Embed Size (px)
Citation preview

Biological Conservation 33 (1985) 133-145
The Rationale for Conservation: Problems from a Virgin Forest
J o h n M i d d l e t o n * & G r a y M e r r i a m t
Department of Biology, Carleton University, Ottawa K1S 5B6, Canada
ABSTRACT
We compared the d&tribution of 111 taxa of plants and animals in 18 forests that had received four very different levels of disturbance. A stand of virgin forest undisturbedJbr over 200 years had the lowest number of taxa of any of the forests examined and contained no rare species not found in the more disturbed forests. The relationship of degree of disturbance to species richness among the forests of this study raises questions about the use of ecological knowledge in rationales Jbr conservation. Ecological knowledge must be used in these arguments but with caution and in conjunction with other arguments such as some based on moral philosophy.
I N T R O D U C T I O N
Less than 200 years ago, extensive cont inuous forest covered much of what is now farmland in eastern Canada. A considerable propor t ion of the Great Lakes-St Lawrence Fores t Region (Rowe, 1977) is still covered by trees. However, forests that have not felt considerable human influence are very rare indeed.
* Present address: Institute of Urban and Environmental Studies, Brock University, St. Catherines, Ontario, LZS 3A1. t Will receive correspondence.
133 Biol. Conserv. 0006-3207/85/$03-30 © Elsevier Applied Science Publishers Ltd, England, 1985. Printed in Great Britain

134 John Middleton, Gray Merriam
The changes made to the forests of eastern Canada by European settlers can be used as a natural perturbation experiment to gain insight into essential properties of these forest systems.
We compared certain characteristics of the virgin forest of Shaw Woods, undisturbed by major external perturbation for at least 200 years, with the equivalent characteristics of several other forests that have been affected by humans to very different degrees.
The purpose of the overall study was to analyse spatial responses of species components of these forests to fragmentation, an apparent major effect of the perturbations. Additional interpretation of the comparative data, particularly species richness and occurrence of rare species, raises questions about some of the rationale often still offered by advocates of conservation. The analysis exemplifies, in particular, the danger of singular use of scientific 'truths' in such arguments and points to consideration of complex rationales for conservation combining scientific knowledge and moral philosophy.
METHODS
We examined the spatial distribution of various types of organisms in four types of forest in the Great Lakes-St Lawrence Forest Region of Canada (Rowe, 1977):
(1) Extensive continuous forest (350 km 2) of Gatineau Park, Quebec. (2) Fifteen remnants of forest (1- to 25-ha) in farmland south of
Ottawa. (3) A 50-ha little-disturbed forest reserve (Burnham Provincial Park,
Peterborough). (4) A 50-ha stand of virgin forest contiguous with a much larger area
of forest (Shaw Woods).
Locations of the sampling sites are shown in Fig. 1. All sampling was carried out in mesic mixed deciduous forest
dominated by Acer saccharum. Open canopy, bare rock, standing water, saturated soil, discontinuous litter, and human artefacts were all rare, covering in total no more than 5 ~o of any transect in any area. Canopy, shrub layer, herb layer, and furniture (fallen logs, boulders, etc.) were each assessed to conclude that the forests were comparable.
Farmland transects were in farm woods of 1 to 25-ha within 30 km of

Problems from a virgin forest 135
QUEBEC
Z ,g Fig. I. MapofsouthemOntarioshowingapproximatelocationsofBurnhamforest(B),
Farm woodlots (F), Oatineau forest (O), Shaw Woods (S).
Ottawa. Each had a closed canopy of Acer saccharum with lesser and variable amounts of Fraxinus americana, Fagus grand(folia, Ostrya virginiana, Tilia americana, and other trees. Ground flora included Trillium grandiflorum, Erythronium americanum, Caulophyllum thalic- troides, Allium tricoccum and other species typical of regional mesic forests. Transects were placed along the longest diagonal, to maximise the number of points in each wood, and were 250 to 1000 m long. Woods were on glacial drumlins or on the slopes of drainage valleys (above the level of flooding), and were on land rockier or steeper than adjacent cultivated fields. All woods were surrounded by farmlarld and obvious signs of human impact such as lanes, paths, evidence of grazing, and stumps. Woods were interconnected to variable degrees by fencerows. All but one were from 300 to 600 m away from the nearest woods.
Gatineau Park is a rolling irregular Precambrian upland, generally unsuitable for farming. Less than 10 ~ of its area remains cleared and almost none of this was in our sample. Its climate differs from that of the farmland studied, with a growing season about 11 days (6 ~ ) shorter (Rowe, 1977), even though the two areas are only 30 km apart. We studied a single continuous transect through an area of several thousand hectares of continuous forest. Forests on the transect were similar in composit ion to the farmland woods except for a constant minor representation of conifers such as Tsuga canadensis, Abies balsamea, and Pinus strobus.
Burnham is a mesic forest on a drumlin near Peterborough, Ontario,

136 John Middleton, Gray Merriam
250 km west of Ottawa. It is part of a 600 ha forest, which is in turn surrounded by farmland. Human impact has always been light and it is now protected as a provincial park. We sampled it by two parallel transects along the drumlin. Vegetation was dominated by Acer saccharum, Fagus grandifolia, and Tsuga canadensis, with an understorey of Trillium spp., Erythronium americanum, Caulophyllum thalictroides, and others.
Shaw Woods Nature Reserve includes two main areas of mesic forest on gravelly glacial soils 100 km northwest of Ottawa. One area had had a history like that of Burnham, while the other has been practically untouched by humans (Dugal, 1980). Each was sampled with a single transect. Very old Acer saccharum, Fagus grandifolia and Tsuga canadensis dominated the canopy over a very sparse ground cover.
Humans have lived in this region of Canada for thousands of years (Jenness, 1932), but no large settlements or extensive clearings were made until about 1800. The maximum extent of clearing was not long before 1900 (Scott, 1979). Fire caused extensive changes to forests adjacent to farmland as late as 1900, and to the forest of Gatineau Park in 1923 (Lopoukhine, 1974). Shaw and Burnham have been neither clearcut nor burned within the approximately 200 years of recorded local history (Berry & O'Higgins, 1973; Dugal, 1980).
We determined the presence or absence of various types of organisms at points spaced 25m apart along approximately 6km of transect in Gatineau, 6.5 km in farmland woods, and 1.25 km in each of Burnham and Shaw. In practice, each point was a small circular quadrat (after Pielou, 1964, 1969). For'trees, herbs and ferns, plots were 5 m in radius. Four diurnal sciurids were recorded by sounds, sight or burrows in a 25 m radius. Ten invertebrate taxa including collembolans, pseudoscorpions and hemiptera were detected from 300cm 3 of leaf litter in Tullgren funnels. Herbs were sampled during the period of maximum growth in the spring, all others were sampled in June-August of two years. The taxonomic authorities used were Maynard (1951), Scott (1961), Marie- Victorin (1964), White & Hosie (1968), Cody (1978), Gillett & White (1978), and, for invertebrates, W. R. Richards (pers. comm.). Operational taxonomic units were species in most cases although definition of species was uncertain for some such as collembolans.
Quadrat size for a given taxon was the same in all forests, so comparisons among forests are not biased, but use of different-sized quadrats for different taxa might be considered a problem when

Problems from a virgin forest 137
comparing among taxa. However, we argue that a squirrel present 20 m from a point is, in ecological fact, also present at that point. The much smaller quadrat size used for invertebrates may or may not have underestimated these taxa, relative to the plants and squirrels. However, comparisons of any taxon among different forests are unaffected by this potential bias.
Curves of cumulative number of taxa against number of points sampled were calculated as a measure of the relative species richness of each area. The calculations were made separately for each of fifteen woods in farmland, fifteen segments of the Gatineau transect, and four segments in each of Burnham and Shaw. Composite curves were calculated for each area by taking the mean of all the segments. Composite curves were calculated for all taxa taken together, and for each type of organism (trees, herbs, ferns, squirrels, invertebrates) separately. Furthermore, each of these curves was calculated in two different ways: using all taxa, and using only those taxa found in both Gatineau and farmland.
RESULTS
The complete data set is available from the Depository of Unpublished Data, CISTI, National Research Council of Canada, Ottawa KIA 0S2, Canada, by reference to Middleton (1982).
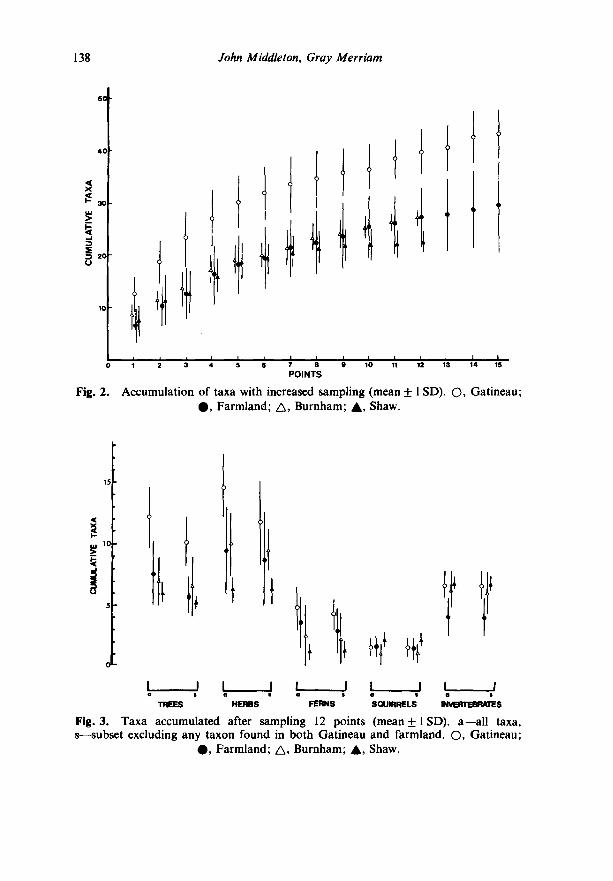
For a given sampling effort, fewer species are found in Shaw Woods than in any other forest examined in this study. This is true for all taxa taken together, and for each of trees, herbs, and ferns taken separately (Figs 2, 3).
Only four species of squirrels were recorded in this study, and they showed no relationship to sampling effort. All but grey squirrel Sciurus carolinensis were seen in Shaw. This squirrel has only recently expanded its range northward in eastern Ontario (Rand, 1945) and was found in only the two most southerly sites of this study (Burnham and farmland).
The soil invertebrates showed a pattern contrary to that of the plants. Shaw yielded a higher number of taxa per unit effort of sampling than any of the other areas. Recall, however, that we did not examine the whole species pool but only a restricted set of ten taxa, so that this result is much less complete than those for the plants and squirrels.
All of the trends reported above remained the same whether calculated on the basis of all taxa, or on the basis of a subset excluding any taxa absent from either of the two largest areas (Gatineau and farmland).
138 John Middleton, Gray Merriam
5 0
4 0
< X
IE 2(
U
0
Fig. 2.
t l I I I
4 I 1 / 1 1 / 110 11 12 13 14 15
P O I N T S
Accumulation of taxa with increased sampling (mean _+ | SD). O, Gatineau; 0 , Farmland; A , Burnham; A , Shaw.
_~ ~c
t '1t I I I I I I I I ~ ~ l 1 a • a a S o •
Fig. 3. Taxa accumulated after sampling 12 points (mean+lSD) . a--all taxa, s--subset excluding any taxon found in both Gatineau and farmland. O, Gatineau;
0 , Farmland; A, Burnham; A, Shaw.

Problems from a virgin forest
TABLE 1 Plant Taxa Recorded from Shaw Woods
Trees Abies balsamea Acer pensyh, anicum and A. spicatum Acer saccharum Betula alleghaniensis Fagus grandifolia Fraxinus americana Ostrya virginiana Tilia americana Tsuga canadensis
Herbs Actaea pachypoda and A. rubra Arisaema triphyllum Caulophyllum thalictroides Erythronium americanum Maianthemum canadense Polygonatum pubescens Ranunculus abortivus Smilacina racemosa Trientalis borealis Trillium erectum Trillium grandiflorum
Fern Dryopteris spinulosa
139
The physical structure of the forests was similar except for ground cover, which was much reduced in Shaw and, to a lesser extent, in Burnham.
All species seen in Shaw were also very common elsewhere. We recorded no species unique to the virgin forest and, in all but five cases, the species seen in Shaw were also seen in each of Burnham, Gatineau, and farmland. The complete list is in Table 1.
DISCUSSION
One of the most obvious differences among the forests examined here is that some are continuous over large areas whereas others are in the form

140 John Middleton, Gray Merriam
of small clumps more or less separate from other nearby woods. However, elsewhere we have shown that this difference in pattern is of little or no significance to the distribution of woodland species in farmland, and cannot be used to explain the difference in the characteristics measured here (Middleton, 1982; Middleton & Merriam, 1983).
All woods examined here are part of the Great Lakes-St Lawrence Forest Region. However, they are found in three different sections of the region (Rowe, 1977). The differences among sections are minor, and mainly concern the relative abundance of the less common species, especially conifers. The more common deciduous species are the same, with Acer saccharum being dominant everywhere. Furthermore, all the results reported here are the same whether they are based on all taxa seen, or on only those found in each of the two most different areas (Gatineau and farmland). Thus, the conclusions reached below are not artefacts of an invalid comparison among incomparable forests.
There was a clear range in the degree of disturbance suffered by these woods, from the heavily disturbed woods in farmland, through the moderately disturbed forests of Gatineau, to the relatively lightly disturbed forests of Burnham Park, and the virgin forest of Shaw Woods.
There are equally clearcut differences in species richness among these forests, with a pronounced maximum in Gatineau (Fig. 2). In addition, of the 98 plant taxa recorded in this study, only 21 were found in Shaw Woods. Eighteen of these were also found in each of the other areas as well (farmland, Gatineau, Burnham), and no species was found only in Shaw. The list of taxa is composed entirely of very common species that could be found with little effort in most forests in the region (Table 1). There is no evidence of accumulation of rare species uniquely adapted to undisturbed forest. On the contrary, the main characteristic of these data is the absence of many taxa found in the more disturbed forests. Furthermore, although we did not record any measure of abundance, our clear impression while working in the virgin forest is that the absolute abundance of plants of any kind, especially ground vegetation, is noticeably lower than in other forests. Vegetation of the virgin forest appears impoverished.
This poverty is not due to any unsuitability of the general area. The more disturbed areas of Shaw Woods immediately adjacent to the virgin stand, but still within the reserve proper, are much richer in species (unpublished observations; Dugal, 1980). The very low richness of the virgin forest is a true feature of the way it has developed.

Problems from a virgin forest 141
The litter invertebrates did not show the same pattern. Although our sampling technique probably underestimated these animals, the results show that the virgin forest is especially rich in species of invertebrates (Fig. 3). The leaf litter of this forest is noticeably thick, and invertebrates are especially numerous compared to samples from other forests. A more detailed survey of invertebrates and other taxa not examined in this study might force modification of our statement that the number of taxa increases less rapidly with increased sampling effort in Shaw than in other, more disturbed, forests.
Responses of these forests to their disturbance regimes are poorly predicted by the classical succession model which increases species diversity as the system matures and more biological specialists accumulate. Rather, maximum species richness is reached at some intermediate successional age, and as the system matures competitive pressures eliminate many species adapted to disturbance and leave a smaller number able to monopolise the resources of the system after a long period free of disturbance.
We conclude that the relationship among the forests examined in this study is well described by the intermediate disturbance hypothesis (Connell, 1978; Huston, 1979). A corollary of this conclusion is that the impoverished vegetation of the virgin forest of Shaw Woods is due to the absence of many species adapted to a more disturbed regimen.
These conclusions have implications for questions of conservation. If we ask why Shaw Woods was considered worthy of protection we find that some of the major rationalisations for conservation fail to justify our concern in this case.
First, parts of Shaw Woods are virgin forest that has remained undisturbed by major catastrophes for at least 200 years, and perhaps for much longer. Undisturbed ecosystems of any kind are a very rare commodity in the more settled parts of Canada these days. The impact of humans on almost all parts of the landscape is very easy to see. Would we not then be justified in arguing that Shaw Woods should be conserved for ecological study and as an archival remnant of what we have destroyed elsewhere?
On consideration of all the facts, we are forced to answer no. Human impacts on the natural world are unquestionably very severe and widespread. However, to a large extent the human impacts have merely replaced equally severe impacts of other sorts. A growing body of evidence demonstrates that frequent disturbance is an integral factor in

142 John Middleton, Gray Merriam
most ecological systems, and that areas left undisturbed for two centuries or more have always been rare (Pickett & Thompson, 1978; Loucks et al., 1981; Pickett & White, in press). Thus we disagree with Dugal (1980) when he suggests that Shaw Woods offers a glimpse of what forest in this region used to be like. Shaw Woods is probably not representative of any widespread ecosystem of the past, and an appeal for its conservation on this basis is ill-founded.
Second, and perhaps the most common plea of conservationists, is for the protection of particular patches of high diversity and for the protection of rare species (e.g. IUCN, 1980; Margules & Usher, 1981; Goode, 1984; Nelson et al., 1984). It is commonly assumed that virgin ecosystems are particularly rich in rare species. Thus, can we not justify the protection of Shaw Woods on the basis of the protection of natural diversity ? Once again the answer is no. The poverty of the vascular plants in the least disturbed sections of Shaw Woods is outstanding. Many other pieces of forest in southern Ontario would be more suitable as a reserve if species richness and rarity of species were the criteria.
For example, we examined a small wood just south of Ottawa, less than a hectare in size. It is a tangle of prickly ash, poison ivy, and dead elms, thoroughly trampled by cattle. We would not want to clear-cut it, but it could be at the bottom of our priorities for conservation. We are confident most naturalists would agree with our assessment. Never- theless, we found more species of plants in this wood, with less than a quarter the sampling effort, than in the virgin forest of Shaw Woods. Furthermore, this little patch had the highest proportion of rarities (although some of them were exotics) of the eighteen woods examined in this study; Shaw Woods had the lowest. We find ,this comparison a powerful reminder of the fact that the protection of diversity is not our main goal in preserving virgin forest. To argue for conservation of species richness would, among the woods studied here, mean choosing this patch of prickly ash, poison ivy and dead elm. It would give the greatest species diversity but it fails to fill the needs underlying the conservation goals of many.
Furthermore, to maximise diversity in each individual woods could mean that every patch of woods could be much like every other patch, on similar sites throughout the region. If, as Connell (1978), Huston (1979), Pickett (1980), Loucks et al. (1981) and others argue, and we have confirmed here, maximum species richness occurs at an intermediate stage of maturity in response to disturbance, then some sort of second

Problems from a virgin forest 143
growth, certainly not virgin or mature forest, would be the general type of high diversity wood. Sampson & Knopf (1982) also recognised the danger in maximising diversity and recommended against it as a management principle.
An alternative conservation policy could take its basis in Whittaker's (1972) discussion of alpha, beta and gamma diversities. Gamma diversity measures across the patchwork of large-scale heterogeneity such as a small region. A conservation policy attempting to maximise gamma diversity would produce a variety of types of forest patches including overmature as well as intermediate and younger stages. Maximising an index of regional heterogeneity related to gamma diversity would ensure a full range of ecological assemblages capable of replacing themselves and with high potential for genetic exchange and ecological coupling.
A justification for conservation that has the ring of objectivity and 'scientific truth' can seem particularly valuable in public discussions. However, it is dangerous to forget the nature of scientific support for a conservation ethic. The justification provided by scientific 'truths' will shift dynamically as scientific knowledge improves through revision, rejection and replacement of views in the light of new findings. Nevertheless, we believe that the current body of ecological knowledge has an important role in conservation arguments by interpreting life processes and by providing large-scale synthesis of systems views.
Other arguments also have importance in justifying the conservation of virgin forest, as an honest examination of motives will reveal. The massive size and broad spacing of trees in Shaw Woods are aesthetically very powerful, quite aside from any scientific significance they may have. However, aesthetic justification alone may not be sufficient. Some ecosystems with strong aesthetic appeal, when interpreted ecologically, may display life processes that complicate justifying their conservation. Consider transparent acidified lakes and their unacidified counterparts.
These arguments based on economic, scientific or aesthetic con- siderations may be called utilitarian because they all relate to human values such as beauty, economic worth or scientific utility. Many of our motives for conservation, however, seem to be rooted at another level, somewhat isolated from such utilitarian or instrumental demands (cf. Peterken, 1981). Attempts to justify such 'deeper', non-anthropocentric motives for saving the natural world can easily be explored in the literature (e.g. Livingston, 1981 ; Skolimowski, 1981 ; Regan, 1982). These

144 John Middleton, Gray Merriam
arguments involve concepts such as the rights of individual components of natural systems, the inherent value of natural processes, and the related notion of health or sustainability of ecological systems. It is true that these arguments, based on moral philosophy, may not provide the simplicity and charisma sought in public discussion. Nevertheless, they should be pursued by all conservationists for ultimately they may be the rationale of our real concern for the protection of the natural world.
ACKNOWLEDGEMENTS
We received excellent advice and direct assistance from J. Wegner and S. Hendler, constructive suggestions from our editor and an anonymous reviewer, and financial support from the Natural Sciences and Engineering Research Council of Canada, Gulf Canada Limited and Carleton University.
REFERENCES
Berry, G. S. & O'Higgins, B. F. (1973). Woodlots, water, wildlife, and wilderness. Peterborough, Ontario, Peterborough Field Naturalists.
Cody, W. J. (1978). Ferns of the Ottawa district. Ottawa, Canada Department of Agriculture.
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science, N.Y., 199, 1302-10.
Dugal, A. (1980). Shaw Woods Nature Reserve. Trail and Landscape, 14, 46-56. Gillett, J. M. & White, D. J. (1978). Checklist of vascular plants of the
Ottawa-Hull region, Canada. Ottawa, National Museum of Natural Sciences.
Goode, D. A. (1984). Conservation and value judgements. In Planning and ecology, ed. by R. D. Roberts and T. M. Roberts, 188-205. London, Chapman and Hall.
Huston, M. (1979). A general hypothesis of species diversity. Am. Nat., 113, 81-101.
IUCN (1980). Worm conservation strategy. Gland, IUCN, UNEP, WWF. Jenness, D. (1932). Indians of Canada. Toronto, University of Toronto Press. Livingston, J. A. (1981). The fallacy of wildlife conservation. Toronto,
McClelland and Stewart. Lopoukhine, N. (1974). The forests and associated vegetation of Gatineau Park,
Quebec. Ottawa, Forest Management Institute Information Report FMR- X-58.

Problems from a virgin forest 145
Loucks, O. L., Ek, A. R., Johnson, W. C. & Monseurd, R. A. (1981). Growth, aging and succession. In Dynamic properties of forest ecosystems, ed. by D. E. Reichle, 37-85. Cambridge, Cambridge University Press.
Margules, C. & Usher, M. B. (1981). Criteria used in assessing wildlife conservation potential: A review. Biol. Conserv., 21, 79-109.
Marie-Victorin (1964). Flore laurentienne. Montr6al, Les Presses de l'Universit6 de Montreal.
Maynard, E. A. (1951). A monograph of the Collembola or springtail insects of New York State. Ithaca, New York, Comstock Publishing Company.
Middleton, J. (1982). On certain spatial characteristics of the distribution of woodland species in farmland. PhD thesis, Ottawa, Carleton University.
Middleton, J. & Merriam, G. (1983). Distribution of woodland species in farmland woods. J. appl. Ecol., 20, 625-44.
Nelson, P. J., Corlett, J. & Swanwick, C. (1984). Environmental impact assessment procedures used in a strategic study of water resource development. In Planning and ecology, ed. by R. D. Roberts and T. M. Roberts, 265. London, Chapman and Hall.
Peterken, G. F. (1981). Woodland conservation and management. London, Chapman and Hall.
Pickett, S. T. A. (1980). Non-equilibrium co-existence of plants. Bull. Torrey bot. Club, 107, 238-48.
Pickett, S. T. A. & Thompson, J. N. (1978). Patch dynamics and the design of nature reserves. Biol. Conserv., 13, 27-37.
Pickett, S. T. A. & White, P. S. (Eds) (in press). Natural disturbance: an evolutionary perspective. New York, Academic Press.
Pielou, E. C. (1964). The spatial pattern of two-phase patchworks of vegetation. Biometrics, 20, 156-67.
Pielou, E. C. (1969). An introduction to mathematical ecology. Toronto, Wiley- Interscience.
Rand, A. L. (1945). Mammals of the Ottawa district. Can. Fld-Nat., 59, 111-35. Regan, T. (1982). All that dwell therein. Essays on animal rights and
environmental ethics. Los Angeles, California, University of California Press.
Rowe, J. S. (1977). Forest regions of Canada. Ottawa, Can. For. Serv. Pubis, No. 1300.
Sampson, F. B. & Knopf, F. L. (1982). In search of a diversity ethic for wildlife management. Trans. N. Am. Wildl. nat. Resour. Conjl, 47th, 421-31.
Scott, H. G. (1961). Collembola: pictorial key to the nearctic genera. Ann. Ent. Soc. Am., 54, 104-13.
Scott, O. R. (1979). Utilizing history to establish cultural and physical identity in the landscape. Landscape Planning, 6, 179-203.
Skolimowski, H. (1981). Eco-philosophy. Boston, Marion Boyars. White, J. H. & Hosie, R. C. (1968). The forest trees of Ontario. Toronto,
Department of Lands and Forests. Whittaker, R. H. (1972). Evolution and measurement of species diversity. Taxon,
21, 213-51.