Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1984 by The American Society of Biological Chemists, Inc
Vol , 259, No. 10, Issue of May 25, pp. 6564-6570 1984 Printed in C.S.A.
The Nuclear-coded Subunits of Yeast Cytochrome c Oxidase I. FRACTIONATION OF THE HOLOENZYME INTO CHEMICALLY PURE POLYPEPTIDES AND THE
IDENTIFICATION OF TWO NEW SUBUNITS USING SOLVENT EXTRACTION AND REVERSED PHASE HIGH PERFORMANCE LIQUID CHROMATOGRAPHY*
(Received for publication, January 26, 1984)
Scott D. Power$, Michael A. LochrieO, Kevin A. Sevarino, Thomas E. Patterson, and Robert 0. PoytonlI From the Department of Molecular, Cellular and Developmental Biology, University of Colorado, Boulder, Colorado 80309
Previously, cytochrome c oxidase from the yeast Sac- charomyces cerevisiae has been thought to be com- posed of seven different polypeptide subunits. Four of these are small polypeptides (4,000-15,000 daltons), subunits IV-VII, which are encoded by nuclear DNA. Studies described here reveal the presence of two new polypeptides in this size range. These polypeptides, designated as subunits VIIa and VIII, co-migrate with subunit VI1 (R. 0. Poyton and G . Schatz (1975) J. Biol. Chem. 250, 762-761) on low resolution sodium dode- cy1 sulfate (SDS) polyacrylamide gels, can be partially resolved on high resolution SDS polyacrylamide gels, and can be completely separated from one another by reversed phase high performance liquid chromatogra- phy.
In order to determine the sequences of each of these six nuclear-coded polypeptides (subunits IV, V, VI, VII, VIIa, and VIII), we have developed new methods for the large scale purification of the holoenzyme and have employed a new strategy for the isolation of each poly- peptide. By using octyl-Sepharose chromatography to isolate holocytochrome c oxidase and by extracting the holoenzyme with aprotic organic solvents and fraction- ating these extracts by reversed phase high perform- ance liquid chromatography, it is possible to isolate several milligrams of each of these subunits. Each sub- unit preparation gives a single peak during reversed phase high performance liquid chromatography, a sin- gle band during SDS-polyacrylamide gel electropho- resis, a single NHderminal sequence, and a unique amino acid composition and tryptic peptide map. Since each purified subunit preparation gives close to a 100% yield of its NH,-terminal amino acid during quantita- tive Edman degradation, we conclude that no subunit has a blocked NH, terminus and that no subunit prep- aration contains either blocked or unblocked contami- nating polypeptides. Thus, each consists of a single unique polypeptide species.
Together, these results demonstrate that yeast cyto- chrome c oxidase contains six, rather than four, small subunit polypeptides. Thus, it appears that these poly-
~
* This study was supported by Grant GM 29838 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
National Institutes of Health Postdoctoral Fellow, 1979-1982. Present address, Genencor, Inc., South San Francisco, CA 94080.
§ Present address, Division of Biology, California Institute of Tech- nology, Pasadena, CA 91125.
IT Established Investigator of the American Heart Association. To whom correspondence should be addressed.
peptides, in combination with the three polypeptides encoded by mitochondrial DNA, constitute a holoen- zyme which contains nine subunits, instead of seven as proposed earlier.
During the past 10 years, cytochrome c oxidase has emerged as a useful system for studying mitochondrial electron trans- port and energy conservation (1-3), the assembly of oligo- meric membrane proteins (4-6), and those interactions be- tween mitochondrial and nuclear genomes which are required for mitochondrial biogenesis (7 ) . While most of the structural and functional data for this enzyme have been derived from studies with bovine heart cytochrome c oxidase, much of the biosynthetic data have come from studies with cytochrome c oxidase from the yeast Saccharomyces cereuisiae. A large part of the rationale for using bovine heart cytochrome c oxidase for physicochemical studies lies in the ease with which it is isolated from bovine heart tissue (3, 8). As a highly oxidative tissue, heart muscle is rich in cytochrome c oxidase. Although S. cereuisiae contains low levels of cytochrome c oxidase, relative to bovine heart, it has been used extensively for biosynthetic and genetic studies because it has easily manip- ulatable mitochondrial and nuclear genetic systems and is easily grown and radiolabeled.
While there are a number of similarities between bovine heart and yeast cytochrome c oxidases, there are also some potentially important differences. Foremost among these is the difference in the apparent number of polypeptide subunits present in each enzyme (9, 10). Previously, yeast cytochrome c oxidase has been reported to have 7 polypeptide subunits (9, 11, 12) while bovine heart cytochrome c oxidase has been reported to have as many as 13 polypeptide subunits (10). The three largest polypeptides (subunits I, 11, and 111) in the enzyme from both organisms are encoded on mitochondrial DNA (7). The sequences of these three polypeptides from yeast are approximately 40% homologous to the sequences of the corresponding subunit polypeptides from bovine heart cytochrome c oxidase. Recent studies with bovine heart cy- tochrome c oxidase have led to the proposal that these three mitochondrially coded polypeptides are solely responsible for the catalytic functions of the holoenzyme ( 1 , l O ) . The remain- ing polypeptide subunits in each enzyme are encoded on nuclear DNA. In the absence of direct biochemical and genetic data, these polypeptides have yet to be assigned a function.
In order to address the function of the nuclear-coded sub- units of eucaryotic cytochrome c oxidases, we are studying these polypeptides from yeast cytochrome c oxidase. Our studies are directed along two lines. In the first, we are using
6564
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase 6565
mutants’ (13) and isolated structural genes (14).’ These ge- netic studies will ultimately allow for the determination of whether these polypeptides are required for enzyme function and the identification of those regions in each polypeptide which are essential. In the second, we are attempting to determine the sequences for each of these polypeptides in order to identify homologous subunits in other eucaryotic cytochrome c oxidases (e.g. bovine heart) for which sequence data are already available (15-21). A comparison of sequences in homologous subunits from different organisms may allow us to identify invariant regions of each subunit polypeptide. To determine the sequences of the nuclear-coded subunits of yeast cytochrome c oxidase, we have had to develop a method for their isolation which provides large quantities of each at a high level of purity. Although methods for the isolation of these polypeptides have been developed previously (12, 22, 23), these are generally small scale and, in some cases, result in subunit preparations which are mixtures of Polypeptides? Therefore, we have developed a new approach to subunit purification. This approach makes use of our previous finding that various organic solvents are capable of removing different subunits from holocytochrome c oxidase (24). In studies re- ported here, we have used these solvent extractions in con- junction with HPLC4 to develop a preparative scale procedure for the rapid isolation of each of the nuclear-coded polypep- tides.
In addition to providing several milligrams of each polypep- tide, this new isolation strategy has revealed the presence of two new, hitherto unrecognized, polypeptide subunits. In the following papers (25, 26), we provide complete or partial sequences for each of these six nuclear-coded polypeptides and use these sequences to identify homologous subunits in bovine heart and Neurospora crassa cytochrome c oxidases.
MATERIALS AND METHODS‘
RESULTS
Large Scale Purification of Yeast Cytochrome c Oxidase A necessary prerequisite for the sequence analysis of the
nuclear-coded subunits of yeast cytochrome c oxidase is the availability of large quantities (several hundred milligrams) of the holoenzyme as starting material. Thus, the first goal of this study was to develop a procedure which would permit us to purifiy 200- to 500-mg batches of holocytochrome c oxidase from commercial bakers’ yeast.
The large scale purification procedure we have developed is diagrammed in Fig. 1A. This procedure involves three steps: first, the extraction of holocytochrome c oxidase from sub-
’ J. McEwen, C. KO, V. Cameron, B. Kloeckener-Gruissem, and R. 0. Poyton, manuscript in preparation.
R. Wright, C. KO, M. Cumsky, and R. 0. Poyton, manuscript in preparation.
C. George-Nascimento and R. 0. Poyton, unpublished observa- tions.
The abbreviations used are: HPLC, high performance liquid chro- matography; SDS, sodium dodecyl sulfate, SDS-PAGE, SDS polyac- rylamide gel electrophoresis; DTT, dithiothreitol; DMSO, dimethyl sulfoxide.
Portions of this paper (including “Materials and Methods” and Figs. 1-6) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Doc- ument No. f”-261, cite the authors, and include a check or money order for $4.40 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
mitochondrial particles with potassium cholate; second, the fractionation of the submitochondrial particle extract with ammonium sulfate; and third, the purification of holocyto- chrome c oxidase by hydrophobic chromatography on octyl- Sepharose. The major differences between this procedure and those developed previously for the purification of yeast cyto- chrome c oxidase (11, 12, 23, 34) are the use of fewer ammo- nium sulfate fractionation steps and the replacement of DEAE-cellulose, hydroxylapatite, or detergent exchange chro- matography by octyl-Sepharose chromatography as the final step in the procedure. The advantages afforded by the use of octyl-Sepharose chromatography as the final step in the pu- rification procedure are its specificity for hydrophobic pro- teins and its high binding capacity. These properties have allowed us to scale up the isolation of yeast cytochrome c oxidase without sacrificing yield or purity. The effectiveness of this step can be seen from SDS-PAGE of the various fractions obtained during the preparation of the enzyme (Fig. 1B). It is of interest that most of the protein in the fraction (111) applied to the column either does not bind or binds only weakly and, therefore, elutes in fractions A and B. Neither of these fractions contains any detectable heme a. Fractions C, D, and E contain low levels of protein and have a relatively simple polypeptide composition (Fig. 1B). The elution of fractions C, D, and E prior to the elution of cytochrome c oxidase (fraction IV) with ME buffer, 3% Triton X-100 is essential because otherwise the polypeptides found in those fractions contaminate the cytochrome c oxidase fraction. The cytochrome c oxidase eluted from the octyl-Sepharose column has a heme a to protein ratio which is routinely between 8 and 9 nmol of heme a per mg of protein and a polypeptide composition (Fig. 1B) which is identical with that reported previously (11, 12, 23). The “low resolution” SDS-PAGE system used in Fig. 1B displays 7 bands. As discussed below, the fastest migrating band, previously called subunit “VII,” consists of three polypeptides, subunits VII, VIIa, and VIII. These can be partially resolved on the “high resolution” SDS- PAGE system (Fig. 5 ) described under “Materials and Meth- ods.” The subunit “VII” band is split into two bands: one containing two polypeptide subunits, labeled VII, VIIa with apparent molecular weights of 4500, and another containing one polypeptide, labeled VIII, with an apparent molecular weight of 3000. These three polypeptides are considered in more detail below.
Cytochrome c oxidase purified by the procedure shown in Fig. 1 has approximately the same heme a to protein ratio (Table I) as that of other yeast holocytochrome c oxidase preparations (11, 12, 23, 34). However, the enzyme loses a substantial portion of its enzymatic activity during octyl- Sepharose chromatography. This loss occurs during elution with ME buffer, 2 M urea. Enzyme prepared in the absence of this step has a specific activity (K(min” mg-’ X lo-’)) of approximately 10.5, a value which is comparable to those reported for previous preparations of yeast cytochrome c oxidase (11, 34). Despite the loss of activity which results from urea treatment, this step is included in the large scale purification procedure because it is very effective in removing residual Tween 20 which is bound to the resin and to the holoenzyme and because it facilitates the subsequent elution of the holoenzyme in a small volume of ME buffer, 3% Triton X-100. The enzyme thus obtained needs no further concen- tration. Since our primary goal was to purify large amounts of the subunits of holocytochrome c oxidase for amino acid sequence determinations, since activity of the enzyme per se was not important for these studies, and since the urea elution step facilitates subsequent handling of the enzyme during its
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

6566 The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase TABLE I
Purification of yeast cytochrome c oxidase by octyl-Sephurose chromatography
Fraction Total protein Total activity Total heme a Heme a/protein Specific activity Purification Yield (heme a) (heme a)
ng K (min”) X IO-’ nmol nmoilmg nml)-l x 10-2 4 - 1 x 10-2 KJheme a (min Klmgpmtein (min
I I1
7673 7071 34.80 5767
I11 3128 21.75 5090 IV
35.15 5936 0.77 5.92 1 100 4.58 6.03 4.92 1.06 97 4.27
245 0.97 2097 8.56 0.463 3.78 6.95 2.12 86
11.12 35
0.82 1.63
large scale purification, the use of the ME buffer-urea step is justified. It should be emphasized, however, that this step is not absolutely essential and is frequently omitted when we purify small amounts (10-100 pg) of the holoenzyme for biosynthetic studies.
Isolation of Subunits IV, V, VI, VII, VIIa, and VIII Our strategy for the large scale purification of the nuclear-
coded subunits of yeast cytochrome c oxidase has been to prefractionate the enzyme into subunit pools using solvent mixtures and then to purify the released polypeptides by reversed phase HPLC (Fig. 2A).
Prefractionation of the Holoenzyme In order to purify subunits IV, V, VI, VII, VIIa, and VI11
by reversed phase HPLC, we first remove them from the oligomer away from the hydrophobic, mitochondrially coded subunits (I, 11, and 111). To accomplish this, we have avoided the use of SDS for dissociating the enzyme into its subunits and have, instead, developed a prefractionation regimen which selectively releases pools of subunits from the holoenzyme. As we have described previously (24), neutral and acidic aqueous solvent mixtures of acetonitrile and propanol are particularly effective at disrupting the holoenzyme and selec- tively solubilizing its component polypeptides. When holocy- tochrome c oxidase is first treated with 40% acetonitrile at neutral pH and subsequently with 50% mixed solvent (ace- tonitri1e:l-propanol, 1:l) at acid pH (0.625% trifluoroacetic acid, 0.625% triethylamine, pH 2.7), two pools of subunits are released (Fig. 2B); the first extract (&) contains subunits IV and VI whereas the second (S57) contains subunit V and a heterogeneous mixture of polypeptides which co-migrate in the subunit “VII” region of low resolution SDS gels (see below). From Fig. 2 B it is clear that the extraction proceeds nearly quantitatively. Since we have found that polypeptide subunits tend to extract optimally from the holoenzyme over a very narrow solvent concentration range (24) and that the polypeptide subunits tend to partition between the enzyme core and the supernatant, we use multiple, low volume ex- tractions, as described under “Materials and Methods,” to maximize yield and avoid excessive dilution. To disaggregate any polypeptide complexes which may be present, both pools are treated with guanidine HC1 under conditions known to dksociate the holoenzyme into nonaggregated subunits (12, 23) prior to their fractionation by reversed phase high per- formance liquid chromatography as outlined below.
Isolation of Subunits IV and VI After treatment with 6 M guanidine HCI and 10 mM dithio-
threitol, the s46 fraction is easily resolved into its two subunit components by reversed phase HPLC (Fig. 3) . This separation employs acetonitrile gradient elution of a pBondapak C18 column in the buffer system: 0.05% triethylamine, 0.05% trifluoroacetic acid, pH 2.7. Denaturation in the presence of reducing agent is required to prevent the formation of subunit
IV and VI homo- and heterodimers which result from cystei- nyl oxidation (24). As seen from Fig. 3, the resolution of the S4, fraction into subunits IV and VI is easily accomplished at both low sample load (0.02 mg of extract) and high sample load (2.5 mg of extract).
Isolation of Subunits V, VIIa, VU, and VIZI Yeast holocytochrome c oxidase has been thought previ-
ously to be composed of seven subunits (11,12,23). However, as revealed by reversed phase HPLC, subunit “VII” appears to be not one, but three unique polypeptides. Through correct choice of the acetonitri1e:propanol ratio in the gradient eluent (l:l, see Ref. 24) during reversed phase chromatography, the S57 fraction (Fig. 2B) can be resolved into six peaks (Fig. 4A). Five of these consist of unique proteins (Fig. 4B) when examined by high resolution SDS-PAGE. The first peak corresponds to the lightly staining band which migrates slightly faster than the “VII” band on SDS-PAGE and has been designated subunit VIII. The second and third peaks correspond to residual subunit VI, which remains after the first extraction, and to subunit V, respectively. The polypep- tides in peaks 4 and 6 have the same apparent molecular weights when analyzed on a variety of SDS-PAGE systems. Despite these identical electrophoretic mobilities, they have been designated VIIa and VII, respectively, because they can be distinguished from one another by their amino acid com- positions, tryptic peptides, and sequences (see below and Ref. 26).
Subunit Yields The yield of each subunit prepared by the above procedure
exceeds 40%, for subunit VII, and approaches 95%, for sub- units IV, VI, and VIII.
Characterization of Purified Subunits IV, V, VI, VII, VIIa, and VIIZ
Amino Acid Compositional Analysis The amino acid composition for each of the subunit prep-
arations is shown in Table 11. For subunits IV, V, and VI, there is good agreement with previously published values (12, 22, 23). However, the compositions of VIIa, VII, and VI11 do not agree with the composition determined previously for subunit “VII” (23). This difference results from the fact that previous subunit “VII?’ preparations were mixtures of subunits VIIa, VII, and VIII. Of special note is the absence of methio- nine in subunits VI1 and VI11 and the lack of cysteine in subunits VIIa and VII. Although these three subunits exhibit different amino acid compositions, they are remarkably sim- ilar in average hydrophobicity (mole per cent Ala, Ile, Leu, Val, Phe, Cys, and Met (36)). With hydrophobicities of 46.5% (VII), 41.4% (VIIa), and 45.9% (VIII), they are more hydro- phobic than the other three nuclear-encoded subunits of the holoenzyme (IV, V, and VI) which have hydrophobicities which range from 32% to 39%. The hydrophobicities of these
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from
The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase 6567
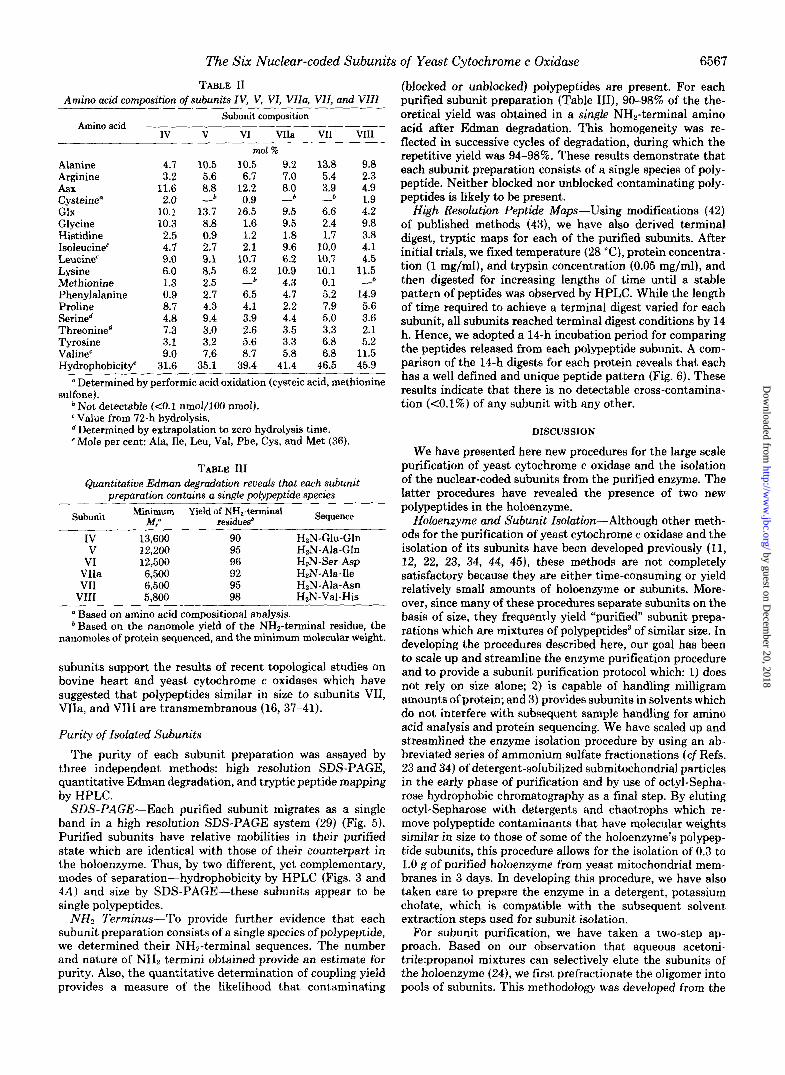
TABLE I1 Amino acid commsitwn of subunits IV. V. VI. Vlla. Vll. and Vlll
Amino acid Subunit composition
IV V VI VIIa VI1 VI11
Alanine Arginine Asx Cysteine" Glx Glycine Histidine Isoleucine' Leucine" Lysine Methionine Phenylalanine Proline Serined Threonined Tyrosine Valinec Hvdroohobicit4
4.1 10.5 3.2 5.6
11.6 8.8 2.0 -b
10.1 13.7 10.3 8.8 2.5 0.9 4.1 2.7 9.0 9.1 6.0 8.5 1.3 2.5 0.9 2.7 8.7 4.3 4.8 9.4 7.3 3.0 3.1 3.2 9.0 7.6
31.6 35.1
mol % 10.5 9.2 13,8 9.8 6.7 7.0 5.4 2.3
12.2 8.0 3.9 4.9 0.9 - b - 1.9
16.5 9.5 6.6 4.2 1.6 9.5 2.4 9.8 1.2 1.8 1.7 3.8 2.1 9.6 10.0 4.1
10.7 6.2 10.7 4.5 6.2 10.9 10.1 11.: - 4.3 0.1 - 6.5 4.7 5.2 14.9 4.1 2.2 7.9 5.6 3.9 4.4 5.0 3.6 2.6 3.5 3.3 2.1 5.6 3.3 6.8 5.2 8.7 5.8 6.8 11.5
39.4 41.4 46.5 45.9
Determined by performic acid oxidation (cysteic acid, methionine
Not detectable ( d . 1 nmo1/100 nmol).
Determined by extrapolation to zero hydrolysis time.
sulfone).
e Value from 72-h hydrolysis.
'Mole per cent: Ala, Ile, Leu, Val, Phe, Cys, and Met (36).
TABLE 111 Quantitative Edman degradation reveals that each subunit
meoaration contains a single polypeptide species
Subunit Minimum Yield of NH,-terminal M/ residuesb
Sequence
IV 13,600 90 H2N-Glu-Gln V 12,200 95 HZN-Ala-Gln
VI 12,500 96 HzN-Ser-Asp VIIa 6,500 92 H2N-Ala-Ile VI1 6,500 95
VI11 HzN-Ala-Asn
5,800 98 HzN-Val-His Based on amino acid compositional analysis.
bBased on the nanomole yield of the NH2-terminal residue, the nanomoles of protein sequenced, and the minimum molecular weight.
subunits support the results of recent topological studies on bovine heart and yeast cytochrome c oxidases which have suggested that polypeptides similar in size to subunits VII, VIIa, and VI11 are transmembranous (16, 37-41).
Purity of Isolated Subunits The purity of each subunit preparation was assayed by
three independent methods: high resolution SDS-PAGE, quantitative Edman degradation, and tryptic peptide mapping by HPLC.
SDS-PAGE-Each purified subunit migrates as a single band in a high resolution SDS-PAGE system (29) (Fig. 5 ) . Purified subunits have relative mobilities in their purified state which are identical with those of their counterpart in the holoenzyme. Thus, by two different, yet complementary, modes of separation-hydrophobicity by HPLC (Figs. 3 and 4A) and size by SDS-PAGE-these subunits appear to be single polypeptides.
NH2 Terminus-To provide further evidence that each subunit preparation consists of a single species of polypeptide, we determined their NH,-terminal sequences. The number and nature of NH2 termini obtained provide an estimate for purity. Also, the quantitative determination of coupling yield provides a measure of the likelihood that contaminating
(blocked or unblocked) polypeptides are present. For each purified subunit preparation (Table III), 90-98% of the the- oretical yield was obtained in a single NH2-terminal amino acid after Edman degradation. This homogeneity was re- flected in successive cycles of degradation, during which the repetitive yield was 94-98%. These results demonstrate that each subunit preparation consists of a single species of poly- peptide. Neither blocked nor unblocked contaminating poly- peptides is likely to be present.
High Resolution Peptide Maps-Using modifications (42) of published methods (43), we have also derived terminal digest, tryptic maps for each of the purified subunits. After initial trials, we fixed temperature (28 "C), protein concentra- tion (1 mg/ml), and trypsin concentration (0.05 mg/ml), and then digested for increasing lengths of time until a stable pattern of peptides was observed by HPLC. While the length of time required to achieve a terminal digest varied for each subunit, all subunits reached terminal digest conditions by 14 h. Hence, we adopted a 14-h incubation period for comparing the peptides released from each polypeptide subunit. A com- parison of the 14-h digests for each protein reveals that each has a well defined and unique peptide pattern (Fig. 6). These results indicate that there is no detectable cross-contamina- tion (<0.1%) of any subunit with any other.
DISCUSSION
We have presented here new procedures for the large scale purification of yeast cytochrome c oxidase and the isolation of the nuclear-coded subunits from the purified enzyme. The latter procedures have revealed the presence of two new polypeptides in the holoenzyme.
Holoenzyme and Subunit Isolation-Although other meth- ods for the purification of yeast cytochrome c oxidase and the isolation of its subunits have been developed previously (11, 12, 22, 23, 34, 44, 45), these methods are not completely satisfactory because they are either time-consuming or yield relatively small amounts of holoenzyme or subunits. More- over, since many of these procedures separate subunits on the basis of size, they frequently yield "purified" subunit prepa- rations which are mixtures of polypeptides3 of similar size. In developing the procedures described here, our goal has been to scale up and streamline the enzyme purification procedure and to provide a subunit purification protocol which: 1) does not rely on size alone; 2) is capable of handling milligram amounts of protein; and 3) provides subunits in solvents which do not interfere with subsequent sample handling for amino acid analysis and protein sequencing. We have scaled up and streamlined the enzyme isolation procedure by using an ab- breviated series of ammonium sulfate fractionations ( c f Refs. 23 and 34) of detergent-solubilized submitochondrial particles in the early phase of purification and by use of octyl-Sepha- rose hydrophobic chromatography as a final step. By eluting octyl-Sepharose with detergents and chaotrophs which re- move polypeptide contaminants that have molecular weights similar in size to those of some of the holoenzyme's polypep- tide subunits, this procedure allows for the isolation of 0.3 to 1.0 g of purified holoenzyme from yeast mitochondrial mem- branes in 3 days. In developing this procedure, we have also taken care to prepare the enzyme in a detergent, potassium cholate, which is compatible with the subsequent solvent extraction steps used for subunit isolation.
For subunit purification, we have taken a two-step ap- proach. Based on our observation that aqueous acetoni- tri1e:propanol mixtures can selectively elute the subunits of the holoenzyme (24), we first prefractionate the oligomer into pools of subunits. This methodology was developed from the
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

6568 The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase
premise that those conditions which are required to elute polypeptides from a hydrophobic oligomeric membrane pro- tein, like cytochrome c oxidase, are similar to those which are required to elute proteins from the bonded C18 phase during reversed phase chromatography. This is reflected in our use of similar solvents for both extraction of subunits from the holoenzyme and their separation by reversed phase HPLC. It is important that the extracted pools be free of the more hydrophob~c subunits (I, 11, and 111) of the holoenzyme be- cause attempts to isolate subunits IV, V, VI, VII, VIIa, and VI11 without first removing them from the holoenzyme were unsuccessful; presumably because subunits I, 11, and I11 irre- versibly adsorb to the C18 matrix and interfere with the fractionation of subunits IV, V, VII, VII, VIIa, and VIII. The combination of these two methods results in the complete separation of all six subunit polypeptides into fractions which are essentially pure. Indeed, these subunit preparations con- tain only one polypeptide species as judged by the following observations: 1) each purified subunit preparation elutes from a reversed phase C18 column as a single peak and runs as a single band on SDS-PAGE gels of several different polyac- rylamide concentrations; 2 ) only one NHz-terminal sequence can be detected in each subunit preparation; and 3) yields from the first Edman degradation reaction for each subunit preparation are quantitative, approaching the yield expected for 100% recovery. The latter two observations demonstrate clearly that none of these subunit polypeptides has blocked NH2 termini and that there is only one type of polypeptide present in each preparation. In view of these observations and the fact that the procedures described here yield milligram amounts of each subunit polypeptide in solvents which are easily removed because they are volatile, it is now possible to initiate sequencing studies on each of these polypeptides.
Subunit Composition-Historically, the subunit composi- tion of eucaryotic cytochrome E oxidases has been linked to the resolving power of polyacrylamide gels during SDS-gel electrophoresis (10, 15, 29, 46). A tendency toward simplifi- cation has led many workers to fit subunit compositions to the seven-subunit model originally proposed by Poyton and Schatz (12) for the yeast enzyme and by Downer et al. (47) for the beef heart enzyme. Subsequently, however, high reso- lution electrophoretic methods (46), in combination with NHz-terminal sequencing studies (15, 48), have revealed a much more complex picture. Where once cytochrome c oxi- dase from bovine heart was thought to be a 7-subunit oligo- mer, it now appears to be a 13-subunit oligomer. Recent evidence has led us to question the conclusion (12) that yeast cytochrome E oxidase is composed of seven subunits. First, high resolution SDS-PAGE has revealed a lightly staining band (VIII) which migrates faster than the subunit "VII" band in the holoenzyme (see Fig. 5 and Refs. 10 and 49). Second, our attempts to sequence subunit "VII" isolated by gel filtration in SDS (23) yielded multiple NH2 termini.3 Clear evidence for heterogeneity in the subunit "VII" band is dem- onstrated in this communication through the use of reversed phase chromatography. This mode of fractionation, based upon relative hydrophobicity, has resolved the "VII" popula- tion into its three components-subunits VIIa, VII, and VIII. Each of these polypeptides, together with subunits IV, V, and VI, has been shown to have different amino acid compositions, tryptic peptides, and NH2-terminal sequences. Thus, it ap- pears that there are six different nuclear-coded polypeptides, instead of four, in yeast cytochrome c oxidase. Studies directed at the stoichiometry and function of these polypeptides are currently in progress.
Ackmwkdgment-We wish to thank AI Smith at the University of California (Davis) for his assistance in all phases of amino acid analysis and protein sequencing.
REFERENCES
1. Wikstrom, M., Krab, K., and Saraste, M. (1981) Cytochrome Oxidose,
2. Racker. E. (1976) A New Look at Mechanism in Bioenereetics. Academic Academic Press, New York
Press, New York - ,
3. Lemberg, R. (1961) Adu. Enzyml. 23,265-321 4. Poyton, R. O., Sevarino, K., George-Nascimento, C., and Power, S. D.
(1980) Ann. N. Y. Acad. Sct. 343,275-290 5. Poyton, R. O., Bellus, G., and Kerner, A. (1982) in Membranes and
Transport (Martonosi, A., ed) Vol. 1, pp. 237-242, Plenum Publishing
6. Poyton, R. 0. (1983) in The Spatsal Organization of Eukaryotic Cells Co., New York
(McIntosh, J. R., ed) pp. 15-72, Alan R. Liss Publishers, New York 7. Poyton, R. 0. (1980) Curr. To Celt. Regul. 7,231-295 8. Hartzeli, C. R., Beinert, H., %an Gelder, B. F., and King, T. E. (1978)
9. Poyton, R. O., McKemmie, E., and George-Nascimento, C. (1978) J. Biol. Methods Enzymol. 53,54-66
Chem. 253,6303-6306 10. Kadenbach, B., and Merle, P. (1981) FEBS Lett. 136.1-11 11. Rubin, M. S., and Tzagoloff, A. (1973) J. Biol. Chem. 248,4269-4274 12. Poyton, R. O., and Schatz, G. (1975) J. Bioi. Chem. 250,752-761 13. McEwen, J., Cumsky, M., KO, C., Power, S. D., and Poyton, R. 0. (1984)
14. Cumsky, M. G., McEwen, J. E., KO, C., and Poyton, R. 0. (1983) J. Bwl.
15. Buse, G. (1980) in Biological Chemistry of Organelle Formation (Bucher, T.,
16. Buse, G., and Steffans, G. J. (1978) Hoppe-Seyler's 2. Physiol. Chem. 3 5 9 ,
J. Cell Biochem. 24, in press
Chem. 2 5 8 , 13418-13421
Sebald, W., and Weiss, H., eds) pp. 59-70, Springer-Verlag, New York
I nn6-1 IKIQ 17. Tanaka, M., Haniu, M., Yasunobu, K. T., Yu, C.-A., Yu, L., Wei, Y.-H.,
18. Biewald, R., and Buse, G. (1982) Hopp-Seyler's Z. Physiol. Chem. 363,
*""" s"""
and King, T. E. (1979) J. BioE. Chem. 254,3879-3885
1141-1153 19. Tanaka,"., Yasunobu, K. T., Wei, Y.-H., and King, T. E. (1981) J. Biol.
Chem. 256,4832-4837 20. Steffens, G. C. M., Steffens, G. J., and Buse, G. (1979) Hoppe-Seyler's Z.
Physiol. Chem. 360,1641-1650 21. Steffens, G . J., and Buse, G. (1979) Hoppe-Seyler's Z. Physiol. Chem. 360,
613419 22. Tzagoloff, A., Akai, A., and Rubin, M. S. (1974) in The Biogenesis of
Mitochondria (Kroon, A. M., and Saccone, C., eds) pp. 405-421, Academic
23. George-Nascimento, C., and Poyton, R. 0. (1981) J. Biol. Chem. 256, Press, New York
24. Power, S. D., Lochrie, M. A,, and Poyton, R. 0. (1983) J. Chromatogr. 266, 9363-9370
5115-5911
25. Power, S. D., Lochrie, M. A., Patterson, T. E., and Poyton, R. 0. (1984) J.
26. Power, S. D., Lochrie, M. A,, and Poyton, R. 0. (1984) J. Biol. Chem. 259,
""" """
Biol. Chem. 259,6571-6574
6575-6578 27. Rosen, S. (1978) Biochim. Biophys. Acta 523,314-320
29. Merle, P., and Kadenbach, B. (1980) Eur. J. Biochem. 105,499-507 28. Cahral, F., and Schatz, G. (1978) Methods Enzymol. 56,314-320
30. Hin, C. H. W. (1967) Methods Enzymol. 11,59-62 31. Edrnan. P.. and Bess. G. 11967) Eur. J. Biochem. 1.80-91 32. Kulbe, K. 16. (1974~"Al.'hioc~m. 59,564-573 ' 33. Pisano, J. J., and Bronzert, T. J. (1969) J. BioZ. Chem. 244,5597-5607 34. Mason. T. L.. Povton. R. 0.. Wharton. D. C.. and Schatz. G. (19731 J. Bioi.
he& 248,1345-i354 . .
35. Bradford, M. M. (1976) Anal. Biochem. 72,248-254 36. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157 , 105-132 37. Ludwig, B., Downer, N. W., and Capaldi, R. A. (1979) Biochemistry 18,
38. Prochaska, L., Bisson, R., and Capafdi, R. A. (1980) Biochemistry 19,3174-
39. Bisson, R, Montecucco, C., Gutweniger, H., and Azzi, A. (1979) J. Biol.
40. Gutweniger, H., Bisson, R., and Montecucco, C. (1981) Biochim. Biophys.
41. Capaldi, R. A. (1982) Biochim. Biophys. Acta 694,291-306 42. Hermondson, M., and Mahoney, W. C. (1980) in Chemical Synthesis and
Sequencing of Peptides and Proteins (Lin, T. Y., Schechler, A. N., Hendrickson, R. L., and Condliffe, P. G., eds) pp 119-130, Elsevier-
43. Fullmer, C . S., and Wasserman, R. H. (1979) J. Biol. Chem 254, 7208- North Holland, Amsterdam
44. Shakespeare, P. G., and Mahler, H. R. (1971) J. Biol. Chem. 2 4 6 , 7649- 7212
45. Eytan, G. D., and Schatz, G. (1975) J. Bwl. Chem. 250,767-774 7655
46. Kadenbach, B., Jarausch, J., Hartmann, R., and Merle, P. (1983) Anal.
47. Downer, N. W., Robinson, N. C., and Capaldi, R. A. (1976) Biochemistry
48. Kadenbach, B., Hartmann, R., Glanville, R., and Buse, G. (1982) FEBS
49. Ludwig, B., and Schatz, G . (1984) Proc. Natl. Acad. Sci. U. S. A. 77, 196-
1401-1407
3179
Chem. 254,9962-9965
Acta 635,187-193
Biochem. 129,517-521
15,2930-2936
Lett. 138,236-238
200
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase 6569
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

6570 The Six Nuclear-coded Subunits of Yeast Cytochrome c Oxidase A
A
. I , r.. :
1 5
""" - B
A E C D E F G H
Vllc, VI1 Vlll
....
11
..._
i i . , . : i i i r ...... - .....
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from

S D Power, M A Lochrie, K A Sevarino, T E Patterson and R O Poytonchromatography.
subunits using solvent extraction and reversed phase high performance liquidholoenzyme into chemically pure polypeptides and the identification of two new
The nuclear-coded subunits of yeast cytochrome c oxidase. I. Fractionation of the
1984, 259:6564-6570.J. Biol. Chem.
http://www.jbc.org/content/259/10/6564Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/259/10/6564.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on Decem
ber 20, 2018http://w
ww
.jbc.org/D
ownloaded from





























