretion by preventing transport from the endoplasmic reticulum. Blood. 81: 683–9.
Li Y. H., and Tian X. (2012). Quorum Sensing and Bacterial Social Interactions in Biofilms.
Sonsors (basel. )12: 2519-2538.
Li Z., Vance D.E. (2008). Phosphatidylcholine and choline homeostasis. J Lipid Res. 49:
1187–94.
Lin P. T., Lee B. J., Chang H. H., Cheng C. H., Tsai A. J., Huang Y. C. (2006). Low-dose
folic acid supplementation reduces homocysteine concentration in hyperhomocysteinemic
coronary artery disease patients.Nutr Res. 26: 460–6.
Liselotte M. S. E. (2003). Hyperhomocysteinemia due to folate deficiency. Impact on
haemostasis and vascular biology. Doctorat Thesis, University of Aarhus, Denmark.
Liu X. J., Qu D., He F., Lu Q., Wang J., Cai D.L. (2007). Effect of lycopene on the
vascular endothelial function and expression of inflammatory agents in
hyperhomocysteinemic rats. Asia Pac J Clin Nutr. 16: 244-248.
Lotfi N., Chahboun N., El Hartiti H., Kabouche Z., El M’Rabet M., Berrabeh M.,
Touzani R., Ouhssine M., Oudda H. (2015). Study of the antibacterial effect of Argan oil
from Bechar region of Algeria on hospital resistant strains. J Mater Environ Sci. 6: 2476-
2482.
Lowy F. D. (1998). Stapylococcus aureus infections. New Eng J Med. 339: 520-535.
Lu S. C. (2009). Regulation of glutathione synthesis. Mol Aspects Med. 30: 42–59.
Luo F., Liu X., Wang S., Chen H. (2006). Effect of homocysteine on platelets activation
induced by collagen. Nutrition. 22: 69–75.
Ma B., Forney L. J., Ravel J. (2012). Vaginal microbiome: rethinking health and disease.
Annu Rev Microbiol. 66: 371e89.
Mack D., Rohde H., Harris L. G., Davies A. P., Horstkotte M. A., Knobloch J. K. (2006).
Biofilm formation in medical device-related infection. Int J Artif Organs. 29: 343–359.
Mack D., Fischer W., Krokotsch A., Leopold K., Hartmann R., Egge H., Laufs R. (1996). The intercellular adhesin involved in biofilm accumulation of Staphylococcus

References
100
epidermidis is a linear beta-1,6-linked glucosaminoglycan: purification and structural
analysis. J Bacteriol. 178:175–183.
Mackness M.I., Abbott C.A., Arrol S. (1993). The role of high-density lipoprotein and lipid
soluble antioxidant vitamins in inhibiting low density lipoprotein oxidation. J Biochem. 294:
829-34.
Madeira S.V., Auger C., Anselm E., Chataigneau M., Chataigneau T., Soares de Moura
R. (2009). eNOS Activation Induced by a Polyphenol-Rich Grape Skin Extract in Porcine
Coronary Arteries. J Vasc Res. 5: 406–16.
Majors A. K., Sengupta S., Jacobsen D.W., Pyeritz R. E. (2002). Upregulation of smooth
muscle cell collagen production by homocysteine-insight into the pathogenesis of
homocystinuria. Mol Genet Metab. 76: 92–9.
Malinowska J., Kolodziejczyk J., Olas B. (2012). The disturbance of hemostasis induced by
hyperhomocysteinemia; the role of anti-oxidants. Acta Biochim Pol. 59: 185-94.
Mangge H., Becker K., Fuchs D., Gostner J. M. (2014). Antioxidants, inflammation and
cardiovascular disease. World J cardiology. 6: 462.
Mansoor M. A., Bergmark C., Svardal A. M., Lonning P. E., Ueland P. M. (1995). Redox
status and protein binding of plasma homocysteine and other aminothiols in patients with
early-onset peripheral vascular disease. Homocysteine and peripheral vascular disease.
Arterioscler Thromb Vasc Biol. 15: 232–40.
Margoliash E., Novogrodsky A. (1958). A study of the inhibition of catalase by 3-amino-
1:2:4:-triazole. Biochem J. 68: 468-75.
Marinou K., Antoniades C., Tousoulis D., Pitsavos C., Goumas G., Stefanadis C. (2005).
Homocysteine: a risk factor for coronary artery disease?. Hellenic J Cardiol. 46: 59 –67.
Maron B. A., Michel T. (2012). Subcellular localization of oxidants and redox modulation
of endothelial nitric oxide synthase. Circ J. 76: 2497-512.
Martinez L. O., Jacquet S., Terce F., Collet X., Perret B., Barbaras R. (2004). New
insight on the molecular mechanisms of high density lipoprotein: cellular interactions. Cell
Mol Life Sci. 61-2343.
Masella R., Giovannini C., Vari R., Di Benedetto R., Coni E., Volpe R. (2001). Effect of
dietary virgin olive oil phenols on low density lipoprotein oxidation in hyperlipidemic
patients. Lipids. 36: 1195–202.
Mato J. M., Lu S. C.(2005). Homocysteine, the bad thiol. Hepatology. 41: 976–979.
Matsumoto-Nakano M., Tsuji M., Inagaki S. (2009). Contribution of cell surface protein
antigen c of streptococcus mutans to platelet aggregation. Oral Microbiol Immunol. 24: 427 –
30.

References
101
Matté C., Mackedanz V., Stefanello F. M., Scherer E. B., Andreazza A. C., Zanotto C.,
et al. (2009). Chronic hyperhomocysteinemia alters antioxidant defenses and increases DNA
damage in brain and blood of rats: protective effect of folic acid. Neurochem Int. 54: 7–13.
Matté C., Stefanello F. M., Mackedanz V., Carolina D., Marcelo L., Carlos L. S. (2009).
Homocysteine induces oxidative stress, inflammatory infiltration, fibrosis and reduces
glycogen/glycoprotein content in liver of rats. J Devl Neuroscience. 27: 337–344.
McCully K. S. (1969).Vascular pathology of homocysteinemia: Implications for the
pathogenesis of arteriosclerosis. Am J Pathol. 56:111–128.
McCully K. S. (1996). Homocysteine and vascular disease. Nature Med. 2(4):386–9.
McCully K. S., Carvalho, A. C. (1987). Homocysteine thiolactone, N-homocysteine
thiolactonyl retinamide, and platelet aggregation. Research communications in chemical
pathology and pharmacology. 56: 349-360.
McDowell I.F., Lang D. ( 2000). Homocysteine and endothelial dysfunction: a link with
cardiovascular disease. J Nutr . 130:369S– 372S.
Mehta G., Kumari. S. (1997). Multi-resistant Staphylococcus haemolyticus in a neonatal
unit in New Delhi. Ann Trop Paediatr. 17:15–20.
Mehta K., Balaraman R., Amin A.H., Bafna P.A., Gulati O.D. (2003). Effect of fruits of
Moringa oleifera on the lipid profile of normal and hypercholesterolaemic rabbits. J
Ethnopharmacol. 86: 191.
Meng B., Weina G., Jingyu W., Jijun Y., Jianquan W., Lingling P., Changjiang G. (2013). Quercetin reduces serum homocysteine level in rats fed a methionine-enriched Diet.
Nutrition. 29: 661–666.
Merritt J. H., Kadouri D. E., O'Toole G. A .(2005). Growing and analyzing static biofilms.
Curr Protoc Microbiol .Chapter 1: Unit 1B 1.
Miller J. W. (2013). Homocysteine. Encyclopedia of Human Nutrition.
http://dx.doi.org/10.1016/B978-0-12-375083-9.00144-6.
Milstone D. S., Ilyama M., Chen M., O’Donnell P., Davis V. M., Plutzky J., Brown J. D.,
Haldar S. M., Siu A., Lau A. C., et al. ( 2015). Differential role of an NF-_B
transcriptional response element in endothelial versus intimal cell VCAM-1 expression. Circ
Res. 117: 166–177.
Milton N.G.N. (1999). Amyloid-ß binds catalase with high affinity and inhibits hydrogen
peroxide breakdown. Biochem J. 344: 293-6.
Milton N.G.N. (2008). Homocysteine inhibits hydrogen peroxide breakdown by catalase.
Open Enzyme Inhibition J. 1: 34–41.
Moat S. J., Bonham J. R., Allen J. C., Powers H. J., McDowel I. F. (1999). Decreased
circulating plasma lipids in patients with homocystinuria. J Inherit Metab Dis. 22: 243–246.

References
102
Moncada S., Palmer R. M., Higgs E. A. (1991). Nitric oxide: physiology, pathophysiology,
and pharmacology. Pharmacol Rev. 43: 109-142.
Msanda F., El Aboudi A., Peltier J. P. (2005). Biodiversité et biogeography de l’arganeraie
marocaine. Cah Agric. 14: 357 -64.
Mudd S.H. (2011). Hypermethioninemias of genetic and non- genetic origin: a review. J Med
GenetC SeminMed Genet. 157: 3–32.
Mudd S. H., Levy H. L., Skovby F. (1995). Disorders of transsulfuration in: The metabolic
and molecular basis of inherited disease. McGraw-Hill Publishing. 1279– 1327.
Musthafa K., Ravi A., Annapoorani A., Packiavathy I., Pandian S. (2010). Evaluation of
anti-quorum-sensing activity of edible plants and fruits through inhibition of the N-acyl-
homoserine lactone system in Chromobacterium violaceum and Pseudomonas aeruginosa.
Chemotherapy. 56: 333- 9.
Nagata E., de Toledo A., Oho T. (2011). Invasion of human aortic endothelial cells by oral
viridans group streptococci and induction of inflammatory cytokine production. Mol Oral
Microbiol . 26: 78 –88.
Naggar M., Mhirit O. (2006). L’arganeraie: un parcours typique des zones arides et semi-
arides marocaines. Science et changements planétaires/Sécheresse. 17 : 314-317.
Nakano K., Fujita K., Nishimura K., Nomura R., Ooshima T. (2005). Contribution of
biofilm regulatory protein A of Streptococcus mutans, to systemic virulence. Microbes Infect.
7: 1246-55.
Nakano K., Inaba H., Nomura R. (2006). Detection of cariogenic Streptococcus mutans in
extirpated heart valve and atheromatous plaque specimens. J Clin Microbiol. 44: 3313–7.
Nakano K., Nemoto H., Nomura R. (2009). Detection of oral bacteria in cardiovascular
specimens. Oral Microbiol Immunol . 24: 64–8.
Namekata K., Enokido Y., Ishii I., Nagai Y., Harada T., Kimura H. ( 2004). Abnormal
lipid metabolism in cystathionine β synthase-deficient mice, an animal model for
hyperhomocysteinemia. J Biol Chem. 279:52961–9.
Nappo F., De Rosa N., Marfella R., De Lucia D., Ingrosso D., Perna A. F. (1999).
Impairment of endothelial functions by acute hyperhomocysteinemia and reversal by
antioxidant vitamins. JAMA . 281: 2113–8.
Narmatha M. P., Karthikumar S., Rajendra N.N., Palanisamy A., Sundara G.,
Murugesh N. (2015). Effect of Folic Acid on Serum Homocysteine Levels in Patients with
Cardiovascular Diseases (CVD). Human Journals. 4: 143-158.
Navarro C. M., Montilla P. M., Martin A., Jimenez J., Utrilla P. M. (1993). Free radicals
scavenger and antihepatotoxic activity of Rosmarinus. Plant Med. 59: 312–314.
Nazzaro F., Fratianni F., Coppola R. (2013). Quorum sensing and phytochemicals. Int J
Mol Sci. 14:12607-19.

References
103
Necib Y., Bahi A., Zerizer S. (2013). Argan oil (Argania spinosa L) Provides protection
against mercuric chloride induced oxidative stress in rat Albinos Wistar. Int J Basic Appli
Scie. 2: 73-80.
Nichols W., Dorrington S., Slack M., Walmsley H. (1988). Inhibition of tobramycin
diffusion by binding to alginate. Antimicrob Agents Chemother. 32: 618-23.
Niu C., Afre S., Gilbert E. (2006). Subinhibitory concentrations of cinnamaldehyde interfere
with quorum sensing. Lett Appl Microbiol. 43: 489-94.
Nomura R., Nakano K., Naka S., et al. (2012). Identification and characterization of a
collagen-binding protein, cbm, in Streptococcus mutans. Mol Oral Microbiol. 27: 308 –23.
Nygard O., Nordrehaug J. E., Refsum H., Ueland P.M., Farstad M., Vollset S. E. (1997).
Plasma homocysteine levels and mortality in patients with coronary artery disease. N Engl J
Med. 337: 230–236.
Nygard O., Vollset S. E., Refsum H., Stensvold I., Tverdal A., Nordrehaug J. E.
(1995).Total plasma homocysteine and cardiovascular risk profile. The Hordaland
Homocysteine Study. 274: 1526–33.
Obeid R., Herrmann W. (2009). Homocysteine and lipids: S-Adenosyl methionine as a key
intermediate. FEBS Letters. 583: 1215–1225.
Olivero J., Pajaro N., Stashenko E. (2011). Antiquorum sensing activity of essential oils
isolated from different species of the genus Piper. Vitae. 18: 77-82.
Olsen. I .(2015). Biofilm-specific antibiotic tolerance and resistance. Eur J Clin Microbiol
Infect Dis. 34: 877-886.
Ott S. J., El Mokhtari N. E., Musfeldt M., Hellmig S., Freitag S., Rehman A. (2006).
Detection of Diverse Bacterial Signatures in Atherosclerotic lesions of patients with coronary
heart disease. Circulation. 113: 929- 937.
Ould safi M. (2014). Caractérisation et état sanitaire de l’Arganier de Tindouf. (Doctoral
dissertation).
Ozer J., Ratner M., Shaw M., Bailey W., Schomaker S. (2008). The current state of serum
biomarkers of hepatotoxicity. Toxicology. 245:194–205.
Pacher P., Beckman J.S., Liaudet L. (2007). Nitric oxide and peroxynitrite in health and
disease, Physiol Rev. 87: 315–424.
Packiavathy I., Agilandeswari P., Musthafa K., Pandian S., Ravi A. (2012) Antibiofilm
and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite
methyl eugenol against gram negative bacterial pathogens. Food Res Int. 45: 8592.
Packiavathy I., Priya S., Pandian S., Ravi A. (2014). Inhibition of biofilm development of
uropathogens by curcumin e an anti-quorum sensing agent from Curcuma longa. Food Chem.
148: 453- 60.

References
104
Palmer R. Jr., White D.C. (1997). Developmental biology of biofilms: implications for
treatment and control. Trends Microbiol . 5: 435– 440.
Papatheodorou L., Weiss N. (2007). Vascular oxidant stress and inflammation in
hyperhomocysteinemia. Antioxid Redox Signal. 9: 1941–1958.
Parra S., Alonso-Villaverde C., Coll B. (2007). Serum paraoxonase-1 activity and
concentration are influenced by human immunodeficiency virus infection. Atherosclerosis.
194:175–181.
Plazar N., Jurdana M. (2010). Hyperhomocysteinemia and the role of B vitamins in cancer.
Radiol Oncol. 44: 79-85.
Poddar R., Sivasubramanian N., DiBello P.M., Robinson K., Jacobsen D. W. (2001).
Homocysteine induces expression and secretion of monocyte chemoattractant protein-1 and
interleukin-8 in human aortic endothelial cells: implications for vascular disease. Circulation.
103: 2717–23.
Polavarapu R., Spitz D.R., Sim J. E., Follansbee M. H., Oberley L. W., Rahemtulla A.,
Nanji A. A. (1998). Increased lipid peroxidation and impaired antioxidant enzyme function is
associated with pathological liver injury in experimental alcoholic liver disease in rats fed
diets high in corn oil and fish oil. Hepatology. 27:1317–1323.
Poloni S., Blom H. J., Schwartz Ida V. D. (2015). Stearoyl-CoA Desaturase-1: Is It the
Link between Sulfur Amino Acids and Lipid Metabolism?. Biology. 4: 383-396.
Poloni S., Mendes R. H., Belló-Klein A., Schwartz I. V. (2012). Is lipid metabolism altered
in classical homocystinuria?. Mol Genet Metab. 106: 382–383.
Prigent-Combaret C., Vidal O., Dorel C., Lejeune P. (1999). Abiotic surface sensing and
biofilm development regulation of gene expression in Escherichia coli. J Bacteriol. 181:
5993-6002.
Putnam C. D., Arvai A. S., Bourne Y., Tainer J. A. (2000). Active and inhibited human
catalase structures: ligand and NADPH binding and catalytic mechanism. J Mol Biol.
296:295-309.
Qureshi S. S., Kgupta J., Upmanyu N. (2016). A review on hyperhomocystenemia and its
risk factors. Innovare J Med Science. 4: 2.
Rabin N., Zheng Y., Opoku-Temeng C., Du Y., Bonsu E., Sintim H. O. (2015). Biofilm
formation mechanisms and targets for developing antibiofilm agents. Future Med Chem. 7:
493–512.
Rabin N., Zheng1 Y., Opoku-Temeng C., du Y., Bonsu E., Sintim H. O. (2015). Agents
that inhibit bacterial biofilm formation. Future med Chem. 7: 647–71.
Raghuveer G., Sinkey C.A., Chenard C., Stumbo P., Haynes W.G. (2001). Effect of
vitamin E on resistance vessel endothelial dysfunction induced by methionine. Am J Cardiol.
88 : 285–90.

References
105
Rahmani M. (2005). Composition chimique de l'huile d'argane «vierge». Cahiers
Agricultures. 14: 461-465.
Ramakrishnan R., Sulochana S., Lakshmi K.N., Selvi S., Angayarkanni R. (2006).
Biochemistry of homocysteine in health and disease. Indian J Biochem Biophys. 43: 275–
283.
Rashid R. M., Salah W., Parada J. P. (2007). "Streptococcus milleri" aortic valve
endocarditis and hepatic abscess. J Med Microbiol. 56: 280-282.
Rasmussen K., Moller J. (2000). Total homocysteine measure-ment in clinical practice. Ann
Clin Biochem. 37: 627-648.
Rasolofo E.A., St-Gelais D., LaPointe G., Roy D. (2010). Molecular analysis of bacterial
population structure and dynamics during cold storage of untreated and treated milk. Int J
Food Microbiol. 138: 108–18.
Refsum H., Nurk E., Smith A.D., Ueland P.M., Gjesdal C.G., Bjelland I. (2006). The
Hordaland Homocysteine Study: a community-based study of homocysteine, its determinants,
and associations with disease. J Nutr. 136(6):1731S–40S.
Refsum H., Smith A.D., Ueland P.M. (2004). Facts and recommendations about total
homocysteine determinations: an expert opinion. Clin Chem . 50: 3–32.
Refsum H., Wesenberg F., Ueland P.M. (1991). Plasma homocysteine in children with
acute lymphoblastic leukemia: changes during a chemotherapeutic regimen including
methotrexate. Cancer Res. 51: 828–35.
Rifai N., Ridker P. M. (2001). High-sensitivity C-reactive protein: a novel and promising
marker of coronary heart disease. Clinical chemistry. 47: 403-411.
Robert K., Nehme J., Bourdon E., Pivert G., Friguet B., Delcayre C., Delabar J.M.,
Janel N. (2005). Cystathionine beta synthase deficiency promotes oxidative stress, fibrosis,
and steatosis in mice liver. Gastroenterology . 128: 1405–1415.
Roblin X., Pofelski J., Zarski J.P. (2007). Steatosis, chronic hepatitis virus C infection and
homocysteine. Gastroenterol Clin Biol. 31: 415–420.
Rochette L., Lorin J., Zeller M., Guilland J. C., Lorgis L., Cottin Y. (2013). Nitric oxide
synthase inhibition and oxidative stress in cardiovascular diseases: Possible therapeutic
targets? Pharmacol Ther. 140: 239-57.
Rolland P.H., Friggi A., Barlatier A. (1995). Hyperhomocysteinemia- induced vascular
damage in the minipig. Captopril-hydrochlorothiazide combination prevents elastic
alterations. Circulation. 91:1161–1174.
Rosenfeld M. E., Campbell L. A. (2011). Pathogens and atherosclerosis: update on the
potential contribution of multiple infectious organisms to the pathogenesis of atherosclerosis.
Thromb Haemost. 106: 858–867.

References
106
Ross R. (1993). The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature. 362:
801-809.
Ross R. (1999) .Atherosclerosis-an inflammatory disease. N Engl J Med. 340:115–126.
Sainani Gs., Talwalkar Pg., Wadia, Rs., Keshvani AA. (2008). Hyperhomocysteinemia and
its implications in atherosclerosis the Indian Scenario. Medicine update. 12–20.
Sakhri F.Z. (2014). Effects of Algerian olive oil on cardiovascular diseases and lipids status
in hyperhomocysteinemia treated mice. Thesis submitted for the degree of magister. Option:
biology and molecular physiology. University of Constantine 1.
Saleh A. A. S . (2015). Lipid profile and levels of homocysteine and total antioxidant capacity
in plasma of rats with experimental thyroid disorders. J Basic Applied Zoolog.72: 173-178.
Samane S., Noel J., Charrouf Z., Amarouch H., Haddad P. S. (2006). Insulin sensitizing
and Anti-proliferative Effects of Argania spinosa Seed Extracts. Evid Based Complement
Alternat Med. 3: 317-327.
Samy R. P., Gopalakrishnakone P. (2010). Therapeutic potential of plants as anti-
microbials for drug discovery. Evid Based Complement Alternat Med. 7: 283–94.
Santulli G., Iaccarino G. (2013). Pinpointing beta adrenergic receptor in ageing
pathophysiology: Victim or executioner? Evidence from crime scenes. Immun Ageing. 10(1):
10.
Sauls D. L., Lockhart E., Warren M. E., Lenkowski A., Wilhelm S. E., Hoffman M. (2006). Modification of fibrinogen by homocysteine thiolactone increases resistance to
fibrinolysis: a potential mechanism of the thrombotic tendency in hyperhomocysteinemia.
Biochemistry. 45: 2480–2487.
Schalinske K. L., Smazal, A. L. (2012). Homocysteine imbalance: a pathological metabolic
marker. Advances in Nutrition: An Inter Review J. 3: 755-762.
Singh P. K., Schaefer A.L., Parsek M. R. (2000). Quorum-sensing signals indicate that
cystic fibrosis lungs are infected with bacterial biofilms. Nature. 407: 762–764.
Škovierová H., Vidomanová E., Mahmood S., Sopková J., Drgová A., Červeňová T., et
al. (2016). The molecular and cellular effect of homocysteine metabolism imbalance on
human health. InterJ molecular scien. 17: 1733.
Sour S., Belarbi M., Sari N., Benammar C. H., Baghdad C.H., Visioli F.(2015). Argan oil
reduces, in rats, the high fat diet-induced metabolic effects of obesity. Nutrition Metabolism
& Cardiovascular Diseases . 1-6.
Spence J.D. (2007).Homocysteine-lowering therapy: a role in stroke prevention?. Lancet
Neurol. 6:830–8.
Steyers C. M., Miller F. J. (2014). Endothelial dysfunction in chronic inflammatory
diseases. Inter J Molecular Sci. 15 : 11324-11349.

References
107
Stewart P. (1996). Theoretical aspects of antibiotic diffusion into microbial biofilms.
Antimicrob Agents Chemother. 40: 2517-22.
Stipanuk M.H. (2004). Sulfur amino acid metabolism: pathways for production and removal
of homocysteine and cysteine. Annu Rev Nutr. 24: 539e77.
Stipanuk M.H., Dominy Jr .J.E., Lee J.I., Coloso R.M. (2006). Mammalian cysteine
metabolism: new insights into regulation of cysteine metabolism. J Nutr. 136:1652Se9S.
Stojković D., Petrović J., Soković M., Glamočlija J., Kukić‐Marković J., Petrović S. (2013). In situ antioxidant and antimicrobial activities of naturally occurring caffeic acid,
p‐coumaric acid and rutin, using food systems. J Scien Food Agricult. 9 : 3205-3208.
Streck E. L., Vieira P. S., Wannmacher C. M., Dutra-Filho C. S., Wajner M., Wyse A. T. (2003). In vitro effect of homocysteine on some parameters of oxidative stress in rat
hippocampus. Metab Brain Dis . 18:147–54.
Stuhlinger M. C., Oka R. K., Graf E. E., Schmolzer I., Upson B. M., Kapoor O., Szuba
A., Malinow M. R., Wascher T. C., Pachinger O., Cooke J. P. ( 2003). Endothelial
dysfunction induced by hyperhomocyst(e)inemia: role of asymmetric dimethylarginine.
Circulation. 108: 933–8.
Sun C. F., Haven T.R., Wu T.L., Tsao K.C., Wu J.T. (2002). Serum total homocysteine
with the rapid proliferation rate of tumor cells and decline upon cell death: A potential new
tumor marker. Clin Chim Acta. 321:55-62.
Sun F., Qu F., Ling Y., Mao P., Xia P., Chen H. (2013). Biofilm-associated infections:
antibiotic resistance and novel therapeutic strategies. Future Microbiol . 8: 877–86.
Sussman J. I., Baron E. J., Tenenbaum M. J., Kaplan M. H., Greenspan J., Facklam R.
R., Tyburski M. B., Goldman M. A., Kanzer B. F., Pizzarello R. A. (1986). Viridans
streptococcal endocarditis: clinical, microbiological, and echocardiographic correlations. J
Infect Dis. 154:597-603.
Tchantchou F. (2006). Homocysteine metabolism and various consequences of folate
deficiency. J Alzheimers Dis. 9: 421–7.
Townsend D. M., Tew K. D., Tapiero H. (2003). The importance of glutathione in human
disease. Biomedicine & Pharmacotherapy, 57: 145-155.
Tsai J. C., Perrella M. A., Yoshizumi M., Hsieh C. M., Haber E., Schlegel R. (1994).
Promotion of vascular smooth muscle cell growth by homocysteine: a link to atherosclerosis.
Proc Natl Acad Sci USA. 91: 6369–73.
Tselmin S., Rodionov R.N., Müller G., Bornstein S. et Julius U. (2013). Homocysteine in
lipoprotein apheresis patients − Retrospective data analysis in apheresis center of a university
hospital. Atherosclerosis Supplements. 14: 123-128.
Tyagi N., Sedoris K. C., Steed M., Ovechkin A. V., Moshal K. S., Tyagi S. C. (2005).
Mechanisms of homocysteine- induced oxidative stress. Am J Physiol Heart Circ Physiol.
289:H2649–H2656.

References
108
Undas A., Brozek J., Szczeklik A. (2005). Homocysteine and thrombosis: from basic
science to clinical evidence. Thromb Haemost. 94: 907–915.
Upadhyay A., Upadhyaya I., Kollanoor-Johny A., Venkitanarayanan K. (2014).
Combating pathogenic microorganisms using plant-derived antimicrobials:
a Minireview of the mechanistic basis. Biomed Res Int. 2014: 761741.
Vallance P. (2001). The asymmetrical dimethylarginine/ dimethylarginine
dimethylaminohydrolase pathway in the regulation of nitric oxide production. Clin Sci. 100:
159–60.
Veeranki S., Tyagi S C. (2013). Defective Homocysteine Metabolism: Potential
Implications for Skeletal Muscle Malfunction. Int. J. Mol. Sci. 14: 15074-15091.
Ventura P., Rosa M. C., Abbati G., Marchini S., Grandone E., Vergura P., Tremosini S.,
Zeneroli M. L. (2005). Hyperhomocysteinaemia in chronic liver diseases: role of disease
stage, vitamin status and methylenetetrahydrofolate reductase genetics. Liver Int. 25:49–56.
Viggianoa A., Viggianoa E., Mondaa M., Ingrossob D., Pernac A.F., Luca B.D. (2012).
Methionine-enriched diet decreases hippocampal antioxidant defences and impairs
spontaneous behaviour and long-termpotentiation in rats . brain research. 1 4 7 1: 6 6– 7 4.
Welch G. N., Upchurch G. R., Farivar R. S., Pigazzi A., Vu K., Brecher P. (1998).
Homocysteine-induced nitric oxide production in vascular smooth-muscle cells by NF-_B-
dependent transcriptional activation of Nos2. Proc Assoc Am Physicians. 110:22–31.
Werner E., Roe F., Bugnicourt A., Franklin M.J. (2004). Stratified growth in
Pseudomonas aeruginosa biofilms. Appl Environ Microbiol. 70: 6188-96.
Werstuck G. H., Lentz S. R., Dayal S., Hossain G. S., Sood S. K., Shi Y. Y., Zhou J.,
Maeda N., Krisans S. K., Malinow M. R. (2001). Homocysteine-induced endoplasmic
reticulum stress causes dysregulation of the cholesterol and triglyceride biosynthetic
pathways. J Clin Investig. 107: 1263–1273.
Whiley R. A., Beighton D. (1991). Emended descriptions and recognition of Streptococcus
constellatus, Streptococcus intermedius, and Streptococcus anginosus as distinct species. Int J
Syst Bacteriol . 41: 1-5.
Whiley R. A., Beighton D., Winstanley T. G., Fraser H. Y., Hardie J. M. (1992).
Streptococcus intermedius, Streptococcus constellatus, and Streptococcus anginosus (the
Streptococcus milleri group): association with different body sites and clinical infections. J
clin microbiol. 30: 243-244.
Wijekoon E. P., Brosnan M. E. (2007). Homocysteine metabolism in diabetes. Biochem Soc
Trans. 35:1175-9.
Willoughby S., Holmes A., Loscalzo J. (2002). Platelets and cardiovascular disease. Eur J
Cardiovasc Nurs. 1: 273–88.

References
109
Wimpenny J., Manz W., Szewzyk U. (2000). Heterogeneity in biofilms. FEMS Microbiol
Rev. 24: 661-71.
Wolcott R. D., Wolcott J. J., Palacio C. A., Rodriguez S. (2012). Possible role of bacterial
biofilm in the pathogenesis of atherosclerosis. J Bacteriol Parasitol. 3: 127.
Woo C. W. H., Prathapasinghe G.A., Siow Y.L. (2006). Hyperhomocysteinemia induces
liver injury in rat: Protective effect of folic acid supplementation. Biochim Biophys Acta.
1762: 656–665.
Woo C. W. H., Siow Y.L. K. (2008). Homocysteine induces monocyte chemoattractant
protein-1 expression in hepatocytes mediated via activator protein- 1 activation. J Biol Chem.
283: 1282–1292.
Wyse A. T. S., Zugno A. I., Streck E. L., Matte C., Calcagnotto T., Wannmacher C. M. (2002). Inhibition of Na+, K+-ATPase activity in hippocampus of rats subjected to acute
administration of homocysteine is prevented by vitamins E and C treatment. Neurochem Res.
27:1685–9.
Xu K. D., Stewart P. S., Xia F., Huang C. T., McFeters G. A. (1998). Spatial physiological
heterogeneity in Pseudomonas aeruginosa biofilm is determined by oxygen availability. Appl
Environ Microbiol. 64: 4035–9.
Yalçınkaya S., Ünlüçerçi Y., Giris M., Olgaç V., Dogru-Abbasoglu S., Uysal M. (2009)
Oxidative and nitrosative stress and apoptosis in the liver of rats fed on high methionine diet:
Protective effect of taurine. Nutrition. 25: 436–444.
Yamada H., Akahoshi N., Kamata S., Hagiya Y., Hishiki T., Nagahata Y., Tomomi
Matsuura. (2012). Methionine excess in diet induces acute lethal hepatitis in mice lacking
cystathionine γ-lyase, an animal model of cystathioninuria. Free Radical Biology & Medicine.
52:1716–1726.
Yang L., Haagensen J., Jelsbak L., Johansen H., Sternberg C., Høiby N. (2008). In situ
growth rates and biofilm development of Pseudomonas aeruginosa populations in chronic
lung infection. J Bacteriol. 190:2767-76.
Yilmaz N. (2012). Relationship between paraoxonase and homocysteine: crossroads of
oxidative diseases. Arch Med Sci. 8: 138-53.
Young D. S. (2001). Effects of disease on clinical Laboratory Tests, 4ème édition. AACC.
Zadoks R. N., Middleton J.R., McDougall S., Katholm J., Schukken Y. H. (2011).
Molecular epidemiology of mastitis pathogens of dairy cattle and comparative relevance to
humans. J Mammary Gland Biol Neoplasia. 16: 357–72.
Zadoks R. N. (2007). Sources and epidemiology of Streptococcus uberis, with special
emphasis on mastitis in dairy cattle. CAB Reviews. 2:30.
Zaki A., Shaaban M., Hashish N., Amer M., Lahloub M. (2013). Assessment of anti-
quorum sensing activity for some ornamental and medicinal plants native to Egypt. Sci
Pharm. 81: 251-8.

References
110
Zarrouk F., Othmani-Mecif K., Khedis L., Chaouad B., Ghoul A., Rezkallah N. (2010).
impact d’une hyperhomocysténémie sur les lipoproteines plasmatiques et sur la structure
hépatique du rat du sable psammomys obesus. Rev sunthése n °22.
Zerizer S, and Naimi D. (2004). Homocysteine: An independent risk factor in atherogenic
Process. Egyptian Pharmaceutical Journal 3: 110-114.
Zerizer S. (2006). Hyperhomocysteinemia, B vitamins and atherogenesis. Clinical and
experimental studies. Doctorat thesis. University of Constantine 1: 33-34.
Zerizer S., Naimi D., Benchaibi Y., Hamdi R. et Heikal O. (2008). Hyperhomocysteimia
and cardiovascular diseases in Algeria people. BULL NRC Egypt. 33: 481-493.
Zhang D., Chen Y., Xie X., Liu J., Wang Q., Kong W. (2012). Homocysteine activates
vascular smooth muscle cells by DNA demethylation of platelet-derived growth factor in
endothelial cells. J Mol Cell Cardiol. 53:487-96.
Zulli A., Buxton B. F., Doolan L., Liu J. J. (1998). Augmented effects of methionine and
cholesterol in decreasing the elastic lamina while thickening the aortic wall in the rat aorta.
Clin Sci (Lond). 95: 589– 93.
Zulli A., Hare D. L. (2009). High dietary methionine plus cholesterol stimulates early
atherosclerosis and late fibrous cap development which is associated with a decrease in
GRP78 positive plaque cells. Int J Exp Pathol. 90: 311–20.

الملخص بالعربية

الملخص
1
المقدمة
عندما الحظ "الشرايينتصلب " قترح فرضية إول من أهو McCully كان 9191في عام
ين يعانون من تراكم ذطفال ال أللدى ا طراف ألشرايين الدماغ و ا تجلط شرايين القلب التاجية و
.ستقالب الميثيونينإمتعلق ب جيني أنتيجة خط ،ينيبيلة الهوموسيست يسمى و ماأ ين في البوليالهوموسيست
ا زاد تركيزه عن ذإلكن ،الخالياو في بالزما الدميوجد ميني طبيعيأن هو حمض يالهوموسيستي
تدمير الجدار الداخلي المبطن إلىدي ؤمما ي ،ن يسبب مشاكل صحيةأمعدله الطبيعي في الدم يمكن
. وعية الدمويةأليعتبر عامل خطر بالنسبة للقلب و ا لك ذل ،صابة بتصلب الشرايينإللى درجة اإللشرايين
إلىالتي تودي حد العوامل أين هو يرتفاع مستويات الهوموسيستإن خدش بطانة الشرايين بسبب إ
ختالل في وظيفة بطانة إلي إدي بدوره ؤمما ي ، atherosclerosis بـو ما يعرف أين بداية تصلب الشراي
.النوبات القلبية و السكتتات الدماغية إليا ما يتسبب في نهاية المطاف ذو ه الشرايين
رتفاع إناتج عن يعتبر عامل مهم في تصلب الشرايين كسدي أجهاد التإلن اإف، لكذلى إضافة إلبا
غشية و ألكسدة دهون و بروتينات األى إدي ؤمما ي ،كسدةألنخفاض مضادات اإ، ينعكس بن يالهوموسيستي
. ة داخل العضويةبيلتهاإلنتاج العوامل اإزيادة
يعتبر عنصر شديد الذيلى ثيوالكتون و إن يتحول أن ييخرى، يمكن للهوموسيستأمن جهة
بروتيناتو التي توثر بدورها على NH-COضافية إمما يسمح بتشكيل معقدات بروتينات الالتفاعل مع
.نزيمات الجسمإو
صابة بأمراض إللتهابات المزمنة تزيد من خطر اإلن بعض اأخرى تشير الدراسات أمن جهة
.تصلب الشرايين فيو غير مباشرة أليات مباشرة آفبعض الجراثيم تساهم ب ،وعية الدمويةالقلب و األ
هي عبارة عن مستعمرات من الكائنات الحية الدقيقة بحيث تكون غشية الحيويةو األأ البيوفيلم
و ما يطلق عليه عديد أفراز نسيج خارجي إبو تتسم ، ملتصقة ببعضها البعض و مرتكزة على سطح
. extracellular polymeric substanceالسكاريديد الخارجي
لتهابات الشغاف المعدية، و إالكيسي، التليف مراض المزمنة مثل ألترتبط البيوفيلمات عادة مع عديد ا
جل تطوير مقاومة بكتيرية بالمضادات الحيوية أا الصدد وجب البحث من ذفي ه ،لخإ....الجروح المزمنة
.مواد الطبيعيةالمستخلصة من ال

الملخص
2
هاما في اكمصدر للمركبات الطبية، و التي لعبت دور النباتاتعتبرت أالعصور القديمة ذمن
ستخدام المكونات الفعالة في الطب إن إف نظمة الصحة العالمية مووفقا ل ،نسانإلالمحافظة على صحة ا
دوية ألمن ا % 08كثر من أن أكما ،من سكان العالم %08 لى عالج ما يقاربإدت أالتقليدي الشعبي
.صل نباتي طبيعيأالحديثة من
Arganiaثير الفعال و المفيد لنبتة طبية جزائرية ألى التعرف على التإه الدراسة ذتهدف ه
spinosa لى عائلةإو التي تنتمي Sapotaceae، ور شجرة ذستخدام المستخلص الخام لبإب سواء
ه النبتة ذن لهأثبتت أستنادا للدراسات السابقة و التي إا ذو ه ،ورذه البذو الزيت المستخلص من هأ رغانألا
لتهابات، مضادة لتكاثر الخاليا إلمخفضة للشحوم و الكوليسترول، مضادة ل: فوائد عالجية على غرار
.لخإ.....السرطانية
: ه الدراسة في ذهداف هأا المنطلق تتمحور ذو من ه
جرعات عالية من باستخداملك ذين في الدم و يالتحفيز على تكوين حالة فرط الهوموسيست
.الميثيونين لدى الفئران
ه الجرعات على وزن الفئرانذتقييم تأثير ه.
لبيوكيميائية مثل قياس ه الجرعات العالية من الميثيونين على بعض التحاليل اذتقييم تأثير ه
، الكوليسترول، البروتينات الدهنية(Triglycerides) ، الدهون الثالثية (Hcy) ن الهوموسيستيي
نزيمات الكبدية إل، ا(HDL-c)، البروتينات الدهنية عالية الكثافة (LDL-c)منخفضة الكثافة
(ALT, AST)مثل كسدةأللى مضادات اإضافة إل، با(Gluthatione ) نشاط الكاتاالز و
Catalase activity.
ورطي و ألالشريان ا ،لك على المقاطع النسيجية لكل من القلبذه الجرعات كذختبار تأثير هإ
.الكبد
ين و باقي التحاليل البيوكيميائيةيرغان على الهوموسيستألور شجرة اذتقييم المستخلص الخام لب.
القلب، الشريان األورطي ) رغان على المقاطع النسيجيةألور شجرة اذتقييم المستخلص الخام لب
(.و الكبد
نواع من البكتيريا و التي قد تسبب مشاكل أ 0ان ضد رغألتقييم المفعول المضاد للبيوفيلم لزيت ا
.وعية الدموية ألللقلب و ا
الطرق و الوسائل المستعملة
:المستخلصات النباتية

الملخص
3
( نرغاألستخدام نبتة طبية جزائرية معروفة في الطب الشعبي و هي اإه الدراسة تم ذخالل ه
(Argania spinosa رغان و الزيت المستخلص ألور شجرة اذالمستخلص الخام لبستعمل كل من أو لقد
.ورذه البذمن ه
:الحيوانات
تم الحصول عليها Mus musculusه الدراسة هي فئران تجارب من نوع ذالحيوانات المستعملة في ه
عطيت فترة للتأقلم مع ظروف أن أبعد ما التجارب فأجريت على الحيوانات أ ،من معهد الصيدلة بقسنطينة
.بل كل تجربةق ألمخبريالعمل
:التجارب
مجموعات بحيث تحتوي كل 4لى إنقضاء فترة التكيف، تم تقسيم الفئران إبعد : ولىألالتجربة ا
:و عوملت كما هو موضح في الجدول التالي ،حيوانات 7مجموعة على
(ميو 19لكل مجموعة لمدة 7: عدد الحيوانات ) .المعاملة التجريبية للفئران : 10الجدول
المجموعة التجريبية
المادة المستخدمة
الجرعة اليومية
(F)المجموعة
مسحوق الدقيق اللين رأف/ مغ 0.5
(M) المجموعة
+مسحوق الدقيق اللين
الميثيونين
+ فأر /مغ 8.0
كغ/مغ088
(MP) المجموعة
+اللين مسحوق الدقيق
+ الميثيونين
(م خ ب ش أ)
+فأر /مغ8.0
(+ميثيونين)كغ /مغ088
كغ /مغ908
(م خ ب ش ا )
(P ) المجموعة
+مسحوق الدقيق اللين
(م خ ب ش أ)
+فأر فا/مغ 8.0
كغ/مغ908
(م خ ب ش ا)

الملخص
4
OD * 1*1.525
GSH (nmol/mg of protein) =
13100*0.8*0.5.mg protein
.ور شجرة األرغانذالمستخلص الخام لب(: أم خ ب ش )
ن ييهوموسيستالخد عينات من الدم لقياس كل من أتم ،(سابيع أ 3) التجريبيةالمدة نقضاء إبعد
(Hcy) الدهون الثالثية ،(Triglycerides) الكوليسترول، البروتينات الدهنية منخفضة الكثافة ،
(LDL-c) البروتينات الدهنية عالية الكثافة ،(HDL-c)نزيمات الكبدية إل، ا(ALT, AST)، كما تم
لى تجميد إروطي و الكبد للدراسة النسيجية باإلضافة ألتخدير الفئران ألخذ عينات من القلب و الشريان ا
Catalase ))نشاط الكاتاالز و ( Gluthatione)كسدة مثل ألجزء من الكبد لمعايرة مضادات ا
activity في المسحوق الخلوي الكبدي.
:التحاليل البيوكيميائية -0
بروتينات الدهنية منخفضة ن ، الدهون الثالثية، الكوليسترول، الييقياس كل من هوموسيستتم
بن إ" نزيمات الكبدية في مخبر التحاليل البيولوجية إل، البروتينات الدهنية عالية الكثافة، و االكثافة
.بقسنطينة" سينا
:كسدة ألمعايرة نشاط مضادات ا -2
قياس الجلوثاثيون المختزل:
لقياس كمية الجلوثاثيون المختزل في الكبد، اتبعت طريقة (Weckbeker و األخرون
. في ذلك( 9100،
بعد إنقضاء فترة التجربة، تم تخدير الحيوانات و تشريحها لنزع الكبد بصفة فورية، في
حالته الرطبة، بعد ذلك تم إستخدام مسحوق الكبد المتجانس لمعايرة كمية الجلوثاثيون
:حسب المعادلة التالية DTNBختزل طيفيا بإستعمال كاشف التلوينالم
:حيث
OD : الكثافة الضوئية.
لى خط إجراء مقارنة إعن طريق ( Bradford) ،9179د تركيز البروتينات بواسطة طريقة تم تحدي -
.BSAالمعايرة

الملخص
5
تاالزقياس نشاط الك:
(Aebi ،9104)نزيم الكتاالز وفق طريقة إمسحوق الكبد المتجانس كذلك لمعايرة نشاط أستخدم
الطريقة ،كسجينأجزيئة ماء و لى إ H2O2 نزيم له القدرة على تحليل بروكسيد الهيدروجينإلا اذفه،
: المتبعة موضحة في الجدول التالي
:12الجدول
µL))العينات µL))الشاهد
الكواشف
088 718 , µM محلول الفوسفات
pH=7.5)988 (
188 188
H2O2
- 98
السائل الطافي لمسحوق الكبد
المتجانس
:كما هو موضح في العالقة التاليةنزيم إلبعدها تم تقدير فعالية ا ،تم قياس نشاط الكتالز طيفيا
:حيث
OD : الكثافة الضوئية

الملخص
6
ε :متصاصية الموليةمعامل اإل (0.043 mM-1.cm -1) .
L :طول كوفيت المستعملة (1 cm). .
X :البروتين كمية (mg/ml).
Fd : التخفيف عامل.
:تحضير القطاعات النسيجية
روطي و جزء من الكبد ووضعها في قارورات ألخد كل من القلب و الشريان اأبعد تشريح الفئران تم
.بغرض تثبيتها % 98بـ صغيرة تحتوي على الفورمول المخفف
مدة % ) 19، %78، %08) يثانول بتراكيز متزايدة إلحواض كحول اأستخدام إلك تم نزع الماء بذبعد
(.مرات 3تعاد كل مرحلة ) د 38كل مرحلة حوالي
د في كل 98لمدة (xylène)بعدها وضعت العينات في البوتانول، ليتم غمرها الحقا مرتين في زيلين
.مرة
مع تغيير البرافين كل و نصف ساعات 4ما في المرحلة الموالية، طمرت العينات في شمع البرافين لمدة أ
.(Hematoxylin eosin) ها خير قطعت العينات بالمقطاع المجهري ليتم تلوينألو في ا ،ساعة و نصف
:التجربة الثانية
: البيوفيلم تشكل رغان علىألتأثير زيت ا
:السالالت المستخدمة
:وهي نواع من السالالت البكتيرية أ 0ستعمال إه الدراسة تم ذفي ه
Staphylococcus haemoliticus
Streptococcus intermedius
Streptococcus anginosus
Streptococcus mutans
Streptococcus uberis
(.إيطاليا) تم الحصول على هذه السالالت من مخبر الميكروبيولوجيا بـكالياري

الملخص
7
و SH agar, Muller-Hinton agar : زرعت كل بكتيريا على حدة في وسط زرع المناسب -
بالنسبة للبكتيريا ساعة 14ثاني أكسيد الكربون لمدة %0و 37°حضنت في درجة حرارة
( Staphylococcus haemoliticus)اما البكتيريا الهوائية . (Streptococcus) الالهوائية
.فحضنت في وسط هوائي
أي وسط زرع سائل يحتوي على بكتيريا، و تم inoculum)) إينوكولوم بعد ذلك تم تحضير -
98الحصول على تركيز 9
و ذلك بإستخدام قياس مل لكل نوع بكتيري، /خلية بكتيرية
(.نانومتر 918) اإلمتصاصية على الطول الموجي
حيث وضعت : كما يلي well microplates-96 ))فيما بعد قمنا بعمل التجربة التالية على -
100 L 100من وسط الزرع و L ليتم بعدها اجراء .من زيت االرغان في الحفرة االولى
من L 988بعدها تمت اضافة ، (g/ml 988الى 8.84من )سلسلة من التخفيفات
. الى كل الحفر االنوكولوم
رت حففي حين احتو ،على وسط الزرع مع االنوكولوم( positive control) احتوت حفر -
( negative control ) على وسط الزرع مع زيت االرغان فقط.
كسيد الكربون لمدة أثاني % 0و 37° درجة حرارة في well microplates-96))ن يحضتتم -
.و في وسط هوائي بالنسبة البكتيريا الهوائية بالنسبة للبكتيريا الالهوائية ساعة 40
.PBSلك ذزالة إليتم فيما بعد PBSثالث مرات بمحلول و غسلت الحفر زيل محتوىألك ذبعد -
،د 98لمدة % 8.9تركيزها ) Crystal violet ( بصبغةالخطوة الموالية تمثلت في تلوين الحفر -
.زالة الصبغةإ بعدها تمت
ستخالص الصبغة الملتصقة بالحفر إنتاج المادة المخاطية نوعيا، تم إولتقدير قدرة البكتيريا على -
.لكل حفرة % 38و تركيز ذمن حامض الخليك (µl)188بإضافة
.نانومتر 918الطول الموجي متصاصية علىإلو من ثم قياس ا -
:تم استعمال العالقة التالية ، و لحساب نسبة تثبيط البيوفيلم -

الملخص
8
:حيث
C :امتصاصية positive control
B :امتصاصية negative control
T :امتصاصية test wells
النتائج المتحصل عليها
و أ( كغ/مغ088)جرعة الميثيونين بأخذسواء يتأثرن وزن الحيوانات لم أولية ألبينت النتائج ا
خالل (F ,M, MP, P)ربعة ألبين المجموعات ا ،(كغ/مغ908) رغانألور شجرة اذالمستخلص الخام لب
. (93)الفترة التجريبية كما هو موضح في الشكل
المنخفض ن كل من الكولسترول و الكولسترول أظهرت النتائج أبالنسبة للتحاليل البيوكيميائية فقد
البروتينات الدهنية عالية الكثافة ما أرتفعت بقيمة معتبرة ، إقد (TG)و الدهون الثالثية (LDL-c)الكثافة
(HDL-c) لك لدى المجموعة الثانية ذنخفضت بقيمة معتبرة كإفقد(M) مقارنة بالمجموعة الشاهدة(F).
(TG)و الدهون الثالثية (LDL-c)خرى، عرفت مستويات كل من الكولسترول أمن جهة
جرعات عالية من الميثيونين عطيتأ، و التى (MP)نخفاضا بقيمة معتبرة لدى المجموعة الثالثة إ
،ا المستخلصذمما يثبت فعالية ه (كغ/مغ908) رغانألور شجرة اذو المستخلص الخام لب( كغ/مغ088)
(.97، 99، 90، 94)شكال ألا
(M)رتفاعا معتبرا لدى المجموعة الثانية إعرفت (AST)نزيمات الكبد إن ألى إتشير النتائج
ن أفي حين ،ي تغير ملحوظألم يشهد ALT) (نزيمإ، بينما (F , MP, P)خرى ألا مقارنة بالمجموعات
ن المستخلص أمما يدل على ، بقيمة معتبرة (AST) نخفضت لديها مستوياتإ (MP)المجموعة الثالثة
(.91،20)الشكلين ،نزيمات الكبدإرغان له دور في تعديل ألور شجرة اذالخام لب
رتفعت بقيمة إن في الدم قد ين مستويات الهوموسيستيألى إلك ذتبين النتائج المحصل عليها ك
ا ما قورنت مع إذ( كغ/مغ088)عطيت جرعات عالية من الميثيونين أمعتبرة لدى المجموعة التي
بينما المجموعة التى تلقت معالجة مضاعفة بالميثيونين و . (F , MP, P)خرى ألالمجموعات ا
ا ذين، و هيمعتبرا في مستويات الهوموسيست نخفاضا إشهدت فقد رغانألور شجرة اذالمستخلص الخام لب
(. 90)الشكل ،ا العاملذرتفاع هإا المستخلص في الوقاية والحد من ذما يثبت نجاعة ه

الملخص
9
ن كمية الجلوثاتيون المختزل في أكسدة، فقد كشفت النتائج المتحصل عليها ألبالنسبة لمضادات ا
إذا، (M)نخفاظا ملحوظا بقيمة معتبرة لدى المجموعة الثانية إنزيم الكتالز قد عرفا إلك نشاط ذالكبد و ك
رتفاعا إفقد سجلت (MP)ن المجموعة أفي حين ،(F MP, P) األخرىما قورنت بالمجموعات
رتفعت لكن ليس بشكل إما كمية الجلوثاثيون لدى نفس المجموعة فقد أ، معتبرا بالنسبة إلنزيم الكتالز
(.11و 19)الشكلين ،(يوم 19)ن المدة التجريبية كانت غير كافية ألملحوظ، ربما
، األخرىما قورنت مع المجموعات إذا ن تحليل المقاطع النسيجية لفئران المجموعة الثانيةإ
نتكون خاليا رغوية في الطبقة الوسطى للشريا كذلكروطي ، ألتقشر البطانة الداخلية للشريان ا يظهر لنا
عضلة القلب تخريبا في بعض المناطق باإلضافة شهدت بينما ،مع تمركز بعض الدهون في حويصالت
خر تغيرات آل، عرف نسيج الكبد هو ا أخرىمن جهة .خالياه الذهتغيير في البنية المورفولوجية ل إلى
نوية، ظهور حويصالت أللبعض الخاليا الكبدية، تضخم بعض اشية البالزمية غألتمثلت في تخريب ا
(. 17، 19، 10، 14، 13)شكال األ .قاطع النسيجيةالمكما هو موضح في ،ليبيدية
تم تغذيتها يروطي و القلب و الكبد لدى المجموعة التألبينما المقاطع النسيجية لكل من الشريان ا
. رغان و الميثيونين معا فقد ظهرت بشكل سليمألور شجرة اذبالمستخلص الخام لب
بالنسبة لألنواع رغان فعاليته في القضاء على تشكل البيوفيلمألزيت اثبت أخر، آفي سياق
: البكتيرية التالية
Staphylococcus haemoliticus
Streptococcus intermedius
Streptococcus anginosus
Streptococcus mutans
و 39، 38، 11، 10)شكال ألا ا بوضوح فيذيتجلى لنا ه ،رغانألتوافقا مع زيادة تركيز زيت اا ذو ه
31.)

الملخص
10
:المناقشة
:خذ جرعات عالية من الميثيونين على وزن الفئرانأتأثير -0
خذ جرعات عالية من الميثيونين لم يوثر على وزن أن تأثير أ بينت النتائج المتحصل عليها
بحاث أ كذلك (.Lamda, ،1888)هذه النتائج تتوافق مع ،األربع المجموعات بين الفئران
(Zarrouk 1898، خرون آو ) على عكس(Zerizer 1889، خرون آو ) رتفاع محسوس إذكرت
، 2013)بحاث أكذلك . يوما 19كغ من الميثيونين لمدة /مغ188 بـالفئران التي عولجت في وزن
Boudebouz; Sakhri، 2014 ) كغ من /مغ 488 ـالفئران التي عولجت بن وزن أشارت أفقد
(Benmebarek بحاثأو على عكس . رتفاعا محسوساإخر عرف هو اآل سابيعأ 3الميثيونين لمدة
3كغ من الميثيونين لمدة /مغ 188 ـالفئران التي عولجت بنخفاض في وزن إين الحظت أ (1893،
.سابيعأ
.الفئران ووزن HHcy بين عالقة هناك نأ نستنتج نأ يمكننا ال ، المتضاربة النتائج لىإ وبالنظر
نزيمات إين، الدهون و يخذ جرعات عالية من الميثيونين على مستويات الهوموسيستأتأثير -1
:كسدة ألنشاط مضادات ا و الكبد
رتفاع مستويات إلى إدى أو جرعات عالية من الميثيونين قد ذ لغذائيان النظام أه الدراسة ذهظهرت أ
كسدة و البروتينات الدهنية ألنزيمات الكبد، بينما عرفت مضادات اإين، الدهون، يكل من الهوموسيست
ضرار في بنية كل من الشريان ألحاق إلى إدى أنخفاضا محسوسا مما إ (HDL-c)عالية الكثافة
.روطي و القلب و الكبد ألا
.كرذالتغيرات السالفة ال رغان فعاليته في تعديل ألور شجرة اذثبت المستخلص الخام لبأخرى أمن جهة
لى إدت أيوم 19للفئران لمدة ( كغ/مغ088)جرعات عالية من الميثيونين ذخأن أظهرت أنتائجنا
. ين في الدم يتكوين حالة فرط الهوموسيست
تنظيم تغير في إلىيرجع أن يمكن العالي من الميثيونين الغذائي النظام من المحتمل التأثير نإ
على أن الحفاظ الواضح ومن. سينثاز β سيستاثيون المثال سبيل على موسيستين،واله ستقالبإ إنزيمات
)ثيولال هذا من فعالة وإزالة نتاجإ لى تنظيمإراجع منخفضة من الهوموسيستين في األنسجة مستويات
Stipanuk ، 1884.)

الملخص
11
ين في الدم يرتفاع الهوموسيستإلى إدى أالغني بالميثيونين الغذائين النظام أبينت عديد الدراسات
(Mudd ،1899 ؛ Viggianoa ؛1891خرون، آو Yamada ؛1891خرون، آو Kirac خرون، آو
1893.)
مسار إلى الهوموسيستيين توجيه يتم ، الميثيونين يكون هناك فائض في عندما
transulfuration .الهوموسيستيين إلىتحويل يتم هذا السياق، في cystathionine ثم ومن cysteine
.الجلوتاثيون تركيب يدخل في عنصر هو pyridoxal 5-phosphate .cysteine قبل من الى
األكسدة ضرار نتيجة أ يسبب أن يمكنالهوموسيستيين تركيز رتفاعإ أن الدراسات أظهرت وقد
وبيروكسيد الحرة الجذور في اإلفراط في سببي مما مؤكسد عامل بمثابة يكون قد والسيستين ،الخلوية
؛ Hogg ، 9111) اتسرطانالتطورو الجينية اتطفرال من مزيد إلى يؤدي أن ويمكن الهيدروجين،
Stipanuk ، 1884 )الدموية واألوعية القلب وأمراض.
لى تشكيل إدى أين يرتفاع مستويات الهوموسيستإن أ( 1884خرون، آو Stanley)بينت دراسة
رتفاع إلك ذلى إ باإلضافة ،وعية الدمويةألكسدة دهون خاليا بطانة اأ، و (ROS)الحرة الجذور
NOنخفاض إ إلى دىأمما eNOSنتاج إو تثبيط iNOSلى تحفيز تكوين إدى أالهوموسيستين في الدم
(Kamal و Thanaa ،1881.)
كسدي أجهاد التإلن و ايمستويات الهوموسيستي رتفاعإثبتت العالقة الموجودة بين أخرى أدراسات
(Streck ؛1883خرون، آو Matté 2009خرون، آو.)
التيين لدى الفئران يالهوموسيست مستوياتنخفاضا كبيرا في إن هناك أظهرت نتائجنا أكما
لى الدور الفعال إمشيرة . المجموعات السابقة عمقارنة مرغان ذور شجرة األبالمستخلص الخام لب تغذت
خرون، أو Hamelet)ه النتائج تتوافق مع ذه. ينيه النبتة الطبية في خفض مستويات الهوموسيستذله
ن كل من أكرت ذ التيو ( 1893خرون، أو Meng ؛1881خرون، أو Yalcınkaya ؛1887
(quercetin )و (catechin) و( taurine )ين في الدم يساهمت في خفض مستويات الهوموسيست
.بالترتيب
رغان يكمل في محتوياته العالية من مضادات ذور شجرة األلمستخلص الخام لبلالوقائي التأثيرن إ
(. 1889خرون، أو Masella)كسدة القوية مثل البوليفينول، و توكوفيرول و ستيروالت ألا
تلقت جرعات عالية من يمستويات الشحوم في الدم لدى المجموعة الترتفاع إفيما يخص
، Herrmenn و Obeid ؛1881، خرونأو Lee)الميثيونين، فنتائجنا تتوافق مع دراسات كل من

الملخص
12
على عكس دراسة (. Sakhri ،1894 ؛Boudebouze ،1893 ؛1893خرون، آو Tselmin ؛1881
(Zerizer ،1889؛ Zerizer 1880خرون، آو ) ين يرتفاع الهوموسيستإلم تجد أي عالقة بين التيو
.رتفاع مستويات الشحومإو
وعية الدموية و خاليا الكبد بطانة األة بالنسبة لخاليا ين و الشحوم سامييعتبر كل من الهوموسيست
(Obeid و Herrmenn ،1881 .) ن ويوجود عالقة عكسية بين الهوموسيستي أثبتتدراسات كثيرة
و Poloni ؛9111خرون، آو Moat)نسان سواء عند اإل HDL-c))البروتينات الدهنية عالية الكثافة
.(1890خرون، آو Poloni)و عند الحيوان أ( 1891خرون، آ
وإفراز phosphatidylcholine نتاجإ من يقلل الميثيلية المجموعة نخفاض في إ أيضا،
VLDL ، الكبد في الثالثية الدهون تراكم في يتسبب مما قد (Li and Vance ، 1880؛ Bravo
يؤدي ، CβS deficiency نقص عن الناجمة فرط الهوموسيستيين في الدم، أنكما (. 1899 وآخرون،
؛9174 ،وآخرون Gaull) الكبد في الدهون وتراكم للدهون الغذائي التمثيل تنظيم في ضطراباتإ إلى
Namekata ،1884 وآخرون.)
جيد بشكل ، HDL وخاصة الدهنية، بين الهوموسيستيين والبروتينات عكسية عالقة وصف تم وقد
وآخرون، Poloni) و عند الحيوانات( 1891 وآخرون، Poloni ؛9111 وآخرون، Moat) عند البشر
وآخرون، Werstuck و) الخاليا داخل الدهون ستقالبإ يغير الهوموسيستيين كما أن(. 1890
أن يبدو و (.1893 وآخرون، Carvalho) الكبدية الدهون تراكم مع ترافقي أن ويمكن ،(1889
hypomethylation األنسجة في الدهون تراكم عن المسؤول هو مع فرط في مستويات الهوموسيستيين .
atherothrombotic (Obeid أمراض بتطور في الدم الكوليستيرول وفرط الهوموسيستيين ويرتبط
and Herrmann ، 1881 .)،توازن أن إلى الدراسات من العديد أشارت فقد وعلى عكس ذلك
(.1890 وآخرون، Koyama) الدموية واألوعية القلب أمراض لقمع رئيسية آلية هو الكوليسترول
تتصدى التي البيولوجية العمليات من العديد في HDL)) البروتينات الدهنية عالية الكثافة تشارك
الدموية بطانة األوعية ،(بيتا)البنكرياس خاليا وظيفة على التأثير خالل من التأكسدي، واإلجهاد لتهابلإل
جزيئات عن التعبير عن فضال األحادية، الخاليا وتنشيط الدموية، المبرمج للبطانة األوعية الخاليا موت ،
(.1899 وآخرون، Chapman)لتصاق اإل

الملخص
13
المشاركة الجزيئات أو اإلنزيمات بتثبيط مرتبطا HDL)) تأثير الهوموسيستيين على يكون وربما
الشرايين تصلب خطر زيادة فإن ولذلك،HDL (Obeid and Herrmann ، 1881 .) تجمع في
Paraoxonase-1 PON1 (Holven نشاط نخفاضإب صلة ذات تكون قد المختلة HDL وجزيئات
thiolactonase (Domagala نشاط ويظهر الكبد من أساسا يفرز PON1(. 1880 وآخرون،
(.1880 وآخرون، Bhattacharyya) الشرايين تصلب من حماية وبالتالي( 1889 وآخرون،
رتفاع مستويات الدهون إقتران مع باإل HDL-c))البروتينات الدهنية عالية الكثافة نخفاض إن إ
لها دور HDL-c))لك جزيئات ذلى إ باإلضافة. بتصلب الشرايين لإلصابةالثالثية يعتبر عامل خطر
(.9113خرون، آو Macknesset) (LDL-c)كسدة الكولسترول الضارأوقائي ضد
التي الجرذانلدى LDL))جزيئات رتفاع إ ، (1898خرون، آو Bhandari)حسب دراسة
في عدد نخفاضإلى إين راجع ربما يميني الهوموسيستآلتعاني من مستويات عالية من الحمض ا
LDL))مستقبالت في تغييرات أو. مع مستقبالته LDL)) نخفاض ترابط إو أ LDL))مستقبالت
.د جرعات عالية من الميثيونينأخرتفاع مستويات الكوليسترول بسبب إدي الى ؤت
رغان في تعديل ألور شجرة اذالمستخلص الخام لبظهرت نتائجنا فعالية أخرى، أمن جهة
الذينخفاض الهوموسيستين لدى نفس المجموعة و إلى إا راجع ذمستويات الدهون السالفة الذكر، و ه
.كسدةأله النبتة بمضادات اذه غنى لى إبدوره راجع
نخفاضا إتالز فقد عرفا انزيم الكإلى كمية الجلوثاثيون المختزل في الكبد و نشاط إبالنسبة
فيما يخص الجلوثاثيون المختزل فنتائجنا . معتبرا لدى الحيوانات التي تلقت جرعات عالية من الميثيونين
لميثيونينعولجت با التيالفئران نفس النتائج لدى تحصلت علىو التي ( Sakhri ،1894) تتوافق مع
.(كغ/مغ488) و لكن بجرعة
نخفاض إن أ، (1899و آخرون، Bravo ؛ Vance ،1880و Li)ظهرت دراسات كل من أ
. ينييضي في تصنيع الهوموسيستأليعتبر المنظم ا الذي (SAM)نخفاض مستوى إالجلوثاثيون يتوافق مع
كسدة القوية التي تلعب دورا هاما في حماية ألمضادات احد أالمختزل هو الجلوثاثيون
(. 1889، آخرون و free radical reactive( )Stipanuk )نسجة من الجذور الحرة ألا
نزيماتاإل وتشمل الخاليا، بين األكسدة تفاعل ساسية للحد منوامل األعحد الأ هو الجلوتاثيون
glutathione peroxidaseو glutathione S-transferase (GST) لها التابعة لألكسدة المضادة

الملخص
14
(GPx)، المؤكسدة واألنواع الحرةللجذور الفعل رد ضد الخلوي الدفاع في أساسيا دورا تلعب والتي
-GSH S و GPx بواسطة العضوية البيروكسيدات تخفيض أيضا يمكن(. Stipanuk ، 1889) األخرى
transferase. في فقط موجود ولكنه الهيدروجين بيروكسيد يقلل أن أيضا يمكن كاتاالزال إنزيم
التأكسدي اإلجهاد من كل ضد الدفاع في خاص بشكل مهما الجلوثاثيون ليجع وهذا. بيروكسيسوم
-Fernándezو Garcia-Ruiz ؛9117 وآخرون، Fernández-Checa) الفسيولوجي والمرضي
Checa ، 1889 .)كثير حدثي الجلوثاثيون مستويات اضخفنإ نأشارت أ التي البحوثالعديد من هناك
بسبب مهيمنا دورا تلعب التأكسدية اإلصابات أن حين في(. 1881 وآخرون، Ballatori) األمراض من
نزيماتإ نتاج إ نخفاضإب سببيا يرتبط وبعضها ضطرابات،اإل هذه من العديد في الجلوثاثيون نخفاضإ
(.Lu ، 1881)نتاج الجلوثاثيون إ
الكبدي لدى التأكسد خطورة من المزيد يسبب قد الكبد في الميثيونين سولفوكسيد تراكم فرط نإ
،جلوثاثيون ذلك في بما السيستين،مثل األكسدة مضادات مستوياتها كانت والتي ، −/−Cth الفئران
taurine/hypotaurine,، غيرمعدلة (Yamada ،1891 وآخرون .)وسيستين فيهومال فرط ويؤدي
لألكسدة المضادة الدفاع أنظمة يضعف الذي ROSنتاج إ طريق عن لتأكسديا اإلجهاد زيادة إلى الدم
(.9110 وآخرون، Welch) أو الخاليا داخل
و Woo) تالز، فنتائجنا تتجه في نفس السياق معانزيم الكإنخفاض نشاط إفيما يخص ما أ
ه الدراسات وجدت ذه(. 1881و آخرون، Matté ؛ 1887و آخرون، Chanson ؛ 1889آخرون،
.نزيم الكتالز في الكبدإنخفاض إين و يرتفاع مستويات الهوموسيستإعالقة عكسية بين
ين بسبب عديد من المثبطات على ينزيم الكتالز من طرف الهوموسيستإنخفاض نشاط إيعود
و Putnam ؛Novogrodsky ،9100و amino-1:2:4: triazole (Margoliash-3غرار
amyloid-ß (Milton ،1999 .)، و (1888خرون،أ
رغان قد ساهم في رفع و ور شجرة األذالمستخلص الخام لب ن أ ه الدراسةذنتائج هظهرت أ
رتفعت لكن ليس بقيمة معتبرة، إخرى ألبينما مستويات الجلوثاثيون هي ا ،تالزانزيم الكإ تحسين مستويات
. فترة التجريبية لراجع لقصر ا ربما اذو ه
نزيم إ نتاجإلى تعديل إدت أه المركبات ذه. الذكرسلفنا أرغان غنية بالبوليفينول كما ألن نبتة اإ
و (1883خرون، آو Floreani )نسيج عضلة القلب نسجة على غرار ألفي عديد ا eNOSتالز و االك
. (1887خرون، آو Kasdallah-Grissa )نسيج الكبد

الملخص
15
رتفاع معتبر في إ، فقد تحصلنا على (AST، ALT)فيما يخص النتائج المتعلقة بإنزيمات الكبد
و التي تتوافق مع دراسة . رتفاعا بقيمة غير معتبرةإشهدت ( ALT)، بينما مستويات (AST)مستويات
(Yalcınkaya 1881خرون، آو )رتفاع إلى إدت أاشهر 9بالميثيونين لمدة الجرذان تغذيةن أثبت أى ذال
(.ALT)، و(AST)ين، و يمستويات كل من الهوموسيست
يدل على تخرب ( AST، ALT) رتفاع هده األنزيمين إن فإ، (1880و آخرون، Ozer)حسب
، Payneو Killip)فهو يعكس تغيرات في نسيج العضلة القلبية (AST)رتفاع نشاط إما أ الكبد، نسيج
9198) .
خرى في تعديل مستوياتأرغان مرة ألور شجرة اذالمستخلص الخام لبظهرت نتائجنا فعالية أ
(. AST) نزيمإ
تأثير اخذ جرعات عالية من الميثيونين على المقاطع النسيجية للقلب و الكبد و الشريان -3
.روطي األ
لية من الميثيونين ، فيتجلى المقاطع النسيجية للمجموعة التي تلقت جرعات عا ما فيما يخص تحليلأ
ل من البنية النسيجية حداث بعض تغيرات لكإن في يرتفاع مستويات الهوموسيستيإبوضوح تأثير لنا
نخفاض مستويات الجلوثاثيون المختزل إعلى الدهون و تأثيرهبسبب . روطي و القلب و الكبدللشريان األ
.كسديجهاد التأو التي تعكس حالة اإل نزيم الكتالز إو نشاط
اللتان ( Sakhri ،1894 ؛Boudebouze ،1893 ( تتوافق مع دراستي كل من ه النتائج فهيذه
.السالفة الذكرنسجة نفس األ بنية لى التأثير علىإدت أجرعات عالية من الميثيونين ن أذكرتا
األوعية، مرونة من والحد البطانية، الخاليا تلف إلى يؤدي ينيرتفاع مستويات الهوموسيستإ أن يعتقد
نهاية و التي تنتهي في (.Kopczynski ،1894 و Baszczuk)توازنها الداخلي عملية كما يغير من
و Papatheodorou ؛1889و آخرون، Gallai) الدماغية والسكتات القلبية النوبات إلى المطاف
Weiss ،1887.)
و التي بدورها تساهم في ( ROS)نتاج الجذور الحرة إلى إن يالهوموسيستي ارتفاع ديؤكذلك ي
و Kanani) وعية الدموية مما يعزز تطور عملية تصلب الشرايينألتالف خاليا بطانة اإتخريب و
(. 9111آخرون،

الملخص
16
ين تساهم في يرتفاع مستويات الهوموسيستإن أ مؤخرا، ( 1890و آخرون، Chen) دراسة أظهرت
في البطانة S-nitrosylation بروتين مستويات نخفاضإ مع ا يتوافق ذو ه الشرايين تصلب تقدم
واألوعية القلب وظائف تنظيم في هاما دورا تلعب وعية الدموية ، و التيألو اأروطي ألالداخلية للشريان ا
. الدموية
والجلطات هي الشرايين تصلب فيين يرتفاع الهوموسيستإخرى التي يساهم فيها ألليات اآلو من بين ا
البروتينات الدهنية كسدة أالتأكسدي، اإلجهاد الخاليا، زيادة العضلية، سمية الخاليا تحفيز تكاثر :
Willoughby) ، و المساهمة في عملية التخثر و تنشيط الصفائح الدموية LDL) ) منخفضة الكثافة
(.1889و آخرون، Luo ؛ 1881و آخرون،
atherogenic ـو ما يعرف بأوجدنا سابقا في هذه الدراسة زيادة في مؤشر تصلب الشرايين كما
index الكوليسترول في الدم وانخفاض نتيجة لزيادةHDL . حسب(Mehta ،1883و آخرون) ،
عضلة القلب، ىلإو تسرب الدهون أ( foam cells) المؤشر وجود الخاليا الرغوية اذيعكس ه
. بها ضرارأالشريان األورطي، الكبد و الكلى ، مما يلحق وعية التاجية، ألا
في عدة نماذج ي، نسيج الكبدالتالف إين يسبب يالهوموسيست فرطن ألك وجد ذلى إباإلضافة
اإلندوبالزمية، الشبكة التأكسدي، إجهاد اإلجهاد زيادة :ليات، و التي تتمثل في أتجريبية عن طريق ثالث
و من بين هده (.1880 وآخرون، Robert ؛Kaplowitz ،1884و Ji) لتهابيةو تنشيط العوامل اإل
و IL-8 (Ji و 1b ، IL-6-(IL)نترلوكينات ، اإلNFkBالعامل النووي : لتهابيةالعوامل اإل
Kaplowitz ،1884؛ Matte ،1881 وآخرون) .
روطي والكبد، ألنسجة القلب، الشريان اأالمقاطع النسيجية لكل من نأه الدراسة ذظهرت هأو قد
بشكل سليم ظهرت رغان، ور شجرة األذعولجت بالميثيونين و المستخلص الخام لب فئران التيلل بالنسبة
.ه النبتةذكسدة التي تحتويها هألا راجع لمضادات اذو ه
الجرذان أن( 1893 وآخرون، Benmebarek) و( Naimi ، 1884 و Zerizer) عمل أثبت
الحقت بأضرار يوما 19 من الميثيونين خالل كغ/ مغ 188 ب التوالي على عولجت التي والفئران
إلى أدى B6 و B9، B12 الحيوانات بالفيتامينات هذه عالج أنكما والقلب األورطي الشريان باألنسجة
. الدموية واألوعية القلبب الملحقة األضرار هذه تصحيح

الملخص
17
البيوفيلم تشكيل تأثير زيت االرغان على -4
أنحاء جميع في والوفيات األمراضفي عديد من هاما سببا البكتيرية المعدية األمراض تمثل
.البكتيرية االلتهاباتلعالج هذه للميكروبات مضادة جديدة عوامل تطوير وجب ولذلك. العالم
، Draelos" )الحيوية العدوى أنواع من نوع" هي البكتيرية المعدية األمراض أن يفترض عادة،
العالجات لمختلف مقاومة أكثر عام بشكل األغشية الحيوية هذه أن األخيرة األبحاث أظهرت وقد ،(1898
(.Olsen ، 1890) للميكروبات المضادة
للميكروبات المضادة لعواملل مقاومتها زيادة هيو البيوفيلم أ الحيوية األغشية سمات أهم من واحدة
(Wimpenny ،1888 وآخرون .)وجود أن المعروف ومن EPS ، من الجافة الكتلة أغلبية يمثل الذي
(. 9119 ستيوارت،) المختلفة الحيوية المضادات فعالية من يقلل مما ميكانيكي، حاجز بمثابة بيوفيلم،ال
مع تفاعلت بيوفيلمال سطح على الموجودة الشحنة سالبةال البوليمرات أن المعروف من ذلك، على وعالوة
Nichols) األدوية هذه تغلغل من يحد مما ،(أمينوغليكوزيد مثل) الموجبة ذات الشحنة الحيوية المضادات
(.9100 وآخرون،
األغشية سطحعلى مثال ، األكسجين تركيزك المتغيرة البيئية الظروف تأثير ذلك، إلى باإلضافة
األغشية جوهر بينما تنخفض هذه النسبة في ،عالية األوكسجين مستويات تكون (وفيلميالب) الحيوية
Yang ؛9114 وآخرون، De Beer) الحيوية مضاداتال وصولل عرضة أقل النواة يجعل مما الحيوية،
(.1880 وآخرون،
تسمى آلية خالل من والفسيولوجية التعاونية أنشطتها تنظيم البكتيريا من العديد أن المعروف ومن
quorum sensing (QS)، اإلفراج طريق عن البعض بعضها مع تتواصل البكتيرية الخاليا حيث
تشمل تشكيل ، quorum sensing آلية. لإلنتشار قابلة صغيرة إشارة لجزيئات واإلستجابة واإلستشعار
و Li) البيوفيلم وتكوين المبرمج، الخاليا وموت الوراثية، والكفاءة البكتريوسين، وإنتاج ، الجراثيم
Tian ، 1891.)
تمتلك والتوابل واألعشاب النباتات في الموجودة الطبيعية المركبات من العديد أن تبين وقد
) األمراض مسببات ضد الميكروبات مضادات لعوامل كمصدر وتعمل الميكروبات مضادات وظائف
Deans and Ritchie ،91107؛ Kumar ،1889 وآخرون.)

الملخص
18
تكون أن يمكن( النباتات) الطبيعي المصدر من المستمدة للبكتيريا المضادة العوامل فإن وهكذا،
ضد نتقائيةإ بمزايا تتمتع أنها المعروف من التي الثانوية، األيضات وجود بسبب وذلك فعاال، بديال
(.Buss ، 1889 و Butler) المقاومة الكائنات
0 ضد الجزائري األرغان لزيت للبيوفيلم المضاد المفعول تقييم لىإهدفت دراستنا ،اقيسال هذا في
: و هي وعية الدمويةألنها تساهم في تشكيل مشاكل للقلب و اأيعتقد بكتيريا أنواع
Streptococcus mutans ، Streptococcus anginosus،Streptococcus intermedius
Streptococcus uberis و Staphylococcus haemolyticus .
و . الذكربكتيرية السالفة أنواع 4رغان كان فعاال في تثبيط ألزيت ا أن بيوفيلمال فحص نتائج أظهرت
حمض كافييك : مثل الطبيعي الفينول من مادة الزيت هذا ثراء ربما بسبب .ا تزامنا مع زيادة تركيزهذه
caffeic acid حمض الفانيليك، vanillic acid تيروسول ، tyrosol و ايبيكاتشين ، epicatechin
(charrouf وGuillaume ،1887.)
زيت من( مل/ ميكروغرام 988) تركيز أول كان اكيز المذكورة ،رالت بينن من أكما بينت النتائج
الزيت تأثيرن أغير . الخاليا لقتل ليس ولكن Streptococcus uberis بيوفيلملتقليص فعال األرغان
Streptococcus mutans ، Streptococcus anginosus على كل من بيوفيلم أعلى كان
،Streptococcus intermedius Staphylococcus haemolyticus كثر من أ بـ نخفاضإ مع
.لتثبيطا نسبةبالنسبة ل 18٪
ضد تنشط و التي النباتية، الكيميائية المواد وجود إلى األرغان لزيت المثبط النشاط يرجع أن ويمكن
الكيميائية المواد من أكثر أو ثنينإل التعاوني للتأثير نتيجة يكون أن يمكن أو البكتيريا من السالالت هذه
المضادات يعتبر من األرغانزيت أن يعني وهذا(. 1899 وآخرون، Da Silva) لهذا الزيت النباتية
الفانيليك، حمض الكافيك، حمض: مثل الطبيعية الفينوالت من الزيت هذا ثراء خالل من ربما الحيوية،
يحتوي انغاألر زيت أن ثبت وقد(. Guillaume ، 1887و Charrouf.... ) إبيكاتشين تيروسول،
و Charrouf ) والصابونين الفالفونويدات العفص، ،ستيروييدال مثل نباتية كيميائية مواد على
Guillaume ، 1887 .)وتمنع البكتيرية بالبروتينات ترتبط النباتية الكيميائية المواد هذه أن ثبت وقد
؛1898 وآخرون، Samy) النقل وبروتينات الخلوي، والمغلف واإلنزيمات، الميكروبات، لتصاقإ
Upadhyay ،1894 وآخرون .)

الملخص
19
. اآلليات هذه من بعض خالل من بيوفيلم تشكيل تثبيطقام ب اناألرغ زيت أن المرجح من وبالتالي،
لتصاقإ تعطل التي والعوامل( 1890 وآخرون، Rabin) بيوفيلم تشكيل أثناء مهم البكتيريا لتصاقإ
.للبيوفيلم مضادة كعوامل العمل على القدرة لديها األسطح إلى البكتيرية
نشاطا ظهرأالكافيك قد ن حمضأ ( 1893 وآخرون، Stojkovic) دراسة أظهرت السياق، هذا في
هيدروكسيناميك، حمض) البوليفينول من عدد على العثور تم أيضا،. للميكروبات ومضاد لألكسدة مضادا
(.1893 وآخرون، Nazzaro) Chromobacterium violaceum لمنع( إبيكاتشين روتين،
كسر طريق عن أو السطح على الحي الكائن تعلق منع طريق عن يتم نأيمكن البيوفيلم تثبيط ن إ
(.1899 وآخرون، Gupta)تشكل إذا البيوفيلم هيكل
النباتية الكيميائية المواد على حتواءهإليرجع قد االرغان في تثبيط البيوفيلم زيت لدور آخر تفسير
تمي عندما أنه الدراسات من العديد أظهرت وقد. البيوفيلم في اإلجهاد ستجابةإ تحفز التي للبكتيريا المضادة
بالبيوفيلم و المرتبطة تحفيزالجينات يتم الحيوية، بالمضادات العالجك الميكروبات، على تطبيق اجهاد
(.1893 وآخرون، Ackart) يوفيلمبلل الظاهري النمط اختفاءبالتالي
ؤ بشار أرغان شجرة ورذب من المستخرج األرغان زيت أن( 1890 آخرون، و Lotfi) خرا، كشفم
ة البكتيريا على إيجابيا في القضاء أثرا أظهر قد ،الجزائر على مقاومةال (البالنكتوني ة)العائمة والحر
العنقودية والمكورات ( Staphylococcus aurus)الذهبية العنقودية المكورات الخصوص وجه
.(Staphylococcus white)البيضاء
Bjarnsholt) الثوم مثل البيوفيلم ثبتت فعاليتها في تثبيط أبهارات ل بحاث أ نتائجنا تتوافق مع عدة
، (1881خرون، أو Khan)القرنفل ، (1889خرون، أو Niu)والقرفة والزنجبيل،، (1880خرون، أو
. ( 1894خرون، أو Packiavathy)والكركم ، ( 1891خرون، أو Packiavathy)الكمون
بخصائصها عرفت النباتية المواد عطرية مستخلصة من زيتية سوائل هي عبارة األساسية ن الزيوتإ
: ثبتت فعاليتها في تثبيط تشكل البيوفيلمأهده الزيوت التي من بين (. Burt ، 1884) للجراثيم المضادة
، Piper bredemeyeri ، Piper brachypodom ، Piper bogotenceلـ الزيت االساسي
Gaultheria procumbensوش دكحزنبل، القرنفل الزعتر و المر، ال(Khan ؛1881خرون، أو
Musthafa ؛ 1898خرون، أو Olivero ؛ 1899خرون، أو Jadhav 1893خرون، ألو.)

الملخص
20
وفي. األسنان لألمراض مسببة بأنها عموما الدراسة هذه في المستخدمة الدقيقة الحية الكائنات تعرف
في للتقدم عرضة أكثر تكون الفموية، البكتيريا تسببها التي تلك ذلك في بما لتهابات،إلا فإن السياق، هذا
البكتيريا بين صلة وجود الدراسات من العديد أثبتت فقد. سابقا يعتقد كان مما الوعائية القلبية األمراض
واحدة آلية من أكثر هناك أن جدا المرجح ومن للغاية، معقد أمر وهو الشرايين، وتصلب الفموية
(Leishman ،1898 وآخرون.)
تتحول جرح، أي ظهور مع. والبكتيريا البطانة الشريان أسطح بين تفاعل هو الشغاف التهابن إ
وآخرون، Kokare) القلب صمامات تتلف أن يمكن التي قوية و بيوفيلمات لىإ نتهازيةإلا الجراثيم
التناسلي البولي الجهاز البلعوم، خالل من الدم مجرى في في تدخل أن لهذه الجراثيم يمكن(. 1881
فإنهالتصاقها إ حالة في ولكن ،ضعيفة الشريان بطانة في جراثيمال تصاقلإ عموما،. الهضمي والجهاز
الحالة، هذه في(. 1881 وآخرون، NBTE( )Kokare) الخثاري الشغاف لتهابإ تلف أوجرح تسبب
فبرونيكتين تفرز البطانية خاليا. اإلصابة موقع في والفيبرين الدموية والصفائح الحمراء الدم خاليا تتراكم
.البكتيريا وكذلك البشرية الخلية الفيبرين، الكوالجين، ربط على القدرة لديها التي
يمكن التي فبرونيكتين مستقبالت لديها والعقدية العنقودية المكورات مثل الميكروباتخرى أمن جهة
وآخرون، Kokare) الصمامات أنسجة تلف وكذلك اإلصابة موقع على الحيوية األغشية تشكل أن
في تغيرات إلى تؤدي قد الفم طريق عن البكتيريا من طرف البطانية الخاليا غزو ولذلك،(. 1881
إلى تؤدي كلها وهي المبرمج،الخاليا موت وكذلك البطانية الخاليا proatherogenicوها مثل خصائص
(.1898 وآخرون، Leishman) البطانية الخاليا في وظيفة خلل
معجون تصنيع في تستخدم أن يمكن رغاناأل ن زيتإف عليها، الحصول تم التي النتائج خالل من
.الدموية بطريقة غير مباشرة واألوعية القلب أمراض يمنع والذي العشبية، األسنان
الخاتمة
ية الفئران تغذ طريق عن الدم في ينالهوموسستي فرطه الدراسة تكوين حالة ذكان هدف ه
يوما، و تقييم التأثير 19 بـخالل مدة تجريبية تراوحت ( كغ/مغ088) بجرعات عالية من الميثيونين
يضية و ألضد التغيرات ا (كغ/مغ908) رغانألور شجرة اام لبذللمستخلص الخالوقائي و العالجي
واع بكتيرية نأ 0ختبار المفعول ضد تشكل البيوفيلم ضد إخرى، أمن ناحية . البنيوية التي سببها الميثيونين
.وعية الدمويةألمقاومة و التي قد تتسبب في مشاكل للقلب و ا
األيضية اإلضطرابات بعض واضح بشكل سبب رتفاع جرعات الميثيونينإ أن الدراسة هذه أظهرت قد
:على غرار

الملخص
21
؛لدى الفئران الدم في نييستيالهوموس فرطتكوين حالة
؛رتفاع مستويات الكوليسترول، الدهون الثالثية إ
يبوبروتينات لختالل في الإ(Dyslipoproteinemia ) نخفاض مستوى إمعHDL-c رتفاع إو
؛LDL-c مستوى
مستوى الدم في الكوليسترول مستوى رتفاعإ بسبب الشرايين، تصلب مؤشر في زيادةHDL-c؛
؛ختالل في مستويات مضادات االكسدة إ
الكبد خاصة نزيماتإ في زيادة ALT.
والكبد والقلب األورطي للشريان بنيوية ضرارأ النسيجية تحليل المقاطع أظهر ذلك، إلى باإلضافة
:بما يلي تميزت
؛األورطي البطانة الداخلية للشريان وتخرب فقدان
؛األورطي الشريان من مختلفة أقسام في رغوية خاليا تشكيل
؛والكبد األبهر الشريان في كل من ظهور حويصالت ليبيدية
؛األورطي للشريان المطاطية األلياف ختفاءإ
؛ القلب لبعض خاليا تغيرات في البنية العامة
الفجوات تشكلنوية، و ألغشية البالزمية لبعض الخاليا الكبدية، تضخم بعض األتخريب ا
. السيتوبالزمية
رغان ألور شجرة اذبالمستخلص الخام لب غني غذائي نظام تباعإ ن أتفيد دراستنا
و البنيوية السالفة األيضية، ضطراباتإلثبت فعاليتة في تحسين اأقد (كغ/مغ908) الجزائرية
. لدى الفئران لميثيونينل عن الجرعات العالية الناجمة الذكر،
المفعول الحيوي و المثبط لتشكل فيما يخص عليها الحصول تم التي النتائج إلى بالنظر و
سالالت 4خرى ضد أثبت نجاعته مرة أقد الجزائري األرغان زيت أن نستنتج أن يمكننا البيوفيلم
. تركيزها توافقا مع زيادة ذبكتيرية مقاومة، و ه
لما تحتويه ربما يعود ( المستخلص الخام للبذور والزيت)رغان ألالوقائي لنبتة ا التأثيرهذا
tocopherolsالتوكوفيرول لتهابات مثل إلعلى العديد من مضادات األكسدة والمركبات المضادة ل
حمض فانيليك ،oleuropein، الوغوبين caffeic acidحمض كافييك ) ، الفينول (Eفيتامين )

الملخص
22
vanillic acid ، تيروسول tyrosolو كاتشين ، catechin) الكاروتينات ،carotenes ،
(.٪ األحماض الدهنية غير المشبعة08)، واألحماض الدهنية، squaleneالسكوالين
:لى القيام بما يليإنطمح في المستقبل الدراسة هذه نتائج على وبناء
؛النباتي مستخلصال في الواردة بيولوجيا النشطة الجزيئات وتحديد ستخالصإ
نشاط مضادات أكسدة أخرى مثل قييمت superoxide dimustaseو glutathione
transférase.
ظهار إو ،النباتي المستخلص يحتويها التي لألكسدة المضادة لإلنزيمات الجيني التعبير دراسة
؛ .الدموية واألوعية القلب عالجات فيدورها
؛اإللتهابية السيتوكينات الجيني ضد التعبير إبراز دور النبتة في تنظيم
؛األرغان لزيت للجراثيم المضادة المركبات عزل
الذهبية ، العنقودية المكورات مثل خرىأبكتيرية نواعأرغان ضد ألختبار فعالية زيت اإ
. الحيوية المضادات بعض معا الزيت ذله التعاوني النشاط إلى باإلضافة

الملخص
23
:االشكال
.يوما 19تأثيرالميثيونين على وزن الفئران خالل فترة : (02)الشكل
تأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات الدهون الثالثية : ( 03)الشكل
.يوما من التجربة 19خالل
تأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات الكوليسترول : (04)الشكل
.يوما من التجربة 19خالل
البروتينات الدهنية المستخلص الخام لبذور شجرة األرغان على مستويات تأثيرالميثيونين و: (05)الشكل
.يوما من التجربة 19خالل منخفضة الكثافة
البروتينات الدهنية تأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات : (06)الشكل
.يوما من التجربة 19خالل عالية الكثافة
تأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات الهوموسيستيين : (07)الشكل
.يوما من التجربة 19خالل
AST نزيم الكبد إتأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات : (08)الشكل
.يوما من التجربة 19خالل
ALT نزيم الكبد إالمستخلص الخام لبذور شجرة األرغان على مستويات تأثيرالميثيونين و : (01)الشكل
.يوما من التجربة 19خالل
تأثيرالميثيونين و المستخلص الخام لبذور شجرة األرغان على مستويات الجلوثاثيون : (21)الشكل
.يوما من التجربة 19المختزل خالل
نزيم الكتاالز إلبذور شجرة األرغان على مستويات تأثيرالميثيونين و المستخلص الخام : (20)الشكل
.يوما من التجربة 19خالل
(P) و (MP) ، (M) ، (F)المقاطع النسيجية للقلب للمجموعات : (22)الشكل
(P) .و (MP) ، (M) ، (F)المقاطع النسيجية للشريان األورطي للمجموعات : (23)الشكل
(P). و (MP) ، (M) ، (F)المقاطع النسيجية األورطي للمجموعات : (24)الشكل

الملخص
24
(P). و (MP) ، (M) ، (F)المقاطع النسيجية األورطي للمجموعات : (25)الشكل
(P). و (MP) ، (M) ، (F)المقاطع النسيجية للكبد للمجموعات : (26)الشكل
S. intermedius.بيوفيلمالرغان على تشكيل ألالتأثيرالمثبط لزيت ا: (27)الشكل
St. haemolyticus بيوفيلم العلى تشكيل األرغانالتأثيرالمثبط لزيت : (28)الشكل
S. mutansبيوفيلم العلى تشكيل األرغانالتأثيرالمثبط لزيت : (21)الشكل
S. anginosus بيوفيلم العلى تشكيل األرغانالتأثيرالمثبط لزيت : (31)الشكل
.S. uberisبيوفيلم العلى تشكيل األرغانالتأثيرالمثبط لزيت : (30)الشكل
.على تثبيط البيوفيلم األرغانتأثير تركيز زيت : (32)الشكل

Paper

THE PROTECTIVE EFFECTS OF ARGANIA SPINOSA SEEDS AGAINST HYPER-HOMOCYSTENEMIA INDUCED BY HIGH METHIONINE DIET IN MICE
Original Article
BADIAA AKLIL1,2, SAKINA ZERIZER*1,2, ZAHIA KABOUCHE1 1.2
Received: 10 Mar 2017 Revised and Accepted: 16 Oct 2017
Université Des Frères Mentouri-Constantine, Département de Biologie Animale, Laboratoire d’Obtention de Substances Thérapeutiques (L. O. S. T), 25000 Constantine, Algeria
Email: [email protected]
ABSTRACT
Objective: Hyperhomocysteinemia (HHCY), oxidative stress and decreased antioxidant capacities lead to several clinical manifestations and particularly, cardiovascular and liver diseases. Our aim in this study was to investigate the protective effects of Argania spinosa powdered seeds against high methionine diet-induced HHCY, oxidative stress and damages in the aorta, and heart of mice.
Methods: Adult male Mus Musculus was systematically divided into four groups of similar mean body weights and fed for 21 d with control and experimental diets. The control group (F) was fed with white bread (0.50 mg/mice), group (M) was fed with L-methionine (500 mg/kg/day), group (MP) was fed with L-methionine (500 mg/kg/day) plus A. spinosa powdered seeds 150 mg/kg), and the group (P) was treated with A. spinosa powdered seeds (150 mg/kg/day). The experimental diets were given in white bread (0.50 mg/mice). After 3 weeks of treatments, homocysteine (HCY) concentrations, hepatic antioxidant status and histological sections of aorta and heart were determined.
Results: Consumption of high methionine diet led to an increase in plasma HCY, reduced the concentrations of GSH, and the enzyme catalase. These were associated with the loss and degeneration of endothelium, fenestration and formation of foam cells of the aorta, also the alteration of the cardiac muscle. However, administration of A. spinosa powdered seeds in combination with methionine decreased the concentration of HCY from (10.04±0.83 μmol/l) to (7.26±0.46 μmol/l), increased catalase activity from (45.82±5.83 m mol/mg protein) to (62.26±3.32 m mol/mg protein), and ameliorated histological changes.
Conclusion: A. spinosa powdered seeds were effectives in decreasing plasma HCY level as induced by methionine-enriched diet in mice, and improved the antioxidants defence.
Keywords: Homocysteine, Argania spinosa, Antioxidant enzymes, Methionine, Oxidative stress, Cardiovascular diseases
© 2017 The Authors. Published by Innovare Academic Sciences Pvt Ltd. This is an open-access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/) DOI: http://dx.doi.org/10.22159/ijpps.2017v9i12.18275
INTRODUCTION
Both cardiovascular disease and liver injury are major public health issues [1]. It has been reported that elevated plasma HCY concentration is an important risk factor for vascular diseases [2-3], and defects in methyl group metabolism, often resulting in HHCY, are among the key molecular events postulated to play a role in liver injury [1]. Plasma HCY levels can be determined by genetic, biological, nutritional, hormonal and lifestyle factors [4].
HHCY caused by excessive methionine intake is a classical attribute cardiovascular [5] and hepatic diseases [6-7]. Due to the presence of the highly reactive sulfhydryl group, HCY can undergo auto-oxidation to generate oxygen radicals [8].
Studies have shown that high HCY concentration could cause oxidative damage to cells [9-10]. HHCY induces endothelial dysfunction [11], which played an important role in the early stages of the atherogenic process by decreasing the availability of NO, stimulating the activation of nuclear factor kappa B and consequently increasing the expression of ICAM-1 [12]. Indeed, the oxidative stress resulting from elevated serum Hcy can oxidize membrane lipids and proteins and stimulate the activation of NF-B, and consequently increase the expression of inflammatory factors in vivo [13].
Hyperhomocysteinemia leads to increased oxidative stress via the generation of reactive oxygen species which weaken intracellular antioxidation defence systems [14]. ROS (including superoxide and hydrogen peroxide) are produced by endothelial cells and the adjacent smooth muscle cells, adventitial fibroblasts and inflammatory cells. ROS can affect the NO pathway [15]. The need for protection against ROS and other reactive molecules has led to
the specious theory that a high intake of exogenous antioxidants protects the body from oxidative stress, which is also widely known as the antioxidant hypothesis [16-17].
A high intake of fruit and vegetables rich in natural antioxidants, such as vitamins C and E, polyphenols, carotenoids, terpenoids, and phytomicronutrients, show an inverse association with the risk of cancer and the development of cardiovascular diseases.
The argan tree (Argania spinosa (L.) Skeels is a tropical plant, which belongs to the Sapotaceae family and is endemic in southwestern Morocco [19], and Algerian region of Tindouf [20]. It is exploited essentially for its fruits. The endosperm seed of fruit constitutes a good potential source of edible oil for human consumption and endowed with important medicinal properties [19].
Considering its rich composition in antioxidant compounds and unsaturated fat, A. spinosa can be used as a nutritional intervention in the CVD diseases prevention [21].
This study was designed to investigate the beneficial effects of powdered seeds of A. spinosa against hyper-homocystenemia, antioxidant status and damages in the heart and aorta induced by high methionine intake in mice.
MATERIALS AND METHODS
Plant material
Seeds of Argania spinosa were collected from a region near Tindouf (Southwest of Algeria). The fruits were cut into pieces to obtain seeds, which were subjected to size reduction to a coarse powder using a mechanical grinder. The powder was then used for treatment preparations.
International Journal of Pharmacy and Pharmaceutical Sciences
ISSN- 0975-1491 Vol 9, Issue 12, 2017

Zerizer et al.
Int J Pharm Pharm Sci, Vol 9, Issue 12, 64-69
65
Chemicals and reagents
L-Methionine purity 98% was obtained from across organics (Belgium). Total HCY levels were measured using the Immulite HCY kit (Siemens, Finland), on Immulite 2000 system. Other chemicals used were of Sigma chemical company.
Animals
Experiments were performed on 28 adult male Albino Mus Musculus mice weighing (30–35g), given from central pharmacy, Algeria. Animals were housed 7 per cage, and maintained under standard laboratory conditions of humidity, temperature (25 °C) and light (12 h day: 12 h night). After the adaptive period, they were divided into four groups of similar mean body weights and fed for 21 d with control and experimental diets. The control group (F) was fed with white bread (0.50 mg/mice), the second group (M) was fed with L-methionine (500 mg/kg/day), the third group (MP) was fed with L-methionine (500 mg/kg/day) in combination with powdered seeds of A. spinosa (150 mg/kg/day), while the group (P) was treated with powdered seeds of A. spinosa (150 mg/kg/day). The experimental diets were given in white bread (0.50 mg/mice) and allowed free access to food and water. After 3 w* of feeding, blood samples were collected after fasting, from the retro orbital plexus into EDTA tubes by using glass capillaries. They were centrifuged immediately, and plasma was frozen under-20 °C until assay time. The experiments were conducted in strict compliance according to ethical principles and provided by the committee for the purpose of control and supervision of Experiments on the animal (CPCSEA).
Determination of homocysteine level
The levels of total homocysteine (t-HCY) were assayed by competitive solid phase chemiluminescence immunoassay.
Tissue homogenate preparation
0.5 g of the liver was homogenized in 2 ml of TBS (Tris 50 mmol, NaCl 150 mmol, pH 7.4). The homogenates were centrifuged at 9000 g for 15 min at 4˚C, and the resultant supernatant was used for determination of: reduced glutathione, the catalase activities and protein concentrations.
Protein quantification
Protein was measured by the method of Bradford (1976) [22], using bovine serum albumin as the standard.
Determination of reduced glutathione (GSH)
The glutathione reduced content in the liver was measured spectrophotometrically by using 5, 5′ -dithiobis-(2 nitrobenzoic acids) (DTNB) as a coloring reagent, following the method described by Weeckbeker and cory (1988) [23].
Determination of catalase (CAT)
Tissue CAT activity was determined according to Aebi’s method (1974) [24]. The principle of the assay based on the determination H2O2 decomposition rate at 240 nm.
Histological sections
After the blood samples collection, the animals were sacrificed and organs designed for morphological analysis (heart, and aortas) were quickly removed, rinsed with saline solution (0.9%), and fixed in formalin 10%. The processed tissues were embedded in paraffin, sectioned at 5 μm thickness, and stained following the haematoxylin eosin staining method.
Statistical analysis
The values obtained were expressed as mean±SEM and subjected to statistical analysis using one-way analysis of variance (ANOVA), followed by Tukey’s post-hoc test (SPSS version 20). P<0.05 values were considered as significantly difference.
RESULTS
Effects of treatments on HCY
At the third week, plasma HCY of the (M) group achieved higher levels, and the average was 10.04±0.83 μmol/l, significantly higher
than that of the control group (F) (6, 84±0.51 μmol/l) and control positive group (P) (7.1±0.88 μmol/l). However, the combined treatments of A. spinosa powdered seeds with methionine (7.26±0.46 μmol/l) showed a significant decline in serum t-HCY, demonstrating that A. spinosa appears to be effective in preventing the increase of t-HCY levels (fig. 1).
Fig. 1: The interaction of L-methionine and A. spinosa seeds on the plasma homocysteine in mice during 21 d of treatment,
Values are the means±SEM (n); *p<0.05 and **p<0.01
Effects of treatments on hepatic anti-oxidative stress parameter
The present data showed that there is a highly significant depletion in reduced glutathione level (4.48±0.55 n mol/mg protein) (P<0.01) and a significant decrease in catalase activity (45.82±5.83 m mol/mg protein) (P<0.05), for the group (M) in comparison with control group (F), where the reduced GSH level was (8.03±0.55 n mol/mg protein) and the concentration of catalase was (61.37±6.39 m mol/mg protein). However the concentration of reduced GSH increased significantly in the group (P) (7.36±1.28 n mol/mg protein) (P<0.01), and not significantly with the group (MP) (6.1±0.5 n mol/mg protein) (P>0.05). On the other hand, the concentration of catalase increased significantly in the group (MP) (62.26±3.32 m mol/mg protein) (P<0.05), but not significantly in the group (P) (52.16±3.19 m mol/mg protein) (P>0.05) (fig. 2).
Histological impact of A. spinosa powdered seeds on heart and aortas tissues
Our data shows various pathological alterations in the heart and aorta of mice induced by the oral methionine administration.
In the second group (M) which had been fed with 500 mg/kg of methionine, Microscopic observation of heart was characterized by the presence of lysis, and architectural changes of cardiomyocytes as shown in (fig. 3-B and 3-C). In the other groups: (F), (MP) and (P), we have not observed any alteration in contrast to methionine intake group (fig. 3-A, 3-D and 3-E).
In the group (M), the aortic intima showed degeneration and desquamation of endothelial cells with fenestration, we also observed in the media lysis, formation of foam cells laden with small lipid droplets and oval nuclei as illustrated in (fig. 4-B and 4-C), (fig. 5-B) and (fig. 6-B, 6-C, 6-D and 6-E).
However, in the control group (F), the aortic sections have intact endothelium and spindle-shaped meiocytes nuclei as shown in (fig. 4-A, 5-A and 6-A). Also, we have observed intact aorta in the group (MP) treated with L-methionine and A. spinosa (fig. 4-D, 5-C and 6-F), and in the group (P) treated with A. spinosa only (fig. 4-E, 5-D and 6-G).
DISCUSSION
Hyper-homocystenemia can arise from nutritional deficiencies of folate, vitamin B6, and vitamin B12 [25]. Several diseases such as renal and thyroid dysfunction cancer, psoriasis, and diabetes as well as various drugs, alcohol, tobacco, coffee, older age and menopause, are believed to be associated with moderately elevated HCY concentrations [26]. There has been an indication towards a

Zerizer et al.
Int J Pharm Pharm Sci, Vol 9, Issue 12, 64-69
66
significant correlation between HHCY and cardiovascular disease and its complications such as heart attacks and strokes [27]. It is believed that hyperhomocysteinemia leads to endothelial cell damage, reduction in the flexibility of vessels, and alters the process of homeostasis [27]. HHCY may lead to an enhancement of
the adverse effects of risk factors like hypertension, smoking, lipid and lipoprotein metabolism, as well as the promotion of the development of inflammation. Physical activity, moderate alcohol consumption, good folate and vitamin B12 status are associated with lower HCY levels.
Fig. 2: The interaction of L-methionine and A. spinosa seeds on the reduced glutathione levels, and catalase activity in mice during 21 d of treatment, values are the means±SEM (n); *p<0.05, and **p<0.01
Fig. 3: Histological sections of the heart of (A) control, (B), (C) treated with Methionine, (D), treated with Methionine and A. spinosa powdered seeds and (E) treated with A. spinosa powdered seeds for 3 w*. Hematoxylin-Eosin Staining (A, C, D and E X100, and B X 400).
ACC. Architectural Changes of Cardiomyocytes, CMF. Cardiac Muscle Fibers, MCN. Muscle Cell Nuclei, L. Lysis
Fig. 4: Histological sections of the arch aorta of (A) control, (B), (C) treated with Methionine, (D), treated with Methionine and A. spinosa powdered seeds and (E) treated with A. spinosa powdered seeds for 3 w*. Hematoxylin-Eosin Staining (A, B, D and E X100, and C X 400).
FC. Foam Cells, FN. Fibroblast Nuclei, IEND. Intact Endothelium, LD. Lipid Droplets, Lu. lumen, ON. Oval Nuclei, SN. Spindle Nuclei

Zerizer et al.
Int J Pharm Pharm Sci, Vol 9, Issue 12, 64-69
67
Fig. 5: Histological sections of the abdominal aorta of (A) control, (B) treated with Methionine, (C) treated with Methionine and A. spinosa powdered seeds and (D) treated with A. spinosa powdered seeds for 3 w*. Hematoxylin-Eosin Staining (A, B, C and D X100). END.
Endolysis, FN. Fibroblast Nuclei, IEND. Intact Endothelium, LU. Lumen, SN. Spindle Nuclei
Fig. 6: Histological sections of the iliac aorta of (A) control, (B), (C), (D) and (E) treated with Methionine, (F), treated with Methionine and A. spinosa powdered seeds and (G) treated with A. spinosa powdered seeds for 3 w*. Hematoxylin-Eosin Staining (A, B, D and E X100, and C X 400). D. Desquamation, END. Endolysis, F. Fenestration, FC. Foam Cells, FN. Fibroblast Nuclei, IEND. Intact Endothelium, LU. Lumen, ON.
Oval Nuclei, SN. Spindle Nuclei
The current study has shown that high Met-diet in mice during 21 d clearly caused a significant increase in HCY level and decreased the reduced glutathione, and catalase concentrations in liver, meanwhile our results confirm that high level of HCY might be an effect of aorta and heart damages.
Several studies have shown that methionine enriched-diet induced a significant increase of plasma t-HCY [28-31].
Further, we found that the content of GSH and catalase activities in liver tissue was significantly decreased in response to the oral methionine administration.
hyperaccumulation of methionine sulfoxide in the liver may induce more serious oxidative hepatotoxicity in Cth−/− mice, whose levels of several antioxidative cysteine metabolites, including GSH, and taurine/hypotaurine, were all downregulated [30]. HHCY leads to increased oxidative stress via the generation of reactive oxygen species (ROS) which weaken intracellular antioxidation defence systems or elicit intracellular redox-controlled inflammation responses [32].
Glutathione is a key buffer of intercellular oxidative reduction reaction, and its dependent antioxidant enzymes include glutathione S-transferase (GST) and glutathione peroxidase (GPx), which play a fundamental role in cellular defense against reactive free radical and other oxidant species [33]. In addition, it has been shown that Hcy can directly act on catalase and inhibit the breakdown of H2O2 by
conversion of the enzyme into the inactive form [34]. Loss of catalase activity is associated with increased susceptibility to oxidative stress [35-36]. The mechanism of HCY inhibition of catalase is shared with a number of inhibitors including 3-amino-1:2:4:-triazole [37-38] and amyloid-ß [39].
Histological analysis showed that HHCY induced by the high methionine intake prompted an angiotoxic activity on the aorta and cardiac tissue damages. This was observed through the loss and degeneration of endothelium, formation of foam cells in the different sections of the aorta, alteration of the cardiac muscle.
In our experimental situation, is due to elevated HCY levels, which decreases the reduced GSH, and catalase activities, the well-known biomarkers of oxidative stress. Our results are in agreement with [40, 31], who reported that HCY-induced injury to the arterial wall is one of the factors that can initiate the process of atherosclerosis, leading to endothelial dysfunction and eventually to heart attacks and strokes [41-42]. Another work of [43] showed that elevated plasma HCY increase cholesterol synthesis, exerts an angiotoxic action direct to the aorta, by the loss of endothelium and degeneration partly with the dissolution of media cells.
Evidence from animal models of HHCY suggests that endothelial dysfunction is largely due to oxidative stress and decreased bioavailability of NO [44], NO may protect against the onset of vascular diseases [45].

Zerizer et al.
Int J Pharm Pharm Sci, Vol 9, Issue 12, 64-69
68
HCY promoted oxidative stress through the production of reactive oxygen species (ROS). ROS disrupts endothelial cell integrity, which in turn, can cause endothelial cell damage predispose affected vessels to the subsequent development of atherosclerosis [46].
Studies in several animal species, including rabbits, baboons, and rats, have demonstrated desquamation of endothelial cells, fragmentation of the internal elastic lamina, disruption of elastic fibers, and focal areas of smooth muscle hyperplasia [47-48] has been reported that the combination of high methionine and cholesterol increased the alterations of the arterial wall structures and the thickness of the aortic wall in animal models.
In methionine-treated animals, it was shown an aortic angiotoxic action with alterations not observed in the arterial vascular system of other organs. [49] have reported that acute elevations in plasma HCY after methionine loading causes vessel endothelial dysfunction and this could be reversed by administration of vitamin E in humans.
It has been documented that HCY can interact with different plasma and cellular proteins and by forming mixed disulfide conjugates, alters the physicochemical properties of the affected proteins. This has been also proposed as a potential mechanism for Hcy induced cellular dysfunction [50].
On the other hand, we found that the diet supplemented with the extract crude of A. spinosa was effective in prevention against HHCY in mice exposed to a Met-enriched diet by lowering the concentrations of HCY and increasing the concentrations of glutathione reduced and catalase, indicating that this medicinal plant has the potential to reduce t-HCY levels in vivo.
In addition, other investigators [51-53] reported that catechin, taurine and quercetin supplementation are effective in attenuating the increase of serum HCY level as induced by a Met-enriched diet in rats and mice respectively. The protective effect of argan oil is probably due to its high contents of powerful antioxidants, particularly polyphenols,tocopherols and sterols, which are known as powerful antioxidants [54]. Indeed we demonstrated that powdered seeds of A. spinosa when given in combination with high methionine diet increased significantly the catalase activity, but not significantly the GSH level, indicating its benefic effect in prevention against oxidative stress in vivo. In accordance with our results, [20] demonstrated that argan oil treatment increased the GSH against mercuric chloride induced oxidative stress in experimental rats. The elevated level of GSH protects cellular proteins against oxidation through glutathione redox cycle and directly detoxifies reactive species [55]. Our results showed an increase of catalase activities in group (MP) which administered with L-methionine (500 mg/kg) and treated with A. spinosa (150 mg/kg), these results are in agreement with those of [56], who showed that the activities of cytosolic CAT were significantly higher in Wistar rats treated with argan oil in comparison with untreated rats.
The work of [57, 40] proved that the rats and mice respectively administered with 200 mg/kg during 21 d could damage the aorta and heart tissue and the treatment of these animals with vitamins B9, B12 and B6 and Stachys mialhesi extract corrected these alterations. Another work of [58] established that the high level of Hcy could stimulated the angiogenesis on the arota of rats, and the treatment with the extracts of medicinal plants Stachys mialhesi and Chrysanthemum Macrocarpum could inhibited the angiogenesis.
Antioxidants are emerging as prophylactic and therapeutic agents [59]. Furthermore, another study of [60] suggested that an increased intake of antioxidants appeared to be protective in cardiovascular diseases. Epidemiological studies have shown that consumption of food and beverages rich in phenols can reduce the risk of heart disease by slowing the progression of atherosclerosis principally by protecting LDL from oxidation [61].
CONCLUSION
The current study has shown that the powdered seeds of A. spinosa were effective in attenuating the increase of HCY level, improved the antioxidants defence and prevented the endothelial, cardiac alterations, as induced by a Met-enriched diet in mice. It may be
interesting in the development of new drugs for cardiovascular diseases induced by hyperhomocysteinemia.
ACKNOWLEDGEMENT
The authors are grateful to the MESRS (Ministery of Scientific Research, Algeria).
AUTHORS CONTRIBUTION
BADIAA Aklil carried out the experimental part of the work, performed data analysis and drafted and revised the manuscript. The design of the work and correction of the manuscript was done by the corresponding author Mrs ZERIZER Sakina and ZAHIA Kabouche carried out the phytochemical part. All authors read and approved the final manuscript.
CONFLICT OF INTERESTS
Declared none
REFERENCES
1. Selicharova I, Korinek M, Demianova Z, Chrudinova M, Mladkova J, Jiracek J. Effects of hyperhomocysteinemia and betaine-homocysteine S-methyltransferase inhibition on hepatocyte metabolites and the proteome. Biochim Biophysica Acta 2013;1834:1596–606.
2. Agoston-Coldea L, Mocan T, Gatfosse M, Lupu S, Dumitrascu DL. Plasma homocysteine and the severity of heart failure in patients with previous myocardial infarction. Cardiol J 2011;18:55–62.
3. Wang X, Cui L, Joseph J, Jiang B, Pimental D, Handy DE. Homocysteine induces cardiomyocyte dysfunction and apoptosis through p38 MAPK mediated increase in oxidant stress. J Mol Cell Cardiol 2012;52:753–60.
4. Faraci FM, Lentz SR. Hyperhomocysteinemia, oxidative stress, and cerebral vascular dysfunction. Stroke 2004;35:345-7.
5. Clarke R, Daly L, Robinson K. Hyperhomocysteinemia: an independent risk factor for vascular disease. N Engl J Med 1999;324:1149–55.
6. Adinolfi LE, Ingrosso D, Cesaro G, Cimmino A, D’Anto M, Capasso R, et al. Hyperhomocysteinemia and the MTHFR C677T polymorphism promote steatosis and fibrosis in chronic hepatitis C patients. Hepatology 2005;41:995–1003.
7. Roblin X, Pofelski J, Zarski JP. Steatosis, chronic hepatitis virus C infection and homocysteine. Gastroenterol Clin Biol 2007;31:415–20.
8. Glushchenko AV, Jacobsen DW. Molecular targeting of proteins by L-homocysteine: mechanistic implications for vascular disease. Antioxid Redox Signal 2007;9:1883–98.
9. Hogg N. The effect of cysteine on the auto-oxidation of homocysteine. Free Radical Biol Med 1999;27:28-33.
10. Stipanuk MH. Sulfur amino acid metabolism: pathways for production and removal of homocysteine and cysteine. Annu Rev Nutr 2004;24:539-77.
11. Eberhardt RT, Forgione MA, Cap A, Leopold JA, Rudd MA, Trolliet M, et al. Endothelial dysfunction in a murine model of mild hyperhomocysteinemia. J Clin Invest 2000;106:483–91.
12. Zhang R, Ma J, Xia M, Zhu H, Ling W. Mild hyperhomocysteinemia induced by feeding rats diets rich in methionine or deficient in folate promotes early atherosclerotic inflammatory processes. J Nutr 2004;134:825–30.
13. Afaf A, Sayed S. Lipid profile and levels of homocysteine and total antioxidant capacity in plasma of rats with experimental thyroid disorders. J Basic Appl Zool 2015;72:173–8.
14. Jacobsen DW. Hyperhomocysteinemia and oxidative stress: time for a reality check? Arterioscler Thromb Vascular Biol 2000;20:1182–4.
15. Landmesser U, Dikalov S, Price SR, McCann L, Fukai T, Holland SM, et al. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J Clin Invest 2003;111:1201–9.
16. Cha JY, Repa J. The liver X receptor (LXR) and hepatic lipogenesis: the carbohydrate-response element-binding protein is a target gene of LXR. J Biol Chem 2007;282:743-51.

Zerizer et al.
Int J Pharm Pharm Sci, Vol 9, Issue 12, 64-69
69
17. Denechaud PD, Bossard P, Lobaccaro JM, Millatt L, Staels B, Girard J. ChREBP, but not LXRs, is required for the induction of glucose-regulated genes in mouse liver. J Clin Invest 2008;118:956-64.
18. Dentin R, Benhamed F, Hainault I, Fauveau V, Foufelle F, Dyck JR. Liver-specifi c Inhibition of ChREBP improves hepatic steatosis and insulin resistance in ob/ob mice. Diabetes 2006;55:2159-70.
19. Charrouf Z, Guillaume D. Ethnoeconomical, ethnomedical, and phytochemical study of Argania spinosa (L.) Skeels. J Ethnopharmacol 1999;67:7–14.
20. Necib Y, Bahi A, Zerizer S. Argan oil (Argania spinosa L) Provides protection against mercuric chloride induced oxidative stress in rat Albinos Wistar. Int J Basic Appl Sci 2013;2:73-80.
21. Cherki M, Berrougui H, Drissi A, Adlouni A, Khalil A. Argan oil: which benefits on cardiovascular diseases? Pharmacol Res 2006;54:1–5.
22. Bradford MA. Rapid and sensitive method for the quantities of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248-54.
23. Weckbercker G, Cory JG. Ribonucleotide reductase activity and growth of glutathione-depended mouse leukaemia L 1210 cells in vitro. Cancer Lett 1988;40:257-64.
24. Aebi H. Catalase: methods for enzymatic analysis. Acadamic Press 1974;2:674- 84.
25. Curro M, Gugliandolo A, Gangemi C, Risitano R, Ientile R, Caccamo D. Toxic effects of mildly elevated homocysteine concentrations in neuronal-like cells. Neurochem Res 2014;39:1485–95.
26. Faeh D, Chiolero A, Paccaud F. Homocysteine as a risk factor for cardiovascular disease: should we (still) worry about it? Swiss Med Wkly 2006;136:745–56.
27. Baszczuk A, Kopczynski Z. Hyperhomocysteinemia in patients with cardiovascular disease. Postepy Hig Med Dosw 2014;68:579.
28. Mudd SH. Hypermethioninemias of genetic and non-genetic origin: a review. J Med Genet C Semin Med Genet 2011;157:3–32.
29. Viggianoa A, Viggianoa E, Mondaa M, Ingrossob D, Pernac AF, Luca BD. Methionine-enriched diet decreases hippocampal antioxidant defences and impairs spontaneous behaviour and long-termpotentiation in rats. Brain Res 2012;1471:66-74.
30. Yamada H, Akahoshi N, Kamata S, Hagiya Y, Hishiki T, Nagahata Y, et al. Methionine excess in diet induces acute lethal hepatitis in mice lacking cystathionine γ-lyase, an animal model of cystathioninuria. Free Radical Biol Med 2012;52:1716-26.
31. Kirac D, Negis Y, Ozer NK. Vitamin E attenuates homocysteine and cholesterol induced damage in rat aorta. Cardiovascular Pathol 2013;22:465–72.
32. Welch GN, Upchurch GR, Farivar RS, Pigazzi A, Vu K, Brecher P. Homocysteine-induced nitric oxide production in vascular smooth muscle cells by NF-kappaB dependent transcriptional activation of Nos2. Proc Assoc Am Physicians 1998;110:22–31.
33. Stipanuk MH, Dominy JE, Lee JI, Coloso RM. Mammalian cysteine metabolism: new insights into the regulation of cysteine metabolism. J Nutr 2006;136:1652S-59S.
34. Milton NGN. Homocysteine inhibits hydrogen peroxide breakdown by catalase. Open Enzyme Inhibition J 2008;1:34–41.
35. Góth L, Rass P, Pay A. Catalase enzyme mutations and their association with diseases. Mol Diagn 2004;8:141-9.
36. Ho YS, Xiong Y, Ma W, Spector A, Ho DS. Mice lacking catalase develop normally but show differential sensitivity to oxidant tissue injury. J Biol Chem 2004;279:32804-12.
37. Margoliash E, Novogrodsky A. A study of the inhibition of catalase by 3-amino-1:2:4:-triazole. Biochem J 1958;68:468-75.
38. Putnam CD, Arvai AS, Bourne Y, Tainer JA. Active and inhibited human catalase structures: ligand and NADPH binding and catalytic mechanism. J Mol Biol 2000;296:295-309.
39. Milton NGN. Amyloid-ß binds catalase with high affinity and inhibits hydrogen peroxide breakdown. Biochem J 1999;344:293-6.
40. Benmebarek A, Zerizer S, Laggoune S, Kabouche Z. Effect of Stachys mialhesi de Noé on the inflammation induced by hyperhomocysteinemia in cardiovascular diseases. Scholar Res Library 2013;5:212-23.
41. Gallai V, Caso V, Paciaroni M, Cardaioli G, Arning E, Bottiglieri T, et al. Mild hyperhomocysteinemia: a possible risk factor for cervical artery dissection. Stroke 2001;32:714–8.
42. Papatheodorou L, Weiss N. Vascular oxidant stress and inflammation in hyperhomocysteinemia. Antioxid Redox Signal 2007;9:1941–58.
43. Lamda S, Aggoun C, Naimi D. The effects of homocysteine on plasma biochemical parameters and aortic matrix metalloproteinases activities. Innov Acad Sci 2014;7:975-491.
44. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev 2007;87:315–424.
45. Cooke JP, Dzau VJ. Nitric oxide synthase: role in the genesis of vascular disease. Annu Rev Med 1997;48:489–509.
46. Kanani PM, Sinkey CA, Browning RL, Allaman M, Knapp HR, Haynes WG. Role of oxidant stress in endothelial dysfunction produced by experimental hyperhomocysteinemia in humans. Circulation 1999;100:1161–8.
47. Rolland PH, Friggi A, Barlatier A. Hyperhomocysteinemia-induced vascular damage in the minipig. Captopril-hydrochlorothiazide combination prevents elastic alterations. Circulation 1995;91:1161–74.
48. Zulli A, Hare DL. High dietary methionine plus cholesterol stimulates early atherosclerosis and late fibrous cap development which is associated with a decrease in GRP78 positive plaque cells. Int J Exp Pathol 2009;90:311–20.
49. Raghuveer G, Sinkey CA, Chenard C, Stumbo P, Haynes WG. Effect of vitamin E on resistance vessel endothelial dysfunction induced by methionine. Am J Cardiol 2001;88:285–90.
50. Barbato JC, Catanescu O, Murray K, DiBello PM, Jacobsen DW. Targeting of metallothionein by L-homocysteine: a novel mechanism for disruption of zinc and redox homeostasis. Arterioscler Thromb Vasc Biol 2007;27:49–54.
51. Hamelet J, Demuth K, Dairou J, Ledru A, Paul JL, Dupret JM. Effects of catechin on homocysteine metabolism in hyperhomocysteinemic mice. Biochem Biophys Res Commun 2007;355:221–7.
52. Yalc S, Unlucerci Y, Giris M, Olgac V, Dogru-Abbasoglu S, Uysal M. Oxidative and nitrosative stress and apoptosis in the liver of rats fed on high methionine diet: protective effect of taurine. J Nutr 2009;25:436–44.
53. Meng MD, Weina MD, Ph D, Jingyu BS, Jijun BS, Jianquan MD. Quercetin reduces serum homocysteine level in rats fed a methionine-enriched Diet. J Nutr 2013;29:661–6.
54. Masella R, Giovannini C, Vari R, Di Benedetto R, Coni E, Volpe R. Effect of dietary virgin olive oil phenols on low-density lipoprotein oxidation in hyperlipidemic patients. Lipids 2001;36:1195–202.
55. Ketterer B. Glutathione-S-transferase and prevention of cellular free radical damage. Free Radical Resear 1998;28:647-58.
56. Benajiba N, Morel S, De Leiris J, Boucher F, Charrouf Z, Mokhtar N. The effect of argan oil on heart function during ischemia and reperfusion. Therapie 2002;57:246-52.
57. Zerizer S, Naimi D. Homocysteine: an independent risk factor in the atherogenic process. Egyptian Pharm J 2004;3:110-4.
58. Zerizer S, Zaher K, Boutaghane N, Laggoune S, Kabouche Z. Comparative effect of Chrysanthemum Macrocarpum and Stachys Mialhesi on the rat's aorta exposed to homocysteine with B Vitamins. Scottish J Arts Soc Sciences Scientific Studies 2008;6:2047-1278.
59. Kale MA, Bindu SM, Khadkikar P. Role of antioxidants and nutrition in oxidative stress: a review. Int J Appl Pharm 2015;7:975-7058.
60. kumar SV, Saritha G, Fareedullah Md. Role of antioxidants and oxidative stress in cardiovascular diseases. A Biol Res 2010;1:158-73.
61. Duffy SJ, Vita JA. Effects of phenolics on vascular endothelial function. Curr Opin Lipidol 2003;14:21–7.

Appendices

1- Treatment dose calculation
A. spinosa crude extract given dose (150 mg/kg)
0.15 g 1000g
X g Mouse weight g
A. spinosa crude extract given dose
Methionine given dose (500 mg/kg)
0.50 g 1000g
X g Mouse weight g
Methionine given dose
2- Hematoxylin eiosin staining:
Dip slides in alcohol for 5 minutes;
Rinse with water;
Stain slides in hematoxylin for 4 minutes;
After rinsing, stain with eosin for 10 minutes;
Rinse with water;
Dip slides in alcohol for 1 minute;
After rinsing and drying, the editing is done using xylene
3- PBS preparation
Nacl : 8g
Kcl : 0,2g
NaH2PO4 : 1,15g
KH2PO4 : 0,2g
Mgcl2 6H 2O: 0,1g
Cacl 2 2H 2O : 0,137g
QS: 1 L of water

Table 1: 21 days of average weight of mice.
Groups
D1
D2
D3
D4 D5 D6 D7 D8 D9 D10 D11 D12 D13 D14 D15 D16 D17 D18 D19 D20 D21
(F) :
Average
weight
(g)
32,10 30,18 28,22 27,82 27,82 27,40 26,98 27,20 27,37 23,88 27,00 26,83 26,85 26,88 26,92 26,92 27,22 26,72 31,70 32,34 32,98
(M) :
Average
weight
(g)
35,01 34,46 33,91 33,67 33,27 33,47 33,88 33,12 33,46 33,81 34,33 33 34,1 34,1 34,39 31,41 28,44 33,59 33,89 34,79 32,04
(MP) :
Average
weight
(g)
31,84 31,74 32,32 32,08 31,7 31,77 32,05 32,33 31,76 32,24 32,49 29,3 28,17 27,03 32,16 31,56 30,41 32,06 32,8 30,45 28,1
(P) :
Average
weight
(g)
30,58 29,97 30,32 30,66 30,74 30 30,81 30,93 31,17 30,86 30,6 31,1 30,2 30,86 30,69 30,51 30,21 29,91 30,33 30,77 31,19

Abstract
Hyperhomocysteinemia (HHcy), oxidative stress and decreased antioxidant capacities
lead to several clinical manifestations and particularly, cardiovascular and liver diseases.
In this study, we evaluated the protective and the preventive effect of the crude extract
of Argania spinosa against HHcy, hyperlipidemia, oxidant status and damages in the aorta,
heart and liver induced by high L-methionine intake in mice.
After 3 weeks of treatments, Hcy concentrations, lipid parameters, liver enzyme
activities, hepatic antioxidant status and histological sections of aorta, liver and heart were
determined. Our results showed that consumption of high L-methionine diet (500mg/Kg) led
to an increase in plasma Hcy, CHO, LDL-c, TG, AST, corresponding with decrease of HDL-
c, reduced GSH, and catalase activity. These were associated with the loss and degeneration
of endothelium, fenestration and formation of foam cells in the media of the aorta, also the
alteration of the cardiac muscle and liver tissue. However, the administration of the crude
extract of A. spinosa (150mg/Kg) in combination with L-methionine ameliorated all these
changes.
Moreover, in the present thesis, we conducted a study in vitro to evaluate the
antibacterial effect of Argan oil on biofilm formation.
The results showed that there was a trend of increasing inhibition of 4 species of
bacteria (Streptococcus mutans, Streptococcus anginosus, Streptococcus intermedius, and
staphylococcus haemolyticus), that belong to gram positive strain as the Argan oil was getting
more concentrated.
Keywords: Homocysteine, Argania spinosa , Antioxidant enzymes, Methionine, Oxidative
stress, Cardiovascular diseases, biofilm.

Résumé
L'hyperhomocystéinémie (HHcy), le stress oxydatif et la diminution des capacités
antioxydantes entraînent plusieurs manifestations cliniques et en particulier des maladies
cardiovasculaires et hépatiques.
Dans cette étude, nous avons évalué l'effet protecteur et préventif de l'extrait brut d’A.
spinosa contre l'HHcy, l'hyperlipidémie, le statut oxydant et des lésions de l'aorte, du cœur
et du foie induits par une forte prise de méthionine chez la souris.
Après 3 semaines de traitement, les concentrations de Hcy, les paramètres lipidiques,
les activités des enzymes hépatiques, le statut antioxydant hépatique et les sections
histologiques de l'aorte, du foie et du cœur ont été déterminés. Nos résultats ont montré que la
consommation d'une diète riche en méthionine (500 mg / kg) peut entraîner une
augmentation des taux de Hcy, CHO, LDL-c, TG, AST, avec une diminution des
concentrations des HDL-c, GSH réduit et des catalases, associés à la perte et à la
dégénérescence de l'endothélium, à la fenestration et à la formation de cellules spumeuses
dans l'aorte, ainsi qu'à l'altération du muscle cardiaque et du tissu hépatique. Cependant,
l'administration de l'extrait brut d’A. spinosa (150 mg / kg) en combinaison avec de la
méthionine a amélioré tous ces changements.
De plus, dans le présent travail, nous avons mené à une étude in vitro pour évaluer
l'effet antibactérien de l'huile d'Argan sur la formation de biofilm.
Les résultats ont montré qu'il y avait une inhibition croissante de 4 espèces de
bactéries (Streptococcus mutans, Streptococcus anginosus, Streptococcus intermedius, and
staphylococcus haemolyticus), appartenant à la souche Gram positif, lorsque l'huile d'Argan
devenait plus concentrée.
Mots-clés: Homocystéine, Argania spinosa, Antioxydant, Méthionine, Stress
oxydant, Maladies cardiovasculaires, Biofilm.

ملخص
وانخفاض مضادات األكسدة تؤدي إلى ظهور سديكاإلجهاد التأ ،فرط الهوموسيستيين في الدم
.األمراض خاصة أمراض القلب واألوعية الدموية والكبد من العديد
A.spinosa رغان ور نبتة األذفي هذه الدراسة، قمنا بتقييم التأثير الوقائي للمستخلص الخام لب
األكسدة باإلضافة إلى مضاداتالهوموسيستيين، ارتفاع مستوى الدهون في الدم وضد ارتفاع مستوي
القلب والكبد الناجمة عن ارتفاع تناول جرعات عالية من ،األضرار النسيجية في الشريان األورطي
.الميثيونين لدى الفئران
،إنزيمات الكبد ،ليبيداتتحاليل ال، الهوموسيستيين بعد ثالثة أسابيع من العالج، تم قياس مستويات
. الكبد والقلب،الشريان األورطي تحضير القطاعات النسيجية لكل من كما تم األكسدةنشاط مضادات
الهوموسيستيين أدى إلى ارتفاع( كغ/مغ/ 055)ميثيونين بالنتائج أن استهالك نظام غذائي غني الأظهرت
، (AST) و (TG) الدهون الثالثية ، (LDL-c)المنخفض الكثافةالكولسترول و الكوليسترول، في الدم
الجلوثاثيون، و نشاط انزيم ، (HDL-c)العالي الكثافةالكولسترول انخفاض في مستوىمع تسجيل
تزامنت هذه النتائج مع فقدان وتدهور البطانة ، نوفذة وتشكيل خاليا رغوية في الشريان . الكاتاالز
كل من اخذ أنفي حين بينت النتائج . وأيضا تم تسجيل تغيير في عضلة القلب وأنسجة الكبد األورطي،
.تحسين كل هذه التغييرات إلى أدىمع الميثيونين معا ( كغ/مغ005)األرغان ور ذالمستخلص الخام لب
من جهة أخرى ، أجرينا دراسة أخرى من اجل تقييم تأثير مضاد للجراثيم لزيت األرغان ضد
.شكيل البيوفيلمت
Streptococcus)أنواع من البكتيريا 4وقد أظهرت النتائج أن زيت األرغان أدى إلى تثبيط
mutans, Streptococcus anginosus, Streptococcus intermedius, and
(Staphylococcus haemolyticus,إيجابية و ذلك كلما كان أكثر غرام كتيرياي، التي تنتمي إلى ب
.تركيزا
:الكلمات المفتاحية
، سديكاإلجهاد التأ ، اإلنزيمات المضادة لألكسدة، ميثيونين،Argania spinosaالهوموسيستيين،
.أمراض القلب واألوعية الدموية، البيوفيلم

Academic year: 2017-2018 Family Name: AKLIL
First Name: BADIAA
Title: The effect of Argania spinosa on plasma Homocysteine, Lipids,
Antioxidant enzymes and Aortas sections in methionine induced
Hyperhomocysteinemia in mice
Thesis submitted for the degree of DOCTORAT IN SCIENCES
Hyperhomocysteinemia (HHcy), oxidative stress and decreased antioxidant capacities
lead to several clinical manifestations and particularly, cardiovascular and liver diseases.
In this study, we evaluated the protective and the preventive effect of the crude
extract of Argania spinosa against HHcy ,hyperlipidemia, oxidant status and damages in
the aorta, heart and liver induced by high L-methionine intake in mice.
After 3 weeks of treatments, Hcy concentrations, lipid parameters, liver enzyme
activities, hepatic antioxidant status and histological sections of aorta, liver and heart were
determined. Our results showed that consumption of high L-methionine diet (500mg/Kg)
led to an increase in plasma Hcy, CHO, LDL-c, TG, AST, corresponding with decrease of
HDL-c, reduced GSH, and catalase activity. These were associated with the loss and
degeneration of endothelium, fenestration and formation of foam cells in the media of the
aorta, also the alteration of the cardiac muscle and liver tissue. However, the administration
of the crude extract of A. spinosa (150mg/Kg) in combination with L-methionine
ameliorated all these changes.
Moreover, in the present thesis, we conducted a study in vitro to evaluate the
antibacterial effect of Argan oil on biofilm formation.
The results showed that there was a trend of increasing inhibition of 4 species of
bacteria (Streptococcus mutans, Streptococcus anginosus, Streptococcus intermedius, and
staphylococcus haemolyticus), that belong to gram positive strain as the Argan oil was
getting more concentrated.
Keywords: Homocysteine, Argania spinosa, Antioxidant enzymes, Methionine, Oxidative
stress, Cardiovascular diseases, biofilm.






![INDEX [decaar.com] · to tone the skin. Finish your daily ... Argania Spinosa Kernel exract, Menthol Retail: ... Perfect application for skin conditions like eczema or damaged skin](https://img.pdfslide.net/doc/110x75/5b23fd1f7f8b9a3f6d8b45a0/index-to-tone-the-skin-finish-your-daily-argania-spinosa-kernel-exract.jpg)




![Serum Levels of Homocysteine, Vitamin B12 and Folate in ... · methyltetrahydrofolate and methyl-Vitamin-B12 are essential factors for methionine synthesis of Hcy [10]. Lacking Vitamin](https://img.pdfslide.net/doc/110x75/5ec906dfa105b02e13239827/serum-levels-of-homocysteine-vitamin-b12-and-folate-in-methyltetrahydrofolate.jpg)