Embed Size (px)
Citation preview

Tp
Ua
b
c
a
ARRAA
KAPVPTP
1
l1pvhas(t2impemtI
0d
Toxicology Letters 186 (2009) 139–145
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
he mycotoxins alternariol and alternariol methyl ether negatively affectrogesterone synthesis in porcine granulosa cells in vitro
. Tiemanna, W. Tomeka, F. Schneidera, M. Müllerc, R. Pöhlanda, J. Vanselowb,∗
Research Unit Reproductive Biology, Research Institute for the Biology of Farm Animals (FBN), Dummerstorf, GermanyResearch Unit Molecular Biology, Research Institute for the Biology of Farm Animals (FBN), Wilhelm-Stahl-Allee 2, 18196 Dummerstorf, GermanyLeibniz-Centre for Agriculture Landscape Research, Institute of Landscape Matter Dynamics, Müncheberg, Germany
r t i c l e i n f o
rticle history:eceived 3 November 2008eceived in revised form 17 December 2008ccepted 12 January 2009vailable online 20 January 2009
eywords:lternaria toxinsig granulosa cells
a b s t r a c t
Mycotoxins as contaminants of animal food can impair fertility in farm animals. In the regulation of femalefertility the ovarian steroid hormone progesterone (P4) plays an important role. In the present study wehave investigated the influence of the mycotoxins alternariol (AOH), alternariol mono-methyl ether (AME),and tenuazonic acid (TeA) on cell viability, P4 synthesis, abundance of the key enzymes of P4 synthesis,P450 cholesterol side-chain cleavage enzyme (P450SCC) and 3-�-hydroxysteroid dehydrogenase (3-�-HSD), and of the corresponding Cyp11a1 and Hsd3b transcripts in cultured pig granulosa cells. Already0.8 �M, AOH and AME inhibited P4 secretion and 1.6 �M also significantly reduced cell viability. Theabundance of P450scc protein but not of Cyp11a1 or Hsd3b transcripts was already significantly reduced
iabilityrogesterone synthesisranscript abundancerotein expression
by 0.8 �M AOH and AME. 1.6 �M AOH but not AME significantly reduced the abundance of �-tubulin andalso clearly affected actin protein concentrations. TeA neither impaired viability nor P4 secretion. Alsomycotoxin extracts isolated from naturally occurring Alternaria strains by HPLC purification inhibited cellviability and P4 synthesis, however at higher concentrations compared to AOH and AME. In conclusion,AOH and AME, but not TeA specifically inhibited P4 secretion in cultured porcine granulosa cells. Alternaria
may
toxin contaminated foodspecies.. Introduction
Mycotoxins are biologically active secondary fungal metabo-ites found as contaminants of food- and feedstuff (Fink-Gremmels,999; King and Schade, 1984). Alternaria toxins are mycotoxinsroduced by Alternaria species, which grow on corn, rice, fruits,egetables and oilseed, and when fed to animals or human beings,ave been shown to exert toxic effects (De Lucca, 2007; Scott etl., 2006; Weidenborner, 2001). Alternaria alternata and A. tenuis-ima are able to produce toxic metabolites which are alternariolAOH), alternariol mono-methyl ether (AME), altenuene (ALT), andenuazonic acid (TeA) (Müller, 1991; Schade and King, 1984; Scott,001). Owing to the lack of toxicological data for these mycotox-
ns, in vitro studies may contribute to hazard identification anday elucidate their mechanisms of action on mammalian cells. In
ig, reproductive performance can be sensitive to high natural lev-
ls of mycotoxins in their diet (Tiemann and Dänicke, 2007). Theycotoxins �- and �-zearalenol inhibit the progesterone (P4) syn-hesis in porcine granulosa cells in vitro (Tiemann et al., 2003a).nteractions of �-zearalenol with cytoplasmic estrogen receptors
∗ Corresponding author. Tel.: +49 38208 68706; fax: +49 38208 68702.E-mail address: [email protected] (J. Vanselow).
378-4274/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.toxlet.2009.01.014
therefore affect reproductive performance in pig and other mammalian
© 2009 Elsevier Ireland Ltd. All rights reserved.
might be responsible for these effects as it has been demonstratedin cultured porcine endometrial cells (Tiemann et al., 2003b). Theinfluence of Alternaria metabolites on viability and steroidogenesisof ovarian cells in pigs is still unknown. Therefore, we have inves-tigated whether the cell viability and P4 production by culturedporcine granulosa cells can be affected by different Alternaria tox-ins administered (1) as pure reference substances AOH, AME, orTeA and (2) as naturally occurring mixtures of toxins isolated fromdifferent Alternaria strains. P4 is an important ovarian hormoneinvolved in preparing the reproductive tract for zygote implanta-tion and maintaining pregnancy (Hadley, 1995). Porcine granulosacells in primary culture are commonly used as a model to studygranulosa cell function. These cell monolayers retain hormonalresponsiveness and physiological functions thus resembling the invivo condition (Albertini et al., 2001).
During P4 biosynthesis, important enzymatic steps are theconversion of cholesterol to pregnenolone by the mitochon-drial cytochrome P450 cholesterol side-chain cleavage enzyme(P450scc) (Lahav et al., 1996) and of pregnenolone to P4 by 3-
�-hydroxysteroid dehydrogenase (3-�-HSD). These enzymes areencoded by the Cyp11a1 and Hsd3b genes, respectively. Directeffects of Alternaria toxins on the content of the P450scc protein andon the abundance of Cyp11a1 and Hsd3b transcripts are unknownand have been investigated in the present study. The role of
1 ogy Le
mh1it
2
2
(fr(G
2
2(sffirptrowwdHwr
(VzAw
wrwT
a
2
dcca(tfstaevae
tcRttmsThas
40 U. Tiemann et al. / Toxicol
icrotubules in regulating ovarian granulosa cell steroidogenesisas been demonstrated earlier (Cameron et al., 1995; Chen et al.,994; Denkova et al., 1992). Therefore, we also investigated thenfluence of AOH and AME on the cytoskeleton components �-ubulin and actin.
. Material and methods
.1. Materials
Chemicals and tissue culture supplies were obtained from Sigma–AldrichDeisenhofen, Germany) if not indicated otherwise. Acrylamide and bisacrylamideor electrophoresis were from Serva (Heidelberg, Germany). All solvents andeagents used in the extraction and high-performance liquid chromatographyHPLC) analysis were analytical or HPLC grade from Mallinckrodt-Baker (Griesheim,ermany).
.2. Preparation of Alternaria extracts from rice
Alternaria strains were isolated from cereals and forage grasses in 2006 and007 and identified as A. alternata (Fr.) Keissler in accordance with the literatureEllis, 1971). A rice medium (5 g polished rice and 5 ml water) was inoculated withmall pieces of a single spore culture of 9 different Alternaria strains and incubatedor 21 days at 22 ◦C. The rice–mycel-mixtures were minced, extracted and puri-ed according to Visconti et al. (1986) and Li and Yoshizawa (2000). Briefly, theice samples were extracted with acetonitrile/KCl solution for 30 min. After proteinrecipitation, the filtrate was divided into two parts. One part was extracted threeimes with dichloromethane. The organic phases were combined, evaporated andedissolved in 2 ml of methanol for HPLC analyses of AOH, AME and ALT. The sec-nd part of the crude extract was acidified (pH 2) with 6N HCl and extracted twiceith dichloromethane. Finally, TeA was extracted from the combined organic phasesith aqueous sodium bicarbonate, acidified to pH 2 again, re-extracted twice withichloromethane, evaporated to dryness and redissolved in 2 ml of methanol. ThePLC apparatus was an Jasco PU 1580 system (Groß-Umstadt, Germany) equippedith a degasser, a quaternary pump, an autosampler as well as an UV/vis and fluo-
escence detector.The toxins were analysed by HPLC after isolation on a reverse phase column
LiChrospher 100 RP 18 with guard RP 18, 5 �m particle size, 250 mm × 4.6 mm i.d.,DS OptiLab Berlin, Germany). Methanol/water (75:25, v/v) containing 300 mg ofinc sulphate l−1 was used as eluent with a flow rate of 0.6 ml min−1. AOH, AME andLT concentrations were estimated by fluorescence detection at excitation/emissionavelengths of 253/415 nm.
TeA was detected by measuring the UV-absorbance at 280 nm. Detection limitsere 0.19 �M or 0.18 �M for AOH and AME and 0.34 �M or 0.44 �M for ALT and TeA,
espectively. Each analysis was performed in duplicate. Photodiodearray detectionas performed to control toxin identity, reference substances of AOH, AME, ALT and
eA were used as external standards.After HPLC analyses, 1 ml of the methanol extracts were evaporated to dryness
nd redissolved in dimethyl sulphoxide (DMSO) for the toxicological tests.
.3. Preparation and culture of pig granulosa cells
Tissue collection and isolation of porcine granulosa cells were carried out asescribed by Tiemann et al. (2007). Ovaries from pigs were collected in a commer-ial slaughterhouse and transferred to PBS. Two hours after slaughter, granulosaells were aspirated from follicles (3–6 mm in diameter) with the help of a syringend the granulosa cells were flushed with complete Hank’s balanced salt solutionHBSS). To disperse cell clusters the cell aggregates were resuspended several timeshrough a Pasteur pipette. The cells were washed with HBSS, centrifuged at 200 × gor 5 min and resuspended in complete TCM199 medium containing 10% fetal calferum (FCS) and 1% ABAM (penicillin, 100 IU/ml; amphotericin B, 250 ng/ml; strep-omycin, 100 �g/ml). The proportion of live cells with intact membranes was ∼70%,s measured by trypan blue exclusion. Aliquots of granulosa cells were culturedither in 96-well plates with approximately 1 × 105 cells per well to estimate celliability and P4 secretion, in 24-well plates to analyse the number of cells and thebundance of proteins and transcripts at 37 ◦C in a humidified 95% air—5% CO2
nvironment.After 24 h, the culture medium was changed and the granulosa cells were cul-
ured in complete medium for additional 24 h. Subsequently, after reaching 50–70%onfluency the cells were rinsed with HBSS and a serum- and phenol red-freePMI-medium was added supplemented with 1% ABAM, 1% ITS (selenium, 4 ng/ml);ransferring, 2.5 �g/ml; insulin, 10 ng/ml), and different additives (see below) and
he cells were cultured for additional 24 h. Control cultures were treated withedium alone. AOH, AME, and TeA reference substances were solubilized as stockolutions at a concentration of 20 mM in DMSO and then diluted in RPMI-medium.he final concentration of DMSO in the medium did not exceed 0.2%, which did notave any detectable effect on the cells. Mycotoxin serial dilutions were preparedt concentrations ranging from 0.8–100 �M. The concentration range used in thistudy corresponded to levels reported by Lehmann et al. (2006).
tters 186 (2009) 139–145
In experiment 1, for estimation of the cell viability and P4 concentration, mono-layers were incubated in the absence or presence of TeA, AOH and AME for 24 h. Totest whether the cells were able to recover from mycotoxin effects on P4 secretion,medium with AME or AOH was removed and the cells were further incubated intoxin-free medium in the presence of 10% FCS. After one day the cells were cen-trifuged at 800 × g for 10 min. The medium was collected and stored at −20 ◦C for P4
quantification.In experiment 2 effects of mycotoxins on the number of cells were determined.
Monolayers of granulosa cells were incubated with or without different concentra-tions of TeA, AOH, and AME at final concentrations ranging from 0.8 �M to 100 �Mfor 24 h.
Experiment 3 was carried out to measure Gapdh (encoding glyceraldehyde-3-phosphate dehydrogenase as a housekeeping control), Cyp11a1 and Hsd3btranscripts, encoding P450scc and 3-�-HSD, respectively. For this, granulosa cellswere incubated in the absence or presence of 0.8 �M, 1.6 �M, and 6.4 �M AOH orAME.
Experiment 4 was performed to determine the expression of P450scc, �-tubulinand actin in the absence or presence of 0.8 �M, 1.6 �M, 6.4 �M AOH or AME. Theproteins were detected by Western blotting. To test whether the cells were able torecover their ability to synthesize P450scc and �-tubulin proteins, AOH containingmedium was removed and the cells were further incubated for one day with toxin-free medium containing 10% FCS.
For the estimation of protein and transcript abundance the medium wasremoved and the attached cells were stored at −80 ◦C until analysis.
Experiment 5 was carried out to investigate effects of mycotoxin extracts derivedfrom nine naturally occurring Alternaria strains grown on a rice medium on theviability and P4 synthesis of cultured granulosa cells. The HPLC analyzed extractswere dissolved in DMSO and diluted with medium to concentrations ranging from0.2 �g to 25 �g/ml AOH, AME, ALT and TeA.
2.4. Cell viability and number
After removing the culture medium for progesterone radioimmunoassay,subsequently granulosa cell viability was measured in 96-well plates with the 3-[4,5-dimethyldiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT, Sigma–Aldrich)assay as described previously (Tiemann et al., 2007). To estimate the number of cells,the cell monolayers were washed with HBSS (without Ca2+and Mg2+) and than incu-bated with 500 �l trypsin–EDTA (0.02–0.05%) solution per well for 30 min to detachthe cells from the plate. The number of detached cells was estimated by a cell counter(Coulter-Multisizer, Krefeld, Germany). For counting, 100 �l of cells were suspendedin 10 ml 0.9% NaCl solution. Each sample was analyzed twice.
2.5. Progesterone concentration
The concentration of P4 in the culture medium of granulosa cells was mea-sured after 24 h by a 3H-RIA previously described by Schneider et al. (2002). Briefly,the main components of the P4 assay were [1,2,6,7-3H)progesterone as tracer andan antibody raised in rabbits by immunization with an 11hydroxy-progesteroneconjugate. The antibodies were purified by affinity chromatography on protein Asepharose. The sensitivity of the RIA was 6 pg/ml. The intra-assay coefficient ofvariations was 7.7% and the inter-assay precision 10.1%.
2.6. RNA preparation and determination of transcript abundance
The abundance of Cyp11a1, Hsd3b and Gapdh transcripts encoding the keyenzymes of P4 synthesis, P450scc and 3-�-HSD, and GAPDH as a housekeeping con-trol, was determined after incubation with or without 0.8 �M, 1.6 �M and 6.4 �MAOH or AME for 24 h.
Total RNA was isolated using QIAshredderTM Homogenizers and the RNeasyMini Kit (both from Qiagen, Hilden, Germany). Reverse transcription was per-formed with M-MLV reverse transcriptase, RNase H Minus, Point Mutant (Promega,Mannheim, Germany) and quantitative real-time PCR (qPCR) with the LC 480 SYBRGreen I Master Kit in a LightCycler® 480 instrument (both from Roche, Mannheim,Germany). The procedures are described in detail in Tiemann et al. (2007). Gene-specific primers for reverse transcription (RT) and PCR were derived from theporcine Cyp11a1 (EMBL/GenBank acc. no., X13768), RT-primer, 5′-CCAGGTCTT-GGTCCTGAACAGAC-3′ , PCR-primers, 5′-TTTACAGGGAGAAGCTCGGCAAC-3′ and 5′-T-TACCTCCGTGTTCAGGACCAAC-3′ , Hsd3b (EMBL/GenBank acc. no AF232699), RT-primer, 5′-CTATGCTGCTGGTGTGGATGAAG-3′ , PCR-primers, 5′-AGGGTTTCTGGGTC-AGAGGATC-3′ and 5′-CGTTGACCACGTCGATGATAGAG-3′ , and Gapdh mRNA sequence(EMBL/GenBank acc. no., U48832), RT-primer, 5′-CAGTCTTCTGGGTGGCAGTGATG-3′ , PCR-primers, 5′-CAGGAGGCATTGCTGATGATCTTG-3′ and 5′-TACATGGTCTACATG-TTCCAGTATG-3′ .
2.7. Expression of P450scc, ˛-tubulin, and actin proteins
Expression of P450scc, �-tubulin, and actin proteins in cultured granulosa cells(106 cells per sample) was analyzed by Western Blotting as described in Tiemann etal. (2007). P450scc was detected with the polyclonal antiserum AB1244 (ChemiconInternational, Temecula, USA) and �-tubulin with monoclonal antibody T-5168. To

ogy Letters 186 (2009) 139–145 141
dc
2
ppcvCm
3
3p(
obtaAw6o
FofofViv
Fig. 2. Effects of AOH and of AME on numbers of porcine granulosa cells. Cells werecultured for 48 h in the presence of 10% fetal calf serum and for additional 24 hin serum- and phenol red-free medium without or with different concentrations
U. Tiemann et al. / Toxicol
etect actin proteins, among them the ubiquitous cytoskeletal �-actin, the mono-lonal antibody A-2066 (Sigma–Aldrich) was used.
.8. Statistical analysis
Data are shown as means ± S.E.M. of two to four independent granulosa cellreparations with one to four replicas each (see figure legends). To facilitate com-arability of measured values from different granulosa cell preparations, data werealculated relative to untreated controls. For statistical analysis one-way analysis ofariance (ANOVA, SigmaStat Statistical Analysis System, Jandel Scientific, San Rafael,A) was used. Comparisons between groups were performed using a protected New-an Keuls test. A value of P < 0.05 was set as the limit of statistical significance.
. Results
.1. Effects of TeA, AOH and AME on viability, number, androgesterone secretion of cultured porcine granulosa cellsexperiments 1 and 2)
Granulosa cells were cultured at doses of 0.8–12.8 �M of AOHr AME and of 6.4–100 �M of TeA for 24 h. Subsequently their via-ility was measured by means of a colorimetric MTT assay andhe media were evaluated for progesterone concentration. Fig. 1A
nd B shows that the cell viability was significantly reduced byOH and AME at concentrations of 1.6 �M and higher comparedith the unexposed control. TeA in the concentration range from.4 �M to 100 �M did not influence cell viability. Inhibitory effectsf AOH and AME on the basal P4 secretion of cultured granulosa
ig. 1. Effects of AOH, AME and of TeA on P4 secretion (bars) and cell viability (lines)f cultured porcine granulosa cells. Cells were cultured for 48 h in the presence of 10%etal calf serum and for additional 24 h in serum- and phenol red-free medium with-ut or with different concentrations of mycotoxins. Culture media were collectedor P4 radioimmunoassay and MTT assays were made with remaining attached cells.alues are relative to untreated controls and are expressed as means ± S.E.M. of four
ndependent granulosa cell preparations with four replicas each. Asterisks denotealues that were significantly different from respective controls (P < 0.05).
of both mycotoxins. Values are relative to untreated controls and are expressedas means ± S.E.M. of two independent granulosa cell preparations with four repli-cas each. Asterisks denote values that were significantly different from respectivecontrols (P < 0.05).
cells were even more pronounced. Mean P4 concentrations in cul-tured cells from different granulosa cell preparations ranged from24 pg/100 �l to 225 pg/100 �l after 24 h, resulting in a mean P4 con-centration of 171 ± 41 pg/100 �l (mean of means ± S.E.M., n = 7) inuntreated control samples. Already with 0.8 �M AOH or AME, theseconcentrations were significantly reduced by 40% and 60%, respec-tively. TeA in the concentration range from 6.4 �M to 100 �M didnot influence P4 secretion. Fig. 2 demonstrates that the number ofadherent cells was only reduced by AOH and AME at concentrationsof 12.8 �M and higher.
To determine whether the inhibitory effects of AME or AOH onP4 synthesis were reversible, granulosa cells were treated with AMEor AOH for 24 h. Subsequently toxins were removed and the cellswere cultured for another 24 h in a toxin-free 10% FCS containingmedium. After preincubation with 0.8 �M and 1.6 �M but not with6.4 �M AOH this resulted in a significant recovery of the P4 secretioncompared with the control (Fig. 3A). The recovery of P4 secretionafter preincubation with 0.8 �M and 1.6 �M AME, however, was notcomplete. After preincubation with 6.4 �M AME, the recovery wasseverely impaired (Fig. 3B).
3.2. Effects of AOH and AME on the abundance of Gapdh, Cyp11a1and Hsd3b transcripts in cultured porcine granulosa cells(experiment 3)
In a separate set of experiments effects of AOH and AME onthe abundance of Cyp11a1 and Hsd3b transcripts, encoding keyenzymes of P4 synthesis, were monitored. Transcripts of the Gapdhhousekeeping gene were also measured and used for normalization.The relative abundance of Cyp11a1 and Hsd3b transcripts was notsignificantly different after treatment with AME and AOH (Fig. 4).AOH tended to slightly decrease the abundance of Gapdh transcriptssuggesting that the basal metabolic activity of the cells might havebeen affected, whereas AME but not AOH tended to affect Cyp11a1and even more pronounced Hsd3b transcripts.
3.3. Effects of AOH and AME on the expression of cytochromeP450scc, ˛-tubulin and actin proteins in cultured porcine
granulosa cellsIn experiment 4 it was investigated if the reduction of basal P4secretion caused by AOH or AME is due to the reduced abundance ofP450scc enzyme. In Western blot analyses P450scc is represented

142 U. Tiemann et al. / Toxicology Letters 186 (2009) 139–145
Fig. 3. Recovery of P4 synthesis after AOH or AME treatment. Cells were culturedfor 48 h in the presence of 10% fetal calf serum and for additional 24 h in serum- andphenol red-free medium without or with 0.8 �M, 1.6 �M, and 6.4 �M AOH (A) orAME (B). To estimate the recovery after treatment, the cells were cultured for fur-ther 24 h in the presence of the complete medium containing 10% fetal calf serum.Culture media were collected before (empty bars) or after recovery for P4 radioim-maff
bcPeto6tanwe
3s
n2nfLctaA
Fig. 4. Effects of AOH and AME on the abundance of Gapdh, Cyp11a1 and Hsd3btranscripts. The abundance of Gapdh transcripts was measured for normalizationand is expressed as number of copies per �g RNA (left abscissas, columns). Theabundance of Cyp11a1 and Hsd3b transcripts is expressed relative to Gapdh (rightabscissas, solid and broken lines). Cells were cultured for 48 h in the presence of 10%fetal calf serum and for additional 24 h in serum- and phenol red-free medium with
ring Alternaria strains significantly reduced cell viability. A dose-dependent inhibition was caused by extracts no. 444 (3.2–25 �g/ml,≈6.4–100 �M) and no. 445 (1.6–25 �g/ml, ≈12.5–100 �M),whereas extract no. 424 significantly decreased the cell viabilityonly at 25 �g/ml (≈100 �M, Fig. 6A). Fig. 6B indicates that extracts
Table 1Concentrations (�g/ml) of the Alternaria toxins TeA, AOH, AME, and ALT in extractsof Alternaria strains. One ml of these extracts quantified by HPLC was evaporated todryness, redissolved in DMSO and used as stock solution for the experiments.
Extract no. TeA (�g/ml) AOH (�g/ml) AME (�g/ml) ALT (�g/ml)
417 13,559 871 942 1634424 27,222 285 0.70 n.d.430 10 n.d. 3 n.d.444 2,654 2797 1,570 1045
unoassay (black bars). Values are relative to the respective untreated controls andre expressed as means ± S.E.M. of two independent granulosa cell preparations withour replicas each. Different letters denote values that were significantly differentrom respective controls (P < 0.05).
y a single band with a molecular mass of 45 kDa (Fig. 5A). At con-entrations of 0.8 �M and more AOH and AME the abundance of450scc was significantly reduced (Fig. 5A and B). The inhibitoryffect of AOH on P450scc protein abundance was not restored inhe recovery experiment (Fig. 5C and D). Also a marked reductionf the �-tubulin protein concentration was observed at 1.6 �M and.4 �M AOH but not AME (Fig. 5A and B). Also the concentrations ofwo additional housekeeping genes (Eisenberg and Levanon, 2003),ctin (Fig. 5A) and eukaryotic translation initiation factor 4A (EIF4a,ot shown) were clearly reduced in a similar way after treatmentith AOH. In recovery experiments the reduction �-tubulin, how-
ver, was largely restored (Fig. 5C and D).
.4. Identification of Alternaria toxins produced in vitro on riceubstratum
To analyze toxic effects of naturally occurring Alternaria toxins,ine different Alternaria strains isolated from cereals in 2006 and007 were grown on rice medium. Corresponding extracts wereumbered according to Alternaria strains of the phytopathogenic
ungi culture collection established at the Centre of Agricultural
andscape Research (Müncheberg, Germany). The major toxin washaracterized as TeA followed by AOH and AME in different quan-ities and relative portions to each other. With one exception,ll strains accumulated considerable amounts of all three majorlternaria toxins AOH, AME and TeA. Alternaria strain 430 producedor without different concentrations of AOH (A) or AME (B). Cells were processedfor transcript quantification as described. Data are means ± S.E.M. of three indepen-dent experiments. There are no significant differences between treatment groups(P > 0.05).
very low concentrations of AME and TeA, whereas AOH could not bedetected. Table 1 shows the various concentrations of AOH, AME,TeA, and ALT in the nine extracts. The concentration of ALT wasvery low or absent in most extracts, therefore, it was neglected asa reference substance in our study.
3.5. Effects of Alternaria toxin extracts on viability andprogesterone secretion in porcine granulosa cells (experiment 5)
Only three of the nine extracts derived from naturally occur-
445 3,000 300 1,300 n.d.480 13,691 96 191 11490 72,252 3009 1,408 n.d.494 58,133 1517 647 n.d.499 53,716 570 966 n.d.
n.d. < the limit of detection.
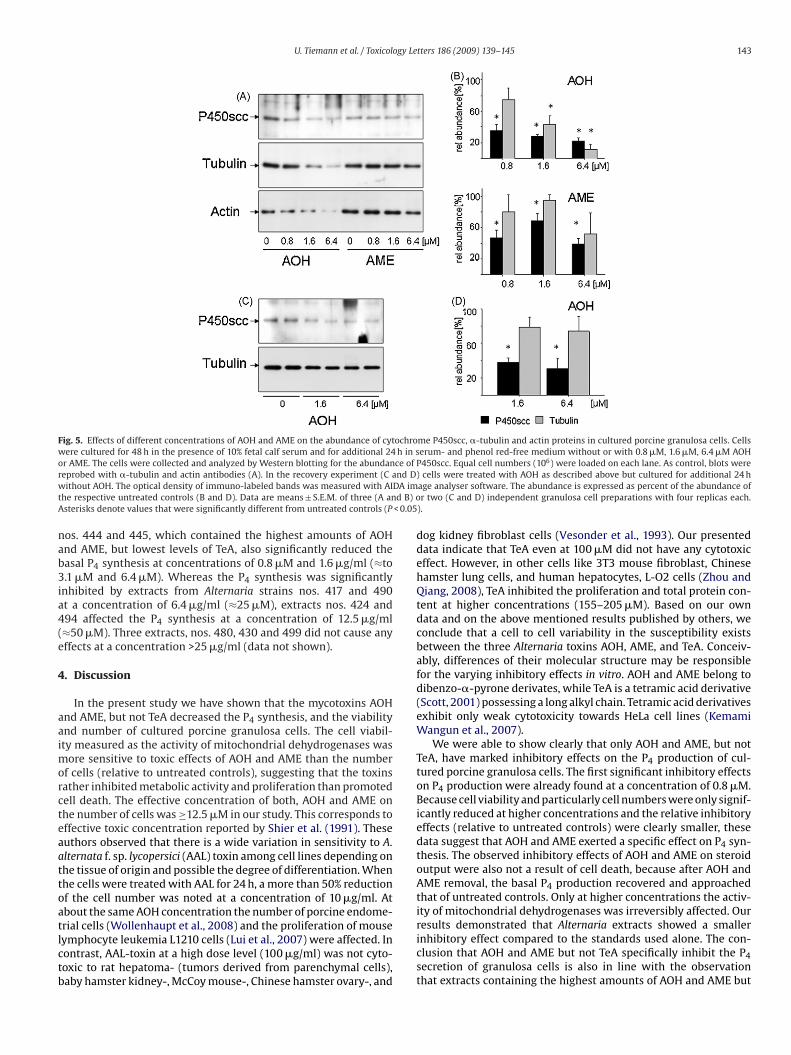
U. Tiemann et al. / Toxicology Letters 186 (2009) 139–145 143
Fig. 5. Effects of different concentrations of AOH and AME on the abundance of cytochrome P450scc, �-tubulin and actin proteins in cultured porcine granulosa cells. Cellswere cultured for 48 h in the presence of 10% fetal calf serum and for additional 24 h in serum- and phenol red-free medium without or with 0.8 �M, 1.6 �M, 6.4 �M AOHor AME. The cells were collected and analyzed by Western blotting for the abundance of P450scc. Equal cell numbers (106) were loaded on each lane. As control, blots werer nd D)w DA imt d B) oA < 0.05
nab3ia4(e
4
aaimorcteaattoatlctb
eprobed with �-tubulin and actin antibodies (A). In the recovery experiment (C aithout AOH. The optical density of immuno-labeled bands was measured with AI
he respective untreated controls (B and D). Data are means ± S.E.M. of three (A ansterisks denote values that were significantly different from untreated controls (P
os. 444 and 445, which contained the highest amounts of AOHnd AME, but lowest levels of TeA, also significantly reduced theasal P4 synthesis at concentrations of 0.8 �M and 1.6 �g/ml (≈to.1 �M and 6.4 �M). Whereas the P4 synthesis was significantly
nhibited by extracts from Alternaria strains nos. 417 and 490t a concentration of 6.4 �g/ml (≈25 �M), extracts nos. 424 and94 affected the P4 synthesis at a concentration of 12.5 �g/ml≈50 �M). Three extracts, nos. 480, 430 and 499 did not cause anyffects at a concentration >25 �g/ml (data not shown).
. Discussion
In the present study we have shown that the mycotoxins AOHnd AME, but not TeA decreased the P4 synthesis, and the viabilitynd number of cultured porcine granulosa cells. The cell viabil-ty measured as the activity of mitochondrial dehydrogenases was
ore sensitive to toxic effects of AOH and AME than the numberf cells (relative to untreated controls), suggesting that the toxinsather inhibited metabolic activity and proliferation than promotedell death. The effective concentration of both, AOH and AME onhe number of cells was ≥12.5 �M in our study. This corresponds toffective toxic concentration reported by Shier et al. (1991). Theseuthors observed that there is a wide variation in sensitivity to A.lternata f. sp. lycopersici (AAL) toxin among cell lines depending onhe tissue of origin and possible the degree of differentiation. Whenhe cells were treated with AAL for 24 h, a more than 50% reductionf the cell number was noted at a concentration of 10 �g/ml. Atbout the same AOH concentration the number of porcine endome-
rial cells (Wollenhaupt et al., 2008) and the proliferation of mouseymphocyte leukemia L1210 cells (Lui et al., 2007) were affected. Inontrast, AAL-toxin at a high dose level (100 �g/ml) was not cyto-oxic to rat hepatoma- (tumors derived from parenchymal cells),aby hamster kidney-, McCoy mouse-, Chinese hamster ovary-, andcells were treated with AOH as described above but cultured for additional 24 hage analyser software. The abundance is expressed as percent of the abundance ofr two (C and D) independent granulosa cell preparations with four replicas each.
).
dog kidney fibroblast cells (Vesonder et al., 1993). Our presenteddata indicate that TeA even at 100 �M did not have any cytotoxiceffect. However, in other cells like 3T3 mouse fibroblast, Chinesehamster lung cells, and human hepatocytes, L-O2 cells (Zhou andQiang, 2008), TeA inhibited the proliferation and total protein con-tent at higher concentrations (155–205 �M). Based on our owndata and on the above mentioned results published by others, weconclude that a cell to cell variability in the susceptibility existsbetween the three Alternaria toxins AOH, AME, and TeA. Conceiv-ably, differences of their molecular structure may be responsiblefor the varying inhibitory effects in vitro. AOH and AME belong todibenzo-�-pyrone derivates, while TeA is a tetramic acid derivative(Scott, 2001) possessing a long alkyl chain. Tetramic acid derivativesexhibit only weak cytotoxicity towards HeLa cell lines (KemamiWangun et al., 2007).
We were able to show clearly that only AOH and AME, but notTeA, have marked inhibitory effects on the P4 production of cul-tured porcine granulosa cells. The first significant inhibitory effectson P4 production were already found at a concentration of 0.8 �M.Because cell viability and particularly cell numbers were only signif-icantly reduced at higher concentrations and the relative inhibitoryeffects (relative to untreated controls) were clearly smaller, thesedata suggest that AOH and AME exerted a specific effect on P4 syn-thesis. The observed inhibitory effects of AOH and AME on steroidoutput were also not a result of cell death, because after AOH andAME removal, the basal P4 production recovered and approachedthat of untreated controls. Only at higher concentrations the activ-ity of mitochondrial dehydrogenases was irreversibly affected. Our
results demonstrated that Alternaria extracts showed a smallerinhibitory effect compared to the standards used alone. The con-clusion that AOH and AME but not TeA specifically inhibit the P4secretion of granulosa cells is also in line with the observationthat extracts containing the highest amounts of AOH and AME but
144 U. Tiemann et al. / Toxicology Le
Fig. 6. Effects of various concentrations of extracts (�g/ml) isolated from differentAlternaria strains on viability (A) and basal P4 secretion (B) of cultured porcine gran-ulosa cells. Cells were cultured for 48 h in the presence of 10% fetal calf serum and foradditional 24 h in serum- and phenol red-free medium without and with differentcaod
limeoteft
tttoHtmacus(iuir
oncentrations of extracts. Culture media were collected for P4 radioimmunoassaynd MTT assays were made with remaining attached cells. Data are means ± S.E.M.f three independent experiments. Asterisks denote values that were significantlyifferent from respective controls (P < 0.05).
owest concentration of TeA (nos. 444, 445) strongly and signif-cantly reduce P4 concentrations. Alternaria extracts are complex
ixtures consisting of several different mycotoxins. Attenuatingffects between them may explain the reduced inhibitory effectsf these complex mycotoxin mixtures on P4 synthesis. In contrasto our results however, King and Schade (1984) even described syn-rgistic effects between alternariol and its methyl ether and theyound relatively high toxicity of crude extracts or cultures comparedo pure standard substances.
In the present study, AOH and AME inhibited the basal P4 syn-hesis in a dose-dependent manner. Additionally, our data revealedhat particular in case of AOH, the inhibitory action on P4 produc-ion can be largely explained by adverse effects on the expressionf P450scc protein, whereas the abundance of Cyp11a1 and ofsd3b transcripts was not significantly reduced. This suggests
hat inhibition can be attributed to post-transcriptional regulatoryechanisms. The precise sites of action on steroidogenic enzymes
nd the mechanisms involved are not fully understood. However, itannot be excluded that AME and AOH also may influence reactionspstream from P4 synthesis. The synthesis of cholesterol precur-ors is catalyzed in the cytoplasm by hydroxy-methyl-glutaryl-CoA
HMG-CoA) reductase (Richards, 1994). Cholesterol transport to thenner mitochondrial membrane is mediated by steroid acute reg-latory (StAR) protein (Stocco and Clark, 1996). If these or othermportant steps of P4 synthesis were affected by AME and AOH,emains to be investigated.
tters 186 (2009) 139–145
Our study also indicated that AOH-mediated suppression of P4secretion was accompanied by a significantly reduced �-tubulinprotein abundance. For this AOH effect, its chemical structure mightbe responsible. In contrast to AME, AOH is a diphenolic compoundsimilar to bisphenol A. Lehmann and Metzler (2004) observed inbisphenol A-treated human fibroblasts structural abnormalities ofthe cytoplasmic microtubules. An optimal steroidogenic responseinvolves the depolymerization of microtubules and microfilamentswhich is important for the transport of steroidogenic substratesand intermediate products (Carnegie and Tsang, 1988). Thereforeincreased levels of AOH may inhibit tubulin polymerization result-ing in a depression of the synthesis of new tubulin monomersthus strengthening the assumption that AOH is an inhibitor ofmicrotubule assembly in cultured porcine granulosa cells. After theremoval of AOH the �-tubulin protein concentration was largelyrestored. This again suggests that AOH rather affects the conditionof cells than promotes cell death. The interference with the twoother so called housekeeping proteins, actin and EIF4a, may alsobe explained by the transiently altered physiological condition ofmycotoxin-treated cells.
In our study AOH and AME concentrations in the low �M rangewere necessary to exert significant inhibitory effects on culturedgranulosa cells. This raises the question if these results are relevantfor the in vivo situation. We assume that Alternaria concentrationstested in our in vitro study are in a similar range compared to toxincontents in feedstuff. In Germany, Poland and Australia levels ofup to 2.3 �M AOH, 1.5 �M AME and 0.5 �M TeA, respectively (seereview Müller et al., 2002; Scott, 2001) have been found in wheat.In weathered wheat from China, Li and Yoshizawa (2000) evendetected maximum levels of 1.3 �M AOH, 5.3 �M AME and 28.2 �MTeA.
In conclusion our data demonstrated that AOH and AME specif-ically inhibited P4 secretion of porcine ovarian cells in vitro andsuggestively may also have adverse effects in vivo. There are nospecific international regulations regarding Alternaria toxins in food(Scott, 2004). In view of the fact that granulosa cells directly influ-ence the metabolic and structural growth of the oocyte (Albertiniet al., 2001), exposure to AOH or AME may eventually affect repro-ductive performance by interfering with follicular developmentin swine and possibly other mammalian species. Feeding shouldtherefore be carefully controlled for the content of Alternaria toxins.
Conflict of interest
None.
Acknowledgements
We gratefully acknowledge the technical assistance of Mrs. P.Reckling, G. Krüger, S. Rodewald and M. Anders in Dummerstorf andMrs. Martina Peters and Grit von der Waydbrink in Müncheberg.
References
Albertini, D.F., Combelles, C.M., Benecchi, E., Carabatsos, M.J., 2001. Cellular basisfor paracrine regulation of ovarian follicle development. Reproduction 121,647–653.
Cameron, M.R., Caudle, M.R., Sullivan Jr., W.R., Peluso, J.J., Wimalasena, J., 1995. Thesteroidogenic and morphological effects of paclitaxel on cultured ovarian cancercells. Oncol. Res. 7, 145–156.
Chen, T.T., Maasey, P.J., Caudle, M.R., 1994. The inhibitory action of taxol on granulosacell steroidogenesis is reversible. Endocrinology 134, 2178–2183.
Carnegie, J.A., Tsang, B.K., 1988. The cytoskeleton and rat granulosa cell steroidoge-nesis: possible involvement of microtubules and microfilaments. Biol. Reprod.
38, 100–108.De Lucca, A.J., 2007. Harmful fungi in both agriculture and medicine. Rev. Iberoam.Micol. 24, 3–13.
Denkova, R., Ivanov, I., Dimitrova, M., 1992. Microtubules and regulation of granu-losa cell steroidogenesis by porcine granulosa cell conditioned medium. Endocr.Regul. 26, 195–199.

ogy Le
E
E
F
H
K
K
L
L
L
L
L
M
M
R
S
S
U. Tiemann et al. / Toxicol
isenberg, E., Levanon, E.Y., 2003. Human housekeeping genes are compact. TrendsGenet. 19, 362–365.
llis, M.B., 1971. Dematiaceous Hyphomycetes. Commonwealth Mycological Insti-tute, Kew, Surrey, UK, 608 pp.
ink-Gremmels, J., 1999. Mycotoxins: their implications for human and animalhealth. Vet. Q. 21, 115–120.
adley, M.E., 1995. Hormones and female reproductive physiology. In: Hadley, M.E.(Ed.), Endocrinology. Prentice Hall, Upper Saddle River, NJ, pp. 476–504.
emami Wangun, H.V., Dahse, H.-M., Hertweck, Ch., 2007. Epicoccamides B–D, gly-cosylated tetramic acid derivatives from an Epicoccum sp. associated with thetree fungus Pholiota squarrosa. J. Nat. Prod. 70, 1800–1803.
ing Jr., A.D., Schade, J.E., 1984. Alternaria toxins and their importance in food. J. FoodProtect. 47, 886–901.
ahav, M., Garmey, J.C., Veldhuis, J.D., 1996. Paradoxical effect of 3-isobutyl-1-methylxanthine on cytochrome P450 cholesterol side-chain cleavage mRNAaccumulation in porcine granulosa cells. Mol. Cell. Endocrinol. 117, 203–210.
ehmann, L., Metzler, M., 2004. Bisphenol A and its methylated congeners inhibitgrowth and interfere with microtubules in human fibroblasts in vitro. Chem.Biol. Interact. 147, 273–285.
ehmann, L., Wagner, J., Metzler, M., 2006. Estrogenic and clastogenic potential ofthe mycotoxin alternariol in cultured mammalian cells. Food Chem. Toxicol. 44,398–408.
i, F., Yoshizawa, T., 2000. Alternaria mycotoxins in weathered wheat from China. J.Agric. Food Chem. 48, 2920–2924.
ui, Z.-Z., Zhu, J., Sun, B., Lui, S., Geng, S., Lui, X., Li, Ch.-L., 2007. Alternol inhibitsproliferation and induces apoptosis in mouse lymphocyte leukemia (L1210) cells.Mol. Cell. Biochem. 306, 115–122.
üller, M., 1991. Untersuchungen zum Alternaria Befall von Silomais und Heu. Zen-tralbl. Mikrobiol. 146, 481–488.
üller, M., von der Waydbrink, G., Peters, M., Umann, K., Seyfarth, W., 2002. Belas-tung von Winterweizen mit Alternaria-Mykotoxinen im Land Brandenburg.Mycotoxin Res. 18A, 217–220.
ichards, J.S., 1994. Hormonal control of gene expression in the ovary. Endocr. Rev.15, 725–751.
chade, J.E., King Jr., A.D., 1984. Analysis of the major Alternaria toxins. J. Food Protect.47, 978–995.
cott, P.M., 2001. Analysis of agricultural commodities and foods for Alternaria Myco-toxins. J. AOAC Int. 84, 1809–1817.
tters 186 (2009) 139–145 145
Scott, P.M., 2004. Other mycotoxins. In: Magan, N., Olsen, M. (Eds.), Mycotoxinsin Food: Detection and Control. Woodhead Publishing Limited, Cambridge, pp.406–440.
Scott, P.M., Lawrence, G.A., Lau, B.P., 2006. Analysis of wines, grape juices and cran-berry juices for Alternaria toxins. Mycotoxin Res. 22, 142–147.
Schneider, F., Bellmann, A., Becker, F., Bambang Poernomo, S., Rehfeldt, C., Nürnberg,G., Kanitz, W., 2002. Gonadotropin release in preovulatory heifers after GnRHanalogs measured by two types of immunoassays. Exp. Clin. Endocrinol. Diab.111, 235–244.
Shier, W.T., Abbas, H.K., Mirocha, C.J., 1991. Toxicity of the mycotoxins fumonisins B1and B2 and Alternaria alternata f. sp. lycopersici toxin (AAL) in cultured mam-malian cells. Mycopathologia 116, 97–104.
Stocco, D.M., Clark, B.J., 1996. Role of the steroidogenic acute regulatory protein(StAR) in steroidogenesis. Biochem. Pharmacol. 51, 197–205.
Tiemann, U., Dänicke, S., 2007. In vivo and in vitro effects of the mycotoxins zear-alenone and deoxynivalenol on different non-reproductive and reproductiveorgans in female pigs. A review. Food Addit. Contam. 24, 306–314.
Tiemann, U., Schneider, F., Vanselow, J., Tomek, W., 2007. In vitro exposure of porcinegranulosa cells to the phytoestrogens genistein and daidze: effects on the biosyn-thesis of reproductive steroid hormones. Reprod. Toxicol. 24, 317–325.
Tiemann, U., Tomek, W., Schneider, F., Vanselow, J., 2003a. Effects of the mycotox-ins �- and �-zearalenol on regulation of progesterone synthesis in culturedgranulosa cells from porcine ovaries. Reprod. Toxicol. 17, 673–681.
Tiemann, U., Viergutz, T., Jonas, L., Schneider, F., 2003b. Influence of the mycotox-ins alpha- and beta-zearalenol and deoxynivalenol on the cell cycle of culturedporcine endometrial cells. Reprod. Toxicol. 17, 209–218.
Vesonder, R.F., Gasdorf, H., Peterson, R.E., 1993. Comparison of the cytotoxicities ofFusarium metabolites and Alternaria metabolite AAL-toxin to cultured mam-malian cell lines. Arch. Environ. Contam. Toxicol. 24, 473–477.
Visconti, A., Logrieco, A., Bottalico, A., 1986. Natural occurrence of Alternaria myco-toxins in olives—their production and possible transfer into the oil. Food Addit.Contam. 3, 323–330.
Weidenborner, M. (Ed.), 2001. Encyclopedia of Food Mycotoxins. Springer-Verlag,
Berlin Heidelberg, pp. 19–22.Wollenhaupt, K., Schneider, F., Tiemann, U., 2008. Influence of alternariol (AOH) onregulator proteins of cap dependent translation in porcine endometrial cells.Toxicol. Lett. 182, 57–62.
Zhou, B., Qiang, S., 2008. Environmental, genetic and cellular toxicity of tenuazonicacid isolated from Alternaria alternata. Afr. J. Biotechnol. 7, 1151–1156.





























