Embed Size (px)
Citation preview

International Journal of Angiology 4:88-93 (1995)
The Role of MHC Class I and Class II Molecules in Susceptibility to Lyme Disease in Inbred B10 H-2 Congenic and Recombinant Mice
Sunitha Reddy, M.S . , 1 Colin R. Young, Ph.D., 2 Regina Hokanson, B.S., z Julie Rawlings, MPH., 3 Chella S. David, Ph.D. 4
Departments of ~Biology and ZVeterinary Pathobiology, Texas A&M University, College Station, Texas; 3Texas Department of Health, Austin, Texas; and 4Department of Immunology, Mayo Clinic, Rochester, Minnesota, USA
Abstract. We have further developed the mouse model of Lyme disease using B10 H-2 congenic and recombinant strains. The influence of MHC class I and class II genes of the mouse H-2 complex on the sus- ceptibility and/or resistance to Lyme disease was stud- ied. We report that B10 strains congenic to the haplo- types, H-2 k, H-2 a, H-2 f, H-2 q, and H-2 b are highly susceptible to Lyme disease as they develop high an- tibody titers and severe disease over mice of other haplotypes used in this study. By using recombinant strains we demonstrate that this susceptibility is ge- netically based and specifically involves the K and D regions (MHC class I genes) and I-A and I-E regions (MHC class II genes) of the H-2 complex.
Introduction
Decades before the causative agent of Lyme disease was discovered, the disease (then not known as Lyme disease) had previously been described as separate en- tities. Description of this disease by Buchwald [1] in Germany dates back as early as 1883 as an idiotypic skin atrophy. In Europe, different manifestations such as erythema chronicum migrans (ECM), lymphadeno- sis benigna cutis (LABC), acrodermatitis chronica atrophicans (ACA), and the lymphocytic meningora- diculitis, later named as Garin-Bujadoux-Bannwarth syndrome, have been recognized since the first de- cades of this century [2-6]. Arthritis was first recog- nized in the 1970s in the United States by Allen Steere [7,8] as a new form of inflammatory arthritis, initially named Lyme arthritis. On further evaluation it was
Presented at the 35th Annual World Congress, International College of Angiology, Copenhagen, Denmark, July 1993 Correspondence to: Colin Young, Ph.D., Department of Veterinary Pathobiology, Texas A&M University, College Station, TX 77843, USA
found that patients also developed other neurological and/or cardiac problems. Due to the involvement of different organs and organ systems, this multisystemic disease was renamed Lyme disease in I971. All the above-mentioned disorders are now recognized as components within the Lyme disease spectrum [7,8]. The organism causing Lyme disease is a bacterial spi-
rochete, Borrelia burgdorferi, and was identified by Willy Burgdorfer in 1982 [9,10]. Several tick species of the genus Ixodes (I. dammini/scapularis, I. ricinus, I. pacificus) are vectors to B. burgdorferi [11-13] includ- ing Amblyomma americanum (Lone-star tick), an- other genus of the Ixodidae family [14].
There may be a genetic susceptibility to chronic in- fection with Lyme disease as there appears to be an association between HLA (Human leukocyte anti- gens) phenotypes and the occurrence of arthritis. The frequency of certain HLA phenotypes is increased in various immunologic disorders with rheumatologic manifestations. Studies indicate that patients with HLA-DR2 and HLA-DR4 antigens may be prone to develop progressive, destructive lyme arthritis that is refractory to antibiotic therapy [15]. It has been hy- pothesized that infectious agents may trigger in genet- ically susceptible persons an immune response that leads to chronicity. Though the linkage of an HLA phenotype to the predilection for the development of late manifestations of Lyme borreliosis remains a pos- sibility, the existence of such an association has yet to be demonstrated conclusively [16].
We have further developed the already documented mouse model by studying the incidence of Lyme dis- ease in 3-week-old B10 H-2 congenic and recombinant mouse strains [17]. The significance of using such strains is that any difference in the response to inoc- ulation with B. burgdorferi would only be due to the different genotype that exists in the H-2 locus and not to other background genes. We have evaluated the antibody responses of the different haplotypes to the B31 isolate ofB. burgdorferi and mapped them to their

S. Reddy et al.: Class 1 and II Molecules and Lyme Disease 89
Table 1. Genetic map of BI0 congenic mouse strains at the K, I-A, I-E, and D regions of the H-2 complex with peak antibody response to experimental inoculation with B31 B. burgdorferi
Alleles at H-2 loci Antibody Mouse H-2 response strain haplotype K I-A I-E D (cpm)
B10.D2 d d d d d 22,000 B10.S s s s s s 20,000 B10.P p p p p p 15,000 BRVR.B10 b b b b b 22,000 B10 b b b b b 21,000 B10.SM v v v v v 14,000 B6/kh b b b b b 20,000 B10.M f f f f f 33,000 B10.K k k k k k 23,000 BRVR k k k k k 22,000 B 10.Q q q q q q 26,000 BI0.Rll l r r r r r 17,000
g e n e t i c m a p s o f t h e H - 2 c o m p l e x [17]. W e s h o w t h a t m o u s e o f d i f f e r e n t h a p l o t y p e s r e s p o n d d i f f e r e n t l y , a n d w i t h t h e u s e o f r e c o m b i n a n t s t r a i n s w e a l s o s h o w t h a t th is d i f f e r e n c e in s u s c e p t i b i l i t y a n d / o r r e s i s t a n c e m a p s to t h e H-2 c o m p l e x , s p e c i f i c a l l y , t h e c l a s s I a n d c l a s s II g e n e s . T h e s e s t u d i e s w e r e c o n d u c t e d in a n t i c i p a t i o n o f d e v e l o p i n g B10 t r a n s g e n i c m i c e e x p r e s s i n g h u m a n C l a s s I a n d C l a s s II H L A a n t i g e n s . E x p e r i m e n t a t i o n u s i n g s u c h m i c e wil l tel l us a b o u t a n y r o l e t h a t c l a s s I a n d c l a s s II m o l e c u l e s m i g h t p l a y in s u s c e p t i b i l i t y a n d / o r r e s i s t a n c e to L y m e d i s e a s e in h u m a n s .
Materials and Methods
Experimental Animals
Groups of five mice consisting of 12 BI0 H-2 congenic strains (BI0.D2, BI0.S, BI0.P, BRVR.BI0, BI0, B10.SM, B6/kh, BI0.M, BI0.K, BRVR, BI0.Q, and B10.RIII) and eight B10 H-2 recombi- nant strains (BI0.TL, BI0.OL, BI0.PL, B10,RKB, B6-CH-2 bin12, BI0,RDD, B10.RKD1, and B10.AQR) were used for experimental inoculation with the Lyme disease spirochete Borrelia burgdorferi. The H-2 haplotype designations of these strains are shown in Tables 1 (haplotype of congenic strains) and 2 (haplotype of recombinant strains). Mice were obtained from Dr. Chella S. David, Dept. of Immunology, Mayo Clinic, Rochester, MN. All mice were pathogen free and shipped in different crates. They were housed separately for the infectivity experiments, five per cage, in isolation cubicles with an air filtered environment maintained at 20-22~ All mice were prebled before inoculation and were found to be sero-negative and culture negative for B. burgdorferi.
Borrelia Isolate
The B31 isolate of B. burgdorferi originally isolated from the deer tick [18] Ixodes dammini, Shelter Island, NY was obtained from Dr. Julie Rawlings, Dept. of Health, Austin, TX. The culture was main- tained in Kelly's liquid medium [10] until inoculation.
Inoculation o f Animals
Five mice from all strains of BI0 H-2 congenic and recombinant mice were injected with either phosphate-buffered saline (PBS) (in case of negative control animals) or 1 • l 0 6 live spirochetes of B31
Table 2. Genetic map of BI0 recombinant mouse strains at the K, I-A, I-E, and D regions of the H-2 complex 22 with peak antibody response to experimental inoculation with B31 B. burgdorferi
Alleles at H-2 loci Antibody Mouse H-2 response strain haplotype K I-A I-E D (cpm)
B10.TL tL s k k d 11,000 B10.OL oL d d d k 10,000 BI0.PL u u u u d 16,000 B10.RKB h k k k b 14,000 B-6-CH-2 bma~ b b mt b b 12,000 BI0.RDD d d d - - d 14,000 B 10.RKD 1 a k k k d 10,000 BI0.AQR yL q k k k 11,000
mt = mutation.
isolate in a single intraperitoneal (ip) or intradermal (id) site. Low passage spirochetes were used, and prior to inoculation the organ- isms were washed twice in PBS buffer. Control and test mice were bled from the tail vein once every 2 weeks over a 10-week period. Mice were observed daily for disease symptoms, and observations were recorded.
Histological Examinat ion
At 10 weeks postinoculation, all mice were sacrificed by carbon- dioxide inhalation. At necropsy, half the brain and spleen organs were removed for spirochete recovery and the rest of the tissue and organ samples such as skin, joints, heart, lungs, liver, spleen, brain, and kidneys were fixed in 10% neutral-buffered formalin (pH 7.2) for histologic study. Joints were decalcified, tissue and organs were processed and paraffin-embedded, sectioned at 5-7 ~m, and stained with hematoxylin and eosin using standard techniques. Selected tis- sues were stained for spirochetes using Steiner's or Warthin-Starry silver staining procedures [19].
Serology
Serum samples were tested for B. burgdorferi antibodies by a pre- viously described radioimmunoassay (R1A) [20] with additional modifications. This assay was recently developed and standardized at Texas A&M University. In the RIA, 96-well PVC flexible plates were coated with 50 ~1 antigen (1 x 105 spirochetes per well) in 0.01% Na azide/PBS buffer solution and incubated overnight at 20- 25~ to allow antigen binding. Plates were then drained and washed with 0.1% gelatin/PBS buffer (pH 7.2) solution. Wells were filled with 0.1% gelatin/PBS and incubated for 30 minutes to block any unbound sites. The plates were then washed with the same solution followed by two washes with PBS buffer. Fifty microliters dilutions of mouse antisera (10- ~ to 10 4) in 0.1% gelatin were added to each well in duplicates and incubated for 2 hours at room temperature. Antiserum was removed and plates were washed three times with 0.1% gelatin/PBS solution and once with PBS buffer. The assay was developed by the addition of protein-A-1125, 50 i~l/well (in 0.1% gel- atin/PBS containing 100,000 cpm/well). Following incubation for 1 hour, the labeled material was removed and wells were washed with PBS buffer. After drying the plates, wells were cut out and counted in a gamma counter. Each plate included previously determined, known positive control sera and known negative control sera. All sera were tested simultaneously. Results are expressed as the counts per minute (cpm) of test samples minus the cpm of the neg- ative control serum samples.
Results
T h e l is t o f B I0 H-2 c o n g e n i c s t r a i n s a l o n g w i t h t h e g e n e t i c m a p s o f t h e H-2 l o c u s a n d t h e p e a k R I A an t i -

90 S. R e d d y e t al . : C l a s s I a n d II M o l e c u l e s a n d L y m e D i s e a s e
25000
~ 15c~
5000 o , , , , i
BI0 Congenir Strains
[ I First Bleed D Second Blce.d I Third Bleed " Fourth Bleed 1
Fig. 1. A n t i b o d y r e s p o n s e (1:10 d i lu t ion ) o f B10 c o n g e n i c m i c e to i n o c u l a t i o n w i t h B31 B. burgdorferi.
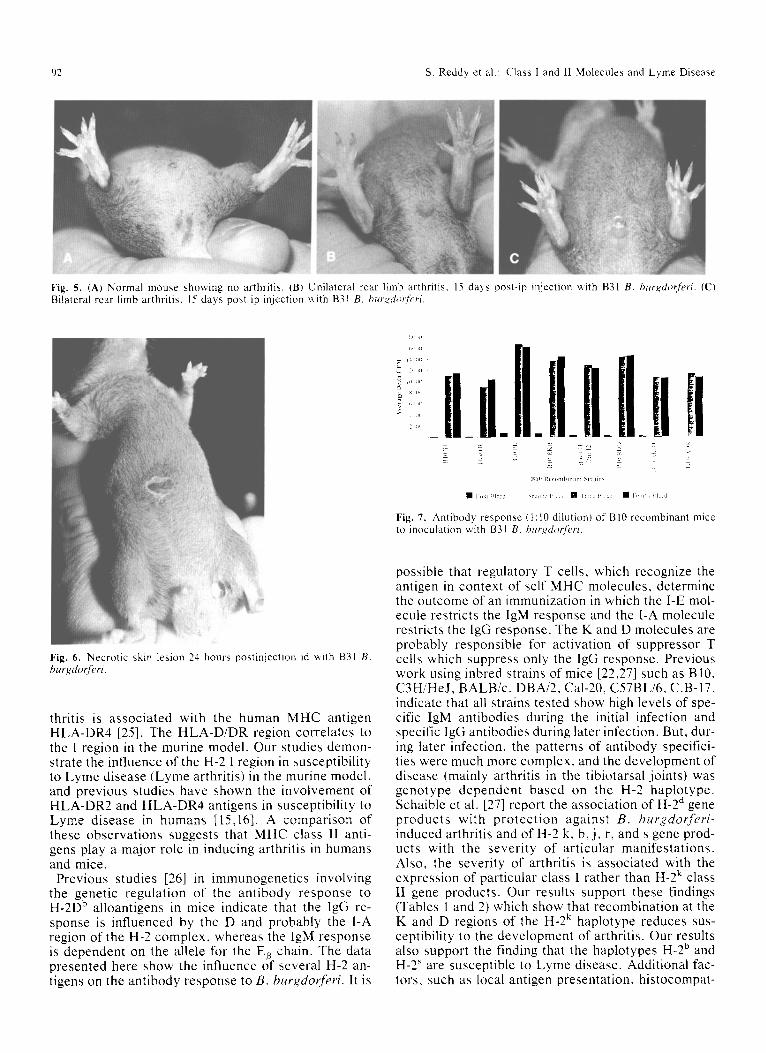
body titers (in cpm) of individual strains is shown in Table 1. Congenic strains of all genotypes developed an immune response to ip inoculation ofB. burgdorferi but the degree of immune response varied with geno- type (Fig. 1). Congenic strains of haplotypes H-2 f (BI0.M), H-2 b (BRVR.B10, B10, and B6/kh), H-2 k (B10.K, BRVR), H-2 q (B 10.Q), H-2 S (BI0.S), and H-2 d (BI0.D2) developed high antibody titers compared with other congenic strains of haplotypes, H-2 p (BI0.P), and H-2 v (B10.SM) which developed low an- tibody titers indicating a genetic influence of the H-2 haplotype. The strains that developed high antibody titers also developed severe disease. Pathologic le- sions were observed in the livers (Fig. 2), lungs (Fig. 3), and spleens (Fig. 4) of these susceptible strains. Warthin-Starry silver staining of the liver showed spi- rochetes in the hepatocytes. Spirochetes were recov- ered from the brain of some of these strains but not from the spleen. Mice showing a high immune re- sponse uniformly developed either unilateral (Fig. 5b) or bilateral rear limb arthritis (Fig. 5c). Based on these clinical and pathologic findings and also the high anti- body titers, the following haplotypes--H-2 f (B10.M), H-2~ B10, and B6/kh), H-2 k (B10.K, and BRVR), H-2 q (B10.Q), and H-2 d (B10.D2)--were cat- egorized as susceptible strains that develop a high im- mune response and severe disease to infection with B. burgdorferi. Splenomegaly was common in all high (susceptible) and low (resistant) responding strains. No other histologic lesions were found nor spirochetes recovered from any of the organs of the low respond- ing strains. Control mice had no lesions and were cul- ture negative.
Susceptible strains (B10.M, BRVR.B10, B6/kh, B10.K, BI0.Q, and BI0.D2) of mice when injected id with B. burgdorferi, developed necrotic skin lesions (Fig. 6) and arthritis. In addition, splenomegaly and kidney enlargement was observed. Spirochetes were recovered from the brains of these mice. No signifi- cant lesions were found in the lungs or livers of any of these mice.
In order to specifically map the gene(s) within the H-2 complex responsible for susceptibility to Lyme dis- ease and Lyme arthritis, similar studies were con- ducted using B10 H-2 recombinant strains. The im- mune response of the individual recombinant strains to
ip inoculation with B. burgdorferi is shown in Fig. 7. Table 2 shows the genetic map of the H-2 locus [17] and the peak antibody levels (in cpm) of the individual strains. It can be seen that there is a significant de- crease in the antibody titers of strains BI0.TL, B I 0 . O L , B I 0 . R K B , B6-CH-2 bm12, B10.RDD, BI0.RKDI, and B10.AQR (Tables 2 and 3) as a result of substitution or deletion in the gene loci of either the K, I-A, I-E, or D regions of the H-2 complex. These recombinant strains are a result of recombination of specific congenic strains (parental strains), as seen in Tables 1 and 2. The influence of the parental H-2 hap- lotype through the class I and/or class II MHC mole- cules on disease susceptibility is summarized in Table 3. H-2K and H-2D regions are MHC class I genes whereas, H-2/I-A and H-2/I-E regions are MHC class II genes. The recombinant strain, B10.TL, shows a 50% reduction in the antibody titer (11,000 cpm) as a result of a recombination in the H-2K and H-2D re- gions of its parental strain B10.K (23,000 cpm), a higher responder indicating the influence of class I genes of the H-2 k haplotype (Tables 1 and 2). Simi- larly, B10.OL (10,000 cpm) differs from its parental strain B10.D2 (22,000 cpm) in the H-2D region and shows a 50% reduction in the immune response sug- gesting a H-2D d influence (class I molecule). Strain B 6 - C H - 2 bm12 is a mutant form of its parental strain B6/kh in the I-A region. Both strains have the H-2 b haplotype but B6-CH-2 bin12 differs in three amino acid residues in the [3 chain of the I-A region (Table 3) and shows a significant decrease in the antibody titer (12,000 cpm) when inoculated with the same dose and culture of B. burgdorferi as B6/kh (20,000 cpm). This shows the influence of class I! genes on the antibody response to B. burgdorferi [17]. Another significant observation was made in the recombinant strain BI0.RDD whose antibody titer is 14,000 cpm. Both B10.D2 and B10.RDD have the H-2 d haplotype but B 10.RDD differs from its parental strain B 10.D2 in the sense that it lacks the I-E gene (Tables 1 and 2). The impact of this deletion is seen on the immune re- sponse. B10.RDD has an antibody titer of 14,000 cpm whereas the parental strain B10.D2 has an antibody titer of 22,000 cpm. This is another indication of the influence of class II genes, i.e., the I-E region in the haplotype H-2 d. No histologic lesions were found in any of the tissue and organ sections of the B10 recom- binant strains except for splenomegaly. Control mice did not seroconvert and had no lesions, and were cul- ture negative for spirochetes.
Discussion
The Major histocompatibility (MHC) restriction of the immune response, whereby foreign antigens can only be recognized by T-lymphocytes when presented in association with class I or class II MHC molecules, leads to the understanding that MHC polymorphism is maintained by different alleles providing varying de- grees of protection against infectious pathogens [21].

S. Reddy et al.: Class I and ll Molecules and Lyme Disease 91
Fig. 2. Warthin Starry silver staining of liver, 4 weeks post-ip injection with B31 B. burgdorferi spirochetes intact in hepatocytes. Spirochetes phagocytosed by Kupffer cells. Fig. 3. Pneumonitis of the lung, 4 weeks post-ip injection with B31 B. burgdorferi. Fig. 4. Spleen, 4 weeks post-ip injection with B31 B. burgdorferi. Splenic follicle shows lymphoid depletion. There is hemorrhagic infiltration in the red pulp area.
An individual who is heterozygous for MHC alleles would have an increased capacity to present antigens from a range of pathogens compared with homozy- gotes. The use of B10 H-2 congenic and recombinant strains of mice as a laboratory model in this study has been a key factor in positively determining the effect of class I and class II genes on the pathogenesis and immune response of mice to B. burgdorferi, the caus- ative agent of Lyme disease. Most of the published mouse models of Lyme disease [22] use strains of mice where the background genes outside the H-2 complex are different amongst the strains studied. The use of B10 H-2 congenic strains has enabled us to conclude that the susceptibility to Lyme disease is due to de- fined H-2-1inked genes and not due to non-H-2-1inked background genes. Using B I0 H-2 recombinant strains of mice we have been able to fine map the specificity of genes within the H-2 complex responsible for the susceptibility to Lyme disease, namely, the H-2K, H-2D (MHC class I genes) and the H-2/I-A, H-2/I-E (MHC class II genes). Congenic and recombinant strains can be used as po-
tential animal models for Lyme disease research be- cause there are reports that indicate that both Lyme disease and Lyme arthritis are associated with human HLA antigens such as HLA-DR2 and HLA-DR4 [15,16]. HLA-DR2 antigens are associated with pe- ripheral facial palsy combined with other cranial nerve impairment and HLA-DR4 antigens are associated
with radiculitis and myeloradiculitis. Further, both HLA-DR2 and HLA-DR4 (class II antigens) were fre- quent in cases of joint pain or arthritis in Lyme dis- ease. Lyme disease spirochetes are known to persist for long periods in the host and trigger new immune responses during later attacks of arthritis [23]. In ear- lier studies of the mouse model [22], susceptible strains (3-week-old C3H/He, SWR, and BALB/c mice) developed higher antibody titers and severe polyar- thritis to infection with B. burgdorferi than did resis- tant mice (C57BL/6 and SJL mice). Our results corre- late with these findings that susceptible strains of mice under the influence of specific H-2 haplotypes devel- oped unilateral and bilateral rear limb arthritis with high antibody titers.
Susceptibility to collagen-induced arthritis (CIA) is associated with a high antibody response to type II collagen and both are controlled by MHC genes in both the rat and the mouse. In mice, the use of recom- binant strains indicated that CIA susceptibility is as- sociated with the I region gene products in H-2 q and the H-2 r haplotype mice [24]. Our results also show that H-2 q and H-2 r haplotypes are susceptible to Lyme disease. This similarity indicates a parallel between CIA and Lyme arthritis and the involvement of MHC antigens. CIA in mice bears a close resemblance to human rheumatoid arthritis in the histologic and clin- ical manifestations of the disease. It is now well estab- lished that increased susceptibility to rheumatoid ar-
92 S. Reddy et al.: Class I and ll Molecules and Lyme Disease
Fig. 5, (A) Normal mouse showing no arthritis, (B) Unilateral rear limb arthritis, 15 days post-ip injection with B31 B, burgdorferi. (C) Bilateral rear limb arthritis, 15 days post-ip injection with B31 B. burgdotferi.
. . . . . . . . . . U.n I II n it Sf,l:,l
, I , l !
4;:',,,
, 2 k Z E
Ill{} Rct mllhinarl l SIT;lirI~
�9 F lagl Bl : :d q::t lr 'd El:~d �9 l~lllkl [~l~d �9 I Lilrll, Iq d
Fig. 7. Antibody response (1:10 dilution) of B10 recombinant mice to inoculation with B31 B. b,rx, dot:f~'ri.
Fig. 6. Necrotic skin lesion 24 hours postinjeclion id with B31 B. burgdotferi.
thritis is associated with the human MHC antigen HLA-DR4 [25]. The HLA-D/DR region correlates to the I region in the murine model. Our studies demon- strate the influence of the H-2 I region in susceptibility to Lyme disease (Lyme arthritis) in the murine model, and previous studies have shown the involvement of HLA-DR2 and HLA-DR4 antigens in susceptibility to Lyme disease in humans [15,16]. A comparison of these observations suggests that MHC class II anti- gens play a major role in inducing arthritis in humans and mice.
Previous studies [26] in immunogenetics involving the genetic regulation of the antibody response to H-2D b alloantigens in mice indicate that the IgG re- sponse is influenced by the D and probably the I-A region of the H-2 complex, whereas the IgM response is dependent on the allele for the E~ chain. The data presented here show the influence of several H-2 an- tigens on the antibody response to B. b.rgdopferi. It is
possible that regulatory T cells, which recognize the antigen in context of self MHC molecules, determine the outcome of an immunization in which the I-E mol- ecule restricts the IgM response and the I-A molecule restricts the IgG response. The K and D molecules are probably responsible for activation of suppressor T cells which suppress only the IgG response. Previous work using inbred strains of mice [22,27] such as B10, C3H/HeJ, BALB/c, DBA/2, Cal-20, C57BL/6, C.B-17, indicate that all strains tested show high levels of spe- cific IgM antibodies during the initial infection and specific IgG antibodies during later infection. But, dur- ing later infection, the patterns of antibody specifici- ties were much more complex, and the development of disease (mainly arthritis in the tibiotarsal joints) was genotype dependent based on the H-2 haplotype. Schaible et al. [27] report the association of H-2 d gene products with protection against B. b,rgdorferi- induced arthritis and of H-2 k, b, j, r, and s gene prod- ucts with the severity of articular manifestations. Also, the severity of arthritis is associated with the expression of particular class I rather than H-2 k class II gene products. Our results support these findings (Tables 1 and 2) which show that recombination at the K and D regions of the H-2 k haplotype reduces sus- ceptibility to the development of arthritis. Our results also support the finding that the haplotypes H-2 6 and H-2 ~ are susceptible to Lyme disease. Additional fac- tors, such as local antigen presentation, histocompat-

S. Reddy et al.: Class I and II Molecules and Lyme Disease 93
Table 3. Representative B 10 strains showing influence of class I and class II MHC genes involved in susceptibility to Lyme disease in mice
Antibody response
H-2 lines (cpm) Characteristics
B6/kh 20,000 B6-CH-2 bin12 12,000
B 10.D2 22,000 B10.RDD 14,000
B10.K 23,000 B10.RKD1 10,000
BI0.AQR 11,000
H-2 b haplotype Similar to B6/kh but differs in only
3 amino acid residues in 13-chain of I-A region
Influence of MHC class II genes H-2 d haplotype Similar to BI0.D2 but lacks I-E
gene Influence of class II MHC H-2 k haplotype Recombinant at the H-2 k D region Influence of MHC class I genes Recombinant at the H-2 k K region Influence of MHC class I genes
ibility antigen expression, and involvement of nonim- mune cells must also be involved because the chronic manifestations in Lyme borreliosis are so variable.
In conclusion, it can be said that limitations in the ability of a particular MHC molecule (haplotype) to associate with an antigen may explain the linkage of histocompatibility antigens to variations in susceptibil- ity to disease, as demonstrated in this study. We have demonstrated that B10 H-2 mice strains congenic to haplotypes H-2 k, H-2 d, H-2 f, H-2 q, and H-2 b are highly susceptible to experimental inoculation with the Lyme disease spirochete Borrelia burgdorferi over several other haplotypes used in this study, and that class I and class II MHC genes of the H-2 complex play an important role in inducing this genetic response to Lyme disease. These findings will help in generating transgenic mice wherein human MHC class I and class II genes will be introduced into mouse B 10 haplotypes which are high or low responders to B. burgdorferi, and the effects of these transgenes will be determined.
Acknowledgment. This work was supported in part by a grant from Texas A&M University.
References
1. Buchwald A (1883) Ein fall von diffuser idiopathischer haut- atrophie. Arch derm syph (Berlin) 15:553-556.
2. Afzelius A (1921) Erythema chronicum migrans. Acta Derm Ve- nereal 2:120-125.
3. Lipschutz B (1913) Uber eine settene erytheform (erythema chronicum migrans). Arch Dermatol 118:34%356.
4. Garin C, Bujadoux (1922) Paralysie pae les Tiques. J Med Lyon 71:765-767.
5. Bannwarth A (1941) Chronische lymphocytare meningitis en- tzundliche polyneuritis and "Rheumatismus" eing beitrag zum problem "Al le rg ic and N e r v e n s y s t e m . " Arch Psychia t r Nervenkr 113:284-376.
6. Bannwarth A (1944) Zur klinik und pathogenese der "chronis- chen lymphocytaire meningitis." Arch Psychiatr Nervenkr 117: 682-716.
7. Steere AC, Malawista SE, Snydman DR, et al (1977) Lyme arthritis: An epidemic of aligoarticular arthritis in children and adults in three Connecticut communities. Arthritis Rheum 20:7.
8. Steere AC, Malawista SE, Hardin JA, et al (1977) Erythema chronicum migrans and Lyme arthritis: The enlarging clinical spectrum. Ann Intern Med 86:685-698.
9. Johnson RC, Schmid GP, Hyde FW, et al (1984) Borrelia burg- dorferi sp. nov: Etiological agent of Lyme disease. Int J Syst Bac 34:496--497.
10. Steere AC, Grodzicki RL, Kornblatt AN, et al (1983) The spi- rochetal etiology of Lyme disease. N Engl J Med 308:773-740.
11. Steere AC, Malawista SE (1979) Cases of Lyme disease in the United States: Locations correlated with distribution of lxodes dammini. Ann Intern Med 91:730-733.
12. Burgdorfer W, Lane RS, Barbour AG, et al (1985) The western black-legged tick Ixodes pacificus: A vector of Borrelia burg- dorferi. Am J Trop Med Hyg 34:925-930.
13. Dekonenko EJ, Steere AC, Beraudi VP, et al (1988) Lyme bor- reliosis in the Soviet Union: A co-operative US-USSR report. J Infect Dis 158:748-753.
14. Schulze TL, Bowen GS, Bolser EM, et al (1988) Amblyyomma americanum: A potential vector of Lyme disease in New Jer- sey. Science 224:601-603.
15. Steere AC (1988) Pathogenesis of Lyme arthritis. Ann NY Acad Sci 539:87-92.
16. Finn AF, Dattwyler MD (1990) The immunology of Lyme bor- reliosis. Lab Med 21(5):305-309.
17. Klein J, Figueroa F, Klein D (1982) H-2 haplotypes, genes, and antigens: Second listing. Immunogenetics 16:285-317.
18. Johnson RC, Schmid GP, Hyde FW, et al (1984) Borrelia burg- dorferi sp. nov: Etiological agent of Lyme disease. Int J Syst Bact 34:496--497.
19. Thompson SW (1966) Modified Warthin-Starry method for the demonstration of spirochetes. In: Selected Histochemical and Histopathological Methods. Charles C. Thomas: Springfield, IL.
20. Clague RB, Shaw MJ, Holt L (1980) Incidence of serum anti- bodies to native Type I and Type II collagen in patients with inflammatory arthritis. Ann Rheum Dis 39:201-206.
21. Hill A, Allsopp C, Kwiatkowski D, et al (1991) Common West African HLA antigens are associated with protection from se- vere malaria. Nature 352:595-600.
22. Barthold SW, Beck DS (1990) Lyme borreliosis in selected strains and ages of laboratory mice. J Inf Dis 162:133-138.
23. Craft JE, Fischer DK, Shimamato GT, et al (1986) Antigen of Borrelia burgdorferi recognized during Lyme disease. J Clin Invest 78:934-939.
24. Wooley PH, Luthra HS, Lafuse WP, et al (t985) Type II colla- gen-induced arthritis in mice. J Immunol 134(4):2366-2374.
25. Statsny P (1978) Association of the B-cell alloantigen DRw4 with rheumatoid arthritis. N Engl J Med 298:869-871.
26. Wernet D (1982) Genetic regulation of the antibody response to H-2D b alloantigens in mice. Immunogenetics 16:71-81.
27. Schaible UE, Kramer MD, Wallich R, et al (1991) Experimental Borrelia burgdorferi infection in inbred mouse strains: Antibody response and association of H-2 genes with resistance and suscep- tibility to development of arthritis. Eur J Immunol 21:2397-2405.





























