Embed Size (px)
Citation preview

Abstract. The balance between inhibition and excitationplays a crucial role in the generation of synchronousbursting activity in neuronal circuits. In human andanimal models of epilepsy, changes in both excitatoryand inhibitory synaptic inputs are known to occur.Locations and distribution of these excitatory andinhibitory synaptic inputs on pyramidal cells play a rolein the integrative properties of neuronal activity, e.g.,epileptiform activity. Thus the location and distributionof the inputs onto pyramidal cells are importantparameters that influence neuronal activity in epilepsy.However, the location and distribution of inhibitorysynapses converging onto pyramidal cells have not beenfully studied. The objectives of this study are toinvestigate the roles of the relative location of inhibitorysynapses on the dendritic tree and soma in the gener-ation of bursting activity. We investigate influences ofsomatic and dendritic inhibition on bursting activitypatterns in several paradigms of potential connectionsusing a simplified multicompartmental model. We alsoinvestigate the effects of distribution of fast and slowcomponents of GABAergic inhibition in pyramidal cells.Interspike interval (ISI) analysis is used for examinationof bursting patterns. Simulations show that the inhib-itory interneuron regulates neuronal bursting activity.Bursting behavior patterns depend on the synapticweight and delay of the inhibitory connection as wellas the location of the synapse. When the inhibitoryinterneuron synapses on the pyramidal neuron, inhibi-tory action is stronger if the inhibitory synapse is closeto the soma. Alterations of synaptic weight of theinterneuron can be compensatory for changes in thelocation of synaptic input. The relative changes in theseparameters exert a considerable influence on whethersynchronous bursting activity is facilitated or reduced.Additional simulations show that the slow GABAergicinhibitory component is more effective than the fastcomponent in distal dendrites. Taken together, these
findings illustrate the potential for GABAergic inhibi-tion in the soma and dendritic tree to play an importantmodulatory role in bursting activity patterns.
1 Introduction
Synchronized firing of neural networks characterizesepileptic seizures in human and experimental models.Sanabria et al. (2001) demonstrated that the develop-ment of temporal lobe epilepsy (TLE) is related tointrinsic bursting in hippocampal CA1 pyramidal cells inthe pilocarpine-treated rat model. They suggested thatthis intrinsic bursting may be responsible for theinitiation of epileptiform events that synchronize allCA1 pyramidal cells. Models of synchronous bursting inneural networks have been described using simple single-compartment neurons assembled in ensembles (Kudelaet al. 1997, 1999a,b, 2000). These networks withsimplified neurons can produce a range of burstingactivity produced by either stimulation or randomexcitatory input. Spread of activity in these networks isat a speed consistent with known biological models. Inthese simplified models, synaptic weight and delay arethe two sole variable parameters of connectivity. Tofurther validate the appropriateness of the simplifiedmodel, we recently studied a simplified multicompart-ment model of three pyramidal neurons, which allowsfor additional variable sites of synaptic input. Using thismodel, synchronized bursting activity can be producedby modifications of synaptic connectivity as well assynaptic weight and delay.
Av-Ron (1994) and Av-Ron et al. (1993) suggest thatchanges in certain parameters influence neuronal firingpatterns. They suggested that neuronal firing patternscan be described by changing a limited set of modelparameters in a bursting neuron model. In the simplifiedsingle-compartment model (Kudela et al. 1997, 1999a,b,2000), the increased excitatory synaptic weight, repre-senting a composite of all synaptic inputs, increases
Correspondence to: K. H. Yang(e-mail: [email protected],Tel.: +1-410-5028059, Fax: +1-410-9550751)
Biol. Cybern. 89, 242–253 (2003)DOI 10.1007/s00422-003-0429-3� Springer-Verlag 2003
The influences of somatic and dendritic inhibition on burstingpatterns in a neuronal circuit model
Keun-Hang Yang, Piotr J. Franaszczuk, Gregory K. Bergey
Department of Neurology, Johns Hopkins Epilepsy Center, Johns Hopkins University School of Medicine,600 North Wolfe Street, Meyer 2-147, Baltimore, MD 21287, USA
Received: 2 January 2003 /Accepted in revised form: 4 June 2003 / Published online: 12 August 2003

synchronized bursting activity. Changes in both excit-atory and inhibitory synaptic organization occur in hu-man and experimental models of epilepsy (Bernard et al.2000). An imbalance between excitatory and inhibitorysynapses is thought to be important for the generation ofbursting activity in pyramidal neurons (Lamsa et al.2000). Lamsa et al. (2000) examined the relative contri-butions of AMPA and GABAA receptors in networkactivity of CA3-CA1 pyramidal cells in the newborn rathippocampus slices. They suggested that AMPA syn-apses play an important role in the generation of spon-taneous bursting activity in pyramidal cells. Velazquezand Carlen (1999) demonstrated that spontaneousbursting activity could be produced by excitatory inputsand GABAergic interactions among interneurons.
GABAergic inhibition influences neuronal excitabil-ity; changes in GABAergic inhibition are important in anumber of models of experimental epilepsy (Prince 1978;Freund and Buzsaki 1996; Bernard et al. 2000). Indeedthe changes in GABAergic inhibition produced by sei-zures can potentially involve receptor number, location,and function (Bernard et al. 2000; Coulter 2001) andhave yet to be fully elucidated. Megias et al. (2001)demonstrated that the properties of pyramidal neuronaloutput strongly depend on the number, proportion, anddistribution of excitatory and inhibitory inputs. Theyobserved that the proportion of inhibitory inputs ondistal dendrites is low (� 3 %), whereas inputs onproximal dendrites is high (70% to 100%) in hippo-campal CA1 pyramidal cells. In one recent study ofexperimental TLE, somatic GABAergic inhibition isincreased, while dendritic GABAergic inhibition is de-creased (Cossart et al. 2001). Such an increase of somaticinhibition could potentially act to reduce epileptiformactivity.
GABA is the main inhibitory transmitter in the CNS,where it acts predominantly at one of two types ofGABA receptors: GABAA receptors and GABAB
receptors. GABAA receptors are ligand-gated receptorsthat hyperpolarize the neuron by increasing inwardchloride conductance and have a fast inhibitory effect.GABAB receptors are G protein-linked receptors thathyperpolarize the neuron by increasing potassium con-ductance. The GABAB receptors decrease calcium entryand have a slow inhibitory effect.
Different locations and distribution of inhibitory in-puts via these GABAA and GABAB receptors maycontrol the properties of neurons (Megias et al. 2001).Although the location and distribution of inputs on thepyramidal cell are important parameters influencingcellular activity, these parameters on pyramidal cellshave not been fully studied. Kapur et al. (1997) sug-gested that GABAA-mediated inhibition has both fastand slow components with different locations on pyra-midal cells in piriform cortex. They demonstrated thatboth fast and slow GABAA components are in bothsoma and dendrites, but a slow component is morecommonly found in dendrites of pyramidal neurons.They also demonstrated that the slow component isregulated by presynaptic GABAB inhibition, whereas thefast component is not. Other studies have demonstrated
that the slow GABAB-mediated component is also in thedendrites of pyramidal cells in piriform cortex (Tsengand Haberly 1988; Kanter et al. 1996). It is important tostudy both the location and the distribution of theinhibitory inputs in order to investigate the properties ofneurons. It has been demonstrated that the slowGABAB-mediated component may be found morecommonly in distal dendrites than the fast GABAA
component.GABAB-mediated inhibition is involved in the mod-
ulation of focal seizures and epileptogenesis (Scanzianiet al. 1991, 1994; Haas et al. 1996; McLean et al. 1996;Velıskova 1996; Sutor and Luhmann 1998; Scanziani2000). Scanziani (2000) suggested that GABAergicinhibition activates postsynaptic GABAB receptors inhippocampus, which results in regulating hippocampalnetwork activity. He showed that postsynaptic GABAB
receptors on pyramidal cells play a role in regulatingtheir oscillation frequency. Scanziani et al. (1994) dem-onstrated a role of GABAB receptors in the generationof bursting activity in hippocampal slice cultures of therat. They showed that a GABAB receptor antagonistelicited the appearance of spontaneous ictal-like dis-charges. Scanziani et al. (1991) suggested that an in-crease of GABAB-receptor-mediated synaptic potentialsmay be important in preventing epileptogenesis. Sutorand Luhmann (1998) suggested that a postsynapticblockade of GABAB receptors induces an increase ofbursting activity in neocortical slices disinhibited byGABAA receptor antagonists.
Interspike interval (ISI) analysis has been used forexamination of bursting patterns (Armstrong-James andFox 1983; Sharp and Green 1994). Armstrong-Jamesand Fox (1983) suggested that ISI analysis could showclear bursting patterns by presenting very short ISIswithin the burst and long interburst intervals. Theyanalyzed bursting patterns in terms of the duration ofbursts and interburst periods. Sharp and Green (1994)demonstrated that examination of the ISIs could classifybursting patterns in single hippocampal cells into burst,nonburst, and depolarized burst patterns. They sug-gested that the burst has ISIs of 2–4 ms and the non-burst has no firing in the 2–4 ms ISIs but showed a laterpeak between 7 and 25 ms instead. The depolarizedburst has an early and a late peak in the ISI histogram,and the early peak is between 1 and 2 ms, while the laterpeak is at 20–40 ms.
The aim of the computational studies reported here isto extend the previous studies (Yang et al. 2002) byinvestigating the influence of changes in the connectivityof inhibition on the patterns of burst activity in modelpyramidal neurons, specifically incorporating two typesof inhibitory interneurons with inputs at varying loca-tions. The range of activity includes nonbursting activ-ity, repetitive bursting, and sustained depolarization.The median and standard deviation values of an ISI areused to classify the patterns of bursting behavior. Sim-ulations were used to find the synaptic weight and delaythat could generate repetitive bursting behavior in amodel of simplified multicompartmental pyramidalneurons incorporating an inhibitory interneuron.
243

Furthermore, we investigate the influences of the locationof inhibitory interneuronal synapses on bursting activityin a simplified pyramidal model. We also investigate theeffect of fast and slow GABAergic inhibition on burstingactivity in a model of simplified multicompartmentalpyramidal neurons. The insights gained from these mul-ticompartmental neuronal networks can provide impor-tant guides for models of large neural networks usingeven more simplified individual neurons.
2 Materials and methods
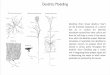
We have built a simplified pyramidal model using thesimulation software GENESIS (http://www.genesis-sim.org/GENESIS/). Three simplified pyramidal neu-rons and an interneuron are modeled in this study: twoneurons synaptically connected with excitatory synapsesforming a loop (neuron 1 and neuron 2 in Fig. 1), aneuron in which random input is applied to generateaction potentials (neuron 0 in Fig. 1), and an inhibitoryinterneuron (neuron 3 in Fig. 1) in a negative feedbackloop with one of the modeled pyramidal neurons(neuron 2 in Fig. 1). The generated action potentialsstimulate neuron 1 in Fig. 1, which is connected toneuron 2 in Fig. 1 by an excitatory synapse as arecurrent loop. Synaptic inputs are connected on themain dendrite of two neurons by an excitatory synapse(Fig. 1a), which is consistent with findings by Megiaset al. (2001) that excitatory inputs in particular are onthe dendrite spines in hippocampal CA1 pyramidal cells.An inhibitory interneuron (neuron 3 in Fig. 1) in anegative feedback loop with one of the two pyramidalneurons (neuron 2 in Fig. 1) is added. This inhibitoryinterneuron then synapses on the soma, main dendritete,or branch dendrites of the modeled pyramidal neurons(Fig. 1b–d). The main dendrite has four compartments,main dendrite location 1–4. Each branch dendrite has sixcompartments, branch dendrite location 110–115 or120–125. The main dendrite 1 compartment is closer tothe soma than main dendrite 4. The branch dendrite 111
or 121 compartment is closer to the main dendrite thanbranch dendrite 115 or 125.
Each cell is comprised of a soma, a main dendrite,and two branch dendrites, with 17 compartments. Thesoma has a fast sodium (INaÞ, delayed potassium (IKDRÞ,transient potassium (IAÞ, high-threshold calcium (ICaÞ,slow calcium-activated potassium (IAHPÞ, and short-duration voltage and calcium-dependent potassium(IKCÞ channel. The equations for these channels are thesame as in the Traub et al. (1991) multicompartmentalCA3 pyramidal cell model. The parameters for synapticconductance and cellular dimensions of the soma anddendrites are the same as in a simplified model of piri-form cortex pyramidal cells (Protopapas et al. 1998). Inthis model, the soma and main and branch dendriteshave excitatory and inhibitory synaptic channels thatconnect two neurons as a loop. The soma and dendriteshave a fast GABAergic inhibitory synapse and a slowGABAergic inhibitory synapse, which has much slowerkinetics than a fast GABAergic component. The kineticsof these fast and slow GABAergic components arethe same as for GABAA inhibitory and GABAB inhib-itory synapses, respectively, in a model of piriformcortex pyramidal cells (Protopapas et al. 1998). Theinhibitory interneuron (neuron 3 in Fig. 1) has a somawith the same channels as in the soma of two neuronssynaptically connected by excitatory synapses. Thesynaptic connection between neurons is modeled by asynaptic channel, Isyn (Bhalla and Bower 1993). Thesynaptic conductance is modeled as an alpha functionwith a maximum value of 0.2 ns for the excitatorysynapse, 0.5 ns for the slow GABAergic inhibitorysynapse, and 0.05 ns for the fast GABAergic inhibitorysynapse. The synaptic weight represents the overallstrength of a connection, and the synaptic delay repre-sents all delays between neurons. Simulations wereperformed for 10 s using GENESIS version 2.2 on aLINUX operating system. The simulation time step was0.05 ms, which is the same value as in the Traub et al.(1991) multicompartmental CA3 pyramidal cell modelprovided with GENESIS.
a
b c d
Fig. 1. Schematic representationof neural connections in themodel. a The connection betweentwo neurons connected by anexcitatory synapse has synapticinputs on the main dendrite oftwo neurons. In b–d, an inhibi-tory interneuron in a negativefeedback loop with one of themodeled pyramidal neurons isadded to the excitatory circuitshown in a. b An inhibitoryinterneuron synapses on the somaof neuron 2. c An inhibitoryinterneuron synapses on the maindendrite of neuron 2. d Aninhibitory interneuron synapseson the branch dendrite of neu-ron 2
244

In additional simulations (results not shown), wemodeled active dendrites by including the same channelsas in the soma with channel densities based on the areaof dendritic compartments. The diameter of main den-dritic compartments is 5:78lm, while for the branchdendritic compartments it is 4:84lm. These diametersare the same as apical and basilar compartmentparameters, respectively, in Traub et al. (1991). In thisstudy, the channel density is the maximum conductanceper unit area of a compartment. Therefore, the maindendritic compartments have slighly larger channeldensities than the branch dendritic compartments. Eachcompartment in the main dendrite has the same channeldensities. Each compartment in the branch dendritesalso has the same channel densities, which are slightlyless than in the main dendritic compartments. Therewere no significant differences in the results betweenmodels with active and passive dendrites. Since thecomputational time is significantly larger in a modelwith active dendrites, we show here only simulationsperformed with the passive dendrites.
The robustness of a simplified multicompartmentalmodel of pyramidal cells is examined. First, the sensi-tivity to parameters is tested. The synaptic delay of thepyramidal neuron is increased to large values in order toexamine the sensitivity of the synaptic delay to the pat-terns of bursting activity. Second, the connectivity be-tween the pyramidal neuron and inhibitory interneuronis changed in Fig. 1. The inhibitory interneuron syn-apses on the pyramidal neuron 1 instead of neuron 2, asin Fig. 1. Additional simulations are run with the largesynaptic weight or different connectivity between thepyramidal neuron and inhibitory interneuron to exam-ine the robustness of the present model on burstingactivity patterns.
It was necessary to define a burst for the analysis ofbursting activity patterns. Sharp and Green (1994) usedthe interspike interval (ISI) analysis to classify thebursting activity patterns in single hippocampal cellsinto burst, nonburst, and depolarized burst. They de-fined a burst using an early and a late peak value of ISIsin the ISI histogram. The present study used ISI medianor standard deviation values for examining burstingpatterns instead. The number of spikes in bursts for allpatterns of activity was used to determine thresholddetection. The spike counts and timing were then used todetermine ISI median and standard deviation valuesusing a program written in MATLAB.
3 Results
In our previous studies, we showed that bursting activitypatterns are dependent upon the synaptic weight anddelay for several locations of excitatory synaptic con-nections in a simplified pyramidal neuron model (Yanget al. 2002). In the present study, synaptic inputs areconnected on the main dendrite of two neurons by anexcitatory synapse (Fig. 1a), which is consistent with asuggestion by Megias et al. (2001) that the excitatoryinputs terminate exclusively on dendrites. We investigate
the influences of the location of inhibitory synapses onbursting activity in this excitable synaptic connection.Various random inputs are applied to the pyramidalneuron 0 in Fig. 1 in order to evaluate the influences ofthe synaptic weight and delay on the bursting activitypatterns shown in the previous study. Figure 2 showsthat repetitive burst activity depends on synapticparameters represented as the synaptic weight and delayin the excitatory synaptic connections between neuronsas shown in Yang et al. (2002).
The threshold ISI median and standard deviationvalues are determined based on results of simulationsused for classification of bursting activity patterns.Figure 3 shows that the median ISI values in neuron 1are constant at 185 ms for synaptic weights greater than700 and synaptic delays between 1 and 9 ms. We choosethe median ISI value of 185 ms as the lowest value forthe definition of nonbursting activity in neuron 1 (mid-dle panel in Fig. 5). The median ISI value of 52.8 ms inthese simulations for the synaptic weight and delay, 900and 5 ms, respectively, clearly represents burstingactivity. The choice of the threshold median values in therange of 53–185 ms results in the same classification ofbursting activity patterns because there are no medianISI values in this range for our simulations. However,the median ISI values are not sufficient to distinguishbetween bursting activity and the state of prolongeddepolarization; thus we use the standard deviation ofISIs to classify prolonged depolarization.
Fig. 2. Bursting behavior patterns in neurons 1 and 2 when thesynaptic inputs are on the main dendrite of the two neurons as shownin Fig. 1a. Bursting activity occurs at a range of 1–9 ms of thesynaptic delay and > 700 of the synaptic weight (closed circles). Nobursting activity occurs at a value of synaptic weight 700 for anyvalues of the synaptic delay (open triangles). Prolonged depolarizationoccurs at synaptic weight values of > 1300 and synaptic delay valuesof > 3 ms (open squares)
245

Figure 4 shows that the ISI standard deviation valuesstabilize for values of the synaptic weights larger than1300 and synaptic delays greater than 3 ms. The trace ofmembrane potentials in neuron 1 (middle panel inFig. 7) for the synaptic weight and delay, 2600 and 2 ms,respectively, with the ISI standard deviation value of20.5 ms clearly represents prolonged depolarization. Ina range of ISI standard deviation values above 30 ms,the next highest value of ISI standard deviation obtainedfrom simulations is 39.6 ms for the synaptic weight of1600 and delay of 2 ms, which clearly represents bursting
activity (data not shown). We choose an ISI standarddeviation value of 30 ms as the lowest value for our defi-nition of bursting activity. Thus we classify our simula-tions, which result in ISI median values of � 185 ms asnonbursting activity, ISI standard deviation values of< 30 ms as prolonged depolarization, and all others asbursting activity. There is nonbursting activity andbursting activity but no prolonged depolarization whenthe synaptic delay is 1 ms. When the synaptic delay is2 ms, prolonged depolarization occurs for the synapticweights 2000, 2400, and 2600.
Figure 3 shows the ISI median values for the synapticdelays between 1 and 9 ms shown in Fig. 2. There is nobursting activity (open triangles in Fig. 2) for thesevalues of the synaptic delay that result in ISI medianvalues of � 185 ms. Figure 4 shows the standard devi-ation values of ISIs when the synaptic delays are be-tween 1 and 9 ms, as shown in Fig. 2. The ISI standarddeviation values almost reach steady state at values lessthan 30 ms when the synaptic delay is greater than 3 ms.The bursting behavior becomes prolonged depolariza-tion (open squares in Fig. 2) for values of synaptic de-lays of > 1 ms that result in ISI standard deviationvalues of less than 30 ms. There is no bursting activity(open triangles in Fig. 2) for values of synaptic delays of� 1 ms that result in constant ISI standard deviationvalues of 270.5 ms.
Figures 5–7 show traces of membrane potentials forall neurons with a synaptic delay value of 2 ms andsynaptic weight values 700, 1200, or 2600 (Fig. 2) whenthe synaptic inputs are on the main dendrite of the twoneurons (Fig. 1a). There is no bursting activity in neu-rons 1 and 2 when the synaptic weight is 700 and thesynaptic delay is 2 ms (Fig. 5). When the synapticweight is increased to 1200 with the same delay of 2 ms,bursting activity occurs in neurons 1 and 2 (Fig. 6).When the synaptic weight is increased to 2600 with thesame delay of 2 ms, prolonged depolarization occurs inneurons 1 and 2 (Fig. 7).
Simulations show that the inhibitory interneuron canregulate neuronal bursting activity. The results of sim-ulations with active and passive dendrites were not sig-nificantly different with respect to the influence of theinhibitory interneuron. A model with passive dendrites,however, produced more robust bursting activityappropriate for modeling of epileptiform activity. Thussimulations in this study were run using a model withpassive dendrites. When the excitatory synaptic inputsare on the main dendrites of two connected neurons(Fig. 1a), the effect of a slow component of GABAergicinhibition is stronger if the inhibitory synapse is close tothe soma for a given synaptic weight (Fig. 8). The valuesof synaptic weight 1200 and synaptic delay 2 ms of theexcitatory synapse, which generate bursting activity withno inhibitory interneuron (Fig. 6), are used to comparebursting activity with and without an inhibitory inter-neuron. Bursting activity is blocked for values of thesynaptic weight of the inhibitory connection greater than100 for the soma (open triangles for the soma in Fig. 8)if a slow component of the inhibitory interneuron syn-apses on the soma (Fig. 1b). When a slow component of
500 1000 1500 2000 2500 30000
40
80
120
160
200
Synaptic Weight of the Pyramidal Neuron
Med
ian
of
the
Inte
rsp
ike
Inte
rval
(m
sec)
1 msec2 msec3 msec5 msec7 msec9 msec
Synaptic Delay
No Bursting Activity
Fig. 3. The median values of interspike intervals (ISIs) when thesynaptic delays are between 1 and 9 ms, as in Fig. 2
Fig. 4. Standard deviation values of ISIs when the synaptic delays arebetween 1 and 9 ms, as shown in Fig. 2. The domain betweennonbursting activity and prolonged depolarization represents burstingactivity
246

the inhibitory interneuron synapses on the main dendrite1 (Fig. 1c), there is no bursting activity for values of thesynaptic weight of the inhibitory connection larger than500 (open triangles for the main dendrite 1 in Fig. 8). Inorder to produce the same inhibitory effect on thebranch dendrite, the synaptic weight of an interneuronneeds to be increased (open triangles for the branchdendrites 111 and 114 in Fig. 8). Therefore, the inhibi-tory effect is stronger when the slow component of theinhibitory synapse is close to the soma.
Additional simulations were run to compare theinhibitory effect of the slow component with the fastcomponent of GABAergic inhibition. Simulations withthe fast component of GABAergic inhibitory interneu-ron also show that the inhibition is stronger when thefast GABAergic inhibitory interneuron synapses on thesoma rather than on the main dendrite, and the sameholds true for the slow component (for the soma andmain dendrites 1 and 4 in Fig 8). However, there is nosignificant change in bursting activity patterns when the
Fig. 5. Traces of membranepotentials for all neurons whenexcitatory synaptic connectionsare between the main dendrite ofneurons 1 and 2, as shown inFig. 1a. In this example, whenthe synaptic weight and synapticdelay are 700 and 2 ms, respec-tively, bursting activity does notoccur in neurons 1 and 2 (the ISImedian and standard deviationvalues for neuron 1 are 185 msand 270.5 ms, respectively)
100
50
0
50
Vm
(m
V)
100
50
0
50
Vm
(m
V)
0 1 2 3 4 5 6 7 8 9 10 100
50
0
50
Time (sec)
Vm
(m
V)
Generated Action Potentials in Neuron 0 from Random Inputs
Bursting Activity in Neuron 1
Bursting Activity in Neuron 2
Fig. 6. Traces of membranepotentials for all neurons whenexcitatory synaptic connectionsare between the main dendrite ofneurons 1 and 2, as shown inFig. 1a. When the synapticweight and synaptic delay are1200 and 2 ms, respectively,bursting activity occurs in neu-rons 1 and 2 (the ISI medianvalue for neurons 1 and 2 is16.3 ms)
247

fast GABAergic inhibitory interneuron with large syn-aptic weights synapses on the branch dendrite of apyramidal neuron (data not shown).
Figure 9 shows the evaluation of bursting activitypatterns, including nonbursting and bursting activity,using the median values of interspike intervals (ISIs) fordifferent locations and weights of the inhibitory synapticconnections from Fig. 8. For locations and values of thesynaptic weight of the inhibitory synaptic connectionsthat result in median values of ISIs of � 185 ms (thesame values as in Fig. 4), there is no bursting behaviorpattern.
Fig. 7. Traces of membranepotentials for all neurons whenexcitatory synaptic connectionsare between the main dendrite ofneurons 1 and 2, as shown inFig. 1a. Here, when the synapticweight and synaptic delay are2600 and 2 ms, respectively,prolonged depolarization occursin neurons 1 and 2 (the ISI stan-dard deviation for neurons 1and 2 is 20.5 ms)
Fig. 8. Influence of the location and weight of a slow component ofGABAergic inhibitory synaptic connection on patterns of burstingactivity. The synaptic delay of the slow component of GABAergicinhibitory synapse is 1 ms. The synaptic weight and delay for theexcitatory synapse are 1200 and 2 ms, respectively, the same as inFig. 6. The inhibitory effect is stronger when the inhibitory synapse isclose to the soma. When the inhibitory synapse is on the branchdendrite, the synaptic weight has to be increased to produce the sameeffect. The main dendrite 1 compartment is closer to the soma thanmain dendrite 4. The branch dendrite 111 compartment is closer thanbranch dendrite 114 to the main dendrite
Fig. 9. The median values of interspike intervals (ISIs) for a differentlocation and weight of the inhibitory synaptic connections fromFig. 8. There is no bursting behavior pattern for locations and valuesof the synaptic weight of the inhibitory synaptic connections thatresults in an ISI median value of � 185 ms, which is the same value asin Fig. 3
248

Figures 10 and 11 show traces of membrane poten-tials for neurons 0 and 2 with the inhibitory connectionwhen the excitatory synaptic inputs are on the maindendrite of the two neurons. The excitatory synapticweight and delay are 1200 and 2 ms, respectively. Whena slow component of inhibitory interneuron is in anegative feedback loop with one of the connectedexcitatory neurons that synapses on the main dendrite 1of neuron 2 (Fig. 8), there is no bursting activity inneuron 2 when the synaptic weight and delay of theinhibition are 2000 and 1 ms, respectively (Fig. 10).When an inhibitory interneuron synapse is on thebranch dendrite 111 of neuron 2 (Fig. 8) with the samesynaptic weight of 2000 and the same delay of 1 ms of
the inhibitory connection, however, there is still burstingactivity in neuron 2 (Fig. 11).
4 Discussion
The present pyramidal cell model with 17 compartmentsincludes both soma and dendrites. This allows modelingof the influences of location of synaptic input andfacilitates studies of interneuronal interactions, which inturn permits modeling of changes in connectivity thatmay accompany pathological conditions such as epi-lepsy and how these changes may affect synchronousburst activity. It is now recognized that the pathology of
Fig. 10. Traces of membrane potentials for neurons 0 and 2 whenexcitatory synaptic connections are between the main dendrite ofneurons 1 and 2, as shown in Fig. 1a. The excitatory synaptic weightand delay are 1200 and 2 ms, respectively, the same as in Fig. 6. Whenthe slow GABAergic component of the inhibitory synapse is on the
main dendrite 1 of neuron 2 (Fig. 1c and Fig. 8), there is no burstingactivity in neuron 2 if the synaptic weight and delay of the inhibitorysynapse are 2000 and 1 ms, respectively (the ISI median value forneuron 2 is 1052.4 ms)
Fig. 11. Traces of membrane potentials for neuron 0 and neuron 2when excitatory synaptic connections are between the main dendriteof neuron 1 and neuron 2 as shown in Fig. 1a. The excitatory synapticweight and delay are 1200 and 2 ms, respectively, the same as inFig. 6. When the slow GABAergic component of inhibitory synapse is
on branch dendrite 111 of neuron 2 (Fig. 1d and Fig. 8) with the samesynaptic weight and delay, 2000 and 1 ms, respectively, there isbursting activity in neuron 2 (the median value of ISIs for neuron 2 is17.8 ms)
249

epilepsy is often not fixed and that synaptic plasticityand reorganization occur in a dynamic fashion.
The balance between inhibition and excitation iscritical in the generation of bursting activity. Changes inthis balance can dramatically alter the behavior of bothindividual neurons and large networks. Cossart et al.(2000) investigated the ratio between excitatory andinhibitory synaptic currents along the somatodendriticaxis of rat hippocampal CA1 pyramidal cells. Theysuggested that more than 90% of the spontaneous cur-rents were GABAergic (inhibitory), whereas less than10% were glutamatergic (excitatory). This observed in-crease in spontaneous somatic GABAergic inhibitionwas postulated to result from hyperactivity of somaticinterneurons. Decreased dendritic GABAergic inhibi-tion was thought to result from the degeneration of asubpopulation of dendritic interneurons in this model ofexperimental TLE.
Our previous (Yang et al. 2002) and present studieshave demonstrated that the presence and patterns ofbursting activity depend heavily on three factors: syn-aptic weight, synaptic delay, and synaptic location. Thepresent model, now incorporating inhibitory interneu-rons, clearly illustrates that inhibition is stronger whenthe inhibitory synapse is located close to the soma. Thusthis model supports known properties of forwardpropagation and attenuation of the synaptic inputs withdistance in passive dendrites. Moreover, this finding isconsistent with an experimental study by Cossart et al.(2001) which showed that dendritic GABAergic inhibi-tion is decreased while somatic GABAergic inhibition isincreased in TLE. Preferential somatic location ofinhibitory inputs occurs in normal local neural networks(e.g., hippocampus, spinal cord), but, as discussedabove, pathological conditions such as epilepsy mayshift the balance of inhibition further to perisomaticlocations. Whether these changes act to reduce epilep-togenic activity is not known.
Simulations were run using the present model withpassive and active properties of dendrites. The activedendrites have the same channels as in the soma, and thechannel densities vary based on the area of dendriticcompartments. There is no difference between passiveand active dendrite properties in the effect of theinhibitory interneuron. In both passive and active den-drites, inhibitory synaptic inputs modeled at more dis-tant sites on the branch dendrites need increasedsynaptic weight to produce the same effect as somaticinputs. However, the passive dendrites have more effecton bursting activity in a model of pyramidal neurons;the duration of the bursts is much longer in the passivedendrites than active dendrites with the same synapticweight and delay, so that bursting activity can be clearlydistinguished from nonbursting activity in the passivedendrites. Thus our simulations do not require activedendritic properties to investigate the influences of theinhibitory interneuron on bursting activity patterns.
Interspike interval (ISI) analysis is used to definebursting activity patterns in the present study. Thethreshold ISI median and standard deviation values aredetermined based on results of simulations; the ISI
median value of 185 ms is the lowest value for the def-inition of nonbursting activity. The standard deviationof ISIs of 30 ms is the lowest value for the definition ofbursting activity. Thus we define simulations that resultin median values of ISIs of � 185 ms as nonburstingactivity, the standard deviation values of ISIs of< 30 ms as prolonged depolarization, and all others asbursting activity. Figures 3, 4, and 9 show that this cri-terion for the classification of bursting activity patternswell support our simulation results.
Our simulations show that slow GABAergic inhibi-tion plays a role in epileptiform activity as shown byGABAB inhibition in several laboratories (Scanzianiet al. 1994; Haas et al. 1996; McLean 1996; Velıskovaet al. 1996; Sutor and Luhmann 1998). The inhibitoryeffect of a slow component of GABAergic inhibition isstronger when the inhibitory interneuron synapses closeto the soma rather than to dendrites (Fig. 8). Theinhibitory effect of a fast component of GABAergicinhibition is also stronger when the inhibitory inter-neuron synapses on the soma rather than on the maindendrite. However, there is no significant change inbursting activity patterns when the fast inhibitoryinterneuron synapses with large synaptic inputs on thebranch dendrite of the pyramidal neurons; when the fastinhibitory interneuron synapses on the branch dendriteof the pyramidal neurons (branch dendrite location 111or 114), there is no inhibition effect for large synapticweights, > 105, of the inhibitory interneuron (data notshown). Kapur et al. (1997) suggested that GABAA-mediated inhibitory postsynaptic currents (IPSCs) aregenerated in apical dendrites and cell bodies in piriformcortex. The generated IPSCs in the neuronal circuit arelargely separated in distal dendrites and somatic regions;GABAA inhibition consists of fast and slow compo-nents, and there is a greater proportion of the slowcomponent in distal dendrites than cell bodies. Theydemonstrated that the slow component of GABAA-mediated IPSCs is regulated by presynaptic GABAB-mediated inhibition but that the fast one is not. Otherstudies in piriform cortex (Kanter et al. 1996; Tseng andHaberly 1988) demonstrated that GABAB-mediatedIPSCs are generated in the dendrites of pyramidal cells.Our simulations with the fast GABAA-mediated andslow GABAB-mediated components also show that theslow component of GABAergic inhibition is strongerthan the fast component at distal dendritic locations.
Additional simulations with large synaptic delays or adifferent connectivity between pyramidal and inhibitoryneurons were run to test the robustness of the simplifiedmodel; the synaptic delay of the pyramidal neuron wasincreased by up to 20 ms; the inhibitory interneuronsynapses on neuron 1 instead of on neuron 2 in Fig. 1.Simulations with large synaptic delays (> 10 ms) of thepyramidal neuron show that bursting activity patternsare insensitive to a parameter, the synaptic delay, in amodel of pyramidal cells. When the synaptic delays are> 10 ms, bursting activity patterns are the same as forthe synaptic delays, 5 ms, 7 ms, or 9 ms (data notshown). Simulations with a different connectivity be-tween pyramidal and inhibitory neurons show the same
250

behavior, i.e., that the inhibitory effect is stronger whenthe inhibitory interneuron synapses close to the somarather than to the dendrites of pyramidal neuron 1, asshown in Fig. 8 (data not shown). These tests help tobuild confidence that the model is valid for investigatingbursting activity patterns modulated by somatic anddendritic inhibition.
There are several similarities and differences betweenour model and other models of pyramidal cell orexperimental studies. Our model and that of Kager et al.(2000) incorporate neither the axon nor dendritic spinesin pyramidal cells. The rate constants that describe thevoltage-dependent channels are based on a model ofhippocampal pyramidal cells described by Traub et al.(1991) in both models of pyramidal cells. However, theKager et al. (2000) model has modified channel kineticsfrom Traub et al. (1991); the activation curve of the fasttransient sodium current (INaÞ was shifted 5 mV in adepolarizing direction based on their own observations(Vreugdenhil et al. 1998). The neuron in the Kager et al.(2000) model was surrounded by an interstitial spacewhere extracellular potassium and sodium concentrationwere incorporated. Our model includes neither anymodification of channel kinetics from Traub et al. (1991)nor extracellular potassium and sodium concentration.Kager et al. (2000) showed that three defects – inherentlyenhanced excitation, a synaptic inhibition defect, orfailure of ion concentration regulation – could causeepileptiform activity. Since they were interested in neu-ronal membrane changes underlying seizure dischargesand spreading depression, they needed to investigateextracellular and intracellular ion concentration changesin detail. Our model demonstrates the role of the exci-tation and inhibition, which is associated with syn-chronized neuronal bursting activity, but not ionconcentration on the epileptic seizures.
In their model of neocortical neurons, Mainen andSejnowski (1996) demonstrated that active dendritesmay play an important role in modifying the firingproperties of the neuron. They found that this effect ispresent only when electrical coupling between the somaand dendrite compartment is relatively weak. Changingthis coupling parameter affects various firing patterns.They suggested that this effect of different coupling andmorphology of the dendritic tree may be very importantfor the specialization of neocortical neurons in per-forming various functions in processing information inneocortical neurons. In this study, we investigate theinfluences of the inhibitory interneuron on epilepticbursting activity. Our model is based on hippocampalpyramidal cells with relatively strong coupling betweencompartments, which is capable of producing very ro-bust bursts. Our simulations with active dendrites werenot significantly different from results with passivedendrites, but they required much longer computationaltime. Some experimental studies have suggested thatobserved attenuation ratios are adequately explained bypassive membrane properties alone (Larkum et al. 1998;Pyapali et al. 1998). Ceci et al. (1999) suggested that thepassive membrane properties of pyramidal neuronsresemble regular spiking or intrinsically bursting activity
in the medial prefrontal cortex of the rat. Also, somerecent modeling studies have shown that active currentsmay not be required to explain observed integrationproperties of neurons (Jaffe and Carnevale 1999). Thus,it may not be necessary to include active properties ofdendrites to investigate the role of the inhibitory inter-neuron on epileptic bursting activity patterns.
In future studies, evaluation of channel kinetics fromTraub et al. (1991) as well as changes in intracellular andextracellular ion concentrations using experimental datawill be needed to study in more detail channel activity inpyramidal cells. Kapur et al. (1997) demonstrated thatGABAA-mediated IPSCs have fast and slow compo-nents in piriform cortex. The fast component has a de-cay time constant of 10 ms and a slow component of40 ms. Our model has fast GABAA-mediated and slowGABAB-mediated components instead. The fast com-ponent has a decay time constant of 7 ms and a slowcomponent of 100 ms, the same as in a model of piri-form cortex pyramidal cells (Protopapas et al. 1998).Future studies will require the evaluation of kinetics aswell as the locations and distribution of GABA-medi-ated receptors using experimental data of GABA inhi-bition in pyramidal cells.
Figure 4 shows that there is nonbursting activity andbursting activity but no prolonged depolarization whenthe synaptic delay is 1 ms. When the synaptic delay is2 ms, prolonged depolarization occurs for the synapticweights 2000, 2400, and 2600. Additional simulationsare run with different kinetics of the slow calcium-acti-vated potassium (IAHPÞ channels to investigate thedependence of the patterns of bursting activity on boththe synaptic delay and IAHP channel dynamics. When theactivation of forward rate constant for the slow calcium-activated potassium (IAHPÞ channels is increased from0.2 to 0.4 with the same synaptic delay of 2 ms, there isno prolonged depolarization for any synaptic weight(data not shown). This may be related to the IAHP
channels in pyramidal cells, as shown in Martin et al.(2001). Our simulations show that bursting activitypatterns may be related to both the IAHP channeldynamics and synaptic delay. Further studies are neededto evaluate interactions of the IAHP channel dynamicsand parameter connectivity in the generation of variousbursting activity patterns.
Recent observations in several laboratories have indi-cated that there may be compensatory mechanisms forthe synapse location of excitatory synapses. Excitatorypostsynaptic potentials (EPSPs) generated at more distalsites may have associated increased conductances thatmay counter the effects of the distal location (Magee andCook 2000; Williams and Stuart 2002). Indeed thesecompensatory conductance changes are thought bythese investigators to be more important than any activedendritic properties. In themodel used in our studies, suchlocation-dependent changes could be readily modeledwith changes in synaptic weights. Whether or not suchsite-dependent differences in conductance are operationalfor inhibitory inputs is not yet known.
In conclusion, our simplified multicompartmentalmodel of pyramidal cell neurons allows for the incor-
251

poration and modification of the important factorsinfluencing synaptic inputs and the generation of syn-chronous bursting activity. Particularly important andrelevant to epilepsy is the ability to incorporate inhibi-tory interneurons into the model and to study the neteffects of changes and reorganization of such inputs. Thestudies reported here with this model reveal the impor-tant balance, not only between excitatory and inhibitoryactivity, but between input location and synaptic weight.Inhibitory modulation of recurrent bursting behavior ismost effective with perisomatic inhibitory inputs; moredistal inputs require significantly greater synapticweights to produce the same effects on bursting behav-ior. Distal dendrites have more slow GABAergic syn-apses than fast GABAergic synapses. The observationsfrom these small networks can provide insight intoinfluences on synchronous bursting behavior in muchlarger networks.
Acknowledgements. This research is supported by NIH grantNS38958. A preliminary version of this work was presented atComputational Neuroscience and American Epilepsy Society 2001Annual Meetings.
References
Armstrong-James M, Fox K (1983) Effects of ionophoresed nor-adrenaline on the spontaneous activity of neurons in rat pri-mary somatosensory cortex. J Physiol 335: 427–447
Av-Ron E (1994) The role of a transient potassium current in abursting neuron model. J Math Biol 33: 71–87
Av-Ron E, Parnas H, Segel LA (1993) A basic biophysical modelfor bursting neurons. Biol Cybern 69: 87–95
Bernard C, Cossart R, Hirsch JC, Esclapez M, Ben-Ari Y (2000)What is GABAergic inhibition? How is it modified in epilepsy?Epilepsia 41 (Suppl 6): S90–S95
Bhalla US, Bower JM (1993) Exploring parameter space in detailedsingle neurons models: simulations of the mitral and granulecells of the olfactory bulb. J Neurophysiol 69: 1948–1965
Ceci A, Brambilla A, Duranti P, Grauert M, Grippa N, Borsini F(1999) Effect of antipsychotic drugs and selective dopaminergicantagonists on dopamine-induced facilitatory activity in pre-limbic cortical pyramidal neurons: an in vitro study. Neuro-science 93(1): 107–115
Cossart R, Hirsch JC, Cannon RC, Dinoncourt C, Wheal HV,Ben-Ari Y, Esclapez M, Bernard C (2000) Distribution ofspontaneous currents along the somato-dendritic axis of rathippocampal CA1 pyramidal neurons. Neuroscience 99: 593–603
Cossart R, Dinocourt C, Hirsch JC, Merchan-Perez A, De Felipe J,Ben-Ari Y, Esclapez M, Bernard C (2001) Dendritic but notsomatic GABAergic inhibition is decreased in experimentalepilepsy. Nat Neurosci 4: 52–62
Coulter DA (2001) Epilepsy-associated plasticity in gamma-amin-obutyric acid receptor expression, function, and inhibitorysynaptic properties. Int Rev Neurobiol 45: 237–252
Freund TF, Buzsaki G (1996) Interneurons of the hippocampus.Hippocampus 6: 347–470
Haas KZ, Sperber EF, Moshe SL, Stanton PK (1996) Kainicacid-induced seizures enhance dentate gyrus inhibition bydownregulation of GABAB receptors. J Neurosci 16: 4250–4260
Jaffe DB, Carnevale NT (1999) Passive normalization of synapticintegration influenced by dendritic architecture. J Neurophysiol82(6): 3286–85
Kager H, Wadman WJ, Somjen GG (2000) Simulated seizuresand spreading depression in a neuron model incorporating
interstitial space and ion concentrations. J Neurophysiol 84:495–512
Kanter ED, Kapur A, Haberly LB (1996) A dendritic GABAA-mediated IPSP regulates facilitation of NMDA-mediatedresponses to burst stimulation of afferent fibers in piriformcortex. J Neurosci 16(1): 307–312
Kapur A, Pearce RA, Lytton WW, Haberly LB (1997) GABAA-mediated IPSCs in piriform cortex have fast and slow compo-nents with different properties and locations on pyramidal cells.J Neurophysiol 78(5): 2531–2545
Kudela P, Franaszczuk PJ, Bergey GK (1997) A simple computermodel of excitable synaptically connected neurons. Biol Cybern77: 71–77
Kudela P, Franaszczuk PJ, Bergey GK (1999a) Model of thepropagation of synchronous firing in a reduced neuron net-work. Neurocomputing 26–27: 411–418
Kudela P, Franaszczuk PJ, Bergey GK (1999b) Decreasing inhi-bition increases synchronization and burst duration in simu-lated neural networks. Epilepsia 40(Suppl 7): 12
Kudela P, Franaszczuk PJ, Bergey GK (2000) Increase in numberof synaptic connections increases bursting activity in simulatedneural networks. Epilepsia 41(Suppl 7): 6
Lamsa K, Palva JM, Ruusuvuori E, Kaila K, Taira T (2000)Synaptic GABA(A) activation inhibits AMPA-kainate recep-tor-mediated bursting in the newborn (P0-P2) rat hippocam-pus. J Neurophysiol 83: 359–366
Larkum ME, Launey T, Dityatev A, Luscher HR (1998) Integra-tion of excitatory postsynaptic potentials in dendrites ofmotoneurons of rat spinal cord slice cultures. J Neurophysiol80(2): 924–935
Magee JC, Cook EP (2000) Somatic EPSP amplitude is indepen-dent of synapse location in hippocampal pyramidal neurons.Nat Neurosci 3(9): 895–903
Mainen ZF, Sejnowski TJ (1996) Influence of dendritic structure onfiring pattern in model neocortical neurons. Nature 382(6589):363–366
Martin ED, Araque A, Buno W (2001) Synaptic regulation of theslow Ca2+-activated K+ current in hippocampal CA1 pyra-midal neurons: implication in epileptogenesis. J Neurophysiol86(6): 2878–2886
McLean HA, Caillard O, Khazipov R, Ben-Ari Y, Gaiarsa JL(1996) Spontaneous release of GABA activates GABABreceptors and controls network activity in the neonatal rathippocampus. J Neurophysiol 76(2): 1036–1046
Megias M, Emri Z, Freund TF, Gulyas AI (2001) Total numberand distribution of inhibitory and excitatory synapses onhippocampal CA1 pyramidal cells. Neuroscience 102(3): 527–540
Prince DA (1978) Neurophysiology of epilepsy. Annu Rev Neu-rosci 1: 395–415
Protopapas AD, Vanier M, Bower JM (1998) Simulating LargeNetworks of Neurons. In: Koch C, Segev I (eds) Methods inneuronal modeling, 2nd edn. MIT Press, Cambridge, pp 461–498
Pyapali GK, Sik A, Penttonen M, Buzsaki G, Turner DA (1998)Dendritic properties of hippocampal CA1 pyramidal neuronsin the rat: intracellular staining in vivo and in vitro. J CompNeurol 391(3): 335–352
Sanabria ER, Su H, Yaari Y (2001) Initiation of network burstsby Ca2+ dependent intrinsic bursting in the rat pilocarpinemodel of temporal lobe epilepsy. J Physiol 532(Pt 1): 205–216
Scanziani M (2000) GABA spillover activates postsynaptic GA-BAB receptors to control rhythmic hippocampal activity.Neuron 25: 673–681
Scanziani M, Gahwiler BH, Thompson SM (1991) Paroxysmalinhibitory potentials mediated by GABAB receptors in par-tially disinhibited rat hippocampal slice cultures. J Physiol 444:375–396
Scanziani M, Debanne D, Muller M, Gahwiler BH, Thompson SM(1994) Role of excitatory amino acid and GABAB receptors in
252

the generation of epileptiform activity in disinhibited hippo-campal slice cultures. Neuroscience 61: 823–832
Sharp PE, Green C (1994) Spatial correlates of firing patterns ofsingle cells in the subiculum of the freely moving rat. J Neurosci14: 2339–2356
Sutor B, Luhmann HJ (1998) Involvement of GABA(B) receptorsin convulsant-induced epileptiform activity in rat neocortex invitro. Eur J Neurosci 10(11): 3417–3427
Traub RD, Wong RK, Miles R, Michelson H (1991) A model of aCA3 hippocampal pyramidal neuron incorporating voltage-clamp data on intrinsic conductances. J Neurophysiol 66: 635–650
Tseng GF, Haberly LB (1988) Characterization of synapticallymediated fast and slow inhibitory processes in piriform cortex inan in vitro slice preparation. J Neurophysiol 59(5): 1352–1376
Velazquez JL, Carlen PL (1999) Synchronization of GABAergicinterneuronal networks during seizure-like activity in the rathorizontal hippocampal slice. Eur J Neurosci 11: 4110–4118
Velıskova J, Velısek L, Moshe SL (1996) Age-specific effects ofbaclofen on pentylenetetrazol-induced seizures in developingrats. Epilepsia 37: 718–722
Vreugdenhil M, Faas GC, Wadman WJ (1998) Sodium currents inisolated rat CA1 neurons after kindling epileptogenesis. Neu-roscience 86: 99–107
Williams SR, Stuart GJ (2002) Dependence of EPSP efficacy onsynapse location in neocortical pyramidal neurons. Science295(5561): 1907–1910
Yang KH, Franaszczuk PJ, Bergey GK (2002) The influence ofsynaptic connectivity on the pattern of bursting behavior inmodel pyramidal cells. Neurocomputing 44–46: 233–242
253