Embed Size (px)
Citation preview
1
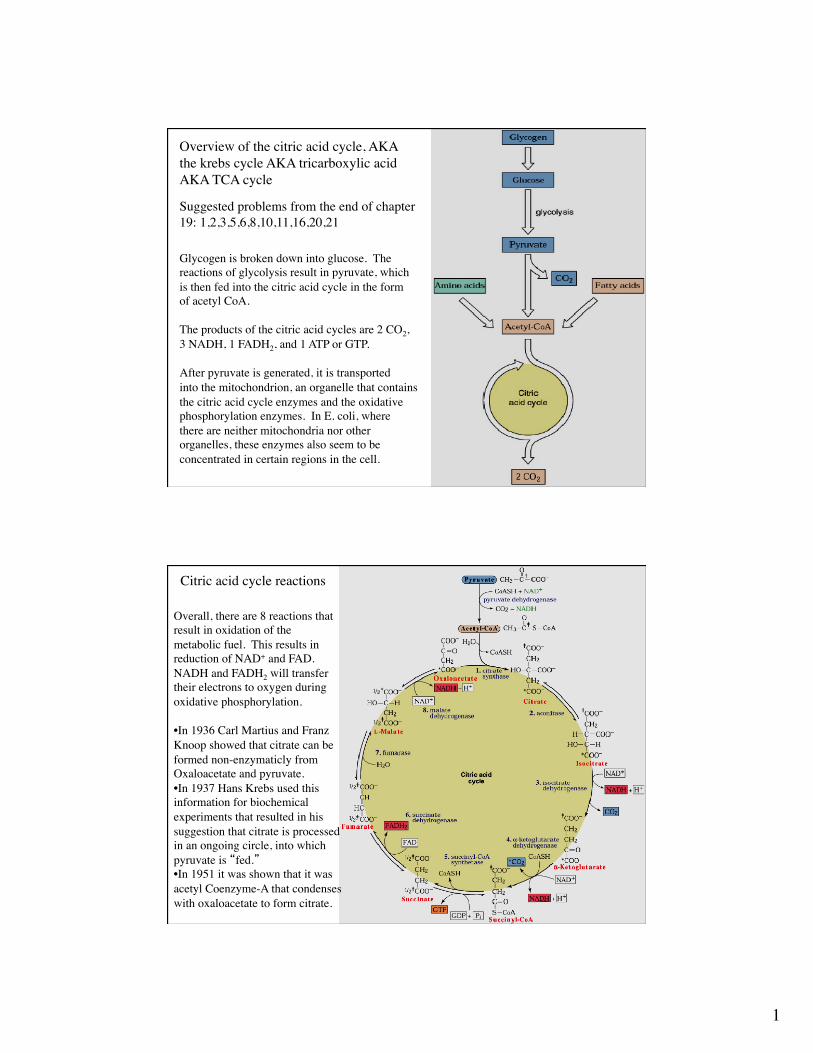
Overview of the citric acid cycle, AKAthe krebs cycle AKA tricarboxylic acidAKA TCA cycle
Glycogen is broken down into glucose. The reactions of glycolysis result in pyruvate, which is then fed into the citric acid cycle in the formof acetyl CoA.
The products of the citric acid cycles are 2 CO2,3 NADH, 1 FADH2, and 1 ATP or GTP.
After pyruvate is generated, it is transportedinto the mitochondrion, an organelle that containsthe citric acid cycle enzymes and the oxidativephosphorylation enzymes. In E. coli, wherethere are neither mitochondria nor other organelles, these enzymes also seem to be concentrated in certain regions in the cell.
Suggested problems from the end of chapter 19: 1,2,3,5,6,8,10,11,16,20,21
Citric acid cycle reactions
Overall, there are 8 reactions thatresult in oxidation of themetabolic fuel. This results inreduction of NAD+ and FAD.NADH and FADH2 will transfer their electrons to oxygen duringoxidative phosphorylation.
• In 1936 Carl Martius and FranzKnoop showed that citrate can beformed non-enzymaticly fromOxaloacetate and pyruvate.• In 1937 Hans Krebs used thisinformation for biochemicalexperiments that resulted in hissuggestion that citrate is processedin an ongoing circle, into whichpyruvate is “fed.”• In 1951 it was shown that it wasacetyl Coenzyme-A that condenses with oxaloacetate to form citrate.

2
The pre-citric acid reaction- pyruvate dehydrogenase
Pyruvate dehydrogenase is a multi-subunit complex, containing three enzymes that associatenon-covalently and catalyze 5 reaction. The enzymes are: (E1) pyruvate dehydrogenase(E2) dihydrolipoyl transacetylase(E3) dihydrolipoyl dehydrogenase
What are the advantages for arranging enzymes in complexes?
E. coli pyruvate dehydrogenase complex
The E. coli complex is a particle of about 300 Å in diameter. The mass is ~4.6 million Daltons (g/mol). There are 60 polypeptidesper particle.The complex contains 24 E2 proteins whichare arranged in a cube and form the so-calledcore complex. 24 E1 proteins and 12 E3proteins are arranged around the core.
3
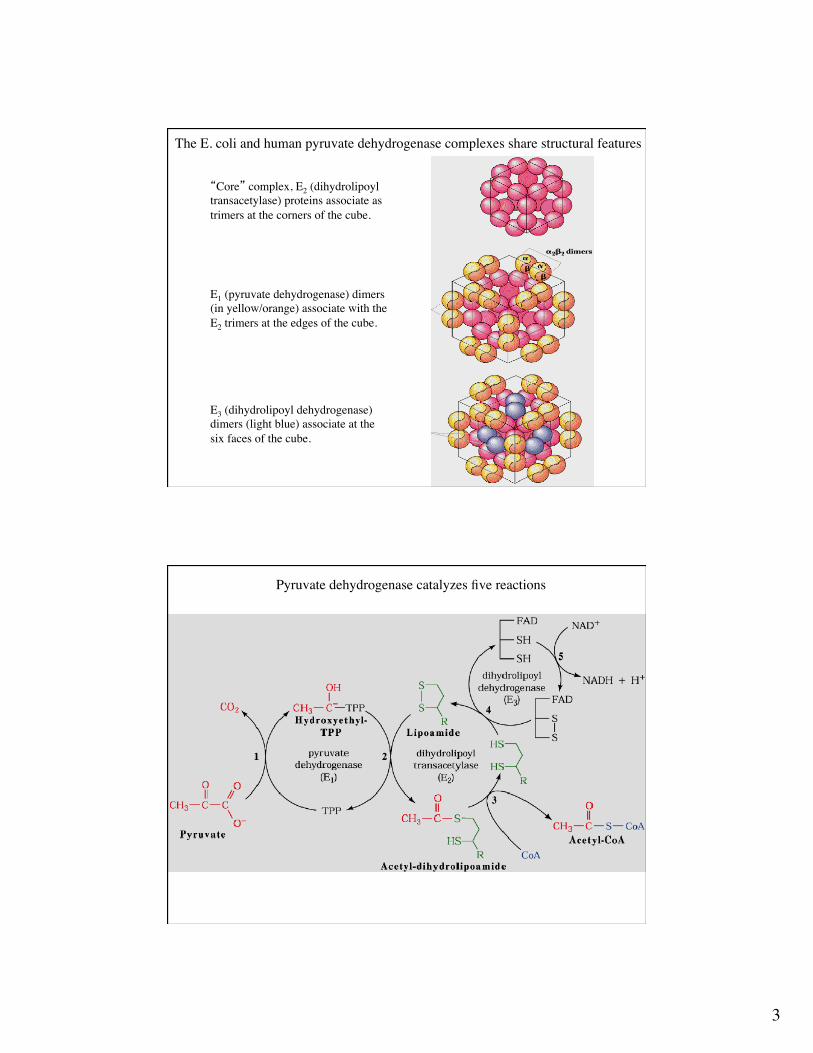
The E. coli and human pyruvate dehydrogenase complexes share structural features
“Core” complex, E2 (dihydrolipoyltransacetylase) proteins associate as trimers at the corners of the cube.
E1 (pyruvate dehydrogenase) dimers(in yellow/orange) associate with the E2 trimers at the edges of the cube.
E3 (dihydrolipoyl dehydrogenase)dimers (light blue) associate at thesix faces of the cube.
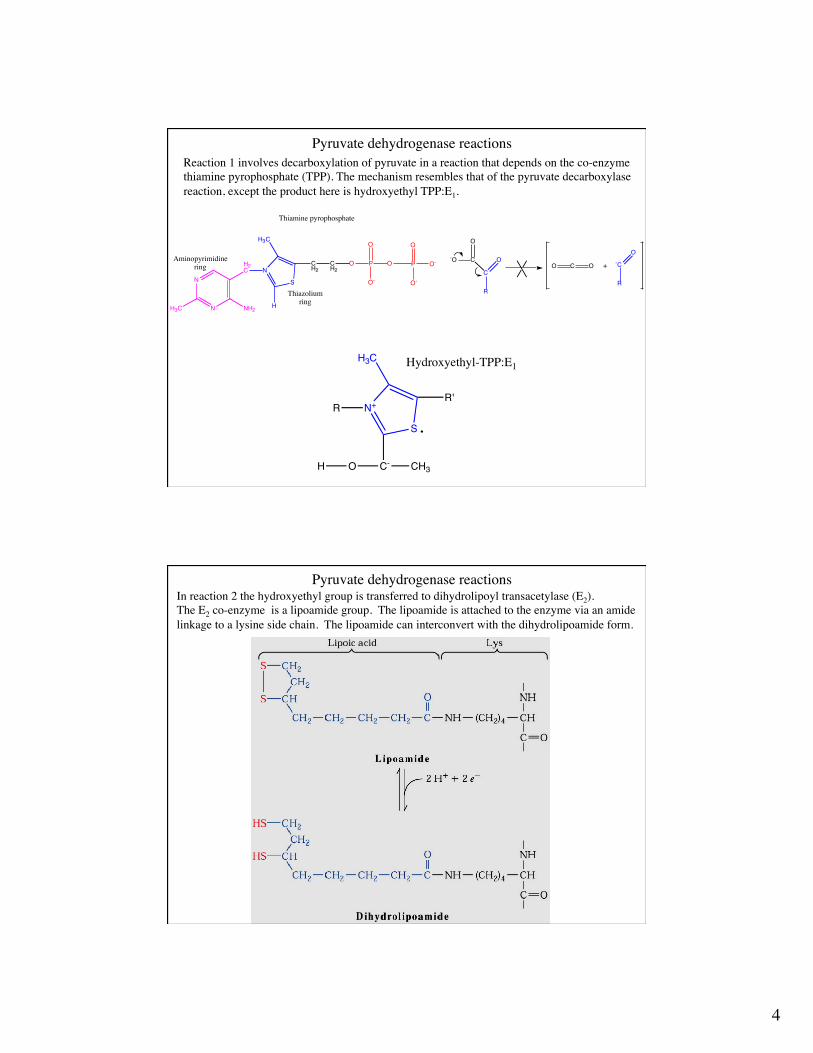
Pyruvate dehydrogenase catalyzes five reactions
4
Pyruvate dehydrogenase reactionsReaction 1 involves decarboxylation of pyruvate in a reaction that depends on the co-enzymethiamine pyrophosphate (TPP). The mechanism resembles that of the pyruvate decarboxylasereaction, except the product here is hydroxyethyl TPP:E1.
N
NH3C NH2
H2C N
S
H3C
H
CH2
CH2
O P
O
O-
O P
O-
O-
Aminopyrimidinering
Thiazoliumring
Thiamine pyrophosphate
O
-O C
O
C
R
O
O C O-C
R
O
+
N+
S
H3C
R
R'
C-O CH3H
Hydroxyethyl-TPP:E1
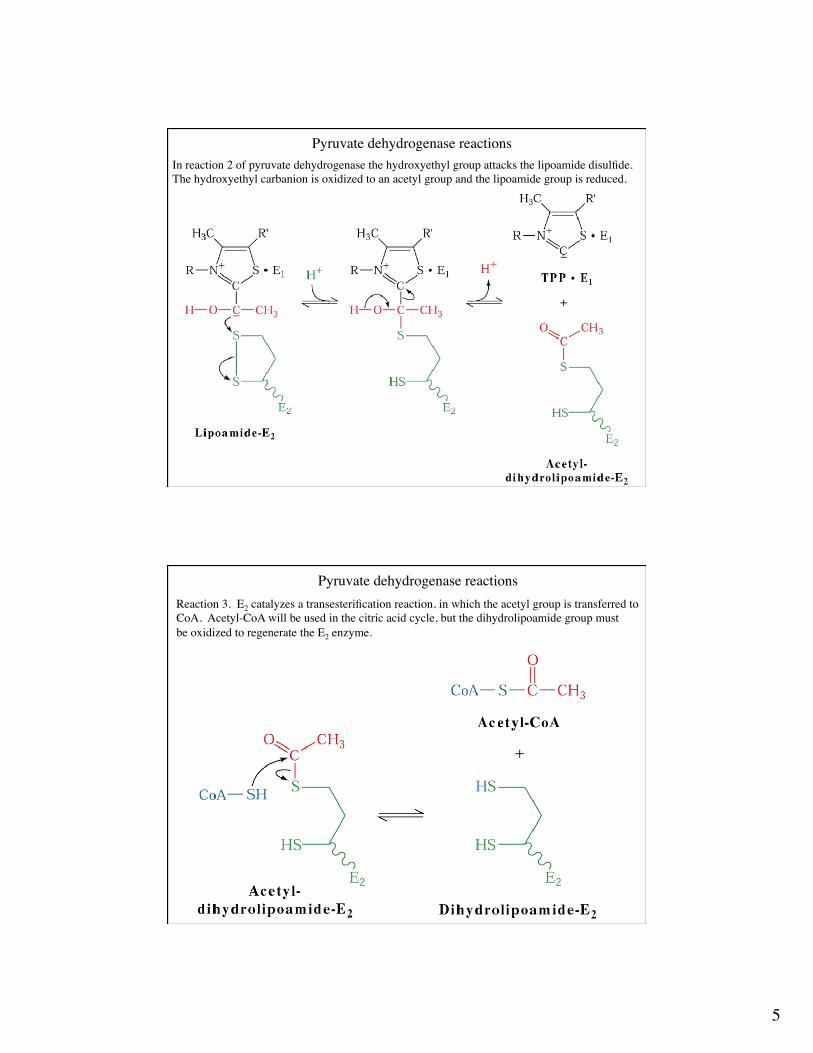
Pyruvate dehydrogenase reactionsIn reaction 2 the hydroxyethyl group is transferred to dihydrolipoyl transacetylase (E2). The E2 co-enzyme is a lipoamide group. The lipoamide is attached to the enzyme via an amidelinkage to a lysine side chain. The lipoamide can interconvert with the dihydrolipoamide form.
5
Pyruvate dehydrogenase reactionsIn reaction 2 of pyruvate dehydrogenase the hydroxyethyl group attacks the lipoamide disulfide.The hydroxyethyl carbanion is oxidized to an acetyl group and the lipoamide group is reduced.
Pyruvate dehydrogenase reactionsReaction 3. E2 catalyzes a transesterification reaction, in which the acetyl group is transferred toCoA. Acetyl-CoA will be used in the citric acid cycle, but the dihydrolipoamide group mustbe oxidized to regenerate the E2 enzyme.
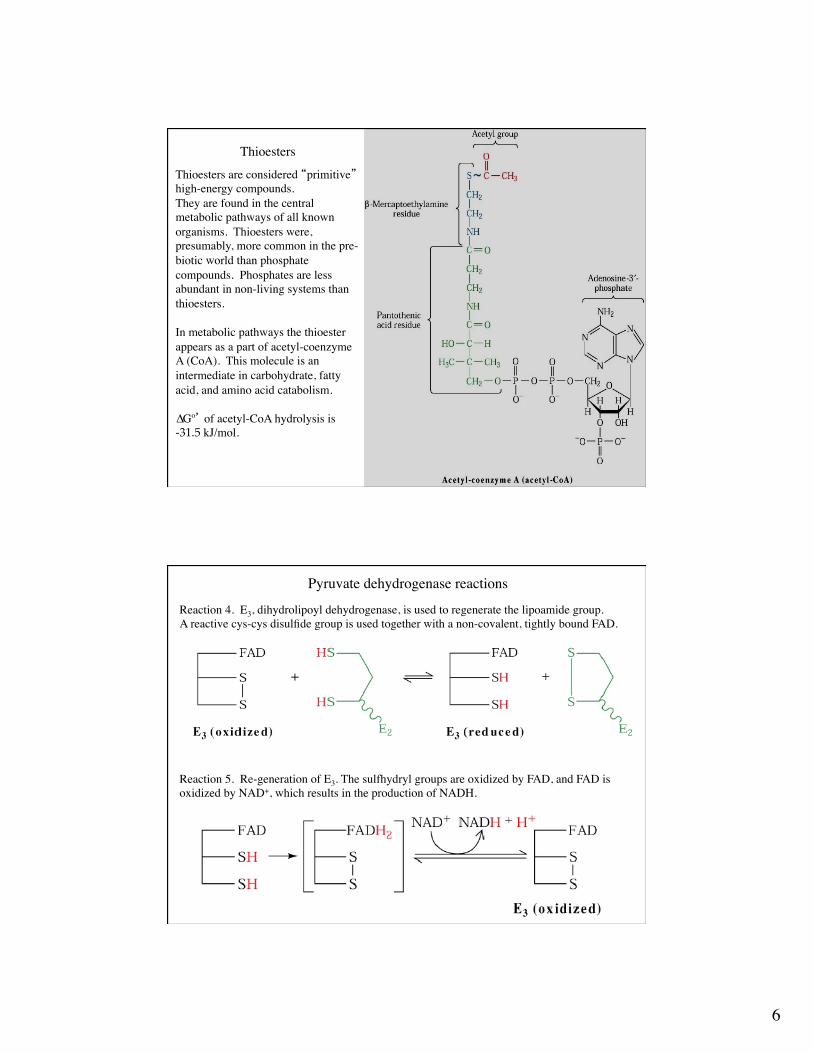
6
Thioesters
Thioesters are considered “primitive” high-energy compounds. They are found in the central metabolic pathways of all known organisms. Thioesters were, presumably, more common in the pre-biotic world than phosphate compounds. Phosphates are less abundant in non-living systems than thioesters.
In metabolic pathways the thioesterappears as a part of acetyl-coenzyme A (CoA). This molecule is an intermediate in carbohydrate, fatty acid, and amino acid catabolism.
∆Gº’ of acetyl-CoA hydrolysis is-31.5 kJ/mol.
Pyruvate dehydrogenase reactions
Reaction 4. E3, dihydrolipoyl dehydrogenase, is used to regenerate the lipoamide group.A reactive cys-cys disulfide group is used together with a non-covalent, tightly bound FAD.
Reaction 5. Re-generation of E3. The sulfhydryl groups are oxidized by FAD, and FAD isoxidized by NAD+, which results in the production of NADH.

7
Pyruvate dehydrogenase reactionsThe E2 protein on the inside of the structure must “channel” intermediates from the E1 and E3 enzymes on the outside of the structure. This apparently is achieved due to the flexibility and length of the lipoyllysyl arm.
This arm (shown on right) may swing between E1 and E3. In terms of the reactions that are catalyzed, this arm can be thought of as a “tether” that brings the reactive disulfidegroup to the active site of E1 (where it reacts to accept the hydroxyethyl group), then swings to the active site of E2 (where CoA accepts the acetyl group), and finally to the active site of E3 (where the disulfide group is re-oxidized).
Support for this idea is the finding that the lipoyllysyl arm is attached to the E2 protein via an alanine-proline rich region that seems to adopt multiple conformations, meaning that it is flexible.If this idea is correct, a single lipoyllysyl arm can reach numerous E3 active sites.
How would you test the idea that a flexible linkage near the lipoamide group is essential for the activity of pyruvatedehydrogenase?
Structure of E2 of pyruvate dehydrogenase
The E2 catalytic domain from thebacterium Azotobacter vinelandii is shown.
The 24 E2 proteins are arrangedin trimers, each trimer occupyingthe corner of a cube.The edge of the cube is ~125 Å.Substrates and co-factors mustfit in the spaces between the E2 proteins and within the interior cavityof the cube. Only 4 “corners” are shown.
Acetyl CoA must diffuse into itsbinding site from the internal spaceof the cube.
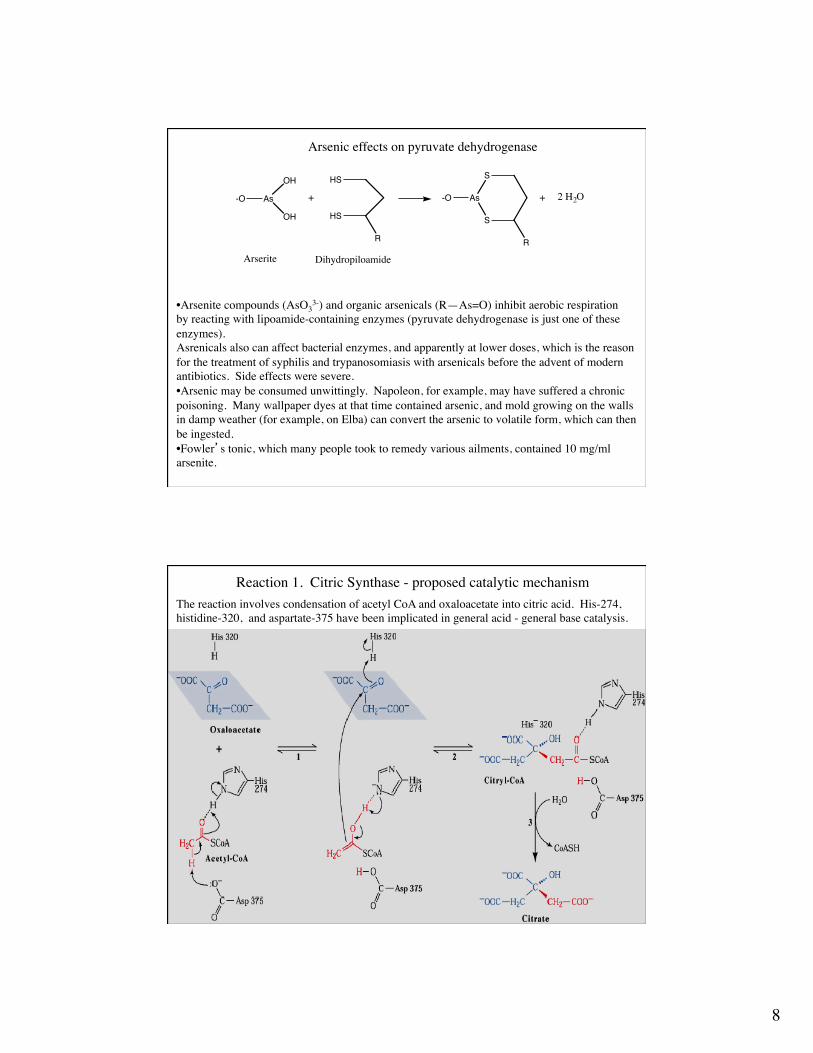
8
Arsenic effects on pyruvate dehydrogenase
-O As
OH
OH
+
HS
HS
R
S
As
S
-O + 2 H2O
R
Arserite Dihydropiloamide
• Arsenite compounds (AsO33-) and organic arsenicals (R—As=O) inhibit aerobic respiration
by reacting with lipoamide-containing enzymes (pyruvate dehydrogenase is just one of theseenzymes).Asrenicals also can affect bacterial enzymes, and apparently at lower doses, which is the reasonfor the treatment of syphilis and trypanosomiasis with arsenicals before the advent of modernantibiotics. Side effects were severe.• Arsenic may be consumed unwittingly. Napoleon, for example, may have suffered a chronicpoisoning. Many wallpaper dyes at that time contained arsenic, and mold growing on the wallsin damp weather (for example, on Elba) can convert the arsenic to volatile form, which can thenbe ingested.• Fowler’s tonic, which many people took to remedy various ailments, contained 10 mg/ml arsenite.
Reaction 1. Citric Synthase - proposed catalytic mechanismThe reaction involves condensation of acetyl CoA and oxaloacetate into citric acid. His-274, histidine-320, and aspartate-375 have been implicated in general acid - general base catalysis.
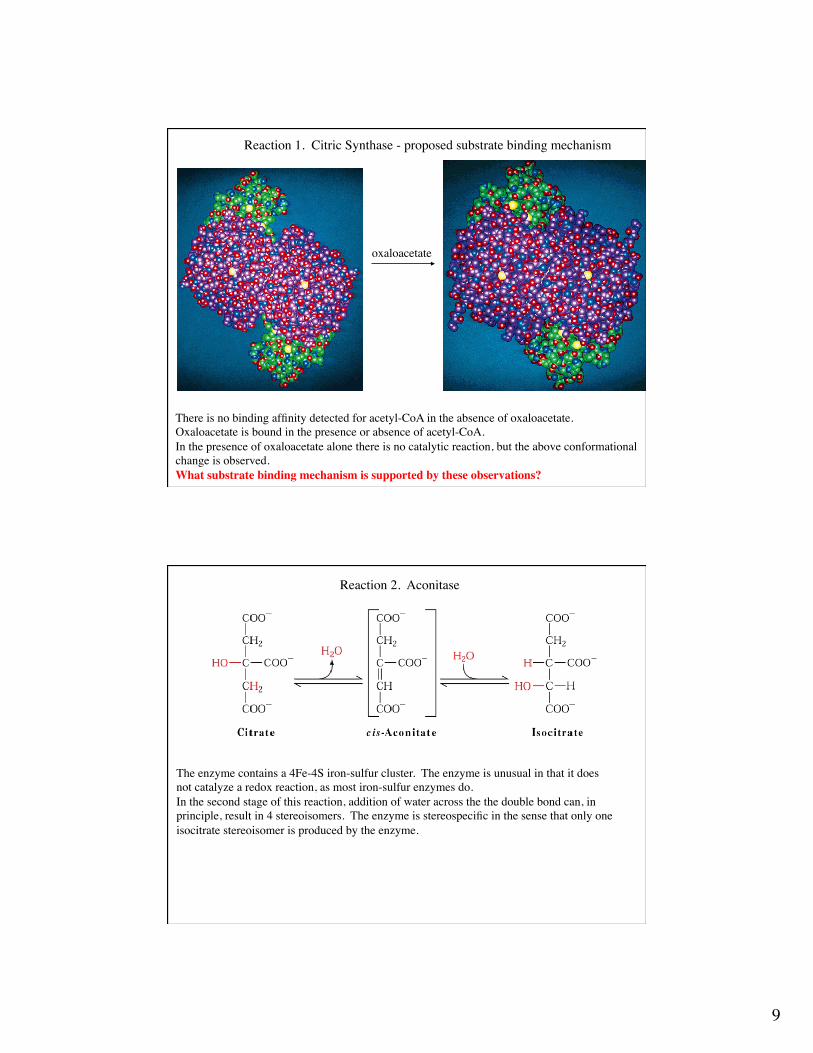
9
Reaction 1. Citric Synthase - proposed substrate binding mechanism
There is no binding affinity detected for acetyl-CoA in the absence of oxaloacetate. Oxaloacetate is bound in the presence or absence of acetyl-CoA.In the presence of oxaloacetate alone there is no catalytic reaction, but the above conformationalchange is observed.What substrate binding mechanism is supported by these observations?
oxaloacetate
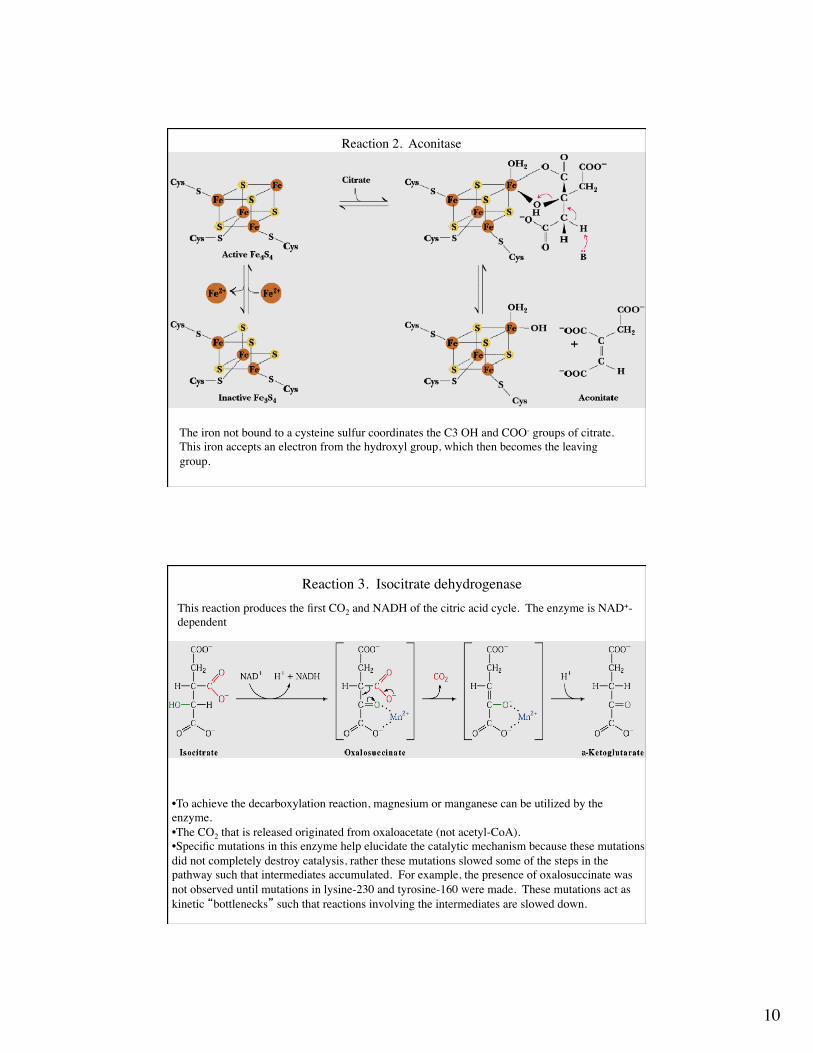
Reaction 2. Aconitase
The enzyme contains a 4Fe-4S iron-sulfur cluster. The enzyme is unusual in that it doesnot catalyze a redox reaction, as most iron-sulfur enzymes do.In the second stage of this reaction, addition of water across the the double bond can, inprinciple, result in 4 stereoisomers. The enzyme is stereospecific in the sense that only one isocitrate stereoisomer is produced by the enzyme.
10
Reaction 2. Aconitase
The iron not bound to a cysteine sulfur coordinates the C3 OH and COO- groups of citrate.This iron accepts an electron from the hydroxyl group, which then becomes the leaving group.
Reaction 3. Isocitrate dehydrogenaseThis reaction produces the first CO2 and NADH of the citric acid cycle. The enzyme is NAD+-dependent
• To achieve the decarboxylation reaction, magnesium or manganese can be utilized by theenzyme.• The CO2 that is released originated from oxaloacetate (not acetyl-CoA).• Specific mutations in this enzyme help elucidate the catalytic mechanism because these mutationsdid not completely destroy catalysis, rather these mutations slowed some of the steps in thepathway such that intermediates accumulated. For example, the presence of oxalosuccinate wasnot observed until mutations in lysine-230 and tyrosine-160 were made. These mutations act askinetic “bottlenecks” such that reactions involving the intermediates are slowed down.

11
Reaction 4. α-ketoglutarate dehydrogenase
This reaction resembles the pyruvate dehydrogenase multi-enzyme complex reaction, down tothe identity of the E3 dihydrolipoyl dehydrogenase. As expected, the mechanisms are, mostlikely, the same, and the product is the high energy thioester, in this case, succinyl CoA.
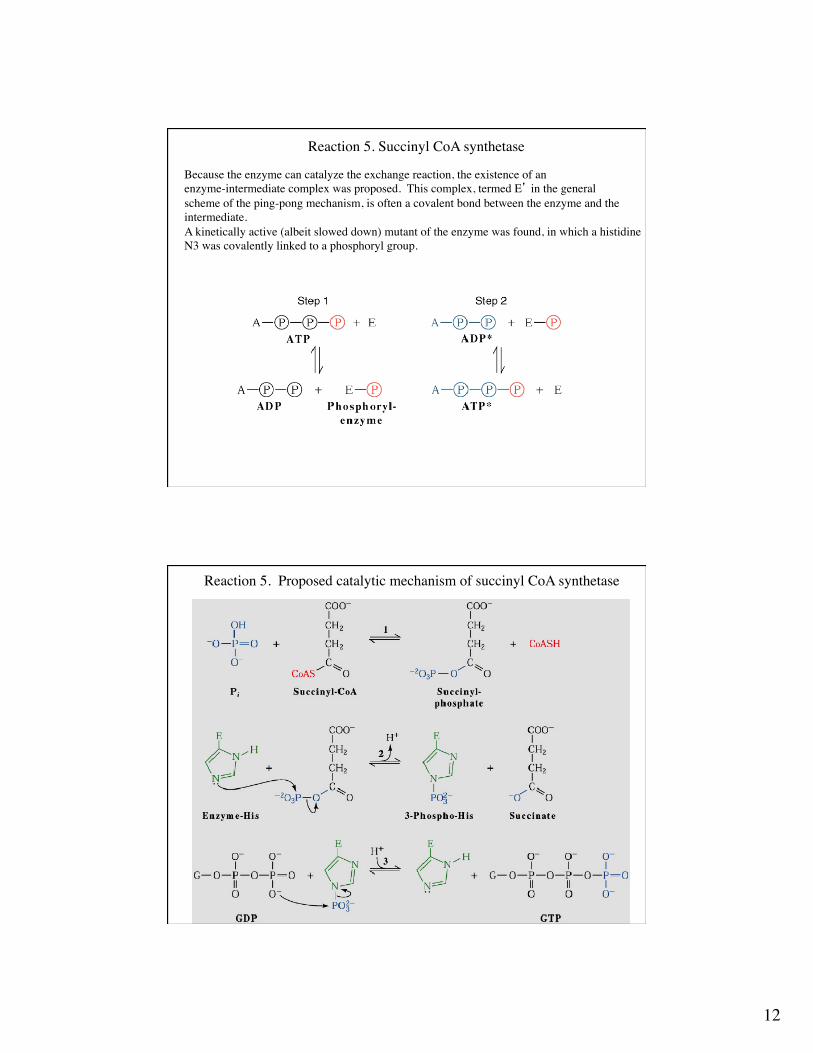
Reaction 5. Succinyl CoA synthetase
This enzyme couples the free energy released from hydrolysis of the high energy thioestersuccinyl CoA to the synthesis of a high energy NTP. GTP is generated in mammalian cells, whereas ATP is generated in bacteria and plant cells.From an energetic standpoint, these reactions essentially are the same because GTP and ATP can be interconverted rapidly through the action of nucleoside diphosphate kinase.
GTP + ADP ↔ GDP + ATP ∆G°’ = 0
Isotopic labeling experiments permit us to “follow” the fate of the labeled group and elucidatethe catalytic steps.In the absence of succinyl CoA, the enzyme transfers the unlabeled γ-phosphoryl group of ATP to a labeled ADP, thus generating labeled ATP.What substrate binding mechanism is supported by this finding?
Succinyl-CoA + ADP + Pi ↔ ATP + succinate + CoASH
12
Reaction 5. Succinyl CoA synthetase
Because the enzyme can catalyze the exchange reaction, the existence of anenzyme-intermediate complex was proposed. This complex, termed E’ in the generalscheme of the ping-pong mechanism, is often a covalent bond between the enzyme and the intermediate.A kinetically active (albeit slowed down) mutant of the enzyme was found, in which a histidineN3 was covalently linked to a phosphoryl group.
Reaction 5. Proposed catalytic mechanism of succinyl CoA synthetase
13
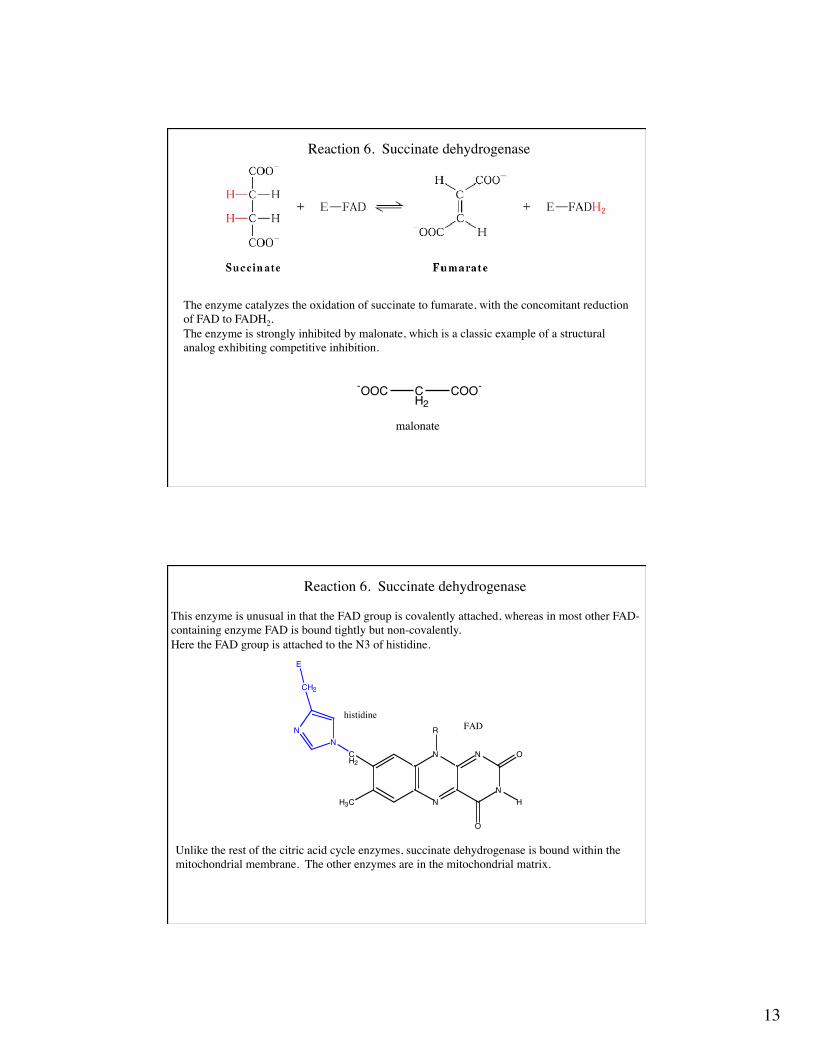
Reaction 6. Succinate dehydrogenase
The enzyme catalyzes the oxidation of succinate to fumarate, with the concomitant reductionof FAD to FADH2.The enzyme is strongly inhibited by malonate, which is a classic example of a structural analog exhibiting competitive inhibition.
-OOC C
H2
COO-
malonate
Reaction 6. Succinate dehydrogenase
This enzyme is unusual in that the FAD group is covalently attached, whereas in most other FAD-containing enzyme FAD is bound tightly but non-covalently.Here the FAD group is attached to the N3 of histidine.
N
N
N
NCH2
H3C
O
R
H
O
N
N
CH2
E
histidine
FAD
Unlike the rest of the citric acid cycle enzymes, succinate dehydrogenase is bound within the mitochondrial membrane. The other enzymes are in the mitochondrial matrix.
14
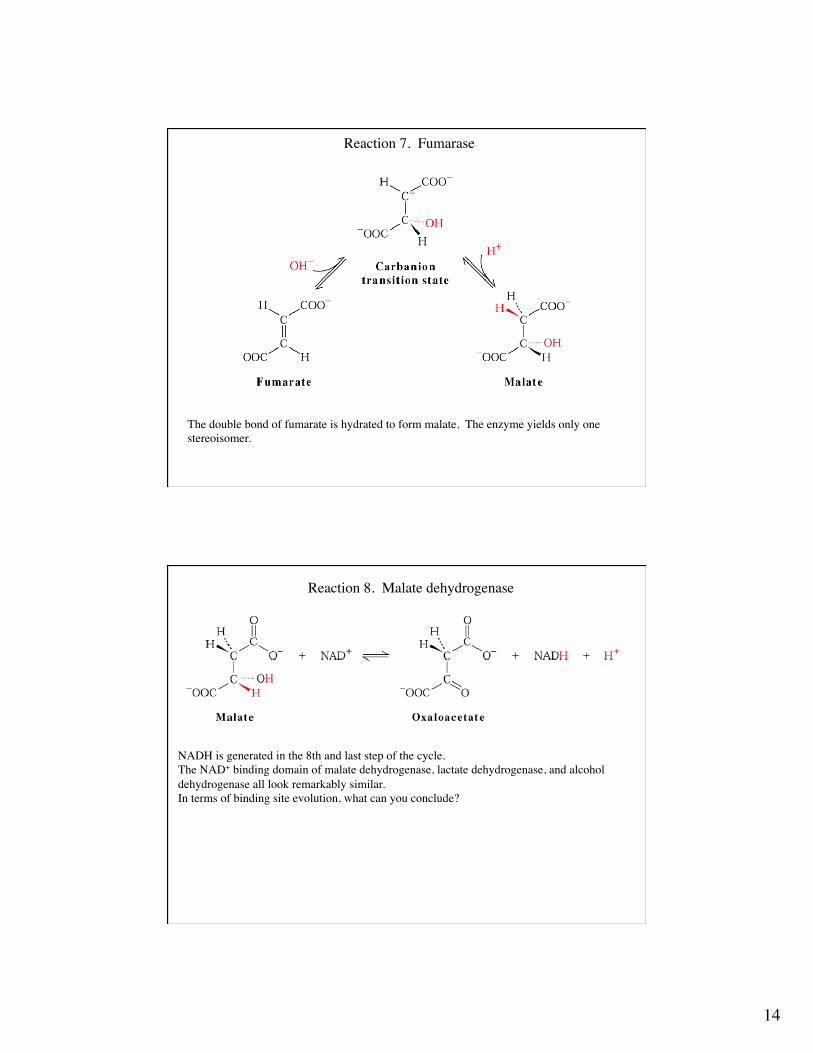
Reaction 7. Fumarase
The double bond of fumarate is hydrated to form malate. The enzyme yields only one stereoisomer.
Reaction 8. Malate dehydrogenase
NADH is generated in the 8th and last step of the cycle.The NAD+ binding domain of malate dehydrogenase, lactate dehydrogenase, and alcoholdehydrogenase all look remarkably similar.In terms of binding site evolution, what can you conclude?

15
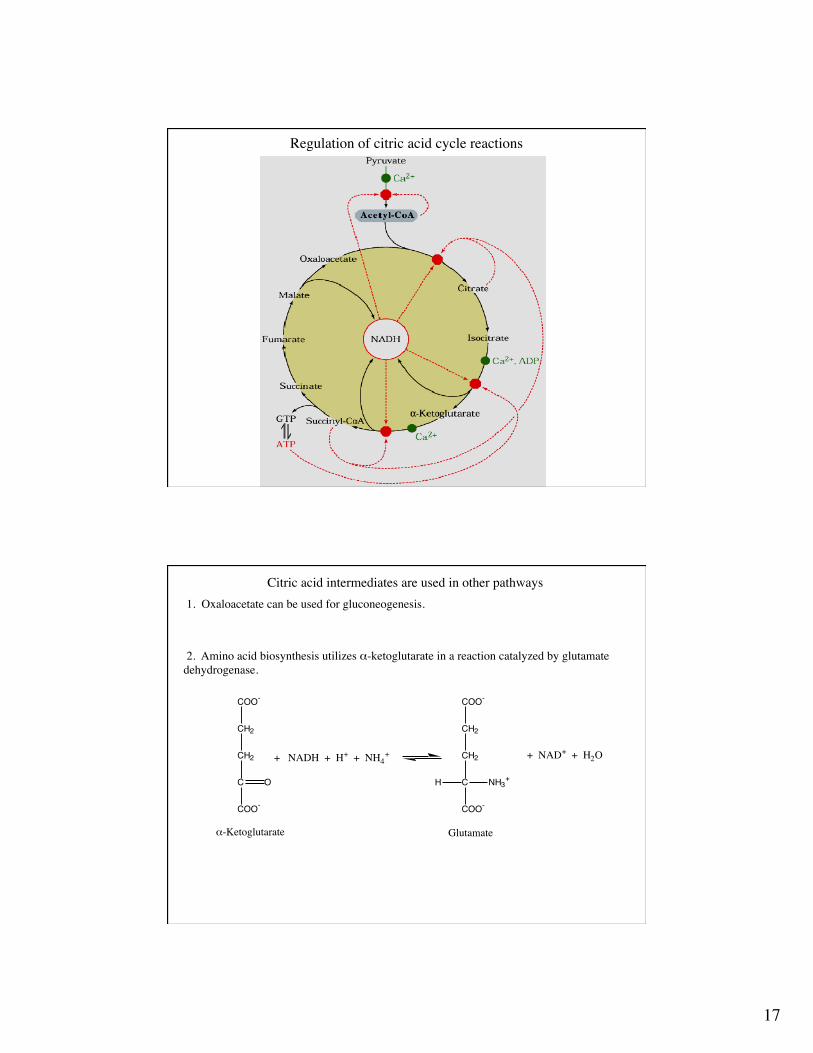
Regulation of the citric acid cycle - is there a metabolon?
According to the metabolon hypothesis, enzymes involved in a pathway might be expected to be physically associated in vivo. The same considerations given for the efficiency of pyruvatedehydrogenase can be used here successfully.At least two criteria must be met: 1. In vitro evidence for the association of proteins must pass the test of specificity. Manyproteins can associate in vitro, but what is the relevance of such data to the situation in vivo?Specificity should be demonstrated. By one criterion, specificity is measured by the resistanceto non-specific competition.
2. Evidence in vivo must be obtained. This can be done through (fluorescent) labeling thatcan demonstrate co-localization of the proteins in the cell. Also, this can be done by isolatingsub-organelle structures containing the pathway enzymes. At least some of the citric acid cycle proteins seem to associate (transiently or weakly) with the inner mitochondrial membrane. Partially purified mitochondrial fractions display a faster turnover rate than completely solubilize fractions, arguing that there is an advantage to keeping the enzymes in their “in vivo”overall arrangement.
Regulation of pyruvate dehydrogenase
The entry of acetyl-CoA into the citric acid cycle results in a large-scale production of ATPthrough oxidative phosphorylation. It is not surprising that the citric acid cycle is regulatedcarefully, especially because there is no other enzyme that will generate acetyl CoA frompyruvate in mammalian cells.The first reaction, the charging of CoA with an acetyl group, via the action of the multi-enzymecomplex pyruvate dehydrogenase, is not reversible (operates far form equilibrium). This is anatural site for enzymatic regulation.
1. Product inhibition by NADH and acetyl CoA. These products compete with NAD and CoA,respectively, for their binding sites. High [NADH]/[NAD+] and [acetyl-CoA]/[CoA] result inenzyme inhibition. E1 is the non-reversible enzyme within this complex.
2. The E1 (pyruvate dehydrogenase) activity is modified by serine phosphorylation and de-phosphorylation, such that when the enzyme is phosphorylated it is inactive.

16
Regulation of pyruvate dehydrogenase (continued)
Insulin is released in response to high glucose levels.Insulin activates pyruvate dehydrogenase phosphatase. This results in the de-phosphorylation of pyruvate dehydrogenase and activation of the citric acid cycle.In addition to activating the glucose metabolic pathways, insulin stimulates the glycogenanabolic pathways, thus helping to maintain steady levels of blood glucose.
3. Pyruvate and ADP inhibit pyruvate dehydrogenase kinase.
Rate-controlling enzymes of the citric acid cycle
The difficulty in defining the ∆G of the citric acid reactions is that many of the metabolitesare present in the cytoplasm and the mitochondria, but the concentrations of these metabolitesin each compartment are not known.Under the assumption that metabolites are free to diffuse into the mitochondria, ∆G can becalculated. Only reactions 1, 3, and 4 show ∆G < 0, which argues that these reactions are therate limiting steps in the pathway.
The citric acid enzymes are controlled via three mechanisms: 1. Substrate availability. 2. Product inhibition. 3. Competitive feedback inhibition by intermediates.Thus most of the regulation depends on the amount of substrates being fed into the cycle (acetyl-CoA) and the cycle products (ATP, NADH).
17
Regulation of citric acid cycle reactions
Citric acid intermediates are used in other pathways 1. Oxaloacetate can be used for gluconeogenesis.
2. Amino acid biosynthesis utilizes α-ketoglutarate in a reaction catalyzed by glutamate dehydrogenase.
COO-
CH2
CH2
C O
COO-
+ NADH + H+ + NH4+
COO-
CH2
CH2
C NH3+
COO-
H
+ NAD+ + H2O
α-Ketoglutarate Glutamate

18
Citric acid intermediates are used in other pathways
COO-
C
CH2
COO-
O
Oxaloacetate
+ C
COO-
H
CH3
+H3N
Alanine
COO-
C
CH2
COO-
+H3N H
Aspartate
+
COO-
C
CH3
O
Pyruvate
3. Aspartate aminotransferase catalyzes the trans-amination reaction that yields aspartatefrom oxaloacetate.





























