Embed Size (px)
Citation preview

The Evolutionary History of YAP and the Hippo/YAP Pathway
Dror Hilman1 and Uri Gat*,1
1Department of Cell and Developmental Biology, The Alexander Silberman Institute of Life Sciences, Edmond Safra Campus atGivat Ram, The Hebrew University, Jerusalem, Israel
*Corresponding author: E-mail: [email protected].
Associate editor: Billie Swalla
Abstract
The Hippo/YAP pathway plays an important role in animal organ size control, which it exerts by regulating tissueproliferation and apoptosis rates as a response to developmental cues, cell contact, and density. With the ever increasingadvance in genome sequencing and analysis tools, our understanding of the animal world and its evolution has greatlyincreased in the recent years. We used bioinformatic tools to study the evolution of the Hippo/YAP pathway focusing onthe transcriptional coactivator YAP, which is a pivotal effector of the pathway. The aim was to establish the origin andmode of development of YAP and its pathway in the animal world. Some pathway members can be already identified insingle-celled eukaryotes like the yeast that have preceded multicellular animals. Interestingly, we can find most of thecomponents that are present in human in the sea-anemone Nematostella, which belongs to a very basal group ofmetazoans, the cnidarians. All the major domains of YAP have been conserved between cnidarians and mammals, and YAPcan be identified even in the more basal placozoan clade. We show a very high degree of conservation in regions such asthe WW and the TEAD-binding domains, TEAD being the major DNA-binding partner of YAP. Remarkably, we found thatthe location of an intron in the WW1 genomic region has been invariant along an evolutionary span of over 700 My. Wehave followed the evolutionary changes in YAP and in other main components of the pathway from the first metazoanssuch as sponges, described the phylogenetic relationships between the YAP genes and indicated where YAP and othercomponents have been secondarily lost. Evidence is provided that YAP and its binding partner TEAD demonstrate strongcoevolution. This gives further support for the importance of the TEAD–YAP association. Beyond contributing to anunderstanding of the evolutionary history of this pathway, we have provided insights into the ‘‘birth’’ of this pathway, itsfunctions and its mode of operation in animals with different body plans, development, and life styles.
Key words: YAP, yorkie, Hippo/YAP pathway, organ size control, evolution, coevolution.
IntroductionA fundamental aspect of the development of multicellularorganisms is the ability to control the size and shape oftheir tissues and organs. In recent years, the abundanceand wealth of genomic data and molecular biology techni-ques have revealed a ‘‘toolkit’’ of developmental genes,which served in generating the remarkable diversity ofthe body plans of organisms (De Robertis 2008; Kinget al. 2008; Degnan et al. 2009; Srivastava et al. 2010). Itappears that all the major animal groups share a subsetof developmental pathways. These pathways are repeatedlyemployed over vast spans of evolutionary time to buildever more complex forms, using similar building blocks(De Robertis 2008). One of the most intriguing phenomenaof metazoan development is their ability to precisely con-trol organ size by coordinating cell proliferation rate duringembryogenesis and maturation and in certain cases duringregeneration and wound healing processes (Dong et al.2007; Kango-Singh and Singh 2009).
In the past decade, a new pathway that controls cell pro-liferation and cell apoptosis in response to developmentalcues has been described (reviewed in Harvey and Tapon2007; Kango-Singh and Singh 2009; Grusche et al. 2010;Oh and Irvine 2010; Sudol and Harvey 2010; Zhao et al.2010). This pathway is termed the Hippo or the Hippo/
YAP pathway, after some of the major components thatconstitute it. The pathway includes proteins involved in re-ception of signals, signaling mediators, a transcription co-activator, and executing transcription factors (see fig. 1 fora simplified scheme; Justice et al. 1995; Xu et al. 1995; Taponet al. 2002; Harvey et al. 2003; Pantalacci et al. 2003; Wuet al. 2003; Huang et al. 2005; Kango-Singh and Singh2009). It has been hypothesized that several upstream reg-ulators are involved in producing growth arrest signalsupon stimulation. These include: 1) The arthropod/mam-malian planar cell polarity protocadherins Fat/Ft1-4 anddachsous/DCHS, 2) The FERM-domain proteins Ex-panded/FRMD6/Willin and Merlin/NF2, 3) The WW-do-main protein Kibra, the Ras-familly protein dRASSF/RASSF1-10, apico-basal polarity proteins Lgl/Lgl1/2, atypi-cal protein kinase C, and Crumbs/Crb1-3. The signals eli-cited are transmitted through the kinases Hpo/Mst1,2and Warts/Lats1,2, with the help of the adaptor proteinsSalvador/WW45 and Mats/Mob to the coactivators Yki/YAP and TAZ. These induce the growth program by acti-vating several transcription factors and micro RNAs. Themajor YAP/TAZ transcriptional partners in the program,which provide the DNA-binding function, were shownto be Scalloped/TEAD1-4, but others like Runt/Runx andp73 have been reported as well (Strano et al. 2001, 2005;Levy et al. 2007; Vitolo et al. 2007). The phosphorylation
© The Author 2011. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. All rights reserved. For permissions, pleasee-mail: [email protected]
Mol. Biol. Evol. 28(8):2403–2417. 2011 doi:10.1093/molbev/msr065 Advance Access publication March 16, 2011 2403
Research
article

of Yki/YAP by Warts/Lats1,2 has been found to enhancethe cytoplasmic localization of Yki/YAP preventing or ter-minating its coactivation activity in the nucleus (reviewedin Oh and Irvine 2010; Zhao et al. 2010). Importantly, it isnow well established that malfunctioning of the signalingpathway, which phosphorylates YAP and prevents its pro-liferative and antiapoptotic activity, can lead to dispropor-tionate organ growth and tumorigenesis, which mayultimately develop into cancer (Hisaoka et al. 2002; Taponet al. 2002).
The conservation of this pathway from mammals to in-sects suggests that its components have deep evolutionaryroots. Here, we examined the evolution of the Hippo/Yki/YAP pathway by following the phylogeny of its core com-ponents focusing on Yki/YAP, the crucial effector switch ofthe pathway. We looked for evidence for the evolutionaryorigin of the pathway, examined the basal form of YAP, itsmost conserved components, and the changes in YAP
across distant phyla. Our results shed light on the sourceof this pathway, its major forms and modifications alonganimal evolution. They also may provide novel insights intowhy the pathway has emerged and into the modes of itsevolution.
Methods
Finding Orthologs for YAP Pathway ProteinsTo find orthologs of Hippo/YAP pathway genes, we em-ployed a BlastP (Altschul et al. 1990) search of the humanproteins: FAT4, DCHS1, CSNK1E, RASSF1 FRMD6, NF2(Merlin), KIBRA, MOB1, MST1/2, LATS1/2, YAP, TAZ,and TEAD1 from refseq (Pruitt et al. 2007; table 1). Otherpathway associated proteins were analyzed as well. Theseincluded p73, RUNX1-3 AJUBA and c-ABL, as well as thehedgehog pathway proteins, SHH, SMO, PTCH, andGLI1-3. The later were used as controls. All human sequen-ces were searched against the refseq protein data from dif-ferent organisms, representing different metazoan andnonmetazoan groups. All blast hits were filtered, and onlysequences with blast score.150 and sequence length.50were examined. We then tested each sequence using recip-rocal blast search and assigned it as an ortholog if the besthit of one blast search matched the best hit of the other(supplementary data 1, Supplementary Material online). Ifthe sequences were not found reciprocally in two genomes,we picked the sequence with the best coverage,with score .150, relative identity .30%, and relative sim-ilarity.40%. If the parameters were lower, we assigned it as‘‘nonortholog.’’ When we did not find an ortholog, we ver-ified the lack of orthologous sequences with tBlastX againstthe genomic sequence and expressed sequence tags (EST)libraries of the relevant organism. The existence or absenceof an ortholog was also checked by blast search of the pro-tein sequence in the Ortho-MCL4 database of orthologs (Liet al. 2003; Chen et al. 2007). Sequence alignment of severalHippo/YAP pathway components is illustrated in supple-mentary data 2, Supplementary Material online.
Sequence Alignment and Phylogenetic AnalysisSequence alignments were performed on orthologous pro-teins using MUSCLE (Edgar 2004) with default parameters(supplementary data 2, Supplementary Material online),which were manually curated if necessary. Conservationof motifs and phosphorylation sites were checked by handusing data from Entrez gene (Maglott et al. 2007) andUniprot (Consortium 2010). Phylogenetic analysis was con-ducted using PhyML (Guindon and Gascuel 2003;Guindon et al. 2005) and FastTree (Price et al. 2010) formaximum likelihood (ML) analysis, with default parame-ters. Neighbor joining (NJ) trees were built using MEGA4(Kumar et al. 2008). Phylogeny support was verified withthe bootstrap consensus tree inferred from 1,000 replicates.In NJ trees, the evolutionary distances were computed us-ing the Poisson correction method and are given in units ofthe number of amino acid substitutions per site. Therate variation among sites was modeled with a gamma
FIG. 1. Model of the core Hippo/YAP pathway. The Hippo/YAPpathway controls tissue growth by regulating the subcellularlocalization of Yki/YAP/TAZ, which, when nuclear, associate withthe transcription factors Sd/TEAD1-4 activating a transcriptionalprogram that drives cell growth and proliferation and inhibitsapoptosis. Other transcriptional partners such as p73, Runx, andSmads have been described as well. Several upstream inputsregulate the activity of the core components Salvador (Sav), Hpo/Mts, Wts/Lats, and Mats/Mob. These components control Yki/YAP/TAZ phosphorylation and localization. Rassf modulate theMst/Hippo kinases activity. The protocadherin Ft/FAT signals tothe core components by influencing the localization and levels ofExpanded (Ex/FRMD6) that forms a complex with Merlin (Mer)and Kibra to facilitate and activate the Hpo/Sav/Mats/Latscomplex. Fat may inhibit Dachs, which represses Lts/Warts proteinlevels. Fat activity is controlled by its ligand Dachsous (Ds), by thekinases Four-jointed (Fj), and Discs overgrown (Dco) and by LowFat (Lft), Approximated (App), and Dachs (Ds).
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2404

distribution using default parameters. Positions containingalignment gaps and missing data were eliminated only inpairwise sequence comparisons so as to keep all majorprotein domains in the alignment.
Testing CoevolutionTo assess the strength of coevolution between two pro-teins, we followed the methodology described in Pazoset al. (2005) and Sato et al. (2006). Pairwise distances be-tween sequences in different taxa were measured inMEGA4 with pairwise deletion and Poisson correctionwas used for amino acids substitution. Distances betweendifferent proteins in diverse organisms were plotted, andPearson’s correlation coefficient (product–moment r)was used to determine the overall correlation between dis-tances of two genes (supplementary data 3, SupplementaryMaterial online).
Retrieving Genomic DataAll the genomic data used in this analysis were retrievedfrom Entrez genome and Entrez nucleotide databases.Protein sequences were retrieved from the NationalCenter for Biotechnology Information Refseq database.Orthologs for comparing YAP and TAZ were obtainedfrom OrthoMCL 4 DB to retrieve a large group ofvertebrate orthologs.
Results and Discussion
YAP Proteins—A Metazoan NoveltyDetecting the evolutionary origin of genes that belong tospecific pathways may give insights into how they havecontributed to the formation of novel structures andmechanisms in animals over the course of evolution.YAP orthologs have previously been explored almost exclu-sively in higher vertebrates and in Drosophila. We set out tofind the origin of YAP and the Hippo/YAP pathway in thetree of life (fig. 2). We first explored the YAP protein, a crit-ical connecting switch between the signaling pathway andthe transcriptional program. We could not detect a bonafide YAP gene in the group that is considered to representthe most basal or ‘‘ancient’’ metazoans—the sponges.Searching for conserved regions of YAP in the demosponge
Amphimedon queenslandica whole-genome shotgunsequencing (Srivastava et al. 2010) and in the homoscler-omorph sponge Oscarella carmela EST database (Nicholset al. 2006; Hemmrich and Bosch 2008) did not yield anobvious ortholog. Actually, proteins with two conservedWW domain proteins in sponges and in the nonmetazoaneukaryotes choanoflagellates Monosiga brevicollis (Kinget al. 2008) usually aligned best with the human geneWWP1–E3 ubiquitin ligase, which seems to be conservedin nearly all eukaryotes we tested. Searching further, YAPorthologs were not found in yeasts (Saccharomyces cerevi-siae) or in plants (Arabidopsis thaliana). The most basal
Table 1. The Hippo/YAP Pathway Protein Components Studied in This Article.
Protein Human Refseq Acc. Drosophila Refseq Acc.
DCHS1/ds NP_003728.1 NP_523446.2FAT4/ft NP_078858.4 NP_477497.1CSNK1/Dco NP_001885.1 NP_733414.1RASSF1/Rassf NP_009113.3 NP_651126.3NF2/Merlin (Mer) NP_000259.1 NP_523413.1WWC1/Kibra NP_001155133.1 NP_001034055.1FRMD6/expanded (ex) NP_001035946.1 NP_476840.2Mst1/2/Hippo NP_006273.1/NP_006272.2 NP_611427.1Salvador (Sav) NP_068590.1 NP_788721.1Mob1/Mats NP_775739.1 NP_651041.3Lats1/2/Warts NP_004681.1/NP_055387.2 NP_733403.1YAP/TAZ/Yorkie NP_001123617.1/NP_001161750.1 NP_001036568.2TEAD1/scalloped (sd) NP_068780.2 NP_001096989.1
FIG. 2. The evolutionary tree of taxa studied in this work. The grossestimated divergence time ranges of the major clades arementioned near each group name. Divergence times are based onOtsuka and Sugaya (2003), Pisani et al. (2004), Nichols andWorheide (2005), Hejnol et al. (2009), and Han et al. (2010).
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2405
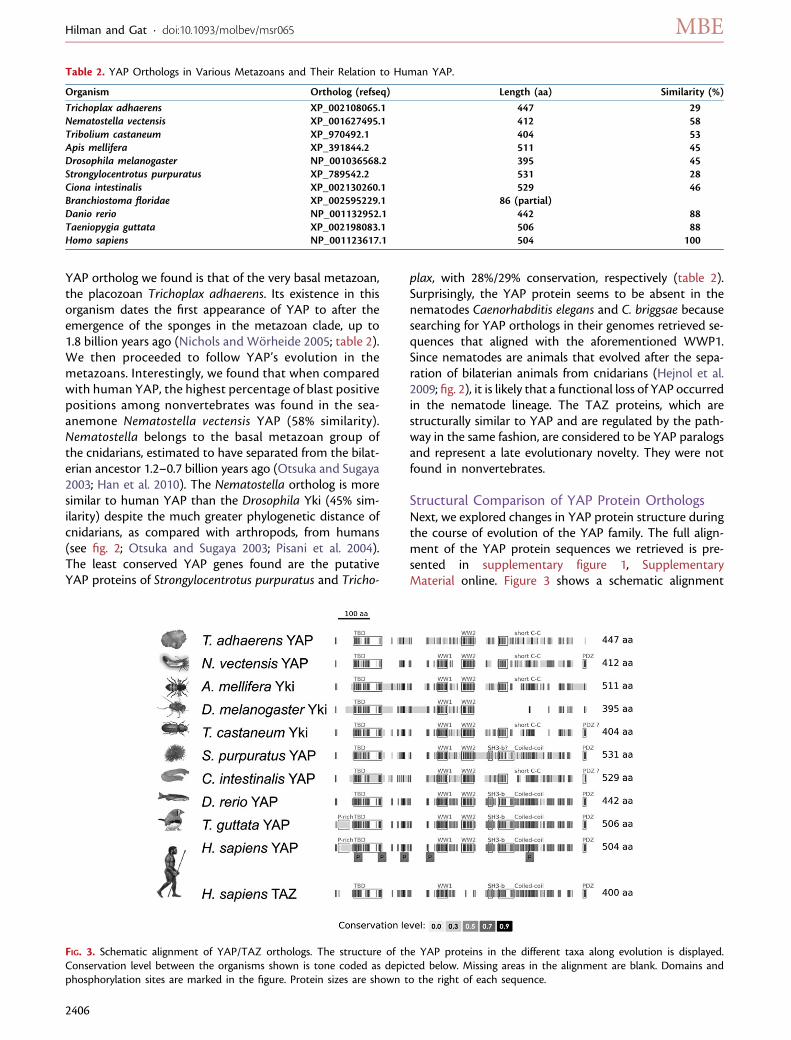
YAP ortholog we found is that of the very basal metazoan,the placozoan Trichoplax adhaerens. Its existence in thisorganism dates the first appearance of YAP to after theemergence of the sponges in the metazoan clade, up to1.8 billion years ago (Nichols and Worheide 2005; table 2).We then proceeded to follow YAP’s evolution in themetazoans. Interestingly, we found that when comparedwith human YAP, the highest percentage of blast positivepositions among nonvertebrates was found in the sea-anemone Nematostella vectensis YAP (58% similarity).Nematostella belongs to the basal metazoan group ofthe cnidarians, estimated to have separated from the bilat-erian ancestor 1.2–0.7 billion years ago (Otsuka and Sugaya2003; Han et al. 2010). The Nematostella ortholog is moresimilar to human YAP than the Drosophila Yki (45% sim-ilarity) despite the much greater phylogenetic distance ofcnidarians, as compared with arthropods, from humans(see fig. 2; Otsuka and Sugaya 2003; Pisani et al. 2004).The least conserved YAP genes found are the putativeYAP proteins of Strongylocentrotus purpuratus and Tricho-
plax, with 28%/29% conservation, respectively (table 2).Surprisingly, the YAP protein seems to be absent in thenematodes Caenorhabditis elegans and C. briggsae becausesearching for YAP orthologs in their genomes retrieved se-quences that aligned with the aforementioned WWP1.Since nematodes are animals that evolved after the sepa-ration of bilaterian animals from cnidarians (Hejnol et al.2009; fig. 2), it is likely that a functional loss of YAP occurredin the nematode lineage. The TAZ proteins, which arestructurally similar to YAP and are regulated by the path-way in the same fashion, are considered to be YAP paralogsand represent a late evolutionary novelty. They were notfound in nonvertebrates.
Structural Comparison of YAP Protein OrthologsNext, we explored changes in YAP protein structure duringthe course of evolution of the YAP family. The full align-ment of the YAP protein sequences we retrieved is pre-sented in supplementary figure 1, SupplementaryMaterial online. Figure 3 shows a schematic alignment
Table 2. YAP Orthologs in Various Metazoans and Their Relation to Human YAP.
Organism Ortholog (refseq) Length (aa) Similarity (%)
Trichoplax adhaerens XP_002108065.1 447 29Nematostella vectensis XP_001627495.1 412 58Tribolium castaneum XP_970492.1 404 53Apis mellifera XP_391844.2 511 45Drosophila melanogaster NP_001036568.2 395 45Strongylocentrotus purpuratus XP_789542.2 531 28Ciona intestinalis XP_002130260.1 529 46Branchiostoma floridae XP_002595229.1 86 (partial)Danio rerio NP_001132952.1 442 88Taeniopygia guttata XP_002198083.1 506 88Homo sapiens NP_001123617.1 504 100
FIG. 3. Schematic alignment of YAP/TAZ orthologs. The structure of the YAP proteins in the different taxa along evolution is displayed.Conservation level between the organisms shown is tone coded as depicted below. Missing areas in the alignment are blank. Domains andphosphorylation sites are marked in the figure. Protein sizes are shown to the right of each sequence.
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2406

of representative YAP orthologs in a wide array of meta-zoans. The most noticeable YAP conserved domains arethe two WW domains and the TEAD interaction domain.These are conserved from placozoans to the late bilaterianslike human. The Trichoplax YAP seems to have only oneWW domain, which aligned best with the second WW do-main of vertebrates and insect YAPs. The WW2 domain ismissing in TAZ proteins which contain only the first WWdomain (WW1).
Another conserved region is the C-terminal transactiva-tion domain. It seems to consist of several separated sub-domains and its composition varies between differentclades (fig. 3). As previously shown (Oka and Sudol2009), a conspicuous conserved PDZ-binding motif canbe found in the most distal sequence of the C-terminiof cnidarians, sea-urchins, and chordates. However, in in-sects, this sequence is divergent (fig. 3). The coiled-coil(c-c) domain that is important for TAZ and YAP dimeriza-tion (Murakami et al. 2005) is also conserved in vertebratesand cnidarians. The c-c domain in the insects Triboliumcastaneum and Apis mellifera is present partially and itis completely absent in Drosophila Yki. This domain ishighly divergent in Ciona intestinalis (fig. 3, supplementaryfig. 1, Supplementary Material online) suggesting an al-tered function with respect to its dimerization and inter-action activities. To conclude the analysis of the domainstructure, we observed that the proline rich region inthe N-terminal area of mammalian and avian YAP is miss-ing in other taxa (fig. 3).This indicates a yet unidentifiedfunction that arose late in evolution.
YAP activity was shown to be regulated by phosphory-lation and indeed several phosphorylation sites found inhumans are highly conserved in all metazoans tested, es-pecially the five sites (S61, S109, S127, S164, and S397; Zhaoet al. 2007; Hao et al. 2008; fig. 3). These are known to bephosphorylated by the Lats kinases in mammals and threeare phosphorylated by the Warts kinase in Drosophila. Thephosphoserine sites that are located prior to the WW do-mains are known to be important for Hpo/Wts-mediatedinhibition in Yki and YAP and this is likely reflected in thehigh degree of conservation of the regions in which theyreside (fig. 3 and supplementary fig. 1, Supplementary Ma-terial online). Mammalian YAP tyrosine phosphorylationwas shown to promote apoptosis in response to DNA dam-age (Levy et al. 2008). However, the tyrosine that is in-volved, Y391, seems to be conserved only in vertebrates.This tyrosine is aligned with phenylalanine in other deuter-ostomes (C. intestinalis, S. purpuratus) and is absent in pro-tostomes (see supplementary fig. 1, SupplementaryMaterial online).
The Genomic Composition of YAP Genes inMetazoansA genomic analysis of the YAP gene family demonstratedthe overall increased complexity of YAP gene structureover evolution (fig. 4). The basal YAP genes have 2 or 3exons, whereas the later forms have 8–9 exons. The secondWW domain appears to reside in a distinct exon in all bi-laterians, which can lead to differential splicing isoformsthat contain only the WW1 domain. In both human
FIG. 4. Genomic structure of YAP in representative metazoans. Major domains are marked with rectangles. The transcript coding region isdrawn to scale and introns positions are illustrated but their size is not drawn to scale.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2407

YAP and Drosophila Yki, there are isoforms documentedthat lack the second WW domain (fig. 4). These isoformsmay mimic the ancient condition of only one WW do-main in the Placozoans. The vertebrate TAZ (WWTR1)contains one WW domain, which is better aligned withthe first WW domain of YAP (figs. 3 and 4, supplementaryfig. 1, Supplementary Material online).
All the YAP (and TAZ) genes that we analyzed, exceptfor the T. adhaerens gene which does not have WW1, con-tain a remarkably conserved intron within theWW1 codingregion (figs. 4 and 5). The intron boundaries fall exactly inthe same codon, which codes for a conserved asparagine(N) residue and always start in the same phase (þ2) of thiscodon. The most diverse form of this intron boundarycan be observed in the sequence of S. purpuratus. Here,the intron is contained in an aspartic acid codon anddiffers from the chordate and the arthropod clades sequen-ces in several other positions around the intron boundaries(fig. 5A) The N. vectensis sequence, on the other hand,shares some features with chordate sequences and somewith arthropods around the intron boundaries. This mayreflect an ancient state before the separation of the majorbilaterian groups. The extreme conservation of thisintron strongly suggests that the same YAP structure ap-peared in a eumetazoan ancestor of both cnidarians and
all bilaterians. Why is this intron so well conserved oversuch a great span of evolutionary time? The reason isnot known, but we speculate that the intron harbors animportant binding site for some as yet undetermined crit-ical factor which conferred strong conservation to its prox-imity. Another hypothesis may be that without thepresence of this intron, this WW domain may be proneto duplication by a reverse transcription–based mechanism.The result may be evolutionary unfavorable.
Phylogeny of YAP and TAZ ProteinsIn order to examine the phylogenetic relations of the YAPfamily orthologs, we have utilized several tree-buildingtechniques. Both NJ and ML trees, which are consideredreliable for the analysis of distant sequences, were ableto distinguish between three major groups, arrangingthe chordates, arthropods, and early nonbilaterians in sep-arate clades (fig. 6A and B). In the ML tree, the nonbilater-ians T. adhaerens and N. vectensis form a clade of their ownand do not form an outgroup to the bilaterians. The NJanalysis, however, positions the T. adhaerens YAP as themost basal, being an outgroup to N. vectensis and all bilat-erians. N. vectensis YAP, on the other hand, clustered as anoutgroup to the bilaterians as could be expected from itsstructure. Surprisingly, the deuterostomate S. purpuratustended to group closer to the protostome clade in bothtrees but with low bootstrap support. Because some partsof the S. purpuratus YAP (Spr-YAP) display more similarityto chordates and other parts to arthropods, the Spr-YAPsequence likely represents an early bilaterian form, differentfrom both the chordates and the arthropods, as it hasemerged before the separation of these major bilaterianclades. In both NJ and ML trees, the C. intestinalis YAP fallsclose to the root of the chordate clade but with a longbranch. This suggests strong divergence of its sequence af-ter the urochordate and chordate lineage separation. Asimilar phenomenon can be observed in insects, where theDrosophila Yki sequence seems to be more diverse (fig. 3,supplementary fig. 1, Supplementary Material online) andprobably represents a more varied and novel sequence whencompared with themore basal-like insect groups which wereincluded in the analysis.
We went on and analyzed the later evolutionary radia-tions of the YAP family of proteins. The origin of the YAPparalogous gene TAZ was explored by aligning the multipleTAZ and YAP proteins that we retrieved from the data-bases and generating their NJ cladogram (fig. 6C). The re-sulting plot clusters the vertebrates’ YAP and TAZ proteinsin different clades. Both, however, have similar distancefrom out groups of vertebrates. This suggests that theTAZ gene probably originated from gene duplication ofYAP in early vertebrates. The conserved nature of theWW1 domain in TAZ and the identical localization ofthe intron in it also alludes to a common origin. We didnot find TAZ in chordates such as the cephalochordateB. lanceolatum or in the urochordate C. intestinalis ge-nomes. This dates the YAP/TAZ duplication to a late stagein chordates evolution, possibly associating it with the
FIG. 5. A conserved intron in the WW1 domain. An intron is foundin most YAP and TAZ orthologs within the WW1 domain. Theintron position and phase inside the codon for ‘‘N’’ is highlyconserved. The illustration shows the genomic sequence flankingthe intron boundaries in different organisms as well as the aminoacids coded by this region.
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2408

whole-genome duplication in the ancestral vertebrate lin-eage (Dehal and Boore 2005). A perplexing question is thereason for the emergence of the TAZ paralogs, especiallybecause a single WW1 harboring splicing isoform seeminglyof the same structure (fig. 4), exists in both vertebrates andDrosophila. Future experiments exploring the exact func-tion of TAZ, as opposed to YAP with its splicing forms,may shed light on the rationale for this late evolutionarydevelopment and on its benefit for vertebrates.
YAP and the Hippo/YAP Pathway—DevelopingComplexityWhat is the evolutionary status of YAP in the context of theentire pathway? How did the pathway as we know itevolve? Did the components appear as a full set along witha novel animal group or did they emerge gradually inseveral steps to establish and elaborate the pathway? Inorder to address these questions and to learn about theevolutionary history of the pathway, we searched for theorthologs of the core pathway components as well as
the upstream members of the pathway in representativesingle-celled eukaryotes and in metazoan groups (table 1).
Single-Celled EukaryotesFigure 7 depicts the composition of the pathway alongevolution from eukaryotes to mammals. We can find bonafide orthologs of TEAD and Mob in the yeast S. cerevisiae,which may represent the most ancient orthologs of thepathway. We did not find them in prokaryotes. Moreover,YAP orthologs were not found in plants, reducing the pos-sibility of pathway loss during the early evolution of eu-karyotes in general. Previously, it has been suggested thatthe roots of the pathway may be represented by the mi-totic exit network (MEN) and the separation initiationnetwork pathways in the budding and fission yeast cor-respondingly (Bardin and Amon 2001; Harvey and Tapon2007). Our results cannot, however, categorically identifythe yeast Cdc15 and Dbf2 as orthologs of Hippo andWarts, respectively, as previously suggested because theytend to align better with other paralogs of these kinases.
FIG. 6. Phylogenetics of YAP. ML (A) and NJ (B) unrooted trees of YAP orthologs from distant metazoans. All sequences were downloaded fromrefseq and aligned with MUSCLE. Trees were constructed as described in Methods. The bootstrap consensus tree inferred from 1,000 replicates.The percentage of replicate trees in which the associated ortholog clustered together in the bootstrap test is shown next to the branches. Thetrees are drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. In theNJ tree, the distances were computed using the Poisson correction method and are in the units of the number of amino acid substitutions persite. The rate variation among sites was modeled with a gamma distribution. All positions containing alignment gaps and missing data wereeliminated only in pairwise sequence comparisons. There were a total of 796 positions in the final data set. (C) The TAZ proteins are paralogs ofvertebrate YAPs. The cladogram demonstrates the relations between the YAP and TAZ paralogs, showing that TAZ is clustered closer to thevertebrates YAP than to the invertebrates’ ortholog. The high similarity of vertebrate and Nematostella YAP (table 2 and fig. 3) relative toDrosophila Yki, puts Nematostella, closer to the vertebrates, when compared mainly with vertebrates proteins. The lack of other insects in thisanalysis artificially clusters Drosophila outside the bilaterians and not as presented in (A) and (B). The cladogram was inferred using the NJmethod. YAP and TAZ sequences were downloaded from the orthoMCL database. The tree was built with MEGA4 using default parameters.The optimal tree is shown and the bootstrap test (1,000 replicates) is shown next to the branches. The tree is drawn to scale, with branchlengths in the same units as those of the evolutionary distances used to infer the phylogeny.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2409

That said, the similarity of the genetic module and thegenes involved, along with the similar interaction withMob like protein in yeast, suggest a deep evolutionaryconnection that warrants further examination. Warts/Lats has been reported to be localized to the centrosomein interphase, translocate to the mitotic spindle in meta-phase and anaphase, and then to localize to the midbodyin telophase in human cells (Nishiyama et al. 1999;Morisaki et al. 2002). Additionally, embryonic cells fromLats2 knockout mice display multiple mitosis defects in-cluding abnormal exit from mitosis (Yabuta et al. 2007).As this expression pattern and function is reminiscent ofthat of Dbf2 in yeast (Bardin and Amon 2001), it may rep-resent an ancient role for the pathway in mitotic exit thathas remained in metazoans up to humans (Praskova et al.2008)
Tec1, the putative yeast TEAD ortholog, regulates signal-ing of one of the mitogen-activated protein (MAP) kinasepathways in the budding yeast, a pathway that is involvedin invasive growth (Wang et al. 2009). We could not finda direct connection between the function of the aboveMEN factors that are related to the Hippo/YAP pathwayand that of TEAD/Tec1. However, the founding memberof the Mst/Hippo family of kinases, Ste-20 in yeast (Raduand Chernoff 2009), is part of the MAP kinase network thatmodulates Tec1 activity. It is thus possible that Ste-20 is theancestral Mst and that the origin of the pathway is relatedto the Tec1-regulated invasive response. In conclusion,there are several candidates in yeast for elements of thecurrent Hippo/Yap pathway. These components may havejoined to evolve the metazoan form of the pathway. In ad-dition, the pathway may have consolidated along
FIG. 7. Summary of the Hippo/YAP pathway evolution. The figure shows the conservation of the core components of the Hippo/YAP signalingpathway in representative metazoans. Orthologs between different organisms are placed in the same relative position. The colors code for theproposed antiquity of the genes in evolution. When orthologs were not found, the ortholog place remained blank with a red circle. In case theortholog identity is not sure, a question mark (?) was added, and the putative paralog that matches this sequence was written above if found.Divergence times of major metazoa groups are presented next to the relevant branches. Our analysis shows that the nearly complete corepathway known from bilaterians is represented in the cnidarian Nematostella vectensis. The pathway is mostly absent in sponges andnonmetazoans and is partially present in placozoans. The nematode Caenorhabditis elegans and its relatives Caenorhabditis brigssae aredeficient in several of the pathway components (represented here by the C. elegans conserved components). This may indicate of a functionalloss of major components in nematodes. The YAP paralogs TAZ were found only in vertebrates and most likely evolved as a result ofduplication in early vertebrates or prevertebrate chordates.
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2410

metazoan evolution due to newly acquired roles of theseprogenitors as well as to the emergence and integration ofnew pathway members.
Analysis of the genome of the unicellular eukaryotechoanoflagellateM. brevicollis, considered to be the closestrelative of metazoans, did not yield any more orthologs ofthe core pathway. However, upstream elements such as theprotocadherins fat/ds may have first emerged in choano-flagellates (King et al. 2003, 2008). This would have allowedcolony formation and further on in evolution they couldhave been utilized for the more intricate and complex tasksof tissue growth control, which necessitated formation ofthe Hippo/YAP pathway by recruitment of other preexist-ing factors and incorporation of novel elements.
Sponges and PlacozoansMoving along into the metazoan clade, we can identifya true ortholog of Mst/Hippo kinase in sponges and allgroups beyond, but still no Sav, Lats or YAP (fig. 7). A hall-mark in the evolution of this pathway is the appearance ofthe dynamic effector YAP. The most basal YAP ortholog wehave detected is that of the placozoan T. adhaerens, whichis a eumetazoan with simple but organized tissues(Srivastava et al. 2008). This YAP ortholog includes theTEAD-binding domain and four out of five of the Lats/Wts phosphorylation sites, but only one WW domain,which is similar to WW2 (see fig. 3 and supplementaryfig. 1, Supplementary Material online). Thus, this YAP is dif-ferent in structure from all the following orthologs. Takinginto account the lack of other components of the pathwayin this organism, such as the Lats/Wts kinase, this suggeststhat the pathway in the placozoans is still in a primordialform and likely functions differently from that in bilater-ians. It will be interesting to find out the mode of actionof this rudimentary and minimal pathway in this littlestudied organism.
The Sea-Anemone NematostellaThe cnidarian N. vectensis is considered to be the closestoutgroup to the bilaterians among the known extant taxa(Finnerty et al. 2004; Dunn et al. 2008; Hejnol et al. 2009;Sperling et al. 2009; Ryan et al. 2010). Our analysis clearlyshows that this group of basal nonbilaterians containsmost of the components of the ‘‘modern’’ Hippo/YAPpathway (fig. 7). This includes the complete core Hippomachinery and all the supposed upstream signaling pro-teins except for the ortholog of Expanded which we couldnot find. Notably, the Nematostella YAP demonstrates al-most all the features recognized in many later bilateriangroups. In addition to the conserved elements that we de-scribed above for T. adhaerens, these include two well-conserved WW domains and a PDZ-binding C-terminalpentapeptide identical to the human form (see fig. 3and supplementary fig. 1, Supplementary Material online).Intriguingly, when compared with different bilateriangroups, the Nematostella YAP shows the highest similarityto the vertebrate YAP (see table 2 and supplementary fig.1, Supplementary Material online). This high level of re-semblance between Nematostella and vertebrates has
been demonstrated before for many developmental genes(Ryan et al. 2007; Hejnol et al. 2009; Saina et al. 2009). Ithas been explained by the basal nature of both the sea-anemone genome and the vertebrate genome, which ismuch less derived than many of the protostome groupslike insects and nematodes. The question then ariseswhether this similarity in protein structure is also re-flected in a similar modi operandi among these very dis-tant animal groups.
The Bilaterian Groups—Conservation and DivergenceWhen we considered the status of the pathway within thebilaterian groups for which genomic data are available, wegenerally found all the components that were establishedearlier. However, as in other major developmental path-ways, the complexity increased and we were able to detectthe appearances of additional factors and more paralogsadded to the preexisting pathway components. Thus,theMst, Lats, TEAD, and YAP genes underwent duplicationevents during deuterostome evolution (fig. 7). This mayhave conferred more flexibility to the pathway as the var-ious paralogs could allow modulation of function in differ-ent organs. Our phylogenetic analysis points out that theancient YAP sequence diverged to two main groups of pro-teins: the arthropod Yki and the deuterostome YAP, whichincludes the chordates. The sequence alignments suggestthat the arthropod Yki forms are much more divergentand harbor many more changes when compared withthe chordate YAP species that are more ‘‘basal’’ and resem-ble the sea-anemone ancestral YAP. Several interaction el-ements that are important in vertebrate YAP/TAZ, such asthe SH3-binding domain (Espanel and Sudol 2001) and theY391 like tyrosine (Levy et al. 2008), are absent in insectsand early metazoans. This may point to the generation ofnovel functions of YAP during deuterostomes and verte-brate evolution. The new phosphorylation site thatappeared on tyrosine can be associated with more elabo-rate apoptosis promotion abilities for which vertebratesYAPs were enlisted (Levy et al. 2008). Interestingly, al-though the site is a vertebrates novelty, the kinase respon-sible for the phosphorylation, c-Abl, is an ancient metazoangene that is already found in the sponge.
Inspection of the Drosophila Yki shows that indeed, itsstructure is different in that the C-terminus is much shorterand is missing many sequences attributed to transcriptionactivation. It also entirely lacks the c-c domain found indeuterostomes and in a partial form also in other insects.Moreover, the PDZ-binding domain, which was recentlyshown to be critical for the function of YAP in vertebrates(Oka and Sudol 2009), is highly modified in Yki. The diver-gent nature of Yki likely indicates that a significant changein the gene function has transpired during arthropods evo-lution. This change may have occurred in order to accom-modate a unique form of embryonic development such asthe whole-body segmentation mode typical to Drosophila.Unlike many other insect and arthropod groups flies lacka growth zone that gradually gives rise to posterior segmentsduring its embryogenesis.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2411

The apparent lack of obvious YAP orthologs in the nem-atodes C. elegans and C. briggasae and the lack of otherpathway components (Ex and Kibra, fig. 7) is another dra-matic example of a major change in the pathway in a pro-tostome group. A similar phenomenon of the absence ofcrucial and conserved developmental genes in nematodeswas also previously described (Aboobaker and Blaxter2003a, 2003b), wherein the authors portrayed the disorga-nization and decay of the Hox gene cluster in C. elegans.The Hpo/Mst orthologs Cst-1/2 in C. elegans are Ste-20 likekinases, which were shown to increase nematode lifespanby phosphorylating Daf-16, the mammalian FoxO orthologthat is a known substrate of Mst kinases (Lehtinen et al.2006) The C. elegans Lats/Warts, Ce-Wts-1, is importantto various developmental functions in C. elegans includingbody length control by the TGF-beta Sma/Mab pathway(Cai et al. 2009). It is also associated with lifespan control(Curran and Ruvkun 2007), whereas the putative TEAD or-tholog, egl-44, is involved in mechanosensory neuron cellfate determination (Wu et al. 2001). Further experimentaldata are required in order to establish the function of theseand the remaining Hippo/YAP pathway elements in nem-atodes and to reveal the changes that the pathway hasundergone in this derived group.
It will also be interesting to explore the status of thepathway in other protostome groups such as mollusksas more sequence data become available.
Coevolution of Genes in the Hippo/YAP PathwayWe were interested in testing whether there is any evidencefor coevolution between components of the pathway,which may indicate the evolutionary origin of a functionalassociation between them. In recent studies, an alignmentof the YAP-binding domain of TEAD1 and the TEAD-binding domain of YAP, along with the solved structureof YAP and TEAD1 (Chen et al. 2010; Li et al. 2010), showedthe putative coevolution of these areas in YAP and TEAD1.Specifically, residues in the TEAD-binding domain of YAP,and in the TEAD partner protein, which physically interactin the 3D model, are also highly conserved within metazo-ans (Li et al. 2010). To test whether YAP and TEAD havecoevolved, we used phylogenetic evidence to compare thecalculated pairwise lengths between all YAP orthologs andTEAD orthologs across distant taxa. This method was usedin earlier work to demonstrate strong protein–protein in-teractions between two proteins (Pazos et al. 2005). Theprinciple of this analysis is that in order to assess evolution-ary connection, one examines how the evolution of onesequence, which is measured by the pairwise distancesof any two orthologs, is correlated to the distances ofthe orthologs of another gene. We expect that if two genesinteracted and thus coevolved, we will observe a correlationbetween the pairwise distances (supplementary data 3,Supplementary Material online).
Our coevolution analysis indeed suggested a very strongcorrelation between YAP and TEAD pairwise distances,with a correlation coefficient r 5 0.85, P 5 1.1 � 10�16 (fig. 8A). This strongly supports a functional association
between these two proteins. Such coevolution was alsoshown in the transcription factor and coactivator pairRunx-CBFb (Sullivan et al. 2008). This coevolution suggestsa strong selection pressure toward the interaction of YAPand TEAD and strengthens the hypothesis that TEAD isa key downstream effector of the pathway (Ota and Sasaki2008; Li et al. 2010). YAP can interact with other transcrip-tional partners and thus we analyzed its coevolution withthe aforementioned Runx and p73 cofactors. AlthoughYAP seems to be highly correlated to p73 (r 5 0.85,P 5 6.2 � 10�7, supplementary data 3, SupplementaryMaterial online), which suggests that they strongly inter-acted during evolution, there was no indication of coevo-lution with the Runx proteins. This in turn could indicatea recent interaction or that the interaction contact regionsare very short.
Extending the analysis we showed that YAP/Yki exhibita high degree of coevolution with Mob1/Mats (fig. 8B) andSav1 (fig. 8C) and a lower but positive correlation withLats1/Wts (fig. 8D) and Mst2/Hpo (fig. 8E). All these func-tion in a close signaling network (Pan 2010). The lower cor-relations seen for the later (fig. 8D and E) could be due tothe multiple other partners of these kinases and of YAP.Some of the other components in the pathway that areknown or suspected to interact with one another also dem-onstrate a positive correlation such as Mob1-Mst2 (Weiet al. 2007; fig. 8F), Mob1-Lats1 (Lai et al. 2005; Weiet al. 2007; fig. 8G), whereas Mer-Ex display a high correla-tion (McCartney et al. 2000; Hamaratoglu et al. 2006; Yuet al. 2010; fig. 8H). As could be expected not all previouslyknown binding partners showed coevolution in this assay,whereas other pairs of proteins, such as YAP/TAZ and Mer-lin, exhibited unexpected high correlation, which could in-dicate that they may physically interact (see supplementarydata 3, Supplementary Material online). Other pairs of pro-teins that are not known to bind or interact show a series ofdistances with much lower correlation. For example, thepredicted correlation between YAP and FAT4 yielded anr 5 0.27 (fig. 8I) and with DCHS1, r 5 �0.02 (8J). The cor-relation between FAT4 and DCHS1, that do interact in Dro-sophila (Cho et al. 2006), is much higher with r 5 0.65(fig. 8K). b-catenin, another transcriptional coactivator thatacts with the same operational logic as YAP, but in a differentpathway, yields an r 5 0.10 for YAP (fig. 8L).
To further test this technique on another signaling path-way, we analyzed the main components of the hedgehogpathway, which is also composed of an assembly of ancienteukaryotic and novel metazoan genes (Matus et al. 2008).We found an r5 0.68 between Hedgehog and its receptor,the tumor suppressor Patched1, (PTCH1), an r 5 0.32 ofPTCH1 to the smoothened receptor and an r 5 0.52 ofPTCH1 to the GLI2 transcription factor of this pathway(supplementary data 3, Supplementary Material online).These correlations point to a general trend of coevolutionof interacting components in developmental pathways.
Overall, our data demonstrate the deep evolutionaryconnections between the components of the Hippo/YAP pathway, which are manifested in their coevolution
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2412
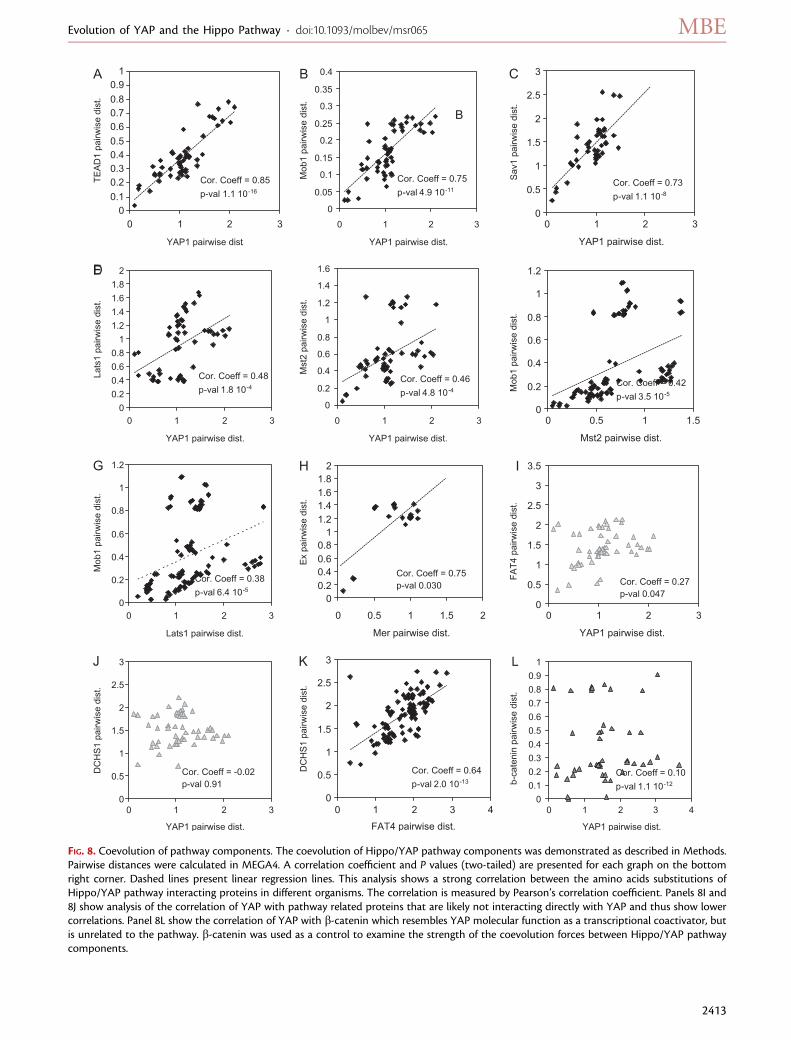
FIG. 8. Coevolution of pathway components. The coevolution of Hippo/YAP pathway components was demonstrated as described in Methods.Pairwise distances were calculated in MEGA4. A correlation coefficient and P values (two-tailed) are presented for each graph on the bottomright corner. Dashed lines present linear regression lines. This analysis shows a strong correlation between the amino acids substitutions ofHippo/YAP pathway interacting proteins in different organisms. The correlation is measured by Pearson’s correlation coefficient. Panels 8I and8J show analysis of the correlation of YAP with pathway related proteins that are likely not interacting directly with YAP and thus show lowercorrelations. Panel 8L show the correlation of YAP with b-catenin which resembles YAP molecular function as a transcriptional coactivator, butis unrelated to the pathway. b-catenin was used as a control to examine the strength of the coevolution forces between Hippo/YAP pathwaycomponents.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2413

and are presumably necessary to maintain the functionalityof the network. In addition, we show the potential of thisapproach for defining possible protein–protein interactionsthat were previously not explored.
Hypotheses Regarding the Origin and EvolutionaryRole of YAP and the Hippo/YAP PathwayFrom our evolutionary survey, we hypothesize that thepathway as we know it in human and in Drosophila, withthe majority of its associated machinery, can be dated tothe last common ancestor of cnidarians and bilaterians, es-timated to exist more than 700 Ma (Otsuka and Sugaya2003; Hejnol et al. 2009; Han et al. 2010). The finding ofother components of the pathway that have much broaderclusters of orthologs in distant phyla, well before the adventof metazoans estimated up to 1.8 billion years ago (Nicholsand Worheide 2005), indicates that the preexisting seg-ments of this pathway likely played roles both in mitoticregulation processes like mitotic exit and in environmentalsignaling. Moreover, they were redirected to add multicel-lular tissue growth control tasks during metazoans bodyplan evolution. This new recruitment was necessary to per-form novel regulatory functions. These, in turn, rewired ormodified some existing components, eventually addingnew components such as YAP. This paradigm of using pre-existing proteins and incorporating components witha new combination of domains, or recruiting altogethernovel domains on top of existing ones as well as the de-ployment of totally new proteins, is increasingly recognizedas the ‘‘evolutionary workshop.’’ It created the new path-ways (e.g., the Wnt pathway; Adamska et al. 2010) and themodified functional complexes (e.g., adhesion junctions;Nichols et al. 2006), necessary for emergence of new lifeforms such as the metazoans and bilaterians (Srivastavaet al. 2010).
Although TEAD and Mob are ancient eukaryotic pro-teins, YAP seems to be a dynamic metazoan novelty inwhich an existing WW domain protein was modifiedand combined with novel domains such as the TEAD-binding domain and the PDZ-interacting domain. Whatis interesting about the evolution of YAP is that it has likelyemerged to form this pathway in ancestral eumetozoanslike the placozoans/cnidarians/bilaterian progenitors,which may suggest that the pathway is necessary for thegrowth control of more complex organized tissue as com-pared with sponge cells. In addition, YAP structure hasbeen modified along the course of evolution with theputative addition of a second WW domain and thePDZ-binding domain in cnidarians. Later, more minor alter-ations of the scheme appeared, such as diversification ofthe PDZ-binding pentapeptide in insects and the formationof the TAZ paralogs in the vertebrates.
What new insights can be gleaned from our evolutionarystudy about the function of the Hippo/YAP pathway? Theapparent lack of a YAP ortholog and several other pathwaycomponents in the ecdysozoan C. elegans demonstratesthat the full pathway may not be essential for all types
of metazoan life forms. The variant insect forms suggestthat the pathway had to be modified and diversified forsome others. The absence of YAP may indicate a dramaticchange in the developmental mode of nematodes towardan early ‘‘deterministic’’ pattern (Schierenberg 2001). In thismode, which is the classical alternative to the ‘‘regulative’’mode of development, a signaling pathway that controlscell growth in response to environmental cues may nolonger be needed. Thus, we speculate that this pathwayhas primarily evolved in order to regulate tissue and organgrowth during metazoan ‘‘regulative’’ phases of develop-ment, which are flexible and rely on cell–cell interactionand environmental signals. However, the full complementof the pathway may not be essential for the ‘‘determina-tive (mosaic)’’ like type of development which is preprog-rammed in the developing organism. Exploration of theHippo/YAP pathway’s function in developmental stagesthat exhibit these types of development, both of whichoften appear at distinct stages in the development ofmodel organisms may show if indeed our speculation isvalid.
The Hippo/YAP pathway’s evolutionary tale may thusprovide insight into the evolution of tissue and organ sizecontrol in multicellular animals. It features YAP as one ofthe most important tools in the metazoan developmentaltoolbox.
ConclusionsWe explored the origin and evolution of the Hippo/YAPpathway, which controls tissue proliferation rates and or-gan size in animals. Some of the pathway’s elements werefound already in protists like yeast and the choanoflage-lates, whereas the first YAP progenitor was detected inthe placozoan T. adhaerens. The full pathway as knownin man can be first identified in the basal metazoan cnidar-ian N. vectensis. Following the evolution of the pathwayalong the bilaterian groups shows an overall increase inits complexity, such as the emergence of additional paral-ogs of the core kinases and the advent of the TAZ genes,the vertebrate YAP paralogs. However, some bilaterians,such as Drosophila show divergence of the YAP effector.This may reflect a unique mode of their embryonic devel-opment pattern. At the same time, other groups, like thenematode C. elegans, show a loss of YAP and several othercomponents, which may hint at their simplified mode ofcellular growth control.
Thus, in reconstructing the origin of this pathway, wecan envision the following scenario: The evolution of orga-nized tissues and organs in metazoans required the inven-tion of a mechanism for tissue growth regulation. For thispurpose, a new genetic network that can connect intertis-sue signals with cellular proliferation control was necessary.This pathway has evolved in accordance with the develop-mental growth control programs of the various animalforms. Our findings indicate that the emergence of the ‘‘ad-vanced’’ Hippo/YAP pathway is associated with the appear-ance of an organized body plan that incorporates distinct
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2414

organs and tissues. Even though the pathway is highly con-served, and its genes have undergone strong coevolution, itis adapted to the developmental program of the organismand shows plasticity that is reflected in divergence of somecritical domains of its components.
Supplementary MaterialSupplementary datas 1–3 and figure 1 are available atMolecular Biology and Evolution online (http://www.mbe.oxfordjournals.org/).
AcknowledgmentsThe authors would like to thank Drs. Ariel Chipman, LiranCarmel, Rachel Green, and Marshall Devor of the HebrewUniversity in Jerusalem for their critical reading of this man-uscript and helpful suggestions. This study was supportedby the Israel Science Foundation (grant 825/07).
ReferencesAboobaker A, Blaxter M. 2003a. Hox gene evolution in nematodes:
novelty conserved. Curr Opin Genet Dev. 13:593–598.Aboobaker AA, Blaxter ML. 2003b. Hox gene loss during dynamic
evolution of the nematode cluster. Curr Biol. 13:37–40.Adamska M, Larroux C, Adamski M, Green K, Lovas E, Koop D,
Richards GS, Zwafink C, Degnan BM. 2010. Structure
and expression of conserved Wnt pathway components in
the demosponge Amphimedon queenslandica. Evol Dev.
12:494–518.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic
local alignment search tool. J Mol Biol. 215:403–410.Bardin AJ, Amon A. 2001. MEN and SIN: what’s the difference? Nat
Rev Mol Cell Biol. 2:815–826.Cai Q, Wang W, Gao Y, Yang Y, Zhu Z, Fan Q. 2009. Ce-wts-1 plays
important roles in Caenorhabditis elegans development. FEBS
Lett. 583:3158–3164.Chen F, Mackey AJ, Vermunt JK, Roos DS. 2007. Assessing
performance of orthology detection strategies applied to
eukaryotic genomes. PloS One. 2:e383.Chen L, Chan SW, Zhang X, Walsh M, Lim CJ, Hong W, Song H.
2010. Structural basis of YAP recognition by TEAD4 in the hippo
pathway. Gene Dev. 24:290–300.Cho E, Feng Y, Rauskolb C, Maitra S, Fehon R, Irvine KD. 2006.
Delineation of a fat tumor suppressor pathway. Nat Genet.
38:1142–1150.Consortium TU. 2010. The universal protein resource (UniProt) in
2010. Nucleic Acids Res. 38:D142–D148.Curran SP, Ruvkun G. 2007. Lifespan regulation by evolutionarily
conserved genes essential for viability. PLoS Genet. 3:e56.De Robertis E. 2008. Evo-devo: variations on ancestral themes. Cell
132:185–195.Degnan BM, Vervoort M, Larroux C, Richards GS. 2009. Early
evolution of metazoan transcription factors. Curr Opin Genet
Dev. 19:591–599.Dehal P, Boore JL. 2005. Two rounds of whole genome duplication in
the ancestral vertebrate. PLoS Biol. 3:e314.Dong J, Feldmann G, Huang J, Wu S, Zhang N, Comerford SA,
Gayyed MF, Anders RA, Maitra A, Pan D. 2007. Elucidation of
a universal size-control mechanism in Drosophila and mammals.
Cell 130:1120–1133.
Dunn CW, Hejnol A, Matus DQ, et al. (18 co-authors). 2008. Broadphylogenomic sampling improves resolution of the animal treeof life. Nature 452:745–749.
Edgar RC. 2004. MUSCLE: multiple sequence alignment withhigh accuracy and high throughput. Nucleic Acids Res. 32:1792–1797.
Espanel X, Sudol M. 2001. Yes-associated protein and p53-bindingprotein-2 interact through their WW and SH3 domains. J BiolChem. 276:14514–14523.
Finnerty JR, Pang K, Burton P, Paulson D, Martindale MQ. 2004.Origins of bilateral symmetry: Hox and dpp expression in a seaanemone. Science. 304:1335–1337.
Grusche FA, Richardson HE, Harvey KF. 2010. Upstream regulationof the Hippo size control pathway. Curr Biol. 20:R574–R582.
Guindon S, Gascuel O. 2003. A simple, fast, and accurate algorithmto estimate large phylogenies by maximum likelihood. Syst Biol.52:696–704.
Guindon S, Lethiec F, Duroux P, Gascuel O. 2005. PHYML online—aweb server for fast maximum likelihood-based phylogeneticinference. Nucleic Acids Res. 33:W557–W559.
Hamaratoglu F, Willecke M, Kango-Singh M, Nolo R, Hyun E, Tao C,Jafar-Nejad H, Halder G. 2006. The tumour-suppressor genesNF2/Merlin and expanded act through Hippo signalling toregulate cell proliferation and apoptosis. Nat Cell Biol. 8:27–36.
Han J, Kubota S, Uchida HO, Stanley GD Jr., Yao X, Shu D, Li Y,Yasui K. 2010. Tiny sea anemone from the lower cambrian ofChina. PloS One. 5:e13276.
Hao Y, Chun A, Cheung K, Rashidi B, Yang X. 2008. Tumorsuppressor LATS1 is a negative regulator of oncogene YAP. J BiolChem. 283:5496–5509.
Harvey K, Tapon N. 2007. The salvador-warts-hippo pathway—anemerging tumour-suppressor network. Nat Rev Cancer.7:182–191.
Harvey KF, Pfleger CM, Hariharan IK. 2003. The Drosophila mstortholog, Hippo, restricts growth and cell proliferation andpromotes apoptosis. Cell. 114:457–467.
Hejnol A, Obst M, Stamatakis A, et al. (17 co-authors). 2009.Assessing the root of bilaterian animals with scalable phyloge-nomic methods. Proc R Soc B. 276:4261–4270.
Hemmrich G, Bosch TCG. 2008. Compagen, a comparativegenomics platform for early branching metazoan animals,reveals early origins of genes regulating stem-cell differentiation.BioEssays 30:1010–1018.
Hisaoka M, Tanaka A, Hashimoto H. 2002. Molecular alterations ofh-warts/LATS1 tumor suppressor in human soft tissue sarcoma.Lab Invest. 82:1427–1435.
Huang J, Wu S, Barrera J, Matthews K, Pan D. 2005. The Hipposignaling pathway coordinately regulates cell proliferation andapoptosis by inactivating Yorkie, the Drosophila homolog ofYAP. Cell. 122:421–434.
Justice RW, Zilian O, Woods DF, Noll M, Bryant PJ. 1995. TheDrosophila tumor suppressor gene warts encodes a homolog ofhuman myotonic dystrophy kinase and is required for thecontrol of cell shape and proliferation. Gene Dev. 9:534–546.
Kango-Singh M, Singh A. 2009. Regulation of organ size: insightsfrom the Drosophila hippo signaling pathway. Dev Dyn.238:1627–1637.
King N, Hittinger CT, Carroll SB. 2003. Evolution of key cell signalingand adhesion protein families predates animal origins. Science301:361–363.
King N, Westbrook MJ, Young SL, et al. (36 co-authors). 2008. Thegenome of the choanoflagellate Monosiga brevicollis and theorigin of metazoans. Nature 451:783–788.
Kumar S, Nei M, Dudley J, Tamura K. 2008. MEGA: a biologist-centric software for evolutionary analysis of DNA and proteinsequences. Brief Bioinform. 9:299–306.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2415

Lai ZC, Wei X, Shimizu T, Ramos E, Rohrbaugh M, Nikolaidis N,Ho LL, Li Y. 2005. Control of cell proliferation and apoptosis bymob as tumor suppressor, mats. Cell. 120:675–685.
Lehtinen MK, Yuan Z, Boag PR, et al. (11 co-authors). 2006. Aconserved MST-FOXO signaling pathway mediates oxidative-stress responses and extends life span. Cell 125:987–1001.
Levy D, Adamovich Y, Reuven N, Shaul Y. 2007. The Yes-associatedprotein 1 stabilizes p73 by preventing Itch-mediated ubiquitinationof p73. Cell Death Differ. 14:743–751.
Levy D, Adamovich Y, Reuven N, Shaul Y. 2008. Yap1 phosphor-ylation by c-Abl is a critical step in selective activation ofproapoptotic genes in response to DNA damage. Mol Cell.29:350–361.
Li L, Stoeckert CJ, Roos DS. 2003. OrthoMCL: identification ofortholog groups for eukaryotic genomes. Genome Res.13:2178–2189.
Li Z, Zhao B, Wang P, Chen F, Dong Z, Yang H, Guan KL, Xu Y. 2010.Structural insights into the YAP and TEAD complex. Genes Dev.24:235–240.
Maglott D, Ostell J, Pruitt KD, Tatusova T. 2007. Entrez gene: gene-centered information at NCBI. Nucleic Acids Res. 35:D26–D31.
Matus DQ, Magie CR, Pang K, Martindale MQ, Thomsen GH. 2008.The Hedgehog gene family of the cnidarian, Nematostellavectensis, and implications for understanding metazoan hedge-hog pathway evolution. Dev Biol. 313:501–518.
McCartney BM, Kulikauskas RM, LaJeunesse DR, Fehon RG. 2000.The neurofibromatosis-2 homologue, Merlin, and the tumorsuppressor expanded function together in Drosophila toregulate cell proliferation and differentiation. Development.127:1315–1324.
Morisaki T, Hirota T, Iida S, et al. (13 co-authors). 2002. WARTStumor suppressor is phosphorylated by Cdc2/cyclin B at spindlepoles during mitosis. FEBS Lett. 529:319–324.
Murakami M, Nakagawa M, Olson EN, Nakagawa O. 2005. A WWdomain protein TAZ is a critical coactivator for TBX5,a transcription factor implicated in Holt-Oram syndrome. ProcNatl Acad Sci U S A. 102:18034–18039.
Nichols S, Worheide G. 2005. Sponges: new views of old animals.Integr Comp Biol. 45:333–334.
Nichols SA, Dirks W, Pearse JS, King N. 2006. Early evolution ofanimal cell signaling and adhesion genes. Proc Natl Acad Sci U S A.103:12451–12456.
Nishiyama Y, Hirota T, Morisaki T, et al. (11 co-authors). 1999. Ahuman homolog of Drosophila warts tumor suppressor, h-warts,localized to mitotic apparatus and specifically phosphorylatedduring mitosis. FEBS Lett. 459:159–165.
Oh H, Irvine KD. 2010. Yorkie: the final destination of Hipposignaling. Trends Cell Biol. 20:410–417.
Oka T, Sudol M. 2009. Nuclear localization and pro-apoptoticsignaling of YAP2 require intact PDZ-binding motif. Genes Cells.14:607–615.
Ota M, Sasaki H. 2008. Mammalian tead proteins regulate cellproliferation and contact inhibition as transcriptional mediatorsof Hippo signaling. Development 135:4059–4069.
Otsuka J, Sugaya N. 2003. Advanced formulation of base pairchanges in the stem regions of ribosomal RNAs; its applicationto mitochondrial rRNAs for resolving the phylogeny of animals.J Theor Biol. 222:447–460.
Pan D. 2010. The Hippo signaling pathway in development andcancer. Dev Cell. 19:491–505.
Pantalacci S, Tapon N, Leopold P. 2003. The Salvador partner Hippopromotes apoptosis and cell-cycle exit in Drosophila. Nat CellBiol. 5:921–927.
Pazos F, Ranea JAG, Juan D, Sternberg MJE. 2005. Assessing proteinco-evolution in the context of the tree of life assists in theprediction of the interactome. J Mol Biol. 352:1002–1015.
Pisani D, Poling LL, Lyons-Weiler M, Hedges SB. 2004. Thecolonization of land by animals: molecular phylogeny anddivergence times among arthropods. BMC Biol. 2:1.
Praskova M, Xia F, Avruch J. 2008. MOBKL1A/MOBKL1B phosphor-ylation by MST1 and MST2 inhibits cell proliferation. Curr Biol.18:311–321.
Price MN, Dehal PS, Arkin AP. 2010. FastTree 2—approximatelymaximum-likelihood trees for large alignments. PloS One.5:e9490.
Pruitt KD, Tatusova T, Maglott DR. 2007. NCBI reference sequences(RefSeq): a curated non-redundant sequence database ofgenomes, transcripts and proteins.Nucleic Acids Res. 35:D61–D65.
Radu M, Chernoff J. 2009. The DeMSTification of mammalian Ste20kinases. Curr Biol. 19:R421–R425.
Ryan JF, Mazza ME, Pang K, Matus DQ, Baxevanis AD,Martindale MQ, Finnerty JR. 2007. Pre-bilaterian origins of thehox cluster and the hox code: evidence from the sea anemone,Nematostella vectensis. PloS One. 2:e153.
Ryan JF, Pang K. Mullikin JC, Martindale MQ, Baxevanis AD. NISCComparative Sequencing Program. 2010. The homeodomaincomplement of the ctenophore Mnemiopsis leidyi suggests thatCtenophora and Porifera diverged prior to the ParaHoxozoa.Evodevo. 1:9.
Saina M, Genikhovich G, Renfer E, Technau U. 2009. BMPs andchordin regulate patterning of the directive axis in a seaanemone. Proc Natl Acad Sci U S A. 106:18592–18597.
Sato T, Yamanishi Y, Horimoto K, Kanehisa M, Toh H. 2006. Partialcorrelation coefficient between distance matrices as a newindicator of protein-protein interactions. Bioinformatics22:2488–2492.
Schierenberg E. 2001. Three sons of fortune: early embryogenesis,evolution and ecology of nematodes. Bioessays 23:841–847.
Sperling EA, Peterson KJ, Pisani D. 2009. Phylogenetic-signaldissection of nuclear housekeeping genes supports the para-phyly of sponges and the monophyly of Eumetazoa. Mol BiolEvol. 26:2261–2274.
Srivastava M, Begovic E, Chapman J, et al. 2008. The Trichoplaxgenome and the nature of placozoans. Nature 454:955–960.
Srivastava M, Simakov O, Chapman J, et al. (33 co-authors). 2010.The Amphimedon queenslandica genome and the evolution ofanimal complexity. Nature 466:720–726.
Strano S, Monti O, Pediconi N, et al. (13 co-authors). 2005. Thetranscriptional coactivator Yes-associated protein drives p73gene-target specificity in response to DNA Damage. Mol Cell.18:447–459.
Strano S, Munarriz E, Rossi M, Castagnoli L, Shaul Y, Sacchi A,Oren M, Sudol M, Cesareni G, Blandino G. 2001. Physicalinteraction with Yes-associated protein enhances p73 transcrip-tional activity. J Biol Chem. 276:15164–15173.
Sudol M, Harvey KF. 2010. Modularity in the Hippo signalingpathway. Trends Biochem Sci. 35:627–633.
Sullivan JC, Sher D, Eisenstein M, Shigesada K, Reitzel AM, Marlow H,Levanon D, Groner Y, Finnerty JR, Gat U. 2008. The evolutionaryorigin of the Runx/CBFbeta transcription factors—studies of themost basal metazoans. BMC Evol Biol. 8:228–2248.
Tapon N, Harvey KF, Bell DW, Wahrer DCR, Schiripo TA, Haber DA,Hariharan IK. 2002. Salvador promotes both cell cycle exit andapoptosis in Drosophila and is mutated in human cancer celllines. Cell 110:467–478.
Vitolo MI, Anglin IE, Mahoney WM, Renoud KJ, Gartenhaus RB,Bachman KE, Passaniti A. 2007. The RUNX2 transcription factorcooperates with the YES-associated protein, YAP65, to promotecell transformation. Cancer Biol Ther. 6:856–863.
Wang Y, Abu Irqeba A, Ayalew M, Suntay K. 2009. Sumoylation oftranscription factor Tec1 regulates signaling of mitogen-activated protein kinase pathways in yeast. PloS One. 4:e7456.
Hilman and Gat · doi:10.1093/molbev/msr065 MBE
2416

Wei X, Shimizu T, Lai ZC. 2007. Mob as tumor suppressor is
activated by hippo kinase for growth inhibition in Drosophila.
EMBO J. 26:1772–1781.Wu J, Duggan A, Chalfie M. 2001. Inhibition of touch cell fate by egl-
44 and egl-46 in C. elegans. Genes Dev. 15:789–802.Wu S, Huang J, Dong J, Pan D. 2003. Hippo encodes a Ste-20 family
protein kinase that restricts cell proliferation and promotes
apoptosis in conjunction with salvador and warts. Cell
114:445–456.Xu T, Wang W, Zhang S, Stewart R, Yu W. 1995. Identifying tumor
suppressors in genetic mosaics: the Drosophila lats gene encodes
a putative protein kinase. Development 121:1053–1063.
Yabuta N, Okada N, Ito A, et al. (12 co-authors). 2007. Lats2 is anessential mitotic regulator required for the coordination of celldivision. J Biol Chem. 282:19259–19271.
Yu J, Zheng Y, Dong J, Klusza S, Deng WM, Pan D. 2010. Kibrafunctions as a tumor suppressor protein that regulates hipposignaling in conjunction with merlin and expanded. Dev Cell.18:288–299.
Zhao B, Li L, Lei Q, Guan KL. 2010. The Hippo-YAP pathway in organsize control and tumorigenesis: an updated version. Genes Dev.24:862–874.
Zhao B, Wei X, Li W, et al. (16 co-authors). 2007. Inactivation of YAPoncoprotein by the hippo pathway is involved in cell contactinhibition and tissue growth control. Genes Dev. 21:2747–2761.
Evolution of YAP and the Hippo Pathway · doi:10.1093/molbev/msr065 MBE
2417





























