Embed Size (px)
Citation preview

University of South FloridaScholar Commons
Graduate Theses and Dissertations Graduate School
January 2011
The invasive cactus moth, Cactoblastis cactorum:Host plant testing, species interactions, and effectson local Opuntia populationsHeather JezorekUniversity of South Florida, [email protected]
Follow this and additional works at: http://scholarcommons.usf.edu/etd
Part of the American Studies Commons, and the Ecology and Evolutionary Biology Commons
This Dissertation is brought to you for free and open access by the Graduate School at Scholar Commons. It has been accepted for inclusion inGraduate Theses and Dissertations by an authorized administrator of Scholar Commons. For more information, please [email protected].
Scholar Commons CitationJezorek, Heather, "The invasive cactus moth, Cactoblastis cactorum: Host plant testing, species interactions, and effects on localOpuntia populations" (2011). Graduate Theses and Dissertations.http://scholarcommons.usf.edu/etd/3731

The invasive cactus moth, Cactoblastis cactorum: Host plant testing, species interactions,
and effects on local Opuntia populations
by
Heather Jezorek
A dissertation submitted in partial fulfillment
of the requirements for the degree of
Doctor of Philosophy
Department of Integrative Biology
College of Arts and Sciences
University of South Florida
Major Professor: Peter Stiling, Ph.D.
Gordon Fox, Ph.D.
Earl McCoy, Ph.D.
Jason Rohr, Ph.D.
Date of Approval:
November 18th, 2011
Keywords: cacti, oviposition, larval performance, associational resistance, predation
Copyright © 2011, Heather Jezorek

ACKNOWLEDGEMENTS
I have been fortunate to have a wonderful support system while at USF. I first and
foremost thank my major professor, Peter Stiling, for his exceptional guidance and
encouragement, not only on my research, but on all aspects of graduate school. I thank
my committee members, Dr. Gordon Fox, Dr, Earl McCoy, and Dr. Jason Rohr for their
insightful comments on my dissertation, and for freely sharing their time and expertise
during the past six years. I would also like to thank Henry Mushinsky and Susan Bell for
help in acquiring funding and for creating an outstanding environment for higher
education. Many people have helped in the production of this work by assisting with field
work, offering advice, and sharing their knowledge. In particular, I would like to thank
the following past and present members of the Stiling lab: Laura Altfeld, Lance
Arvidson, Kerry Bohl, Tatiana Cornelissen, Christy Foust, Jennifer Maul, and Keith
Stokes. I thank the Department of Integrative Biology, the Fred L. and Helen M. Tharp
Endowed Scholarship Fund, the Biology Graduate Student Organization and USF
Student Government for providing funding for summer research and travel. I thank the
Florida Department of Environmental Protection Southwest District, in particular Terry
Hingten, and Pinellas County Parks, for allowing me use of their parks for field work. I
thank Jim Carpenter, Stephen Hight, and Susan Drawdy for providing live Cactoblastis
cactorum specimens and the opportunity to collaborate. Additional thanks to Jim for
agreeing to serve as an external chair for my doctoral defense. I thank my parents, John

and Susan Jezorek, for their moral and financial support, as well as their superior
grandparenting skills, which permitted me to finish writing this work. They have been
generous, encouraging, and patient throughout all of my life decisions, both major and
minor. I thank my daughter Chessa for timing her arrival impeccably, and for bringing a
new level of love and delight to my life during the final phase of writing. Lastly, I thank
my husband PJ for being a truly amazing person and partner. He has been there for me
each and every day, through tears and joy. I cannot fully express my gratitude, but am
certain that this work would not have been completed without him by my side.

i
TABLE OF CONTENTS
LIST OF TABLES ............................................................................................................. iv
LIST OF FIGURES .............................................................................................................v
ABSTRACT .................................................................................................................. viii
CHAPTER 1- INTRODUCTION ........................................................................................1
Invasive Species .......................................................................................................1
Cactaceae: Opuntioideae ..........................................................................................9
Cactoblastis cactorum ...........................................................................................10
Biology .......................................................................................................10
History: Australia, South Africa, and the Caribbean .................................12
History: U.S. and Mexico ..........................................................................15
References cited .....................................................................................................17
CHAPTER 2- OVIPOSITION PREFERENCE AND LARVAL
PERFORMANCE OF CACTOBLASTIS CACTORUM ON NORTH
AMERICAN OPUNTIOID CACTI ............................................................................26
Synopsis .................................................................................................................26
Introduction ............................................................................................................27
Materials and methods ...........................................................................................32
Oviposition experiment ..............................................................................32
Larval performance experiment .................................................................34
Results ....................................................................................................................36
Oviposition experiment ..............................................................................36
Larval performance experiment .................................................................37
Discussion ..............................................................................................................46
Acknowledgements ................................................................................................50
References cited .....................................................................................................50
CHAPTER 3- ASSOCIATIONAL EFFECTS OF CHAMAECRISTA
FASCICULATA ON ANT PREDATION OF CACTOBLASTIS CACTORUM
AND HERBIVORY OF OPUNTIA.............................................................................56
Synopsis .................................................................................................................56
Introduction ............................................................................................................57
Study system and methods .....................................................................................61
Insects ........................................................................................................61
Plants ..........................................................................................................62

ii
Damage and growth surveys ......................................................................63
Predation experiment .................................................................................66
Results ....................................................................................................................68
Damage and growth surveys ......................................................................68
Predation experiment .................................................................................69
Discussion ..............................................................................................................77
Acknowledgments........................................................................................................82
References cited ...........................................................................................................82
CHAPTER 4- FINDINGS FROM A SIX YEAR STUDY: EFFECTS OF
CACTOBLASTIS CACTORUM ON THE SURVIVAL AND GROWTH OF
LOCAL OPUNTIA .....................................................................................................90
Synopsis .................................................................................................................90
Introduction ............................................................................................................91
Methods..................................................................................................................94
Study system ..............................................................................................94
Data collection and analysis.......................................................................95
Q1: Are plants attacked by C. cactorum more likely to die than
unattacked plants? ......................................................................................97
Q2: How does frequency of attack by C. cactorum affect survival
of plants? ....................................................................................................98
Q3: For surviving plants, does C. cactorum attack affect growth
rate? ..........................................................................................................100
Results ..................................................................................................................101
Q1: Are plants attacked by C. cactorum more likely to die than
unattacked plants? ....................................................................................101
Q2: How does frequency of attack by C. cactorum affect survival
of plants? ..................................................................................................103
Q3: For surviving plants, does C. cactorum attack affect growth
rate? ..........................................................................................................104
Discussion ............................................................................................................115
Acknowledgments................................................................................................120
References cited ...................................................................................................120
CHAPTER 5- LACK OF ASSOCIATIONAL EFFECTS BETWEEN TWO
COMMON HOSTS ON ATTACK RATES BY CACTOBLASTIS
CACTORUM ..............................................................................................................125
Synopsis ...............................................................................................................125
Introduction ..........................................................................................................126
Methods................................................................................................................129
Study system ............................................................................................129
Data collection and analysis.....................................................................131
Results ..................................................................................................................134
Discussion ............................................................................................................143
Acknowledgments................................................................................................146
References cited ...................................................................................................146

iii
APPENDIX I: TAXONOMIC INFORMATION FOR THE OPUNTIOIDEAE
TRIBES CYLINDROPUNTIEAE AND OPUNTIEAE ...........................................154

iv
LIST OF TABLES
Table 2.1: Eggstick and spine data for the fourteen opuntioid taxa...................................40
Table 2.2: Mean values (± SE) for recorded plant characteristics .....................................42
Table 2.3: Results from the no choice larval performance experiment .............................43
Table 2.4: Adult weights and number of eggs per female (mean ± SE) ............................44
Table 2.5: Results from regressions of larval performance variables on epidermal
toughness of the fourteen tested opuntioids .................................................................45
Table 3.1: Native Opuntia herbivores found in central Florida and their
characteristic feeding damage ......................................................................................71
Table 3.2: Relative abundance of ant species in a sample (n=99) collected from
Opuntia individuals ......................................................................................................74
Table 4.1: Summary of attack and survival rates for each survey year ...........................105
Table 4.2: Results from likelihood ratio tests ..................................................................108
Table 4.3: Results from log linear models .......................................................................109
Table 4.4: Results from logistic regression models with final survival status as the
dependent variable .....................................................................................................110
Table 4.5: Results from multiple regressions of ln(proportional cladode
change/generation of C. cactorum) on attack frequency and ln(initial cladode
number) ..................................................................................................................112
Table 5.1: The four native Opuntia herbivores examined in the study ...........................137
Table 5.2: Results from a Mann-Whitney test for differences between O. humifusa
(N=94) and O. stricta (N=96) ....................................................................................138

v
LIST OF FIGURES
Figure 3.1: Results from C. cactorum damage surveys .....................................................72
Figure 3.2: Results from native herbivore and ant surveys ...............................................73
Figure 3.3: Results from a growth survey from April-2009 to Oct-2009 of O.
humifusa plants (n=40) near to and far from C. fasciculata ..............................................75
Figure 3.4: Results from the predation experiment ...........................................................76
Figure 4.1: Histogram of C. cactorum attack and survival rates .....................................106
Figure 4.2: The proportion of plants that died each survey year grouped by C.
cactorum attack status for that same year ..................................................................107
Figure 4.3: Mean proportional change in initial cladode number per generation of
C. cactorum for all surviving plants ..........................................................................111
Figure 4.4: Partial regression plots for O. humifusa ........................................................113
Figure 4.5: Partial regression plots for O. stricta.............................................................114
Figure 5.1: Total cladodes for the four plant species/neighborhood groups
surveyed at HM in 2006 .............................................................................................139
Figure 5.2: Proportion of cladodes with damage from the weevil G. hubbardi for
the four plant species/neighborhood groups surveyed at HM in 2006 ......................140
Figure 5.3: Proportion of cladodes with C. cactorum larvae from the three O.
humifusa neighborhoods surveyed at HM in 2009 ....................................................141
Figure 5.4: Proportion of cladodes with old C. cactorum damage from three O.
humifusa neighborhoods surveyed at HM in 2009 ....................................................142

vi
ABSTRACT
The invasive cactus moth, Cactoblastis cactorum, poses a threat to opunitoid cacti
species of North America. The following work contains four separate studies
investigating C. cactorum host plant preference and performance, predation and
parastitism of C. cactorum, effects of C. cactorum on local Opuntia populations, and
associational effects of host and non-host plants on C. cactorum and native Opuntia-
feeding herbivores. We found that, among southwestern and Mexican opuntioid taxa,
moths preferred O. engelmannii var. linguiformis and var. engelmannii for oviposition,
while Consolea rubescens and O. streptacantha were superior larval hosts. Oviposition
was best predicted by number of cladodes and degree of spininess; epidermal toughness
was a significant predictor of most larval fitness parameters. In general, oviposition
preference was not correlated with larval performance. A lack of co-evolutionary history
between C. cactorum and North American opuntioid species may help explain this
disconnect. We placed irradiated C. cactorum eggsticks and pupae on Opuntia plants in
the field to test for predation. We found evidence of predation, most likely from ants, on
~16% of eggsticks and ~18% of pupae. Predation rates, ant abundance, and cladode
growth were higher, and C. cactorum damage lower, on Opuntia located near the
extrafloral nectar-producing legume Chamaecrista fasciculata. We attribute these
associational effects to the ability of C. fasciculata to attract ants to its extrafloral nectar.
Over the course of six years, ~78% of nearly 600 tagged Opuntia were attacked by C.

vii
cactorum at least once and ~76% of the plants survived. Two separate studies found that
Opuntia stricta was more likely to be attacked by C. cactorum than O. humifusa; we also
found that O. stricta was more likely to die following an attack. A plant‟s odds of
survival decreased as C. cactorum attack frequency increased, but plants that did survive
had positive growth rates, regardless of attack status. We did not find significant evidence
of associational effects for O. humifusa and O. stricta, either for C. cactorum or native
Opuntia-feeding herbivores. It could be that present herbivore densities are low enough,
and host plants plentiful enough, to avoid mechanisms that usually lead to associational
resistance or susceptibility. Overall, our results suggest that the presence and spread of C.
cactorum should be taken seriously, especially for rare opuntioids and the Opuntia-rich
deserts of North America. However, for more common opuntioid host species, there may
be enough resistant or tolerant individuals, and sufficient top down control through ant
predation, for populations to persist at current C. cactorum densities. We acknowledge
that information on Opuntia reproduction and recruitment rates are needed to confirm this
suggestion, and see this as an excellent opportunity for future research.

1
CHAPTER 1
INTRODUCTION
Invasive species
Inarguably, the spread and establishment of non-indigenous species has had an enormous
impact worldwide. Much of this impact has been positive; introduced crops and livestock
make up 98% of the US food industry and bring in approximately $800 billion annually
(Pimentel et al. 2005). However, non-indigenous species can also have far-reaching
negative impacts. It has been suggested that the introduction and spread of such species
ranks second only to habitat loss as a threat to imperiled species in the United States
(Wilcove et al. 1998). Of particular concern is the small percentage of non-indigenous
species that rise to the status of „invasive‟. According to Executive Order 13112 (1999), a
non-indigenous species is considered invasive when its “introduction does or is likely to
cause economic or environmental harm or harm to human health.
It is nearly impossible to accurately quantify the number of non-indigenous
species that quickly perish after arriving in a new habitat, the number that survive for a
brief period before going extinct, and the number that successfully establish (Mack et al.
2000). Based upon past studies of the British Isles, Williamson and Fitter (1996a)
suggest the “tens rule”: 1 out of 10 imported species (accidentally or deliberately) will
appear in the wild, or naturalize; 1 out of those 10 will become established; 1 out of those
10 will become a pest (have a negative economic affect). Of course, this rule is an

2
approximation and is subject to taxonomic variation and exceptions. For example,
biological control insects are deliberately released into the wild and tend to follow more
of a “threes rule” for the proportion that establish and the proportion that lead to control
of the target species (analogous to reaching pest status, as both have an economic effect)
(Williamson and Fitter 1996a). In addition, the rule does not consider those species that
may have negative ecological effects without inflicting any obvious economic effects.
When trying to predict the number of biological invasions that will occur out of a
given number of non-indigenous arrivals, it is only natural to attempt to determine if
there are certain biological attributes shared by successful invaders. Similarly, there may
be biotic and/or abiotic factors of communities that make them more prone to invasion.
This topic has been the subject of many studies (Ehrlich 1989, Noble 1989, Simberloff
1989, Williamson and Fitter 1996a, Williamson and Fitter 1996b, Williamson 1999,
Kolar and Lodge 2001, Colautti et al. 2006) and several theories have been developed to
explain the success of invaders (Tsutsui et al. 2000, Keane and Crawley 2002, Colautti et
al. 2004) and the invasibility of a community (Lonsdale 1999, Davis et al. 2000,
Kennedy et al. 2002). In plants, short juvenile period, short time between large seed
crops and small seed mass were found to be significantly, positively correlated with
invasion success (Rejmanek and Richardson 1996). Ehrlich (1989) suggests that for
vertebrates, most of the characteristics of successful invaders can be grouped under
“broad ecological amplitude”; this includes qualities like large native range, generalist
diet, and ability to tolerate a large range of physical conditions. Today, most scientists
agree that multiple factors act simultaneously on the probability of an introduced species
establishing and becoming invasive. The complexity of the interactions of these factors,

3
coupled with the bias of differential opportunities to receive non-indigenous species, may
make it impossible for us to ever form a predictive theory of invasion (Williamson 1999,
Mack et al. 2000).
Population-, community-, and ecosystem-level effects have been attributed to the
arrival of invasive species. Although there is much more information available for
population- and community-level effects, in the last two decades there has been an
increase in the number of studies investigating the alteration of ecosystem processes by
exotics (Vitousek et al. 1987, Vitousek 1990, Heath et al. 1995, Ehrenfeld et al. 2001,
Schwindt et al. 2004). Effects at any level can be direct, such as the introduction of a
virulent pathogen, or indirect, such as reduced pollination of a plant species following the
introduction of an organism that preys upon one of the plant‟s important pollinators.
Some of the most devastating consequences of invasive species are brought about by the
diseases they carry. A well-known example of this type of direct effect is the elimination
of the American chestnut, Castanea dentata, in the first half of the century by the Asian
chestnut blight, Cryphonectria parasitica, a fungus thought to have arrived via imported
Japanese chestnuts (Anagnostakis 2001, Davelos and Jarosz 2004). Introduction of avian
malaria and avian poxvirus by the mosquito species Culex pipiens quinquefasciatus has
caused substantial declines in Hawaii‟s native avian populations and has contributed to
the extinctions of at least twenty species of honeycreepers and the Hawaiian crow
(extinct in the wild) (Freed et al. 2005). A similar situation could develop on the
Galapagos Islands; Gottdenker et al. (2005) found 13 pathogens in domestic poultry
stocks that are known to adversely affect wild birds. Prevalence of bird pox in Darwin‟s
ground finches rose 33% between 2000 and 2004, and male finches that carried the virus

4
were less likely to find a mate (Kleindorfer and Dudaniec 2006). The rise in disease
could be due to recent increases in the number of domestic chickens on inhabited islands
(Gottdenker et al. 2005), to increasing populations of introduced arthropod vectors, such
as C. pipiens quinquefasciatus (Kleindorfer and Dudaniec 2006), or to a combination of
factors. Declines in some of Galapagos‟ endemic avian species have already been
observed (Wikelski et al. 2004).
Invasive species can also cause the loss of native species through predation or
herbivory. The brown tree snake, Boiga irregularis, was introduced to Guam in 1949 and
has since been responsible for the extirpation of the majority of the island‟s native bird
and bat species, as well as several species of lizards. The loss of these vertebrates will
inevitably lead to indirect ecological effects due to decreased seed dispersal (Fritts and
Rodda 1998, Rodda and Savidge 2007). Boiga irregularis has also caused severe
economic losses and social disturbances in the form of electrical outages, death of pets
and poultry, infant bites, and a decrease in tourism (Rodda and Savidge 2007).
Introduced red deer locally eliminated the understory species Aristotelia chilensis in
Chilean rainforests and drastically reduced its numbers in Patagonian mesic forests
(Veblen et al. 1989). Areas of Nahuel Huapi National Park in Argentina that have dense
red deer and cattle populations have significantly less seedlings and saplings of native
canopy trees, suggesting that browsing and grazing are hindering recruitment (Veblen et
al. 1992). Hybridization of native and introduced congeners, post-introduction evolution,
and competition (to be discussed in more detail later) are still other ways that invasive
species can have detrimental outcomes on native flora and fauna (Mack et al. 2000,
Manchester and Bullock 2000).

5
Most of the above examples deal with population or community level effects.
However, in some instances, invasive species can cause ecosystem-level effects, leading
to even more severe economic and environmental consequences. Alteration by invasive
species of processes like nutrient and energy cycling, water flow, and fire regime have
been found to occur (Vitousek 1990, Mack et al. 2000, Manchester and Bullock 2000,
Crooks 2002). In Hawaii, the nitrogen fixer Myrica faya has been shown to alter primary
succession after volcanic ash fall by adding significant amounts of available fixed
nitrogen to the soil (Vitousek et al 1987, Vitousek 1990). The invasive paper bark tree,
Melaleuca quinquenervia, has totally changed the structure of some of Florida‟s native
sawgrass prairies and cypress swamps. The large scale transformations are thought to be
due to the tree‟s tendency to form dense stands that exclude native plants and provide
poor habitat for native animals, its large water intake, and its ability to increase fire
intensity and frequency while being extremely fire-tolerant itself (Myers 1983, Gordon
1998, Mack et al. 2000, Crooks 2002). Species considered to be “ecosystem engineers”
can have ecosystem-level effects by altering the physical structure of a habitat, which
can, in turn, alter the processes mentioned above by directly or indirectly changing the
availability of resources to other species (Jones et al. 1994). The reef-building polychaete
Ficopomatus enigmaticus is an ecosystem engineer that is considered invasive or exotic
in most of the world‟s temperate, brackish waters. It has clogged intake pipes in power
plants and has also been found to alter sediment deposition and transport and relative
water flow in an Argentinean coastal lagoon (Schwindt et al. 2004).
Many pathways exist for the arrival of new species. Some of these pathways can
be considered natural; air, ocean, and river currents can carry seeds, spores and plant

6
propagules to a new location (Mack 2003); insects can disperse aerially over long
distances (Kiritani and Yamamura 2003). However, in modern times, natural
introductions have a negligible effect compared to human-assisted introductions, both
purposeful and accidental. Some of the most common accidental pathways include the
arrival of non-indigenous organisms via ballast water or soil, encrusted ship hulls, and
cargo. In 1986, Dreissena polymorpha, the zebra mussel, was unintentionally released
into the Great Lakes from ballast water from the Caspian Sea. It has spread throughout
the eastern US and into parts of Canada and now threatens many native invertebrates, as
well as some deep water fish (Vanderploeg et al. 2002, Stiling 2004). “Stowaway”
species are commonly found in shipments of the agriculture and nursery industries, but
packing materials (straw, wood crates) of any industry can provide a pathway for the
introduction of a new plant, animal, or pathogen.
Intentional introductions occur most often through the pet and nursery industries
and often result in unintentional escapes of non-indigenous species (Frank and McCoy
1995, Kraus 2003, Mack 2003). The deliberate release of pets into natural systems can
cause large scale invasions. In some areas of southern Florida, Black Spiny tail iguanas
have become invasive due to a single release event of three individuals by a pet owner
(Krysko et al. 2003). Their omnivorous diet means they not only pose an ecological
threat via consumption of hatchling sea turtles and bird eggs, but are also a nuisance to
residents, eating the flowers and fruits in their gardens. They may possibly be competing
with gopher tortoises for burrowing space and native vegetation (Krysko et al. 2003).
Vertebrates, particularly fish, birds, and mammals, have often been deliberately
introduced to increase hunting or sport fishing opportunities. Micropterus salmoides, the

7
largemouth bass, and Oncorhynchus mykiss, the rainbow trout, have been widely
introduced for recreational fishing; both now claim spots on the „One Hundred of the
World‟s Worst Invasive Alien Species‟ list compiled by the Global Invasive Species
Database (Cambray 2003).
Organisms of all taxa have been released intentionally in the hopes of providing
biological control of other invasive species. Although there are many successful cases of
biological control, the risks to non-target species are difficult to fully assess, and have
prompted much debate (Simberloff and Stiling 1996b, Louda et al. 2003, Louda and
Stiling 2004). Despite the existing potential, there are relatively few documented
examples of adverse effects on native or non-target species by biological control agents,
but this is very likely due to lack of monitoring for these effects (Simberloff and Stiling
1996a, Frank and McCoy 2007). Cactoblastis cactorum, the primary study organism of
this dissertation, is one of ten documented cases of non-target impacts reviewed by
Louda et al (2003). Cactoblastis cactorum has been used as a successful biological
control agent of Opuntia (prickly pear) species in Australia, South Africa, and the
Caribbean. It eventually arrived in Florida where it currently threatens three endemic
opuntioids in the Keys as well as native, non-pest Opuntia populations throughout the
southeastern US.
The rate at which exotic species have been introduced over the past several
hundred years reflects our increasing global trade and travel. Insects, in particular,
provide a good example of this. The number of exotic insects introduced annually into
the U.S. increased from 1640 to 1980 with the rate of increase becoming exponential
around 1920 (Sailer1983, in Mooney and Drake 1989). A study by Frank and McCoy

8
(1992) found that the rate of insect immigration to the US, and perhaps specifically to the
state of Florida, increased markedly in the 1980s, although the increase may partially be
a product of better detection methods. Levine and D‟Antonio (2003) predicted an
increase of 3-60% in the number of non-indigenous insect species present in the U.S.
from 2000 to 2020. The large range in the percent increase demonstrates the difficulty in
making accurate, quantitative, long-term predictions. It is not only our travel and trade-
related activities that lead to establishment of exotics. Humans continue to pollute and
alter the atmosphere, climate, waterways, and terrestrial landscapes, leading to even
greater opportunities for invasive species to establish (Mooney and Drake 1989).
The state of Florida, where this doctoral research took place, is particularly prone
to the establishment of non-indigenous species because the subtropical portion of the
peninsula is bordered by water on three sides and by frost to the north, making it a
“habitat island”. Similar to oceanic islands, it has relatively poor native species richness
(Simberloff 1997). In addition, Florida‟s many miles of coastline, lakes, and rivers ease
the spread of introduced species, as do the nearly 50 million tourists that visit annually
(Invasive Species Working Group 2003). The large ornamental plant business contributes
significantly to the problem; the Port of Miami receives 85% of the live nonindigenous
plant shipments that arrive each year in the US (Gordon 1998). The Florida Exotic Pest
Plant Council lists 147 species of invasive plants on its website, 73 of which are altering
native plant communities (FLEPPC 2009). Florida has the largest number of established
non-indigenous herpetofaunal species in the entire world (Krysko et al. 2011) and is
second only to Hawaii in number of breeding exotic vertebrates (Forys and Allen 1999).

9
While certainly bad news for native flora and fauna, the statements above make Florida
an ideal place for undertaking research of invasive species.
Cactaceae: Opuntioideae
The subfamily Opuntioideae in the family Cactaceae contains five tribes and between
fifteen and eighteen genera (Griffith and Porter 2009). The taxa in this work come from
two tribes, Cylindropuntieae and Opuntieae, and four genera, Consolea, Cylindropuntia,
Nopalea, and Opuntia (see Appendix I).
All opuntioid species are native to the New World. Members of Opuntieae can be
found as far north as British Columbia, in nearly all the contiguous United States, the
Caribbean, and south to the Strait of Magellan. Cylindropuntieae has a narrower
distribution, mainly Mexico and the southwestern US (Britton and Rose 1937, USDA-
NRCS 2010). Florida has six native opuntioids and three species which are considered
naturalized (Wunderlin and Hansen 2008). Taxa in both Opuntieae and Cylindropuntieae
have jointed stems called cladodes. In Opuntieae the cladodes are generally flattened
while in Cylindropuntieae they are cylindrical or globular. Both tribes have species that
are low-growing and shrub-like and species that are tree-like. The cladodes often bear
larger, fixed spines as well as small, barbed, deciduous bristles called glochids.
Opuntioids are both ecologically and economically important. Songbirds, rodents,
snakes, and lizards often take shelter in their spiny cladodes in order to receive shade and
protection (Soberon et al. 2001). The plants also provide nesting structures for birds such
as the Northern bobwhite, reddish egrets, cactus wrens and curve-billed thrashers
(Chavez-Ramirez et al. 1997, Slater et al. 2001, Hernandez et al. 2003, NatureServe

10
2011). The fruits, seeds, and pads are eaten by a wide variety of animals, including deer,
javelinas, lagomorphs, rodents, birds, and tortoises (Macdonald and Mushinsky 1988,
Hoffman et al. 1993, Chavez-Ramirez et al. 1997, Soberon et al. 2001, Birkhead et al.
2005). Many arthropods feed upon the cladodes and make use of their complex structure
for oviposition sites or web construction, while the flowers provide nectar and pollen to
bees, beetles, and butterflies. Prickly pears also prevent soil erosion and desertification,
particularly in arid regions (Soberon et al. 2001).
In Mexico, approximately 250,000 ha of Opuntia are cultivated, bringing in over
80 million dollars annually, and another 3 million ha of wild populations are used for
fodder (Soberon et al. 2001). The fruit is used in jams and juices and the cladodes are
eaten as a vegetable (nopales), used as fodder, burned as an energy source, and used in
manufacturing soap, cosmetics, fertilizer, and adhesives (Soberon et al. 2001, Vigueras
and Portillo 2001). Clearly, there are ample ecological and economical reasons to protect
opuntioid diversity.
Cactoblastis cactorum
Biology
Cactoblastis cactorum is a cactus-feeding moth belonging to the Lepidopteran family
Pyralidae and the subfamily Phycitinae. It is native to northern Argentina, Paraguay,
Uruguay, and southern Brazil, where it is distributed over a wide range of climates.
Larvae are oligophagous, feeding on at least 30 different hosts in the genera Opuntia,
Consolea, and Nopalea (Dodd 1940, Mann 1969, Zimmermann et al. 2004).

11
Adult females are pale brown and grey and have a wingspan of 27-40mm; males
are lighter in color and smaller (23-32mm). The sex ratio is normally 1:1 (Legaspi et al
2009, J. Carpenter, personal communication), but shifts to favor males when conditions
for larvae are suboptimal (Dodd 1940, Zimmermann et al 2004) . The average adult lives
about 9 days and does not feed. Moths emerge from pupae at dusk and mate at dawn on
the first or second day after emergence; females begin laying eggs the next night.
Females lay their eggs one on top of the other to form an eggstick that is attached to the
tip of a cactus spine or directly to the cladode. An average eggstick has 70-90 eggs and a
female may lay several sticks for a total of 200-300 eggs. All eggs from a particular
eggstick hatch at the same time, usually within three or four hours. The neonates work
together to chew a communal entrance hole into the cladode, allowing them to better
overcome the mucilage produced by the cactus when its epidermis is penetrated. They
tunnel inside the cladode, feeding as a group until only the cuticle and vascular bundles
remain. At this point they may either tunnel internally into an adjacent segment or exit
the damaged cladode to crawl over the plant‟s surface to a fresh one (Dodd 1940, Mann
1969). The eaten cladodes are left with a hollowed out, rotting appearance and are more
susceptible to secondary microbial infections (Starmer et al. 1987). Larvae go through
five instars. Early instars are pinkish-cream with dark red dots on the back of each
segment. Later instars become bright orange, and the dots expand and join to become a
dark band across each segment. The caterpillars reach a length of about 1.5 cm. By the
time the larvae are full-grown, the colony has split up and each caterpillar separately
exits the plant, drops to the ground below, and pupates in the soil or leaf litter (Dodd
1940, Zimmermann et al. 2004).

12
In both its native range in and temperate areas of introduction C. cactorum has
two generations per year; the shorter summer generation has an average lifecycle of 100-
120 days while the longer winter generation averages 235-265 days. In warmer areas,
such as Florida, central Queensland, and the Caribbean, there are three generations per
year with a shorter average lifecycle length of about 90 days (Zimmermann et al. 2004,
Hight and Carpenter 2009). The host plants of C. cactorum in Florida include all six
native opuntioid species (Consolea coralicolla, O. stricta, O.humifusa, O. pusilla, O.
triacantha, and O. cubensis) and several naturalized species (O. ficus-indica, O.
cochenillifera, and O. leucotricha). There is concern for all the native species, but
especially for C. coralicolla, O. cubensis, and O. triacantha because in the U.S. they are
found only in the Florida Keys. Consolea coralicolla is an endangered, endemic species
that is restricted to two populations. The Little Torch Key population contains just seven
mature stems (Stiling et al. 2000), while a larger population on Swan Key in Biscayne
National Park contains 586 individuals that have not thus far been attacked by C.
cactorum (Cariaga et al. 2005).
History: Australia, South Africa, and the Caribbean
C. cactorum has been used in several countries to control Opuntia species. The
most famous instance of its use as a biological control agent took place in Australia,
specifically in Queensland and New South Wales (NSW). The first two introductions of
C. cactorum failed to establish, but the third attempt in 1925 was immensely successful
due to intensive mass rearing and distribution efforts by the Commonwealth Prickly Pear
Board. The intial 1925 shipment of 2,750 eggs was increased to 2,540,000 after two

13
generations of cage-rearing. Mass rearing, collection from field sites, and distribution
throughout Opuntia-infested areas continued until 1930, with an estimated total of 3
billion eggs released (Dodd 1940). By 1933, C. cactorum had eradicated 90% of the 25
million hectares infested with nonnative O. inermis and O. stricta, allowing the land to
be reclaimed for agricultural purposes. However, persistent, dense stands of Opuntia still
exist in some coastal regions; water and nutrient stress and dispersal of females to nearby
islands have been suggested as explanations for the lack of control in these areas (Monro
1975, White 1980, Hosking et al. 1994).
After seeing the impressive results produced in Australia, several other countries
imported C. cactorum eggsticks in an effort to control prickly pears. In 1932, South
Africa received 112,600 eggs from Australia, mainly to use against O. ficus-indica.
Although nearly 580 million eggsticks were distributed in the eastern provinces, the moth
did not afford the same level of control as in Queensland (Pettey 1948). The difference in
success is thought to be due to several factors, including the less succulent, woodier
texture of O. ficus-indica, ant predation, and South Africa‟s colder temperatures that
inhibit egg-laying and prolong development times (Pettey 1948, Robertson 1989,
Robertson and Hoffmann 1989). Although it has not completely cleared areas of O. ficus-
indica, C. cactorum has helped to thin out dense stands and reduce its spread by killing
small plants and destroying terminal segments of larger plants. Less than 100,000
hectares of the original 900,000 remain, but much of this success has been attributed to
the scale insect Dactylopius opuntiae, which is considered the primary biological control
agent (Zimmermann and Moran 1991). The species O. stricta and O. aurantiaca are also
considered pest weeds in South Africa. Opuntia stricta is particularly problematic in

14
Kruger National Park, where C. cactorum has decreased the average size of plants, but
increased the density of plants through fragmentation, resulting in no net reduction in
fruiting (Moran and Zimmermann 1991, Hoffmann et al. 1998, Foxcroft et al. 2007).
In 1957, C. cactorum was released on the Caribbean island of Nevis to control
several Opuntia species that had reached abnormal densities due to overgrazing. Despite
their status as weeds, the targeted cacti were all native species. Non-target effects and the
possible dispersal of the moth to neighboring islands were apparently not enough of
concern for the release to be contested. Following the very successful control of the
Opuntia species, especially of O. triacantha, on Nevis, C. cactorum was purposely
introduced on Montserrat and Antigua in 1960 and Grand Cayman in 1970. It also spread
to St. Kitts, the US Virgin Islands, Puerto Rico, Haiti, the Dominican Republic,
Hispaniola, the Bahamas, and Cuba. Whether C. cactorum spread among these islands
via natural dispersal, unintentional human activity, or intentional, but illegal transport by
humans is not known for sure, but it is likely that all three factors played a role.
Although many of the weedy, target species of Opuntia were brought under
control in the Caribbean, non-target species, such as O. repens in Puerto Rico, were also
affected (Tuduri et al. 1971, Bennett and Habeck 1995, Zimmermann et al. 2005). The
status of C. cactorum and Opuntia in the Caribbean was the subject of a 2005 joint report
by the FAO and the IAEA (Zimmermann et al. 2005). Perhaps the most surprising piece
of information from this report was the fact that, except for Puerto Rico, none of the
personnel from quarantine or plant health agencies on the surveyed islands knew of the
presence and effects of C. cactorum on their islands prior to this survey. The authors
attribute this ignorance to a general lack of interest in Opuntia species in the Caribbean,

15
where they are not actively used or cultivated and now mostly exist only as minor pests.
On many of the islands surveyed in 2005 (Montserrat, Antigua, St. Kitts, Grand Cayman,
Guadeloupe, Dominique, Grenada, St. Lucia and Martinique) Opuntia populations were
sparse and a high proportion of the plants that were located had been attacked by C.
cactorum. The first confirmed record of the moth on Jamaica was made during the 2005
survey. Cactoblastis cactorum damage was found on five species and botanists described
a recent decline in several Opuntia species, including the already rare O. sanguinea. The
moth is apparently also abundant in Cuba and the Dominican Republic, although time
and political constraints prevented a thorough survey of these islands. A 2002 survey of
Nevis (Pemberton and Liu 2007), the original site of C. cactorum introduction in the
Caribbean, found the moth on two native (O. stricta (dillenii) and O. triacantha) and one
naturalized species (N. cochenillifera), although less than 30% of plants were attacked.
As has been found in previous studies, larger plants were more likely to be attacked, but
smaller plants had a greater proportion of attacked cladodes, indicating that they may
suffer a higher mortality risk (Pemberton and Liu 2007).
Other locales where C. cactorum has been successfully introduced for control of
non-native Opuntia include Mauritius, Hawaii, St. Helena Island, and Ascension Island.
It has been introduced, but has not established in Pakistan, Kenya, and Israel
(Zimmermann et al. 2004).

16
History: U.S. and Mexico
In 1989, after being warned of C. cactorum‟s presence in Cuba (still unconfirmed
at that point), scientists from the Fairchild Tropical Gardens in Miami visited the Florida
Keys to check for the moth. It was found on O. stricta on Big Pine Key and, less than
one year later, in Key Biscayne State Park near Miami. The large distance (200 km)
between these two locations, coupled with records of C. cactorum interceptions in Miami
between 1981 and 1986, suggest that the moth may have arrived via imported Opuntia
from the Dominican Republic (Pemberton 1995, Stiling 2002). Regardless of whether the
moth‟s arrival was human-mediated, or it flew the 145 km from Cuba to the Keys, C.
cactorum has since spread up the east coast to Bull Island, SC, which has remained its
northern leading edge since 2004. The farthest western detection occurred in 2009 in
Terre Bonne Parrish, south of New Orleans, LA (USDA-APHIS-PPQ 2009b).
Ongoing efforts in the southeastern states are being made to carefully monitor the
spread of C. cactorum and to push back the western leading edge of its distribution.
Mechanical control (removal of eggsticks by hand) is not practical and biological and
chemical control methods could have detrimental non-target effects on native
lepidopterans (Stiling 2002). Therefore, genetic control via the release of sterile males is
the best option for halting the spread of the moth. This technique is currently used on
barrier islands along the AL and MS coast (Carpenter et al. 2001, Hight et al. 2005).
Pheromone traps are also being used to monitor the moth‟s current range and to survey
for new outbreaks (USDA-APHIS-PPQ 2010).
In 2006 C. cactorum was discovered on Isla Mujeres in the Mexican state of
Quintana Roo; this was followed by detection in 2007 on nearby Isla Contoy. The most

17
likely arrival path is thought to be from Cuba, Haiti, or the Dominican Republic
(Zimmermann et al. 2004). An eradication program on Isla Mujeres, focusing on host
removal, pheromone trapping, and outreach, was quickly put into place. Subsequently,
the eradication program was extended to Isla Contoy, where infested Opuntia cladodes
were removed and sterile moths from the USDA mass rearing program in Tifton, GA
were released. Traps were also set up on the Mexican mainland in the Cancun area
(Rodriguez 2007). Both islands have since been declared eradicated and as of this
writing, no other Mexican infestations have been found (USDA-APHIS-PPQ 2008,
2009a)
References cited
Anagnostakis, S. L. 2001. American chestnut sprout survival with biological control of
the chestnut-blight fungus population. Forest Ecology and Management
152:225-233.
Bennett, F. D. and D. H. Habeck. 1995. Cactoblastis cactorum: A successful weed
control agent in the Caribbean, now a pest in Florida? Pages 21-26 in E. S.
Delfosse and R. R. Scott, editors. Biological control of weeds. Proceedings of the
8 International Symposium on Biological Control of Weeds. Lincoln University,
Canterbury, New Zealand, 2-7 February 1992.
Birkhead, R. D., C. Guyer, S. M. Hermann, and W. K. Michener. 2005. Patterns of
folivory and seed ingestion by gopher tortoises (Gopherus polyphemus) in a
southeastern pine savanna. American Midland Naturalist 154:143-151.
Britton, N. L. and J. N. Rose. 1937. The Cactaceae: Descriptions and Illustrations of
Plants of the Cactus Family. Dover Publications, New York.
Cambray, J. A. 2003. Impact on indigenous species biodiversity caused by the
globalisation of alien recreational freshwater fisheries. Hydrobiologia 500:217-
230.
Cariaga, K. A., C. E. Lewis, J. Maschinski, S. J. Wright, and J. Francisco-Ortega. 2005.
Patterns of genetic diversity in the critically endangered Florida key endemic

18
Consolea corallicola Small (Cactaceae): Evidence from inter-simple sequence
repeat (ISSRs) DNA polymorphisms. Caribbean Journal of Science 41:225-233.
Carpenter, J. E., K. A. Bloem, and S. Bloem. 2001. Applications of F-1 sterility for
research and management of Cactoblastis cactorum (Lepidoptera : Pyralidae).
Florida Entomologist 84:531-536.
Chavez-Ramirez, F., X. G. Wang, K. Jones, D. Hewitt, and P. Felker. 1997. Ecological
characterization of Opuntia clones in South Texas: Implications for wildlife
herbivory and frugivory. Journal of the Professional Association for Cactus
Development 2:9-19.
Colautti, R. I., I. A. Grigorovich, and H. J. MacIsaac. 2006. Propagule pressure: A null
model for biological invasions. Biological Invasions 8:1023-1037.
Colautti, R. I., A. Ricciardi, I. A. Grigorovich, and H. J. MacIsaac. 2004. Is invasion
success explained by the enemy release hypothesis? Ecology Letters 7:721-733.
Crooks, J. A. 2002. Characterizing ecosystem-level consequences of biological
invasions: The role of ecosystem engineers. Oikos 97:153-166.
Davelos, A. L. and A. M. Jarosz. 2004. Demography of American chestnut populations:
Effects of a pathogen and a hyperparasite. Journal of Ecology 92:675-685.
Davis, M. A., J. P. Grime, and K. Thompson. 2000. Fluctuating resources in plant
communities: A general theory of invasibility. Journal of Ecology 88:528-534.
Dodd, A. P. 1940. The biological campaign against prickly pear. Commonwealth Prickly
Pear Board Bulletin:1-177. Brisbane, Australia.
Ehrenfeld, J. G., P. Kourtev, and W. Z. Huang. 2001. Changes in soil functions following
invasions of exotic understory plants in deciduous forests. Ecological
Applications 11:1287-1300.
Ehrlich, P. R. 1989. Attributes of invaders and the invading process: Vertebrates. Pages
315-328 in J. A. Drake et al, editors. Biological Invasions: A Global Perspective.
Wiley and Sons, Chichester, U.K.
Executive Order No. 13112. 1999. Federal Registry 64:6183
FLEPPC. 2009. List of Invasive Plant Species. Wildland Weeds 12:13-16.
Forys, E. A. and C. R. Allen. 1999. Biological invasions and deletions: Community
change in south Florida. Biological Conservation 87:341-347.
Foxcroft, L. C., J. H. Hoffmann, J. J. Viljoen, and J. J. Kotze. 2007. Factors influencing
the distribution of Cactoblastis cactorum, a biological control agent of Opuntia
stricta in Kruger National Park, South Africa. South African Journal of Botany
73:113-117.

19
Frank, J. H. and E. D. McCoy. 1995. Introduction to insect behavioral ecology: The
good, the bad, and the beautiful: Nonindigenous species in Florida. Invasive
adventive insects and other organisms in Florida. Florida Entomologist 78:1-15.
Frank, J. H. and E. D. McCoy. 2007. The risk of classical biological control in Florida.
Biological Control 41:151-174.
Freed, L. A., R. L. Cann, M. L. Goff, W. A. Kuntz, and G. R. Bodner. 2005. Increase in
avian malaria at upper elevation in Hawai'i. Condor 107:753-764.
Fritts, T. H. and G. H. Rodda. 1998. The role of introduced species in the degradation of
island ecosystems: A case history of Guam. Annual Review of Ecology and
Systematics 29:113-140.
Gordon, D. R. 1998. Effects of invasive, non-indigenous plant species on ecosystem
processes: Lessons from Florida. Ecological Applications 8:975-989.
Gottdenker, N. L., T. Walsh, H. Vargas, J. Merkel, G. U. Jimenez, R. E. Miller, M.
Dailey, and P. G. Parker. 2005. Assessing the risks of introduced chickens and
their pathogens to native birds in the Galapagos Archipelago. Biological
Conservation 126:429-439.
Griffith, M. P. and J. M. Porter. 2009. Phylogeny of Opuntioideae (Cactaceae).
International Journal of Plant Sciences 170:107-116.
Heath, R. T., G. L. Fahnenstiel, W. S. Gardner, J. F. Cavaletto, and S. J. Hwang. 1995.
Ecosystem-level effects of zebra mussels (Dreissena polymorpha): An enclosure
experiment in Saginaw Bay, Lake Huron. Journal of Great Lakes Research
21:501-516.
Hernandez, F., S. E. Henke, N. J. Silvy, and D. Rollins. 2003. The use of prickly pear
cactus as nesting cover by northern bobwhites. Journal of Wildlife Management
67:417-423.
Hight, S. D. and J. E. Carpenter. 2009. Flight phenology of male Cactoblastis cactorum
(Lepidoptera: Pyralidae) at different latitudes in the southeastern United States.
Florida Entomologist 92:208-216.
Hight, S. D., J. E. Carpenter, S. Bloem, and K. A. Bloem. 2005. Developing a sterile
insect release program for Cactoblastis cactorum (Berg) (Lepidoptera :
Pyralidae): Effective overflooding ratios and release-recapture field studies.
Environmental Entomology 34:850-856.
Hoffman, M. T., C. D. James, G. I. H. Kerley, and W. G. Whitford. 1993. Rabbit
herbivory and its effect on cladode, flower, and fruit production of Opuntia
violacea var. macrocentra (Cactaceae) in the Northern Chihuahuan Desert, New-
Mexico. Southwestern Naturalist 38:309-315.

20
Hoffmann, J. H., V. C. Moran, and D. A. Zeller. 1998. Evaluation of Cactoblastis
cactorum (Lepidoptera : Phycitidae) as a biological control agent of Opuntia
stricta (Cactaceae) in the Kruger National Park, South Africa. Biological Control
12:20-24.
Hosking, J. R., P. R. Sullivan, and S. M. Welsby. 1994. Biological control of Opuntia
stricta (Haw) Haw var. stricta using Dactylopius opuntiae (Cockerell) in an area
of New South Wales, Australia, where Cactoblastis cactorum (Berg) is not a
successful biological control agent. Agriculture Ecosystems & Environment
48:241-255.
Jones, C. G., J. H. Lawton, and M. Shachak. 1994. Organisms as ecosystem engineers.
Oikos 69:373-386.
Keane, R. M. and M. J. Crawley. 2002. Exotic plant invasions and the enemy release
hypothesis. Trends in Ecology & Evolution 17:164-170.
Kennedy, T. A., S. Naeem, K. M. Howe, J. M. H. Knops, D. Tilman, and P. Reich. 2002.
Biodiversity as a barrier to ecological invasion. Nature 417:636-638.
Kiritani, K. and K. Yamamura. 2003. Exotic insects and their pathways for invasion.
Pages 44-67 in G. M. Ruiz and J. T. Carlton, editors. Invasive Species: Vectors
and Management Strategies. Island Press, Washington D.C.
Kleindorfer, S. and R. Y. Dudaniec. 2006. Increasing prevalence of avian poxvirus in
Darwin's finches and its effect on male pairing success. Journal of Avian Biology
37:69-76.
Kolar, C. S. and D. M. Lodge. 2001. Progress in invasion biology: Predicting invaders.
Trends in Ecology & Evolution 16:199-204.
Kraus, F. 2003. Invasion pathways for terrestrial vertebrates. Pages 68-92 in G. M. Ruiz
and J. T. Carlton, editors. Invasive Species: Vectors and Management Strategies.
Island Press, Washington D.C.
Krysko, K. L., J. P. Burgess, M. R. Rochford, C. R. Gillette, D. Cueva, K. M. Enge, L.
A. Somma, J. L. Stabile, D. C. Smith, J. A. Wasilewski, G. N. Kieckhefer, M. C.
Granatosky, and S. V. Nielsen. 2011. Verified non-indigenous amphibians and
reptiles in Florida from 1863 through 2010: Outlining the invasion process and
identifying invasion pathways and stages. Zootaxa Issue 3028 (1-64).
Krysko, K. L., F. W. King, K. M. Enge, and A. T. Reppas. 2003. Distribution of the
introduced black spiny-tailed iguana (Ctenosaura similis) on the southwestern
coast of Florida. Biological Sciences 66:74-79.
Legaspi, J.C., I. Baez, B. C. Legaspi. 2009. Phenology and egg production of the cactus
moth (Lepidoptera: Pyralidae): comparison of field census data and life stage
development in the field. Journal of Entomological Science 44: 341-352.

21
Lonsdale, W. M. 1999. Global patterns of plant invasions and the concept of invasibility.
Ecology 80:1522-1536.
Louda, S. M., R. W. Pemberton, M. T. Johnson, and P. A. Follett. 2003. Nontarget
effects - The Achilles' Heel of biological control? Retrospective analyses to
reduce risk associated with biocontrol introductions. Annual Review of
Entomology 48:365-396.
Louda, S. M. and P. Stiling. 2004. The double-edged sword of biological control in
conservation and restoration. Conservation Biology 18:50-53.
Macdonald, L. A. and H. R. Mushinsky. 1988. Foraging ecology of the gopher tortoise,
Gopherus polyphemus, in a sandhill habitat. Herpetologica 44:345-353.
Mack, R. N. 2003. Global plant dispersal, naturalization, and invasion: Pathways,
models, and circumstances. Pages 3-30 in G. M. Ruiz and J. T. Carlton, editors.
Invasive Species: Vectors and Management Strategies. Island Press, Washington,
D.C.
Mack, R. N., D. Simberloff, W. M. Lonsdale, H. Evans, M. Clout, and F. A. Bazzaz.
2000. Biotic invasions: Causes, epidemiology, global consequences, and control.
Ecological Applications 10:689-710.
Manchester, S. J. and J. M. Bullock. 2000. The impacts of non-native species on UK
biodiversity and the effectiveness of control. Journal of Applied Ecology 37:845-
864.
Mann, J. 1969. Cactus-feeding Insects and Mites. United States National Museum
Bulletin 256.
Monro, J. M. 1975. Environmental variation and the efficiency of biological control -
Cactoblastis in the southern hemisphere. Pages 204-212 in Kikkawa, J. and H.A.
Nix (Eds.). Proceedings of the Ecological Society of Australia, Vol. 9. Managing
Terrestrial Ecosystems. Symposium. Brisbane, Australia., May 15-16, 1975.
Mooney, H. A. and J. A. Drake. 1989. Biological Invasions: A SCOPE program
overview. Pages 491-503 in J. A. Drake et al, editors. Biological invasions: A
global perspective. Wiley and Sons, Chichester, U.K.
Moran, V. C. and H. G. Zimmermann. 1991. Biological control of jointed cactus,
Opuntia aurantiaca (Cactaceae), in South Africa. Agriculture Ecosystems &
Environment 37:5-27.
Myers, R. L. 1983. Site susceptibility to invasion by the exotic tree Melaleuca-
quinquenervia in Southern Florida. Journal of Applied Ecology 20:645-658.
NatureServe. 2011. NatureServe Explorer: An online encyclopedia of life, Version 7.1
NatureServe.

22
Noble, I. R. 1989. Attributes of invaders and the invading process: Terrestrial and
vascular plants. Pages 301-313 in J. A. Drake et al, editors. Biological invasions:
A global perspective. Wiley and Sons, Chichester, UK.
Pemberton, R. W. 1995. Cactoblastis cactorum (Lepidoptera: Pyralidae) in the United
States: An immigrant biological control agent or an introduction of the nursery
industry? American Entomologist 41:230-232.
Pemberton, R. W. and H. Liu. 2007. Control and persistence of native Opuntia on Nevis
and St. Kitts 50 years after the introduction of Cactoblastis cactorum. Biological
Control 41:272-282.
Pettey, F. W. 1948. The biological control of prickly pear in South Africa. Science
Bulletin. Department of Agriculture of the Union of South Africa.
Pimentel, D., R. Zuniga, and D. Morrison. 2005. Update on the environmental and
economic costs associated with alien-invasive species in the United States.
Ecological Economics 52:273-288.
Rejmanek, M. and D. M. Richardson. 1996. What attributes make some plant species
more invasive? Ecology 77:1655-1661.
Robertson, H. G. 1989. Seasonal temperature effects on fecundity of Cactoblastis
cactorum (Berg) (Lepidoptera: Pyralidae): Differences between South Africa and
Australia. Journal of the Entomological Society of Southern Africa 52:71-80.
Robertson, H. G. and J. H. Hoffmann. 1989. Mortality and life-tables of Cactoblastis
cactorum (Berg) (Lepidoptera: Pyralidae) compared on two host-plant species.
Bulletin of Entomological Research 79:7-17.
Rodda, G. H. and J. A. Savidge. 2007. Biology and impacts of pacific island invasive
species. 2. Boiga irregularis, the Brown Tree Snake (Reptilia: Colubiridae).
Pacific Science 61:307-324.
Rodriguez, R. Z. 2007. Detection and eradication campaign on Isla Mujeres, Quintana
Roo, Mexico. International Cactoblastis cactorum Conference, Phoenix, AZ.
Schwindt, E., O. O. Iribarne, and F. I. Isla. 2004. Physical effects of an invading reef-
building polychaete on an Argentinean estuarine environment. Estuarine Coastal
and Shelf Science 59:109-120.
Simberloff, D. 1989. Which insect introductions succeed and which fail? Pages 61-75 in
J. A. Drake et al, editors. Biological invasions: A global perspective. Wiley and
Sons, Chichester, UK.
Simberloff, D. 1997. The biology of invasions. Pages 3-20 in D. Simberloff, D. C.
Schmitz, and T. C. Brown, editors. Stranger in Paradise: Impact and Management
of Nonindigenous Species in Florida. Island Press, Washington, D.C.

23
Simberloff, D. and P. Stiling. 1996a. How risky is biological control? Ecology 77:1965-
1974.
Simberloff, D. and P. Stiling. 1996b. Risks of species introduced for biological control.
Biological Conservation 78:185-192.
Slater, S. C., D. Rollins, R. C. Dowler, and C. B. Scott. 2001. Opuntia: A "prickly
paradigm" for quail management in west-central Texas. Wildlife Society Bulletin
29:713-719.
Soberon, J., J. Golubov, and J. Sarukhan. 2001. The importance of Opuntia in Mexico
and routes of invasion and impact of Cactoblastis cactorum (Lepidoptera :
Pyralidae). Florida Entomologist 84:486-492.
Starmer, W. T., V. Aberdeen, and M. Lachance. 1987. The yeast community associated
with decaying Opuntia stricta (Haworth) in Florida with regard to the moth.
Cactoblastis cactorum (Berg). Florida Scientist 51:1-6.
Stiling, P. 2002. Potential non-target effects of a biological control agent, prickly pear
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), in North America,
and possible management actions. Biological Invasions 4:273-281.
Stiling, P. 2004. Ecology: theories and applications. 4th edition. Prentice Hall India, New
Delhi.
Stiling, P., A. Rossi, and D. Gordon. 2000. The difficulties of single factor thinking in
restoration: replanting a rare cactus in the Florida Keys. Biological Conservation
94:327-333.
Tsutsui, N. D., A. V. Suarez, D. A. Holway, and T. J. Case. 2000. Reduced genetic
variation and the success of an invasive species. Proceedings of the National
Academy of Sciences of the United States of America 97:5948-5953.
Tuduri, J. C. G., L. F. Martorell, and S. M. Gaud. 1971. Geographical distribution and
host plants of the cactus moth, Cactoblastis cactorum (Berg) in Puerto Rico and
the US Virgin Islands. Journal of the Agricultural University Puerto Rico 58:130-
134.
USDA-APHIS-PPQ. 2008. Cactus moth (Cactoblastis cactorum) program update. United
States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine.
USDA-APHIS-PPQ. 2009a. Cactus moth (Cactoblastis cactorum) program update.
United States Department of Agriculture, Animal and Plant Health Inspection
Service, Plant Protection and Quarantine.
USDA-APHIS-PPQ. 2009b. Cactus moth program annual report, Eastern region. United
States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine.

24
USDA-APHIS-PPQ. 2010. Cactus moth program annual report, Eastern region. United
States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine.
USDA-NRCS. 2011. The PLANTS Database (http://plants.usda.gov), 1 October 2011.
National Plant Data Team, Greensboro, NC 27401-4901 USA.
Vanderploeg, H. A., T. F. Nalepa, D. J. Jude, E. L. Mills, K. T. Holeck, J. R. Liebig, I.
A. Grigorovich, and H. Ojaveer. 2002. Dispersal and emerging ecological
impacts of Ponto-Caspian species in the Laurentian Great Lakes. Canadian
Journal of Fisheries and Aquatic Sciences 59:1209-1228.
Veblen, T. T., M. Mermoz, C. Martin, and T. Kitzberger. 1992. Ecological impacts of
introduced animals in Nahuel-Huapi-National-Park, Argentina. Conservation
Biology 6:71-83.
Veblen, T. T., M. Mermoz, C. Martin, and E. Ramilo. 1989. Effects of exotic deer on
forest regeneration and composition in northern Patagonia. Journal of Applied
Ecology 26:711-724.
Vigueras, A. L. and L. Portillo. 2001. Uses of Opuntia species and the potential impact
of Cactoblastis cactorum (Lepidoptera : Pyralidae) in Mexico. Florida
Entomologist 84:493-498.
Vitousek, P. M. 1990. Biological invasions and ecosystem processes - towards an
integration of population biology and ecosystem studies. Oikos 57:7-13.
Vitousek, P. M., L. R. Walker, L. D. Whiteaker, D. Muellerdombois, and P. A. Matson.
1987. Biological Invasion by Myrica faya alters ecosystem development in
Hawai‟i. Science 238:802-804.
White, G. C. 1980. Current status of prickly pear control by Cactoblastis cactorum in
Queensland. Pages 609-616 in Proceedings of the Vth International Symposium
on Biological Control of Weeds, Brisbane, Australia.
Wikelski, M., J. Foufopoulos, H. Vargas, and H. Snell. 2004. Galapagos birds and
diseases: Invasive pathogens as threats for island species. Ecology and Society 9.
Wilcove, D. S., D. Rothstein, J. Dubow, A. Phillips, and E. Losos. 1998. Quantifying
threats to imperiled species in the United States. Bioscience 48:607-615.
Williamson, M. 1999. Invasions. Ecography 22:5-12.
Williamson, M. and A. Fitter. 1996a. The varying success of invaders. Ecology 77:1661-
1666.
Williamson, M. H. and A. Fitter. 1996b. The characters of successful invaders.
Biological Conservation 78:163-170.

25
Wunderlin, R. P. and B. F. Hansen. 2008. Atlas of Florida Vascular Plants. Institute for
Systematic Botany, University of South Florida, Tampa.
Zimmermann, H. G., S. Bloem, and H. Klein. 2004. Biology, history, threat, surveillance
and control of the cactus moth, Cactoblastis cactorum. International Atomic
Energy Agency/Food and Agriculture Organization, Vienna, Austria.
Zimmermann, H. G., M. P. Cuen, and A. Rivera. 2005. The status of Cactoblastis
cactorum (Lepidoptera: Pyralidae) in the Caribbean and the likelihood of its
spread to Mexico. IAEA, Vienna.
Zimmermann, H. G. and V. C. Moran. 1991. Biological control of prickly pear, Opuntia
ficus-indica (Cactaceae), in South Africa. Agriculture Ecosystems &
Environment 37:29-35.

26
CHAPTER 2
OVIPOSITION PREFERENCE AND LARVAL PERFORMANCE OF
CACTOBLASTIS CACTORUM ON NORTH AMERICAN OPUNTIOID CACTI
Synopsis
Cactoblastis cactorum Berg (Lepidoptera: Pyralidae), the cactus moth, is a well-known
biological control agent of prickly pear cactus (Cactaceae: Opuntia Miller). The arrival
of the moth in Florida and its subsequent spread through the southeastern United States
poses a threat to opuntioid diversity in North America. Of particular concern are the
ecological and economic impacts the moth could have in the southwestern United States
and Mexico, where both native and cultivated Opuntia species are important resources. It
is unknown which species would best support larval development if the moth were to
spread further westward in North America. This study aimed to determine if ovipositing
females demonstrate preferences for any of fourteen common opuntioids native to or
naturalized in Mexico and the southwestern United States; which of these opuntioids best
support larval development; and if oviposition preference correlates with larval
performance, as predicted by simple adaptive models. Results from a field experiment
showed that female moths preferred O. engelmannii Salm-Dyck ex Engelmann var.
linguiformis (Griffiths) Parfitt and Pinkava and O. engelmannii var. engelmannii for
oviposition. A generalized linear model showed number of cladodes and degree of
spininess to be significant predictors of oviposition activity. Results from a no-choice

27
larval survival experiment found Consolea rubescens (Salm-Dyck ex de Candolle.)
Lemaire and O. streptacantha Lemaire to be the best hosts among the taxa tested.
Epidermal toughness was a significant predictor of most larval fitness parameters. In
general, oviposition preference was not correlated with larval performance. A lack of co-
evolutionary history between C. cactorum and North American opuntioid species may
help explain this disconnect.
Introduction
The cactus moth, Cactoblastis cactorum Berg (Lepidoptera: Pyralidae), is well known
for its role in the biological control of Opuntia Miller species, including the highly
successful campaign in Australia during the 1920s and 1930s, the less successful
campaign begun in the late 1930s in South Africa, and the campaign against native cacti
in the Caribbean begun in the 1950s (Dodd 1940, Pettey 1948, Bennett et al. 1995,
Zimmermann et al. 2004, Zimmermann et al. 2005). However, with the 1989 discovery
of C. cactorum in the Florida Keys (Habeck and Bennett 1990, Dickel 1991), the moth
has become a potential threat to Opuntia biodiversity in continental North America
(Bennett et al. 1995, Zimmermann et al. 2000, Stiling 2002). Of particular concern are:
1) populations of rare opuntioids (members of the subfamily Opuntioideae: Cactaceae),
for example Consolea corallicola Small and Opuntia triacantha (Willdenow) Sweet in
the Florida Keys (Stiling et al. 2004, Stiling 2010) and O. sanguinea Proctor and O.
jamaicensis Britton in Jamaica (Zimmermann et al. 2005); 2) the possible adverse effects
on wild opuntioid species diversity found in the southwestern United States and Mexico
(Perez-Sandi 2001, Soberon et al. 2001, Vigueras and Portillo 2001, Zimmermann et al.

28
2004); and 3) the economic devastation to Mexico, where cultivated Opuntia is a
significant commodity (Soberon et al. 2001, Vigueras and Portillo 2001, Simonsen et al.
2005).
The Opuntioideae are both ecologically and economically important. In the desert
regions of the southwestern US and Mexico, wild Opuntia and Cylindropuntia
(Engelmann) Kreuzinger species provide nesting sites for birds, reptiles, and rodents,
cladodes and fruit for a variety of wildlife, and contribute to soil stability (Chavez-
Ramirez et al. 1997, Hervert et al. 2005). In Mexico, approximately 250,000 ha of
Opuntia and Nopalea Salm-Dyck species are cultivated for food, livestock fodder, fuel,
and industrial manufacturing; another 3 million ha of wild populations are used for
fodder (Soberon et al. 2001, Vigueras and Portillo 2001). The estimated annual value of
opuntioid production in Mexico is over 80 million dollars (Soberon et al. 2001). If C.
cactorum is able to establish populations in the southwestern United States or in Mexico,
the impacts on opuntioid diversity and agriculture could be severe.
Presently, C. cactorum is established in the southeastern United States and has
been found as far north as Bull Island, South Carolina and as far west as southeastern
Louisiana (Simonsen et al. 2005, USDA-APHIS-PPQ 2009). Two populations of the
moth were also detected on islands off the Yucatan peninsula, the first on Isla Mujeres
(detected April 2006) and the second on Isla Contoy (detected May 2007) (USDA-
APHIS-PPQ 2009). A combination of tactics including the sterile insect technique (SIT),
in particular F1 or inherited sterility (Carpenter et al. 2001b, Carpenter et al. 2001a,
Hight et al. 2005, Tate et al. 2007), sanitation activities (removal of infested plant parts),
and host plant removal has been used in western coastal Alabama since March 2005 and

29
at Isla Mujeres and Isla Contory in 2008. Both sites in Mexico have since been declared
eradicated and are under surveillance by the Mexican government (USDA-APHIS-PPQ
2009). These tactics have been successful in reducing moth populations along the U.S.
Gulf Coast, but recent detections of C. cactorum in Mississippi (in January and April
2008 and August 2009) and Louisiana (in June and July 2009) (USDA-APHIS-PPQ
2009) attest to the continued threat of this pest to Opuntia-rich areas of North America.
The long-term effects of C. cactorum on cacti populations in the United States and
Mexico are uncertain, but the moth is known to kill small plants and decrease biomass of
larger plants, leading to the expectation of extinctions and reduced population sizes
(Dodd 1940, Johnson and Stiling 1996, Hoffmann et al. 1998, Zimmermann et al. 2000,
Baker and Stiling 2009).
Determining the potential effects of a C. cactorum invasion involves verifying its
potential host range by testing possible host species from the threatened region for
oviposition preference and larval performance. Among the five species of Cactoblastis
Ragonot, all of which are native to South America, C. cactorum has the widest
geographic and host range. It is distributed through southern Brazil, Uruguay, Paraguay,
northern Argentina and has at least 30 known host species from three genera within the
subfamily Opuntioideae (Opuntia, Consolea Lemaire, and Nopalea Salm-Dyck), many
of which occur outside its native range (Mann 1969, Zimmermann et al. 2004). Opuntia
and Nopalea spp. can be tree- or shrub-like, with determinate, segmented trunks and/or
branches and flattened cladodes. Consolea spp. are endemic to the Caribbean Basin and
are usually tree-like, with an indeterminate, unsegmented trunk from which flattened
cladodes branch off in an asymmetrical fashion (Anderson 2001). Members of Consolea

30
have a unique breeding system in that they are cryptically dioecious/subdioecious, with
functionally staminate flowers that occasionally produce fruit (Strittmatter et al. 2006). It
is unclear how risk would vary among those hosts native to Mexico and the western
United States. Cactoblastis cactorum has been shown to attack two economically
important species native to Mexico, O. ficus-indica (L.) Miller and O. streptacantha
Lemaire, but is not thought to attack Cylindropuntia species, although low levels of
damage are seen when such species grow in close proximity to preferred hosts
(Zimmermann et al. 2004). In the wild, the moth would encounter many opuntioid
species for the first time, and risks to these “new association” species are largely
unknown. Host range testing in the southwestern United States or Mexico is not practical
because of concerns about unintentional releases or escapes. To minimize the risk of
escape, tests can be done in laboratories under strict quarantine, but the host range
determined from laboratory tests can sometimes be a poor indicator of the host range in
the field (Hajek et al. 1996, Louda et al. 1997, Louda et al. 2003). For potential
biological control agents, an emerging idea is to perform field studies in the invaded
region with sterilized insects; this is particularly promising for lepidopteran studies
because of the process of F1 sterility (Tate et al. 2009). In C. cactorum, it has been
shown that dispersal, mating frequency, and host preference are not significantly
different between untreated moths and moths treated with 200 Gy of gamma radiation,
the most effective dose for F1 sterility in C. cactorum (Hight et al. 2005, Tate et al.
2007, Marti and Carpenter 2009, Tate et al. 2009). An alternative option is to assess risks
to target host species by moving them from the threatened region into regions where the
insect is native or well-established.

31
The first part of this study tests C. cactorum oviposition preference among
fourteen common opuntioid taxa, most of which are native to the southwestern United
States and/or Mexico, but which were moved to Florida for the experiment. Our
assumption is that C. cactorum females, if they were to establish in the desert southwest,
would use the same oviposition cues as they do in Florida, and would thereby exhibit
similar preferences to those found in this study. We included plant height, cladode
number, epidermal toughness, and spininess in a generalized linear model to determine
which, if any, of these characteristics could be useful predictors of oviposition
preference. The second part of this study tested larval performance in a laboratory
setting, using the same cactus taxa from the oviposition experiment. Here we assume that
larval performance in the laboratory will be similar to larval performance in the desert
southwest, although this assumption is made cautiously. Finally, we compare larval
performance with oviposition preference, as it is often thought that these two attributes
should be correlated among various hosts, especially in cases where the larvae are
relatively immobile (Levins and Macarthur 1969, Jaenike 1978, 1990).
Opuntioid taxonomy is not fully agreed upon and remains highly confusing, in
part because of the tendency for hybridization and polymorphism among its species
(Chavez-Ramirez et al. 1997). Here, the classifications of Anderson (2001) and the
USDA-ARS Germplasm Information Resources Network (USDA-ARS 2009) were
followed when classifying the fourteen potential hosts used in the study. The taxa vary in
their native and introduced ranges and conservation status (see Supplemental taxa list for
detailed species information).

32
Methods
Oviposition experiment
The oviposition study took place at Fort DeSoto Park (hereafter FDS), Pinellas County,
Florida. Three distinct flight periods of C. cactorum occur at FDS: a spring flight from
February to May, a summer flight from June to August, and a fall flight from September
to November (Hight and Carpenter 2009 and pers. obsv.). Although specific densities of
C. cactorum at FDS are unknown, Baker and Stiling (2009) found that approximately
44% of Opuntia plants and 6% of cladodes at three west Florida coastal sites were
damaged by C. cactorum. No effort was made to either increase or control the naturally
occurring population of C. cactorum at FDS. Within the park, the native species Opuntia
stricta Haworth and O. humifusa Rafinesque are found in sand dunes, scrub, and
pinelands. In August of 2006, potted plants of fourteen common opuntioids were sunk
into the ground, with six individuals of each taxa placed in a randomized block design.
After the cacti had been in the field for approximately two weeks, censusing was begun.
During each census, all plants were inspected for eggsticks, which were then removed.
The number of eggsticks per plant and the number of eggs per eggstick were recorded.
Censusing occurred biweekly until August 2007, with the exception of December 2006
and January 2007 when plants were censused once per month. Plant height and cladode
number were recorded at the beginning and end of the experiment, and the mean values
were used in analyses. Epidermal toughness, measured as grams of pressure per mm2
needed to puncture the epidermal layer, was recorded at the end of the experiment using
an Effegi penetrometer (Model FT-011, International Ripening Company, Norfolk, VA).
Neither primary cladodes, i.e., highly lignified, “woody” cladodes, nor tertiary cladodes,

33
i.e., young cladodes with ephemeral true leaves, were used for toughness measurements,
as they were not available on all species and, in the case of primary cladodes, are not
typically eaten by C. cactorum larvae. Three secondary cladodes, i.e., cladodes that are
mature, but not woody, were randomly selected from each plant and three penetrometer
readings were taken per cladode. Starting from the point of attachment to the plant (or the
“joint” of the cladode), a proximal, medial, and distal reading were taken for a mean
toughness reading for that cladode. As C. cactorum females often oviposit on the tips of
cactus spines or on glochids at areoles where spines might appear, each opuntioid was
assigned a level for a categorical variable representing density of glochids and spines
(termed spininess). Levels for spininess were found by giving each taxa separate scores
for glochid and spine density (0=none to very sparse, 1=sparse, 2=moderate, 3=dense,
4=very dense), and then summing the two scores. Scores in each category were based
upon personal observation and on descriptions in Anderson (2001).
Statistical analyses were performed in STATISTICA 5.5 (StatSoft 1984-2000).
There were no significant effects of block, so data were pooled among blocks. Data for
proportions of total eggsticks (the sum of eggsticks laid on a plant over the course of the
year divided by the sum of eggsticks laid on all plants over the course of the year) were
highly non-normal, violated the homogeneity of variance assumption, and did not
respond to transformations. These data were therefore compared among taxa and levels
of spininess using a Kruskal-Wallis ANOVA by ranks (Kruskal and Wallis 1952).
Dunn‟s test for significant differences of mean rank (as described in Zar 1999) was used
for post hoc comparisons. Numbers of eggs per eggstick met assumptions of normality
and homogeneity of variance and were compared among host taxa and levels of spininess

34
using a single factor ANOVA; only those taxa that received two or more eggsticks were
used in these analyses. Epidermal toughness, plant height and number of cladodes were
log transformed to meet assumptions of homogeneity of variance. These variables were
then used in single factor ANOVAs, grouped by taxa, and in regressions of numbers of
eggs per eggstick. Means were separated by Bonferroni post-hoc tests. A generalized
linear model (StatSoft 2010) was used to determine if the plant characteristics of
toughness, cladode number, height, or the categorical variable spininess, could be used as
predictors of oviposition preference. Number of eggsticks was used as the Poisson
distributed response variable and a log link was used. Continuous variables used in the
generalized linear model were examined for tolerance and found to be non-redundant (all
tolerance values ≥0.83). A Type III log likelihood test was performed on the full model,
and all subsets of models were compared using Akaike‟s Information Criterion (AIC)
(Akaike 1974).
Larval performance experiment
The plants used in the oviposition experiment were removed from Fort DeSoto in
August 2007 and brought to University of South Florida Botanical Garden. On
September 5, 2007, freshly laid C. cactorum eggsticks were obtained from the mass
rearing facility located at the USDA-ARS Crop Protection and Management Research
Unit in Tifton, Georgia. Eggsticks containing between 40 and 60 eggs were placed upon
cladodes of each of the fourteen opuntioids used in the oviposition experiment and kept
in acrylic cages with ventilated lids (22x18x16cm). This experiment was designed to be

35
of the “no-choice” type, therefore only one eggstick and one type of cactus were placed
in each cage, with five individuals of each taxa arranged in a randomized block design.
Larvae were kept at ~25°C and a 14L:10D photoperiod. The cages were checked daily
and cladodes were replaced as needed. At the final instar, newspaper was placed in cages
for pupation media. Pupae were removed after completion of a cocoon and moved to
individual vials. Vials were checked daily for adult emergence and adults were placed in
a freezer overnight. Variables recorded were: initial number of eggs that hatched,
penetration success (into the cladode), number of penetration attempts, pupation rate,
adult emergence rate, adult sex ratio (proportion of males), adult weight, number of eggs
per female, and development times. Adults were sexed so that weight could be compared
separately for males and females.
Analyses were performed in Statistica 5.9 (StatSoft 1984-2000). Initial numbers
of larvae were not correlated with any fitness variables, and were therefore not included
as co-variates. To meet assumptions of normality and homogeneity of variance, pupation
and emergence rates were arcsine square root transformed, and weights and number of
eggs per female were either log- or square root-transformed. Durations of larval and
pupal stage were log transformed to meet assumptions of homogeneity of variance.
Pupation rate, emergence rate, weights, number of eggs per female, and development
times were compared among host taxa using a single factor ANOVA. Means were
separated by Bonferroni post-hoc tests. For development times and adult variables, only
host taxa in which two or more replicates produced adults were used. Proportions of
females were highly non-normal and did not respond to transformations; they were
compared among species using a Kruskal-Wallis ANOVA by ranks (Kruskal and Wallis

36
1952). To determine if epidermal toughness could predict larval fitness variables, least
squares linear regressions of pupation rate, emergence rate, adult weights, number of
eggs per female, and development times were performed.
Results
Oviposition experiment
Ranks of proportions of eggsticks were significantly different among all taxa (Kruskal-
Wallis H = 39.59, df = 13, p = 0.0002) and when taxa receiving zero eggsticks were
excluded (Kruskal-Wallis H = 20.42, df = 9, p = 0.0155). Ranks of proportions of
eggsticks were significantly different among all levels of spininess (Kruskal-Wallis H =
30.35, df = 7, p = 0.0001) and when levels of spininess that received zero eggsticks were
excluded (Kruskal-Wallis H = 11.34, df = 5, p = 0.0450). When comparing ranks of all
levels of spininess, level 0 was significantly different from levels 6 and 7 under Dunn‟s
test (Q = 3.69, Q0.05, 8 = 3.12). No other pair wise differences in ranks were found to be
significant. Opuntia engelmannii Salm-Dyck ex Engelmann var. linguiformis (Griffiths)
Parfitt and Pinkava received the highest proportion of eggsticks, followed by O.
engelmannii var. engelmannii Engelmann (Table 2.1). Four species [O. leucotricha de
Candolle, O. streptacantha, Cylindropuntia acanthocarpa (Engelmann and Bigelow)
F.M. Knuth, and C. spinosior (Engelmann) F.M. Knuth] received no eggsticks over the
course of the year. Opuntioids given a score of 5 for spininess received the most
eggsticks, followed by those with a score of 3. Those given scores of 6 or 7 did not
receive any eggsticks over the course of the year (Table 2.1).

37
Numbers of eggs per eggsticks were not significantly different between taxa (F7,
57 = 0.94, p = 0.49) or between levels of spininess (F5, 62 = 1.28, p = 0.29), although
unequal sample sizes (n = 2-17) reduced the power of the analysis. Numbers of eggs per
eggstick were highest for O. macrocentra Engelmann, followed by O. engelmannii var.
engelmannii, and lowest for Consolea rubescens (Salm-Dyck ex de Candolle) Lemaire
(Table 2.1). Host taxa differed significantly in epidermal toughness (Table 2.2; F13, 70 =
19.07, p < 0.0001), cladode number (Table 2.2; F13, 70 = 9.93, p < 0.0001), and height
(Table 2.2; F13, 70 = 17.10, p < 0.0001). Consolea rubescens was significantly different in
epidermal toughness from all other taxa except Nopalea cochenillifera (L.) Salm-Dyck.
Regression of number of eggs per eggsticks on plant characteristics did not show an
effect (r2
= 0.0008, 0.0041, and 0.0036; p = 0.82, 0.60, and 0.62, for toughness, cladode
number and height, respectively).
Dispersion statistics from the generalized linear model indicated that the model
was a good fit for the data (deviance/df = 0.9743, Pearson Χ2/df = 0.8809, log likelihood
= −76.6720). The Type III log likelihood test showed that number of cladodes and
spininess were significant predictors of number of eggsticks (Χ2
1 = 9.3699, p = 0.0022
and Χ2
7 = 71.8413, p < 0.0001, respectively), while height and epidermal toughness were
not. This was confirmed by using AIC to compare between all subsets of models. AIC
values ranged from 172.14 to 233.65; the model that included only the effects of cladode
number and spininess had the lowest AIC value (df = 8, p < 0.0001). The predicted mean
(±SE) number of eggsticks from this model was highest for a spininess score of 5,
followed by a score of 3 (1.097 ± 0.24 and 0.359 ± 0.22, respectively).

38
Larval performance experiment
Overall mean (±SE) pupation rates and adult emergence rates were low (0.22 ± 0.03 and
0.11 ± 0.02, respectively) and were significantly different between taxa (F13, 56 = 3.99, p
< 0.001 and F13, 56 = 3.97, p < 0.001, respectively). The highest pupation rate was found
on C. rubescens, followed by O. streptacantha (Table 2.3). Consolea rubescens also had
the highest adult emergence rate, followed by O. dillenii (Ker Gawler) Haworth (Table
2.3). Opuntia macrocentra Engelmann had zero percent pupation and O. engelmannii
var. lindheimeri (Engelmann) Parfitt and Pinkava and C. spinosior had emergence rates
of less than one percent (Table 2.3). Opuntia macrocentra had significantly different
pupation and emergence rates from C. rubescens, O. ficus-indica, and O. dillenii.
Consolea rubescens had significantly different emergence rates from C. spinosior, O.
engelmannii var. lindheimeri, O. microdasys, and O. santa-rita.
Overall mean (±SE) development time (hatch to adult emergence) was 75.9 ± 1.3
days. Overall mean (±SE) durations of the larval and pupal stages were 54.7 ± 1.3 days
and 21.6 ± 1.0 days. Total development time and duration of the larval stage were
significantly different among taxa (F10, 26 = 2.35, p = 0.039 and F10, 26 = 9.165, p < 0.001,
respectively). Total development time was least on O. microdasys (Lehmann) Pfeiffer
(although only 2% of larvae survived to adult emergence on this species), followed by C.
rubescens, and greatest on O. engelmannii var. lindheimeri (Table 2.3). Opuntia
engelmanni var. lindheimeri was significantly different from O. dillenii and C. rubescens
for total development time, and was significantly different from all taxa except the other
two O. engelmanni varieties and C. acanthocarpa for duration of the larval stage.

39
The overall mean (±SE) proportion of females was low (0.28 ± 0.04) and was not
significantly different among taxa (Kruskal-Wallis H=13.45, df = 9, p = 0.143). Opuntia
microdasys, O. engelmannii var. lindheimeri, and C. acanthocarpa produced only males
(Table 2.3). Male and female weights were significantly different among taxa (F12, 214 =
5.78, p < 0.0001 and F9, 150 = 2.42, p = 0.014, respectively), but number of eggs per
female was not different among taxa (F9, 150 = 1.64, p = 0.109). Number of eggs was
significantly correlated with female weight (rS = 0.52, p < 0.0001), but was not
significantly correlated with development times. Consolea rubescens had the heaviest
females and the highest numbers of eggs per female, followed by O. streptacantha
(Table 4). Opuntia santa-rita (Griffiths and Hare) Rose had both the lightest females and
the lightest males (Table 2.4).
Regressions of pupation rate, emergence rate, number of eggs per female,
duration of larval stage, and total development time on epidermal toughness were
significant, but r2 values were low overall, indicating that untested factors have
considerable influence on the response variables (Table 2.5).

40
Table 2.1. Eggstick and spine data for the fourteen opuntioid taxa. Columns show proportion of eggsticks, mean number of
eggsticks (± SE), mean number of eggs per eggstick (± SE), glochid and spine scores and summed spininess score
Taxa
Proportion
of
eggsticks
Number of
eggsticksa
Number of eggs per
eggstick (n)b
Glochid
score
Spine
score
Spininess
scorec
Consolea rubescens 0.0294 0.33 ± 0.21 36.0 ± 3.0 (2) 1 0 1
Cylindropuntia
acanthocarpa 0 0 -- 3 4 7
Cylindropuntia spinosior 0 0 -- 3 3 6
Nopalea cochenillifera 0.0735 0.83 ± 0.31 49.4 ± 6.8 (5) 0 0 0
Opuntia dillenii 0.1471 1.67 ± 0.33 47.7 ± 4.5 (10) 3 0 3
Opuntia engelmannii var.
engelmannii 0.1618 1.83 ± 0.65 57.5 ± 7.0 (11) 2 2 4
Opuntia engelmannii var.
lindheimeri 0.0147 0.17 ± 0.17 -- 2 2 4
Opuntia engelmannii var.
linguiformis 0.2500 2.83 ± 0.79 48.0 ± 3.3 (17) 2 3 5
Opuntia ficus-indica 0.0588 0.67 ± 0.49 56.0 ± 10.7 (4) 2 0 2
Opuntia leucotricha 0 0 -- 3 4 7
Opuntia macrocentra 0.1176 1.33 ± 0.49 60.5 ± 7.4 (8) 2 1 3

41
Taxa
Proportion
of
eggsticks
Number of
eggsticksa
Number of eggs per
eggstick (n)b
Glochid
score
Spine
score
Spininess
scorec
Opuntia microdasys 0.0147 0.17 ± 0.17 -- 4 0 4
Opuntia santa-rita 0.1324 1.5 ± 0.50 53.9 ± 4.7 (9) 3 1 4
Opuntia streptacantha 0 0 -- 2 4 6
a
n = 6 for all taxa. b Mean number of eggs per eggstick is shown only for those taxa receiving ≥2 eggsticks.
c The categorical variable spininess, used in a generalized linear model to predict oviposition by C. cactorum, is the sum of a
particular taxa‟s glochid density score and its spine density score. Scores were assigned prior to the experiment and were based
upon descriptions in Anderson (2001) and personal observation.

42
Table 2.2 Mean values (± SE) for recorded plant characteristics
Species/variety Height
(cm)a
Number of
cladodesa
Epidermal toughness
(g/mm2)a
Consolea rubescens 46.8 ± 3.6 7.2 ± 1.7 330.40 ± 16.41
Cylindropuntia acanthocarpa 59.6 ± 2.1 17.3 ± 2.6 583.53 ± 40.49
Cylindropuntia spinosior 69.8 ± 8.2 15.4 ± 2.4 675.33 ± 65.09
Nopalea cochenillifera 34.4 ± 3.7 9.8 ± 1.6 417.60 ± 12.52
Opuntia dillenii 32.1 ± 2.2 9.1 ± 0.9 536.40 ± 29.27
Opuntia engelmannii var.
engelmannii 49.7 ± 2.8 10.1 ± 1.1 787.2 ± 32.29
Opuntia engelmannii var.
lindheimeri 40.0 ± 1.4 3.6 ± 0.6 979.07 ± 38.57
Opuntia engelmannii var.
linguiformis 71.3 ± 5.3 8.5 ±1.1 732.33 ± 36.26
Opuntia ficus-indica 59.3 ± 4.7 6.3 ± 0.7 709.8 ± 15.17
Opuntia leucotricha 52.0 ± 4.5 8.5 ± 0.9 596.73 ± 25.87
Opuntia macrocentra 25.7 ± 1.4 2.9 ± 0.3 959.80 ± 72.31
Opuntia microdasys 24.0 ± 2.1 6.8 ± 1.7 535.73 ± 23.12
Opuntia santa-rita 40.2 ± 6.1 10.6 ± 1.6 775.40 ± 97.19
Opuntia streptacantha 86.0 ± 3.7 7.3 ± 0.6 685.27 ± 58.44
a n = 6 for all taxa
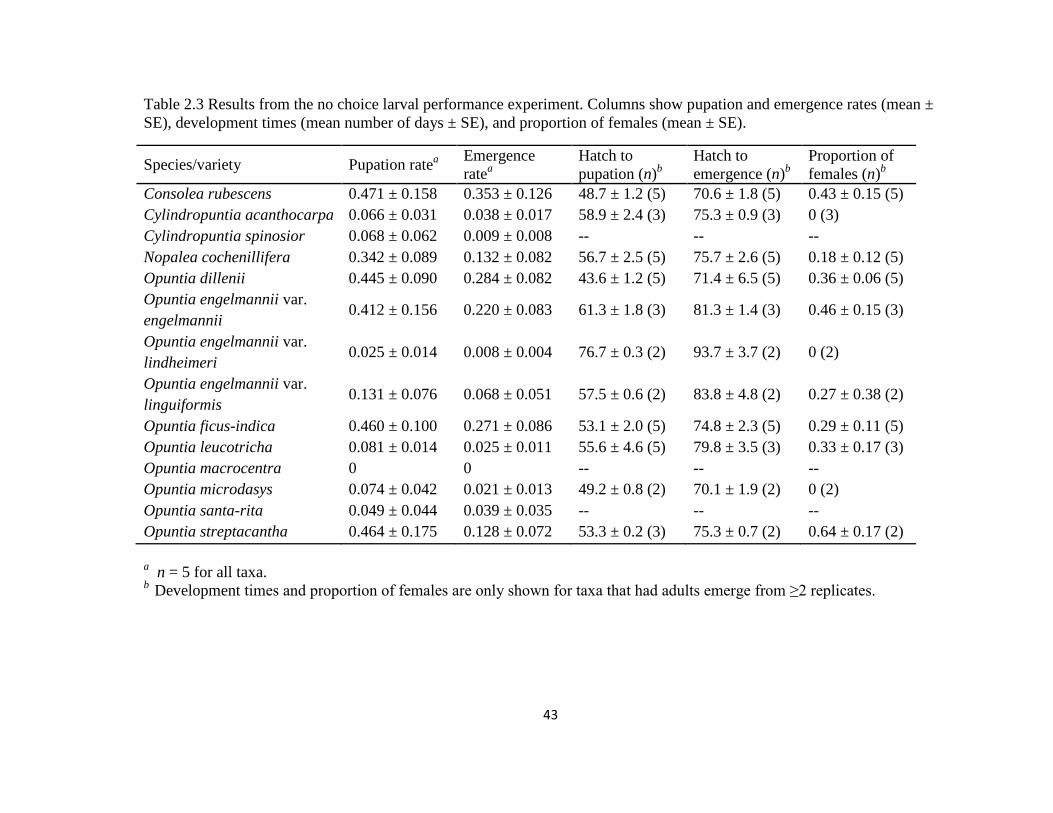
43
Table 2.3 Results from the no choice larval performance experiment. Columns show pupation and emergence rates (mean ±
SE), development times (mean number of days ± SE), and proportion of females (mean ± SE).
Species/variety Pupation ratea
Emergence
ratea
Hatch to
pupation (n)b
Hatch to
emergence (n)b
Proportion of
females (n)b
Consolea rubescens 0.471 ± 0.158 0.353 ± 0.126 48.7 ± 1.2 (5) 70.6 ± 1.8 (5) 0.43 ± 0.15 (5)
Cylindropuntia acanthocarpa 0.066 ± 0.031 0.038 ± 0.017 58.9 ± 2.4 (3) 75.3 ± 0.9 (3) 0 (3)
Cylindropuntia spinosior 0.068 ± 0.062 0.009 ± 0.008 -- -- --
Nopalea cochenillifera 0.342 ± 0.089 0.132 ± 0.082 56.7 ± 2.5 (5) 75.7 ± 2.6 (5) 0.18 ± 0.12 (5)
Opuntia dillenii 0.445 ± 0.090 0.284 ± 0.082 43.6 ± 1.2 (5) 71.4 ± 6.5 (5) 0.36 ± 0.06 (5)
Opuntia engelmannii var.
engelmannii 0.412 ± 0.156 0.220 ± 0.083 61.3 ± 1.8 (3) 81.3 ± 1.4 (3) 0.46 ± 0.15 (3)
Opuntia engelmannii var.
lindheimeri 0.025 ± 0.014 0.008 ± 0.004 76.7 ± 0.3 (2) 93.7 ± 3.7 (2) 0 (2)
Opuntia engelmannii var.
linguiformis 0.131 ± 0.076 0.068 ± 0.051 57.5 ± 0.6 (2) 83.8 ± 4.8 (2) 0.27 ± 0.38 (2)
Opuntia ficus-indica 0.460 ± 0.100 0.271 ± 0.086 53.1 ± 2.0 (5) 74.8 ± 2.3 (5) 0.29 ± 0.11 (5)
Opuntia leucotricha 0.081 ± 0.014 0.025 ± 0.011 55.6 ± 4.6 (5) 79.8 ± 3.5 (3) 0.33 ± 0.17 (3)
Opuntia macrocentra 0 0 -- -- --
Opuntia microdasys 0.074 ± 0.042 0.021 ± 0.013 49.2 ± 0.8 (2) 70.1 ± 1.9 (2) 0 (2)
Opuntia santa-rita 0.049 ± 0.044 0.039 ± 0.035 -- -- --
Opuntia streptacantha 0.464 ± 0.175 0.128 ± 0.072 53.3 ± 0.2 (3) 75.3 ± 0.7 (2) 0.64 ± 0.17 (2)
a n = 5 for all taxa.
b Development times and proportion of females are only shown for taxa that had adults emerge from ≥2 replicates.

44
Table 2.4 Adult weights and number of eggs per female (mean ± SE)
Species/variety Male weights
(mg) (n)a
Females weights
(mg) (n)a
Number of eggs per
female (n)a
Consolea rubescens 19.58 ± 1.11 (45) 46.1 ± 3.1 (41) 60.8 ± 3.9 (41)
Cylindropuntia acanthocarpa 10.09 ± 1.38 (9) -- --
Cylindropuntia spinosior 14.00 ± 4.12 (5) 34.2 ± 11.3 (3) 30.0 ± 10.6 (3)
Nopalea cochenillifera 15.06 ± 2.61 (14) 32.3 ± 4.4 (13) 43.7 ± 5.3 (13)
Opuntia dillenii 19.27 ± 1.24 (39) 43.5 ± 3.0 (28) 51.7 ± 4.2 (28)
Opuntia engelmannii var.
engelmannii 16.18 ± 1.09 (30) 37.2 ± 3.9 (23) 44.7 ± 4.4 (23)
Opuntia engelmannii var.
lindheimeri 22.00 ± 5.70 (2) -- --
Opuntia engelmannii var.
linguiformis 32.48 ± 5.64 (9) 28.6 ± 5.7 (8) 42.6 ± 6.3 (8)
Opuntia ficus-indica 15.12 ± 0.81 (42) 40.9 ± 4.3 (17) 54.1 ± 7.6 (17)
Opuntia leucotricha 30.72 ± 11.81 (6) 44.9 ± 11.8 (3) 43.3 ± 8.7 (3)
Opuntia macrocentra -- -- --
Opuntia microdasys 16.85 ± 1.83 (6) -- --
Opuntia santa-rita 10.06 ± 1.40 (8) 21.9 ± 2.5 (3) 43.3 ± 1.3 (3)
Opuntia streptacantha 20.08 ± 2.69 (12) 21.9 ± 2.5 (21) 59.6 ± 6.8 (21)
a
Data were pooled among the six replicates. Means are only shown for those taxa that
produced ≥2 adults of the respective gender.

45
Table 2.5 Results from regressions of larval performance variables on epidermal
toughness of the fourteen tested opuntioids
Variable Coefficient r2 F
Pupation rate -0.0008 ± 0.0002 0.1665 13.58***
Emergence rate -0.0005 ± 0.0001 0.1379 10.87**
Duration of larval stage 0.0194 ± 0.0064 0.1983 9.15**
Duration of pupal stage 0.0024 ± 0.0055 0.0051 0.19ns
Total development time 0.0218 ± 0.0062 0.2519 12.45**
Number of eggs -0.0023 ± 0.0007 0.0608 10.22**
Female weights -0.0009 ± 0.0006 0.0142 2.27ns
Male weights -0.0002 ± 0.0002 0.0094 2.12ns
** Significant at p < 0.01.
*** Significant at p < 0.001
ns Non-significant

46
Discussion
Plant structure, both in the form of spine density and cladode number, had a clear effect
on oviposition preference of female C. cactorum moths on the North American
opuntioids tested here. Taxa with intermediate spininess ratings (3-5 on a scale of 0-7)
received the most eggsticks. Females may prefer to “camouflage” their eggsticks on the
tips of spines, rather than lay them directly on the cladode. At the same time, too many
spines and glochids interfere with the process of landing, probing the surface of the
cladode, and oviposition (Stange 1997). Plants with moderately high numbers of
cladodes received more eggsticks than those with fewer cladodes or very high numbers
of cladodes. However, cladode number was not separated from true biomass, or
controlled for within species, so our conclusions about the effect of plant size on
oviposition behavior are tentative. Past studies have shown an inconsistent response to
plant size by C. cactorum females (Myers et al. 1981, Robertson 1987, Johnson and
Stiling 1998, Tate et al. 2009). In South Africa, females have been shown to prefer
oviposition sites that are sheltered from wind (Robertson 1987), so some of the cladode
effect found in our study could be due to the fact that higher numbers of cladodes offer
sites that are better sheltered from wind. However, we would also therefore expect taxa
with the most cladodes to have the highest number of eggsticks, which we did not find.
This inconsistency was most likely driven by the two cylindropuntia species, which had
more cladodes than the platyopuntia species used in the study. Clearly, oviposition
preferences cannot be explained by external plant characteristics alone. It is likely that
volatile organic compounds (VOCs) play a large role in determining interspecific
oviposition preferences of C. cactorum, as female moths have been shown to respond to
several VOCs, including nonanal, an alkyl aldehyde produced by O. stricta (Pophof et

47
al. 2005). More work is needed to identify the exact compounds to which C. cactorum
females are attracted.
Epidermal toughness was a good predictor of most larval fitness parameters, and
it is interesting to note that the two toughest hosts, O. engelmannii var. lindheimeri and
O. macrocentra, were also the two species with the lowest pupation and emergence rates.
Although pre-penetration mortality was not quantified, larvae on these two species often
failed to penetrate the cladode, and perished after getting stuck in the mucilage secreted
from their attempts. It seems likely that the apparently thicker mucilage, along with
epidermal toughness, contributed to the low pupation rates found on O. engelmannii var.
lindheimeri and O. macrocentra.
This study confirms the finding of past studies that members of the genus
Cylindropuntia are likely to be at less risk of C. cactorum attack than other opuntioid
species (Dodd 1940, Zimmermann et al. 2004). Neither C. acanthocarpa nor C.
spinosior received any eggsticks, and both proved to be submarginal hosts. However,
overall, our findings did not support the idea that ovipositing females prefer host species
that maximize larval fitness. The two taxa most preferred by females, O. engelmannii
vars. linguiformis and engelmannii, were not superior hosts for larvae. Out of the
fourteen opuntioids, they ranked seventh and fourth, respectively, for percent emergence
and had the second and third longest development times. Additionally, O. streptacantha
received no eggsticks over the course of a year in the field, yet ranked second for
pupation rate, female weights, and number of eggs, and had the highest proportion of
emerging females in the laboratory. Of the opuntioids tested, the best overall host for
larvae appeared to be C. rubescens, although it received less than half of the expected

48
proportion of eggsticks laid in the field (3.1 ± 0.02% observed versus 7.14% expected if
eggsticks were distributed evenly among the tested species).
Cactoblastis cactorum neonates generally enter the cladode upon which the
eggstick was laid, and therefore larval host choice in a natural setting is dictated by
ovipositional behavior. However, in no choice experiments, larvae have been shown to
develop normally on some species that are not selected by females (Dodd 1940), and
females have been shown to prefer species that do not maximize fitness. For example,
Johnson and Stiling (1996) found that when given a choice of Florida cactus species,
larval preference seemed to reflect ovipositional preference; both groups preferred
Consolea corallicola over three other native Florida species. However, survivorship was
significantly lower on C. corallicola than on the less preferred hosts O. stricta and O.
humifusa. Mafokoane et al. (2007) found a similar disparity between preference and
performance; C. cactorum larvae on O. imbricata had the lowest survivorship, yet the
second highest number of eggsticks out of the six species tested.
It is often shown that explicit tests of the preference-performance hypothesis do
not support its expectation (Thompson 1988a, Mayhew 1998, Forister 2004, Forister et
al. 2009), and a variety of reasons have been proposed to explain this lack of support.
These include mechanistic and phylogenetic constraints (Courtney and Kibota 1990,
Jaenike 1990, Mayhew 1997), the exclusion of the effects of adult behavior and natural
enemies from the models (Thompson 1988b, Scheirs et al. 2000, Ballabeni et al. 2001a,
Forister et al. 2009), a lack of distinction between host plants that provide protection
versus those that provide nutrition (Ballabeni et al. 2001b), and lack of sufficient time for
adaptation to have taken place (Mayhew 1997, Mayhew 2001, Forister et al. 2009).

49
Cactoblastis cactorum adults do not feed, and the larvae feed internally, therefore adult
behavior and natural enemies of larvae are not predicted to be influential factors of
oviposition behavior. A more likely explanation for the discrepancies between preference
and performance is the lack of a co-evolutionary history between C. cactorum and its
ever-growing assemblage of “new association” hosts. It seems that when encountering
novel hosts, C. cactorum mothers do not always “know best” (Courtney and Kibota
1990). In the case of C. cactorum’s potential invasion of the southwestern United States
and Mexico, we can only draw tentative conclusions about the opuntioids that will be at
highest risk, namely O. streptacantha, in the event that larval fitness drives oviposition
preference, and O. engelmannii vars. engelmannii and linguiformis, in the event that
oviposition preference drives larval adaptation. The best overall host from our study, C.
rubescens, is native to the Caribbean and West Indies, where C. cactorum is known to
kill seedlings and small plants (Zimmermann et al 2005). It is not native to the
southwestern United States or Mexico, but is naturalized in some areas, including
Arizona, Nevada, and California. Although clearly not a conservation priority,
naturalized populations of C. rubescens in the southwest could help sustain C. cactorum
populations in areas with other, native hosts. The second best host from our study, O.
streptacantha, is native to northern and central Mexico and is widely cultivated for its
fruits (tunas) and cladodes (nopales) (Zimmermann et al 2001). Although ignored by
ovipositing females in this study, O. streptacantha is a known host of C. cactorum and
did receive C. cactorum eggsticks in recent caged choice trials of three different groups
of cacti (Tate et al. 2009). Additionally, the superior performance of C. cactorum larvae
on O. streptacantha raises concerns about the moth becoming an agricultural pest in

50
Mexico. The opuntioid most preferred by females, O. engelmannii var. linguiformis, is
reportedly native to Texas (Anderson 2001) and is very rare in the wild. It is cultivated in
the United States for use in landscaping because of its uniquely shaped pads. There are
therefore both conservation and economic risks posed to this species. Although O.
engelmannii var. linguiformis had very slow development of larvae and a low proportion
of females, the fact that there are so few individuals in the wild suggests they could be
particularly at risk. For now, it is our hope that efforts to control C. cactorum will be
successful, preventing both predicted and unforeseen consequences of its range
expansion to the opuntioid-rich west.
Acknowledgements
We gratefully thank Susan Drawdy (USDA-ARS, Crop Protection and Management
Unit, Tifton, GA) for help in acquiring eggsticks for the larval performance experiment,
Amanda Baker and Mountain States Wholesale Nursery, Litchfield Park, AZ, for
providing cactus plants, and Rachael Vistein and Leah Pope for their assistance during
the larval experiment.
References Cited
Akaike, H. 1974. New look at statistical model identification. IEEE Trans Automat
Contr. AC19: 716-723.
Anderson, E. F. 2001. The cactus family. Timber Press, Portland, OR.
Baker, A. J., and P. Stiling. 2009. Comparing the effects of the exotic cactus-feeding
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae) and the native
cactus-feeding moth, Melitara prodenialis (Walker) (Lepidoptera: Pyralidae) on
two species of Florida Opuntia. Biol. Invasions.11: 619-624.

51
Ballabeni, P., M. Wlodarczyk, and M. Rahier. 2001a. Does enemy-free space for eggs
contribute to a leaf beetle's oviposition preference for a nutritionally inferior host
plant? Funct. Ecol. 15: 318-324.
Ballabeni, P., D. Conconi, S. Gateff, and M. Rahier. 2001b. Spatial proximity between
two host plant species influences oviposition and larval distribution in a leaf
beetle. Oikos 92: 225-234.
Bennett, F. D., D. H. Habeck, E. S. Delfosse, and R. R. Scott. 1995. Cactoblastis
cactorum: A successful weed control agent in the Caribbean, now a pest in
Florida? Biological control of weeds, pp. 21-26. In E.S. Delfosse and R.R. Scott
(eds.), Proceedings, VIII International Symposium on Biological Control of
Weeds, 2-7 February 1992, Lincoln University, Canterbury, New Zealand.
CSIRO Publications, East Melbourne, Victoria, Australia.
Carpenter, J. E., S. Bloem, and K. A. Bloem. 2001a. Inherited sterility in Cactoblastis
cactorum (Lepidoptera: Pyralidae). Fla. Entomol. 84: 537-542.
Carpenter, J. E., K. A. Bloem, and S. Bloem. 2001b. Applications of F-1 sterility for
research and management of Cactoblastis cactorum (Lepidoptera: Pyralidae). Fla.
Entomol. 84: 531-536.
Chavez-Ramirez, F., X. G. Wang, K. Jones, D. Hewitt, and P. Felker. 1997. Ecological
characterization of Opuntia clones in South Texas: implications for wildlife
herbivory and frugivory. J.Prof. Assoc. Cactus. 2: 9-19.
Courtney, S. P., and T. T. Kibota. 1990. Mother doesn't know best: selection of hosts by
ovipositing insects, pp. 161-188. In E. A. Bernays (ed.), Insect-plant interactions,
vol. 2. CRC Press, Inc., Boca Raton, FL.
Dickel, T. S. 1991. Cactoblastis cactorum in Florida. (Lepidoptera: Pyralidae:
Phycitinae). Trop. Lepid. 2: 117-118.
Dodd, A. P. 1940. The biological campaign against prickly pear. Commonwealth Prickly
Pear Board Bulletin:1-177. Brisbane, Australia.
Forister, M. L. 2004. Oviposition preference and larval performance within a diverging
lineage of lycaenid butterflies. Ecol. Entomol. 29: 264-272.
Forister, M. L., C. C. Nice, J. A. Fordyce, and Z. Gompert. 2009. Host range evolution is
not driven by the optimization of larval performance: the case of Lycaeides
melissa (Lepidoptera: Lycaenidae) and the colonization of alfalfa. Oecologia.
160: 551-561.
Habeck, D. H., and F. D. Bennett. 1990. Cactoblastis cactorum Berg (Lepidoptera:
Pyralidae), a phycitine new to Florida. Florida Department of Agriculture and

52
Consumer Services, Division of Plant Industry Entomology Circular, Gainesville,
FL.
Hajek, A. E., L. Butler, S. R. A. Walsh, J. C. Silver, F. P. Hain, F. L. Hastings, T. M.
Odell, and D. R. Smitley. 1996. Host range of the gypsy moth (Lepidoptera:
Lymantriidae) pathogen Entomophaga maimaiga (Zygomycetes:
Entomophthorales) in the field versus laboratory. Environ. Entomol. 25: 709-721.
Hervert, J. J., J. L. Bright, R. S. Henry, L. A. Piest, and M. T. Brown. 2005. Home-range
and habitat-use patterns of Sonoran pronghorn in Arizona. Wildl. Soc. Bull. 33: 1.
Hight, S. D., and J. E. Carpenter. 2009. Flight phenology of male Cactoblastis cactorum
(Lepidoptera: Pyralidae) at different latitudes in the southeastern United States.
Fla. Entomol. 92: 208-216.
Hight, S. D., J. E. Carpenter, S. Bloem, and K. A. Bloem. 2005. Developing a sterile
insect release program for Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae):
effective overflooding ratios and release-recapture field studies. Environ.
Entomol. 34: 850-856.
Hoffmann, J. H., V. C. Moran, and D. A. Zeller. 1998. Evaluation of Cactoblastis
cactorum (Lepidoptera: Phycitidae) as a biological control agent of Opuntia
stricta (Cactaceae) in the Kruger National Park, South Africa. Biol.Control. 12:
20-24.
Jaenike, J. 1978. Optimal oviposition behavior in phytophagous insects. Theor. Popul.
Biol. 14: 350-356.
Jaenike, J. 1990. Host specialization in phytophagous insects. Annu. Rev. Ecol. Syst. 21:
243-273.
Johnson, D. M., and P. D. Stiling. 1996. Host specificity of Cactoblastis cactorum
(Lepidoptera: Pyralidae), an exotic Opuntia-feeding moth, in Florida. Environ.
Entomol. 25: 743-748.
Johnson, D. M., and P. D. Stiling. 1998. Distribution and dispersal of Cactoblastis
cactorum (Lepidoptera: Pyralidae), an exotic Opuntia-feeding moth, in Florida.
Fla. Entomol. 81: 12-22.
Kruskal, W. H., and W. A. Wallis. 1952. Use of ranks in one-criterion variance analysis.
J. Am. Stat. Assoc. 47: 583-621.
Levins, R., and R. Macarthur. 1969. An hypothesis to explain the incidence of
monophagy. Ecology. 50: 910-911.

53
Louda, S. M., D. Kendall, J. Connor, and D. Simberloff. 1997. Ecological effects of an
insect introduced for the biological control of weeds. Science 277: 1088-1090.
Louda, S. M., R. W. Pemberton, M. T. Johnson, and P. A. Follett. 2003. Nontarget
effects - The Achilles' Heel of biological control? Retrospective analyses to
reduce risk associated with biocontrol introductions. Annu. Rev. Entomol. 48:
365-396.
Mafokoane, L. D., H. G. Zimmermann, and M. P. Hill. 2007. Development of
Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae) on six North American
Opuntia species. Afr. Entomol. 15: 295-299.
Mann, J. 1969. Cactus-feeding insects and mites. Smithsonian Institution, Washington,
D.C.
Marti, O. G., and J. E. Carpenter. 2009. Effect of irradiation on the incidence of mating
in Cactoblastis cactorum. Fla. Entomol. 92: 159-160.
Mayhew, P. J. 1997. Adaptive patterns of host-plant selection by phytophagous insects.
Oikos 79: 417-428.
Mayhew, P. J. 1998. Testing the preference-performance hypothesis in phytophagous
insects: lessons from chrysanthemum leafminer (Diptera: Agromyzidae).
Environ. Entomol. 27: 45-52.
Mayhew, P. J. 2001. Herbivore host choice and optimal bad motherhood. Trends Ecol.
Evol. 16: 165-167.
Myers, J. H., J. Monro, and N. Murray. 1981. Egg clumping, host plant selection and
population regulation in Cactoblastis cactorum (Lepidoptera). Oecologia 51: 7-
13.
Perez-Sandi, M. 2001. Addressing the threat of Cactoblastis cactorum (Lepidoptera:
Pyralidae), to Opuntia in Mexico. Fla. Entomol. 84: 499-502.
Pettey, F. W. 1948. The biological control of prickly-pear in South Africa. Sci. Bull.
Dept. Agric. Union South Africa. 271: 1-163.
Pophof, B., G. Stange, and L. Abrell. 2005. Volatile organic compounds as signals in a
plant-herbivore system: electrophysiological responses in olfactory sensilla of the
moth Cactoblastis cactorum. Chem. Senses 30: 51-68.
Robertson, H. G. 1987. Oviposition site selection in Cactoblastis cactorum
(Lepidoptera): constraints and compromises. Oecologia. 73: 601-608.

54
Scheirs, J., L. De Bruyn, and R. Verhagen. 2000. Optimization of adult performance
determines host choice in a grass miner. Proc. R. Soc. Lond., Ser. B: Biol. Sci.
267: 2065-2069.
Simonson, S. E., T. J. Stohlgren, L. Tyler, W. Gregg, R. Muir, and L. Garrett. 2005.
Preliminary assessment of the potential impacts and risks of the invasive cactus
moth, Cactoblastis cactorum Berg, in the U.S. and Mexico. Final Report to the
International Atomic Energy Agency, Vienna, Austria.
Soberon, J., J. Golubov, and J. Sarukhan. 2001. The importance of Opuntia in Mexico
and routes of invasion and impact of Cactoblastis cactorum (Lepidoptera:
Pyralidae). Fla. Entomol. 84: 486-492.
Stange, G. 1997. Effects of changes in atmospheric carbon dioxide on the location of
hosts by the moth, Cactoblastis cactorum. Oecologia. 110: 539-545.
StatSoft, Inc. 1995. STATISTICA for Windows. StatSoft Incorportated, Tulsa, OK.
StatSoft, Inc. 2010. Electronic Statistics Textbook. StatSoft Incorporated, Tulsa, OK.
Stiling, P. 2002. Potential non-target effects of a biological control agent, prickly pear
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), in North America,
and possible management actions. Biol. Invasions. 4: 273-281.
Stiling, P. 2010. Death and decline of a rare cactus in Florida. Castanea.75: 190-197.
Stiling, P., D. Moon, and D. Gordon. 2004. Endangered cactus restoration: mitigating the
non-target effects of a biological control agent (Cactoblastis cactorum) in
Florida. Restor. Ecol. 12: 605-610.
Strittmatter, L. I., V. Negron-Ortiz, and R. J. Hickey. 2006. Comparative
microsporangium development in male-fertile and male-sterile flowers of
Consolea (Cactaceae): when and how does pollen abortion occur? Grana 45: 81-
100.
Tate, C. D., J. E. Carpenter, and S. Bloem. 2007. Influence of radiation dose on the level
of F-1 sterility in the cactus moth, Cactoblastis cactorum (Lepidoptera:
Pyralidae). Fla. Entomol. 90: 537-544.
Tate, C. D., S. D. Hight, and J. E. Carpenter. 2009. Oviposition preference of
Cactoblastis cactorum (Lepidoptera: Pyralidae) in caged choice experiments and
the influence on risk assessment of F-1 sterility. Biocontrol Sci. Technol. 19: 317-
333.

55
Thompson, J. N. 1988a. Evolutionary ecology of the relationship between oviposition
preference and performance of offspring in phytophagous insects. Entomol. Exp.
Appl. 47: 3-14.
Thompson, J. N. 1988b. Coevolution and alternative hypotheses on insect plant
interactions. Ecology. 69: 893-895.
USDA-APHIS-PPQ. 2007. Cactoblastis cactorum activities report for Jan-Feb-Mar
2007. United States Department of Agriculture, Animal and Plant Health
Inspection Service, Plant Protection and Quarantine.
USDA-APHIS-PPQ. 2009. Cactus moth program annual report, Eastern region. United
States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine.
USDA-ARS, National Genetic Resources Program. Germplasm Resources Information
Network - (GRIN) [Online Database].National Germplasm Resources Laboratory,
Beltsville, Maryland.URL: http://www.ars-grin.gov/cgi-bin/npgs/html/taxweed.pl
(01 October 2011)
Vigueras, A. L., and L. Portillo. 2001. Uses of Opuntia species and the potential impact
of Cactoblastis cactorum (Lepidoptera: Pyralidae) in Mexico. Fla. Entomol. 84:
493-498.
Zar, J. H. 1999. Biostatistical analysis. Prentice Hall, Upper Saddle River, N.J.
Zimmermann, H. G., V. C. Moran, and J. H. Hoffmann. 2000. The renowned cactus
moth, Cactoblastis cactorum (Lepidoptera: Pyralidae): Its natural history and
threat to native Opuntia floras in Mexico and the United States of America. Fla.
Entomol. 84: 543-551.
Zimmermann, H. G., S. Bloem, and H. Klein. 2004. Biology, history, threat, surveillance
and control of the cactus moth, Cactoblastis cactorum. Joint FAO/IAEA Division
of Nuclear Techniques in Food and Agriculture. International Atomic Energy
Agency, Vienna, Austria.
Zimmermann, H. G., M. P. Cuen, and A. Rivera. 2005. The status of Cactoblastis
cactorum (Lepidoptera: Pyralidae) in the Caribbean and the likelihood of its
spread to Mexico. Report to the International Atomic Agency, Joint FAO/IAEA
Division of Nuclear Techniques in Food and Agriculture and the Plant Health
General Directorate, Mexico (DGSVB/SAGARPA) as part of the TC Project
MEX/5/029. International Atomic Energy Agency, Vienna, Austria.

56
CHAPTER 3
ASSOCIATIONAL EFFECTS OF CHAMAECRISTA FASCICULATA ON ANT
PREDATION OF CACTOBLASTIS CACTORUM AND HERBIVORY OF OPUNTIA
Synopsis
The legume Chamaecrista fasciculata attracts ants to its extrafloral nectar (EFN) which
can lead to reduced herbivory and increased fecundity for the plant. In Florida, Opuntia
stricta and O. humifusa, hosts of the invasive moth Cactoblastis cactorum, are often
found growing in close association with C. fasciculata. We tested the hypothesis that O.
stricta and O. humifusa individuals have higher ant abundance, lower levels of herbivore
damage, and increased growth when growing in close association with C. fasciculata
compared with individuals not growing near the plant. We also experimentally placed C.
cactorum eggsticks and pupae on Opuntia individuals to see if ant predation of these
stages occurred, and if so, whether predation rates were higher on individuals growing
close to C. fasciculata. Opuntia plants near C. fasciculata were less likely to be attacked
by C. cactorum and had higher ant abundance than plants far from C. fasciculata. Field
surveys also showed that Opuntia plants near C. fasciculata had a lower proportion of
cladodes with C. cactorum damage of any type. Proportions of cladodes with damage
from five native herbivores were not significantly different between treatments. In
addition, Opuntia individuals growing near C. fasciculata added proportionately more

57
cladodes during the growing season. We found evidence of ant predation on 15.9% of C.
cactorum eggsticks and 17.6% of pupae. In August and October of 2008, there was
significantly more evidence of predation on eggs and pupae placed on Opuntia
individuals near C. fasciculata. No effect of distance to C. fasciculata was seen in
November of 2008, potentially because plants were no longer producing EFN at this
time. Our finding that Opuntia plants close to C. fasciculata show reduced herbivory
from invasive C. cactorum, but not from the native herbivores examined, suggests that
patterns of associational resistance may be influenced by the co-evolutionary history of
the organisms in question.
Introduction
The susceptibility of a plant to herbivory can be affected by the characteristics of its plant
neighbors. Neighboring plants can increase or decrease the probability of a target plant
being detected and consumed by an herbivore. The former phenomenon is termed
associational susceptibility (hereafter, AS) (Brown and Ewel 1987; Wahl and Hay 1995),
the latter associational resistance (hereafter, AR) or associational defense (Atsatt and
Odowd 1976; Hay 1986; Tahvanainen and Root 1972). AS and AR have been
demonstrated in both terrestrial (Brown and Ewel 1987; Finch et al. 2003; Hamback et
al. 2000; Karban 1997; Russell et al. 2007; Stenberg et al. 2007; Thomas 1986; White
and Whitham 2000) and marine (Gagnon et al. 2003; Hay 1986; Pfister and Hay 1988;
Wahl and Hay 1995) habitats. Several mechanisms have been invoked to explain these
plant-plant interactions, including the repellant-masking plant hypothesis (Atsatt and
Odowd 1976; Tahvanainen and Root 1972), the attractant-decoy plant hypothesis (Atsatt

58
and Odowd 1976), the resource concentration hypothesis (Root 1973), and the enemies
hypothesis (Root 1973). The concepts of AR and AS have also been shown among
herbivore species (sometimes termed apparent competition), where they are often
mediated by shared natural enemies (Barbosa and Caldas 2007; Hamback et al. 2006;
Holt and Lawton 1994; Settle and Wilson 1990; Stiling et al. 2003). Oviposition
preference (Shiojiri et al. 2002), differences in plant chemistry (Redman and Scriber
2000; Shiojiri et al. 2001), and habitat modification (White and Andow 2006) have also
been shown to drive AR/AS among herbivore species. Various AR/AS mechanisms, and
their effects on plant and herbivore populations, have been the subject of several reviews
(Agrawal et al. 2006; Andow 1991; Barbosa et al. 2009; Hamback and Beckerman 2003;
Milchunas and Noy-Meir 2002; Russell 1989; Sheehan 1986) and therefore only the
enemies hypothesis, which the present study tests, will be detailed here.
The enemies hypothesis predicts that a higher availability of alternative energy
sources (e.g., nectar, pollen, or alternative prey) can lead to higher densities of predators
and parasites in polycultures, as compared with monocultures (Root 1973). For these
“insectary plants” (Atsatt and Odowd 1976) to actually provide AR, their alternative
energy sources must increase predator and/or parasitoid efficiency, longevity, or
herbivore encounter rate, leading to increased herbivore mortality and a subsequent
decrease in damage on the target plant. Support for the enemies hypothesis comes from
both agricultural (Harmon et al. 2000; Mathews et al. 2007; Spellman et al. 2006) and
natural systems (Stenberg et al. 2007; Stiling et al. 2003). Harmon et al (2000) found
that density of common dandelion, Taraxacum officinale, in alfalfa fields was negatively
correlated with density of pea aphid, Acyrthosiphon pisum, and positively correlated with

59
density of the lady bird beetle Coleomegilla maculata. Caged laboratory experiments
showed that C. maculata predation of A. pisum was twice as high in alfalfa-dandelion
patches compared with alfalfa only patches. The authors posit that dandelion pollen
provides an alternative energy source for C. maculata, increasing tenure time in alfalfa
patches with higher dandelion densities and potentially increasing control of pea aphids
across generations. Such studies show that understanding the effects of plant neighbors
on natural enemies can lead to more effective control of pest herbivore species.
Extra-floral nectar (hereafter, EFN) is an important alternative energy source for
natural enemies of herbivores, particularly ants. Numerous studies have shown a
protective benefit of EFN-tending ants to plants in natural systems (Barton 1986; Bentley
1977; DelClaro et al. 1996; Heil and McKey 2003; Janzen 1966; Koptur 1984; Pickett
and Clark 1979; Tilman 1978). In addition, conservation biological control studies have
demonstrated that interplanting crops with species or cultivars that produce EFN can
increase the performance of biological control agents (Lewis et al. 1998; Limburg and
Rosenheim 2001; Mathews et al. 2007). EFN-producing plants have also been studied in
the context of invasive ant-plant interactions, which have the potential to either benefit or
harm the plant (Fleet and Young 2000; Lach 2003; Ness 2003; Stiles and Jones 2001).
For example, it has been suggested that EFN-producing plants can “fuel” the growth of
invasive ant populations, thereby reducing local native ant diversity (Ness and Bronstein
2004; Savage et al. 2009). However, to our knowledge, there have been no studies
documenting EFN-mediated AR to an invasive herbivore in a natural setting.
Cactoblastis cactorum, the cactus moth, is an invasive pest in coastal regions of
the southeast US. Ant predation of C. cactorum has been documented in its native South

60
America (Lobos and de Cornelli 1997) and in areas where it was released for biological
control of Opuntia spp. (e.g. Australia: Dodd 1940; Hawaii: Fullaway 1954; South
Africa: Pettey 1947; Robertson and Hoffman 1989). However, there has been almost no
investigation into the potential for ant predation of C. cactorum in the southeast US (but
see Bennett and Habeck 1996 and Miller et al. 2010). In this region the C. cactorum
hosts Opuntia stricta and O. humifusa frequently co-occur with the legume
Chamaecrista fasciculata, which attracts ants to its EFN. Such a system provided us the
opportunity to answer the following questions: (1) In Florida, are any ant species preying
upon C. cactorum eggs or pupae? (2) If so, what are the predation rates? (3) Do Opuntia
spp. receive AR from C. fasciculata via ants collecting its EFN? We predicted that
evidence of ant predation would be found for both egg and pupal stages (the larval stage
was not examined because C. cactorum feed internally as larvae). We also predicted that
C. fasciculata would provide AR to Opuntia spp. growing in close association, and tested
this prediction with damage and growth surveys of Opuntia individuals and manipulative
ant predation experiments.

61
Study system and methods
Insects
Cactoblastis cactorum is a cactus-feeding, pyralid moth native to northern Argentina,
Paraguay, Uruguay, and southern Brazil. It has been released intentionally in a number of
countries, most notably Australia, to control pest Opuntia species, and was discovered in
Florida in 1989. Cactoblastis cactorum has since spread along the Atlantic and Gulf
coasts of the southeastern United States. Females lay about 40-100 eggs one on top of the
other to form an eggstick that is attached to the tip of a cactus spine or directly to a
cladode. The larvae feed internally on a number of opuntioid species, during which time
they relatively protected from predation. Larval damage results in hollowed out cladodes
and can lead to secondary infections and in some cases, the death of entire plants
(Zimmermann et al 2004). After completing six instars, they drop to the ground to pupate
in the soil or hollowed out cladodes. In central Florida, there are three non-overlapping
flight periods per year: a spring flight beginning in mid- February, a summer flight
beginning in early June, and a fall flight beginning in late August (Hight and Carpenter
2009).
Five native species of Opuntia-feeding insects were included in damage surveys
(Table 3.1). All of these species are specialists on opuntioids, with the exception of
Diaspis echinocacti, which feeds of a variety of cactus species (Hunter 1912; Mann
1969). Dactylopius confusus and D. echinocacti are primarily sessile and are therefore
easily quantified. The remaining three species leave characteristic signs of damage
(Table 3.1), allowing damage levels to be assessed even when insects are not directly
observed. The ant species Monomorium minimum has been reported as an egg predator of

62
Chelinidea vittiger (Hamlin 1924). Pickett and Clark (1979) found that C. vittiger
individuals spent less time on Opuntia acanthocarpa plants occupied by Crematogaster
opuntiae; they also observed C. opuntiae attack and kill C. vittiger nymphs. The
remaining four insect species included in damage surveys have no accounts in the
literature of interactions with ants (either through predation or tending behavior), nor
were any such interactions observed during the course of the study.
Plants
Two Opuntia species native to Florida were used in this study, O. stricta and O.
humifusa. Both are common hosts of C. cactorum, although O. stricta is attacked more
often than O. humifusa (Baker and Stiling 2009). They exhibit slightly different
morphologies; O. stricta has a more erect growth form and more spines, while O.
humifusa has a low, spreading growth form and fewer spines. Both species produce EFN
at the areoles of newly developing cladodes and flower buds, which are mainly present in
April and May in the Tampa Bay area (H. Jezorek pers. obsv.). EFN is produced both
day and night (Oliveira et al. 1999); data on the volume and composition of EFN
produced by O. stricta and O. humifusa were not found in the literature and were not
collected in the present study. Studies from the native range of O. stricta recorded 16 ant
species, mainly belonging to the subfamilies Myrmicinae and Formicinae, associated
with its EFN (Oliveira et al. 1999; Miller at al. 2010; Robbins and Miller 2009). To the
best of our knowledge there are no studies of ant-EFN associations on O. humifusa.
Chamaecrista fasciculata is a native legume that is common throughout the mid-
western and eastern regions of North America. It co-occurs with O. humifusa throughout

63
most of its range and co-occurs with O. stricta in coastal regions of the southeast and
Gulf coast states. Chamaecrista fasciculata produces EFN at the base of each leaf petiole
and EFN is produced both day and night throughout the growing season (Kelly 1986).
Plants can have over 200 active nectaries and secrete up to 3 µl of EFN per day (Rutter
and Rausher 2004). Ants collect EFN from two weeks after plant emergence to pod
senescence (Barton 1986). In central Florida, this period lasts from mid-February to late
October or early November (H. Jezorek pers. obsv.). At least 28 ant species have been
recorded collecting EFN from C. fasciculata (Barton 1986; Boecklen 1984; Kelly 1986;
Rios et al. 2008; Stiles and Jones 2001); the vast majority of these species are found in
the subfamilies Myrmicinae (14 species) or Formicinae (10 species). Two studies
performed in northern Florida found the most common visiting ants to be Forelius (=
Iridomyrmex) pruinosus, Camponotus floridanus, and Crematogaster ashmeadi (Barton
1986; Boecklen 1984).
Damage and growth surveys
Surveys were conducted at Honeymoon Island (hereafter, HI) and Caladesi Island
(hereafter, CI) State Parks (Pinellas County, FL). Both parks lie within Florida‟s west-
central barrier island chain and are separated by approximately 4.5 km. Opuntia stricta,
O. humifusa, and C. fasciculata are common in the sand dune and coastal scrub habitats
of these islands.
Four surveys of naturally occurring Opuntia were conducted: Aug-2008 at HI
(n=42 plants) and Nov-2008, May-2009, and Sept-2009 at CI (n=28, 40, and 42,
respectively). Surveys were conducted between 0900 and 1200. For each survey, Opuntia

64
plants were chosen haphazardly such that half were ≥3 m from any C. fasciculata
individual (“far from” category) and half were ≤ 0.25 m from a C. fasciculata individual
(“near to” category). Distances were measured from the center of each plant, with the
result being that the “near to” Opuntia were usually touching C. fasciculata leaves, i.e.
vegetation bridges were present. Each survey represented a new sample of plants;
distances between the surveyed plants varied, but were ≥5 m. The HI survey included
both O. stricta (n=16) and O. humifusa (n=26) while the CI surveys included only O.
humifusa. The following parameters were measured for each Opuntia individual during
each survey: number of primary, secondary, and tertiary cladodes, number of cladodes
with old C. cactorum damage, number of cladodes with active, i.e. feeding, C. cactorum
larvae, number of cladodes with C. cactorum eggsticks, number of cladodes with native
insect damage (for each of five species in Table 3.1), and ant activity. Ant activity was
measured by counting the number of ants on a plant during a one minute period and then
dividing by the total number of cladodes. Most plants had 1-2 ant species present during
the observation period, therefore relative abundance of ant species was not directly
quantified during surveys. Instead, when a survey plant had ants present, one ant per
observed species was aspirated immediately after the observation period and transferred
to ethanol for off-site species identification (total ants collected=99). Any plant with a C.
cactorum eggstick was checked four weeks later for larval entrance holes or active
larvae. For the second CI sample (n=40), number of cladodes and height (cm) were
measured in April-2009 and again in Oct-2009 in order to estimate growth. Growth was
not quantified for the other three samples, as they were not surveyed during the Opuntia
growing season.

65
We conducted separate analyses for each survey, as each represented a unique
combination of plants, site, and date, and our questions were not aimed at comparing
between surveys, but rather between plant categories within surveys. Data for insect
response variables included many zeros and could not be transformed to meet the
assumptions of parametric analysis. Therefore, proportions of pads with C. cactorum
damage and native insect damage were compared between “near to” and “far from”
categories with Mann-Whitney tests for independent samples. For the HI survey, data
from O. stricta and O. humifusa were pooled, as Kolmogorov-Smirnov tests found no
significant differences in the distribution of insect response variables (P-values ranged
from 0.289 to 1.00). Fisher‟s exact tests were used to compare the frequency of plants in
each category with 1) any type of C. cactorum damage and 2) active larvae one month
after discovery of an eggstick. Growth variables (proportional change in cladode number
and change in height from April-2009 to Oct-2009) were compared between “near to”
and “far from” categories using single factor ANOVA, as these data met normality and
homogeneity of variance assumptions. All analyses were conducted in STATISTICA 5.5
(StatSoft 2000).

66
Predation Experiment
Cactoblastis cactorum eggsticks and pupae, obtained from the USDA-ARS Crop
Protection and Management Research Unit in Tifton, GA, were irradiated at 200 Gy with
a Cobalt 60 Gamma cell 220 irradiator. Irradiation at this level results in sterility of any
F1 progeny (Carpenter et al. 2001; Tate et al. 2007) and was taken as a precaution in
case of early hatch, early emergence, or unrecovered eggsticks or pupae.
In Aug-2008 and Oct-2008, eggsticks containing an average of 56.1(standard
error ±1.2) eggs were placed on Opuntia plants (n=42) at HI, with 3-6 eggsticks per
plant. These months correspond to the summer and fall flights, respectively, of C.
cactorum at similar latitude in Florida (Hight and Carpenter 2009). The same plants (16
O. stricta and 26 O. humifusa) and categories (“near to” and “far from” C. fasciculata)
from the Aug-2008 damage and growth survey were used, but the survey was completed
prior to placement of experimental eggsticks or pupae. We were unable to replicate the
usual oviposition behavior of C. cactorum females, i.e. eggsticks laid on the tip of cactus
spines. Instead, eggsticks were placed on the margins of secondary Opuntia cladodes by
inserting their tips into small holes created with a dissecting probe. This method resulted
in the first 3-5 eggs of the eggstick being inaccessible to predators, as mucilage from the
created hole dried around the end of the eggstick to hold it in place. The location of
eggsticks was marked to enable recovery after 14 days in the field. In Sept-2008 and
Nov-2008, when moths from the summer generation are naturally pupating, five pupae
were placed around the same Opuntia plants. In order to mimic natural pupation
behavior, pupae were covered with sand and/or leaf litter, or placed in dead, hollowed
out cladodes at the base of the plant. The location of each pupa was marked to enable

67
recovery after 10-14 days in the field. We recognize that our methods of placing
eggsticks and pupae do not perfectly imitate the natural behavior of C. cactorum.
However, using experimentally placed, sterile eggsticks and pupae allowed us to obtain
adequate sample sizes; this would be extremely difficult with natural eggsticks and
pupae.
Recovered eggsticks and pupae were placed in individual, ventilated vials, taken
to the lab, and examined under a dissecting microscope for evidence of predation. They
were then returned to ventilated vials and kept at ~25°C and a 14L:10D photoperiod in
order to rear out any parasitoids. Eggsticks were placed into one of four groups:
“missing” (not recovered from HI), “intact” (no breaks or rips), “evidence of predation”
(distinct jagged break or eggs ripped open), or “undetermined” (clean break or break not
sufficiently jagged to infer predation). Eggsticks placed in the “undetermined” category
were excluded from the analysis. Pupae were also placed into one of four groups:
“missing” (not recovered from HI), “intact” (no holes in cocoon and pharate present),
“evidence of predation” (cocoon appeared ripped open; the placement and size of hole in
cocoon was atypical for C. cactorum emergence and the pupal case was absent), or
“emerged” (the placement and size of hole in cocoon was typical for C. cactorum
emergence and the pupal case usually present).
For each replication of the experiment, proportions of eggsticks or pupae in each
group for each plant were compared among categories and species using Mann-Whitney
tests for independent samples, as residuals were again highly non-normal. A Fisher‟s
exact test was used to compare the frequency of eggsticks or pupae showing evidence of

68
predation between the “near to” and “far from” C. fasciculata treatments. All analyses
were conducted in STATISTICA 5.5 (StatSoft 2000).
Results
Damage and growth surveys
The proportion of cladodes with any stage of C. cactorum damage (eggsticks, active
larvae, or old damage) was higher on “far from” plants than “near to” plants in all
surveys except Nov-2008 (Fig. 3.1a). The difference was significant in Aug-2008 (Mann-
Whitney U=291.0, p=0.035) and Sept-2009 (U= 282.0, P=0.024). The proportion of
cladodes with eggsticks was not significantly different between plant categories for any
surveys (Fig. 3.1b). The proportion of cladodes with active larvae was higher on “far
from” plants than “near to” plants in all surveys except Nov-2008 (Fig. 3.1c), but the
difference was only significant for the Sept-2009 survey (U=273.0, P=0.019) when no
active larvae were found on “near to” plants. The proportion of cladodes with old C.
cactorum damage was higher on “far from” plants than “near to” plants in all surveys
(Fig. 3.1d), with significant differences in Aug-2008 (U=296.0, P=0.008) and May-2009
(U=248, P=0.050). Fisher‟s exact tests revealed that “far from” Opuntia plants had C.
cactorum damage significantly more often than expected in Sept-2009 (one-tailed
P=0.022), with marginally significant results from the Aug-2008 and Nov-2008 surveys
(one-tailed P=0.090 and 0.063, respectively). Additionally, “near to” Opuntia plants
were significantly less likely than expected to contain active larvae one month after
discovery of an eggstick (one-tailed P=0.0048).

69
Proportions of cladodes damaged by the five native insect species were not
significantly different between “near to” and “far from” plants, although a slight trend for
higher damage on the “far from” plants was observed (Fig. 3.2a-e). For all surveys, more
ants per cladode were found on plants in the “near to” group (Fig. 2f); this result was
significant in Aug-2008 (U=130.0, P=0.015), May-2009 (U=125.0, P=0.032), and Sept-
2009 (U=123.5, P=0.012). Six species of ants were identified from the sample collected
from survey plants: Monomorium viride, Forelius pruinosus, Solenopsis invicta,
Dorymyrmex bureni, Pseudomyrmex gracilis, and Camponotus planatus (Table 3.2). M.
viride, F. pruinosus, and D. bureni are native to Florida, the remaining three species are
non-native (Deyrup et al 2000).
Opuntia individuals in the “near to” category added significantly more cladodes
than those in the “far from” category (F=4.61, P=0.038); the difference for growth by
height was only marginally significant (F=3.28, P=0.078) (Fig. 3.3).
Predation experiment
In the field, ants were frequently observed searching the tips of spines of O. stricta and
O. humifusa plants (H. Jezorek pers. obsv.), but predation of eggsticks was not directly
observed. Solenopsis invicta were observed investigating C. cactorum cocoons nearly
immediately upon placement at the base of Opuntia plants, prior to concealment with
sand or leaf litter, and two S. invicta individuals were found stuck in the silk of a C.
cactorum cocoon that was ripped open and emptied.
In Aug-2008, “near to” and “far from” plants differed significantly in the
proportion of eggsticks displaying evidence of predation (U= 145, P=0.030) (Fig. 3.4a).

70
In Oct-2008 this difference was marginal (U=153.5, P=0.062) and the difference in the
proportion of intact eggsticks was significant (U=130.0, P=0.020) (Fig. 3.4b). There
were no significant differences between O. stricta and O. humifusa in either month.
Results from the Fisher‟s exact test showed that eggsticks placed on “near to” plants
showed evidence of predation significantly more often than expected in both Aug-2008
and Oct-2008 (one-tailed P=0.017 and 0.023, respectively). Of the intact eggs recovered,
92.1% hatched and no parasitoids were found.
In Sept-2008, all categories of pupae were significantly different between “near
to” and “far from” plants (Intact, U=325, P=0.006; Missing, U=116, P=0.006; Emerged,
U=315, P=0.010; Evidence of predation, U=134, P=0.021) (Fig. 3.4c). No significant
differences were found in Nov-2008 (Fig. 3.4d) or between O. stricta and O. humifusa
for either month. The Fisher‟s exact test revealed that pupae placed on “near to” plants
showed evidence of predation significantly more often than expected in Sept-2008, but
not in Nov-2008 (one-tailed P=0.017 and 0.531, respectively).
Five chalcid wasps emerged from pupae placed on “near to” plants and another
five emerged from pupae placed on “far from” plants. Three of the wasps were recovered
from the Sept-2008 experiment and seven from the Nov-2008 experiment. Nine of the
individuals were identified as Brachymeria pschye; the remaining individual was
identified as Conura side. For recovered, intact pupae the parasitism rate was 5.29%; for
all pupae placed in the field the rate was 2.38%.

71
Table 3.1: Native Opuntia herbivores found in central Florida and their characteristic feeding damage
Insect species Order: Family Common
name Description of feeding/damage
Chelinidea vittiger McAtee Hemiptera: Coridae cactus bug mobile sap sucker; leaves pale half or full
circles on surface of cladode; high levels of
infestation lead to desiccation of cladodes
Dactylopius confusus
Cockerell
Hemiptera:
Dactylopiidae
Cochineal sessile sap sucker; exudes fluffy white
substance on surface of cladodes
Diaspis echinocacti Bouché Hemiptera:
Diaspididae
cactus scale sessile sap sucker; circular, slightly convex,
whitish to tan scale; high levels of infestation
lead to chlorotic patches and desiccation of
cladodes
Gerstaeckeria hubbardi
LeConte
Coleoptera:
Curculionidae
cactus weevil internal feeder; leaves light brown circular
cell; cladode appears pierced by tiny “bullet
hole”
Marmara opuntiella Busck Lepidoptera:
Gracillariidae
cactus stem
miner
internal feeder; pale green to white serpentine
mines on cladodes

72
Figure 3.1: Results from C. cactorum damage surveys. Proportion of cladodes with (a) any type of Cactoblastis cactorum
damage, (b) C. cactorum eggsticks, (c) C. cactorum larvae, and (d) old C. cactorum damage for surveyed Opuntia plants near
to C. fasciculata (unfilled bars) and far from C. fasciculata (filled bars). For ease of interpretation, bars show mean (+SE), but
asterisks represent significant results from a non-parametric Mann-Whitney test (**, P<0.05, ***, P<0.01).
0
0.004
0.008
0.012
0.016
b. Eggsticks
0
0.01
0.02
0.03
0.04
0.05
Aug. '08 Nov. '08 May '09 Sept. '09
Pro
po
rtio
n o
f cl
ado
des
c. Larvae
**
0.00
0.02
0.04
0.06
0.08
0.10
0.12
Aug. '08 Nov. '08 May '09 Sept. '09
d. Old damage
***
**
0
0.04
0.08
0.12
0.16
Pro
po
rtio
n o
f cl
ado
des
a. Any damage
**
**

73
Figure 3.2: Results from native herbivore and ant surveys. Proportion of cladodes with
damage from (a) Chelinidea vittiger, (b) Dactylopius confusus, (c) Diaspis echinocacti,
(d) Gerstaeckeria hubbardi, and (e) Marmara opuntiella, and (f) number of ants per
cladode counted during one minute observation periods for surveyed Opuntia plants near
to Chamaecrista fasciculata (unfilled bars) and far from C. fasciculata (filled bars). For
ease of interpretation, bars show mean (+SE), but asterisks represent significant results
from a non-parametric Mann-Whitney test (**, P<0.05)
0
0.02
0.04
0.06
0.08
Pro
po
rito
n o
fcla
do
des
c. D. echinocacti
0.00
0.10
0.20
0.30
0.40
0.50
Aug. '08 Nov. '08 May '09 Sept. '09
Num
ber
per
cla
do
de
f. Ants
**
** **
0.00
0.02
0.04
0.06
0.08
0.10
b. D. confusus
0.00
0.05
0.10
0.15
0.20
0.25
d. G. hubbardi
0.00
0.02
0.04
0.06
0.08
0.10
0.12
Pro
po
rtio
n o
f cl
ado
des
a. C. v. aequoris
0.0000
0.0050
0.0100
0.0150
Aug. '08 Nov. '08 May '09 Sept. '09
Pro
po
rtio
n o
f cl
ado
des
e. M. opuntiella
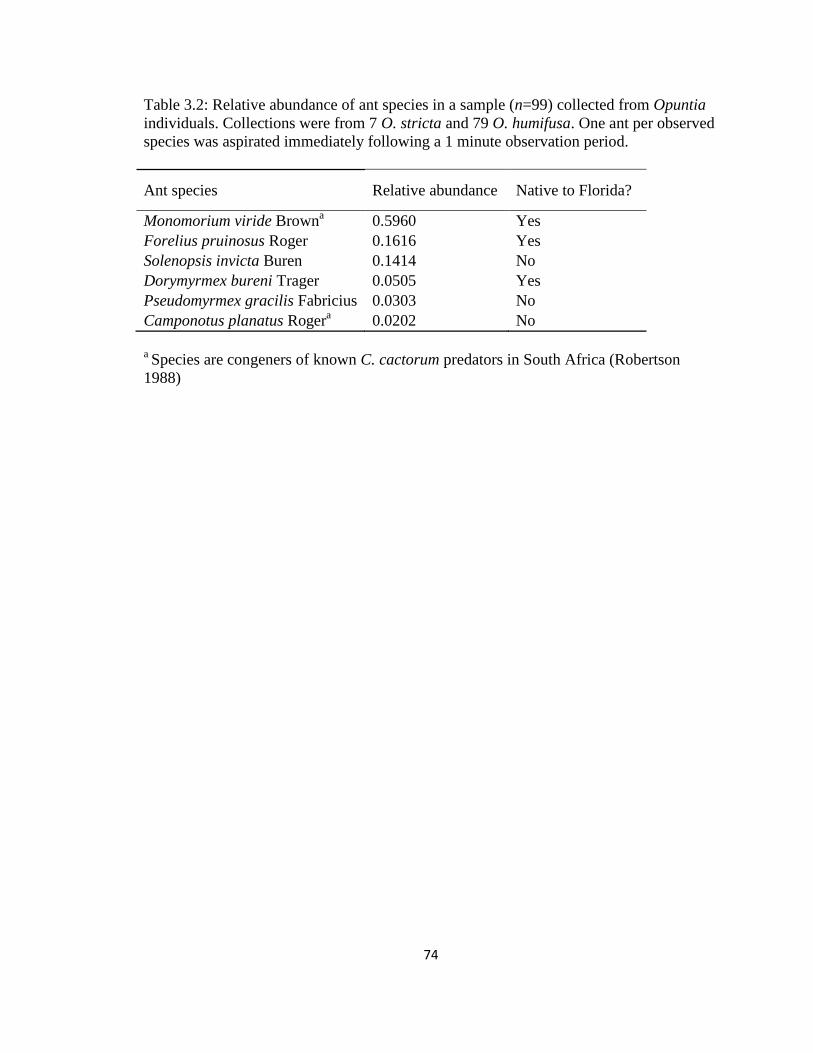
74
Table 3.2: Relative abundance of ant species in a sample (n=99) collected from Opuntia
individuals. Collections were from 7 O. stricta and 79 O. humifusa. One ant per observed
species was aspirated immediately following a 1 minute observation period.
Ant species Relative abundance Native to Florida?
Monomorium viride Browna 0.5960 Yes
Forelius pruinosus Roger 0.1616 Yes
Solenopsis invicta Buren 0.1414 No
Dorymyrmex bureni Trager 0.0505 Yes
Pseudomyrmex gracilis Fabricius 0.0303 No
Camponotus planatus Rogera 0.0202 No
a Species are congeners of known C. cactorum predators in South Africa (Robertson
1988)

75
Figure 3.3: Results from a growth survey from April-2009 to Oct-2009 of O. humifusa
plants (n=40) near to and far from C. fasciculata. a) Mean (+SE) proportional change in
cladodes; b) Mean (+SE) change in height.
0
1
2
3
4
5
6
7
Near C.f. Far from C.f.
Gro
wth
(cm
)
b.
p=0.078
-0.05
0
0.05
0.1
0.15
0.2
0.25
Near C.f. Far from C.f.
Pro
po
rtio
n o
f cl
ado
des
add
ed
a. p=0.038

76
Figure 3.4: Results from predation experiment. Proportion of eggsticks (a-b) or pupae (c-
d) in each category after collection from Opuntia plants far from Chamaecrista
fasciculata (n=21, unfilled boxes) and near to C. fasciculata (n=21, patterned boxes). I=
intact, M= missing, E= emerged, PR= evidence of predation. Boxes indicate interquartile
ranges, bold lines indicate medians, whiskers indicate inner fences, points indicate
outliers, and asterisks indicate extreme values.
a = P<0.05, b = P<0.01
b. October 2008
d. November 2008 c. September 2008
a. August 2008
a a
Pro
port
ion o
f pupae
P
rop
ort
ion
of
eggst
ick
s
a b b b

77
Discussion
We found evidence to support our prediction that ants prey upon C. cactorum eggsticks
and pupae in Florida. Approximately 16% of our experimental eggsticks and 17.5% of
our experimental pupae displayed breaks and/or rips that are consistent with descriptions
of ant predation (Dodd 1940; Pettey 1947; Robertson and Hoffmann 1989). The rates of
pupal predation we found are within the range reported from the Florida Keys (0- 36%,
Bennett and Habeck 1996) and South Africa (13-34%, Robertson and Hoffmann 1989).
However, substantially higher rates of egg mortality from predation have been reported
from both Argentina (30%) (Lobos and de Cornelli 1997) and South Africa (55-78%)
(Robertson and Hoffmann 1989). It should be noted that predation rates from the present
study have the potential to be underestimates, for two reasons. First, eggsticks were
collected after 14 days and pupae after 10-14 days, although the average duration of these
life stages is 30 and 20 days, respectively (USDA-APHIS 2007). They were therefore not
vulnerable to predation for the full development period. Second, we were conservative
when determining whether specimens showed evidence of predation and it is conceivable
that missing eggsticks and/or pupae could have been carried away by ants. Pettey (1947)
reported that in some South Africa C. cactorum colonies, up to 59% of eggsticks were
removed by ants. Future studies that include experimental manipulation of ants and
extended observation periods could reveal that actual predation rates are higher than what
we have reported. The lower rates found in our study could also be due in part to the
newer association of C. cactorum and ants in Florida than in the moth‟s native range or in
South Africa or Hawaii, where the moth has been present since 1933 and 1950,
respectively (Zimmermann et al. 2004).

78
Of the six ant species identified from our sample, half were non-native species,
accounting for about 20% of individuals (Table 3.2). The two most abundant species, M.
viride and F. pruinosus, are native to Florida and accounted for >75% of individuals.
Two congeners of M. viride (M. albopilosum and M. minutum) are known egg predators
of C. cactorum in South Africa (Robertson and Hoffmann 1989). More extensive
surveys and experiments are needed before statements can be made about the relative
importance of various ant species to C. cactorum predation in Florida. For example, S.
invicta accounted for only 14% of ants in our sample, yet its aggressive nature (Deyrup et
al. 2000; Ness 2003; Porter and Savignano 1990) could mean that it accounts for a
greater proportion of predation.
The absence of egg parasitism in our recovered eggsticks is consistent with
previous findings in Florida, where the only recorded instance comes from two eggsticks
parasitized by Trichogramma sp. (Bennett and Habeck 1996). Accounts of egg parasitism
from other regions vary widely (Pemberton and Cordo 2001). Approximately 5% of our
recovered pupae were parasitized; this agrees with what Bennett and Habeck (1996)
found during the same times of year in Florida (10% in September and 6% in November),
although their rates for other months were considerably higher (e.g. 30% parasitism in
July). Although both of the parasitoid species we found represent new records for C.
cactorum, they have relatively broad host ranges and are therefore not good candidates
for inundative biological control.
We found partial support for our prediction that Opuntia plants receive AR when
closely associated with C. fasciculata. Results from our Aug-2008, May-2009, and Sept-
2009 surveys provide evidence that Opuntia plants growing close to C. fasciculata may

79
experience less C. cactorum damage than plants far from C. fasciculata (Fig. 3.1) and
that this benefit could potentially be attributed to higher ant activity (Fig. 3.2f). However,
damage by five native insect species was not affected by distance to C. fasciculata,
indicating that AR benefits may not extend to all herbivores. The lack of an effect may be
explained by the life histories of the insects surveyed. The two scale insects, D.
echinocacti and D. confusus, may be vulnerable to predation during their crawler phases,
but nymphs in both species settle and begin producing waxy or wooly coverings,
respectively, within 24 hours (Mann 1969; Oetting 1984). Egg, larval, and pupal stages of
Gerstaeckeria hubbardi are passed inside cladodes and adults drop to the ground and
feign death if disturbed (Mann 1969; Woodruff 2009). Egg and larval stages of Marmara
opuntiella occur beneath the epidermis of a cladode (Hunter et al. 1912). In contrast to
these native insects, C. cactorum eggsticks and pupae are accessible to predators for a
relatively long duration. The absence of a significant difference in damage by C. vittiger
is more difficult to explain, as eggs of this species are known suffer from ant predation
(Hamlin 1924), and at least the early nymph stages would seem to be vulnerable to
predation. Adults may be protected from ant predation because of their relatively large
size of up to 13mm long and 4.5mm wide (Hamlin 1924).
Miller et al (2010) found that excluding ants from Opuntia spp. did not deter
oviposition by C. cactorum or a native cactus-feeding moth, Melitara prodenialis. Low
numbers of eggsticks found in our surveys make it difficult to draw definite conclusions,
but we also found no evidence that higher ant activity on the “near to” plants affected C.
cactorum oviposition. It is interesting to note that eggsticks found on “near to” Opuntia
were far less likely to result in larval damage than eggsticks found on “far from” Opuntia.

80
Again, this effect could be mediated by differences in ant activity, whereby higher ant-
eggstick encounter rates occur on Opuntia growing in close to C. fasciculata.
“Near to” Opuntia plants added more cladodes, but not height, than “far from”
plants (Fig. 3.3). However, only one set of plants was surveyed for growth and all
individuals were O. humifusa. This species has a low, sprawling growth form and
proportionate change in cladode number may be a more informative measure of growth
than height. The fact that O. humifusa plants far from C. fasciculata actually lost
cladodes over the growth season (Fig. 3a) could be due to increased C. cactorum damage,
as cladodes hollowed out by larvae often fall off the plant. Vegetative reproduction is
extremely common in Opuntia (Anderson 2001). Gimeno and Vila (2002) found that
vegetative recruitment accounted for close to 50% of O. stricta juveniles in Spanish olive
groves. However, the relative importance of vegetative versus sexual recruitment in
Opuntia is known to vary by habitat (Del Carmen Mandujano et al 1998), so studies that
examine both methods are necessary to determine the full effects of plant neighbors and
predation on Opuntia fitness. For example, Oliveira et al (1999) found that excluding
ants from O. stricta branches translated into a 50% decrease in fruit set. Local
environmental factors should also be examined. C. fasciculata fixes nitrogen (Naisbitt et
al 1992), so it is possible that differences in soil chemistry contribute to its observed
effect on Opuntia growth and insect damage levels.
Our results from the predation experiments provided additional support for our
prediction of AR, in that more eggsticks from Opuntia close to C. fasciculata showed
evidence of predation. The fact that eggsticks from these same plants had fewer intact
eggsticks is also of interest. C. cactorum neonates work together to enter a cladode, often

81
making several attempts to overcome mucilage before successfully penetrating (Jezorek
et al. 2010; Robertson and Hoffmann 1989; Zimmermann et al. 2004). As a result of this
communal behavior, the loss of even a few eggs of an eggstick could end up being fatal
to the entire cohort if they are unable to successfully penetrate the cladode. The minimum
number of neonates needed for successful penetration is unknown, but is a potential topic
for further study. Results from our Sept-2008 pupal experiment were consistent with the
AR prediction, but distance to C. fasciculata did not have an effect in our Nov-2008
experiment. This could be attributed to the fact that C. fasciculata has generally stopped
producing EFN by this point in the year (H. Jezorek, pers. obsv.) meaning that ants are
equally likely to encounter C. cactorum on Opuntia plants near to or far from C.
fasciculata.
In summary, C. cactorum is very likely subject to ant predation of its egg and
pupal stages in Florida, although it is not yet known which species function as the main
predator(s). Overall, our results support the enemies hypothesis (Root 1973).
Chamaecrista fasciculata may be acting as an “insectary plant” (Atsatt and Odowd 1976)
for nearby Opuntia individuals by “sharing” the anti-herbivore effects of its EFN
production and its well-documented attractiveness to predacious ants (Abdala-Roberts
and Marquis 2007; Barton 1986; Kelly 1986; Rios et al. 2008; Rutter and Rausher 2004).
What is not clear from our study is whether this AR is due solely to differences in ant
abundance, or also to differences in ant species assemblages, which we did not measure.
Although the effects of AR detailed here may only last as long as EFN is produced, i.e.
mid-February to October in our study sites, this period overlaps with all three C.
cactorum flight periods (Hight and Carpenter 2009), and most of the three pupation

82
periods. Land managers and others concerned with protecting Opuntia from C. cactorum
may want to consider adopting practices, such as frequent prescribed burning, that
encourage the growth of C. fasciculata (Galloway and Fenster 2000) or other EFN-
producing plants.
Acknowledgements
We gratefully thank Susan Drawdy (USDA-ARS, Crop Protection and Management
Unit) for help in acquiring C. cactorum eggsticks and pupae, Mark Deyrup (Archbold
Biological Station) for ant identification, Michael Gates (USDA-ARS, Systematic
Entomology Laboratory) for parasitoid identification, Terry Hingtgen (Florida
Department of Environmental Protection, Southwest Division) for his assistance in
acquiring the necessary permits for this work, and University of South Florida for
providing funding.
References cited
Abdala-Roberts L, Marquis RJ (2007) Test of local adaptation to biotic interactions and
soil abiotic conditions in the ant-tended Chamaecrista fasciculata (Fabaceae).
Oecologia 154:315-326
Agrawal AA, Lau JA, Hamback PA (2006) Community heterogeneity and the evolution
of interactions between plants and insect herbivores. Q Rev Biol 81:349-376
Anderson EF (2001) The Cactus Family. Timber Press, Portland, OR
Andow DA (1991) Vegetational diversity and arthropod population response. Annu Rev
Entomol 36:561-586
Atsatt PR, Odowd DJ (1976) Plant Defense Guilds. Science 193:24-29
Baker AJ, Stiling P (2009) Comparing the effects of the exotic cactus-feeding moth,
Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae) and the native cactus-

83
feeding moth, Melitara prodenialis (Walker) (Lepidoptera: Pyralidae) on two
species of Florida Opuntia. Biol Invasions 11:619-624
Barbosa P, Caldas A (2007) Do larvae of species in macrolepidopteran assemblages share
traits that influence susceptibility to parasitism? Env Entomol 36:329-336
Barbosa P, Hines J, Kaplan J, Martinson H, Szczepaniec A, Szendrei Z (2009)
Associational resistance and associational susceptibility: Having right or wrong
neighbors. Annu Rev Ecol Evol S 40:1-20
Barton AM (1986) Spatial variation in the effect of ants on an extrafloral nectary plant.
Ecology 67:495-504
Bennett FD, Habeck DH (1996) Cactoblastis cactorum: A successful weed control agent
in the Caribbean, now a pest in Florida? In: Delfosse ES, Scott RR, (eds)
Biological control of weeds: Proceedings of the VIII international symposium on
biological control of weeds. CSIRO Publishing, Melbourne pp 21-26
Bentley BL (1977) Extrafloral nectaries and protection by pugnacious bodyguards. J Ecol
65:27-38
Boecklen WJ (1984) The role of extrafloral nectaries in the herbivore defense of Cassia
fasciculata. Ecol Entomol 9:243-249
Brown BJ, Ewel JJ (1987) Herbivory in complex and simple tropical successional
ecosystems. Ecology 68:108-116
Carpenter JE, Bloem S, Bloem KA (2001) Inherited sterility in Cactoblastis cactorum
(Lepidoptera: Pyralidae). Fla Entomol 84:537-542
Del Carmen Mandujano M, Montana C, Mendez I, Golubov J (1998) The relative
contribution of sexual reproduction and clonal propagation in Opuntia rastrera
from two habitats in the Chihuahuan desert. J Ecol 86: 911-921
DelClaro K, Berto V, Reu W (1996) Effect of herbivore deterrence by ants on the fruit set
of an extrafloral nectary plant, Qualea multiflora (Vochysiaceae). J Trop Ecol
12:887-892
Deyrup M, Davis L, Cover S (2000) Exotic ants in Florida. T Am Entomol Soc 126:293-
326
Dodd AP (1940) The biological campaign against prickly pear. Commonwealth Prickly
Pear Board Bulletin: 1-177. Brisbane, Australia.

84
Finch S, Billiald H, Collier RH (2003) Companion planting - do aromatic plants disrupt
host-plant finding by the cabbage root fly and the onion fly more effectively than
non-aromatic plants? Entomol Exp Appl 109:183-195
Fleet RR, Young BL (2000) Facultative mutualism between imported fire ants
(Solenopsis invicta) and a legume (Senna occidentalis). Southwest Nat 45:289-
298
Fullaway DT (1954) Biological control of cactus in Hawaii. J Econ Entomol 47:696-700
Gagnon P, Himmelman JH, Johnson LE (2003) Algal colonization in urchin barrens:
defense by association during recruitment of the brown alga Agarum cribrosum. J
Exp Mar Biol Ecol 290:179-196
Galloway LF, Fenster CB (2000) Population differentiation in an annual legume: local
adaptation. Evolution 54:1173-1181
Gimeno I, Vila M (2002) Recruitment of two Opuntia species invading abandoned olive
groves. Acta Oecologica 23: 239-246
Hamback PA, Agren J, Ericson L (2000) Associational resistance: Insect damage to
purple loosestrife reduced in thickets of sweet gale. Ecology 81:1784-1794
Hamback PA, Beckerman AP (2003) Herbivory and plant resource competition: A
review of two interacting interactions. Oikos 101:26-37
Hamback PA, Stenberg JA, Ericson L (2006) Asymmetric indirect interactions mediated
by a shared parasitoid: Connecting species traits and local distribution patterns for
two chrysomelid beetles. Oecologia 148:475-481
Hamlin JC (1924) A review of the genus Chelinidea (Hemiptera-Heteroptera) with
biological data. Ann Entomol Soc Am 17:193-208
Harmon JP, Ives AR, Losey JE, Olson AC, Rauwald KS (2000) Colemegilla maculata
(Coleoptera: Coccinellidae) predation on pea aphids promoted by proximity to
dandelions. Oecologia 125:543-548
Hay ME (1986) Associational plant defenses and the maintenance of species-diversity -
turning competitors into accomplices. Am Nat 128:617-641
Heil M, McKey D (2003) Protective ant-plant interactions as model systems in ecological
and evolutionary research. Annu Rev Ecol Evol S 34:425-453
Hight SD, Carpenter JE (2009) Flight phenology of male Cactoblastis cactorum
(Lepidoptera: Pyralidae) at different latitudes in the southeastern United States.
Fla Entomol 92:208-216

85
Hight SD, Carpenter JE, Bloem S, Bloem KA (2005) Developing a sterile insect release
program for Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae): effective
overflooding ratios and release-recapture field studies. Environ Entomol 34:850-
856.
Holt RD, Lawton JH (1994) The ecological consequences of shared natural enemies.
Annu Review of Ecol Syst 25:495-520
Hunter WD, Pratt FC, Mitchell JD (1912) The principle cactus insects of the United
States. United States Department of Agriculture, Bureau of Entomology,
Washington DC
Janzen DH (1966) Coevolution of mutualism between ants and acacias in Central
America. Evolution 20:249-275
Jezorek HA, Stiling PD, Carpenter JE (2010) Targets of an invasive species: oviposition
preference and larval performance of Cactoblastis cactorum (Lepidoptera:
Pyralidae) on 14 North American opuntioid cacti. Environ Entomol 39:1884-1892
Karban R (1997) Neighbourhood affects a plant's risk of herbivory and subsequent
success. Ecol Entomol 22:433-439
Kelly CA (1986) Extrafloral nectaries: ants, herbivores and fecundity in Cassia
fasciculata. Oecologia 69:600-605.
Koptur S (1984) Experimental evidence for defense of Inga (Mimosoideae) saplings by
ants. Ecology 65:1787-1793
Lach L (2003) Invasive ants: unwanted partners in ant-plant interactions? Ann Mo Bot
Gard 90:91-108
Lewis WJ, Stapel JO, Cortesero AM, Takasu K (1998). Understanding how parasitoids
balance food and host needs: importance to biological control. Biol Control
11:175-183
Limburg DD, Rosenheim JA (2001) Extrafloral nectar consumption and its influence on
survival and development of an omnivorous predator, larval Chrysoperla
plorabunda (Neuroptera: Chrysopidae). Environ Entomol 30:595-604
Lobos E, de Cornelli JO (1997) Observations on Cactoblastis cactorum (Berg) as a pest
of cactus pear (Opuntia ficus-indica) in Argentina with suggestions on possible
control methods. J Prof Assoc Cactus 2:97-102
Mann J (1969) Cactus-feeding insects and mites. Smithsonian Institution Bulletin 256,
Washington, D.C.

86
Mathews CR, Brown MW, Bottrell DG (2007) Leaf extrafloral nectaries enhance
biological control of a key economic pest, Grapholita molesta (Lepidoptera:
Tortricidae), in peach (Rosales: Rosaceae). Environ Entomol 36:383-389
Milchunas DG, Noy-Meir I (2002) Grazing refuges, external avoidance of herbivory and
plant diversity. Oikos 99:113-130
Miller TEX, Legaspi JC, Legaspi B (2010) Experimental test of biotic resistance to an
invasive herbivore provided by potential plant mutualists. Biol Invasions 12:3563-
3577
Naisbitt T, James EK, Sprent JI (1992) The evolutionary significance of the legume
genus Chamaecrista, as determined by nodule structure. New Phytol 122:487-492
Ness JH (2003) Contrasting exotic Solenopsis invicta and native Forelius pruinosus ants
as mutualists with Catalpa bignonioides, a native plant. Ecol Entomol 28:257-
251.
Ness JH, Bronstein IL (2004) The effects of invasive ants on prospective ant mutualists.
Biol Invasions 6:445-461
Oliveira PS, Rico-Gray V, Diaz-Castelazo C, Castillo-Guevara C (1999) Interaction
between ants, extrafloral nectaries and insect herbivores in Neotropical coastal
sand dunes: herbivore deterrence by visiting ants increases fruit set in Opuntia
stricta (Cactaceae). Funct Ecol 13:623-631
Pemberton RW, Cordo HA (2001) Potential and risks of biological control of
Cactoblastis cactorum (Lepidoptera: Pyralidae) in North America. Fla Entomol
84:513-526
Pettey FW (1947) The biological control of prickly pears in South Africa. Department of
Agriculture of the Union of South Africa
Pfister CA, Hay ME (1988) Associational plant refuges: Convergent patterns in marine
and terrestrial communities result from differing mechanisms. Oecologia 77:118-
129
Pickett CH, Clark WD (1979) Function of extrafloral nectaries in Opuntia acanthocarpa
(Cactaceae). Am J Bot 66:618-625
Porter SD, Savignano DA (1990) Invasion of polygyne fire ants decimates native ants
and disrupts arthropod community. Ecology 71:2095-2106
Redman AM, Scriber JM (2000) Competition between the gypsy moth, Lymantria dispar,
and the northern tiger swallowtail, Papilio canadensis: interactions mediated by
host plant chemistry, pathogens, and parasitoids. Oecologia 125:218-228

87
Rios RS, Marquis RJ, Flunker JC (2008) Population variation in plant traits associated
with ant attraction and herbivory in Chamaecrista fasciculata (Fabaceae).
Oecologia 156:577-588
Robbins M, Miller TEX (2009) Patterns of ant activity on Opuntia stricta (Cactaceae), a
native host-plant of the invasive cactus moth, Cactoblastis cactorum
(Lepidoptera: Pyralidae). Fla Entomol 92:391-393
Robertson HG, Hoffmann JH (1989) Mortality and life-tables of Cactoblastis cactorum
(Berg) (Lepidoptera: Pyralidae) compared on two host-plant species. B Entomol
Res 79:7-17
Root RB (1973) Organization of a plant-arthropod association in simple and diverse
habitats: fauna of collards (Brasscia oleracea). Ecol Monogr 43:95-120
Russell EP (1989) Enemies hypothesis: A review of the effect of vegetational diversity
on predatory insects and parasitoids. Environ Entomol 18:590-599
Russell FL, Louda SM, Rand TA, Kachman SD (2007) Variation in herbivore-mediated
indirect effects of an invasive plant on a native plant. Ecology 88:413-423
Rutter MT, Rausher MD (2004) Natural selection on extrafloral nectar production in
Chamaecrista fasciculata: The costs and benefits of a mutualism trait. Evolution
58:2657-2668
Savage AM, Rudgers JA, Whitney KD (2009) Elevated dominance of extrafloral nectary-
bearing plants is associated with increased abundances of an invasive ant and
reduced native ant richness. Divers Distrib 15:751-761
Settle WH, Wilson LT (1990) Invasion by the variegated leafhopper and biotic
interactions: Parasitism, competition, and apparent competition. Ecology 71:1461-
1470
Sheehan W (1986) Response by specialist and generalist natural enemies to
agroecosystem diversification: A selective review. Environ Entomol 15:456-461
Shiojiri K, Takabayashi J,Yano S, Takafuji A (2001) Infochemically mediated tritrophic
interaction webs on cabbage plants. Popul Ecol 43:23-29
Shiojiri K, Takabayashi J, Yano S, Takafuji A (2002). Oviposition preferences of
herbivores are affected by tritrophic interaction webs. Ecol Lett 5:186-192
Spellman B, Brown MW, Mathews CR (2006) Effect of floral and extrafloral resources
on predation of Aphis spiraecola by Harmonia axyridis on apple. Biocontrol
51:715-724

88
StatSoft, Incorporated (2000) STATISTICA for Windows, Version 5.5. StatSoft,
Incorporated, Tulsa, OK
Stenberg JA, Heijari J, Holopainen JK, Ericson L (2007) Presence of Lythrum salicaria
enhances the bodyguard effects of the parasitoid Asecodes mento for Filipendula
ulmaria. Oikos 116:482-490
Stiles JH, Jones RH (2001) Top-down control by the red imported fire ant (Solenopsis
invicta). Am Midl Nat 146:171-185
Stiling P (2002) Potential nontarget effects of a biological control agent, prickly pear
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), in North America,
and possible management actions. Biol Invasions 4:273-281
Stiling P, Rossi AM, Cattell MV (2003) Associational resistance mediated by natural
enemies. Ecol Entomol 28:587-592
Tahvanainen JO, Root RB (1972) Influence of vegetational diversity on population
ecology of a specialized herbivore, Phyllotreta cruciferae (Coleoptera:
Chrysomelidae). Oecologia 10:321-346
Tate CD, Carpenter JE, Bloem S (2007). Influence of radiation dose on the level of F-1
sterility in the cactus moth, Cactoblastis cactorum (Lepidoptera: Pyralidae). Fla
Entomol 90:537-544
Thomas CD (1986) Butterfly larvae reduce host plant survival in vicinity of alternative
host species. Oecologia 70:113-117
Tilman D (1978) Cherries, ants and tent caterpillars: Timing of nectar production in
relation to susceptibility of caterpillars to ant predation. Ecology 59:686-692
United States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine (2007) Life history of the cactus moth.
http://www.aphis.usda.gov/plant_health/plant_pest_info/cactoblastis. Accessed 25
March 2011
Wahl M, Hay ME (1995) Associational resistance and shared doom: effects of epibiosis
on herbivory. Oecologia 102:329-340
White JA, Andow DA (2006) Habitat modification contributes to associational resistance
between herbivores. Oecologia 148:482-490
White JA, Whitham TG (2000) Associational susceptibility of cottonwood to a box elder
herbivore. Ecology 81:1795-1803

89
Woodruff RE (2009) Cactus Weevils, Gerstaeckeria hubbardi (LeConte) and
Gerstaeckeria fasciata Pierce (Insecta: Coleoptera: Curculionidae). University of
Florida Institute of Food and Agricultural Sciences, Department of Entomology
and Nematology EENY-82
Zimmermann HG, Bloem S, Klein H (2004) Biology, history, threat, surveillance and
control of the cactus moth, Cactoblastis cactorum. International Atomic Energy
Agency/Food and Agriculture Organization, Vienna

90
CHAPTER 4
FINDINGS FROM A SIX YEAR STUDY: EFFECTS OF CACTOBLASTIS
CACTORUM ON THE SURVIVAL AND GROWTH OF LOCAL OPUNTIA
Synopsis
Cactoblastis cactorum is known for being both a biological control agent and an invasive
pest of opuntioid cacti. The spread of C. cactorum in the southeastern United States may
threaten the biological and physical integrity of desert, scrub, and coastal habitats.
However, the effects of invasive species are known to vary spatially and temporally, and
C. cactorum‟s efficacy as a biological control agent varies considerably from region to
region. Therefore, the long term effects of C. cactorum within its U.S. range are still
uncertain. We surveyed tagged Opuntia stricta (n=253) and O. humifusa (n=327) plants
along the west coast of Florida for six years to determine the effects of C. cactorum
attack on survival and growth rate of plants, and to examine host species differences and
the effects of plant size. 78.11% of the Opuntia plants were attacked at least once by C.
cactorum during the six year study and overall survival rate was 75.8%. Plants attacked
by C. cactorum were more likely to die than unattacked plants and a plant‟s odds of
surviving the six year period decreased as C. cactorum attack frequency increased.
However, plants that survived the six year period showed, on average, positive growth
and there was no significant difference in growth rates between surviving attacked and
unattacked plants. O. stricta plants were more likely to be attacked, were attacked more

91
frequently, and were more likely to die after being attacked than were O. humifusa plants.
Plant size did not predict plant survival, but larger surviving plants lost proportionally
more pads over the six years than smaller surviving plants. Although C. cactorum should
still be considered a threat, particularly for rare opuntioids, overall survival at our sites is
currently high and plants that are able to survive C. cactorum attack are not being
reduced in size, possibly because they possess traits that render them more tolerant of C.
cactorum damage. Our findings suggest that an assumption of severe negative effects of
an invasive species, based on its effects in other regions or over short periods of time,
may not always be justified.
Introduction
Invasive species are often responsible for negative effects on native biodiversity which
can result from a variety of direct and indirect mechanisms (Mack et al. 2000, Snyder and
Evans 2006, Kenis et al. 2009). The severity of these negative effects certainly varies
from species to species, but is also expected to vary spatially and temporally for any
particular invasive species, leading to variation in its effects on the community (Parker et
al. 1999, Strayer et al. 2006). Despite the expectation of temporal variation, most studies
of invasive species take place over relatively short time scales. A review of studies of
invasive species from 2001-2005 found that approximately 80% were of durations of 3
years or less (Strayer et al. 2006). Studies of invaders over longer durations or over
multiple points in time post-invasion can reveal patterns that would otherwise be missed.
Severe initial impacts during early exponential growth of an invasive species, i.e., the log
phase (Mack et al. 2000), may be followed by a decline in its abundance, allowing for at

92
least partial recovery of resilient species and communities. Surveys from central Texas
early in the invasion stage of Solenopsis invicta showed that infested sites had
dramatically lower native ant and non-ant arthropod diversity (Porter and Savignano
1990). Twelve years later, however, ant and arthropod diversity at the same sites had
returned to pre-invasion levels and S. invicta abundance had declined (Morrison 2002).
Long term studies of invasive species can also reveal an increase in the severity of an
invader‟s effects. Wilson et al (2004) analyzed 19 years of data from a Wisconsin lake
undergoing dispersal by the invasive Orconectes rusticus (rusty crayfish). They found
that macroinvertebrates, macrophytes, and native crayfish experienced dramatic negative
effects very quickly after exposure to O. rusticus invasion, while negative impacts on
more mobile, longer-lived species took several more years to become evident and large
piscivorous fish such as bass had not yet suffered declines (Wilson et al. 2004). In both of
these cases, data from the first few years post-invasion leads to different conclusions, and
therefore different control and management decisions, than data from a decade post-
invasion. Continuing research for multiple decades post-invasion could lead to yet
different conclusions regarding the severity of an invader‟s effects, although logistical
and monetary constraints make this difficult in more cases than not.
Classical biological control provides some of the best evidence for spatial and
temporal variation in the impacts of non-native species, as the efficacy of a biological
control agent often varies across the invasive range of the target species (Monro 1975,
Phillips and Baird 2001, Denslow and D'Antonio 2005, Romeis et al. 2005, Shea et al.
2005). The pyralid moth Cactoblastis cactorum was a phenomenally successful
biological control agent of Opuntia stricta (erect prickly pear) in Australia (Dodd 1940,

93
Zimmermann et al. 2000). However, in South Africa, C. cactorum has not achieved the
same level of control of Opuntia spp. (Hoffmann et al. 1999, Zimmermann et al. 2004)
and in particular, has failed to control O. stricta in Kruger National Park (Hoffmann et al.
1998). Even within regions where C. cactorum is considered successful, levels of control
are variable, with persistent populations of target Opuntia spp. remaining decades after
the moth‟s introduction (Monro 1975, White 1980, Pemberton and Liu 2007).
In North America, C. cactorum is an invasive pest which currently ranges from
the Florida Keys north to Bull Island, SC and has been detected as far west as Terrebonne
Parish, LA (Rose et al. 2011). Although it is known to feed on nearly all opuntioid
species in its current U.S. range (Johnson and Stiling 1996, Jezorek et al. 2010, Rose et
al. 2011), the long term effects of C. cactorum are as yet unknown. Certainly for rare
species such as Consolea corallicola, which is endemic to the Florida Keys, the risk of
extinction is grave (Stiling 2010). For more common opuntioid species, quantitative data
on mortality rates within C. cactorum‟s U.S. range are lacking. Johnson and Stiling
(1998) found a mortality rate of 15% for O. stricta, but their data were from a small
sample size and gathered from 1991-1993, soon after the first U.S. detection of C.
cactorum in 1989. A second study found even lower mortality rates (~3.5%) for plants
damaged by C. cactorum (Baker and Stiling 2009); this study used data gathered more
recently and from a much larger sample size, but was of relatively short duration (2 years,
2003-2005). Such studies suggest that although C. cactorum is capable of high levels of
damage, an assumption that this damage will result in extirpation of common species of
Opuntia may not be warranted. Based upon the known variation in the efficacy of C.
cactorum as a biological control agent, and of the general tendency for impacts of

94
invasive species to vary over time, we wanted to document mortality and growth of local
Opuntia populations over a longer duration. We used data collected over a six year period
to determine: (1) Are plants attacked by C. cactorum more likely to die than unattacked
plants? (2) How does rate of C. cactorum attack affect survival of plants? (3) For
surviving plants, does C. cactorum attack affect growth rate? We also examined
differences between two host species, O. humifusa and O. stricta, and the relationship of
plant size to C. cactorum attack, survival, and growth rate.
Methods
Study system
Cactoblastis cactorum is a cactus-feeding, pyralid moth native to South America. During
the twentieth century, it was intentionally released to control pest species of opuntioid
cacti in Australia, South Africa, Mauritius, Hawaii, and the Caribbean. The moth was
first discovered in Florida in 1989 and has since spread along the Atlantic and Gulf coasts
of the southeastern United States. Females oviposit ~50-100 eggs one on top of the other
to form an eggstick that is attached to the tip of a cactus spine or directly to a cladode.
The gregarious larvae feed internally, completing five instars, and then drop to the
ground to pupate in the soil or hollowed out cladodes. In central Florida, there are three
non-overlapping flight periods per year: a spring flight beginning in mid- February, a
summer flight beginning in early June, and a fall flight beginning in late August (Hight
and Carpenter 2009). Cactoblastis cactorum has a wide host range within the genera
Opuntia, Consolea, and Nopalea. At our study sites, it feeds on the species O. stricta and
O. humifusa. Surveys were conducted at Fort deSoto Park (Pinellas County; hereafter

95
FDS), Honeymoon Island State Park (Pinellas County; hereafter, HI), and Lido Key
(Sarasota Country; hereafter LK). All three sites are part of Florida‟s west-central barrier
island chain and share similar plant communities; O. stricta and O. humifusa are common
in the dune and coastal scrub areas found on these islands.
Data collection and analysis
In March of 2003, Opuntia plants were marked at the three sites. Each plant was flagged
and had a numbered metal tag driven into the ground at its base. Sample sizes by site and
species were as follows: 140 plants (92 O. humifusa and 48 O. stricta) at FDS, 253 plants
(176 O. humifusa and 77 O. stricta) at HI, and 200 plants (100 O. humifusa and 100 O.
stricta) on LK. In April of 2006, an additional 43 O. stricta were marked at HI. Fifty-six
plants that were marked in 2003 were lost in prescribed burns and were removed from
analyses, leaving a total sample size of 580 (327 O. humifusa and 253 O. stricta).
Plants were surveyed monthly from April 2003 until July 2005 and tri-annually
from November 2005 until March 2009. Tri-annual surveys took place in March/April,
July, and October/November in order to correspond with peak larval activity. During a
survey, the number of live primary (old, woody), secondary (green, mature), and tertiary
(new growth with true leaves) cladodes and the number of cladodes with active C.
cactorum damage and eggsticks were counted for each plant. Damage was considered
active if cladodes were not yet fully hollowed out and had signs of larval infestation
including entrance holes, dripping frass, and a yellow or black “mushy” appearance.
When in doubt, cladodes were cut open to check for live larvae. Larvae and eggsticks of
the native cactus boring moth, Melitara prodenialis, are easily distinguishable from C.

96
cactorum, and were never observed at our sites. Plants were scored as dead when they
had zero live cladodes for a period of a year (i.e., three consecutive tri-annual surveys).
We tested for differences between sites using log likelihood tests for categorical
variables and ANOVA for continuous variables. No significant differences between sites
were found, therefore, data from the three sites were pooled for all analyses.
As some plants were marked in 2003 and other in 2006, data were analyzed
according to „survey year‟, rather than calendar year. For example, „survey year 2‟ is
calendar year 2004 for plants marked in 2003 and 2007 for plants marked in 2006. The
2006 sample consisted of O. stricta plants only and was therefore tested for heterogeneity
against O. stricta from the 2003 sample. We used log likelihood tests for categorical
variables and Student‟s t- tests for unequal sample sizes for continuous and scaled
variables. For categorical variables, no significant differences were found for survey
years 1 and 3; no plants from the 2006 sample died in survey year 2 so this comparison
could not be made. For continuous and scaled variables, there were no significant
differences between the two samples, although a Levene‟s test revealed unequal
variances for the variable attack frequency. A Welch‟s t-test, which does not assume
equal variance or sample size, was then performed and was non-significant. Therefore,
for survey years 1-3 data were pooled between the two samples.
All analyses were performed in SPSS 19 (SPSS Inc. 2010) unless otherwise
indicated. Prior to performing ANCOVA, assumptions of linearity of the covariate and
homogeneity of variance and slopes were checked. The variable attack frequency was
arcsine square root transformed to meet assumptions. Prior to performing linear
regression, assumptions of linearity, homogeneity of variance, normality of residuals, and

97
collinearity of predictors were checked. The variables proportional change in
cladodes/generation and initial cladode number were natural log-transformed to meet
these assumptions.
Q1:Are plants attacked by C. cactorum more likely to die than unattacked plants?
For each survey year, we used a likelihood ratio test (G2 test) of the null hypothesis that
attack status, survival status, and species were independent. We defined attack status for
any survey year as positive if a plant was attacked during at least one of the three C.
cactorum generations of that year and negative if a plant was not attacked at all that year.
We defined survival status for any survey year as positive for plants that were alive at the
end of the year, negative for plants that died that year, and undetermined for plants whose
fate was unknown at the end of that year. Only plants with known fates at the end of each
year were included in the test. We tested for homogeneity of odds ratios between species
with Tarone‟s X2 test. If odds ratios were found to be homogeneous, we used a Mantel-
Haenszel test for conditional independence of attack and survival status while controlling
for species, and found the Mantel-Haenszel common odds ratio estimate, which in this
case represents the ratio of the odds of dying for attacked versus unattacked plants, taking
into account species differences.
We also used a likelihood ratio test of the null hypothesis that final attack status,
final survival status, and species were independent, and determined relative risk and odds
ratios. We defined final attack status as positive if a plant was attacked by C. cactorum at
least once over the duration of the study and negative if a plant was never attacked by C.
cactorum. Final survival status was positive for plants that survived the duration of the

98
study, negative for those that did not survive, and undetermined for plants whose fate was
unknown at the end of the study. Only plants with known fates at the end of the study
were included in the test (n=532). We included the same Tarone‟s and Mantel-Haenszel
tests as described above. In order to test significance and find confidence intervals of
odds ratios, we performed backward elimination model selection with the same plants
and factors. Model selection indicated that all two way interactions and lower order
effects were significant (p=0.000-0.021), but the three way interaction was non-
significant and was not included in the model. Based on the results of the first likelihood
ratio test, and because nearly all O. stricta plants were positive for final attack status, we
performed a second likelihood ratio test and used a second log linear model to analyze
just those plants with both a positive final attack status and a known final survival status
(n=415). The purpose of these analyses was to determine if there were species differences
in survival specifically after C. cactorum attack.
Q2: How does frequency of attack by C. cactorum affect survival of plants?
Attack frequency for each plant over the duration of the study was defined as follows:
Number of C. cactorum generations for which active larvae were recorded
Total number of C. cactorum generations surveyed
For example, a plant surveyed from March 2006 to March 2009 was surveyed for ten C.
cactorum generations (3 per year for 2006-2008, plus one in 2009). If this plant had
active larvae during six of those generations, its attack frequency would be 6/10=0.600.
To determine if attack frequency differed between host species, we performed an

99
ANCOVA on all plants with species as a fixed factor and initial cladode number as a
covariate. This covariate was included because plants with more cladodes clearly have an
advantage in that they can “last longer”, that is, they provide a greater number of
opportunities for attack before suffering 100% cladode loss and therefore, death. Initial
cladode number refers to the number of live secondary and tertiary cladodes present on a
plant during its first survey; primary cladodes are generally not used by C. cactorum and
therefore were not included in the variable.
To determine the effects of attack frequency on plant survival, we used backward
stepwise selection to compare between logistic regression models. Inclusion in the model
was based upon a significant (p≤0.05) change in deviance (-2*log likelihood) when a
predictor term was removed from the model. The dependent variable was final survival
status and potential predictor variables were: attack frequency, host species, initial
cladode number, attack frequency*host species, and attack frequency*initial cladode
number. We performed the procedure with two sets of plants. First, we used all plants
with known fates at the end of the study (n=532); second, we used plants with known
fates and an attack frequency greater than zero (positive attack status, n=415). Model fit
was checked with a Hosmer-Lemeshow test.

100
Q3: For surviving plants, does C. cactorum attack affect growth rate?
Growth rate over the duration of the study was measured as proportional change in live
cladodes as follows:
Final number of cladodes – initial number of cladodes
Initial number of cladodes
This proportional change in cladodes was then divided by the number of C. cactorum
generations for which the plant was followed. For example, a plant with an initial cladode
number of 200 in April 2003 and a final cladode number of 100 in March of 2009 would
have a proportional cladode change of -0.500 over 19 C. cactorum generations (3 per
year for 2003-2008, plus a spring generation of 2009), which would give a proportional
cladode change per generation (hereafter PC/G) of -0.026. Surviving O. humifusa and O.
stricta were found to have equal variances for PC/G (Levene‟s F=0.019, p=0.890), so a
Student‟s t-test for independent samples was used to test for differences in mean PC/G
between host species. Attacked and unattacked survivors had unequal variance (Levene‟s
F=7.13, p=0.008) and unequal sample size (attacked, n=331; unattacked, n=109), so a
Welch‟s t-test was used to test for differences in mean PC/G between plants with positive
and negative final attack status. To determine if attack frequency or starting size of a
plant predicted PC/G, we performed multiple linear regression of ln(PC/G) on attack
frequency and ln(initial cladode number), both for all surviving plants and for each host
species separately.

101
Results
Attack rates for each survey year ranged from 27% to 41.5%, but survival rates were high
in all survey years (Table 4.1). Of the 580 plants used in analyses, 78.10% were attacked
by C. cactorum at least once during the study and 75.87% survived until March of 2009.
A higher percentage of O. stricta were attacked and died over the course of the study than
of O. humifusa (Fig.4.1).
Q1: Are plants attacked by C. cactorum more likely to die than unattacked plants?
For survey years 3-6, the proportion of plants that were attacked in the year they died was
lower than the proportion that were unattacked in the year they died. Survey years 1 and
2 showed the opposite trend (Fig. 4.2). We could not reject the null hypothesis of
independence of attack and survival status for survey years 1-4 and 6 (p-values for all G2
and Mantel-Haenszel X2 statistics >0.100). For survey year 5, we rejected the null
hypothesis for the total sample (both species pooled, G2=6.31, p=0.012) and for O.
humifusa (G2=6.28, p=0.012), but the relationships were relatively weak (Theil‟s
U=0.044 and 0.085, respectively) and we could not reject the null hypothesis for O.
stricta (G2=3.80, p=0.052). When controlling for species, our rejection of the null held
(Mantel-Haenszel X2=6.09, p=0.014), and the common odds ratio was 0.136, indicating
that the odds of a plant dying during survey year 5 were lower if it experienced at least
one C. cactorum attack than if it did not experienced an attack. For all survey years, the
odds ratio of dying for attacked versus unattacked plants was homogeneous between
species (p-values for all Tarone‟s X2 statistics >0.250).

102
We rejected the null hypothesis that final attack and final survival status were
independent for the total sample (both species pooled) or for O. humifusa. However, the
associations were weak, as evidenced by low values of Theil‟s U (Table 4.2). We could
not reject the null hypothesis for O. stricta. This non-intuitive result is due to the fact that
92.6% of O. stricta plants in the sample were attacked. The very low frequency of
unattacked O. stricta made comparisons between attacked and unattacked categories
somewhat problematic, and led us to perform a second test examining species differences
in survival after C. cactorum attack (see below and Table 4.2 and 4.3). When controlling
for species, our rejection of the null held (Mantel-Haenszel X2 statistic = 4.14, df=1,
p=0.043). For both species, the relative risk of dying was higher for attacked plants than
unattacked plants; the increase in risk was higher for O. humifusa but the difference in
risk between species was not significant (Table 4.2c). The Mantel-Haenszel common
odds ratio was 2.33 (p=0.033), indicating that the odds of a plant dying during the study
if it experienced at least one C. cactorum attack was 133% higher than if it experienced
no attacks.
The log linear model using all two-way interactions and lower order effects of
final attack status, final survival status, and species adequately fit the data (G2=1.92,
df=1, p=0.166). Plants were significantly more likely to survive than die (Z=4.29,
p=0.000) and significantly more likely to be have a positive attack status than a negative
one (Z=8.753, p=0.000). Interaction parameters confirmed that plants with a positive final
attack status had a significantly higher risk of death over the course of the study, and
revealed that O. humifusa plants had lower risk of death, regardless of attack status, and a
lower risk of being attacked by C. cactorum (Table 4.2).

103
The second likelihood ratio test revealed that species and survival after attack by
C. cactorum are not independent, although their association was weak (Table 4.2). The
relative risk of dying after an attack was lower for O. humifusa than O. stricta (Table
4.2); the parameter estimate from the second log linear model showed that this was
significant (Table 4.3).
Q2: How does frequency of attack by C. cactorum affect survival of plants?
The mean attack frequency for all plants of both species was 16.67%. The ANCOVA
interaction term between host species and initial cladode number was non-significant
(F=1.52, p=0.219), showing homogeneity of the coefficient for cladode number across
species. Opuntia stricta had a higher mean attack frequency (24.13%) than O. humifusa
(10.90%) and ANCOVA indicated that this difference was significant (F=150.24,
p=0.000) with a relatively large effect size (η2=0.207). Initial cladode number also
accounted for a significant proportion of variation in attack frequency (F=80.82,
p=0.000), but the effect size was smaller than that of host species (η2=0.123).
For both sets of plants analyzed, the variables attack frequency, host species and
attack frequency*host species were retained in the logistic regression model as significant
predictors of final survival status (Table 4.4). Initial cladode number and attack
frequency*initial cladode number were not found to be significant predictors of final
survival status. Partial coefficients (β, Table 4.4) indicated that the predicted log odds of
survival decrease as attack frequency increases, and that this effect is much greater for O.
stricta than for O. humifusa.

104
Q3:For surviving plants, does C. cactorum attack affect growth rate?
The mean PC/G for all plants of both species was -0.015 ± 0.004, meaning plants lost on
average 1.5% of their initial cladodes per generation of C. cactorum. For surviving plants
of both species, the mean PC/G was 0.016 ± 0.005, meaning surviving plants added about
1.6% of their initial cladodes per generation of C. cactorum. Surviving O. humifusa had a
higher (more positive) mean PC/G than surviving O. stricta (Fig. 4.3), but this difference
was not significant (t=1.01, p=0.311). Surviving attacked plants had a higher (more
positive) mean PC/G than unattacked plants (Fig. 4.3), but again, this difference was not
significant (Welch‟s t=1.80, p=0.074). Multiple regression models showed that both
ln(initial cladode number) and attack frequency were significant predictors of ln(PC/G)
for surviving plants (Table 4.5); results were similar for O. humifusa (Fig.4.4) and O.
stricta (Fig.4.5). Coefficients indicated that proportional growth rate of survivors (as
defined by PC/G), decreased as initial cladode number increased and increased as attack
frequency increased. However, standardized coefficients, partial correlations, and partial
regression plots revealed that initial cladode number was the stronger predictor of the two
(Table 4.5, Figs. 4.4 and 4.5).

105
Table 4.1 Summary of attack and survival rates for each survey year.
Survey year Start N Number attacked (%) Number Surviving (%)
1 580 180 (31.0) 573 (98.8)
2 573 238 (41.5) 564 (98.4)
3 564 184 (32.6) 496 (87.9)
4 459 a 124 (27.0) 442 (96.3)
5 442 170 (38.5) 424 (95.9)
6 424 169 (39.9) 403 (95.0)
Finalb 580 453 (78.1) 440 (75.9)
a The lower „Start N‟ for survey year 4 reflects that fact that some plants were only
surveyed for three years (see Methods)
b„Final‟ row displays the total number of plants that were attacked at least once by C.
cactorum during the study and the total number of plants that survived for all years they
were surveyed

106
Figure 4.1: Histogram of C. cactorum attack and survival rates. Data are from those
plants whose fate was known at the end of the study (n=532)

107
Figure 4.2: The proportion of plants that died each survey year grouped by C. cactorum
attack status for that same year
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
1 2 3 4 5 6
Pro
port
ion
of
dea
d p
lan
ts
Survey year
Attacked
Unattacked

108
Table 4.2 Results from likelihood ratio tests
Sample Variables tested df Species G2, p Theil‟s U
a Relative risk
b
1: All plants with known
final survival status
(N=532)
Final attack status,
final survival status,
species
1
O. humifusa 7.19, 0.007 0.035 3.28 c
O. stricta 0.068, 0.795 0.000 1.12c
Total 13.52, 0.000 0.028 2.96
2: Only plants with
positive final attack status
and known final survival
status (N=415)
Final survival status
and species 1 --- 14.57, 0.000 0.035 0.465
a Theil‟s U, also called the uncertainty coefficient, is a directional measure representing a percent reduction in uncertainty in
predicting one variable when the value of a second variable is known. For Test 1 it represents the reduction in error in
predicting final survival status if final attack status is known. For Test 2 it represents the reduction in error in predicting final
survival status for an attacked plant if species is known
b Relative risk is a ratio of the probabilities of an event. For Test 1 it represents the ratio of the probability of death for an
attacked plant to an unattacked plant. For Test 2, it represents the ratio of the probability of death after a C. cactorum attack for
O. humifusa to O. stricta
c A test for homogeneity of the relative risk between species was non-significant (Tarone‟s X
2=2.04, p=0.153)

109
Table 4.3 Results from log linear models
Sample Predictor Parameter estimate a SE Z p 95% CI
1: All plants with known
final survival status (N=532)
Survival*Attack 0.857 b 0.400 2.14 0.032 0.073 − 1.64
Species*Survival -1.07 b 0.248 -4.29 0.000 -1.55 − -0.578
Species*Attack -1.66 b 0.290 -5.74 0.000 -2.30 − -1.09
2: Only plants with positive
final attack status and known
final survival status (N=415)
Species*Survival -0.945 c 0.255 -3.71 0.000 -1.45 − -0.445
a Only coefficients for the interaction terms are presented, as lower order coefficients are conditional upon reference categories
and therefore, more difficult to interpret. The interaction coefficient is an estimate of a conditional log odds ratio.
b Interpretation of parameter estimates for Model 1: Survival*Attack = the log of the ratio of odds of dying for an attacked
versus an unattacked plant; Species*Survival = the log of the ratio of odds of dying for O. humifusa versus O. stricta;
Species*Attack = the log of the ratio of odds of experiencing at least one attack for O. humifusa versus O. stricta
c Interpretation of parameter estimate for Model 2: Species*Survival = the log of the ratio of odds of dying for an attacked O.
humifusa versus an attacked O. stricta

110
Table 4.4 Results from logistic regression models with final survival status as the dependent variable
Sample
Hosmer-Lemeshow
goodness of fit test Nagelkerke R2b
Predictor β SE pc Exp(β)
Χ2 df p
a
1: All plants with known
final survival status
(N=532)
2.29 7 0.942 0.138
Attack frequency -3.79 0.962 0.000 0.023
Host species -2.19 0.575 0.000 0.113
Attack frequency*host
species 3.15 1.20 0.009 23.34
2: Only plants with
positive final attack status
and known final survival
status (N=415)
10.35 8 0.241 0.097
Attack frequency -4.54 1.39 0.001 0.011
Host species -2.45 0.836 0.003 0.087
Attack frequency*host
species 3.70 1.66 0.025 40.55
a A Hosmer-Lemeshow statistic with p≤0.05 indicates a poor fit for the data
b The Nagelkerke R
2 is a pseudo-R
2 statistic which attempts to estimate a coefficient of determination. It has a scale of 0 (the
predictor variables do not explain any of the variation in the dependent variable) to 1 (the predictor variables perfectly explain
variation in dependent variable)
c Significance of the predictors is based upon the Wald statistic, which is the partial coefficient (β) divided by its SE and
follows a Χ2 distribution

111
Figure 4.3 Mean proportional change in initial cladode number per generation of C.
cactorum for all surviving plants. n=440, error bars represent ± 1 SE.

112
Table 4.5 Results from multiple regressions of ln(proportional cladode change/generation of C. cactorum) on attack frequency
and ln(initial cladode number)
Sample R2 F, p Tolerance
a Coefficients B
b SE β
c p
Partial
correlation
All surviving plants
(n=440) 0.204
55.88,
0.000 0.881
Constant 0.130 0.012 0.000
ln(Initial cladode
number) - 0.040
d 0.004 - 0.481 0.000 - 0.452
Attack frequency 0.073
e 0.024 0.140 0.002 0.146
Surviving O. humifusa
(n=284) 0.210
37.12,
0.000 0.863
Constant 0.131 0.015 .000
ln(Initial cladode
number) - 0.040
d 0.005 - 0.493 0.000 - 0.458
Attack frequency 0.120
e 0.037 0.183 0.002 0.187
Surviving O. stricta
(n=156) 0.250
25.42,
0.000 0.780
Constant 0.136 0.020 0.000
ln(Initial cladode
number) - 0.050
d 0.007 - 0.566 0.000 - 0.500
Attack frequency 0.122
e 0.040 0.244 0.003 0.215
a Tolerance is an indication of the proportion of variance in a predictor that cannot be explained by other predictors; values
<0.20 indicate problematic collinearity
b Unstandardized coefficients
c Standardized coefficients
d Due to ln transformations, B coefficients are interpreted as the % change in the dependent variable for a 1% increase in the
predictor variable
e Due to ln transformation of the dependent variable, B coefficients are interpreted as the B*100% change in the dependent
variable for a one unit change in the predictor

113
a) b)
Figure 4.4: Partial regression plots for O. humifusa. Plots are of a multiple regression of ln(proportional cladode
change/generation of C. cactorum) on (a) attack frequency and (b) ln(initial cladode number) and indicate that initial cladode
number is a stronger predictor of proportional growth rate than attack frequency.

114
a) b)
Figure 4.5: Partial regression plots for O. stricta. Plots are of a multiple regression of ln(proportional cladode
change/generation of C. cactorum) on (a) attack frequency and (b) ln(initial cladode number) and indicate that initial cladode
number is a stronger predictor of proportional growth rate than attack frequency.

115
Discussion
Among scientists, a common argument is that we should adapt a “guilty until proven
innocent” stance on non-native species (Ruesink et al. 1995, Mack et al. 2000, Van
Driesche and Van Driesche 2001, Simberloff 2003). However, others argue that the
assumption that alien status implies harmful effects is misplaced, and that we need to be
more objective in our terminology, assessment of, and policies concerning non-natives
(Brown and Sax 2004, Colautti and MacIsaac 2004, Schlaepfer et al. 2011). Lively
debate over how we should define and regulate non-native and invasive species, and
whether they generally have harmful effects, has ensued over recent years (Brown and
Sax 2005, Cassey et al. 2005, Sagoff 2005, Simberloff 2005, Brown et al. 2007, Larson
2007). Many invasion biologists are now pushing for a system in which evaluation of
species is based more upon documented species‟ impacts, and less on native versus non-
native status (Davis et al. 2011). Such a principle seems to be particularly relevant in
light of the rapid changes to ecosystems caused by everything from local land use change
to global climate change. However, policies and programs based on species‟ impacts in
the past may not be appropriate for the current era, and those based on impacts in one
region may not work well in another region, especially if conclusions about said impacts
are drawn mainly from qualitative observations or inconclusive data (Hager and McCoy
1998). Variation in the establishment and efficacy of biological control agents clearly
demonstrates this notion (McFadyen 1998).
Habitat differences are an evident explanation for the varying success of non-
native species, biological control agent or otherwise. Both abiotic factors, for example,
distance to urban areas (Niemela and Spence 1991, Holway et al. 2002), soil moisture

116
(Holway et al. 2002), and stream discharge (Pintor and Sih 2011), and biotic factors, for
example, species diversity (Morrison and Porter 2003) and prey biomass (Pintor and Sih
2011), have been found to correlate with variation in the density of invasive species. The
lack of effective control of Opuntia by C. cactorum in South Africa can be at least
partially explained by higher levels of predation than in Australia, both by ants
(Robertson and Hoffmann 1989) and baboons (Pettey 1948, Hoffmann et al. 1998).
Variation in the success of non-native species can also result from differing source
populations, which can display variable fecundity (Phillips and Baird 2001) or abiotic
tolerances (Romeis et al. 2005), among other differences. Often, explanations will be
more complex. Shea et al (2005) found that population growth of invasive Carduus
nutans (musk thistle) in New Zealand is driven by early life stage fecundity, while in
Australia it is driven by survival of later life stages, even though the plant spread to
Australia from New Zealand. Differences in life history may explain the lack of success
of Rhinocyllus conicus (seed head weevil) in Australia as compared to North America,
and lead to the prediction of more effective thistle control by Urophora solstitialis (a
seed-reducing agent) in New Zealand and by Trichosirocalus horridus (a rosette feeder)
in Australia (Shea et al. 2005).
We found that C. cactorum attack and Opuntia survival were statistically
independent when each survey year was analyzed separately, but as the detrimental
effects of attack may be delayed, examining attack and survival status in terms of plants‟
final status after six years probably gives a better indication of their relationship. When
doing so, we found that C. cactorum-attacked plants had a higher risk of death, and this
risk increased with increasing frequency of attack. Encouragingly, plants were still more

117
likely to survive than die over the six years, and when they did survive, most showed
positive cladode growth, in agreement with findings by Johnson and Stiling (1998). This
suggests that there may be substantial variation in the Opuntia population for tolerance to
C. cactorum attack.
Our finding that O. stricta is more likely to be attacked and to experience a higher
attack frequency than O. humifusa is not unique, but our larger sample size and longer
study duration provide superior support of the notion that O. stricta is the „preferred‟ of
these two hosts of C. cactorum. More novel is the fact that even when an O. humifusa
was attacked, it was more likely to survive than an attacked O. stricta. Together, these
findings indicate that host species differ not only in resistance to C. cactorum attack, but
also in tolerance to attack. Therefore, local and regional variation in the composition and
density of various host communities may lead to dramatic differences in the effects of C.
cactorum in North America.
We also found interesting relationships between plant size, C. cactorum attack,
and plant survival. Plants with a larger initial cladode number had a higher attack
frequency; this result is expected and agrees with findings of Pemberton and Liu (2007)
and Johnson and Stiling (1998). However, we also found a negative relationship between
cladode number and growth rate of surviving plants, such that larger plants lost
proportionally more cladodes than smaller plants. This is in contrast to Pemberton and
Liu (2007) who found that larger plants have proportionally fewer cladodes attacked by
C. cactorum. Despite this, initial cladode number was not found to predict survival of our
plants.

118
Perhaps most surprising were our findings concerning Opuntia growth rate.
Surviving plants that were attacked by C. cactorum added proportionally more cladodes
than those that were unattacked, although this difference was not significant, and there
was a significant positive relationship between attack frequency and proportional cladode
change. One could argue that this is a by-product of the positive correlation between
plant size and attack frequency; C. cactorum may selectively attack larger plants because
they are growing more vigorously (Strauss 1988), leading to an apparent higher growth
rate for attacked plants. However, we found the correlation between size and attack
frequency to be relatively weak. Additionally, as stated above, larger surviving plants
actually lost proportionally more cladodes when controlling for attack frequency (Table
4.5), leading us to conclude that the positive relationship between attack frequency and
growth rate is not driven by initial plant size. This raises the intriguing possibility of
compensatory growth as a topic for further study.
Given the widespread destruction of O. stricta in Australia and the Caribbean, our
finding of high survival rates (87% for O. humifusa, 62% for O. stricta) in conjunction
with high C. cactorum attack rates may seem unexpected. However, a closer look at the
situation in these regions decades after the release of C. cactorum reveals that our results
may be less surprising than at first glance Although C. cactorum provided effective
control of Opuntia over an enormous area of Queensland, it proved ineffective in the
southwest and coastal areas (Monro 1975, White 1980) and in New South Wales
(Hosking et al. 1994). White (1980) proposed that the dense stands of Opuntia in these
areas are largely resistant to C. cactorum because they are under water and nutrient
stress, resulting in lower plant quality and higher levels of mucilage than unstressed

119
plants. It has also been suggested that loss of female moths through dispersal could
provide an explanation for the lack of control on small coastal islands off of Australia
and Mauritius (Monro 1975), although to our knowledge this has not been further
explored.
A survey of the Caribbean islands of Nevis and St. Kitts nearly 50 years after the
initial release of C. cactorum exposed a similar situation (Pemberton and Liu 2007).
Opuntia triacantha and O. stricta populations have been greatly reduced, but not
eradicated, even though C. cactorum remains established on the islands. A previous
survey (Bennett and Simmonds 1966) found plants to be very scarce and small, but
Pemberton and Liu (2007) found that plants were “abundant enough to be easily located”
and documented plants with up to 100 cladodes. The authors state that C. cactorum
populations appear lower today than they were soon after its introduction, allowing some
Opuntia to escape attack and recover, which again emphasizes the importance of
assessing data from multiple points in time (or from longer term studies) before drawing
definitive conclusions on species‟ effects.
Our objective is not to trivialize the threat of C. cactorum. It is obvious that it
could extirpate rare or less resistant opuntioid species, and even low mortality rates of
more common species could have complex, negative effects on the community. We
acknowledge that without data on plant reproduction and recruitment, definite statements
on the long term viability of these Opuntia populations cannot be made. We also
acknowledge that as conclusions about an invasive species‟ effects often change through
space or time, our results may not hold true for other North American regions, or for
studies done in west Florida ten years from now. In light of this we adamantly support

120
control and regulation policies of C. cactorum. Still, we feel our study provides some
support of Pemberton and Liu‟s (2007) hopeful prediction that the effects of C. cactorum
in North America will not necessarily be catastrophic.
Acknowledgements
We would like to thank Debi Tharp and Kerry Bohl for their enormous contribution to
data collection, Terry Hingtgen (Florida Department of Environmental Protection,
Southwest Division) for his assistance in acquiring the necessary permits for field work
at Honeymoon Island State Park, and the employees of Fort deSoto County Park and
Lido Key beaches for their support during the course of the study.
References cited
Baker, A. J. and P. Stiling. 2009. Comparing the effects of the exotic cactus-feeding
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), and the native
cactus-feeding moth, Melitara prodenialis (Walker) (Lepidoptera: Pyralidae), on
two species of Florida Opuntia. Biological Invasions 11:619-624.
Bennett, F. D. and F. J. Simmonds. 1966. Biological control of Opuntia spp. by
Cactoblastis cactorum in the Leeward Islands (West Indies). BioControl 11:183-
189.
Brown, J. H. and D. F. Sax. 2004. An essay on some topics concerning invasive species.
Austral Ecology 29:530-536.
Brown, J. H. and D. F. Sax. 2005. Biological invasions and scientific objectivity: Reply
to Cassey et al. (2005). Austral Ecology 30:481-483.
Brown, J. H., D. F. Sax, D. Simberloff, and M. Sagoff. 2007. Aliens among us.
Conservation Magazine 8:14-21.
Cassey, P., T. M. Blackburn, R. P. Duncan, and S. L. Chown. 2005. Concerning invasive
species: reply to Brown and Sax. Austral Ecology 30:475-480.

121
Colautti, R. I. and H. J. MacIsaac. 2004. A neutral terminology to define 'invasive'
species. Diversity and Distributions 10:135-141.
Davis, M., M. K. Chew, R. J. Hobbs, A. E. Lugo, J. J. Ewel, G. J. Vermeij, J. H. Brown,
M. L. Rosenzweig, M. R. Gardener, S. P. Carroll, K. Thompson, S. T. A. Pickett,
J. C. Stromberg, P. Del Tredici, K. N. Suding, J. G. Ehrenfeld, J. P. Grime, J.
Mascaro, and J. C. Briggs. 2011. Don't judge species on their origins. Nature
474:153-154.
Denslow, J. S. and C. M. D'Antonio. 2005. After biocontrol: assessing indirect effects of
insect releases. Biological Control 35:307-318.
Dodd, A. P. 1940. The biological campaign against prickly pear. Commonwealth Prickly
Pear Board Bulletin 1-177. Brisbane, Australia.
Hager, H. A. and K. D. McCoy. 1998. The implications of accepting untested
hypotheses: a review of the effects of purple loosestrife (Lythrum salicaria) in
North America. Biodiversity and Conservation 7:1069-1079.
Hight, S. D. and J. E. Carpenter. 2009. Flight phenology of male Cactoblastis cactorum
(Lepidoptera: Pyralidae) at different latitudes in the southeastern United States.
Florida Entomologist 92:208-216.
Hoffmann, J. H., V. C. Moran, and D. A. Zeller. 1998. Evaluation of Cactoblastis
cactorum (Lepidoptera : Phycitidae) as a biological control agent of Opuntia
stricta (Cactaceae) in the Kruger National Park, South Africa. Biological Control
12:20-24.
Hoffmann, J. H., V. C. Moran, and H. G. Zimmermann. 1999. Integrated management of
Opuntia stricta (Haworth) Haworth (Cactaceae) in South Africa: an enhanced
role for two renowned insect agents. African Entomology:15-20.
Holway, D. A., A. V. Suarez, and T. J. Case. 2002. Role of abiotic factors in governing
susceptibility to invasion: A test with argentine ants. Ecology 83:1610-1619.
Hosking, J. R., P. R. Sullivan, and S. M. Welsby. 1994. Biological control of Opuntia
stricta (Haw) Haw var. stricta using Dactylopius opuntiae (Cockerell) in an area
of New South Wales, Australia, where Cactoblastis cactorum (Berg) is not a
successful biological control agent. Agriculture Ecosystems & Environment
48:241-255.
Jezorek, H. A., P. D. Stiling, and J. E. Carpenter. 2010. Targets of an invasive species:
oviposition preference and larval performance of Cactoblastis cactorum
(Lepidoptera: Pyralidae) on 14 North American opuntioid cacti. Environmental
Entomology 39:1884-1892.

122
Johnson, D. M. and P. D. Stiling. 1996. Host specificity of Cactoblastis cactorum
(Lepidoptera: Pyralidae), an exotic Opuntia-feeding moth, in Florida.
Environmental Entomology 25:743-748.
Johnson, D. M. and P. D. Stiling. 1998. Distribution and dispersal of Cactoblastis
cactorum (Lepidoptera : Pyralidae), an exotic Opuntia-feeding moth, in Florida.
Florida Entomologist 81:12-22.
Kenis, M., M. A. Auger-Rozenberg, A. Roques, L. Timms, C. Pere, M. Cock, J. Settele,
S. Augustin, and C. Lopez-Vaamonde. 2009. Ecological effects of invasive alien
insects. Biological Invasions 11:21-45.
Larson, B. M. H. 2007. An alien approach to invasive species: Objectivity and society in
invasion biology. Biological Invasions 9:947-956.
Mack, R. N., D. Simberloff, W. M. Lonsdale, H. Evans, M. Clout, and F. A. Bazzaz.
2000. Biotic invasions: Causes, epidemiology, global consequences, and control.
Ecological Applications 10:689-710.
McFadyen, R. E. C. 1998. Biological control of weeds. Annual Review of Entomology
43:369-393.
Monro, J. M. 1975. Environmental variation and the efficiency of biological control -
Cactoblastis in the southern hemisphere. Pages 204-212 in Kikkawa, J. and H.A.
Nix (Eds.). Proceedings of the Ecological Society of Australia, Vol. 9. Managing
Terrestrial Ecosystems. Symposium. Brisbane, Australia., May 15-16, 1975.
Morrison, L. W. 2002. Long-term impacts of an arthropod-community invasion by the
imported fire ant, Solenopsis invicta. Ecology 83:2337-2345.
Morrison, L. W. and S. D. Porter. 2003. Positive association between densities of the red
imported fire ant, Solenopsis invicta (Hymenoptera : Formicidae), and
generalized ant and arthropod diversity. Environmental Entomology 32:548-554.
Niemela, J. and J. R. Spence. 1991. Distribution and abundance of an exotic ground
beetle (Carabidae) - a test of community impact. Oikos 62:351-359.
Parker, I. M., D. Simberloff, W. M. Lonsdale, K. Goodell, M. Wonham, P. M. Kareiva,
M. H. Williamson, B. Von Holle, P. B. Moyle, J. E. Byers, and L. Goldwasser.
1999. Impact: toward a framework for understanding the ecological effects of
invaders. Biological Invasions 1:3-19.
Pemberton, R. W. and H. Liu. 2007. Control and persistence of native Opuntia on Nevis
and St. Kitts 50 years after the introduction of Cactoblastis cactorum. Biological
Control 41:272-282.

123
Pettey, F. W. 1948. The biological control of prickly pear in South Africa. Science
Bulletin. Department of Agriculture of the Union of South Africa.
Phillips, C. B. and D. B. Baird. 2001. Geographic variation in egg load of Microctonus
hyperodae (Hymenoptera : Braconidae) and its implications for biological control
success. Biocontrol Science and Technology 11:371-380.
Pintor, L. M. and A. Sih. 2011. Scale dependent effects of native prey diversity, prey
biomass and natural disturbance on the invasion success of an exotic predator.
Biological Invasions 13:1357-1366.
Porter, S. D. and D. A. Savignano. 1990. Invasion of polygene fire ants decimates native
ants and disrupts arthropod community. Ecology 71:2095-2106.
Robertson, H. G. and J. H. Hoffmann. 1989. Mortality and life-tables of Cactoblastis
cactorum (Berg) (Lepidoptera: Pyralidae) compared on 2 host-plant species.
Bulletin of Entomological Research 79:7-17.
Romeis, J., D. Babendreier, F. L. Wackers, and T. G. Shanower. 2005. Habitat and plant
specificity of Trichogramma egg parasitoids: Underlying mechanisms and
implications. Basic and Applied Ecology 6:215-236.
Rose, R., R. Weeks, and S. Usnick. 2011. Cactus moth, Cactoblastis cactorum, 2011
survey plan for PPQ and state cooperators. United States Department of
Agriculture, Animal and Plant Health Inspection Service.
Ruesink, J. L., I. M. Parker, M. J. Groom, and P. M. Kareiva. 1995. Reducing the risks of
nonindigenous species introductions - guilty until proven innocent. Bioscience
45:465-477.
Sagoff, M. 2005. Do non-native species threaten the natural environment? Journal of
Agricultural & Environmental Ethics 18:215-236.
Schlaepfer, M. A., M. Pascal, and M. A. Davis. 2011. How might science misdirect
policy? Insights into the threats and consequences of invasive species. Journal of
Consumer Protection and Food Safety 6:27-31.
Shea, K., D. Kelly, A. W. Sheppard, and T. L. Woodburn. 2005. Context-dependent
biological control of an invasive thistle. Ecology 86:3174-3181.
Simberloff, D. 2003. Confronting introduced species: A form of xenophobia? Biological
Invasions 5:179-192.
Simberloff, D. 2005. Non-native species do threaten the natural environment! Journal of
Agricultural & Environmental Ethics 18:595-607.
Snyder, W. E. and E. W. Evans. 2006. Ecological effects of invasive arthropod generalist
predators. Annual Review of Ecology Evolution and Systematics 37:95-122.

124
Stiling, P. 2010. Death and decline of a rare cactus in Florida. Castanea 75:190-197.
Strauss, S. Y. 1988. Determining the effects of herbivory using naturally damaged plants.
Ecology 69:1628-1630.
Strayer, D. L., V. T. Eviner, J. M. Jeschke, and M. L. Pace. 2006. Understanding the
long-term effects of species invasions. Trends in Ecology & Evolution 21:645-
651.
Van Driesche, J. and R. Van Driesche. 2001. Guilty until proven innocent: Preventing
nonnative species invasions. Conservation in Practice 2:8-18.
White, G. C. 1980. Current status of prickly pear control by Cactoblastis cactorum in
Queensland. Pages 609-616 in Proceedings of the Vth International Symposium
on Biological Control of Weeds, Brisbane, Australia.
Wilson, K. A., J. J. Magnuson, D. M. Lodge, A. M. Hill, T. K. Kratz, W. L. Perry, and T.
V. Willis. 2004. A long-term rusty crayfish (Orconectes rusticus) invasion:
Dispersal patterns and community change in a north temperate lake. Canadian
Journal of Fisheries and Aquatic Sciences 61:2255-2266.
Zimmermann, H. G., S. Bloem, and H. Klein. 2004. Biology, history, threat, surveillance
and control of the cactus moth, Cactoblastis cactorum. International Atomic
Energy Agency/Food and Agriculture Organization, Vienna, Austria.
Zimmermann, H. G., V. C. Moran, and J. H. Hoffmann. 2000. The renowned cactus
moth, Cactoblastis cactorum (Lepidoptera : Pyralidae): its natural history and
threat to native Opuntia floras in Mexico and the United States of America.
Florida Entomologist 84:543-551.

125
CHAPTER 5
LACK OF ASSOCIATIONAL EFFECTS BETWEEN TWO COMMON HOSTS ON
ATTACK RATES BY CACTOBLASTIS CACTORUM
Synopsis
Associational susceptibility occurs when herbivory on a target plant is increased due to
the influence of plant neighbors. It has been suggested that associational susceptibility
often arises when a plant neighbor is more palatable to a particular herbivore, resulting in
attraction and spill over of the herbivore onto the target plant. We tested this prediction
on a two common hosts of Cactoblastis cactorum in Florida: Opuntia humifusa,
generally considered less preferred by C. cactorum, and O. stricta, considered to be the
preferred host. We used a combination of observational surveys and experimental
plantings to test for differences in C. cactorum and native herbivore damage and cladode
growth on isolated O. humifusa plants, O. humifusa in interspecific pairs, and O.
humifusa in conspecific pairs. We found that O.stricta had significantly more cladodes
with C. cactorum larvae, eggsticks, and old damage than O. humifusa, lending support to
our assumption that it is preferred by C. cactorum over O. humifusa. Three of the four
native herbivores tested were found at higher levels on O.humifusa than O.stricta,
suggesting that O.humifusa may experience decreased damage from these herbivores,
that is, associational resistance, when paired with O. stricta. However, we found little
support for associational resistance or susceptibility. Opuntia humifusa plants in

126
interspecific pairs did have more C. cactorum larvae and old damage than plants in
conspecific pairs, but did not have more than isolated O. humifusa plants, meaning this
result could be driven by density, as opposed to diversity, of host plants. Plants from the
site used for the experimental plantings had little to no C. cactorum damage, regardless
of plant neighbors, and revealed no associational effects for native herbivores. We
believe that the lack of support for associational susceptibility may be because a spillover
effect requires higher densities of herbivores, and lower densities of their preferred host
species, than were present at our sites.
Introduction
The risk of herbivory for a plant is influenced by the diversity and spatial distribution of
its surrounding plant community. A host plant may gain protection from its herbivores
when found in a diverse, versus simple, community or when growing in close association
with non-host or less palatable species. This phenomenon is termed „associational
resistance‟ (hereafter AR) and was first used to describe reduced herbivory on crops
grown in polycultures versus those grown in monocultures (Tahvanainen and Root 1972,
Root 1973). AR was later extended to include reduced herbivory, predation, or parasitism
when a focal host/prey is associated with a more resistant or less preferred host/prey
(Wahl and Hay 1995, Hjalten and Price 1997, Ostfeld and Keesing 2000, LoGiudice et
al. 2003, Stiling et al. 2003, Stenberg et al. 2007, Raffel et al. 2008). Conversely, if a
focal host/prey experiences increased herbivory, predation, or parasitism due to
association with a less resistant or more preferred host/prey, the phenomenon is termed

127
„associational susceptibility‟ (hereafter AS) (Thomas 1986, Brown and Ewel 1987, Wahl
and Hay 1995, White and Whitham 2000). Multiple mechanisms and hypotheses have
been proposed to explain AR and AS, including the resource concentration hypothesis
(Root 1973), enemies hypothesis (Root 1973), plant defense guilds (Tahvanainen and
Root 1972, Atsatt and Odowd 1976), plant „eavesdropping‟ (Karban 2001, 2007,
Himanen et al. 2010), shared natural enemies (or apparent competition) (Holt and
Lawton 1994), and biotic or abiotic habitat modification (White and Andow 2006,
Barbosa et al. 2009); these mechanisms and hypotheses have been reviewed by Russell
(1989), Andow (1991), Agrawal et al (2006), Barbosa et al (2009), and Letourneau et al
(2011).
Many of the mechanisms listed above could, in theory, lead to either AR or AS.
For example, a plant neighbor that serves as an attractant could reduce damage on the
focal plant by drawing away shared herbivores (AR). This is the conceptual basis for trap
cropping in agricultural systems, which involves mixing attractive, but less economically
valuable, plant species with crops in order to lure herbivores away from the crops.
However, if an herbivore species increases in abundance due to feeding on the attractive
neighbor(s), spill over onto the focal plant could occur, thereby increasing damage (AS).
In general, AR is more likely when a neighbor is less palatable relative to the
focal plant (Pfister and Hay 1988, Hjalten and Price 1997, Barbosa et al. 2009), while AS
is more likely when a neighbor is more palatable (Brown and Ewel 1987, White and
Whitham 2000). For example, White and Whitman (2000) found significantly higher
cankerworm densities and percentage defoliation on potted cottonwoods placed under the
worm‟s preferred host, box elder, then on isolated cottonwoods or cottonwoods placed

128
under conspecifics. Agrawal (2006) refines this prediction for specialist herbivores, by
observing that for specialists, AS should become more likely as plant neighbors offer
increasingly essential resources. However, actual occurrence of AR or AS will clearly be
linked to other factors including herbivore density and motility (Brown and Ewel 1987,
White and Whitham 2000, Russell et al. 2007), presence of other herbivores and natural
enemies (Stiling et al. 2003, Hamback et al. 2006, White and Andow 2006) and distance
between focal and neighbor plants (White and Whitham 2000, Stiling and Moon 2001,
Stiling et al. 2004, Russell et al. 2007).
The primary objective of this study was to determine whether Opuntia humifusa,
a host of the invasive cactus moth, Cactoblastis cactorum Berg (Lepidoptera: Pyralidae),
experiences any associational effects when growing near to O. stricta, a second host that
is generally considered more preferred than O. humifusa (Baker and Stiling 2009). For
insect herbivores, AS has been more commonly observed than AR (Barbosa et al. 2009),
and as described above, is generally expected when a neighbor is more preferred than a
focal plant. Cactoblastis cactorum females normally disperse very little and tend to
oviposit very near to their emergence site, leading to a clumped distribution of eggsticks
(Dodd 1940, Myers et al. 1981, Robertson 1987b, Zimmermann et al. 2004). The general
observations and expectations for AS, coupled with the specific behaviors of C.
cactorum females, suggest that spill over from O. stricta is likely to lead to AS for O.
humifusa growing in close proximity. We therefore predicted that O. humifusa would
have higher levels of C. cactorum damage when growing near O. stricta, as compared to
damage levels when growing near conspecifics or isolated. Our secondary objective was
to determine if there were differences in damage levels or abundance of native herbivores

129
that could be attributed to associational effects of the two Opuntia species. We examined
four native herbivores: Chelinidea vittiger, Dactylopius confusus, Diaspis echinocacti,
and Gerstaeckeria hubbardi (Table 5.1). Preferences for O. humifusa versus O. stricta
have not been documented for any of these species, therefore specific predictions
concerning the occurrence of AR or AS were not made. In order to address our two
objectives, we conducted both observational surveys and experimental plantings.
Methods
Study system
Cactoblastis cactorum Berg (Lepidoptera: Pyralidae), is well known for its role in the
biological control of Opuntia species, including the highly successful campaign in
Australia during the 1920s and 1930s, the less successful campaign begun in the late
1930s in South Africa, and the campaign against native cacti in the Caribbean begun in
the 1950s (Dodd 1940, Pettey 1948, Bennett and Simmonds 1966, Zimmermann and
Moran 1991). However, since its discovery in the Florida Keys in 1989, the moth has
become a potential threat to North American opuntioid biodiversity (Bennett and Habeck
1995, Zimmermann et al. 2000, Stiling 2002). Presently, C. cactorum is established in
the southeastern United States as far north as Bull Island, South Carolina and as far west
as southeastern Louisiana (Simonsen et al. 2008, USDA-APHIS-PPQ 2009). Female
moths lay about 40-100 eggs one on top of the other to form an eggstick that is attached
to the tip of a cactus spine or directly to a cladode. The larvae feed internally and
gregariously on a number of opuntioid species. Larval damage results in hollowed out
cladodes and can lead to secondary infections and in some cases, the death of entire

130
plants (Zimmermann et al. 2004). After completing six instars, larvae drop to the ground
to pupate in the soil or in hollowed out cladodes. In central Florida, there are three non-
overlapping flight periods per year: a spring flight beginning in mid- February, a summer
flight beginning in early June, and a fall flight beginning in late August (Hight and
Carpenter 2009).
Four native species of Opuntia-feeding insects were included in damage surveys:
C.vittiger, D. confusus, D. echinocacti, and G. hubbardi. Diaspis echinocacti feeds on a
variety of cactus species, while the remaining three species are specialists on opuntioid
cacti (Hunter et al. 1912, Mann 1969). Dactylopius confusus and D. echinocacti are
primarily sessile and live adult stages are therefore easily quantified. Chelinidea vittiger
nymphs and adults are motile and G. hubbardi larvae feed internally, making accurate,
direct counts of these two species more difficult. However, both leave characteristic signs
of feeding (Table 5.1) which allow for quantification of damage even if insects are not
directly observed.
Two Opuntia species native to Florida were used in this study, O. stricta and O.
humifusa. Both are common hosts of C. cactorum, although O. stricta is attacked more
often than O. humifusa (Baker and Stiling 2009). They exhibit slightly different
morphologies; O. stricta has a more erect growth form and more spines, while O.
humifusa has a low, spreading growth form and fewer spines. However, both species can
range in size from a few cladodes to several hundred cladodes.
Surveys were conducted at Honeymoon Island State Park, Pinellas County
(hereafter, HI) and experimental plantings were conducted at Fort deSoto Park, Pinellas
County (hereafter, FDS). Both sites are part of Florida‟s west-central barrier island chain

131
and have similar plant and herbivore communities. O. stricta and O. humifusa are
common in the dune and coastal scrub areas and C. cactorum is well-established.
Data collection and analysis
In September of 2006, 190 Opuntia plants were marked at HI as follows: Neighborhood
type 1 = 50 O. humifusa alone; Neighborhood type 2 = 52 O. stricta alone;
Neighborhood type 3 = 44 interspecific pairs. Plants in “alone” neighborhoods were >5m
from any other Opuntia individual, while plants in “paired” neighborhoods were ≤1m
apart, but >5m from any other Opuntia individuals, with distances measured between the
two closest cladodes of any two plants at the time of marking. All plants were surveyed
in September 2006 and December 2006. At each survey, the following variables were
recorded: number of live, non-woody cladodes, proportion of cladodes with active C.
cactorum larvae, proportion of cladodes with old C. cactorum damage, number of C.
cactorum eggsticks per cladode, and proportion of cladodes with D. confusus, D.
echinocacti (live insects present), C. vittiger, and G. hubbardi ( feeding marks present).
Data were highly non-normal so to check for differences in herbivore damage
between O. stricta and O. humifusa, a Mann-Whitney test was performed with species as
the grouping factor. To check for associational effects on herbivore damage, a non-
parametric ANOVA by ranks was used to compare between plant species/neighborhood
combinations. All analyses were conducted using IBM SPSS Statistics 19 and where
appropriate, Dunn‟s test for multiple contrasts was used for pair wise comparisons.
The design described above allows for testing of O. humifusa versus O. stricta for
cladode number and herbivore damage levels, but does not allow for a comparison to

132
tease out any associational effects due to density of Opuntia, regardless of host species,
versus associational effects due explicitly to the species of Opuntia. For example, if O.
humifusa in interspecific pairs were found to have significantly more C. cactorum larvae
than O. humifusa alone, the effect could be due to a higher density of hosts in the paired
neighborhood or to a specific effect of O. stricta. Therefore a second set of surveys were
conducted at HI in 2009 with 100 Opuntia plants marked as follows: Neighborhood type
1 =20 O. humifusa alone, Neighborhood type 3 = 20 interspecific pairs, and
Neighborhood type 4 =20 conspecific pairs. Distances for “alone” and “paired”
neighborhoods were as described for 2006 surveys. All plants were surveyed in May,
July, and November of 2009 with the same variables as in 2006 recorded, plus maximum
height. Data were again highly non-normal, so similar methods were used for analysis.
Herbivore-related variables were averaged over the three months, but as this survey
encompassed the latter part of the Opuntia growth season, the change in height and
proportional change in cladode number from May to November were used instead of the
average. A non-parametric ANOVA by ranks was used to compare O. humifusa plants
with plant neighborhood as the grouping factor.
Analysis of data in the above manner results in a “snapshot” assessment of
associational effects. However, the time since establishment in a neighborhood was
unknown for each plant and initial damage levels were not controlled for, so changes
over time were not of primary interest for the observational surveys. In order to control
for these factors we used Opuntia plants raised in a greenhouse at the University of South
Florida Botanical Garden for an out planting experiment. Cladodes from FDS were
collected in April of 2007 and a single cladode was planted in a six inch pot using a mix

133
of sand and potting soil. Plants were fertilized twice a year, and watered as needed. By
April of 2009 the experimental (greenhouse raised) plants had multiple cladodes and
were used to create two plant neighborhoods at FDS: Neighborhood type 3 = 18
interspecific groups, each consisting of two experimental O. stricta planted ≤0.5m from
one naturally occurring O. humifusa, and Neighborhood type 4 = 16 conspecific pairs,
each consisting of one experimental O. humifusa planted ≤0.5m from one naturally
occurring O. humifusa. We also marked plants for Neighborhood type 1 = 15 naturally
occurring O. humifusa alone. Experimental plants were of similar size (2-6 cladodes) and
were initially free of damage from C. cactorum, D. confusus, C. vittiger, and G.
hubbardi, but due to an outbreak in the greenhouse, all experimental plants had D.
echinocacti. Naturally occurring plants were initially free of C. cactorum larvae and had
little or no old C. cactorum damage (maximum=2 cladodes with old damage); initial
damage levels of the four native herbivores varied as finding plants free of all types of
native herbivores proved impossible. All plants were surveyed monthly from April 2009
to November 2009, and the same variables as in the 2009 observational survey were
recorded. Although changes over time were of inherent interest, data could not be
transformed to meet assumptions of repeated measures ANOVA. Instead, we calculated
the difference in height, the proportional difference in cladode number, and the
difference in each herbivore-related variable between the April and November surveys. A
non-parametric ANOVA by ranks was used to compare O. humifusa plants with plant
neighborhood as the grouping factor.

134
Results
Mann-Whitney tests of all O. stricta versus all O. humifusa from the 2006 survey showed
that O. stricta plants had significantly higher proportions of cladodes with larvae and old
damage, and more eggsticks per cladode. Opuntia humifusa had higher proportions of
cladodes with C. vittiger, D. confusus, and G. hubbardi, as well as more total cladodes
(Table 5.2). ANOVA by ranks for plant species/neighborhood combinations were
significant for all variables except proportion of cladodes with D. echinocacti, however,
pair wise comparisons revealed that this was driven almost entirely by interspecific
differences. Opuntia stricta alone and O. stricta in pairs were not significantly different
for any variables. The sole intraspecific differences between Opuntia humifusa alone and
O. humifusa in pairs were for total cladodes (p=0.005) and proportion of cladodes with
G. hubbardi (p=0.000). In both cases O. humifusa alone had the highest value of the four
groups and was significantly different than all other groups (Figs 5.1 and 5.2).
The 2009 data were used to test among O. humifusa plants alone, in conspecifc
pairs, and in interspecific pairs. Plants that were in interspecific pairs had the greatest
proportion of cladodes with C. cactorum larvae and old damage (Figs 5.3 and 5.4). The
difference between neighborhoods was significant for old damage (p=0.036), but was
marginal for larvae (p=0.080). Pair wise comparisons for old damage showed that O.
humifusa from interspecific pairs had significantly more old damage than O. humifusa in
conspecific pairs (adjusted p=0.030), but neither neighborhood was significantly
different than O. humifusa alone (Fig 5.4). Opuntia humifusa plants alone added
significantly more height than O. humifusa in interspecific or conspecific pairs (adjusted
p=0.004 for both pair wise comparisons), yet lost proportionally more cladodes than the

135
paired plants, although this result was not significant. No other variables were
significantly different among the three plant neighborhoods.
Cactoblastis cactorum damage levels were extremely low for the FDS site. Over
the seven months, only one plant had active larvae and only one eggstick was found.
Eight plants showed old damage, but half of these were plants with 1-2 cladodes of pre-
existing old damage. Therefore, C. cactorum variables were not included in the analysis.
Results from the ANOVA by ranks indicated significant differences between plant
neighborhoods for change in height, proportional change in cladodes, and change in D.
echinocacti damage (p=0.016, 0.023, and 0.004, respectively). Further examination
revealed that results for the latter two variables were driven solely by differences
between experimentally planted O. humifusa and naturally occurring O. humifusa. The
experimentally planted O. humifusa had a significantly higher proportional change in
cladodes when compared to naturally occurring O. humifusa alone, in conspecific pairs,
and in interspecific pairs (adjusted p=0.000 for all three pair wise comparisons).
Experimental O. humifusa also significantly decreased the proportion of cladodes with D.
echinocati as compared to all three groups of naturally occurring plants (adjusted
p=0.000 for all three pair wise comparisons). Experimental O. humifusa had D.
echinocacti colonies present at the time of planting and the large decrease in cladodes
with D. echinocacti reflects the insects‟ tendency to reach much higher densities in
greenhouse or plantation settings than in naturally occurring Opuntia populations
(Hunter et al. 1912, H. Jezorek personal observation).

136
Table 5.1 The four native Opuntia herbivores examined in the study
Insect species Order: Family Common
Name
Description of feeding/damage
Chelinidea
vittiger McAtee
Hemiptera:
Coridae
Cactus bug Mobile sap sucker; leaves pale half
or full circles on surface of cladode;
severe damage can lead to
desiccation of cladode
Dactylopius
confusus
Cockerell
Hemiptera:
Dactylopiidae
Cochineal Sessile sap sucker; exudes fluffy
white substance on surface of
cladodes
Diaspis
echinocacti
Bouché
Hemiptera:
Diaspididae
Cactus
scale
Sessile sap sucker; circular, slightly
convex, whitish to tan scale; severe
damage causes chlorotic patches and
desiccation of cladode
Gerstaeckeria
hubbardi
LeConte
Coleoptera:
Curculionidae
Cactus
weevil
Internal feeder; leaves light brown
circular cell; cladode appears pierced
by tiny “bullet hole”

137
Table 5.2 Results from a Mann-Whitney test for differences between O. humifusa
(N=94) and O. stricta (N=96). Plants were surveyed in September and December of 2006
and values for all variables were averaged across the two surveys prior to analysis.
Negative values of the standardized test statistic indicate that the variable was greater for
O. humifusa, positive values indicate that the variable was higher for O. stricta.
a Values for these variables were recorded as the proportion of cladodes on a plant
containing the herbivore or its characteristic damage
Variable O. humifusa rank O. stricta rank Standardized test
statistic
p
Total cladodes 104.27 86.91 -2.176 0.030
C. cactorum larvae a 82.45 108.28 4.54 0.000
C. cactorum old damage a 74.80 115.79 5.62 0.000
Eggsticks/cladode 92.50 98.44 2.46 0.014
D. confusus a 107.63 83.62 -5.04 0.000
G. hubbardi a 121.26 70.28 -7.19 0.000
C.vittiger a 121.30 70.23 -6.56 0.000
D. echinocacti a 97.85 93.20 -0.85 0.394
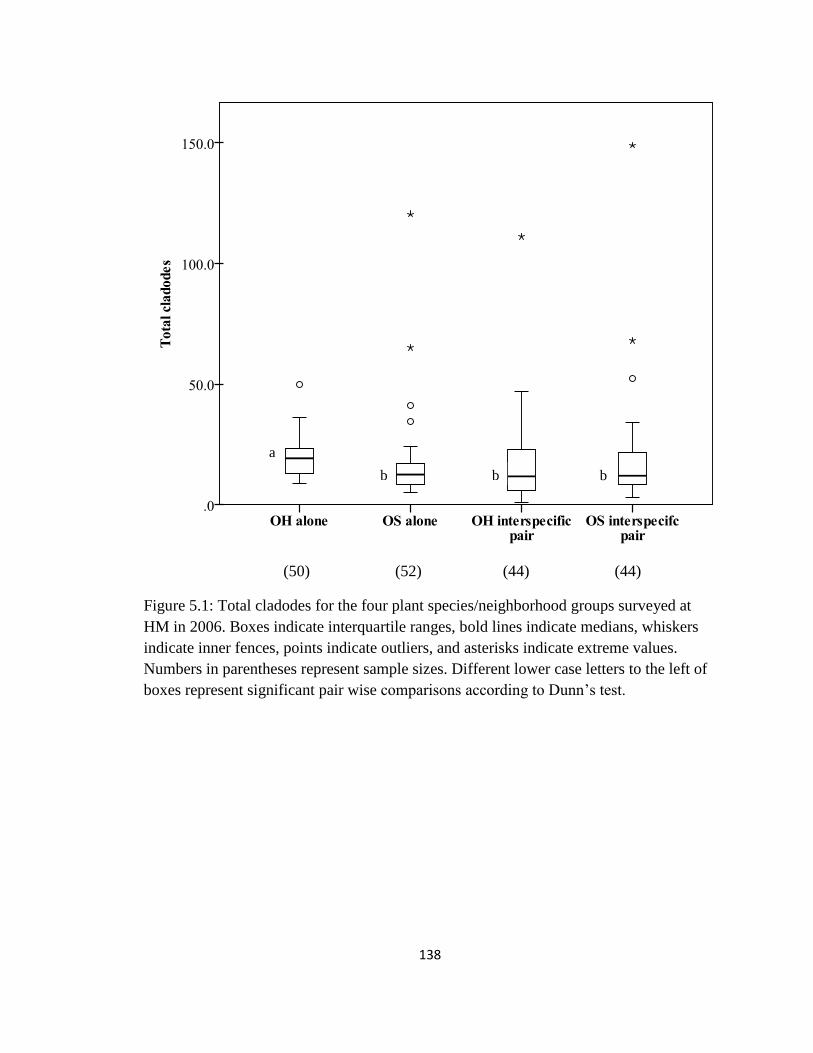
138
(50) (52) (44) (44)
Figure 5.1: Total cladodes for the four plant species/neighborhood groups surveyed at
HM in 2006. Boxes indicate interquartile ranges, bold lines indicate medians, whiskers
indicate inner fences, points indicate outliers, and asterisks indicate extreme values.
Numbers in parentheses represent sample sizes. Different lower case letters to the left of
boxes represent significant pair wise comparisons according to Dunn‟s test.
a
b b b
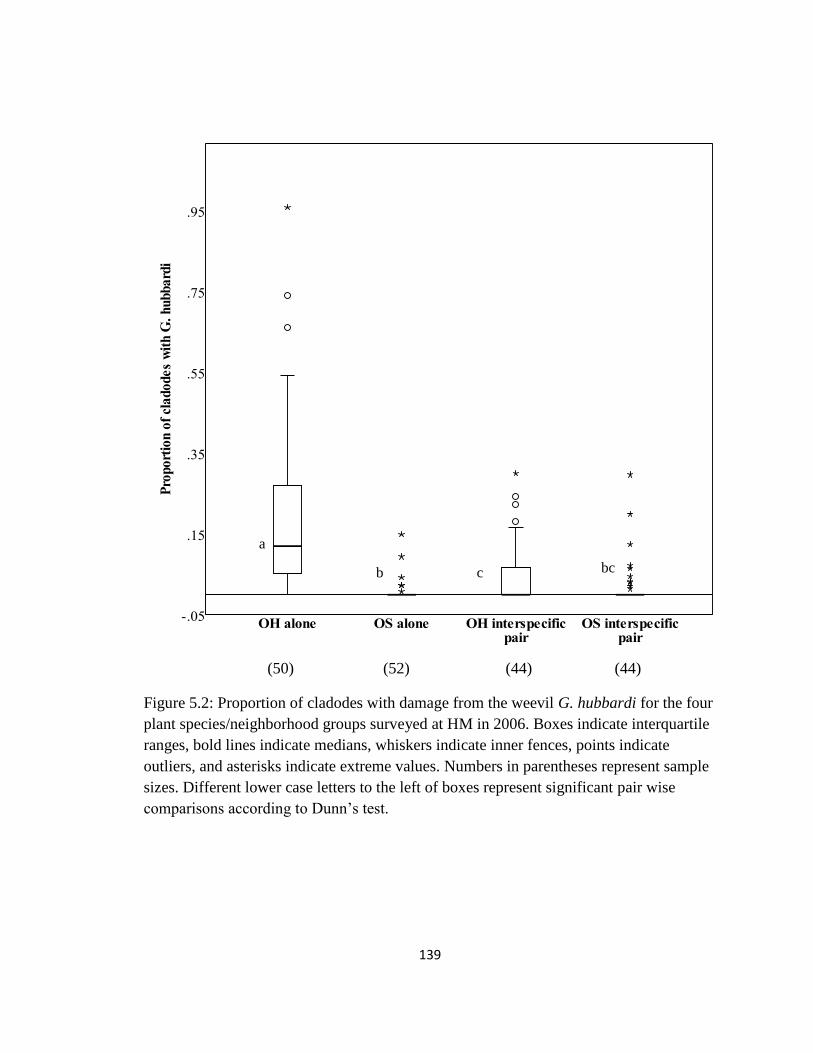
139
(50) (52) (44) (44)
Figure 5.2: Proportion of cladodes with damage from the weevil G. hubbardi for the four
plant species/neighborhood groups surveyed at HM in 2006. Boxes indicate interquartile
ranges, bold lines indicate medians, whiskers indicate inner fences, points indicate
outliers, and asterisks indicate extreme values. Numbers in parentheses represent sample
sizes. Different lower case letters to the left of boxes represent significant pair wise
comparisons according to Dunn‟s test.
b
a
c bc

140
(20) (40) (20)
Figure 5.3: Proportion of cladodes with C. cactorum larvae from the three O. humifusa
neighborhoods surveyed at HM in 2009. Boxes indicate interquartile ranges, bold lines
indicate medians, whiskers indicate inner fences, points indicate outliers, and asterisks
indicate extreme values. Numbers in parentheses represent sample sizes.

141
(20) (40) (20)
Figure 5.4: Proportion of cladodes with old C. cactorum damage from the three O.
humifusa neighborhoods surveyed at HM in 2009. Boxes indicate interquartile ranges,
bold lines indicate medians, whiskers indicate inner fences, points indicate outliers, and
asterisks indicate extreme values. Numbers in parentheses represent sample sizes.
Different lower case letters to the left of boxes represent significant pair wise
comparisons according to Dunn‟s test.
ab b
a

142
Discussion
Our findings agreed with previous reports that O. stricta is preferred over O. humifusa by
C. cactorum, as O. stricta plants had a higher proportion of damaged cladodes and
cladodes with larvae present. In cases where a neighboring species is preferred by an
herbivore, the expectation is of AS for the focal species, provided that focal species is an
acceptable host. However, we found very little evidence of AS to C. cactorum for O.
humifusa growing in close association with O. stricta. The 2006 survey revealed no
intraspecific differences for C. cactorum-related variables, while the 2009 survey
produced limited support for our prediction of AS. From the 2009 survey, we found that
O. humifusa plants paired with O. stricta had higher proportions of cladodes with C.
cactorum larvae and old damage than did O. humifusa plants alone or paired with
conspecifics. Although the result for larvae was marginal, we interpreted these findings
as partial support for our prediction of AS because observation of active larvae is
infrequent compared to that of old damage, leading to lower power for the larval
variable. The significant difference for old damage between plants in interspecific and
conspecifc pairs is of particular interest because the comparison between these two
neighborhoods controls for density of host plants, making it more likely that the observed
difference was due to diversity of hosts (that is, to the specific effect of O. stricta).
However, a significant difference between the interspecific plants and the alone plants,
which would have strengthened the support for AS, was not observed.
Our study found significantly higher levels of the native insects C.vittiger, D.
confusus, and G. hubbardi on O. humifusa than on O. stricta. Although the reason for C.
cactorum‟s preference for O. stricta over O. humifusa is unknown, native herbivores may

143
play a role. Plants that have actively feeding herbivores, or eggs of herbivores, on them
can produce feeding- or oviposition-induced surface compounds and volatiles that serve
to repel gravid females from oviposition (Blaakmeer et al. 1994, Harmon et al. 2003,
Hilker and Meiners 2011). Cactoblastis cactorum females are known to respond to slight
variations in the CO2 concentration at the boundary layer of plants, a behavior
hypothesized to enable them to choose more actively growing, healthier plants for their
offspring (Stange et al. 1995, Stange 1997). It is also possible that they respond to
induced surface compounds or volatiles to avoid Opuntia individuals that are relatively
more damaged and seek out individuals that are relatively less damaged. Given that all
four of the native insects we examined were found in higher levels on O. humifusa than
O. stricta (with three of the four being significantly higher), such behavior in our system
would result in C. cactorum females choosing an O. stricta individual more often than an
O. humifusa individual.
If, based on our finding for G. hubbardi damage, we assume it prefers O.
humifusa over O. stricta, we would expect to see AR for O. humifusa when paired with
O. stricta (Barbosa et al. 2009). Indeed, the 2006 data showed that O. humifusa plants
alone had a higher proportion of cladodes with G. hubbardi damage than did O. humifusa
in interspecific pairs. In theory, this could result from O. stricta acting as a
repellent/masking plant (Atsatt and Odowd 1976) by interfering with the ability of G.
hubbardi to locate its preferred host , or from spill over from O. humifusa onto O. stricta
(Agrawal et al. 2006, Barbosa et al. 2009). If the mechanism at work were
repellency/masking, we would not necessarily expect a difference between O. stricta
alone and in interspecific pairs. However, if spill over were occurring, we would expect

144
higher levels of G. hubbardi on O. stricta in interspecific pairs than O. stricta alone, as it
would be receiving AS from being paired with O. humifusa. Our actual results cannot
distinguish the mechanism for three reasons. First, although O. stricta in pairs did have
higher G. hubbardi damage than O. stricta alone, the difference was not significant
(p=0.126). Second, we did not mark conspecific pairs of O. humifusa in 2006, so we
cannot rule out a density effect, whereby G. hubbardi damage is diluted or spread out
when host plants are more densely arrayed, regardless of their species. Third, we did not
find any evidence for associational effects of G. hubbardi from our 2009 survey or from
the experimental plantings.
Although our study did not set out to document direct competition between the
two Opuntia species, the finding from the 2006 survey that O. humifusa plants alone had
significantly more cladodes than O. humifusa plants in interspecific pairs raises this
possibility by suggesting that plants growing alone are released from this competition
and can grow larger. However, the same effect was not seen for O. stricta plants alone
versus those in interspecific pairs. Further, the 2009 survey produced a contradictory
result in that O. humifusa plants alone actually lost proportionally more cladodes than
those in conspecifc or interspecific pairs. Exploration of direct and indirect competition
between the two Opuntia species is a topic for future studies dedicated expressively to
that purpose.
There are several possible explanations for the lack of evidence for AS/AR, one
of which is our choice of distances when determining plant neighborhoods. Russell et al
(2007) found that egg loads of Rhinocyllus conicus on the native thistle Cirsium
undulatum decreased significantly as distance from Carduus nutans, the invasive musk

145
thistle, increased. Similarly, White and Whitham (2002) found that both cankerworm
densities and defoliation rates on cottonwoods decreased as a function of distance from
box elder, the worm‟s preferred host. We based our distances on the behavior of C.
cactorum, as documenting AS to this herbivore was our primary objective. Larvae are
able to disperse very short distances, but only do so when no cladodes remain on their
original host (Dodd 1940). Females are reluctant to fly and seldom disperse from their
emergence site prior to first oviposition (Myers et al. 1981, Robertson 1987a). They will
disperse over larger distances, but only as hosts become sparser; the longest flight
recorded is 24km (Zimmermann et al. 2004). We reasoned that plants >5m from another
Opuntia would be sufficiently isolated, but now consider that this distance may not be
large enough to observe differences between alone plants and paired plants. Stiling et al
(2004) found no significant difference in C. cactorum attack rates for O. corallicola
plants ≤5m from an O. stricta and those >20m from an O. stricta. However, they found
that O. corallicola plants that were ≥500m from an O. stricta suffered no attacks by C.
cactorum. Perhaps the distance needed for a host plant to be truly isolated in terms of
normal C. cactorum dispersal is more on the order of tens or hundreds of meters. At our
sites, there are few to no plants that meet this criterion, but it is possible that
associational effects for C. cactorum could be documented at other, larger sites.
The occurrence of a spillover effect depends on the herbivore in question
reaching a high enough density to actually begin using the less preferred host. For
example, White and Whitham (2000) note that AS of cottonwoods, due to spillover from
the attractant “sink” plant, box elder, was only seen in areas with high cankerworm
densities. In areas with low densities, cankerworm completed their full life cycle on box

146
elder. Although we observed C. cactorum and native insect damage on both Opuntia
species, the density of these herbivores may simply be too low to result in a true spillover
effect onto their less preferred hosts. In the future, if the density of any of the herbivores
increases, or the density of one of their host species decreases, associational effects could
be more readily documented. Findings from the previous chapter show that mortality of
O. stricta is higher than that of O. humifusa, and that O. humifusa plants are more likely
to survive a C. cactorum attack. If O. stricta plants are dying faster than they are being
recruited, they may become scarce enough to consistently suffer complete “defoliation”
by C. cactorum within a single generation, a phenomenon which is currently rarely seen.
If this were the case, O. humifusa plants close to O. stricta could end up as the most
likely oviposition sites for the next generation of moths.
The phenomenon of AR and AS for mixtures of host plants with shared
herbivores has been fairly well-documented. Despite this, we found, at best, weak
support for our prediction of AS to C. cactorum for O. humifusa plants near O. stricta
and virtually no support for AR from or AS to four native opuntioid-feeding insects.
While the lack of support could be attributed to an imperfect experimental design, we
feel it is more likely due to ecological factors, particularly such as host plant and
herbivore density. Future studies that explicitly measure densities of Opuntia plants and
their associated herbivores are needed to confirm this, but we can cautiously conclude
that at our sites, the herbivore species are sparse enough, and both Opuntia species are
dense enough, for associational effects to be non-existent or undetectable.

147
Acknowledgements
We would like to thank Natalie and Brandon O‟Brien and Diane Harshbarger for their
help with field work, Terry Hingtgen (Florida Department of Environmental Protection,
Southwest Division) for his assistance in acquiring the necessary permits for field work
at Honeymoon Island State Park, and the employees of Fort deSoto County Park for their
support during the course of the study.
References cited
Agrawal, A. A., J. A. Lau, and P. A. Hamback. 2006. Community heterogeneity and the
evolution of interactions between plants and insect herbivores. Quarterly Review
of Biology 81:349-376.
Andow, D. A. 1991. Vegetational diversity and arthropod population response. Annual
Review of Entomology 36:561-586.
Atsatt, P. R. and D. J. Odowd. 1976. Plant defense guilds. Science 193:24-29.
Baker, A. J. and P. Stiling. 2009. Comparing the effects of the exotic cactus-feeding
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), and the native
cactus-feeding moth, Melitara prodenialis (Walker) (Lepidoptera: Pyralidae), on
two species of Florida Opuntia. Biological Invasions 11:619-624.
Barbosa, P., J. Hines, I. Kaplan, H. Martinson, A. Szczepaniec, and Z. Szendrei. 2009.
Associational resistance and associational susceptibility: Having right or wrong
neighbors. Annual Review of Ecology Evolution and Systematics 40:1-20.
Bennett FD, Habeck DH (1996) Cactoblastis cactorum: A successful weed control agent
in the Caribbean, now a pest in Florida? In: Delfosse ES, Scott RR, (eds)
Biological control of weeds: Proceedings of the VIII international symposium on
biological control of weeds. CSIRO Publishing, Melbourne pp 21-26
Bennett, F. D. and F. J. Simmonds. 1966. Biological control of Opuntia spp. by
Cactoblastis cactorum in the Leeward Islands (West Indies). BioControl 11:183-
189.

148
Blaakmeer, A., D. Hagenbeek, T. A. Vanbeek, A. E. Degroot, L. M. Schoonhoven, and J.
J. A. Vanloon. 1994. Plant response to eggs vs. host marking pheromone as
factors inhibiting oviposition by Pieris brassicae. Journal of Chemical Ecology
20:1657-1665.
Brown, B. J. and J. J. Ewel. 1987. Herbivory in complex and simple tropical successional
ecosystems. Ecology 68:108-116.
Dodd, A. P. 1940. The biological campaign against prickly pear. Commonwealth Prickly
Pear Board Bulletin:1-177. Brisbane, Australia.
Hamback, P. A., J. A. Stenberg, and L. Ericson. 2006. Asymmetric indirect interactions
mediated by a shared parasitoid: connecting species traits and local distribution
patterns for two chrysomelid beetles. Oecologia 148:475-481.
Harmon, J. P., J. A. White, and D. A. Andow. 2003. Oviposition behavior of Ostrinia
nubilalis (Lepidoptera : Crambidae) in response to potential intra- and
interspecific interactions. Environmental Entomology 32:334-339.
Hight, S. D. and J. E. Carpenter. 2009. Flight phenology of male Cactoblastis cactorum
(Lepidoptera: Pyralidae) at different latitudes in the southeastern United States.
Florida Entomologist 92:208-216.
Hilker, M. and T. Meiners. 2011. Plants and insect eggs: How do they affect each other?
Phytochemistry 72:1612-1623.
Himanen, S. J., J. D. Blande, T. Klemola, J. Pulkkinen, J. Heijari, and J. K. Holopainen.
2010. Birch (Betula spp.) leaves adsorb and re-release volatiles specific to
neighbouring plants - a mechanism for associational herbivore resistance? New
Phytologist 186:722-732.
Hjalten, J. and P. W. Price. 1997. Can plants gain protection from herbivory by
association with unpalatable neighbours? A field experiment in a willow-sawfly
system. Oikos 78:317-322.
Holt, R. D. and J. H. Lawton. 1994. The ecological consequences of shared natural
enemies. Annual Review of Ecology and Systematics 25:495-520.
Hunter, W. D., F. C. Pratt, and J. D. Mitchell. 1912. The principle cactus insects of the
United States. USDA, Bureau of Entomology.
Karban, R. 2001. Communication between sagebrush and wild tobacco in the field.
Biochemical Systematics and Ecology 29:995-1005.

149
Karban, R. 2007. Associational resistance for mule's ears with sagebrush neighbors. Plant
Ecology 191:295-303.
Letourneau, D. K., I. Armbrecht, B. Salguero Rivera, J. Montoya Lerma, E. Jimenez
Carmona, M. Constanza Daza, S. Escobar, V. Galindo, C. Gutierrez, S. Duque
Lopez, J. Lopez Mejia, A. M. Acosta Rangel, J. Herrera Rangel, L. Rivera, C.
Arturo Saavedra, A. Marina Torres, and A. Reyes Trujillo. 2011. Does plant
diversity benefit agroecosystems? A synthetic review. Ecological Applications
21:9-21.
LoGiudice, K., R. S. Ostfeld, K. A. Schmidt, and F. Keesing. 2003. The ecology of
infectious disease: Effects of host diversity and community composition on Lyme
disease risk. Proceedings of the National Academy of Sciences of the United
States of America 100:567-571.
Mann, J. 1969. Cactus-feeding Insects and Mites. Smithsonian Institution Bulletin 256,
Washington, D.C.
Myers, J. H., J. Monro, and N. Murray. 1981. Egg clumping, host plant selection and
population regulation in Cactoblastis cactorum (Lepidoptera). Oecologia 51:7-13.
Ostfeld, R. S. and F. Keesing. 2000. Biodiversity and disease risk: The case of Lyme
disease. Conservation Biology 14:722-728.
Pettey, F. W. 1948. The biological control of prickly pear in South Africa. Science
Bulletin. Department of Agriculture of the Union of South Africa.
Pfister, C. A. and M. E. Hay. 1988. Associational plant refuges - Convergent patterns in
marine and terrestrial communities result from differing mechanisms. Oecologia
77:118-129.
Raffel, T. R., L. B. Martin, and J. R. Rohr. 2008. Parasites as predators: Unifying natural
enemy ecology. Trends in Ecology & Evolution 23:610-618.
Robertson, H. G. 1987. Oviposition site selection in Cactoblastis cactorum
(Lepidoptera): Constraints and compromises. Oecologia 73:601-608.
Root, R. B. 1973. Organization of a plant-arthropod association in simple and diverse
habitats – Fauna of collards (Brassica-Oleracea). Ecological Monographs 43:95-
120.
Russell, E. P. 1989. Enemies hypothesis – A review of the effect of vegetational diversity
on predatory insects and parasitoids. Environmental Entomology 18:590-599.

150
Russell, F. L., S. M. Louda, T. A. Rand, and S. D. Kachman. 2007. Variation in
herbivore-mediated indirect effects of an invasive plant on a native plant. Ecology
88:413-423.
Simonsen, T. J., R. L. Brown, and F. A. H. Sperling. 2008. Tracing an invasion:
Phylogeography of Cactoblastis cactorum (Lepidoptera : Pyralidae) in the United
States based on mitochondrial DNA. Annals of the Entomological Society of
America 101:899-905.
Stange, G. 1997. Effects of changes in atmospheric carbon dioxide on the location of
hosts by the moth, Cactoblastis cactorum. Oecologia 110:539-545.
Stange, G., J. Monro, S. Stowe, and C. B. Osmond. 1995. The CO2 sense of the moth
Cactoblastis cactorum and its probable role in the biological control of the CAM
plant Opuntia stricta. Oecologia 102:341-352.
Stenberg, J. A., J. Heijari, J. K. Holopainen, and L. Ericson. 2007. Presence of Lythrum
salicaria enhances the bodyguard effects of the parasitoid Asecodes mento for
Filipendula ulmaria. Oikos 116:482-490.
Stiling, P. 2002. Potential nontarget effects of a biological control agent, prickly pear
moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae), in North America,
and possible management actions. Biological Invasions 4:273-281.
Stiling, P., D. Moon, and D. Gordon. 2004. Endangered cactus restoration: Mitigating the
non-target effects of a biological control agent (Cactoblastis cactorum) in Florida.
Restoration Ecology 12:605-610.
Stiling, P. and D. C. Moon. 2001. Protecting rare Florida cacti from attack by the exotic
cactus moth, Cactoblastis cactorum (Lepidoptera : Pyralidae). Florida
Entomologist 84:506-509.
Stiling, P., A. M. Rossi, and M. V. Cattell. 2003. Associational resistance mediated by
natural enemies. Ecological Entomology 28:587-592.
Tahvanainennen, J. O. and R. B. Root. 1972. Influence of vegetational diversity on
population ecology of a specialized herbivore, Phyllotreta cruciferae
(Coleoptera-Chrysomelidae). Oecologia 10:321-&.
Thomas, C. D. 1986. Butterfly larvae reduce host plant survival in vicinity of alternative
host species. Oecologia 70:113-117.

151
USDA-APHIS-PPQ. 2009. Cactus moth program annual report, Eastern region. United
States Department of Agriculture, Animal and Plant Health Inspection Service,
Plant Protection and Quarantine.
Wahl, M. and M. E. Hay. 1995. Associational resistance and shared doom – Effects of
epibiosis on herbivory. Oecologia 102:329-340.
White, J. A. and D. A. Andow. 2006. Habitat modification contributes to associational
resistance between herbivores. Oecologia 148:482-490.
White, J. A. and T. G. Whitham. 2000. Associational susceptibility of cottonwood to a
box elder herbivore. Ecology 81:1795-1803.
Zimmermann, H. G., S. Bloem, and H. Klein. 2004. Biology, history, threat, surveillance
and control of the cactus moth, Cactoblastis cactorum. International Atomic
Energy Agency/Food and Agriculture Organization, Vienna, Austria.
Zimmermann, H. G. and V. C. Moran. 1991. Biological control of prickly pear, Opuntia
ficus-indica (Cactaceae), in South Africa. Agriculture Ecosystems &
Environment 37:29-35.
Zimmermann, H. G., V. C. Moran, and J. H. Hoffmann. 2000. The renowned cactus
moth, Cactoblastis cactorum (Lepidoptera : Pyralidae): its natural history and
threat to native Opuntia floras in Mexico and the United States of America.
Florida Entomologist 84:543-551.

152
APPENDIX I
TAXONOMIC INFORMATION FOR OPUNTIOIDEAE TRIBES
CYLINDROPUNTIEAE AND OPUNTIEAE
Family: Cactaceae
Subfamily: Opuntioideae
Tribe: Cylindropuntieae
Cylindropuntia acanthocarpa (Engelmann and J.M. Bigelow) F.M. Knuth
native to US (AZ, CA, NM, NV, UT) and Mexico
*Cylindropuntia spinosior (Engelmann) F.M. Knuth
native to US (AZ, NM) and Mexico
Tribe Opuntieae
Consolea rubescens (Salm-Dyck ex de Candolle) Lemaire
native to West Indies and Caribbean; said to be naturalized in AZ, CA, NV
Nopalea cochenillifera (L.) Salm-Dyck
introduced in US (FL); cultivated in West Indies, Caribbean, Mexico, Central America;
probable origin is Mexico or Central America (Panama)
*Opuntia dillenii (Ker Gawler) Haworth
native to US (FL, SC,), West Indies, Central America, Mexico; introduced Australia, S.
Africa, Madagascar

153
APPENDIX I (continued)
Opuntia engelmannii Salm-Dyck ex Engelmann var. engelmannii
native to US (AZ, CA, NV, NM, T, UT) and Mexico
*Opuntia engelmannii Salm-Dyck ex Engelmann var. linguiformis (Griffiths)
Parfitt and Pinkava
reportedly native in Bexar County, TX, but possibly extirpated; cultivated and occasionally
naturalized in AZ, TX, Mexico
Opuntia engelmannii Salm-Dyck ex Engelmann var. lindheimeri (Engelmann)
Parfitt and Pinkava
native to US (LA, MO, MS, NM, OK, TX) and Mexico; introduced S. Africa
Opuntia ficus-indica (L.) Miller
native to Mexico; introduced in US (reports of naturalized populations in AL, AZ, CA,
FL, GA, HI, NM, TX); cultivated worldwide
Opuntia leucotricha de Candolle
native to Mexico; naturalized in FL
*Opuntia macrocentra Engelmann
native to US (AZ, NM, TX) and Mexico

154
Opuntia microdasys (Lehmann) Pfeiffer
native to Mexico; introduced in US (cultivated and naturalized in AZ)
Opuntia santa-rita (Griffiths and Hare) Rose
native to US (AZ , NM, and TX) and Mexico
Opuntia streptacantha Lemaire
native to northern and central Mexico; widely cultivated in Mexico
* These species are of conservation concern in their native range
Taxonomic, distribution, and conservation information obtained from:
Anderson, E.F. 2001. The Cactus Family. Timber Press, Portland, OR.
http://plants.usda.gov
http://www.ars-grin.gov
http://www.natureserve.org














![Allure~ MD Tropical warehouse moth (cocoa moth) [Ephestia cautel/a], Tobacco moth [Ephestia elute/la], Raisin moth [Cadra figulilel/a] 3. APPLICATION: Place dispensers every 20 to](https://img.pdfslide.net/doc/110x75/5fc80c76c4631870720794eb/allure-md-tropical-warehouse-moth-cocoa-moth-ephestia-cautela-tobacco-moth.jpg)














