Embed Size (px)
Citation preview

European Journal of Pharmacology, 181 (1990) 71-82 71 Elsevier
EJP 51309
The melanocortin receptor in the rat lacrimal gland: a model system for the study of M S H (melanocyte stimulating hormone)
as a potential neurotransmitter
H a n a Leiba a, Ni ra B. Gar ty , Josepha Schmidt-Sole, Or ly Pi te rman, A n a t Az rad and Y o r a m
Sa lomon
Department of Hormone Research, The Weizmann Institute of Science, Rehovot 76100, Israel
Received 4 January 1990, accepted 27 February 1990
The melanocortin receptors in intraorbital and extraorbital rat lacrimal glands were studied with [125I][Nle4,D- PheT]aMSH as radioligand and with several unlabeled melanocortin peptides. The pharmacological properties of the melanocortin receptor in both tissues appeared to be essentially identical. Receptor binding was studied in a membrane fraction sedimented at 12 000-100 000 × g, establishing for [125I][Nlea,D-PheV]aMSH a K o of 0.76 and 2.2 nM for the intra- and extraorbital glands, respectively. Binding of the radioligand was competitively inhibited by aMSH (a-melanocyte stimulating hormone) and ACTH-(1-24) with ICs0 values in the submicromolar range. MSH binding in both tissues was abolished by EGTA and was increased dose dependently with elevation of free Ca 2+ ion concentration. The half-maximal effect on MSH binding was obtained around 200 /aM Ca 2÷ and maximal binding was reached at nearly 2 mM free Ca 2+ in membrane preparations from both tissues. The calmodulin-binding peptides, melittin, mastoparan and M5, the latter being the 18-amino acid synthetic analogue of the C-terminal calmodulin- binding domain of myosin light chain kinase, inhibited MSH binding in the concentration range of 1-20 /~M. Macroscopic autoradiographic analysis of cryosections prepared from either lacrimal gland to which [125I][Nle4,D- PheT]aMSH was subsequently bound, showed the melanocortin receptor to be uniformly distributed within the acinar lobes. At the microscopic level, MSH was found to be associated with the acinar cells, primarily at the basal perinuclear region. Peroxidase secretion from extraorbital lacrimal slices was stimulated by MSH, ephinephrine and carbamylcholine to a similar extent. The response of the tissue to stimulation by MSH was however not blocked by a/fl-adrenoceptor blockers or by atropine, suggesting that MSH acts as a primary secretagogue in this tissue. Thus, this system seems to be uniquely suited to serve as a model for the study of both the molecular and pharmacological details of the action of MSH and other melanocortins in a non-melanogenic tissue.
Melanocortins; MSH (melanocyte stimulating hormone) receptors; Lacrimal glands; Secretion; Neurotransmitters
I. Introduction
In recent years melanocortin peptides have emerged as regulators of an increasing number of
1 Present address: Department of Ophthalmology, Kaplan Hospital Rehovot, Israel.
Correspondence to: Yoram Salomon, Department of Hormone Research, The Weizmann Institute of Science, P.O. Box 26, Rehovot 76100, Israel.
processes that are unrelated to their classical role as pituitary hormones (reviewed by Eberle, 1988). Furthermore, the synthesis of pro-opiomelanocor- tin was shown to take place in several distinct regions of the central (Liotta et al., 1980) and peripheral nervous system (reviewed by Smith and Funder, 1988; Bardin et al., 1985), and not exclu- sively in the pituitary (Mains et al., 1977) as has been thought previously. When mice and rats were injected in vivo with [125I][Nlea,D-PheT]a-
0014-2999/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

72
melanocyte-stimulating hormone (aMSH), the peptide was shown to accumulate specifically in several glandular organs, white adipose tissue, bladder, duodenum, skin, spleen and hypothala- mus. The highest specific binding was observed in the lacrimal and Harderian glands (Tatro and Reichlin, 1987). These observations suggested a functional role for MSH in these tissues. Further- more, independent observations indicated that melanocortins act directly and influence respective target tissues in a way that is neither related to nor mediated by steroidogenesis or melanogenesis. Ef- fects on animal and human behavior (De Wied and Ferrari, 1986; De Wied and Jolles, 1982), facilitation of nerve regeneration (Strand et al., 1981; Bijlsma et al., 1981) and recovery of motor performance following crush denervation (Saint- Come et al., 1982; Bijlsma et al., 1983), induction of long-lasting potentiation of neurotransmitter release (Johnston et al., 1983) and increase of miniature end-plate potentials in the neuromuscu- lar junction (Strand et al., 1981), stimulation of lacrimal secretion (Jahn et al., 1982), acceleration of lipolysis (Tanaka et al., 1962) and stimulation of aromatase and plasminogen activator in rat Sertoli cells (Boitani et al., 1989) exemplify the diversity of melanocortin functions. The actual presence of specific melanocortin receptors in many of these tissues, their basic properties and mechanism of action, as well as the physiological significance of their effects have not been ad- dressed.
Stimulation of lacrimal secretion in the rat by the melanocortins, MSH and adrenocorticotropic hormone (ACTH) was first demonstrated in 1982 by Jahn et al. (1982). These investigators sug- gested that MSH-stimulated secretion was media- ted by elevation of intracellular 3',5' cyclic AMP (cAMP) levels. However, it was not known whether MSH actually acts as a primary secretagogue, or through the release of endogenous neurotrans- mitters. The presence of MSH receptors in lacrimal tissue has been recently reported by us and others (Salomon et al., 1989; H a n n e t al., 1989).
Therefore, we first studied some of the pharmacological properties of the melanocortin receptors in the rat lacrimal intra- and extraorbital glands (Salomon et al., 1989). Using [125I][Nle4,D-
PheV]aMSH as radioligand, we employed method- ologies and compared aspects previously studied by us on a mouse melanoma cell MSH receptor, including peptide specificity, the effects of Ca 2+ ions and calmodulin (CaM)-binding peptides (Gerst et al., 1986; 1987; Gerst and Salomon, 1987; 1988). Secondly, we determined the micro- scopic distribution of the MSH receptors in the lacrimal gland. Thirdly, we investigated whether MSH acts as a primary secretagogue in this tissue, independently of catecholamines and acetylcho- line. The data indicate that lacrimal tissue is a good model system for the study of the biochem- ical mechanism of action of MSH in processes unrelated to its classical role as a melanocyte stimulating hormone.
2. Materials and methods
2.1. Materials
aMSH, porcine /3MSH, (L-)epinephrine, car- bamylcholine, dithiothreitol (DTT), atropine, pro- pranolol, phentolamine, melittin, mastoparan, 2,2'-azino-bis(3-ethylbenzthi-azoline-6-sulfonic acid) and para-aminobenzamidine (PABA), bovine serum albumin (BSA) were from Sigma Chemical Co. Gonadotropin-releasing hormone (GnRH) and prolactin were from NIH. [Nle4,D-PheV]aMSH (Sawyer et al., 1980) was a gift from Dr. Lerner (Yale University). ACTH(1-24) was a gift from Organon (Oss, Holland). Lys-Arg-Arg-Trp-Lys- Lys-Asn-Phe-Ile-Ala-Val-Ser-Ala-Ala-Asn-Arg- Phe-Gly-NH 2 (M5) peptide (Kennelly et al., 1987) and vasoactive intestinal peptide (VIP) were synthesized at the Weizmann Institute. Carrier-free iodine 125I was from Amersham. All other materi- als were of analytical grade.
2.2. Animals and surgical procedures
Male Wistar rats (200 _+ 10 g) raised in the departmental facility were fed at libitum and kept at 25°C with 14 h light, 10 h dark. The intraor- bital lacrimal gland as referred to in this study was excised along with the eyeball. The entire glandular tissue immediately surrounding the globe

including the Harderian gland was dissected. The extraorbital gland was removed by an external approach.
2.3. Preparation of rat lacrimal cell membranes
The rats were killed by cervical dislocation (experiments 1-7) or by ether anaesthesia (experi- ment 8), and the glands were excised, collected and homogenized in 10 volumes of ice-cold homo- genization medium (0.3 M sucrose, Tris acetate 20 mM, pH 7.6, 0.1 mM EDTA, 1 mM PABA), using a loosely fitted Dounce homogenizer. The homo- genate was filtered through a No. 130 nylon mesh and centrifuged for 10 rain at 12000 × g. The pellet was discarded and the supernatant was centrifuged further for 60 rain at 105× g. The resulting pellet was resuspended in 10 mM Tris acetate pH 7.4 1 mM DTT and frozen in liquid nitrogen in aliquots that were thawed once shortly before use.
2.4. Iodination of [Nle 4-,D-PheT-] a M S H
[Nle4,D-Phe7]aMSH (5/~g) was iodinated with chloramine-T, as previously described for iodina- tion of f lMSH (Gerst et al., 1986), but using 0.025% chloramine-T and incubating at room tem- perature for 45 s only. After addition of DTT to 0.75 M, and KI to 36 mM, the peptide was im- mediately separated from free iodine by gel filtra- tion on Biogel P-2. The [a25I]iodo-[Nle4,D- PheV]aMSH concentration was determined by its ability to activate MSH-sensitive adenylate cyclase in M2R mouse melanoma cells, as previously de- scribed (Gerst et al., 1986) using a calibration curve made with authentic [Nle4,D-Phe7]aMSH. The specific radioactivity of the labeled peptide was 1 000-3 000 c.p.m./fmol.
2.5. [125I]Iodo-[Nle 4,D-Phe7 ] a M S H binding to rat lacrimal membranes
Binding of hormone to lacrimal gland mem- branes was carried out essentially as described by us for [1251]iodo-flMSH binding to M2R mouse melanoma cell membranes (Gerst et al., 1987). The incubation medium in phosphate-buffered
73
saline (PBS) conta ined [125I]Iodo-[Nle4,D- PheV]aMSH (0.4-2.0 x 105 c.p.m.), 1 m g /m l BSA, 10 ~tg/ml leupeptin, 1 mM PABA, 10/xg/ml soya bean trypsin inhibitor, 1 mM CaC12 and 1 mM MgC12 (binding incubation cocktail) and test sub- stances or varying concentrations of unlabeled [Nle4,D-PheT]aMSH as specified in the individual experiments. Binding was initiated by the addition of 20-90/~g of lacrimal cell membrane protein to a final volume of 100 /~1. Binding was linear with membrane concentration in the range tested and incubation was for 20 min at 30°C, conditions under which equilibrium binding was attained. The binding reaction was terminated by the ad- dition of 2 ml ice-cold PBS containing 1.0 m g /m l BSA, 1 mM CaC12 and 1 mM MgC12. The mem- brane suspension was filtered through 0.22 ~m OE-66 Millipore filters (2.5 cm diameter) pre- soaked in PBS containing 1 m g / m l BSA, and washed twice with 2 ml of ice-cold PBS containing 1 m g /m l BSA. The filters were counted in an autogamma spectrometer. Non-specific binding was determined in the presence of 1/~M unlabeled [Nle4,D-PheV]aMSH and subtracted from the val- ues for total binding. Specific binding (>/80% of total binding) is expressed as fmole of [Nle4,D- PheV]aMSH bound / rag membrane protein, means _+ S.E.M. of triplicate determinations.
2. 6. Calculation of free Ca 2 + concentrations
The free calcium concentrations were set with an EGTA buffer system, as described by us earlier (Gerst et al., 1987), using a computer program kindly made available to us by Dr. T. Abrams (constants derived from Tsien and Rink (1980)).
2.7. [125I]Iodo[Nle4 D-Phe7 ] a M S H binding to rat lacrimal tissue slices for autoradiographic analysis
Freshly excised rat lacrimal glands were quickly frozen on aluminum foil placed on dry ice and stored at - 7 0 ° C until sectioned. For sectioning, the frozen tissue was equilibrated to - 2 0 o C, and mounted onto the cryostat stage. Sections, 10 ~m were prepared using a Reichert-Jung Frigocut N- 2800 cryostat. Sections placed on gelatin-coated

74
microscope slides were stored at - 7 0 ° C until further processed.
For [125I]iodo-[Nle4,D-PheT]aMSH binding to the tissue sections, the slides were brought to room temperature and the sections were covered with a sufficient volume of binding incubation cocktail ( - 100 000 c.p.m./100 ~tl). Incubation was for 30 min at 37 °C at saturated humidity. Subse- quently, the slices were washed twice (15 min each) with ice-cold PBS containing 5 mg /ml BSA.
Further treatment of the sections was as fol- lows. (a) Autoradiography for macroscopic analy- sis: the sections were dried and exposed to Kodak XAR-5 X-ray film with intensifier screen for the period of time indicated and developed. (b) Auto- radiography for microscopic analysis: the wet sec- tions were washed twice with ice-cold PBS and post-fixed with 2% glutaraldehyde. The specimens were further washed with PBS, dehydrated with ethanol, dried and coated with photographic em- ulsion (Ilford, K-5) and exposed for the period of time indicated. After development of the photo- graphic layer, the sections were stained with Toluidine blue, dried and covered with coverslips. Light microscopy was performed with a Zeiss pho- tomicroscope III.
2.8. Determination of protein secretion from lacrimal gland slices
Protein secretion from rat extraorbital lacrimal gland slices was determined as described by Fried- man et al. (1981). Fresh extraorbital glands were collected in Krebs Ringer Bicarbonate (KRB) sup- plemented with 25 mM HEPES, pH 7.4, and 5 mM fl-hydroxybutyric acid (HflKRB) and gassed with 95% 02 - 5% CO 2 mixture at 37°C. The glands were then collected and sliced (450 /~m) with a Mickel tissue slicer. The slices were in- cubated in fresh Hf lKRB for 10 min, collected on a nylon mesh No. 130 and washed extensively with the same solution. Duplicate vials containing tissue-slice equivalents of about 0.75-1 gland were incubated with test substances in a final volume of 2 ml HflKRB, briefly gassed and stoppered. In- cubation (60 min at 37 ° C) was carried out with shaking, and gassing was repeated every 15 min. Under these conditions, secretion is progressive
and has not leveled off within 60 min. Samples of the incubation medium (0.1 ml) were removed at zero time and 60 min and stored in the cold. The remaining tissue was homogenized and kept on ice. Peroxidase activity released during the incuba- tion was taken as a measure of protein secretion and was expressed as percentage of the total activ- ity of enzyme in each vial. The total amount of peroxidase in each vial was taken as 100% and calculated from the sum of peroxidase activity secreted at the end of the experiment plus the remaining activity determined in the tissue homo- genate.
2.9. Peroxidase assay
Peroxidase was determined under conditions modified from those described by Engvall (1980). Enzyme activity was assayed in a final volume of 0.2 ml, in the presence of 25 mM citric acid, 1.46 mM H202, 50 mM sodium phosphate and 0.45 mM 2,2'-azino-bis(3-ethylbenthi-azoline-6-sulfonic acid) at pH 4.6. Assays were performed in dupli- cate with incubation at room temperature; optical density at 600 nm was recorded at 15 min using an ELIZA reader (Bio-Tek Instruments).
2.10. Protein determination
Protein was determined according to Bradford (1976).
3. Results
3.1. [I:5I][Nle4-,D-PheZ]aMSH binding to rat lacrimal cell membranes
We measured MSH binding to cell membranes prepared from both intra- and extraorbital lacrimal glands of the rat. In a representative experiment (fig. 1), binding was shown to be dose-dependent, reaching saturation at 10 nM [125I][Nle4,D- PheV]aMSH. The dissociation constant, KD, for the MSH receptor as calculated from a double reciprocal plot was very similar, 2.0 and 1.3 nM for the intra- and extraorbital gland membranes, respectively (fig. 1). The average values obtained
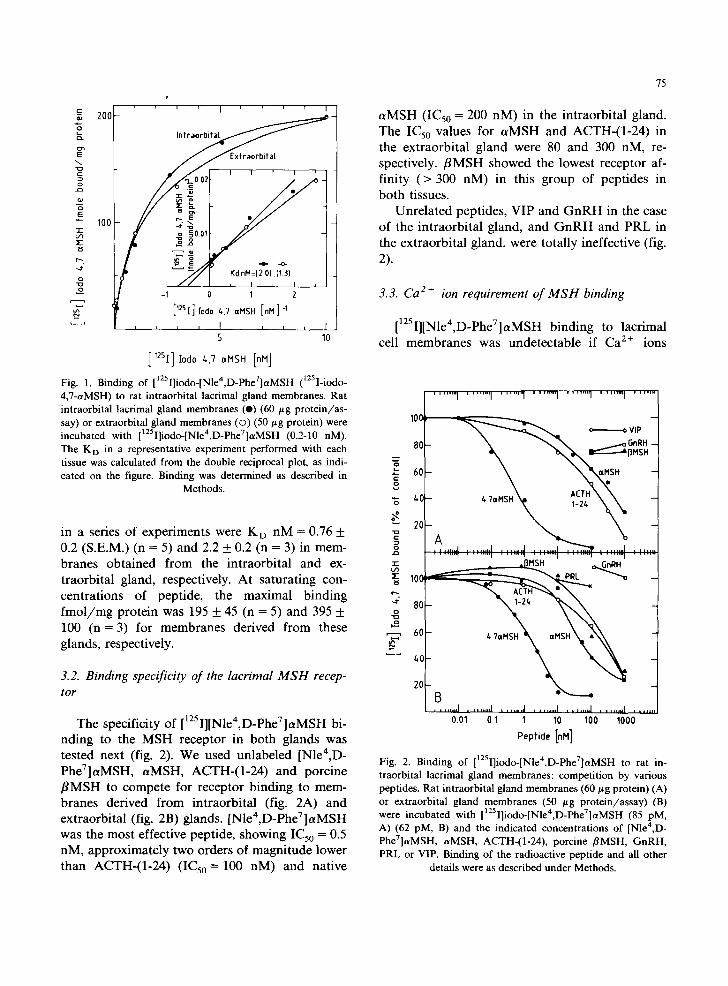
s
__. ' ' I ' , r I
o 200 o ~ - ~. Intra
\ J J Extraorbital
I I i I '
o / S , ' ' / )
+ / -io E ~ E •
100 .., - r , . ~ 0 1
-4" ~ Kd nM={2 O),(13) o [ / / ¢ / , I L I +
"9'° -1 1 2
['2SI]Iod0 t, 7 aHSH [ni l ] - '
, , J , I , L , , l
5 10
[12SI] Iodo ~,7 aHSH [riM]
Fig. 1. Binding of [1251]iodo-[Nle4,D-PheV]c~MSH (tzsI-iodo- 4,7-aMSH) to rat intraorbital lacrimal gland membranes. Rat intraorbital lacrimal gland membranes (e) (60 btg protein/as- say) or extraorbital gland membranes ((3) (50 #g protein) were incubated with []2sI]iodo-[Nle4,D-Phe7]aMSH (0.2-10 nM). The K D in a representative experiment performed with each tissue was calculated from the double reciprocal plot, as indi- cated on the figure. Binding was determined as described in
Methods.
in a series of experiments were K D nM = 0.76 + 0.2 (S.E.M.) (n = 5) and 2.2 + 0.2 (n = 3) in mem- branes obtained from the intraorbital and ex- traorbital gland, respectively. At saturating con- centrations of peptide, the maximal binding fmol /mg protein was 195 + 45 (n = 5) and 395 + 100 (n = 3) for membranes derived from these glands, respectively.
3.2. Binding specificity of the lacrimal M S H recep- tor
The specificity of [125I][Nle4,D-Phe7]aMSH bi- nding to the MSH receptor in both glands was tested next (fig. 2). We used unlabeled [Nle4,D - PheT]aMSH, aMSH, ACTH-(1-24) and porcine f lMSH to compete for receptor binding to mem- branes derived from intraorbital (fig. 2A) and extraorbital (fig. 2B) glands. [Nlea,D-PheT]aMSH was the most effective peptide, showing IC50 = 0.5 nM, approximately two orders of magnitude lower than ACTH-(1-24) (IC50 = 100 nM) and native
75
aMSH (IC50 = 200 nM) in the intraorbital gland. The IC50 values for aMSH and ACTH-(l-24) in the extraorbital gland were 80 and 300 riM, re- spectively, f lMSH showed the lowest receptor af- finity (> 300 nM) in this group of peptides in both tissues.
Unrelated peptides, VIP and G n R H in the case of the intraorbital gland, and G n R H and PRL in the extraorbital gland, were totally ineffective (fig. 2).
3.3. Ca 2 + ion requirement of M S H binding
[]25I][Nle4,D-PheV]aMSH binding to lacrimal cell membranes was undetectable if Ca 2÷ ions
........ I ........ 1 ....... 1 . . . . . . . I ........ I . . . . . . I .......
1 0 ¢ -
=o \
" " : " " " " I . . . . . . . . I . . . . . . . ~ . . . . . . . . I . . . . . . I . . . . . . . . I " : " " ~ " "
• - r ,,r~MSH . ~ GnRH 100
. . t 8 0 - o
'-9'° 60 - l , . 7 a M S H H f
20
. . . . . . . . I . . . . . . . . I . . . . . . . l . . . . . . . . I , , , . . . I . . . . . . . I . . . . .
0 . 0 1 0 1 1 1 0 1 0 0 1 0 0 0
P e p f i d e [nM]
Fig. 2. Binding of [1251]iodo-[Nle4,D-Phe7]aMSH to rat in- traorbital lacrimal gland membranes: competition by various peptides. Rat intraorbital gland membranes (60 #g protein) (A) or extraorbital gland membranes (50 /~g protein/assay) (B) were incubated with [12Sl]iodo-[Nle4,D-Phe7]aMSH (85 pM, A) (62 pM, B) and the indicated concentrations of [Nlea,D - Phe7]aMSH, aMSH, ACTH-(1-24), porcine flMSH, GnRH, PRL or VIP. Binding of the radioactive peptide and all other
details were as described under Methods.

76
c
o '--° -g . . . . . . . . . . . . . . . I I I . . . . . . . . r " . . . . . . ~_
o o E E ~ 20- ~
o o
"1- 212
m I n t r
10
o o
. i t , I . . . . . . I . . . . . . . . I . . . . . . . . I . . . . . . . , - , , - +
3 10 100 1000 A • o
- [Ca2"] ,tee .M
Fig. 3. Calcium-dependent binding of [NIe4,D-Phe7]aMSH to rat lacrimal gland plasma membranes. Rat intraorbital lacrimal gland membranes (O) (30 /Lg proteins/assay) or extraorbital gland membranes (©) (50 ~g/assay) were incubated with [12sI]-[Nle4,D- PheT]aMSH 85 and 62 pM, respectively at varying concentrations of free calcium, as set by means of a calcium/EGTA-Tris buffer
system. [12sI][NIe4,D-PheT]aMSH binding and all other details were as described under Methods.
were omitted and E G T A (10 mM) was included in the incubation medium. The dependence o f [1251] iodo-[Nle4,D-PheV]aMSH binding to lacrimal cell membranes of both sources on ambient free Ca 2÷ was therefore determined (fig. 3). We found that [125I][Nlea,D-PheT]aMSH binding increased over 20-fold in the range of 0.1-2000 /~M free Ca 2÷. The major positive effect ( - 5 - f o l d increase in MSH binding capacity) was seen in the range of 10 to 2000 /~M free Ca 2+, and comprised ap- proximately 80% of the total binding obtained at the saturating Ca 2÷ concentration. The half-maxi- mal Ca 2+ ion concentrat ion was 0.21 m M ___ 0.02 (n = 4) in the intraorbital and 0.17 m M + 0.01 (n = 3) in the extraorbital gland. A smaller effect, comprising only - 20% of the total binding, was seen in the range of 0 .1-10/ tM free Ca 2+ and may represent an additional Ca 2 ÷ dependence site with higher affinity.
3.4. Inhibition of MSH binding by calmodulin-bind- ing peptides
The effect of several CaM-binding peptides on MSH binding was tested next. It was found that M5 (IC50 = /~M 1.5 + 0.1, n = 4), melittin (IC50 =
ttM 3.4 + 0.4, n = 3) and mas toparan (IC50 = /~M 12 + 2, n---3) , all inhibited and at 50 /~M nearly abolished M S H binding to intraorbital gland membranes as shown in a representative experi- ment (fig. 4A). Similar results were observed with the extraorbital gland, but at higher peptide con- centrations, M5 (ICs0 / z M = 13 + 6, n = 3) and melittin (IC50 /~M = 22 + 3, n = 3) as shown in a representative experiment (fig. 4B). As in the case of the melanoma M S H receptor, M5 was found to be the most potent inhibitor in this group of peptides.
3.5. A utoradiographic analysis of MSH-receptor dis- tribution in rat lacrimal glands
Freshly frozen tissue was cryosectioned in order to examine the distr ibution of MSH-receptors in the rat intraorbital lacrimal gland. The 10-ttm- thick slices were collected on microscope slides and incubated for 30 min at 3 7 ° C with [a2sI][Nlea,D-PheV]aMSH under conditions other- wise identical to those used in the binding assay. After washing, the dried slices were exposed to X-ray film (fig. 5). Macroscopic examinat ion of the densi togram revealed that M S H binding sites

( 3 L .
E 0
" I - L ~
I , i i ~ , [ i I I ~ r i i , , , I ~ ,
100 - 8 0 ~ 6O
Q
IoO,8oso 2o'° B
I I I L I I l l l I J I I [ I I I I J I
0.1 1 10 Pepf,de [pM]
1 2 5 4 7 Fig. 4. Inhibition of [ I]iodo-[Nle ,D-Phe ]aMSH binding to rat lacrimal gland membranes by calmodulin-binding peptides. Rat intraorbital lacrimal gland membranes (60/~g protein/as- say) (A) or extraorbital gland membranes (50 /~g protein/as- say) (B) were incubated with calmodulin-binding peptides at the concentrations indicated. []2~I][Nle4,D-PheV]aMSH (85 pM A) (56 pM B). Binding was carried out as described under
Methods.
are homogeneously distributed on the entire slice with clear imprints of the boundaries of the acinar lobes (fig. 5a). The presence of excess (1 /~M) unlabeled [Nlea ,D-PheT]aMSH reduced the bind- ing of the radiolabeled peptide substantially (fig. 5b).
Binding of MSH to the tissue slices was Ca 2+- dependent and reduced to control levels by inclu- sion of 10 m M E G T A in the binding step (fig. 5c), in agreement with the results shown in fig. 3. Similarly, [125I][Nle4,D-Phe7]aMSH binding to the tissue slices was found to be inhibited to near the control levels by 10 /~M melittin (fig. 5d) or 10 /zM M5 (fig. 5e), in agreement with the results shown in fig. 4A. Thus, we took it that the MSH
77
receptor distr ibution in the slice is uniform. Fur- thermore, the basic pharmacology of the MSH receptor, as studied in tissue slices, reflects results obtained in the s tandard binding assays using cell membrane preparations.
For quanti tat ive evaluation of these experi- ments, the specific radioactivity bound to the sec- tions was also counted directly. In early experi- ments, groups of 10 slices each were incubated in test tubes under condit ions otherwise used in the direct b inding assays. Incubat ions were in the presence or absence of 1 /~M unlabeled [Nle4,D - PheV]aMSH. The amount of bound radioactivity was determined by Millipore filtration as de- scribed for binding assays with lacrimal cell mem- branes. We found that the ratio between total []25I][NIe4,D-PheV]aMSH binding to non-specific binding using such material was - 10, with 700 and 900 c.p.m, specifically bound per slice in two experiments. Consequently, in each of the experi- ments carried out with intraorbital gland sections
c
i !
1cm
b
d e
Fig. 5. Binding of []2sI]iodo-[Nle4,D-PheV]aMSH to rat intra- orbital lacrimal gland: autoradiography of freshly frozen cryo- sections. Frozen rat intraorbital lacrimal gland cryosections (10 ttm) were mounted on microscope slides and incubated with [125Iliodo-[Nle4,Phe7]aMSH for 30 rain at 37 °C in the binding assa5 lp_ixture in the absence (a) or presence of 1/tM unlabeled peptide (b), 10 mM EGTA (c), 10 ~M melittin (d) or 10 /.tM M5 peptide (e). The sections were then washed three times with PBS, 5 mg/ml BSA, dried and exposed to X-ray film for 4
days. All other details were as described under Methods.

78
in the first phase of the experiments, three s l ices /group/exper iment were scraped off the mi- croscope slide at the end of the washing step. The radioactivity in this material was counted directly with no further processing, to determine the amount of total and non-specifically bound [125I][NIe4,D-PheT]aMSH. The ratio of specific to non-specific binding obtained in these studies with intraorbital gland slices was 7.2 + 1.0 (n -- 8) with 385-1676 c.p.m./slice specifically bound in the different experiments. The total amount of radio- activity bound/s l ice varied with the surface area of the slices and the amount of radioactivity used in each experiment but varied little within each experiment.
In order to examine the subacinar and cellular MSH-receptor distribution, tissue slices were processed as described above but were post-fixed with glutaraldehyde, covered with photographic emulsion and subjected to microscopic analysis following exposure for 9 days. Autoradiography showed that the radioactive hormone as revealed by silver grains on the photographic plate is dis- tributed over all acinar cells in both the intraor- bital (fig. 6A) and extraorbital (fig. 7A) glands. Grain density was significantly lower in control slices in which incubation with [125I][Nle4,D- Phe7]aMSH was carried out in the presence of excess (1 ~tM) unlabeled [Nle4,D-Phe7]aMSH, in the intraorbital (fig. 6B) and extraorbital (fig. 7B) gland slices. The acinar regions can be identified by the concentric array of individual cell nuclei (Nu). Furthermore, in regions where a clear lumen (L) can be seen the receptors seem to be associ- ated with the perinuclear regions, i.e. at the basal aspect of the acinar cell. These experiments show that MSH receptors are present in both glands in high concentrations and, therefore, are likely to play an important role in lacrimal physiology.
3.6. Stimulation of protein secretion in lacrimal gland slices by MSH
We next examined whether the stimulatory ac- tion of MSH on protein secretion in the rat lacrimal gland results from direct stimulation of the acinar cells by MSH or whether stimulation by the peptide could be mediated by the release of
Fig. 6. Binding of [125I]iodo-[Nle4,D-Phe7]aMSH to rat intra- orbital lacrimal gland: autoradiography of cryosections. Sec- tions were prepared and incubated as described in fig. 5, but were then post-fixed with glutaraldehyde 2% and further washed with PBS, dried and coated with photographic emul- sion (Ilford, K-5) and exposed for 9 days. Incubations were: (A) [12~I]iodo-[Nle4,D-PheV]aMSH alone; (B) control: [12~I]iodo-[Nle4,D-PheT]aMSH+l /~M unlabeled [NIe4,D - PheT]aMSH. The patterns of acini can be identified by the concentric array of acinar cell nuclei (Nu) located at the basal side of each cell and in some cases the actual formation of the acinar lumen (L) can be seen adjacent to the apical side of the acinar cells. All other details were as described under Methods.
acetylcholine or catecholamines endogenously pre- sent in nerve endings. In such a situation, the MSH receptors would have been expected to be distributed on neuronal components within the gland. We stimulated peroxidase secretion in ex- traorbital gland slices by using supramaximal con- centrations of epinephrine (10/~M), carbamylcho- line (10/~M) or [Nlea,D-PheV]aMSH (1 /~M) (fig. 8). The three stimulants increased protein dis-

charge 9-, 11- and 7-fold, respectively, as com- pa red to the uns t imula ted cont ro l (basal) . Pro- p rano lo l (10 /~M) and phen to l amine (10 /~M), b locking /3 and a act ions of ep inephr ine , respec- tively, and a t rop ine (10/~M), b locking the ac t ions of ca rbamylchol ine , had not effect on p ro te in secret ion when a d d e d alone, but effect ively b locked the s t imula tory activit ies of the neuro- t ransmit ters . However , these c o m p o u n d s fai led to affect the secre t ion s t imu la t ed b y [Nlen,D - PheT]aMSH. These results therefore suggested that the ac t ion of M S H as a secretagogue in this t issue
79
40
N 3o
z
~ 2O
r n
CD
~ 0 -4
-IF
II
r ~ r--
Fig. 8. Peroxidase secretion from rat extraorbital gland slices. Extraorbital gland slices were incubated for 60 min in dupli- cates in the absence (Basal) or presence of either epinephrine (10 #M), carbamylcholine (10 ~M), [Nle4,D-Phe7]aMSH (1 /~M), propranolol (10 /~M), phentolamine (10 #M), atropine (10 #M) or combinations of the following at the same final concentrations: epinephrine + propranolol + pbentolamine, carbamylchol ine+ atropine, [NIe4,D-PheT]aMSH + propranolol, [Nle4,D-PheT]aMSH + phentolamine and [Nle4,D-Phe7]aMSH + atropine. Peroxidase activity in the in- cubation medium, as percentage of total enzyme activity (medium + tissue) in each incubation vial, is presented as per- cent secretion. All other details are as described under Meth-
ods.
is exer ted d i rec t ly on the secret ing cells and is not l ikely to be m e d i a t e d b y the release of these en- dogenous classical neuro t ransmi t te r s .
Fig. 7. Binding of [x25I]iodo-[Nle4,D-PheT]aMSH to rat ex- traorbital lacrimal gland: autoradiography of cryosections. As in fig. 6, but using rat extraorbital glands. (A) Total binding of radiolabeled peptide; (B) control for non-specific binding in the presence of 1/~M unlabeled peptide. The patterns of acini can be identified by the concentric array of acinar cell nuclei (Nu) located at the basal side of each cell and in some cases the actual formation of the acinar lumen (L) can be observed on the apical side of the acinar cells. All other details were as
described under Methods.
4. Discuss ion
Our results show that the M S H - r e c e p t o r in bo th rat l ac r imal g lands exhibi ts s imilar pha rma - cological proper t ies . Fu r the rmore , these proper t ies are bas ica l ly s imi lar wi th respect to recep tor affin- it ies to wha t we (Sa lomon et al., 1989; Gers t et al., 1986; 1987) and others (Eberle , 1988; Sawyer et al., 1980; Scimonel l i and Ebefle , 1987) r epor t ed for m e l a n o m a cells.
Whi le a M S H and ACTH-(1 -24 ) have bas ica l ly s imi lar IC50 s in these glands, f l M S H seems to

80
have the lowest relative affinity in this group, being essentially unable to compete with [Nle4,D- PheV]aMSH in the case of the intraorbital gland. This is consistent with the inability of f lMSH (1 I~M) to stimulate cAMP production in intraorbital gland slices (data not shown). There was a previ- ous report (Gerst et al., 1986) that ACTH-(1-24), /3MSH and aMSH were essentially equal in their affinity for the M2R melanoma cell receptor. The question of whether this difference qualifies for a separate classification of the glandular MSH-re- ceptor vis-h-vis the melanoma-receptor remains to be tested. The generally similar response of the lacrimal tissue receptor system towards native aMSH and ACTH-(1-24) warrants its designation as a melanocortin receptor. Due to operational considerations, we refer to this receptor as an MSH receptor also even though this semantic choice may be a matter for some debate.
The interesting Ca 2+ requirement for MSH binding, as first reported by us for M2R melanoma cells (Gerst et al., 1987) with porcine flMSH, seems to hold equally well in the case of the lacrimal cell MSH-receptors and to be indepen- dent of the type of peptide used, since [Nle4,D- PheV]aMSH was used in the present study (fig. 3). Ca 2+ was found to increase the affinity of MSH for receptor binding in melanoma cells 20-fold. This effect was reversible since the peptide-recep- tor complex dissociated readily upon addition of EGTA and re-associated following replenishment of free Ca 2+ ion to millimolar concentrations (Gerst et al., 1987). While the molecular basis for this requirement is not yet clear, it appears to be a general property of this class of receptors. The possible physiological significance of this peculiar ionic requirement may indeed be more relevant to the acute and dynamic control of protein secre- tion, which itself is a CaZ+-requiring process, than in the case of the long-lasting trophic control of melanogenesis. Also, in the case of the M2R cell MSH receptors, we observed two CaZ+-depen - dence sites saturating in the micromolar and sub- millimolar ranges, respectively. This may implicate two independent Ca 2 + binding sites in the control of MSH receptor activity. The similarity continues with respect to the inhibition of receptor binding activity by various CaM-binding peptides (fig. 4)
which we have described previously for the melanoma cell MSH-receptor (Gerst and Salomon, 1987; 1988). Even though the effect of these peptides implies the possible involvement of CaM or a similar calcium-binding protein in MSH-re- ceptor function, no direct link has been estab- lished between this line of evidence and the Ca 2÷ requirement of receptor activity. It is logical to assume that melanocortins regulate lacrimal func- tion in the rat in vivo, although direct evidence in proof of this assumption is lacking. In this regard, the observation made by Tatro and Reichlin (1987), that [Nlen,D-PheV]aMSH uptake by lacrimal tissue of the rat and mouse was the highest as compared to that of other organs, is most interesting and rather surprising if viewed according to classical concepts and terminology that earmark MSH peptides for the control of pigmentation. Whether MSH acts in lacrimal tis- sue as a hormone or may be released from nerve endings as a neurotransmitter also remains to be established. It should be mentioned that VIP has been shown to be present in "nerve endings in the cat lacrimal gland (Uddman et al., 1980) and to act as secretagogue in the rabbit lacrimal gland (Dartt et al., 1988). Moreover, the involvement of several peptides/neuropeptides in the regulation of exocrine glands, such as salivary glands (VIP and substance P) (Ekstrom et al., 1983; Martinez and Martinez, 1981) and the exocrine pancreas (cholecystokinin, gastrin, substance P and bombe- sin, VIP and PHI) (Dimaline and Dockray, 1980; Gardner and Jensen, 1980), has been described. Their coexistence along with classical neurotrans- mitters and their exact role in controlling exocrine function has to be evaluated further.
We have now shown MSH to be a primary secretagogue in the rat lacrimal gland, as its activ- ity was unaffected by the presence of specific blockers (phentolamine, propranolol and atropine) of the classical neurotransmitters (fig. 8). This result eliminated the possibility that MSH stimu- lates secretion indirectly by release of endogenous neurotransmitters. Our study also showed for the first time that MSH-receptors heavily populate the lacrimal gland in its entirety (fig. 5), and that MSH receptors are located on practically all acinar cells and are not confined to a selected population

or g roup of cells w i t h i n the g land . T h e a s soc i a t i on
o f the M S H recep to r s w i th the basa l a spec t s o f the
a c i n a r cells is sugges ted f r o m o b s e r v a t i o n s o f re-
g ions in the t issue sec t ion in wh ich a c lea r l u m e n ,
l imi t ed by the ap ica l s ide of the ac ina r cells can be
seen (figs. 6, 7). A d is t inc t c o n c l u s i o n in this
regard , however , m u s t awai t the d e v e l o p m e n t of
t e c h n i q u e s p e r m i t t i n g r e c e p t o r i m a g i n g in m u c h
t h inne r t issue sec t ions or, a l t e rna t ive ly , the c o m -
p a r i s o n of M S H - r e c e p t o r c o n t e n t on basa l a n d
ap ica l a c ina r m e m b r a n e s p r e p a r e d d i f fe ren t ia l ly ,
as p r o p o s e d by M i r c h e f f et al. ( M i r c h e f f et al.,
1983). W e be l i eve tha t the ins t an t and acu te secre-
to ry r e sponse o f l ac r ima l t issue to M S H , p r e sen t s
a h igh ly a p p r o p r i a t e m o d e l for the s tudy of the
d y n a m i c aspec ts of M S H ac t ion in n o n - m e l a n o -
gen ic t issue wh ich seems to r e s e m b l e those o f
n e u r o p e p t i d e s ac t ing as n e u r o t r a n s m i t t e r s r a the r
t h a n h o r m o n e s .
A c k n o w l e d g e m e n t s
We wish to thank Ms. Rachel Benjamin for her devoted secretarial assistance, and M. Tosky and J. Schultz for their critical review of the manuscript. Y.S. is the incumbent of the Charles W. and Tillie K. Lubin Professorship of Hormone Research.
References
Bardin, C.W., C. Shaha, J.P. Mather, Y. Salomon, A.N. Margioris, A.S. Liotta, I. Gerendai, C.-L. Chen and D.T. Krieger, 1985, Identification and possible function of pro- opiomelanocortin-derived peptides in the testis, Ann. N.Y. Acad. Sci. 438, 346.
Bijlsma, W.A., F.G.I. Jennekens, P. Schotman and W.H. Gispen, 1981, Effects of corticotropin (ACTH) on recovery of sensorimotor function in the rat: structure-activity study, European J. Pharmacol. 76, 73.
Bijlsma, W.A., F.G.I. Jennekens, P. Schotman and W.H. Gispen, 1983, Stimulation by ACTH(4-10) of nerve fibre regeneration following sciatic nerve crush, Muscle Nerve 6, 104.
Boitani, C., D. Farini, R. Canipari and C.W. Bardin, 1989, Estradiol and plasminogen activator secretion by cultured rat Sertoli cells in response to melanocyte-stimulating hormones, J. Andol. 10.
Bradford, M.M.A., 1976, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem. 72, 248.
81
Dartt, D.A., M. Shulman, K.L. Graw, S.R. Rossi, C. Matkin and J.P. Gilbard, 1988, Stimulation of rabbit lacrimal gland secretion with biologically active peptides, Am. J. Physiol. G300.
De Wied, D. and W. Ferrari, 1986, Central Actions of ACTH and Related Peptides, Symposia in Neuroscience, Vol. 4 (Springer-Verlag, Berlin).
De Wied, D. and J. Jolles, 1982, Neuropeptides derived from proopiomelanocortin: behavioral, physiological, and neuro- chemical effects, Physiol. Rev. 62, 976.
Dimaline, R. and G.J. Dockray, 1980, Actions of a new peptide from porcine intestine (PHI) on pancreatic secretion in the rat and turkey, Life Sci. 27, 1947.
Eberle, A.N. 1988, The Melanocortins: Chemistry, Physiology and Mechanisms of Action (Karger, Basel).
Ekstrom, J., B. Mansson and G. Tobin, 1983, Vasoactive intestinal peptide evoked secretion of fluid and protein from rat salivary glands and the development of supersensi- tivity, Acta Physiol. Scand. 119, 169.
Engvall, E., 1980, Enzyme immunoassay: ELISA and EMIT, Meth. Enzymol. 70, 419.
Friedman, Z.Y., M. Lowe and Z. Selinger, 1981, fl-Adrenergic receptor stimulated peroxidase secretion from rat lacrimal gland, Biochim. Biophys. Acta 675, 40.
Gardner, J.D. and R.T. Jensen, 1980, Receptor for secreta- gogues on pancreatic acinar cells, Am. J. Physiol. 238, G63.
Gerst, J.E. and Y. Salomon, 1987, Inhibition by melittin and fluphenazine of melanotropin receptor function and adenylate cyclase in M2R melanoma cell membranes, En- docrinology 121, 1766.
Gerst, J.E. and Y. Salomon, 1988, A synthetic analog of the calmodulin binding domain of mymosin light chain kinase inhibits melanotropin receptor function and activation of adenylate cyclase, J. Biol. Chem. 263, 7073.
Gerst, J.E., J. Sole, J.P. Mather and Y. Salomon, 1986, Regu- lation of adenylate cyclase by/3-melanotropin in the M2R melanoma cell line, Mol. Cell. Endocrinol. 46, 137.
Gerst, J.E., J. Sole and Y. Salomon, 1987, Dual regulation of receptor function and adenylate cyclase by calcium and guanosine nucleotides in the M2R melanoma cell line, Mol. Pharmacol. 31, 81.
Hann, L.E., J.B. Tatro and D.A. Sullivan, 1989, Morphology and function of lacrimal gland acinar cells in primary culture, Invest. Ophthalmol. Vis. Sci. 30, 145.
Jahn, R., U. Padel, P.H. Porsch and H.D. SOling, 1982, Adren- ocorticotropic hormone and a-melanocyte-stimulating hormone induce secretion and protein phosphorylation in the rat lacrimal gland by activation of a cAMP-dependent pathway, European J. Biochem. 126, 623.
Johnston, M.F., E.A. Kravitz, H. Meiri and R. Rahamimoff, 1983, Adrenocorticotropic hormone causes long-lasting potentiation of transmitter release from frog motor nerve terminals, Science 220, 1071.
Kennelly, P.J., A.M. Edelman, D.K. Blumenthal and E.G. Krebs, 1987, Rabbit skeletal muscle myosin light chain kinase: the calmodulin binding domain as a potential active site-directed inhibitory domain, J. Biol. Chem. 262, 11958.

82
Liotta, A.S., C. Loudes, J.F. McKelvy and D.T. Krieger, 1980, Biosynthesis of precursor corticotropin/endorphin-, corti- cotropin-, a-melanotropin-, fl-lipotropin-, and fl-en- dorphin-like material by cultured neonatal rat hypo- thalamic neurons, Proc. Natl. Acad. Sci. U.S.A. 77, 1880.
Mains, R.E., B.A. Eipper and N. Ling, 1977, Common pre- cursor to corticotropins and endorphins, Proc. Natl. Acad. Sci. U.S.A. 74, 3014.
Martinez, J.R. and A.M. Martinez, 1981, Stimulatory and inhibitory effects of substance P on rat submandibular secretion, J. Dent. Res. 60, 1031.
Mircheff, A.K., C.C. Lu and C.N. Conteas, 1983, Resolution of apical and basal-lateral membrane populations from rat exorbital gland, Am. J. Physiol. 245, G661.
Saint-Come, C., G.R. Acker and F.L. Strand, 1982, Peptide influences on the development and regeneration of motor performance, Peptides 3, 439.
Salomon, Y., H. Leiba and N.B. Garty, 1989, the MSH-recep- tor in the rat lacrimal gland: location and pharmacology, in: 71st Annual Meeting of the Endocrine Society Meeting, Seattle, Washington, June 21-24 (abstract).
Sawyer, T.K., P.J. Sanfilippo, V.J. Hruby, M.H. Engel, C.B. Heward, J.B. Burnett and M.E. Hadley, 1980, 4-Norleucine, 7-D-phenylalanine-a-melanocyte-stimulating hormone: a highly potent a-melanotropin with ultralong biological ac- tivity, Proc. Natl. Acad. Sci. U.S.A. 77, 5754.
Scimonelli, T. and A.N. Eberle, 1987, Photoaffinity labelling of melanoma cell MSH receptors, FEBS Lett. 226, 134.
Smith, A.I. and J.W. Funder, 1988, Proopiomelanocortin processing in the pituitary, central nervous system, and peripheral tissues, Endocrinol. Rev. 9, 159.
Strand, F.L., T.T. Kung and C. Saint-Come, 1981, Regenera- tive ability of spinal motor systems influenced by ACTH/MSH peptides, in: Functional Recovery from Brain Damage, eds. M.W. Van Hof and G. Mohn (Elsevier, North Holland) p. 369.
Tanaka, A., B.T. Pickering and C.H. Li, 1962, Relationship of chemical structure to in vitro lipolytic activity of peptides occurring in adrenocorticotropic and melanocyte-stimulat- ing hormones, Arch. Biochem. Biophys. 99, 294.
Tatro, J.B. and S. Reichlin, 1987, Specific receptors for a- melanocyte-stimulating hormone are widely distributed in tissues of rodents, Endocrinology 121, 1900.
Tsien, R.Y. and T.J. Rink, 1980, Neutral carrier ion-selective microelectrodes for measurement of intracellular free calcium, Biochim. Biophys. Acta 599, 623.
Uddman, R., J. Alumets, B. Ehinger, R. H~lkanson, I. Loren and F. Sundler, 1980, Vasoactive intestinal peptide nerves in ocular and orbital structures of the cat, Invest. Ophthal- mol. Vis. Sci. 19, 878.





























