Embed Size (px)
Citation preview

The fine structure of the respiratory tree in Cucumaria
By WILLIAM L. DOYLE and G. FRANCES McNIELL(From the Department of Anatomy, University of Chicago)
With 6 plates (figs, i to 6)
SummaryThe delicate tubules of the respiratory tree consist of 4 layers: a lining epithelium,a thick mucoid layer containing collagenous filaments, a smooth muscle net, and acoelomic epithelium. The free surfaces of both epithelia have well developed plas-modesms. Amoebocytes are present in all layers and the spherules of one type areconsidered to be precursors of the mucoid substance; another amoebocyte may be afibroblast. Perpendicularly oriented smooth muscle fibres, as well as those parallel toeach other, are linked by desmosomes ensuring synchronous contraction. Secretoryactivity is evident in distended cisternae of the endoplasmic reticulum of certainepithelial cells and in the vacuoles of the lining epithelium.
IntroductionT H E respiratory trees of some holothurians such as Cucumaria frondosa arehighly branched thin-walled diverticula of the rectal portion of the gut. Thesedelicate tubules fill the coelomic cavity and surround the internal organs.Slow rhythmic pulsations of the body wall result in alternate filling andemptying of the respiratory trees. It has been demonstrated that they functionin aerating the tissues and that they are permeable to water in both directions.On the other hand several different investigators have concluded that they areimpermeable to common ions and to sugars (Hyman, 1955). The coelomicfluid is reported to have the same ionic composition as sea water but witha lower pH and a small amount of nitrogenous material. Dyes injected intothe coelom are rapidly eliminated and various investigators have attributedthis transport to the activity of amoebocytes passing through the epithelia.Hyman (1955) concludes that 'from what is known of the physiology of holo-thurians it would seem that these animals operate on a very primitive basis,that each organ system covers more than its usual function, and that amoe-bocytes play a remarkable role in the economy'. The present study addsfurther support to these conclusions and provides some new observations onthe nature of the barrier between the coelom and sea water.
Materials and methodsCucumaria frondosa were collected from rocks about 6 feet below low-tide
levels in Frenchman Bay, Maine, adjacent to the Mt. Desert Island BiologicalLaboratory. The body wall was excised and branches of the respiratory treestied off to prevent collapse of the distended tubules. Short segments, suitablyligated, were fixed in Palade's osmium tetroxide mixture containing 2%[Quart. J. micr. Sci., Vol. 105, pt. 1, pp. 7-11, 1964.]

8 Doyle and McNiell—Respiratory tree in Cucumaria
additional sodium chloride. This was followed by secondary fixation in 10%formalin in the same buffer and salt mixture. After dehydration in alcoholthe material was embedded in epoxy resin (Shell-epon 812). For light micro-scopy the fixative was formalin or formalin with cetyl trimethyl ammoniumbromide added to preserve mucopolysaccharides.
ObservationsThe respiratory trees have the same basic structure as other portions of the
gut, with certain simplifications. When normally distended the tubules are3 to 4 mm in diameter and are colourless and transparent. When contractedthey are less than 1 mm in diameter and are opaque and light tan or yellowowing to the presence of lipid droplets. When distended the wall is 15 to20/x thick, consisting of 4 layers. There is a coelomic epithelium coveringa sparse mesh of smooth muscle-fibres, a thick (8 /JL) mucoid later, and a liningepithelium. In the contracted state the epithelial layers are thrown intovillus-like projections resembling transitional epithelia. In distended tubulesthe epithelia are not much thicker than the nuclei they contain and in thelight microscope appear to be a single layer of squamous cells. In the electronmiscroscope the cells are seen to possess long processes extending laterallyunder adjacent cells to give the appearance of a multi-layered epithelium (p,figs. 1, A; I , B; 4, B).
Scattered through the epithelia and the mucoid layer are amoebocytes of3 types. In light microscope studies Boolootian (1962) has recognized 14fairly distinct types of wandering cells in echinoderm body fluids. One ofthese is packed with large spherical granules which we observed to stainmetachromatically with azure A like the substance of the mucoid layer. Inthe electron miscroscope the individual spherules have dense centres withpale haloes after fixation with osmium tetroxide (fig. 2, A). When stained withphosphotungstic acid the haloes are intensely blackened. We have observedseveral instances of these amoebocytes disintegrating in the mucoid layer. Insuch cells the spherules show an increased halo and diminished central densearea.
Another type of amoebocyte is found to contain a variety of vacuoles withheterogeneous contents and is obviously a phagocyte (fig. 3, A). Portions ofa third type of cell found free in the mucoid layer are shown at / fig. 1, Band in fig. 3, B. These cells vary in the degree of development of the
FIG. 1 (plate). A, lining epithelium of distended tubule. Finger-like processes on the freesurface at upper right with large vacuoles, v, adjacent to Golgi lamellae, g. Processes (p) ofcells adjacent to the mucoid layer (lower right) containing fine collagenous filaments.
B, section of lining epithelium and mucoid layer including portion of a free cell, / , and theend of a smooth muscle fibre, m, containing mitochondria. Vacuoles, v, and cell processes,p, as in A.
FIG. 2 (plate). A, amoeboid cell with large spherules and compressed nucleus. The in-dividual spherules have dark centres and light haloes.
B. prominent endoplasmic reticulum with dense contents, c, in a cell of the lining epi-thelium.

m
Fie. i
W. L. DOYLE and G. F. MCNIELL

FIG. 2
W. L. DOYLE and G. F. MCNIELL

FIG. 3
W. L. DOYLE and G. F. MCNIELL
BFIG. 4
W. L. DOYLE and G. F. MCNIELL

Doyle and McNiell—Respiratory tree in Cucumaria 9
endoplasmic reticulum but all of them show a prominent Golgi apparatus,ribosomes, and coarser granules and more mitochondria than the other freecells. Their cytoplasmic features are similar to those found in fibroblasts inhigher forms. What may be developmental stages of these cells are seen in thelining epithelium (figs. 2, B; 6, B) described below.
The lining epithelium consists of a superficial layer of foamy cytoplasmfilled with vacuoles 1 -o to 1 • 5 fi in diameter (v, fig. 1, A, B) . In sections of con-tracted tubules this produces a lacy pattern. There are widely spaced finger-like processes extending about ifx from the free surface (figs. 1, A; 6, B).The large vacuoles appear to fuse with each other and they contain strandsof precipitated material (v, figs. 1, A; I , B). Deeper in the cells there are pro-minent Golgi membranes surrounded by small vacuoles {g, fig. 1, A). Beneaththe Golgi region is the nucleus and then the basal processes of the cells. Allthe cells of the lining epithelia are alike in their superficial (apical) portionsbut the basal portions show differences. The central and basal portions mayhave a prominent endoplasmic reticulum bordered with ribosomes. The con-tents of the endoplasmic reticulum exhibit varying degrees of electron density(figs. 2, B; 6, B), but the reticulum is homogeneous in a given cell. Mito-chondria are more numerous in the basal portions of the cells.
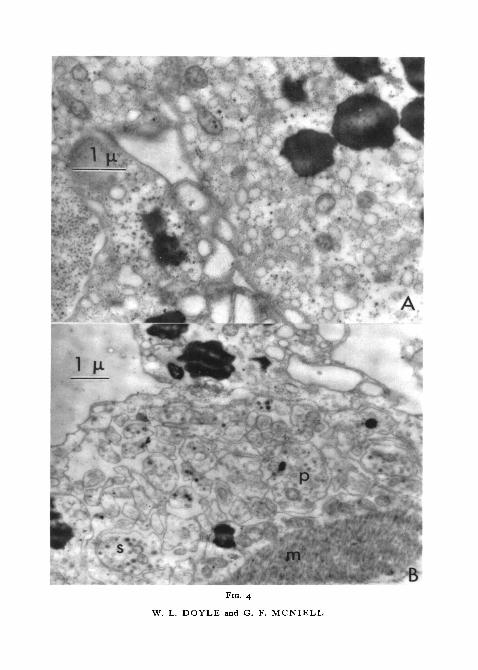
About half the thickness of the wall of the distended tubule of the respiratorytree consists of a mucoid layer which stains metachromatically with azure.In the electron microscope after staining with phosphotungstic acid numerousfine collagenous filaments are seen (figs. 1, A; I , B). The filaments are uni-formly about 0-02p. in diameter and at least 10/x long; they do not aggregateinto larger fibres and have not been seen more closely packed than o-i fj, apart.In some areas they are randomly oriented but in distended tubules they tendto be arranged circumferentially. The fibrils appear to offer no substantialresistance to the movement of amoebocytes in the mucoid layer. The coelomicepithelium contains prominent lipid droplets in the abundant superficialcytoplasm (fig. 4, A). Nuclei are centrally located and the basal portion ofthe cell consists of numerous long processes (p) containing membrane-bounded secretion granules (s, fig. 4, B). These long thin processes are inter-mixed with processes of adjacent cells and they extend to the level of thesmooth muscle-fibres. Mitochondria are scattered through the cytoplasm,which is filled with small vesicles and occasional areas of fine rough-surfacedendoplasmic reticulum (fig. 4, A). In the superficial region of coelomic epi-thelium, desmosomes are prominent and they are also occasionally seen in the
FIG. 3 (plate), A, part of the cytoplasm of a phagocytic amoeboid cell.B, portion of an amoeboid cell in the mucoid layer. Nucleus at bottom with Golgi lamellae
(g) just above. Ribosomes and coarse granules in the cytoplasm.FIG. 4 (plate). A, portion of the cytoplasm of a cell of the coelomic epithelium with typicalblackened large lipid droplets, small vacuoles, a few mitochondria, and coarse and finegranules.
B, partially contracted coelomic epithelium. Muscle, m, at lower right with a row of mito-chondria. Numerous cell processes, p, with typical components; in some there are densemembrane-bound granules, s.

io Doyle and McNiell—Respiratory tree in Cucumaria
basal portions between the fine cell processes. In contracted tubules thespaces between desmosomes may become spherical, giving rise, in sections,to a beaded appearance of the membranes bounding the intercellular space.
The smooth muscle-fibres run circularly and longitudinally with respectto the lumen and occur as groups forming strands rather than as definitelayers. Tangential sections show an open mesh pattern. Cross-sections ofthe strands often show 3 or 4 fibres in the same sarcoplasm. In longitudinalsections one may observe a circular fibre {cm, fig. 6, A) originating as a per-pendicular branch of a longitudinal fibre. Plasmodesms occur betweenadjacent parallel fibres and also between longitudinal and circular fibres, thuscross-linking the mesh-work (fig. 5, A). The fibres lie in an abundantsarcoplasm which is often scalloped in outline along the fibre (fig. 6, A).The fibres terminate in cytoplasmic processes which extend up into the regionof the epithelial cell processes. These processes are well supplied with mito-chondria and with characteristic vesicles (about o-i6/x in diameter) arrangedlinearly along the muscle-fibre. Within the fibrous portion of the muscle-cellsthere are regions of closely packed small granules. In appearance and inaggregation these granules are similar to those found immediately adjacentto or surrounding some of the large lipid droplets of the coelomic epithelium.The small granules apparently result from the breakdown of the large drop-lets and their occurrence among the muscle filaments suggests a nutritiverole (fig. 5, B).
DiscussionThe wall of the respiratory tree is found to consist of a substantial mucoid
layer containing fine collagenous filaments. It is bounded by a thin liningepithelium and a coelomic epithelium overlying an open mesh of circular andlongitudinal smooth muscle-fibres. The presence of numerous large vacuolesin the lining epithelium adjacent to the sea-water surface implies a consider-able transfer of substance across the free surface. Intercellular space is promi-nent between the lateral and basal portions of the cells but there are prominentplasmodesms along all the apical boundaries. The superficial levels of thecoelomic epithelium similarly show plasmodesms but the cytoplasm containsfewer vacuoles as large as 1 fx. In the basal regions of these cells, consistingof long thin processes, the intercellular space is prominent but occasionalplasmodesms are seen. Thus the cells of each of the free surfaces showprominent terminal bars with appreciable intercellular space at basal levels.The mucoid layer extends between the basal portions of such cell processes.
The amoebocytes with large spherical granules have been found in bothepithelia but more commonly in the coelomic layer, where developmental
FIG. S (plate), A, smooth muscle-fibres with prominent desmosomes between the upperfibres which are oriented in parallel and, at arrow, between fibres oriented perpendicularlyto each other.
B, disintegrating lipid droplet, d, in smooth muscle, and aggregates of coarse granules sur-rounding the droplet and among the fibrils.

FIG. S
W. L. DOYLE and G. F. MCNIELL

FIG. 6
W. L. DOYLE and G. F. MCNIELL

Doyle and McNiell—Respiratory tree in Cucumaria 11
stages are seen. When fully packed with spherules so as to distort the nucleusthey are frequently found in the mucoid layer and are observed with dis-continuous cell membranes. In this stage the spheres have a diminished cen-tral dark region and their periphery blends indistinguishably with thesubstance of the mucoid layer. It seems probable that this kind of amoebocyteforms the mucous substance of the mucoid layer. The origin of the fibrouselements of the mucoid layer is less evident. There are some cells of the liningepithelium which develop a prominent endoplasmic reticulum so that togetherwith other cytoplasmic constituents they resemble a free cell found in themucoid layer. These free cells have many of the features associated withfibroblasts in higher forms and it is possible that they may play a similar rolein this organism.
The presence of desmosomes at points of apposition between perpendicu-larly oriented fibres of the smooth muscle mesh is an interesting structuralfeature which seems to ensure synchronous contraction of the circular andlongitudinal elements. They may also be of significance in preventing localizedweakness of the delicate membranous wall of the respiratory tubule.
This work was supported by grants G-21016 from the National ScienceFoundation and RG 5627 from the United States Public Health Service.
ReferencesBoolootian, R. A., 1962. Amer. Zool., 2, 275.Hyman, L., 1955. The invertebrates, Vol. 5, Echinodermata. New York (McGraw-Hill).
FIG. 6 (plate). A, tangential section of tubule of respiratory tree at level of muscle-fibres.A single circular fibre, cm, is seen branching from a fibre oriented longitudinally.
B, lining epithelium with finger-like cell process at upper right. At lower left, part of a cellwith prominent endoplasmic reticulum of low density, c.