Embed Size (px)
Citation preview

0 1994 by The American Society for Biochemistry and Molecular Biology, Inc THE JOURNAL OF BIOLOGICAL CHEhllSTRY Vol. 269, No. 39, Issue of September 30, pp. 2448&24485, 1994
Printed in U.S.A.
The POXl Gene Encoding Peroxisomal Acyl-CoA Oxidase in Saccharomyces cereuisiae Is under the Control of Multiple Regulatory Elements*
(Received for publication, June 3, 1994, and in revised form, July 21, 1994)
Tongwen Wang$§, Yi Luo§, and Gillian M. Small1 From the Department of Cell Biology and Anatomy, Mount Sinai School of Medicine, New York, New York 10029
Transcription of POX1, the gene encoding peroxiso- mal acyl-CoA oxidase in the yeast Saccharomyces cerevi- siae, is controlled by the carbon source given for cell growth. The gene is repressed in glucose, derepressed in glycerol, and induced in oleate. This regulation is con- trolled by cis-acting elements in the 5' region of the gene, which bind regulatory proteins. By deletion anal- ysis and DNA band shift assays, we have characterized three such elements in the POXl upstream sequence, two upstream repression sequences and an upstream activating sequence. Each was able to regulate the tran- scription of a heterologous gene construct iso-l-cyto- chrome-c (CYCl)/ZacZ.
The biosynthesis of peroxisomes and regulation of peroxiso- mal enzymes is highly regulated in both mammals and yeast. In mammals, peroxisome proliferation is induced by adminis- tration of various compounds that include hypolipidemic drugs, plasticizers, and pesticides (1, 2). In the yeast Saccharomyces cereuisiae, peroxisome number is regulated in response to growth conditions; few peroxisomes are present when the yeast are provided glucose for growth, but many peroxisomes are induced when a fatty acid such as oleate is the sole carbon source (3). In both cases the peroxisomal P-oxidation enzymes are either repressed or induced; this regulation occurs at the transcriptional level (4, 5) .
Regulation of transcription occurs by the binding of regula- tory proteins to specific DNA sequences. These proteins can either activate or repress transcription by RNA polymerase at specific promoters. There is much evidence that major portions of the transcription apparatus are conserved among eukaryotes and that proteins involved in transcription, as well as promoter elements with which such proteins interact, are evolutionarily related between yeast and mammals (6, 7). Yeast upstream activating or repressing sequences (UASs' or URSs), like mam- malian upstream elements, function in both orientations and at variable distances from the transcription initiation sites (8). In order to understand the mechanisms involved in the cellular events leading to peroxisome proliferation and enzyme induc-
* This work was supported by American Heart Association Grant AHA 92-850. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Pediatric Surgical Lab., Massachusetts General Hospital, Boston, MA 02114.
1 American Heart Association Established Investigator. To whom cor- $ Contributed equally to this work.
respondence should be addressed. Tel.: 212-241-0981; Fax: 212-860- 1174. ' The abbreviations used are: UAS, upstream activating sequence;
URS, upstream repressing sequence; PCR, polymerase chain reaction; bp, base pairb).
tion, we initiated studies on the regulation of peroxisomal acyl- CoA oxidase in s. cereuisiae. It is likely that some of the factors involved in this process may be similar in yeast and mammals.
Acyl-CoA oxidase is the first and rate-limiting enzyme in the peroxisomal P-oxidation cycle. In S. cerevisiae, the acyl-CoA oxidase is a single copy gene (POX1) that has been cloned and sequenced (9). We previously described the characterization of one negative regulating element in the promoter region of this gene (10). Recent studies make it clear that multiple proteins may be associated with the functioning of relatively short cis- acting elements of yeast genes (ll), and multiple sites are com- monly found to be required for the regulation of gene expres- sion (12). Here we report further analysis of the POXl promoter and demonstrate that at least three elements and two different proteins are involved in the complex regulation of this gene.
MATERIALS AND METHODS Yeast Strains and Culture Conditions
S. cereuisiae strain BWG1-7a (MATa adel-100 his4519 leu2-112 ura3-52) was used throughout these studies.
Yeast were grown in rich (YPD) or minimal (SD) media, as required. YPD medium contains 2% yeast extract, 1% peptone, 2% glucose; SD medium contains 0.67% yeast nitrogen base without amino acids, 2% glucose. Induction media was either YPG, which contained 1% yeast extract, 2% peptone, 3% glycerol, or YPGO, which contained 1% yeast extract, 2% peptone, 3% glycerol, 0.1% oleic acid, and 0.2% Tween-40.
Plasmid Constructions pP13570, pP13571, and pP1353 have been described previously (10).
To obtain other promoter fragments of POXl in frame with lacZ, a PCR approach was taken. DNA primers were synthesized as shown in Table I.
pP13532"Primers 1 and 2 were used in the PCR reaction with pP13570 DNA. The resulting PCR fragment was treated with DNA polymerase I to render the ends blunt and was then ligated into pBlue- script I1 at the SmaI site. The fragment was then excised from this plasmid by digesting with PstI and HindIII and was subcloned into the PstIIHindIII sites ofYIp353 (13). This resulted in a plasmid containing 349 nucleotides 5' of the ATG and 602 coding bases of POXl fused in-frame with la&.
pP13533-pP13533 was prepared in the same manner as described above, but with primers 1 and 3 in the PCR.
pP13534"Primers 1 and 4 were used in the PCR, the resulting fragment was treated with DNA polymerase I, digested with HindIII, and subcloned into the SmaIIHindIII sites of Mp353.
pP13535-pP13535 was prepared in the same manner as pP13534, but with primers 1 and 5 in the PCR.
pNG plasmids were prepared by cloning fragments of the POXl promoter into pNG15 or pNG22 (14). pNG15 contains the CYCl pro- moter, without the CYCl upstream activation elements, fused in-frame with lacZ. pNG22 is the same construct except that the CYCl UAS elements are also present. The POXl fragments were cloned such that any regulatory elements present would control the expression of CYCU lacZ.
pNG1599-pNG1599 was prepared using primers 6 and 7 in a PCR with DNA from pP13570. The resulting 99-bp fragment was treated with T4 DNA polymerase to render the ends blunt and was cloned into
24480

Regulation of Acyl-CoA Oxidase in S . cereuisiae 24481
TABLE I DNA primers used for preparing constructs
Primer Sequence Region of POX1
1 5' CTGCAGCTGGGCAACATTGGA 3' 582 to 602 2 5' GGTACCGCGTTCAAACCCTGA 3' -349 to -335 3 5' GGTACCTTGGCGTTGTTAAGT 3' -213 to -199 4 5' GGTACCTATAAAGGTCTAACCT 3' -164 to -149 5 5' GGTACCGGTCTAACCTGTAA 3' -158 to -145 6 5' AACGGCTATTAGCCGCTAATGACCG 3' -275 to -251 7 5' GGTACCGCGTTCAAACCCTGA 3' -349 to -335 8" 5' GGGGTCGACGGTGATTTACCCCAACG 3' -238 to -254 9 5' GGGGTCGACTTTACGGTATTAGTTGA 3' -316 to -300 10 5' GGGCTCCGAAGCGAAAGGAAT 3' -294 to -277 11" 5' GGGTCTAGAGGTGATTTACCCCAACGG 3' -238 to -255 12 5' GGGGCTCGAGAGATCTCGACCAAAAAAA 3' -454 to -436 13 5' GGGGTCTAGAAAAAAAGGAGGTTTATAA 3' -350 to -368
., Note that primers 8 and 11 differ in the restriction site introduced at the 5' end.
the SmaI site of pBluescript I1 to create p n l . The fragment was then excised by digesting with XhoI and EagI and subcloned into the SalY EagI sites of pNG15.
pNG1556-pNG1556 was prepared as described above except that primers 10 and 11 were used in the PCR, and the 56-bp fragment was subcloned into pBluescript I1 to form pYL2.
pNG1580-pNG1580 was prepared using primers 8 and 9 in the PCR. The resulting 80-bp fragment was subcloned into pT7 Blue (Novagen) creating pYL3. The fragment was excised by digestion with SalI, and was subcloned into the SalI site of pNG15.
pNG2256-The XhoYEagI fragment was excised from p n 2 and sub- cloned into the SalYEagI sites of pNG22 to create pNG2256.
pNG2ZURSI-Primers 12 and 13 were used in the PCR to generate a 105-bp fragment that was digested with XhoI and XbaI and subcloned into pBluescript I1 a t homologous sites. An XhoI-EagI fragment, con- taining the 105-mer, was excised from this construct and was subcloned into pNG22 to create pNG22URS1.
pNG22URS2"TWo complementary oligonucleotides (5' GGCCG-
CCGAAATATAGGGCTTAAAATGTGTCAGGGTTTGAACGCC 3', con- sisting of bases -349 to -314 containing flanking EagI sites) were synthesized and annealed by heating to 65 "C and slowly cooling to room temperature. The annealed DNA was cloned into the EagI site of pNG22.All constructs were sequenced to verify their identity.
GCGTTCAAACCCTGACACATT'ITAAGCCCTATAWI'C 3' and 5' GG-
Dunsformation Standard protocols were used for transformation of Escherichia coli
and yeast (15). The POX1::lacZ fusion constructs were integrated in the URA3 locus. Selection for uracil prototrophs was carried out on SD medium supplemented with the appropriate amino acids.
Preparation of Yeast Extracts Yeast cells were grown to stationary phase in YPD medium and then
transferred to YPG, YPGO, or YPD and allowed to grow for another 18 h. Cells were harvested, washed, and resuspended in extraction buffer
EDTA, 10% glycerol, 7 mM 2-mercaptoethanol, 1 IIIM phenylmethylsul- fonyl fluoride). The cells were disrupted by vortexing in the presence of glass beads (0.5 m~ diameter), and cell debris was removed by centrifu- gation at 10,000 x g for 1 h. Cell extracts were divided into aliquots and stored at -80 "C.
(200 mM Tris-HC1, pH 8.0, 400 mM (NH,),SO,, 10 mM MgCl,, 1 IIIM
Assays P-galactosidase was measured as described previously (10) with the
exception that the specific activity was calculated according to the Worthinggton Manual. Protein concentrations were determined by the method of Bradford (16), using y-globulin as standard.
Gel Retardation Assays Gel retardation assays were performed as described by Fried and
Crothers (171, with some modifications (18). Cell extracts (approxi- mately 30 pg) were mixed with 1 p1 of 32P-end labeled DNA fragment (approximately 5000 cpm) in a volume of 20 pl of binding buffer (12 mM Hepes pH 7.5, 60 mM KCl, 5 m~ MgCl,, 4 mM Tris-HC1, pH 8.0, 0.6 mM dithiothreitol, 10% glycerol, 0.26 pg/pl poly(d1-dC), 0.3 pg/pl bovine serum albumin). The binding reactions were incubated at room tem- perature for 20 min, and protein-DNA complexes were resolved on 5% polyacrylamide gels at 150 V. For competition experiments, 30-60 mo-
TABLE I1 DNA fragments used in band shift assays
Fragment identification Region of POXl
184-mer -349 to -165 105-mer -454 to -349 99-mer -349 to -251 80-mer -316 to -238 70-mer -349 to -279
uRs2 29-mer -279 to -251
-349 to -314
lar excess of unlabeled competitor DNA relative to probe DNA were added to the cell extract 10 min before the addition of the probe. The DNA fragments used for band shift assays are shown in Table 11 (also see Fig. 2).
The 184-mer was obtained by PCR as described previously (IO), the resulting DNA fragment was cloned into pBluescript I1 and was subse- quently excised by digesting the plasmid with BamHI and HindIII. The 99- and 80-mer were excised from pYLl and pYL3, respectively, by digesting with BamHI and HindIII. The 70- and 29-mers were obtained by digesting the 99-mer with EcoRI, and PL and PR (see Fig. 2) were obtained by digesting the 80-mer with EcoRI. The fragments were la- beled with [a-32PldATP using Klenow fragment of DNA polymerase I.
All oligonucleotides used in this study were synthesized on an Applied Biosystems DNA synthesizer.
RESULTS
Identification of Further Regulatory Elements in the POXl Promoter-Our previous studies had identified a negative regu- lating element in the 5"flanking region of POXl (-434 to -410) that is involved in controlling the transcription of this gene (10). We have continued our analysis of the POXl promoter region to determine other sequences that are involved in the complex regulation of acyl-CoA oxidase in S. cerevisiae. We had previously prepared a fusion construct between the 5' region of POXl and the lac2 gene from E. coli. We have now prepared a series of deletion constructs in order to determine other regu- latory elements in this promoter (Fig. 1). All constructs were introduced into S. cerevisiae, and @-galactosidase activity was measured in extracts from the cells grown in different media. The production of @-galactosidase by the cells expressing these plasmids showed a complex pattern (Fig. 1).
In order to characterize the URS at -434 to -410, we previ- ously introduced point mutations into this sequence (construct P1353) (10). Such mutations caused an increase in @-galacto- sidase activity in the cells when grown in galactose or oleate media, confirming the functionality of this URS as a negative regulating element. Deletion of the region containing this URS plus some extra sequence (-454 to -349, ~13532) led to an additional increase of @-galactosidase activity in cells grown in each media. This result suggests that either the point mutation introduced into the U R S region did not completely abolish its function or that there is another URS element between -410 and -349. Alternatively, deletion of this 61-bp region may have interfered with the functioning of a putative URS. Further deletions from -349 to -213 caused a general increase of @-ga- lactosidase activity in glucose-grown cells and a decreased ac- tivity in oleate-grown cells, suggesting that this region contains a glucose-responsive element and an oleate-activating element. Cells transformed with pP13534, which contained the proximal promoter TATAA box (-164 to -1601, but no other regulatory elements, had @-galactosidase activities that were similar in all media tested, indicating that the TATAA box alone is sufficient for basal expression of POX1. Deletion of the TATAA box (P13535) resulted in the abolishment of POXl transcription (Fig. 1). The possibilities discussed above were further exam- ined by carrying out DNA band shift analysis of this region.
Specific Protein-DNA Interactions Correlate with the Regu- lation of POXl Expression-Our deletion analysis of the 5'
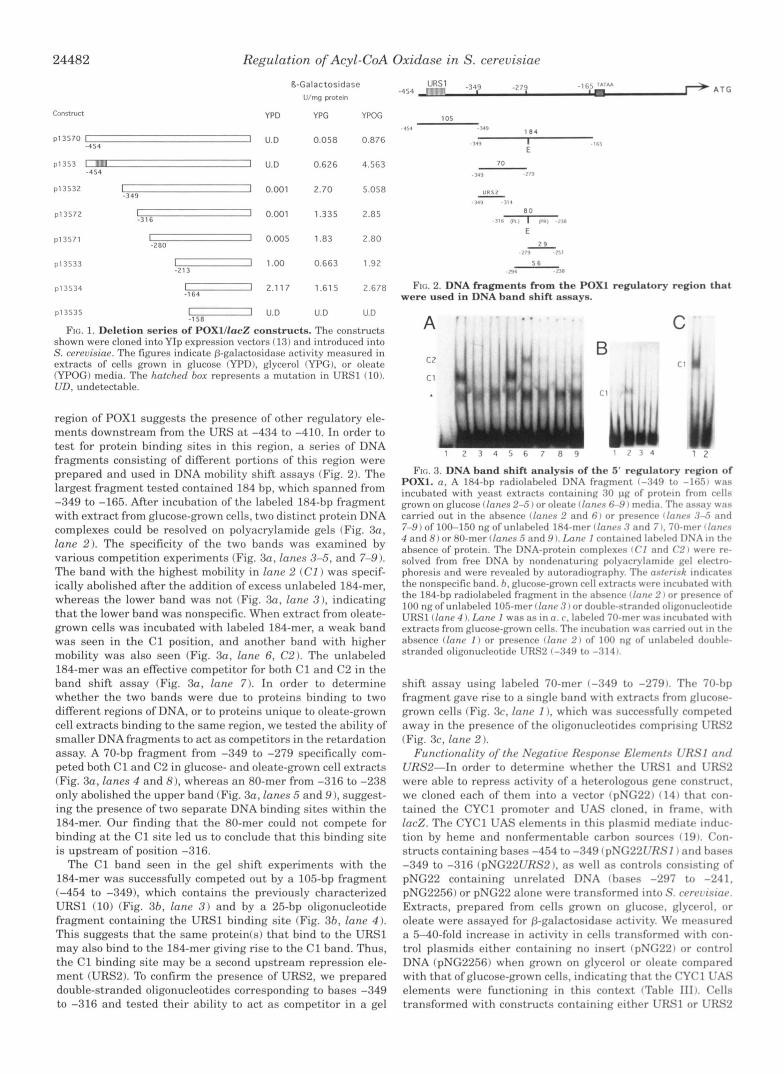
24482 Regulation of Acyl-CoA Oxidase in S. cerevisiae
Consttnrct
p13570 I 1 - 4 5 4
p13532 I I -349
~ 1 3 5 7 2 7 P I 3 5 7 1 - ~ 1 3 5 3 3 - 1113534 - ~ 1 3 5 3 5
R-Galactosidase V/mg potein
YPD YPG YPOG
U.D 0.058 0.876
U.D 0.626 4.563
0.001 2.70 5.058
0.001 1.335 2.85
0.005 1.83 2.80
1.00 0.663 1.92
2.117 1.615 2.678
U.D U.D U.D
FIG. 1. Deletion series of POXl/lacZ constructs. The constructs shown were cloned into YIP expression vectors ( 13) and introduced into S. cereuisinr. The figures indicate [+galactosidase activity measured in extracts of cells grown in glucose (YPD), glycerol IYPG), or oleate (YPOG) media. The hntrhrd ho.r represents :I mutation in URSl (10). UTI, undetectnhle.
region of POXl suggests the presence of other regulatory ele- ments downstream from the URS at -434 to -410. In order to test for protein binding sites in this region, a series of DNA fragments consisting of different portions of this region were prepared and used in DNA mobility shift assays (Fig. 2). The largest fragment tested contained 184 bp, which spanned from -349 to -165. After incubation of the labeled 184-bp fragment with extract from glucose-grown cells, two distinct protein DNA complexes could be resolved on polyacrylamide gels (Fig. 3a, lane 2). The specificity of the two bands was examined by various competition experiments (Fig. 3a, lanes 3-5, and 7-9). The band with the highest mobility in lane 2 (CI ) was specif- ically abolished after the addition of excess unlabeled 184-mer, whereas the lower band was not (Fig. 3a, lane 3 ) . indicating tha t t he lower band was nonspecific. When extract from oleate- grown cells was incubated with labeled 184-mer, a weak band was seen in the C1 position, and another band with higher mobility was also seen (Fig. 30, lane 6, C 2 ) . The unlaheled 184-mer was an effective competitor for both C1 and C2 in the band shift assay (Fig. 3a, lane 7) . In order to determine whether the two bands were due to proteins binding to two different regions of DNA, or to proteins unique to oleate-grown cell extracts binding to the same region, we tested the ability of smaller DNA fragments to act as competitors in the retardation assay. A 70-bp fragment from -349 to -279 specifically com- peted both C1 and C2 in glucose- and oleate-grown cell extracts (Fig. 3a, lanes 4 and 8), whereas an 80-mer from -316 to -238 only abolished the upper band (Fig. 3a, lanes 5 and 9 ), suggest- ing the presence of two separate DNA binding sites within the 184-mer. Our finding that the 80-mer could not compete for binding at the C1 site led us to conclude that this binding site is upstream of position -316.
The C1 band seen in the gel shift experiments with the 184-mer was successfully competed out by a 105-bp fragment (-454 to -349), which contains the previously characterized URSl (10) (Fig. 3h, lane 3 ) and by a 25-bp oligonucleotide fragment containing the URSl binding site (Fig. 3h, lane 4 ) . This suggests that the same protein(s) that bind to the URSl may also bind to the 184-mer giving rise to the C1 band. Thus, the C1 binding site may be a second upstream repression ele- ment (URS2). To confirm the presence of URS2, we prepared double-stranded oligonucleotides corresponding to bases -349 to -316 and tested their ability to act as competitor in a gel
8 0 116 ii,, 1 . Y ..I1
E
- 111 . T . I
2 9
n a 5 6
.'I"
FIG. 2. DNA fragmentn from the POXl regulatory region that were used in DNA hand shift assays.
A
CZ
c1
C E
FIG. 3. DNA band shift analysis of the 5' regulatory region of POXl. n. A 184-hp rnr l iolalwld I)NA f r a ~ m r n t 1 - : < 4 9 to - I I ; . - I I w a s incuhated with y w s t rxtracts containing :<I) pg of protrin from crlls grown on glucose clnnrs 2-5 I or olcntr rlnrtrs 6-9 I mrdin. Thc assay was carried out in the ahsence fIonc*s 2 and 6 1 or prrwnccx i l n r r r - s :l;i nnrl 7-9) of 100-150 ng of unlaheletl 184-rner f Innrs 3 and 7 I. 70-mrr 1lrrrrr.s 4 and 8) or 80-mer llnnrs 6 nntl 9 I. IAnr I containrd Iahrlrd D N A in t h r absence of protein. The DNA-protein complexrs ( ( ' I and ('2 I w r r r rv- solved from free DNA hy nondenaturing polyacrylnrnidr ~ r l rlrctro- phoresis and were revealed hy autoracliogr;lphy. Thr nstvrisf: intlic:rtw the nonspecific hand. h, glucose-grown crll extracts wrrv inruhntrd wl th the 184-hp radiolaheled fragment in the nhscnce rlrrnr 2 I nr p r r s r n w of 100 ng of unlahrled 105-mer I lnnr .'I I or douhlr-strandrd oliponuclrotidr UKSl (Ionr 4 1. IAnr I was :IS inn. r . lahrlrtl 70-rnrr was incuhntc*d w l t h extracts from glucose-grown cells. The incuhation was carrwd nut In t h v ahsence f lnnr I ) or presence (Innr 2 ) of 100 ng of unlahrlrd douhlv- stranded oligonucleotidr lJRS2 (-349 to -314 I.
shift assay using laheled 70-mer (-349 to -279). The 70-hp fragment gave rise to a single hand with extracts from glucosr- grown cells (Fig. 3c, fanr I ) , which was successfully compctcd away in the presence of the oligonucleotides comprising LJKS2 (Fig. 3c, lanr 2 ).
Functionality of the Nrgativr Rwponsr Elrmcnts U R S 1 nnrl URS2"In order to determine whether the URSl and URS2 were able to repress activity of a heterologous gene construct. we cloned each of them into a vector lpNG221 ( 14) that con- tained the CYCl promoter and UAS cloned, in frame. with la&. The CYCl UAS elements in this plasmid mediate induc- tion by heme and nonfermentable carhon sources (19). Con- structs containing bases -454 to -349 (pNG221JR.91 ) and bases -349 to -316 (pNG22URS2). as well as controls consisting of pNG22 containing unrelated DNA (bases -297 to -241, pNG22.56) or pNG22 alone were transformed into S. crrcrqisiac. Extracts, prepared from cells grown on glucose. glycerol, or oleate were assayed for P-galactosidase activity. We measured a 5-40-fold increase in activity in cells transformed with con- trol plasmids either containing no insert (pNG22) or control DNA fpNG22.56) when grown on glycerol or oleate comparcBd with that of glucose-grown cells, indicating that the CYCl UAS elements were functioning in this context (Table 1111. Cells transformed with constructs containing either URSl or URS2
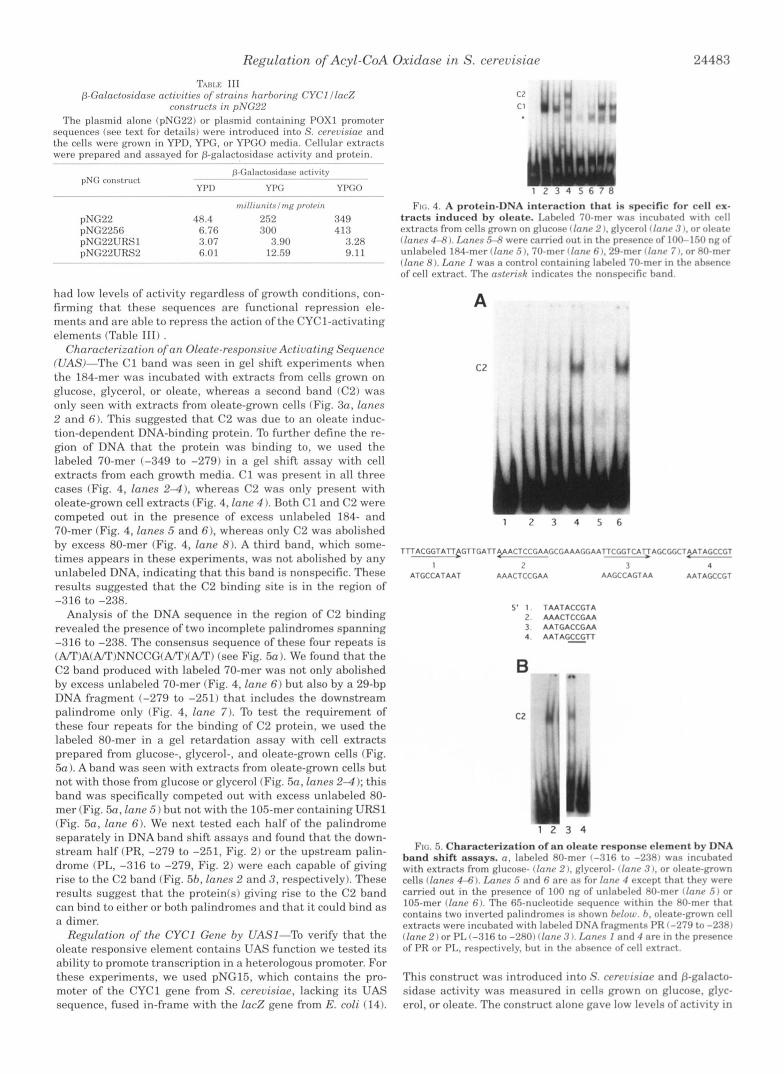
Regulation of Acyl-CoA Oxidase in S. cerevisiae 24483
TAIW 111 P-Golnctonirlosr nctioilirs of strains hnrhoring CYCl l lnc j !
constructs i n pNG22 The plasmid alone (pNG22) or plasmid containing POX1 promoter
sequences (see text for details) were introduced into S. crreoisine and the cells were grown in YPD, YPG, or YPGO media. Cellular extracts were prrpared and assnyrd for ~-galactosidasr activity and protein. ~~ ~ ~~~ ~~ -
p-Cnlnctosidaw activity
YPI) YPG YPCO
r n i l l i u n ~ f s / n ~ g pmlrin
pNG construrt ~ ~~~
~- -" ~ - ~~~ -
pNG22 48.4 252 349 pNG2256 6.76 300 4 18 pNG22URS1 3.07 8.90 3.28 pNG22URS2 6.01 12.59 9.1 1
~~ ~~ ~~ ~~~ ~ ~ ~
had low levels of activity regardless of growth conditions, con- firming that these sequences are functional repression ele- ments and are able to repress the action of the CYC1-activating elements (Table 111) .
Charactcrization ofan Oleate-responsive Activating Sequence WAS)-The C1 band was seen in gel shift experiments when the 184-mer was incubated with extracts from cells grown on glucose, glycerol, or oleate, whereas a second band (C2) was only seen with extracts from oleate-grown cells (Fig. 3n, lanes 2 and 6). This suggested that C2 was due to an oleate induc- tion-dependent DNA-binding protein. To further define the re- gion of DNA that the protein was binding to, we used the labeled 70-mer (-349 to -279) in a gel shift assay with cell extracts from each growth media. C1 was present in all three cases (Fig. 4, lanes 2-4). whereas C2 was only present with oleate-grown cell extracts (Fig. 4, lane 4 ). Both C1 and C2 were competed out in the presence of excess unlabeled 184- and 70-mer (Fig. 4, lanes 5 and 6). whereas only C2 was abolished by excess 80-mer (Fig. 4, lane 8). A third band, which some- times appears in these experiments, was not abolished by any unlabeled DNA, indicating that this band is nonspecific. These results suggested that the C2 binding site is in the region of
Analysis of the DNA sequence in the region of C2 binding revealed the presence of two incomplete palindromes spanning -316 to -238. The consensus sequence of these four repeats is (A/*r)A(A/T)NNCCG(A/T)(A/"I') (see Fig. 5a ). We found tha t the C2 band produced with labeled 70-mer was not only abolished by excess unlabeled 70-mer (Fig. 4, lane 6 ) but also by a 29-bp DNA fragment (-279 to -251) that includes the downstream palindrome only (Fig. 4, lane 7). To test the requirement of these four repeats for the binding of C2 protein, we used the labeled 80-mer in a gel retardation assay with cell extracts prepared from glucose-, glycerol-, and oleate-grown cells (Fig. 5n ). A band was seen with extracts from oleate-grown cells but not with those from glucose or glycerol (Fig. 5a, lanes 2 4 ); this band was specifically competed out with excess unlabeled 80- mer (Fig. 50, lane .5) but not with the 105-mer containing URSl (Fig. sa, lane 6). We next tested each half of the palindrome separately in DNA band shift assays and found that the down- stream half (PR, -279 to -251, Fig. 2 ) or the upstream palin- drome (PL, -316 to -279, Fig. 2) were each capable of giving rise to the C2 band (Fig. 5h, lanes 2 and 3 , respectively). These results suggest that the protein(s) giving rise to the C2 band can bind to either or both palindromes and that it could bind as a dimer.
Regulation of the CYCl Gene hy UASl-To verify tha t the oleate responsive element contains UAS function we tested its ability to promote transcription in a heterologous promoter. For these experiments, we used pNG15, which contains the pro- moter of the CYCl gene from S. cerevisine, lacking its UAS sequence, fused in-frame with the lac%: gene from E. coli (14).
-316 to -288.
C
C
Ftc:. 4. A protein-DNA intc-rartion t h n t is specific for cell cx- tracts induced hy oleate. 1 , : ~ t ~ v l d 70-tnI.r was ~ncuhatr t l w l t h crll extracts from cells LTown nn glucow 1 lrrnr 2 I , glycrrol 1 inn^ 3 I, or olratrn Ilnnrs 44l J. Ixznrs 54 wrre carricd out in the prrsrnce of 100-150 ng o f unlabeled 184-mer I lnnr .5 I , 70-mer (lane 6 1. 29-mrr clone 7 1, or 80-mrr ( fane 8). Lanr I was a control containing Iabrlwi 70-mrr in thr nbsrncr of cell extract. The nstcrisk indicate9 thr nonspecific hand.
A
c2
1 <' < 4 ', 6
T T T A C G G T A ~ ~ C T T G A T T ~ A C T C C G A A G C G A A A G G A A T T C ~ T C A ~ A G C G G C T ~ T A G C C G T
1 4 ATGCCATAAT
2 AAACTCCGAA
3 AAGCCACTAA AATACCCGT
5' 1 TAATACCCTA 2. AAACTCCGAA 3 AATGACCGAA 4. A A T A G S T T
i 1 3 4
FIG. 5. Characterization of an oleate renponse element by DNA hand shift assays. 0 , Iahelrtl XO-mrr (-316 to -23x1 wax tncuhatrd
cells ( fanes 4-6) . Ixznrs 5 and fi arr as for lnnr 4 rxcrpt that thry wrrr with extracts from glucose- ( lone 2 ), Rlycerol- (Innr .'{ I. or olratr-grown
c a m e d out in the presence of 100 ng of unlahrled XO-mer Ilnnr 5 ) or 105-mer (Innr fi). The 65-nucleotidr srqurncr within thr 80-mrr that contains two inverted palindromes is shown hrlotr. h, olrate-pown crl l extracts were incubated with lahrled DSA f r a p e n t n PK 1-27:) to -2W I (Inn@ 2 ) or PL (-316 to -280) ( I n n r .? ). Imncs 1 and 4 nrr in th r p rwrnce of PR or PL, resprctivr1.v. hut in t h r ahnrnrr of rrll extract.
This construct was introduced into S. cc.rrl,isiae and p-galacto- sidase activity was measurrd in cells grown on glucose, glyc- erol, or oleate. The construct alone gavr low Irvels of activity in

24484 Regulation of Acyl-CoA Oxidase in 5'. cerevisiae TABLE IV
P-Galactosidase activities of strains harboring CYCl / lac2 constructs in pNGl5
Cells were prepared for Table 111.
P-Galactosidase activity
YPD W G YPGO
milliunitslmg protein
pNG construct
pNG15 0.375 0.623 0.611 pNG1556 pNG1580 pNG1599
0.269 0.211 15.62
8.93 61.1 130
0.738 0.967 33.63
all conditions tested (Table IV). When a 56-bp fragment from the POXl promoter region that contained one of the two pal- indromes (-294 to -238) was inserted in front of the CYCl/ZacZ gene fusion (pNG1556), the activity was increased by 100-fold in oleate-grown cells. The 80-mer encompassing bp -316 to -238, which includes the complete putative UAS (pNG1580), gave a 200-fold increase over background under these condi- tions. The activity was reduced to 50-fold above background in pNG1599, which contains the complete UAS, but also URS2.
DISCUSSION
The experiments described above detail our continued anal- ysis of the promoter region of the POXl gene. These studies indicate that at least three cis-acting elements mediate the regulation of acyl-CoA oxidase in S. cerevisiae. All of these putative elements were found in a region at least 200 bp up- stream of the POXl transcribed region, and are clustered in a 182-bp region. The results shown in Fig. 1 indicate that the transcription of POXl is repressed when cells are grown in glucose, derepressed in glycerol, and highly induced when a long chain fatty acid such as oleate is supplied as the carbon source. A similar regulation has been shown for the gene en- coding peroxisomal 3-oxoacyl-CoA thiolase in S. cerevisiae (201, indicating that this is a general mechanism acting to regulate peroxisomal P-oxidation in this yeast. We have shown that this regulation of POXl is controlled by the presence of two up- stream repression sequences (URN and URS2) and by a UAS. URSl consists of a direct inverted repeat AGGGTWI')(A/")(AI T)(APT)(A/T), whereas URSS consists of a direct repeat
AGGG^l"(APT)N(AIT), which is separated by seven nucleo- tides. Our previous studies on URSl demonstrated that one half of this repeat is sufficient for protein binding and that AGGG are critical nucleotides for protein binding to occur (10). Our finding that interactions between the trans-acting factor with URSS is competed out with an excess of URSl fragment suggests that the same protein is binding to these two repres- sion sequences (Fig. 3b). Furthermore, we demonstrated that both fragments are equally capable of repressing the action of the CYCl UAS (Table 111).
Transcription of the POX1-ZacZ fusion gene increased 5-10- fold in glycerol- and in oleate-grown cells when URSl was mutated, and a further increase was seen when it was com- pletely deleted (Fig. 1). However, when URSS was deleted (con- struct ~13572) the P-galactosidase activity of glucose-grown cells did not change (compare p13532 with ~13572 , Fig. 1). It is possible that a functional part of URS2 is already deleted in ~13532, this would also explain why the activities of ~ 1 3 5 3 2 are higher than p1353. It is also possible that repression mediated through URSS may require an interaction with other cis-ele- ments such as URSl that are not present in ~13532, but such elements may exist in the CYCl reporter construct as CYCl itself has an upstream URS. We also noted that when URS2 was deleted, the activity of oleate-grown cells was decreased. It is possible that in order to function optimally as an activating
C
sequence, the UAS may require adjacent cis-elements. The UAS sequence that we identified in POXl is similar to a
sequence present in several genes encoding peroxisomal pro- teins that has been termed the "P-oxidation box" (20). This was proposed to be the potential binding site for a trans-acting factor responsible for the oleate-specific induction. The posi- tions of a single palindrome consisting of two repeats were suggested in the 5' upstream sequences of the genes encoding the trifunctional enzyme and thiolase. Our finding of two pal- indromes consisting of four repeats in the POXl promoter may reflect the important regulatory role of this protein, the rate- limiting enzyme of the peroxisomal 6-oxidation pathway. This complex regulation of POXl bears some resemblance to the regulation of the GAL genes in S. cerevisiae. The regulation of these genes, which encode the enzymes responsible for convert- ing galactose to glucose 6-phosphate, has been studied exten- sively (21). The GAL4 protein is a trans-acting factor with a consensus binding site that is an inverted repeat of CGGA(G/ CIGAC, the most highly conserved and important bases appear to be the GGC (22). Some genes such as GALUGALlO have four such binding sites of which just two are required for gene ex- pression (21).
In our deletion studies, we observed that deletion of the UAS caused a decrease in activity in cells grown on oleate media, but we also observed an unexpected increase in activity of glucose grown cells (compare ~ 1 3 5 7 2 with ~13533 , Fig. 1). This sug- gests that the deleted region, -316 to -213, can mediate both oleate induction and glucose repression. I t is possible that there is another URS element in this region; however, we have no evidence for protein binding in our gel shift assays. An alternative possibility is that a protein, which may be present in low levels, could bind to the defined UAS in glucose-grown cells and cause glucose repression. In oleate-grown cells, a dif- ferent factor would bind to the same region and activate tran- scription by an "anti-repression'' mechanism. Thus, this regu- latory element may have a dual function. This theory is supported by our finding that the inserted 56-mer in the con- struct pNG2256, which contains part of the UAS, caused re- pression in glucose-grown cells (compare activities of pNG22 with pNG2256, Table 111).
Our next goal is to characterize the proteins that are respon- sible for this regulation. Genes implicated in the regulation of peroxisomal proteins thus far include ADRl (231, SNF1, and SNF4 (24). It has been suggested that these genes affect per- oxisome biogenesis via separate signal pathways, and it is likely that many more genes are involved in these pathways. Experiments are underway in our laboratory to isolate genes responsible for the regulation of POX1.
Acknowledgments-We thank Murl Casey for technical assistance and for help in preparing the figures for this manuscript.
REFERENCES
2. Reddy, J. K., and Krishnakantha, T. P. (1975) Science 190, 787-789 1. Rao, M. S., and Reddy, J. K. (1987) Carcinogenesis 8 , 6 3 1 4 3 6
3. Veenhuis, M., Mateblowski, M., Kunau, W. H., and Harder, W. (1987) East 3,
4. Reddy, J. K., Goel, S. K., Nemali, M. R., Carrino, J. J., Laffler, T. G.. Reddy, M. 77-84
K, Sperbeck, S. J., Osumi, T., Hashimoto, T., Lalwani, N. D., and Rao, M.
5. Skoneczny, M., Chelstowska, A., and Rytka, J. (1988) Eur. J . Biochem. 174,
6. Guarente, L. (1987) Annu. Rev. Genet. 2 1 , 4 2 5 4 5 2 7. Chodosh, L. A., Olesen, J., Hahn, S., Baldwin, A. S., Guarente, L., and Sharp,
8. Guarente, L., Yocum, R., and Gifford, P. (1982) Proc. Natl. Acad. Sci. U. S. A.
S. (1986) P ~ o c . Natl. Acad. Sei. U. S. A. 88, 1747-1751
297-302
P. A. (1988) Cell 63, 25-35
9. Dmochowska, A,, Dignard, D., Maleszka, R., and Thomas, D. Y. (1990) Gene 79,7410-7414
10. Wang, T. W., Lewin, A. S., and Small, G. M. (1992) Nucleic Acids Res. 20,
12. Yoo, H. S., and Cooper, T. G. (1989) Mol. Cell. Biol. 9, 32314243 11. Olesen, J., Hahn, S., and Guarente, L. (1987) Cell 61, 953-961
(Amst. ) 88, 247-252
3495-3500

Regulation of Acyl-CoA Oxidase in S. cerevisiae 24485
13. Myers, A. M., Tzagoloff, A., Kinney, D. M., and Lusty, C. J. (1986) Gene (Amst. ) 45,299-310
14. Kovari, L., Sumrada, R., Kovari, I., and Cooper, T. G. (1990) Mol. Cell. Biol. 10, F , O R ~ - F , ~ W
15. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
. .~
16. Bradford, M. (1976) Anal. Biochem. 72,248-254
18. Ausubel, E M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, 17. Fried, M., and Crothers, D. M. (1981) Nucleic Acids Res. 9, 65054525
J. A,, and Struhl, K. (eds) (1987) Current Protocols in Molecular Biology,
19. Guarente, L., Lalonde, B., Gifford, P., and Alani, E. (1984) Cell 36, 503-
20. Einerhand, A. W. C., Voorn-Brouwer, T. M., Erdmann, R., Kunau, W.-H., and
21. Johnston, M. (1987) Microbiol. Reu. 51, 458476 22. Giniger, E., Varnum, S., and Ptashne, M. (1985) Cell 40, 767-774 23. Simon, M., Adam, G., Rapatz, W., Spevak, W., and Ruis, H. (1991) Mol. Cell.
Greene Publishing Associates and Wiley-Interscience, New York
511
Tabak, H. F. (1991) Eur. J . Biochem. 200, 113-122
Biol. 11, 699-704
303309 24. Simon, M., Binder, M., Adam, G., Hartig, A,, and Ruis, H. (1992) East 8,



















![UNOFFICIAL/UNAUTHENTICATED TRANSCRIPT 26 July 2019.]€¦ · UNOFFICIAL/UNAUTHENTICATED TRANSCRIPT UNOFFICIAL/UNAUTHENTICATED TRANSCRIPT 24485 [The R.M.C. 803 session was called to](https://img.pdfslide.net/doc/110x75/5f07260f7e708231d41b8c44/unofficialunauthenticated-transcript-26-july-2019-unofficialunauthenticated.jpg)





![KNOX Protein OSH15 Induces Grain Shattering by Repressing Lignin Biosynthesis Genes1[OPEN] · KNOX Protein OSH15 Induces Grain Shattering by Repressing Lignin Biosynthesis Genes1[OPEN]](https://img.pdfslide.net/doc/110x75/5e3064817026f92f9235ddb5/knox-protein-osh15-induces-grain-shattering-by-repressing-lignin-biosynthesis-genes1open.jpg)



