Embed Size (px)
Citation preview

The Female Genital Morphology of the Orb WeavingSpider Genus Agriognatha (Araneae, Tetragnathidae)
Dimitar Dimitrov,* Fernando Alvarez-Padilla, and Gustavo Hormiga
Department of Biological Sciences, The George Washington University, Washington, D.C. 20052
ABSTRACT The female genital morphology of the spi-ders in the araneoid genus Agriognatha (Tetragna-thidae) is described and illustrated. The female genitaliaof Agriognatha is characterized by a strong reduction ofthe sperm storage organs (spermathecae) and by the pres-ence of a specialized distal compartment of the medianmembranous chamber that functions as a sperm storageorgan (the posterior sac). The genital morphology ofAgriognatha species is unique among Tetragnathidae andit provides robust synapomorphic evidence for the mono-phyly of genus. We discuss the phylogenetic implicationsof these new findings for the placement and monophylyof Agriognatha and for the monophyly of Tetragnathinae.J. Morphol. 268:758–770, 2007. � 2007 Wiley-Liss, Inc.
KEY WORDS: Tetragnathinae; spermathecae; homology;sperm storage; haplogyne; reduction
It has been known since the seventeenth centurythat different spider species differ in their genitalmorphology. The first author to document spidergenitalia in a taxonomic work was Clerck (1757),who illustrated the male palp of some, but not all,the species he described in his monograph onSwedish spiders. Although the morphology of thefemale genitalia is also diagnostic for the species,Clerck did not illustrate it. The morphology of cop-ulatory organs in spiders is widely used now asone of the most important diagnostic charactersystems, especially at the species level (e.g., Plat-nick, 1975; Griswold, 1993; Foelix, 1996; see alsoHuber, 2004). The morphology of the copulatoryorgans is also used in phylogenetic studies and inmost of the recently published cladistic matrices itaccounts for more than half of the character data(e.g., Griswold, 1990, 1993; Hormiga, 1994, 2000;Wang, 2002; Ramırez, 2003; Miller and Hormiga,2004; Dimitrov and Ribera, in press; see Huber,2004 for further discussion).
Despite their central role in spider systematics,the copulatory organs of many species are poorlystudied and their morphology is not well under-stood. This problem is not exclusive of old taxo-nomic treatments: it is not uncommon in moderntaxonomic revisions to find genital illustrationsthat are insufficient to understand the basic mor-phology of the species in question. Illustrating gen-ital diagnostic characters does not necessarily
imply an understanding of the basic genital mor-phology. Most taxonomic illustrations are donewith the goal of depicting species diagnostic fea-tures and do not necessarily provide the informa-tion needed to understand genital morphology.This is particularly true for species with complexgenital structures. This problem is also commonfor the descriptive work of species in which thefemales present a low degree of sclerotization ofthe genitalia and do not have a well sclerotizedepigynum (the sclerotized region that covers theinternal genitalia), although there are a number ofnotable exceptions (e.g., Uhl, 1994, 2000; Michaliket al., 2005; Burger et al., 2003, 2006). It is notuncommon in taxonomic descriptions of thefemales of such species to have their genital mor-phology insufficiently described and poorly illus-trated. The spider genus Agriognatha Cambridge,1896 (Tetragnathidae) is a good example of thisproblem. Agriognatha (Fig. 1) is a small genus ofTetragnathinae spiders known from Central andSouth America. Currently ten species are knownto belong in Agriognatha, but ongoing research onthe taxonomy on this genus will increase the num-ber to over 20 species (Dimitrov and Hormiga, per-sonal observation). Agriognatha females do nothave an epigynum and the internal genital struc-tures are weakly sclerotized, a condition that istypical of the members of the subfamily Tetragna-thinae. This low degree of sclerotization makesmorphological observations of the female genitaliaof Agriognatha species difficult. Despite the avail-ability of female specimens in the original descrip-tions of several Agriognatha species the femalegenital morphology is not discussed or illustratedat all (Bryant, 1940, 1945). Furthermore, two spe-
Contract grant sponsor: U.S. National Science Foundation; Con-tract grant numbers: DEB-0328644, EAR-0228699; Contract grantsponsors: Research Enhancement Fund and The George Washing-ton University.
*Correspondence to: Dimitar Dimitrov, Department of BiologicalSciences, The George Washington University, WA, D.C. 20052.E-mail: [email protected]
Published online 30 May 2007 inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10543
JOURNAL OF MORPHOLOGY 268:758–770 (2007)
� 2007 WILEY-LISS, INC.

cies have been described from single female speci-mens but without providing a description or illus-trations of the internal female genitalia (Cam-bridge, 1889; Mello-Leitao, 1947).
Spiders in the large Araneoclada lineage (a cladethat includes about 90% of the known spider spe-cies; see (Coddington et al., 2004) have been classi-cally divided into two groups: Haplogynae andEntelegynae (Simon, 1892; Wiehle, 1967; Platnicket al., 1991; Uhl, 2002; but see Griswold et al.,2005). Support for these two groups, since theywere first proposed by Simon (1892), has come inpart from the genital morphology. In haplogynesthe female sperm storage organs (spermathecae)open to only one set of ducts, which function bothas copulatory and fertilization ducts (or in somespecies, there are no ducts at all, as in the familyPholcidae). In entelegyne spiders the spermathe-cae open to the exterior by means of the so-calledcopulatory ducts and a separate set of ducts (thefertilization ducts) connect the spermathecae tothe uterus externus. Haplogynes and entelegyneshave been said to have ‘‘cul-de-sac’’ and ‘‘conduit’’spermathecae, respectively, because in the firsttype of spermathecae the seminal fluid enters andexits through the same duct and in the second
type there are separate ducts for the seminal fluidto enter and leave (Austad, 1984). The haplogynecondition is primitive and consequently a symple-siomorphy in Haplogynae. The entelegyne condi-tion is derived (apomorphic) and thus it providessupport to the monophyly of Entelegynae. Never-theless this ‘‘division’’ is far from perfect: a numberof taxa within Entelegynae have evolved haplo-gyne genitalia (e.g., some tetragnathines; seebelow) and some members of Haplogynae have fer-tilization ducts, such as some pholcids (Huber,1997) or some tetrablemmids (Burger et al., 2006).It seems clear that the notion that these two line-ages have somewhat uniform female genitalia isnot empirically supported (Uhl, 2002).
Most species of the family Tetragnathidae, aswell as the rest of araneoid spiders, have entele-gyne genitalia (but note that the exact circum-scription of Araneoidea awaits resolution; see Gris-wold et al., 2005). In the tetragnathid subfamilyTetragnathinae (which includes the genera Tetrag-natha, Agriognatha, Glenognatha, and Pachygna-tha, among others; see Hormiga et al., 1995) thehaplogyne condition has evolved independently ofthat found in the Haplogynae clade and the basalaraneomorph lineages: the fertilization ducts have
Fig. 1. Agriognatha rucilla in its web. Dominican Republic. Photo by Gustavo Hormiga.
FEMALE GENITAL MORPHOLOGY OF AGRIOGNATHA 759
Journal of Morphology DOI 10.1002/jmor
been secondarily lost and there has been a generalsimplification of the female genital structures.Such simplification of the entelegyne genitalia to asecondary haplogyny condition is a rare evolution-ary event. Secondary haplogyny probably evolvedindependently several times within the entele-gyne group, e.g., in the families Archaeidae andHuttoniidae. However, the phylogenetic place-ment of these two families is still unresolved and
the results under certain character weightinganalyses suggest that they may be primitivelyhaplogyne (Griswold et al., 2005: Fig. 219). Primi-tive haplogyny had been previously proposed forArchaeidae (Forster and Platnick, 1984) but inthe light of the new evidence presented by Gris-wold et al. (2005) both hypotheses are plausible.Despite the enormous advances brought by thework of Griswold et al. (2005), the answer to this
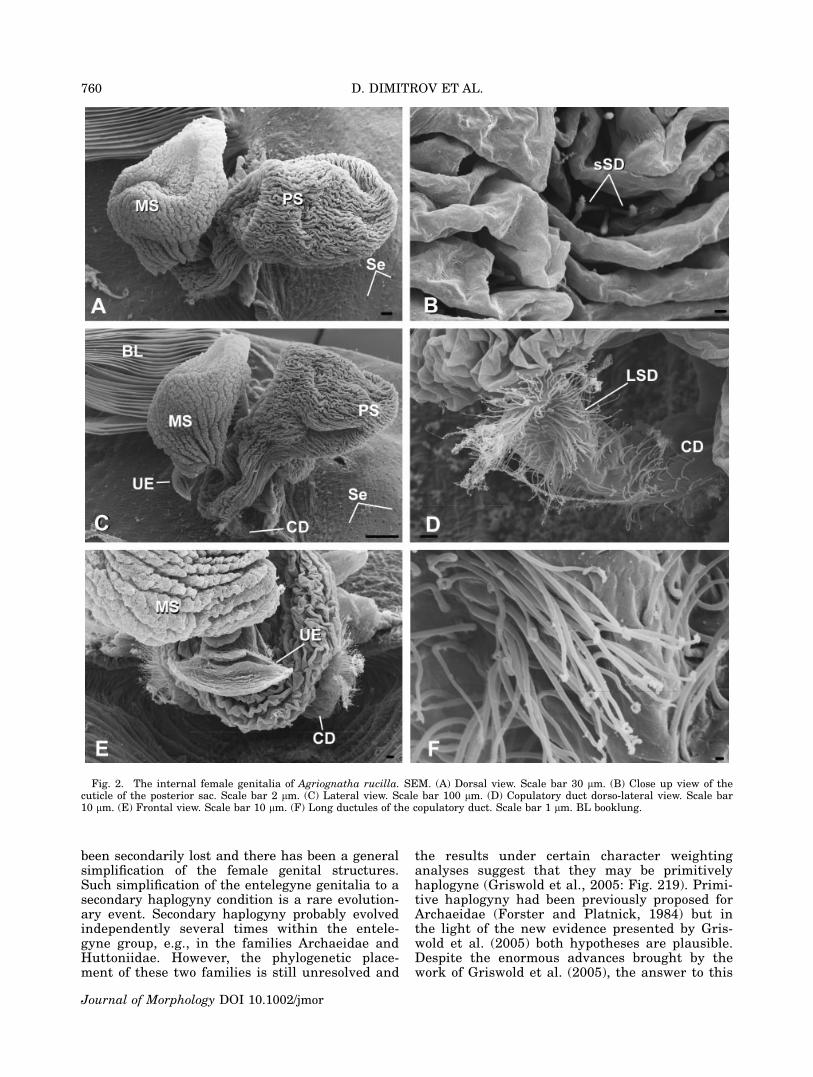
Fig. 2. The internal female genitalia of Agriognatha rucilla. SEM. (A) Dorsal view. Scale bar 30 lm. (B) Close up view of thecuticle of the posterior sac. Scale bar 2 lm. (C) Lateral view. Scale bar 100 lm. (D) Copulatory duct dorso-lateral view. Scale bar10 lm. (E) Frontal view. Scale bar 10 lm. (F) Long ductules of the copulatory duct. Scale bar 1 lm. BL booklung.
760 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor
question requires a more resolved and robust phy-logenetic hypothesis.
During the course of systematic studies of tet-ragnathid spiders we have examined in somedetail the genital morphology of several Agriogna-tha species as well as some other closely relatedgenera. We were very surprised to discover that inAgriognatha species the spermathecae were so dra-matically reduced that they are unlikely to be func-tional (as sperm storage organs) and that the inter-
nal female genitalia exhibited a very atypical mor-phology. The most unusual observation, togetherwith the reduced spermathecae, was the presence ofan unpaired sac that holds the sperm. The goal ofthis article is to describe and illustrate the peculiarmorphology of the female genitalia in several repre-sentatives of Agriognatha and to provide some com-ments on their possible phylogenetic significance.
We did not make histological preparations andsome questions regarding the morphology and the
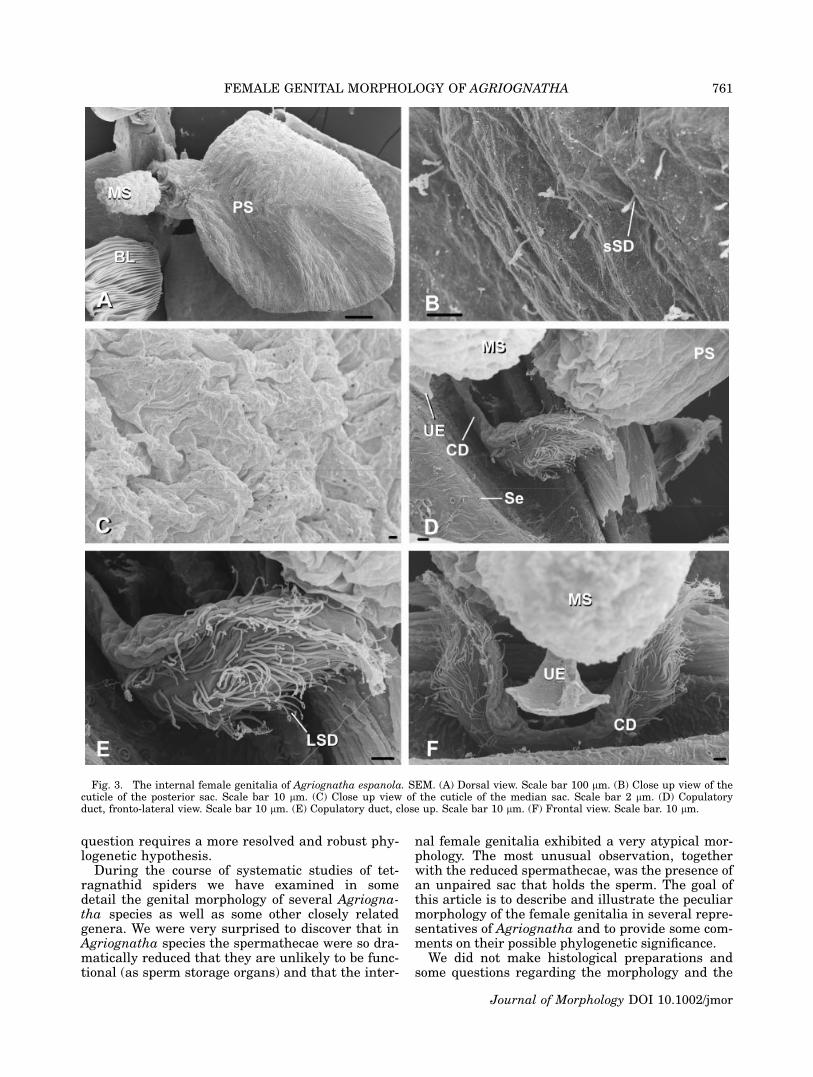
Fig. 3. The internal female genitalia of Agriognatha espanola. SEM. (A) Dorsal view. Scale bar 100 lm. (B) Close up view of thecuticle of the posterior sac. Scale bar 10 lm. (C) Close up view of the cuticle of the median sac. Scale bar 2 lm. (D) Copulatoryduct, fronto-lateral view. Scale bar 10 lm. (E) Copulatory duct, close up. Scale bar 10 lm. (F) Frontal view. Scale bar. 10 lm.
FEMALE GENITAL MORPHOLOGY OF AGRIOGNATHA 761
Journal of Morphology DOI 10.1002/jmor
function of the documented structures remainunanswered. Despite this, the results presentedhere provide a starting point for future studies onthese spiders. Additionally, some of the featuresreported herein are documented for the first timein araneoid spiders and we hope that they will cat-alyze future investigations to help us better under-stand the process of copulation and sperm transferin this group.
MATERIALS AND METHODS
Female genitalia were dissected and the nonchitinous abdom-inal tissue was digested with SIGMA Pancreatin LP 1750enzyme complex (Alvarez Padilla and Hormiga, in press), in asolution of sodium borate prepared following the concentrationsdescribed by Dingerkus and Uhler (1977). After removing anyremaining tissues with needles, the preparations were washedin distilled water and transferred to 75% ethanol. Cleaned geni-talia were observed and illustrated in 75% ethanol using a
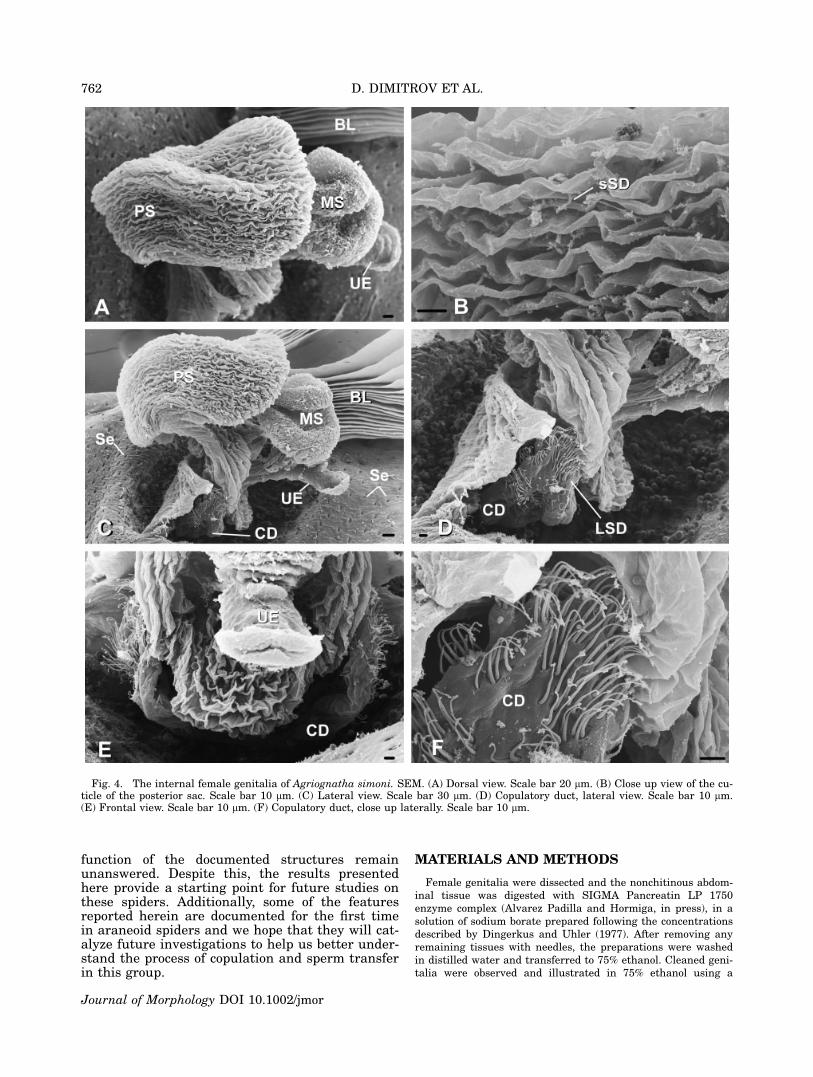
Fig. 4. The internal female genitalia of Agriognatha simoni. SEM. (A) Dorsal view. Scale bar 20 lm. (B) Close up view of the cu-ticle of the posterior sac. Scale bar 10 lm. (C) Lateral view. Scale bar 30 lm. (D) Copulatory duct, lateral view. Scale bar 10 lm.(E) Frontal view. Scale bar 10 lm. (F) Copulatory duct, close up laterally. Scale bar 10 lm.
762 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor
Leica MZ16 stereoscopic microscope with a camera lucida. High
resolution digital photos of the preparations were taken with a
Nikon DXM1200F digital camera attached to a Leica MZ16A
stereoscope. Additional details were checked with a Leica
DMRM compound microscope. After illustrating and photo-
graphing the cleaned genitalia, the preparations were critically
point dried and sputter coated with gold-palladium for SEM ex-
amination with a LEO 1430VP scanning electron microscope.
RESULTS
We examined specimens of four species ofAgriognatha (A. rucilla Bryant, 1945; A. simoniBryant, 1940, A. insolita Chickering, 1956 and A.espanola Bryant, 1945). In addition we also exam-ined specimens of closely related tetragnathinetaxa: Tetragnatha versicolor (Walckenaer, 1842),
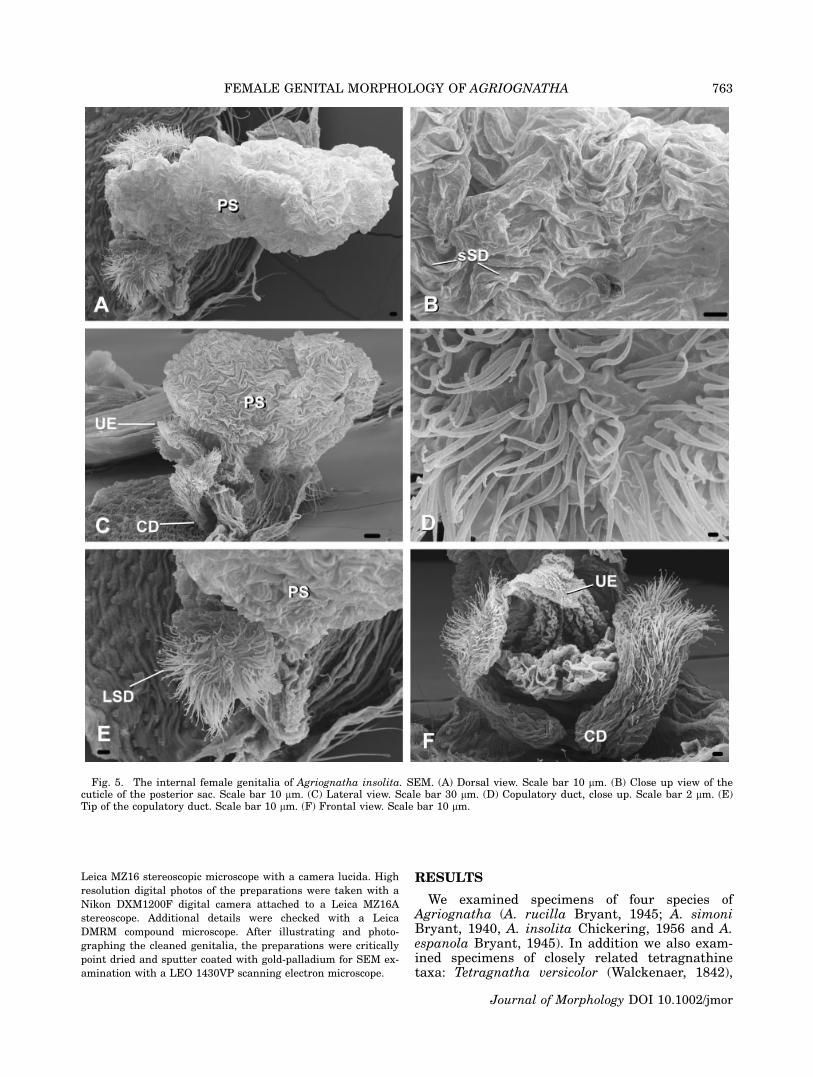
Fig. 5. The internal female genitalia of Agriognatha insolita. SEM. (A) Dorsal view. Scale bar 10 lm. (B) Close up view of thecuticle of the posterior sac. Scale bar 10 lm. (C) Lateral view. Scale bar 30 lm. (D) Copulatory duct, close up. Scale bar 2 lm. (E)Tip of the copulatory duct. Scale bar 10 lm. (F) Frontal view. Scale bar 10 lm.
FEMALE GENITAL MORPHOLOGY OF AGRIOGNATHA 763
Journal of Morphology DOI 10.1002/jmor
Tetragnatha sp. (from the Dominican Republic),Glenognatha foxi (McCook, 1894) and Pachygnathaautumnalis (Marx, 1884) (see Appendix). Theexternally visible genital opening in all the studiedspecies is formed by a ventral lobe of the abdomi-nal cuticle, which overhangs the actual opening(which is more internal) and leads into a chamberwhose dorsal wall is a continuation of the ventralabdominal cuticle. In all the species examined forthis study the dorsal cuticle in this area still car-ries the abdominal setae (Se in Figs. 2A, 4C, 6D,9C). In Agriognatha species this chamber con-nects with a pair of short ducts anteriorly (thecopulatory ducts or ‘‘connecting ducts’’ sensu Levi,1980) and with another wider duct (or chamber),which runs dorsally and divides into one or twosacs (two in A. rucilla, A. simoni, and A. espa-nola; Figs. 2A, C, E, 4A, C, E and 3A, D, F,respectively) and one in A. insolita (Fig. 5A, C,F). These sacs are medially arranged, along thelongitudinal axis of the abdomen. We have namedthese two sacs according to their position as me-dian and posterior sacs (MS and PS, respectively).The copulatory ducts (CD) share a wide common
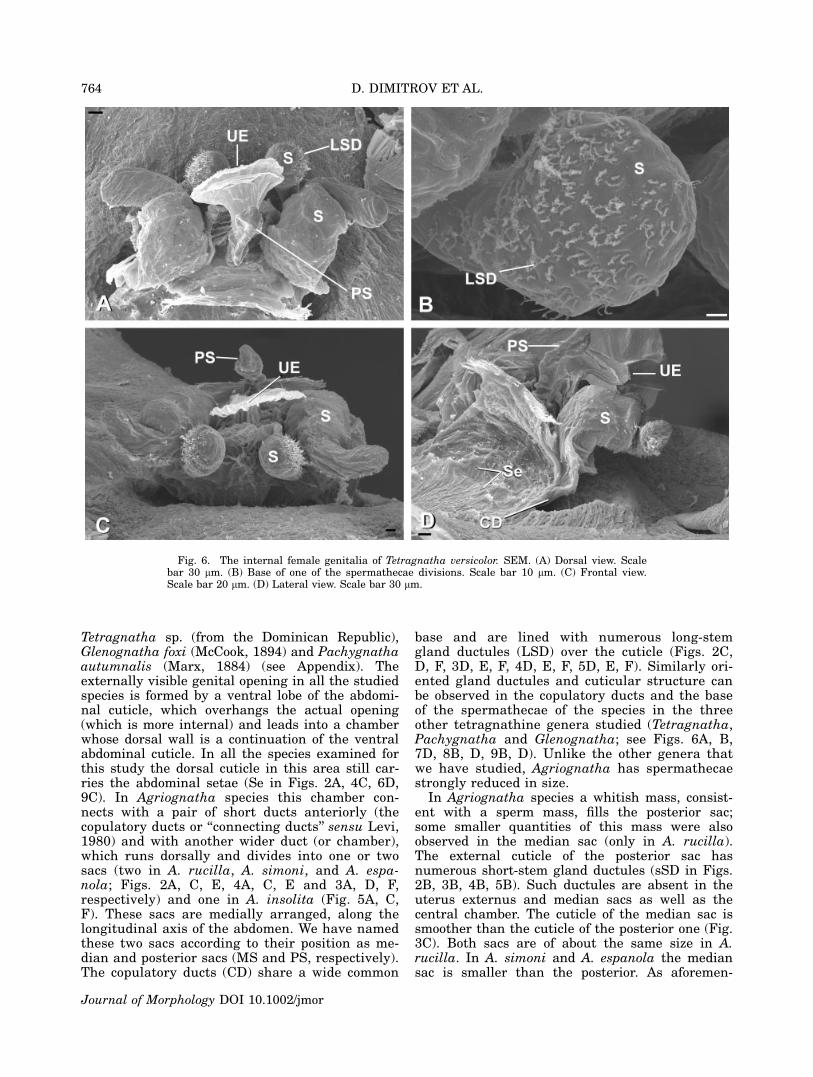
base and are lined with numerous long-stemgland ductules (LSD) over the cuticle (Figs. 2C,D, F, 3D, E, F, 4D, E, F, 5D, E, F). Similarly ori-ented gland ductules and cuticular structure canbe observed in the copulatory ducts and the baseof the spermathecae of the species in the threeother tetragnathine genera studied (Tetragnatha,Pachygnatha and Glenognatha; see Figs. 6A, B,7D, 8B, D, 9B, D). Unlike the other genera thatwe have studied, Agriognatha has spermathecaestrongly reduced in size.
In Agriognatha species a whitish mass, consist-ent with a sperm mass, fills the posterior sac;some smaller quantities of this mass were alsoobserved in the median sac (only in A. rucilla).The external cuticle of the posterior sac hasnumerous short-stem gland ductules (sSD in Figs.2B, 3B, 4B, 5B). Such ductules are absent in theuterus externus and median sacs as well as thecentral chamber. The cuticle of the median sac issmoother than the cuticle of the posterior one (Fig.3C). Both sacs are of about the same size in A.rucilla. In A. simoni and A. espanola the mediansac is smaller than the posterior. As aforemen-
Fig. 6. The internal female genitalia of Tetragnatha versicolor. SEM. (A) Dorsal view. Scalebar 30 lm. (B) Base of one of the spermathecae divisions. Scale bar 10 lm. (C) Frontal view.Scale bar 20 lm. (D) Lateral view. Scale bar 30 lm.
764 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor
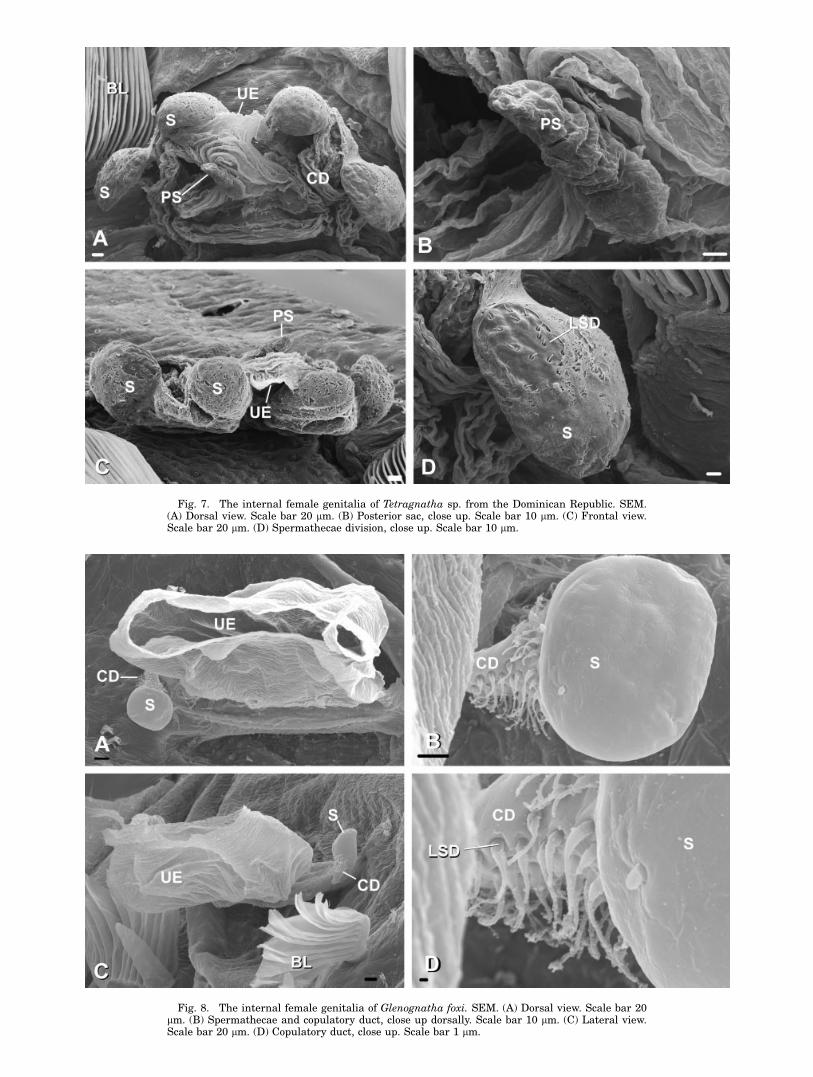
Fig. 7. The internal female genitalia of Tetragnatha sp. from the Dominican Republic. SEM.(A) Dorsal view. Scale bar 20 lm. (B) Posterior sac, close up. Scale bar 10 lm. (C) Frontal view.Scale bar 20 lm. (D) Spermathecae division, close up. Scale bar 10 lm.
Fig. 8. The internal female genitalia of Glenognatha foxi. SEM. (A) Dorsal view. Scale bar 20lm. (B) Spermathecae and copulatory duct, close up dorsally. Scale bar 10 lm. (C) Lateral view.Scale bar 20 lm. (D) Copulatory duct, close up. Scale bar 1 lm.
tioned, A. insolita does not appear to have a me-dian sac.
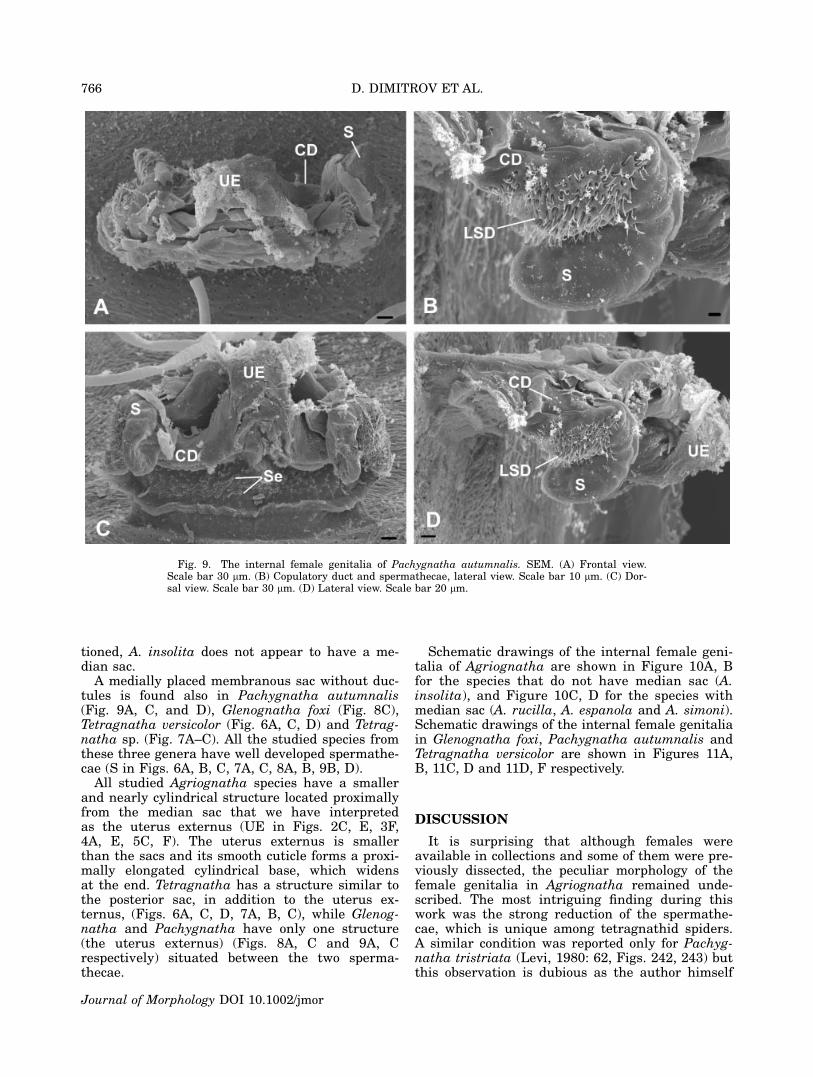
A medially placed membranous sac without duc-tules is found also in Pachygnatha autumnalis(Fig. 9A, C, and D), Glenognatha foxi (Fig. 8C),Tetragnatha versicolor (Fig. 6A, C, D) and Tetrag-natha sp. (Fig. 7A–C). All the studied species fromthese three genera have well developed spermathe-cae (S in Figs. 6A, B, C, 7A, C, 8A, B, 9B, D).
All studied Agriognatha species have a smallerand nearly cylindrical structure located proximallyfrom the median sac that we have interpretedas the uterus externus (UE in Figs. 2C, E, 3F,4A, E, 5C, F). The uterus externus is smallerthan the sacs and its smooth cuticle forms a proxi-mally elongated cylindrical base, which widensat the end. Tetragnatha has a structure similar tothe posterior sac, in addition to the uterus ex-ternus, (Figs. 6A, C, D, 7A, B, C), while Glenog-natha and Pachygnatha have only one structure(the uterus externus) (Figs. 8A, C and 9A, Crespectively) situated between the two sperma-thecae.
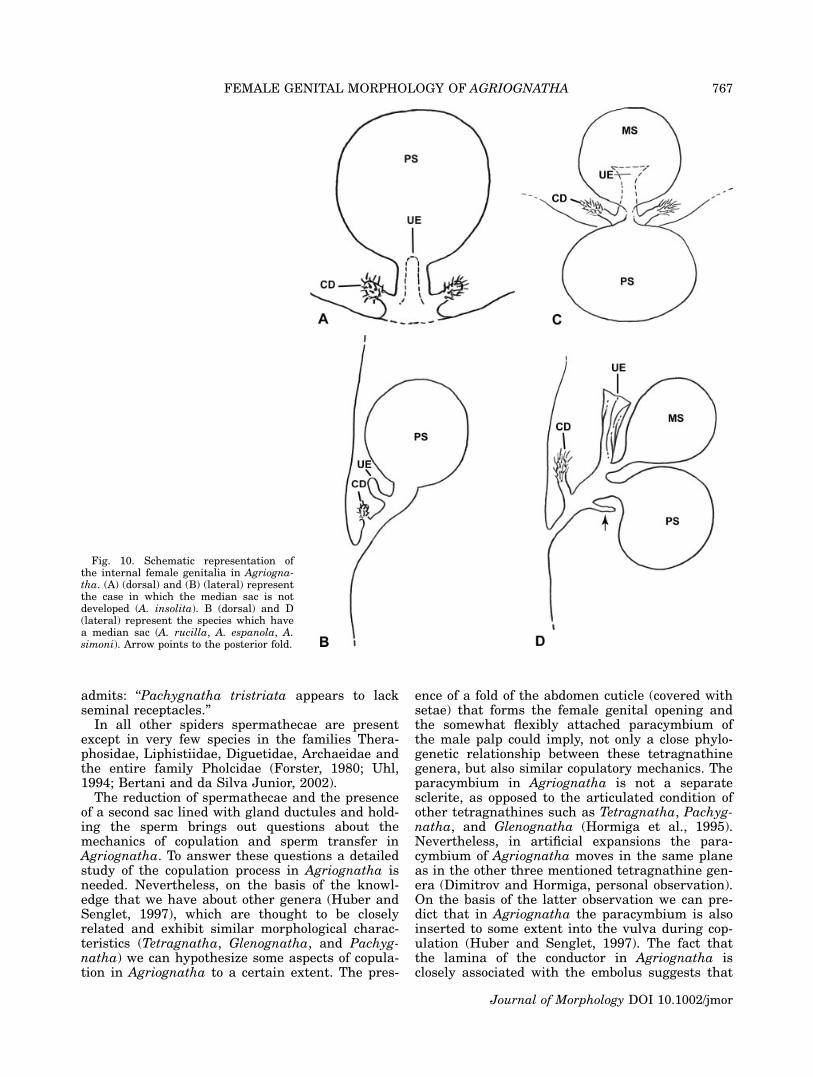
Schematic drawings of the internal female geni-talia of Agriognatha are shown in Figure 10A, Bfor the species that do not have median sac (A.insolita), and Figure 10C, D for the species withmedian sac (A. rucilla, A. espanola and A. simoni).Schematic drawings of the internal female genitaliain Glenognatha foxi, Pachygnatha autumnalis andTetragnatha versicolor are shown in Figures 11A,B, 11C, D and 11D, F respectively.
DISCUSSION
It is surprising that although females wereavailable in collections and some of them were pre-viously dissected, the peculiar morphology of thefemale genitalia in Agriognatha remained unde-scribed. The most intriguing finding during thiswork was the strong reduction of the spermathe-cae, which is unique among tetragnathid spiders.A similar condition was reported only for Pachyg-natha tristriata (Levi, 1980: 62, Figs. 242, 243) butthis observation is dubious as the author himself
Fig. 9. The internal female genitalia of Pachygnatha autumnalis. SEM. (A) Frontal view.Scale bar 30 lm. (B) Copulatory duct and spermathecae, lateral view. Scale bar 10 lm. (C) Dor-sal view. Scale bar 30 lm. (D) Lateral view. Scale bar 20 lm.
766 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor
admits: ‘‘Pachygnatha tristriata appears to lackseminal receptacles.’’
In all other spiders spermathecae are presentexcept in very few species in the families Thera-phosidae, Liphistiidae, Diguetidae, Archaeidae andthe entire family Pholcidae (Forster, 1980; Uhl,1994; Bertani and da Silva Junior, 2002).
The reduction of spermathecae and the presenceof a second sac lined with gland ductules and hold-ing the sperm brings out questions about themechanics of copulation and sperm transfer inAgriognatha. To answer these questions a detailedstudy of the copulation process in Agriognatha isneeded. Nevertheless, on the basis of the knowl-edge that we have about other genera (Huber andSenglet, 1997), which are thought to be closelyrelated and exhibit similar morphological charac-teristics (Tetragnatha, Glenognatha, and Pachyg-natha) we can hypothesize some aspects of copula-tion in Agriognatha to a certain extent. The pres-
ence of a fold of the abdomen cuticle (covered withsetae) that forms the female genital opening andthe somewhat flexibly attached paracymbium ofthe male palp could imply, not only a close phylo-genetic relationship between these tetragnathinegenera, but also similar copulatory mechanics. Theparacymbium in Agriognatha is not a separatesclerite, as opposed to the articulated condition ofother tetragnathines such as Tetragnatha, Pachyg-natha, and Glenognatha (Hormiga et al., 1995).Nevertheless, in artificial expansions the para-cymbium of Agriognatha moves in the same planeas in the other three mentioned tetragnathine gen-era (Dimitrov and Hormiga, personal observation).On the basis of the latter observation we can pre-dict that in Agriognatha the paracymbium is alsoinserted to some extent into the vulva during cop-ulation (Huber and Senglet, 1997). The fact thatthe lamina of the conductor in Agriognatha isclosely associated with the embolus suggests that
Fig. 10. Schematic representation ofthe internal female genitalia in Agriogna-tha. (A) (dorsal) and (B) (lateral) representthe case in which the median sac is notdeveloped (A. insolita). B (dorsal) and D(lateral) represent the species which havea median sac (A. rucilla, A. espanola, A.simoni). Arrow points to the posterior fold.
FEMALE GENITAL MORPHOLOGY OF AGRIOGNATHA 767
Journal of Morphology DOI 10.1002/jmor

at least in some cases the conductor is inserted to-gether with the embolus as in Tetragnatha (Huberand Senglet, 1997).
The female genitalia of all four tetragnathinegenera studied here have more or less complexmembranous unpaired structures located medially.At least in Agriognatha the most distal chamber(the posterior sac) appears to be responsible forsperm storage (Fig. 12A, B). The function of themedian sac remains unknown. The presence ofmembranous median sacs was previously reportedin Pachygnatha and Tetragnatha by Whiele (1963)and Levi (1980, 1981). Levi (1981) suggested thatthe ‘‘median seminal receptacles’’ are used to holdthe conductor when mating. Later, Huber and
Senglet (1997), studying copulation of Tetragnathamontana, demonstrated that the conductoractually reached the entrance of the spermatheca.Neither the conductor nor other palpal scelriteswere inserted in the median sac. On the otherhand, Huber and Senglet (1997) observed that theparacymbia of T. montana, T. extensa and Pachyg-natha clerki are also inserted in the genital open-ing and that it is pressed against the posteriorwall of the genital opening.
An important question that directly affects theevolutionary interpretation of the newly describedstructures in Agriognatha is hypothesizing its ho-mologous counterparts in closely related taxa. Thesmall cylindrical sac situated posteriorly to the
Fig. 11. Schematic representation ofthe internal female genitalia in other Tet-ragnathinae genera. Glenognatha foxi. (A)dorsal, (B) lateral. Pachygnatha autum-nales; (C) dorsal, (D) lateral. Tetragnathaversicolor; (E) dorsal, (F) lateral. Arrowpoints to the posterior fold.
768 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor
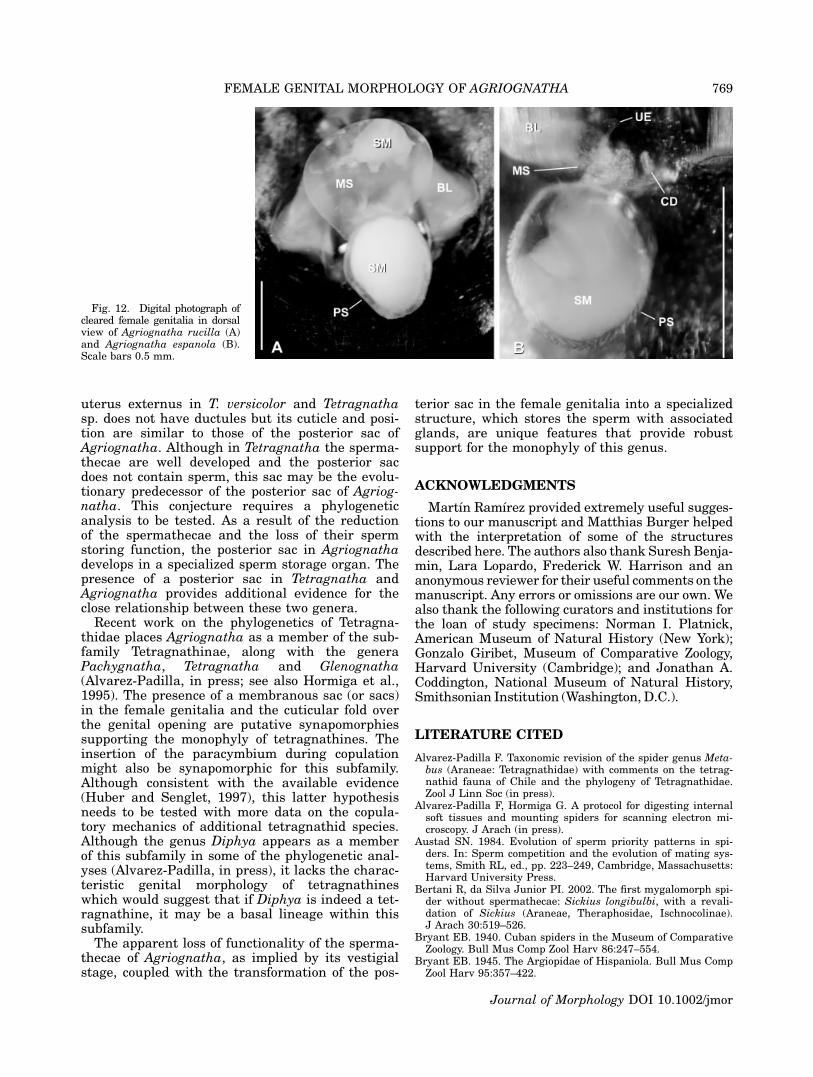
uterus externus in T. versicolor and Tetragnathasp. does not have ductules but its cuticle and posi-tion are similar to those of the posterior sac ofAgriognatha. Although in Tetragnatha the sperma-thecae are well developed and the posterior sacdoes not contain sperm, this sac may be the evolu-tionary predecessor of the posterior sac of Agriog-natha. This conjecture requires a phylogeneticanalysis to be tested. As a result of the reductionof the spermathecae and the loss of their spermstoring function, the posterior sac in Agriognathadevelops in a specialized sperm storage organ. Thepresence of a posterior sac in Tetragnatha andAgriognatha provides additional evidence for theclose relationship between these two genera.
Recent work on the phylogenetics of Tetragna-thidae places Agriognatha as a member of the sub-family Tetragnathinae, along with the generaPachygnatha, Tetragnatha and Glenognatha(Alvarez-Padilla, in press; see also Hormiga et al.,1995). The presence of a membranous sac (or sacs)in the female genitalia and the cuticular fold overthe genital opening are putative synapomorphiessupporting the monophyly of tetragnathines. Theinsertion of the paracymbium during copulationmight also be synapomorphic for this subfamily.Although consistent with the available evidence(Huber and Senglet, 1997), this latter hypothesisneeds to be tested with more data on the copula-tory mechanics of additional tetragnathid species.Although the genus Diphya appears as a memberof this subfamily in some of the phylogenetic anal-yses (Alvarez-Padilla, in press), it lacks the charac-teristic genital morphology of tetragnathineswhich would suggest that if Diphya is indeed a tet-ragnathine, it may be a basal lineage within thissubfamily.
The apparent loss of functionality of the sperma-thecae of Agriognatha, as implied by its vestigialstage, coupled with the transformation of the pos-
terior sac in the female genitalia into a specializedstructure, which stores the sperm with associatedglands, are unique features that provide robustsupport for the monophyly of this genus.
ACKNOWLEDGMENTS
Martın Ramırez provided extremely useful sugges-tions to our manuscript and Matthias Burger helpedwith the interpretation of some of the structuresdescribed here. The authors also thank Suresh Benja-min, Lara Lopardo, Frederick W. Harrison and ananonymous reviewer for their useful comments on themanuscript. Any errors or omissions are our own. Wealso thank the following curators and institutions forthe loan of study specimens: Norman I. Platnick,American Museum of Natural History (New York);Gonzalo Giribet, Museum of Comparative Zoology,Harvard University (Cambridge); and Jonathan A.Coddington, National Museum of Natural History,Smithsonian Institution (Washington, D.C.).
LITERATURE CITED
Alvarez-Padilla F. Taxonomic revision of the spider genus Meta-bus (Araneae: Tetragnathidae) with comments on the tetrag-nathid fauna of Chile and the phylogeny of Tetragnathidae.Zool J Linn Soc (in press).
Alvarez-Padilla F, Hormiga G. A protocol for digesting internalsoft tissues and mounting spiders for scanning electron mi-croscopy. J Arach (in press).
Austad SN. 1984. Evolution of sperm priority patterns in spi-ders. In: Sperm competition and the evolution of mating sys-tems, Smith RL, ed., pp. 223–249, Cambridge, Massachusetts:Harvard University Press.
Bertani R, da Silva Junior PI. 2002. The first mygalomorph spi-der without spermathecae: Sickius longibulbi, with a revali-dation of Sickius (Araneae, Theraphosidae, Ischnocolinae).J Arach 30:519–526.
Bryant EB. 1940. Cuban spiders in the Museum of ComparativeZoology. Bull Mus Comp Zool Harv 86:247–554.
Bryant EB. 1945. The Argiopidae of Hispaniola. Bull Mus CompZool Harv 95:357–422.
Fig. 12. Digital photograph ofcleared female genitalia in dorsalview of Agriognatha rucilla (A)and Agriognatha espanola (B).Scale bars 0.5 mm.
Journal of Morphology DOI 10.1002/jmor
FEMALE GENITAL MORPHOLOGY OF AGRIOGNATHA 769

Burger M, Nentwig W, Kropf C. 2003. Complex genital struc-tures indicate cryptic female choice in a haplogyne spider(Arachnida, Araneae, Oonopidae, Gamasomorphinae). J Mor-phol 255:80–93.
Burger M, Michalik P, Graber W, Jacob A, Nentwig W, Kropf C.2006. The complex genital system of a haplogyne spider(Arachnida, Araneae, Tetrablemmidae) indicates internal fer-tilization and full female control over transferred sperm.J Morphol 267:166–186.
Cambridge OP. 1889. Arachnida. Araneida. In Porter RH, editor.Biologia Centrali-Americana, Zoology, Vol 1. London. pp 1–56.
Cambridge OP. 1896. Arachnida. Araneida. In Porter RH, edi-tor. Biologia Centrali-Americana, Zoology, Vol 1. London.pp 161–224.
Chickering AM. 1956. A new species of Agriognatha (Araneae,Argiopidae) from Panama. Breviora 65:1–7.
Clerck C. 1757. Svenska spindlar, uti sina hufvud-slagter indeltesamt under nagra och sextio sarskildte arter beskrefne ochmed illuminerade figurer uplyste. Stockholmiae, 154 pp.
Coddington JA, Giribet G, Harvey MS, Prendini L, Walter DE.2004. Arachnida. In Cracraft J and Donoghue MJ, editors.Assembling the Tree of Life. New York: Oxford UniversityPress. pp 296–318.
Dimitrov D, Ribera C. The genus Pholcus (Araneae, Pholcidae)in the Canary Islands. Zool J Linn Soc (in press).
Dingerkus G, Uhler LD. 1977. Enzyme clearing of alcian blue-stained whole small vertebrates for demonstration of carti-lage. Stain Technol 52:229–232.
Foelix RF. 1996. Biology of spiders, 2nd ed. New York: OxfordUniversity Press.
Forster RR. 1980. Evolution of the tarsal organ, the respiratorysystem and the female genitalia in spiders. In Gruber J, edi-tor. Verhandlungen des 8. Internationalen Arachnologen Kon-gress, Wien 1980. pp. 269–284.
Forster RR, Platnick NI. 1984. A review of the archaeid spidersand their relatives, with notes on the limits of the superfam-ily Palpimanoidea (Arachnida, Araneae). Bull Am Mus NatHist 178:1–106.
Griswold CE. 1990. A revision and phylogenetic analysis of thespider subfamily Phyxelidinae (Araneae, Amaurobiidae). BullAmer Mus Nat Hist 196:1–206.
Griswold CE. 1993. Investigations into the phylogeny of thelycosoid spiders and their relatives (Arachnida: Araneae:Lycosoidea). Smiths Contr Zool 539:1–39.
Griswold CE, Ramırez MJ, Coddington JA, Platnick NI. 2005.Atlas of phylogenetic data for entelegyne spiders (Araneae:Araneomorphae: Entelegynae) with comments on their phy-logeny. Proc Calif Acad Sci 56 (Suppl II):1–324.
Hormiga G. 1994. A revision and cladistic analysis of the spiderfamily Pimoidae (Araneoidea: Araneae). Smith Contr Zool549:1–157.
Hormiga G. 2000. Higher level phylogenetics of erigonine spi-ders (Araneae, Linyphiidae, Erigoninae). Smiths Contr Zool609:1–160.
Hormiga G, Eberhard WG, Coddington JA. 1995. Web construc-tion behavior in Australian Phonognatha and the phylogenyof nephiline and tetragnathid spiders (Araneae, Tetragnathi-dae). Aust J Zool 43:313–364.
Huber BA. 1997. On American ‘Micromerys’ and Metagonia(Araneae, Pholcidae), with notes on natural history and geni-tal mechanics. Zool Scripta 25:341–363.
Huber BA. 2004. The significance of copulatory structures inspider systematics. In: Schult J, editor. Studien zur Theorieder Biologie, Band 5, Biosemiotik—Praktische Anwendungund Konsequenzen fuer die Einzeldisziplinen. VWB-Verlagfuer Wissenschaft und Bildung. Berlin.
Huber BA, Senglet A. 1997. Copulation with contralateral inser-tion in entelegyne spiders (Araneae: Entelegynae: Tetragna-thidae). Neth J Zool 47:99–102.
Levi HW. 1980. The orb-weaver genus Mecynogea, the subfam-ily Metinae and the genera Pachygnatha, Glenognatha andAzilia of the subfamily Tetragnathinae north of Mexico (Ara-neae: Araneidae). Bull Mus Comp Zool Harv 149:1–74.
Levi HW. 1981. The American orb-weaver genera Dolichogna-tha and Tetragnatha north of Mexico (Araneae: Araneidae,Tetragnathinae). Bull Mus Comp Zool Harv 149:271–318.
Mello-Leitao CF. de. 1947. Aranhas de Carmo do Rio Claro(Minas Gerais) coligidas pelo naturalista Jose C. M. Carvalho.Bolm Mus nac. Rio de J. (N.S., Zool) 80:1–34.
Michalik P, Reiher W, Suhm-Tintelnot M, Coyle FA, Alberti G.2005. The female genital system of the folding-trapdoor spiderAntrodiaetus unicolor (Hentz, 1842) (Antrodiaetidae, Araneae)-an ultrastructural study of form and function with notes onreproductive biology of spiders. J Morphol 263:284–309.
Miller JA, Hormiga G. 2004. Clade Stability and the Addition ofData - A Case Study from Erigonine Spiders (Araneae: Liny-phiidae, Erigoninae). Cladistics 20:385–442.
Platnick NI. 1975. A revision of the palpimanid spiders of thenew subfamily Otiothopinae (Araneae, Palpimanidae). AmerMus Novitates 2562:1–32.
Platnick NI, Coddington JA, Forster RR, Griswold CE. 1991.Spinneret morphology and the phylogeny of haplogyne spiders(Araneae, Araneomorphae). Amer Mus Novitates 3016: 1–73.
Ramırez MJ. 2003. The spider subfamily Amaurobioidinae (Ara-neae, Anyphaenidae): A phylogenetic revision at the genericlevel. Bull Am Mus Nat Hist 277:1–262.
Uhl G. 1994. Genital morphology and sperm storage in Pholcusphalangioides (Fuesslin, 1775) (Pholcidae; Araneae). ActaZool Stockholm 75:1–12.
Simon E. 1892. Histoire naturelle des araignees. Paris 1:1–256.Uhl G. 2000. Two distinctly different sperm storage organs in
female Dysdera erythrina (Araneae: Dysderidae). ArthropodStruct Dev 29:163–169.
Uhl G. 2002. Female genital morphology and sperm prioritypatterns in spiders. In Toft S, Scharff N, editors. EuropeanArachnology. Aarhus N, Denmark: Aarhus University Press.pp. 146–156.
Walckenaer CA. 1842. Histoire naturelle des Insects. Apteres.Paris. 2:1–549.
Wang X.-P. 2002. A generic-level revision of the spider subfam-ily Coelotinae (Araneae, Amaurobiidae). Bull Amer Mus NatHist 269:1–150.
Wiehle H. 1963. Spinnentiere order Arachnoidea (Araneae). XII.Tetragnathidae-Streckspinnen und Dickkiefer. Die TierweltDeutschlands, Jena. pp 1–76.
Wiehle H. 1967. Meta,-eine semientelegyne Gattung der Ara-neae. Senckenberg Biol 48:183–196.
APPENDIX
Species list and collection data of the studied specimens. Mu-seum abbreviations: AMNH (American Museum of Natural His-tory, New York); MCZ (Museum of Comparative Zoology, Har-vard University, Cambridge); USNM (National Museum of Nat-ural History, Smithsonian Institution, Washington, D.C.).
Agriognatha rucilla (Bryant, 1945)—Dominican Republic,Barahona province, Reserva Natural Cachote, cloud forest andsecondary growth. N 18805054.8@: W 71811022.0@, 1220 m, 6-9.iv.2005, G. Hormiga leg. (MCZ).
Agriognatha espanola (Bryant, 1945) Dominican Republic,Barahona province, Reserva Natural Cachote, cloud forest andsecondary growth. N 18805054.8@: W 71811022.0@, 1220 m, 6-9.iv.2005, G. Hormiga, F. Alvarez and S. Benjamin leg. (MCZ).
Agriognatha simoni (Bryant, 1940) - Cuba, Pico Turquino,Cuba. vi.1936, P.J. Darlington leg. (MCZ).
Tetragnatha versicolor (Walckenaer, 1842)-USA VA, Waterlik 5.5miles S of 678 rd. 04.vi.2005, F. Alvarez and S. Rood leg. (MCZ).
Tetragnatha sp.—Dominican Republic, La Vega Province, ReservaCientifica Valle Nuevo, pine forest, N 18846039.2@: W 70838024.4@,2277 m, 11.iv.2005, F. Alvarez and S. Benjamin leg. (MCZ).
Pachygnatha autumnalis (Marx, 1884)-USA MA Quisset Co.24-03-1989 R.L. Edwards (USNM).
Glenognatha foxi (McCook, 1894)-USA LA, Hamburg Co. 29Apr 1963 (AMNH).
770 D. DIMITROV ET AL.
Journal of Morphology DOI 10.1002/jmor





























