Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Jan. 2003, p. 60–70 Vol. 185, No. 10021-9193/03/$08.00�0 DOI: 10.1128/JB.185.1.60–70.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
The Streptococcus pneumoniae cia Regulon: CiaR Target Sitesand Transcription Profile Analysis
Thorsten Mascher,1† Dorothea Zahner,2‡ Michelle Merai,1 Nadege Balmelle,1§Antoine B. de Saizieu,1§ and Regine Hakenbeck1*
Department of Microbiology, University of Kaiserslautern, D-67663 Kaiserslautern, Germany,1
and F. Hoffmann-La Roche Ltd., CH-4070 Basel, Switzerland2
Received 28 May 2002/Accepted 12 September 2002
The ciaR-ciaH system is one of 13 two-component signal-transducing systems of the human pathogenStreptococcus pneumoniae. Mutations in the histidine protein kinase CiaH confer increased resistance tobeta-lactam antibiotics and interfere with the development of genetic competence. In order to identify the genescontrolled by the cia system, the cia regulon, DNA fragments targeted by the response regulator CiaR wereisolated from restricted chromosomal DNA using the solid-phase DNA binding assay and analyzed by hybrid-ization to an oligonucleotide microarray representing the S. pneumoniae genome. A set of 18 chromosomalregions containing 26 CiaR target sites were detected and proposed to represent the minimal cia regulon. Theputative CiaR target loci included genes important for the synthesis and modification of cell wall polymers,peptide pheromone and bacteriocin production, and the htrA-spo0J region. In addition, the transcriptionprofile of cia loss-of-function mutants and those with an apparent activated cia system representing the off andon states of the regulatory system were analyzed. The transcript analysis confirmed the cia-dependent expres-sion of seven putative target loci and revealed three additional cia-regulated loci. Five putative target regionswere silent under all conditions, and for the remaining three regions, no cia-dependent expression could bedetected. Furthermore, the competence regulon, including the comCDE operon required for induction ofcompetence, was completely repressed by the cia system.
Bacterial two-component signal-transducing systems (TC-STS) mediate adaptive responses to environmental signals.They typically consist of two modular proteins: a histidineprotein kinase that acts as a sensor for external stimuli and acytoplasmic response regulator that translates the signal into acellular response by changing the expression profiles of tar-geted genes (40, 51). The TCSTS ciaRH was the first of 13 suchsystems identified in the human pathogen Streptococcus pneu-moniae (19). Two membrane-spanning regions in CiaH areproposed to separate the N-terminal external sensor domainfrom the cytoplasmic kinase domain. The response regulatorCiaR contains a typical DNA binding domain characteristic ofthis subclass of regulators (35). The two genes are arranged inan operon with a 8-bp overlap (19).
Mutations in the histidine protein kinase ciaH conferredincreased resistance to beta-lactam antibiotics, revealing anovel pathway for resistance development in this organism andsuggesting that ciaR may control genes that are involved in thebiochemistry of the bacterial cell wall (19, 60). Other pheno-types in cia mutants have since been described, such as growthdefects associated with the tendency for early lysis (17, 22, 27;
T. Mascher, M. Merai, and D. Zahner, unpublished results)and attenuation of virulence (53), again indicating that the ciasystem plays an important role in the maintenance of theoverall integrity of the cell wall.
Curiously, ciaH mutants were affected in the development ofgenetic competence as well (19, 20), an effect which has sincebeen confirmed in several recent publications (14, 34, 58). ThecomCDE system, which is responsible for the induction ofgenetic competence, is a complex regulatory network: the CSPpeptide, the processed secreted product of comC, is recognizedby the histidine protein kinase ComD, and the response regu-lator ComE is responsible for autoinduction of the comCDEoperon and other early genes, among which are the transcrip-tional activators ComX1 and ComX2, which in turn induce thelate competence genes (43). Using crude preparations of so-called competence factor, competence could not be induced innoncompetent ciaH mutants; however, after the identificationof the competence factor as a small unmodified peptide, CSP(24), chemically synthesized CSP could be used in higher con-centrations and complemented this defect in all ciaH mutantsanalyzed (60). This suggested that either the export, the sta-bility, or the sensing of CSP is affected in cia mutants.
What are the genes controlled by the ciaRH system, andwhat is the link between the two TCSTS, ciaRH and comDE?It has been suggested that the cia system is related to thebiosynthesis of cell wall components at steps prior to biosyn-thetic functions of penicillin binding proteins functioning dur-ing the final assembly of the peptidoglycan (22). However,neither the molecular nature of the signal sensed by the CiaHkinase nor cia-regulated genes are known. Genes directly con-trolled by the response regulator CiaR should be located in the
* Corresponding author. Mailing address: Department of Microbi-ology, University of Kaiserslautern, Paul Ehrlich Straße 23, D-67663Kaiserslautern, Germany. Phone: 49-631-2052353. Fax: 49-631-2053799. E-mail: [email protected].
† Present address: Department of Microbiology, Cornell University,Ithaca, NY 14853-8101.
‡ Present address: Department of Microbial Pathogenesis and Vac-cine Research, German Research Centre for Biotechnology, D-38124Braunschweig, Germany.
§ Present address: Roche Vitamins AG, CH-4070 Basel, Switzer-land.
60
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

vicinity of CiaR binding sites. In order to identify such CiaRtarget regions, we have used a DNA binding assay based on amethod established for the Escherichia coli DNA binding pro-tein DnaA (48). In the solid-phase DNA binding (SPDB) as-say, an overexpressed, biotinylated fusion derivative of a DNAbinding protein is purified via its interaction with streptavidin-coated magnetic beads, and after incubation with DNA frag-ments, those that show specific interaction with the protein canbe isolated and analyzed further. We have modified this pro-cedure for the response regulator CiaR and used an oligonu-cleotide microarray for identifying the set of DNA fragmentsobtained from restricted chromosomal DNA of S. pneumoniae.
The results are in agreement with the hypothesis that severalcia-regulated genes are involved in the biochemical makeup ofthe cell envelope, including modifications of peptidoglycan,surface-exposed proteins, and a variety of transport systems.Transcript analysis of cia mutants in comparison with wild-typecells was used to confirm the cia-dependent transcription ofsuch loci. In addition to cia-dependent expression of some ofthe putative target loci, repression of the entire competenceregulon by the cia system could be documented.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. S. pneumoniae strainswere routinely grown in Todd-Hewitt broth (THB; Difco) or in the semidefinedC medium (26) supplemented with 0.2% yeast extract (Difco) at 37°C withoutaeration. S. pneumoniae R6 is a transformable, unencapsulated laboratory strainderived from the type 2 strain D39 (50). The genomic sequence of the encap-sulated type 4 strain KNR.7/87 (1, 52) served as the basis for the oligonucleotidemicroarray used for this study. R6ciaHC306 is a transformant obtained with theciaH gene of the mutant C306 which carries the mutation T230�P (19). E. coliWM1704 (46) is a pMC9-cured derivative of strain Y1089 (57). E. coli wasroutinely grown aerobically in Luria-Bertani medium at 37°C; exceptions arestated in the text. pBEX5BA (46) and its derivatives were selected in E. coli with100 �g of ampicillin/ml.
Construction of lacZ reporter strains and �-galactosidase assay. To monitorlacZ expression, a �-galactosidase-negative derivative of the S. pneumoniae R6strain (R6bga::erm) was used (59). The plasmid pXF520 carrying part of thecomC leader peptide and upstream sequences, with a fusion to the lacZ reportergene at the 15th codon of the comC gene, has been described (41). R6 Cmr
transformants were isolated, and a blue colony was used for the introduction ofcia mutations. Insertion-duplication mutants in ciaR produced dark-blue colo-nies, whereas introduction of the ciaHC306 allele resulted in white colonies; nomixed populations were observed for the cia mutants as has been described forstrains grown under competence-preventing conditions (58). Chromosomal fu-sion of the lacZ� gene under the control of the lic and dlt promoter regions wasperformed using the lacZ reporter plasmid pEVP3 (11). The dlt promoter regionwas amplified by PCR using the oligonucleotide pair 5�-AAAGTGTAGATCTTCAGGAAACAGTAGAGG and 5�-GCGGCATTCTAGACAGGATAGCTAGGCTG, covering the region from position �704 to �62 in respect to the A1TGstart codon; the lic promoter region was obtained with the primers 5�-CCAATGCATCGGTCCAAGACACGCGCG and 5�-CGGGATCCGCGTGTGCCAGTTCCACC, covering the region �447 to �50. The fragments were transcrip-tionally fused to the lacZ� gene of pEVP3, and the resulting plasmids wereintegrated into the chromosome upstream of the operons by homologous re-combination, resulting in strains R6bga::erm,dltA::pEVP3 and R6bga::erm,licA::pEVP3. Correct integration was confirmed by PCR using pairs of oligonu-cleotides priming within pEVP3 and in the respective promoter region. The�-galactosidase assay was based on the classical procedure (36) and was per-formed as described previously (41).
Construction of ciaR loss-of-function mutants. The spectinomycin resistancegene aad9 from pDL278 (29) was used for disruption of the cia genes. The ciaRgene was cloned into the vector pCR2.1 (Invitrogen). The aad9 gene was am-plified with the oligonucleotide pair 5�-gctagcATCGATTTTCGTTCGTGAATAC and 5�-gctagcCCAATTAGAATGAATATTCCC containing flanking NheIsites (lowercase). The gene was then recovered by NheI digestion and ligated intothe single AvrII site of pCR2.1 carrying ciaR. The resulting construct, ciaR::aad9,
was amplified using the oligonucleotide pair 5�-GCTTTGTTATCCTTCCTGCTC and 5�-cccaagcttGAGAAGGGCCTGAATCCGCAT (the lowercase lettersindicate the HindIII site) and transformed into R6 using 80 mg of spectinomy-cin/ml for selection. Transformation of S. pneumoniae strains was performedessentially according to published procedures, with 30 min of incubation in thepresence of DNA at 30°C followed by a 2-h phenotypic-expression period at 37°C(19). All constructs were confirmed by PCR using pairs of oligonucleotide prim-ers flanking the insertion. The resulting mutant was named R6-R1 (ciaR::aad9).
Construction and isolation of bCiaR. The vector pBEX5BA (46) was used forthe construction of a biotinylated derivative of CiaR (bCiaR). The vector con-tains the gene encoding the 12.5-kDa subunit of the transcarboxylase complexfrom Propionibacterium shermanii directly upstream from the multiple cloningsite. This subunit harbors a target sequence for the E. coli enzyme biotin-apoprotein ligase, which transfers a biotin residue to this peptide (38). Thecomplete ciaR gene of S. pneumoniae was amplified by PCR (nucleotides 219 to889 according to GenBank accession number X77249) using the oligonucleotidepair 5�-cgggatccATAAAAATCTTATTGGTTGAGG and 5�-cccaagcttACTGAACATCTTTTAAAAGAT (terminal restriction sites [lowercase] were included).The fragment obtained was translationally fused with its 5� end to the 3� end ofthe gene coding for the 12.5-kDa subunit of the transcarboxylase complex of P.shermanii, leading to the fusion gene bciaR. The resulting plasmid, pDZ9, wastransformed in E. coli WM1704. The DNA sequence of bciaR was confirmed byDNA sequencing. E. coli WM1704(pDZ9) was grown in Luria-Bertani mediumsupplemented with 2 �M biotin in the presence of 100 �g of ampicillin/ml at28°C to prevent the formation of inclusion bodies. Expression of bCiaR wasinduced with IPTG (isopropyl-�-D-thiogalactopyranoside; 100 �M final concen-tration) at an optical density at 560 nm of 0.7 for 2.5 h. The expression of theCiaR derivative was confirmed by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis and Coomassie staining of the proteins and on Western blotsusing an anti-CiaR specific rabbit antiserum; biotinylation of bCiaR was con-firmed by near-Western blotting with a streptavidin-alkaline phosphatase conju-gate.
Crude preparation of bCiaR. After induction with IPTG for 2.5 h as describedabove, E. coli WM1704(pDZ9) cells were harvested by centrifugation and resus-pended in 1/20 of the culture volume in 20 mM Tris-HCl (pH 7.3)–10 mMMgCl2–100 mM KCl; 500-�l aliquots of this suspension were frozen in liquidnitrogen and stored at �80°C. Immediately before use, the aliquots were thawedon ice and the cells were disintegrated on ice by sonication (three intervals of 1min each). Intact cells and cell envelope fragments were removed by centrifu-gation (Heraeus Biofuge 28RS; 24,000 rpm [�40,000 � g]; 15 min; 4°C), and thesupernatant containing bCiaR was used directly in the SPDB assay.
DNA sequence analysis. Nucleotide sequencing was performed using the ABIPrism dRhodamine Terminator Cycle Sequencing Ready Reaction kit (PerkinElmer-ABI).
Preparation of chromosomal restriction fragments. Chromosomal DNA of S.pneumoniae KNR.7/87 was prepared as previously described (23) and digestedwith either HindIII, RsaI, or Sau3A1 in a 200-�l final volume. The enzymes wereheat inactivated, and salt was removed by dialysis on a membrane filter (type VS;0.025-�m pore size; Millipore, Eschborn, Germany).
SPDB assay. The SPDB assay was performed according to published proce-dures with modifications (48). Strepavidin-coated magnetic beads (20 �l) (Dyna-beads M-280; Dynal AS, Oslo, Norway) were washed four times with 40 �l of 20mM Tris-HCl (pH 7.3)–100 mM KCl (BB buffer) supplemented with completeprotease inhibitor mix (PIM) (Roche, Mannheim, Germany) as described by themanufacturer and finally resuspended in 40 �l of BB-PIM. The beads were mixedwith 80 �l of crude extract of WM1704(pDZ9) and incubated for 5 min at roomtemperature. No phosphorylation reaction was performed for bCiaR. The pro-tein-coated beads were vigorously washed three times with 40 �l of BB-PIMsupplemented with 0.4% IGEPAL CA-630 (Sigma-Aldrich, Munich, Germany)and five times with 40 �l of BB. Finally, the beads were resuspended in 80 �l ofBB, mixed with 20 �l of restricted chromosomal DNA of S. pneumoniae, andincubated at room temperature for 10 min. Nonspecifically bound DNA wasremoved by washing the beads with 40 �l of BB (three times). DNA fragmentsspecifically interacting with bCiaR were eluted after incubation of the coatedbeads with 1.3 M NaCl (40 �l) for 5 min. The eluted DNA was precipitated withethanol-glycogen and collected by centrifugation before further use. The speci-ficity of binding was verified using 10 ng of the PCR-amplified putative targetfragment to be tested together with a mixture of unrelated DNA fragments (125ng of competitor DNA [DNA molecular weight marker X; Roche]) in the SPDBassay as described above. The eluted DNA fragments were separated on 1%agarose gels and visualized on a Fluorimager (Molecular Dynamics) after beingstained with SYBRgreen (Molecular Probes Inc., Eugene, Oreg.).
VOL. 185, 2003 STREPTOCOCCUS PNEUMONIAE cia REGULON 61
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

DNA fragmentation and labeling. The detection limit for DNA fragments usedfor hybridization on the microarray was tested for five probe sets and estimatedto be �5 ng per kb. SPDB-derived DNA fragments from 5 to 7 assays (RsaI- andSau3A-restricted chromosomal DNA) or 20 assays (HindIII-restricted DNA)were combined, and the amount finally used was estimated to represent between10 and 20 ng per kb of DNA fragmen per assay. The pooled DNA fragmentswere diluted in 200 �l of water and sonicated on ice three times for 1 min eachtime, resulting in fragments of an average size of 300 to 500 bp. Using a relativelysmall amount of hybridizing DNA of low complexity required adjustment of thefragmentation conditions used previously (21). After ethanol precipitation, theDNA was resuspended in 89 �l of water and further fragmented by partialDNase treatment for 10 s at 37°C using 0.005 U of DNase I in RQ1 buffer(Promega) in a final volume of 100 �l. The reaction was stopped by the additionof 3 �l of proteinase K (20 mg/ml), and incubation continued for another 3 minat 37°C. The average size of the resulting fragments decreased during this periodto between 50 and 100 bp. The DNA was ethanol precipitated, resuspended in 85�l of H2O, and quantified spectrophotometrically. DNA labeling was performedwith biotin-labeled ddATP incorporated at the 3� end of the fragmented DNAusing Terminal-deoxy-Transferase (Roche Molecular Biochemicals) as describedpreviously (21).
RNA extraction and cDNA labeling. The preparation of RNA was done aspreviously described (13). In brief, 30-ml cultures grown in the absence ofantibiotics were harvested in late exponential growth phase by centrifugation,and the cell pellet was frozen immediately in liquid nitrogen. Each experimentwas done in duplicate with two different cultures with all preparation stepsperformed independently. RNA was isolated using the hot-phenol method (33)and finally purified through Qiagen RNeasy columns as instructed by the man-ufacturer in order to remove small RNA molecules. The hybridization target wassynthesized by an optimized reverse-transcription reaction from total RNA in thepresence of random hexanucleotides and biotin-labeled dATP during overnightincubation at 37°C in a 100-�l volume according to the published procedure (13).
Microarray design. The oligonucleotide microarray used in this study repre-sents the two genomes of S. pneumoniae and Haemophilus influenzae. It wascustom designed by Affymetrix (Santa Clara, Calif.), and details have beendescribed previously (13, 21). In summary, the S. pneumoniae genome of strainKNR.7/87, represented by a total of 2,291 probe sets covering 1,968 predictedopen reading frames and 323 intergenic regions larger than 200 bp, was consid-ered in the final analysis. The oligonucleotide probe selection (25-mers) andarray fabrication were performed by Affymetrix according to published proce-dures (31, 56). In general, each gene represented on the ROEZ06a microarrayhas 25 probe pairs, with at least 20 probe pairs for very short genes. A probe pairconsists of a perfect-match probe and a mismatch probe that is identical exceptfor a single base change in the central position. The position of the oligonucle-otide on each gene is determined by sequence uniqueness criteria based onempirical rules for the selection of oligonucleotides likely to hybridize with highspecificity and sensitivity (31).
Hybridization and staining procedures. Hybridization and staining of DNAfragments were carried out essentially as described previously (13, 21). Thesolutions contained 100 mM MES (N-morpholinoethane-sulfonic acid), 1 MNa�, 20 mM EDTA, and 0.01% Tween 20. In addition, unlabeled fragmentedyeast RNA (3 mg/ml; Roche Molecular Biochemicals), acetylated bovine serumalbumin (1.5 mg/ml), and TOP Block (20 mg/ml; Juro, Lucerne, Switzerland)were used. In brief, the microarrays were washed and prehybridized for 10 minat 40°C. Biotin-labeled DNA fragments were hybridized overnight at 40°C on arotisserie. After removal of the hybridization mixture, the arrays were rinsed andwashed with 6� SSPE (1� SSPE is 0.18 M NaCl, 10 mM NaH2PO4, and 1 mMEDTA [pH 7.7])–0.01% Tween 20 (40°C for 20 min). A stringent wash was thenperformed with 0.5� SSPE–0.01% Tween 20 for 15 min at 45°C. The hybridizedDNA was labeled with streptavidin-phycoerythrin conjugate (3 �g/ml; MolecularProbes) and acetylated bovine serum albumin (2 mg/ml) in 6� SSPE–0.01%Tween 20 for 10 min at 40°C. Further signal amplification was also performedwith a biotinylated anti-streptavidin antibody and subsequent staining withstreptavidin-phycoerythrin conjugate. A final washing step was performed in 6�SSPE–0.01% Tween 20 for 10 min at 40°C prior to scanning. All hybridizationswith the microarray were performed at least in duplicate.
Data processing. The microarrays were scanned at 570 nm (3-�m resolution)with a gene chip scanner (Affymetrix) and analyzed as previously described (31).The signal intensity for each gene is calculated as the average intensity difference,represented by the following expression: (PM MM)/number of probe pairs,where PM and MM denote perfect-match and mismatch probes. Due to thebackground caused by DNA fragments present in the SPDB assay that are notspecifically associated with bCiaR, the minimum average intensity difference foreach gene was set to 100, which corresponds to the noise; for the experiments
using cDNA, the minimum average intensity difference was set to 20. Probe setswith an average intensity difference of at least fivefold above background wereextracted and assembled according to the available genome sequence (http://www.tigr.org).
Computational analysis. Protein homology and genetic analyses of open read-ing frames were performed using the ORFfinder and BLAST algorithms (http://www.ncbi.nlm.nih.gov) provided by the National Center for Biotechnology In-formation.
Nucleotide sequence accession numbers. GenBank accession numbers are asfollows: S. pneumoniae genome KNR.7/87 (TIGR4), AE005672; S. pneumoniaeR6 genome, AE007317 (25, 52).
RESULTS
Isolation of CiaR DNA target regions. In the SPDB assay forisolation of CiaR DNA target fragments, the derivative bCiaRwas expressed in E. coli, where the CiaR DNA binding domainremains free at the C terminus and the N terminus has beenfused to a peptide that becomes biotinylated in E. coli (seeMaterials and Methods for details). The purification of bCiaRwas achieved in one step, incubating a crude E. coli cell lysatecontaining the overproduced, biotinylated bCiaR with strepta-vidin-coated magnetic beads. The carefully washed protein-loaded beads were then incubated with restricted pneumococ-cal chromosomal DNA, and DNA fragments that remainedassociated with the beads at elevated salt concentrations wereassumed to interact specifically with CiaR. DNA of the S.pneumoniae strain KNR.7/87 was used in the binding assay,since the oligonucleotide microarray was based on the genomicsequence of that strain, which differs from the R6 strain byalmost 10% of the genes (21). The ciaR gene of KNR.7/87 isidentical to that of the R6 strain and is also highly conservedwithin the pneumococcal population (our unpublished results).Therefore, the target sequences could also be expected to beconserved. Since it is possible that a CiaR binding site is lost ifit is located close to a restriction site, independent assays wereperformed using three distinct restriction digests. The enzymesRsaI and Sau3A produced relatively small fragments (5 kb),whereas those generated with HindIII ranged from 1 to ap-proximately 20 kb (Fig. 1). In all three cases, a reproduciblepattern of specific fragments eluted from the bCiaR-coatedbeads with between 11 and 17 bands clearly identifiable (Fig.1). The size of the targeted fragments ranged from 1 to �15 kbfor HindIII and from 4.5 kb and smaller for Sau3A and RsaI.
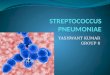
Characterization of CiaR target regions. After hybridiza-tion to the oligonucleotide microarray, the DNA fragmentsequences corresponding to the fluorescent signals resultingfrom each of the three restriction digests were aligned accord-ing to the published KNR.7/87 genome sequence (52). Positivesignals that covered exclusively repetitive elements, such astransposons and insertion sequence elements, were not takeninto account. The 41 regions detected with at least two or allthree sets of restriction fragments were investigated in detail.Using the published genome of strain KNR.7/87 as the basisfor a restriction map, the region of interest was examined tosee whether the fluorescent signals coincided with the pre-dicted restriction fragments. As an example, the region thatincludes the manLMN operon encoding an orthologue of themannose-specific PTS system previously described in Strepto-coccus salivarius (32) is shown in Fig. 2. The signals obtainedwith the HindIII and RsaI restriction fragments were located inthe man genes, whereas the Sau3A1-derived signal was located
62 MASCHER ET AL. J. BACTERIOL.
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

upstream of the man operon. One should keep in mind that theprobe sets on the microarray do not necessarily cover thefull-length gene, and the promoter region of the man operon isnot represented on the microarray. The restriction map forHindIII, Sau3A1, and RsaI deduced from the KNR.7/87 ge-nome shows that the fluorescent signals match the predictedrestriction fragments and that the only area overlapping in allthree fragments covers the entire intergenic region locatedupstream of the manL gene. It was thus concluded that a CiaRbinding site is located in the man operon regions, suggestingthat manLMN may be one of the cia-regulated gene loci.
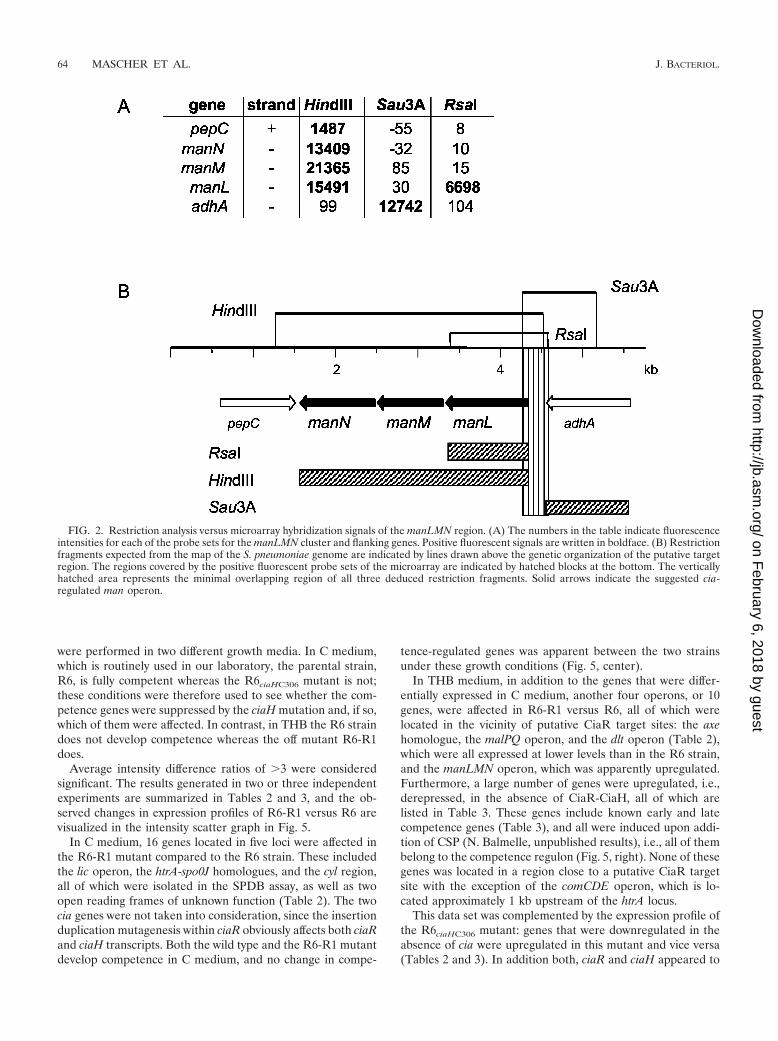
Specificity of binding was confirmed for several fragments bytesting them individually in the SPDB assay in the presence ofcompetitor DNA as shown in Fig. 3 for the region upstream ofthe uppS gene. The Sau3A fragment covering the region up-stream of the uppS gene and the gene downstream of ruvB (seeFig. 5) was PCR amplified and shown to contain a unique DraIsite. Only the smaller 266-bp DraI fragment immediately up-stream of uppS was retained by bCiaR, whereas neither thelarger DraI fragment nor the competitor DNA remained
bound (Fig. 3). Other regions tested for CiaR binding includedthose identified on fragments 14 and 15, and in both casesbinding specificity could be confirmed in the SPDB assay (notshown).
Altogether, 26 of the 41 analyzed fragments with positivehybridization signals for all three restriction digests containedmatching restriction sites. The final list of 18 chromosomallocations, 7 of which contain more than one CiaR binding site,is shown in Table 1, and the genetic organization of a 10-kbregion covering these sites is presented in Fig. 4. For mostregions, the functions of the predicted cia target genes wereknown or could be inferred from significant sequence ho-mologies (identity in all cases was �40% for the completecoding sequence [Table 1]). Many of the genes, some of whichare essential, relate to cell wall polysaccharide metabolism.There are systems required for sugar uptake and utilization(manLMN and malPQ), glycosyltransferases of unknown spec-ificity (region I), and modifying enzymes, such as a homo-logue of a xylan esterase, Axe. The gene products of deoAC areinvolved in ribose metabolism and are important for maintain-ing the nucleotide pool required for activation of sugar com-pounds in the cytoplasm. The undecaprenyl pyrophosphatesynthetase UppS is a key enzyme required for translocation ofcell wall polysaccharide precursors through the cytoplasmicmembrane. FibB (also called MurN) is involved in interpeptidebridge formation in the murein. The lic locus is involved inphosphoryl choline metabolism, with choline being a pneumo-coccus-specific component of the wall teichoic acid and lipo-teichoic acid. The dlt cluster is responsible for D alanylation oflipoteichoic acid in Streptococcus mutans. There are severaltransport systems (regions 8 and 15) and gene clusters involvedin the production of biologically active peptides, such as thebacteriocin cluster spi (region 5) and a cluster encoding homo-logues of the Enterococcus faecalis cytolysin locus cyl (region14). HtrA functions as an extracellular protease responsible forthe degradation of abnormally folded export proteins in theclosely related organism Lactococcus lactis. None of these geneproducts has been related to genetic competence so far, and noobvious link to competence could be deduced from their pu-tative functions.
cia-dependent gene expression. In order to prove that theregions identified above include cia-regulated gene loci, thetranscript profiles of cia mutants that had been generated inthe laboratory strain R6 were compared to that of the parentalR6 strain. In mutant R6-R1, the ciaR gene was disrupted byinsertion of a spectinomycin resistance cassette (see Materialsand Methods). This mutant produces neither a functionalCiaR nor CiaH, since both genes are transcribed as an operonand CiaH-specific antibodies failed to detect the protein (17,60).
The second mutant, R6ciaHC306, contains a point mutation inCiaH, T230�P, close to the conserved His226 (19); it wasobtained by transformation of the ciaHC306 allele into the R6strain. It has been suggested that this mutation affects thephosphatase activity of CiaH, resulting in overphosphorylationof CiaR (19). Thus, both mutants should allow a comparison ofthe expression profile of a cia-off situation (R6-R1) to that ofa cia-on situation (R6ciaHC306).
In order to also be able to test the effects of cia mutations onthe expression of the competence regulon, the experiments
FIG. 1. Visualization of CiaR target fragments obtained with RsaI-,Sau3A-, and HindIII-restricted chromosomal DNA of S. pneumoniae.DNA fragments obtained after specific elution from bCiaR-coatedmagnetic beads were separated on a 1% agarose gel, stained withSYBRgreen, and visualized with the help of a fluoroimager. Threedifferent restriction digests were used: Sau3A1, HindIII, and RsaI.Lanes: a, total restricted chromosomal DNA; b, DNA fragments ob-tained after specific elution in the SPDB assay; c, high-mass DNAladder marker; d, size ladder marker.
VOL. 185, 2003 STREPTOCOCCUS PNEUMONIAE cia REGULON 63
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
were performed in two different growth media. In C medium,which is routinely used in our laboratory, the parental strain,R6, is fully competent whereas the R6ciaHC306 mutant is not;these conditions were therefore used to see whether the com-petence genes were suppressed by the ciaH mutation and, if so,which of them were affected. In contrast, in THB the R6 straindoes not develop competence whereas the off mutant R6-R1does.
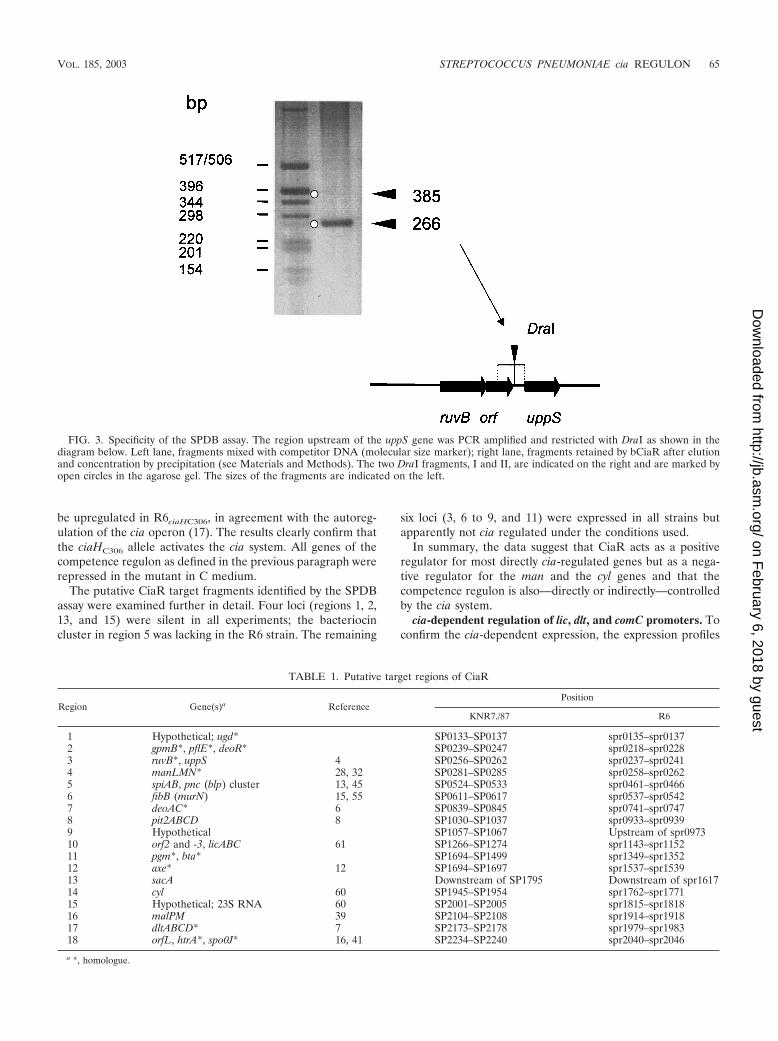
Average intensity difference ratios of �3 were consideredsignificant. The results generated in two or three independentexperiments are summarized in Tables 2 and 3, and the ob-served changes in expression profiles of R6-R1 versus R6 arevisualized in the intensity scatter graph in Fig. 5.
In C medium, 16 genes located in five loci were affected inthe R6-R1 mutant compared to the R6 strain. These includedthe lic operon, the htrA-spo0J homologues, and the cyl region,all of which were isolated in the SPDB assay, as well as twoopen reading frames of unknown function (Table 2). The twocia genes were not taken into consideration, since the insertionduplication mutagenesis within ciaR obviously affects both ciaRand ciaH transcripts. Both the wild type and the R6-R1 mutantdevelop competence in C medium, and no change in compe-
tence-regulated genes was apparent between the two strainsunder these growth conditions (Fig. 5, center).
In THB medium, in addition to the genes that were differ-entially expressed in C medium, another four operons, or 10genes, were affected in R6-R1 versus R6, all of which werelocated in the vicinity of putative CiaR target sites: the axehomologue, the malPQ operon, and the dlt operon (Table 2),which were all expressed at lower levels than in the R6 strain,and the manLMN operon, which was apparently upregulated.Furthermore, a large number of genes were upregulated, i.e.,derepressed, in the absence of CiaR-CiaH, all of which arelisted in Table 3. These genes include known early and latecompetence genes (Table 3), and all were induced upon addi-tion of CSP (N. Balmelle, unpublished results), i.e., all of thembelong to the competence regulon (Fig. 5, right). None of thesegenes was located in a region close to a putative CiaR targetsite with the exception of the comCDE operon, which is lo-cated approximately 1 kb upstream of the htrA locus.
This data set was complemented by the expression profile ofthe R6ciaHC306 mutant: genes that were downregulated in theabsence of cia were upregulated in this mutant and vice versa(Tables 2 and 3). In addition both, ciaR and ciaH appeared to
FIG. 2. Restriction analysis versus microarray hybridization signals of the manLMN region. (A) The numbers in the table indicate fluorescenceintensities for each of the probe sets for the manLMN cluster and flanking genes. Positive fluorescent signals are written in boldface. (B) Restrictionfragments expected from the map of the S. pneumoniae genome are indicated by lines drawn above the genetic organization of the putative targetregion. The regions covered by the positive fluorescent probe sets of the microarray are indicated by hatched blocks at the bottom. The verticallyhatched area represents the minimal overlapping region of all three deduced restriction fragments. Solid arrows indicate the suggested cia-regulated man operon.
64 MASCHER ET AL. J. BACTERIOL.
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
be upregulated in R6ciaHC306, in agreement with the autoreg-ulation of the cia operon (17). The results clearly confirm thatthe ciaHC306 allele activates the cia system. All genes of thecompetence regulon as defined in the previous paragraph wererepressed in the mutant in C medium.
The putative CiaR target fragments identified by the SPDBassay were examined further in detail. Four loci (regions 1, 2,13, and 15) were silent in all experiments; the bacteriocincluster in region 5 was lacking in the R6 strain. The remaining
six loci (3, 6 to 9, and 11) were expressed in all strains butapparently not cia regulated under the conditions used.
In summary, the data suggest that CiaR acts as a positiveregulator for most directly cia-regulated genes but as a nega-tive regulator for the man and the cyl genes and that thecompetence regulon is also—directly or indirectly—controlledby the cia system.
cia-dependent regulation of lic, dlt, and comC promoters. Toconfirm the cia-dependent expression, the expression profiles
FIG. 3. Specificity of the SPDB assay. The region upstream of the uppS gene was PCR amplified and restricted with DraI as shown in thediagram below. Left lane, fragments mixed with competitor DNA (molecular size marker); right lane, fragments retained by bCiaR after elutionand concentration by precipitation (see Materials and Methods). The two DraI fragments, I and II, are indicated on the right and are marked byopen circles in the agarose gel. The sizes of the fragments are indicated on the left.
TABLE 1. Putative target regions of CiaR
Region Gene(s)a ReferencePosition
KNR7./87 R6
1 Hypothetical; ugd* SP0133–SP0137 spr0135–spr01372 gpmB*, pflE*, deoR* SP0239–SP0247 spr0218–spr02283 ruvB*, uppS 4 SP0256–SP0262 spr0237–spr02414 manLMN* 28, 32 SP0281–SP0285 spr0258–spr02625 spiAB, pnc (blp) cluster 13, 45 SP0524–SP0533 spr0461–spr04666 fibB (murN) 15, 55 SP0611–SP0617 spr0537–spr05427 deoAC* 6 SP0839–SP0845 spr0741–spr07478 pit2ABCD 8 SP1030–SP1037 spr0933–spr09399 Hypothetical SP1057–SP1067 Upstream of spr097310 orf2 and -3, licABC 61 SP1266–SP1274 spr1143–spr115211 pgm*, bta* SP1694–SP1499 spr1349–spr135212 axe* 12 SP1694–SP1697 spr1537–spr153913 sacA Downstream of SP1795 Downstream of spr161714 cyl 60 SP1945–SP1954 spr1762–spr177115 Hypothetical; 23S RNA 60 SP2001–SP2005 spr1815–spr181816 malPM 39 SP2104–SP2108 spr1914–spr191817 dltABCD* 7 SP2173–SP2178 spr1979–spr198318 orfL, htrA*, spo0J* 16, 41 SP2234–SP2240 spr2040–spr2046
a *, homologue.
VOL. 185, 2003 STREPTOCOCCUS PNEUMONIAE cia REGULON 65
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

of the lic and dlt promoters were examined in detail during thegrowth cycle in the wild type versus cia mutants (R6-R1 andciaHC306). These two promoters were chosen because both areassumed to encode functions related to teichoic acid chemistry,
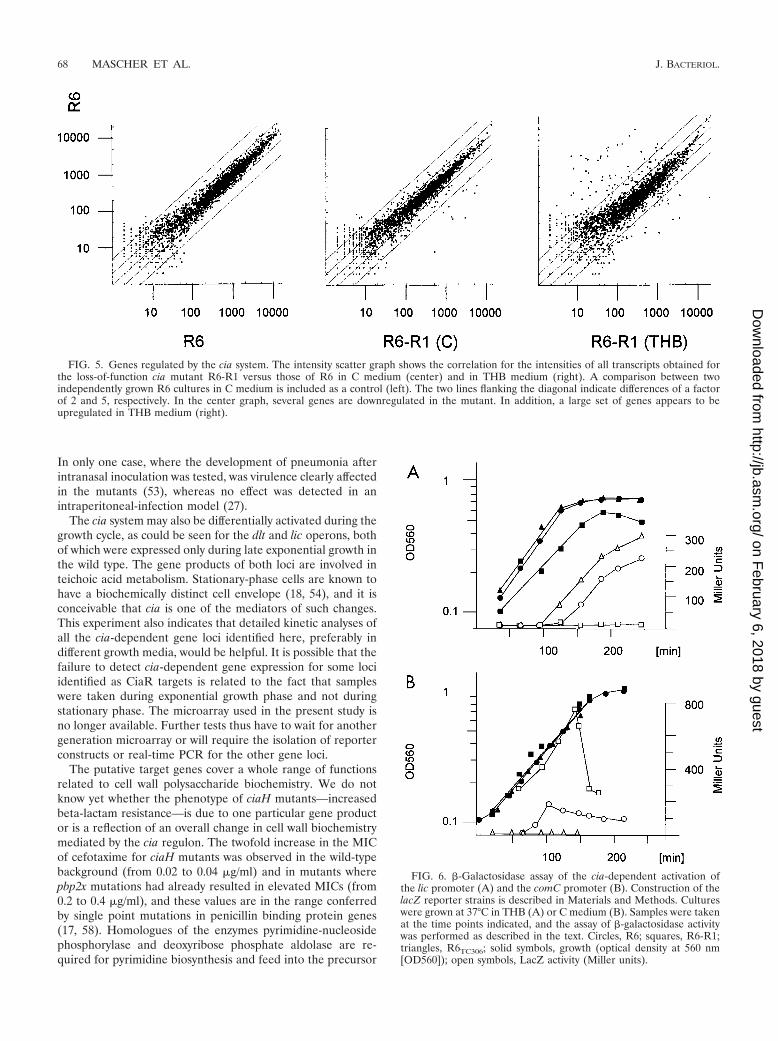
using a lacZ reporter system as described previously (11, 59).The expression of lacZ production was monitored throughoutthe growth cycle. The lic promoter was induced at the end ofthe exponential growth phase in the wild-type R6 strain (Fig.6A), whereas the loss-of-function mutants showed no detect-able expression. In contrast, the R6ciaHC306 mutant showedexpression at earlier time points and at higher levels than theR6 strain. Similar results were obtained for dlt constructs (notshown).
The comC promoter was also investigated in detail in theR6-R1 background, as well as in the ciaHC306 mutant, underconditions where R6 develops competence (Fig. 6B). The ex-pression of comC::lacZ was followed throughout the growthcycle. The R6 strain develops a broad peak of competenceunder these conditions, whereas the ciaHC306 mutant remainscompletely noncompetent throughout the growth cycle (11). Inthe present experiment, transformation efficiency was con-trolled at two cell densities at which competence in wild-typeR6 is high. In the R6 background, comC-dependent LacZactivity remained low early during exponential growth andincreased rapidly, consistent with the appearance of compe-tence. Maximal expression was reached after approximately 20min; it then decreased and remained at an elevated level untilthe onset of the stationary phase. In the ciaHC306 mutant thatremained completely noncompetent, LacZ activity was not de-tectable throughout the growth cycle. In contrast, the loss-of-function mutant R6-R1, which was as competent as the R6strain, showed a very high level of lacZ expression even at lowcell density, reaching a maximum during late exponentialgrowth.
DISCUSSION
The regulon of the TCSTS ciaR-ciaH was analyzed by acombination of two experimental approaches: DNA targetfragments of the response regulator CiaR were isolated in asolid-phase DNA binding assay, and the gene expression pat-
FIG. 4. Putative CiaR target regions. The putative CiaR targetregions are listed according to their positions on the S. pneumoniaegenome (see Table 1 for details). The regions representing the set ofreactive probe sets that are present in the different restricted DNAsamples are represented by bars above the genetic organization. Pu-tative cia target genes are represented by solid arrows. The boxedsegments indicate cia-dependently regulated genes (see Table 2).
TABLE 2. cia-regulated gene loci
Clustera Geneb SPDBc
Gene expressioncompared to R6
R6-R1 R6ciaHC306
4 man (3) � More Less10 lic (5) � Less Samed
12 axe (1) � Less More14 cyl (7) � More Same16 mal (2) � Less More17 dlt (4) � Less More18 htrA-spo0J (2) � Less More
1 cia (2) � NDe More2 spr0782 (SP0879) � Less More3 spr0931 (SP1027) � Less MoreMany f Competence regulon � More Less
a Number refers to nomenclature used in Table 1.b Number in parentheses are numbers of genes in the operons.c Gene identified (�) or not identified (�) an DNA fragments isolated in
SPDB assay.d Identical within the threefold threshold (see the text).e ND, not determined; due to the insertion in the ciaR gene, the negative value
obtained in R6-R1 is not meaningful.f See table 3.
66 MASCHER ET AL. J. BACTERIOL.
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

tern of cia mutants representing off and on stages of the ciasystem were compared to that of the parental R6 strain. Inboth cases, oligonucleotide microarray technology was used toidentify the isolated DNA fragments and the transcripts, re-spectively. The overall picture revealed that the cia system ispart of a complex regulatory network: it not only regulatesgenes directly in a positive or negative way but also controls theentire competence regulon. The com regulon is composed ofthe quorum-sensing system comCDE, which not only mediatesits own autoregulation but also induces early competencegenes, among which are the ComX regulators controlling latecompetence genes (30).
The method used here for identification of CiaR bindingsites—a modified procedure developed for isolating targetDNA fragments of DNA binding proteins—is certainly appli-cable to other response regulators and transcription regulatorsin general, provided that the protein construct is sufficientlystable during the incubation and washing steps necessary toobtain the desired specificity for DNA binding. The high re-producibility of the DNA fragment pattern obtained after the
SPDB assay supports the specificity of the DNA-protein inter-action, which was confirmed using isolated target fragments inthe presence of competitor DNA (Fig. 3). The number ofputative target regions identified after hybridization to theoligonucleotide microarray corresponds well to the number ofbands contained in the SPDB-eluted fraction. We used a highlevel of stringency for discrimination of DNA fragments spe-cifically interacting with the CiaR protein. Due to the presenceof nonspecific background after the SPDB assay, the noiselevel was set relatively high compared to previous studies (21).The use of three distinct restriction digests in the SPDB assayallowed the easy discrimination of apparent positive regions: insome cases, positive signals were restricted to only one of therestriction digests and further discrimination was achieved bycomparing the restriction map of the apparent positive frag-ments with the locations of the positive probe sets.
Although many of the putative cia target loci identified bydirect interaction of DNA fragments with CiaR were detectedin the transcription-profiling approach, some were not (andseen in the transcription analysis were not revealed as targetgenes by the SPDB assay). Five of the gene loci were notexpressed under any conditions or were absent in the R6 strain.It has been shown recently that different methods employed instudies defining the Bacillus subtilis W-regulated genes nevercovered the entire regulon and never overlapped completely(10), and a similar conclusion can be drawn from the presentstudy with the cia regulon. Two gene loci were not detected inthe SPDB assay although they were clearly expressed in acia-dependent manner. It is possible that the putative bindingsite for CiaR is restricted by all three enzymes used in thisassay or that they are indirectly controlled by the cia system. Itshould also be pointed out that we used nonphosphorylatedbCiaR in the assay. Attempts to phosphorylate the proteinwere unsuccessful due to the instability of the biotinylatedfusion derivative bCiaR under these conditions. However, al-though phosphorylation may result in a higher affinity to thebinding sites, the specificity should not be affected, and the factthat cia-dependent expression was observed for targeted loci isin agreement with this conclusion. Computer searches wereunsuccessful in identifying a consensus binding site for CiaRfor all cia-regulated genes, which may not be surprising giventhe low level of strictly conserved nucleotides in several knownbinding sites of response regulators, and one has to await directbinding studies, e.g., footprint analyses.
Loss-of-function mutants are often without significant phe-notype, and since they represent constitutive situations, tran-scription analyses do not distinguish between primary and sec-ondary regulated genes. It required the combination of the twoexperimental approaches, direct DNA binding and transcriptanalyses, as well as two sets of mutants, the R6-R1 mutant witha nonfunctional cia system and the kinase mutant R6ciaHC306
reflecting an on situation, in order to be able to distinguishbetween primary and secondary regulated genes and to con-firm gene induction or repression in the cia mutants relative tothe wild-type situation. By using two different growth media (Cmedium and THB), it became apparent that the regulationmachinery is also modulated under different environmentalconditions and thus perhaps in different in vivo situations aswell. This could be the reason for distinct results obtained withloss-of-function cia mutants using two different murine models.
TABLE 3. Competence-related cia-regulated genes
Operon Genea ReferencebGenomic position
KNR.7/87 R6
1 ccs1 43, 47 SP0124 spr0127ccs15 SP0125 spr0128
2 clpB SP0338 spr03073 orfA SP0428 spr0387
orfB SP0429 spr0388orfC SP0430 spr0389
4 celA (cilE) 2, 9, 30, 42 SP0954 spr0856celB 3, 30, 37, 41, 42 SP0955 spr0857orfD SP0956 spr0858orfE SP0957 spr0859orfF SP0958 spr0860
5 coiA 3, 30, 37, 41, 42 SP0978 spr08816 radC 47 SP1088 spr09967 smf (cilB) SP1266 spr11448 pilD 47 SP1808 spr16289 ssbB (cilA) 2, 9, 30, 42 SP1908 spr172410 lytA SP1937 spr1754
dinF SP1939 spr1756recA 2, 9, 30, 42 SP1940 spr1757cinA 3, 30, 37, 41, 42 SP1941 spr1758
11 orfG SP1945 spr176212 comX2 5, 43 SP2006 spr181913 CPIP788 5, 43 SP2017 spr183014 orf1 SP2047 spr1857
cglE 3, 30, 37, 41, 42 SP2048 spr1859cglD SP2050 spr1861cglC SP2051 spr1862cglB SP2052 spr1863cglA (cilD) 2, 3, 9, 30, 37, 41, 42 SP2053 spr1864
15 cbpD 43, 47 SP2201 spr200616 comFC (cflB) SP2207 spr2012
comFA (cflA) 3, 30, 37, 41, 42 SP2208 spr201117 comE 43 SP2235 spr2041
comD SP2236 spr204218 comX1 3, 5, 30, 37, 41, 42 SP0014 spr001319 orfK SP0029 spr0030
ccs16 43 SP0030 spr003120 comA 3, 5, 30, 37, 41–43, 47 SP0042 spr0043
comB SP0043 spr0044
a orfA to orfK encode hypothetical proteins of unknown function.b Competence-related expression studies.
VOL. 185, 2003 STREPTOCOCCUS PNEUMONIAE cia REGULON 67
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
In only one case, where the development of pneumonia afterintranasal inoculation was tested, was virulence clearly affectedin the mutants (53), whereas no effect was detected in anintraperitoneal-infection model (27).
The cia system may also be differentially activated during thegrowth cycle, as could be seen for the dlt and lic operons, bothof which were expressed only during late exponential growth inthe wild type. The gene products of both loci are involved inteichoic acid metabolism. Stationary-phase cells are known tohave a biochemically distinct cell envelope (18, 54), and it isconceivable that cia is one of the mediators of such changes.This experiment also indicates that detailed kinetic analyses ofall the cia-dependent gene loci identified here, preferably indifferent growth media, would be helpful. It is possible that thefailure to detect cia-dependent gene expression for some lociidentified as CiaR targets is related to the fact that sampleswere taken during exponential growth phase and not duringstationary phase. The microarray used in the present study isno longer available. Further tests thus have to wait for anothergeneration microarray or will require the isolation of reporterconstructs or real-time PCR for the other gene loci.
The putative target genes cover a whole range of functionsrelated to cell wall polysaccharide biochemistry. We do notknow yet whether the phenotype of ciaH mutants—increasedbeta-lactam resistance—is due to one particular gene productor is a reflection of an overall change in cell wall biochemistrymediated by the cia regulon. The twofold increase in the MICof cefotaxime for ciaH mutants was observed in the wild-typebackground (from 0.02 to 0.04 �g/ml) and in mutants wherepbp2x mutations had already resulted in elevated MICs (from0.2 to 0.4 �g/ml), and these values are in the range conferredby single point mutations in penicillin binding protein genes(17, 58). Homologues of the enzymes pyrimidine-nucleosidephosphorylase and deoxyribose phosphate aldolase are re-quired for pyrimidine biosynthesis and feed into the precursor
FIG. 5. Genes regulated by the cia system. The intensity scatter graph shows the correlation for the intensities of all transcripts obtained forthe loss-of-function cia mutant R6-R1 versus those of R6 in C medium (center) and in THB medium (right). A comparison between twoindependently grown R6 cultures in C medium is included as a control (left). The two lines flanking the diagonal indicate differences of a factorof 2 and 5, respectively. In the center graph, several genes are downregulated in the mutant. In addition, a large set of genes appears to beupregulated in THB medium (right).
FIG. 6. �-Galactosidase assay of the cia-dependent activation ofthe lic promoter (A) and the comC promoter (B). Construction of thelacZ reporter strains is described in Materials and Methods. Cultureswere grown at 37°C in THB (A) or C medium (B). Samples were takenat the time points indicated, and the assay of �-galactosidase activitywas performed as described in the text. Circles, R6; squares, R6-R1;triangles, R6TC306; solid symbols, growth (optical density at 560 nm[OD560]); open symbols, LacZ activity (Miller units).
68 MASCHER ET AL. J. BACTERIOL.
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

pool of XTP-activated sugar compounds. The enzymes phos-phoglycerate mutase and pyruvate formate lyase play key rolesin the glycolytic pathway. It has been shown that glycolyticintermediates differ in exponential- and stationary-phasegrowth and that these changes correlate with the production ofexopolysaccharide in L. lactis (44). It is curious that two cia-regulated loci encode enzymes involved in teichoic acid bio-synthesis or modification: the lic locus is required for phospho-ryl choline metabolism, and the dlt locus is involved in Dalanylation in related bacteria. No D alanylation of teichoicacids has been observed in S. pneumoniae (W. Fischer, per-sonal communication). The activation of the dlt operon oc-curred only in certain media and only at the end of exponentialgrowth, and it is possible that the failure to detect D alanyla-tion of teichoic acids is related to analyzing cells grown underconditions where the dlt operon is not active.
What is the link between the directly cia-regulated genes andthe cia-mediated effect on competence? None of the compe-tence-regulated gene loci was targeted by CiaR, whereas thetranscript analyses clearly documented a complete repressionof competence genes in the ciaHC306 mutant background,which was reflected in more detail in the comC promoter-lacZfusion (Fig. 6B). The competence deficiency mediated by ciaHmutations could be complemented by the addition of CSP, thecomC gene product responsible for the induction of the com-petence regulon (60). It has recently been suggested that HtrAis involved in cia-mediated inhibitory control over the compe-tence pathway (49). However, a loss-of-function htrA mutantdid not restore competence in the R6 strain when tested inTHB medium, where the cia system appears to be activated(M. Merai, unpublished results), suggesting that it is not theHtrA protease itself that is responsible for the inhibition ofcompetence. The comCDE operon region is located approxi-mately 1 kb upstream of the htrA-spo0J locus, which contains aCiaR binding site. It is possible that the repression of compe-tence under cia-activating conditions is due to interferencewith ComE binding to the comC promoter region. Alterna-tively, competence induction might be prevented by cia-medi-ated altered surface properties that could interfere with suffi-cient CSP production or recognition.
ACKNOWLEDGMENTS
We thank Walter Messer for the gift of plasmid pBEX5BA, Jean-Pierre Claverys for plasmid pEVP3, and Don Morrison for pXF520.
This work was supported by grants from the Deutsche Forschungs-gemeinschaft and the Schwerpunkt fur Biotechnologie of the Univer-sity of Kaiserslautern.
REFERENCES
1. Aaberge, I. S., J. Eng, G. Lermark, and M. Lovik. 1995. Virulence of Strep-tococcus pneumoniae in mice: a standardized method for preparation andfrozen storage of the experimental bacterial inoculum. Microb. Pathog. 18:141–152.
2. Alloing, G., C. Granadel, D. A. Morrison, and J.-P. Claverys. 1996. Compe-tence pheromone, oligopeptide permease, and induction of competence inStreptococcus pneumoniae. Mol. Microbiol. 21:471–478.
3. Alloing, G., B. Martin, C. Granadel, and J.-P. Claverys. 1998. Developmentof competence in Streptococcus pneumoniae: pheromone autoinduction andcontrol of quorum sensing by the oligopeptide permease. Mol. Microbiol.29:75–83.
4. Apfel, C. M., B. Takacs, M. Fountoulakis, M. Stieger, and W. Keck. 1999.Use of genomics to identify bacterial undecaprenyl pyrophosphate syn-thetase: cloning, expression, and characterization of the essential uppS gene.J. Bacteriol. 181:483–492.
5. Bartilson, M., A. Marra, J. Christine, J. S. Asundi, W. P. Schneider, and
A. E. Hromockyj. 2001. Differential fluorescence induction reveals Strepto-coccus pneumoniae loci regulated by competence stimulator peptide. Mol.Microbiol. 39:126–135.
6. Bolotin, A., O. Wincker, S. Mauger, O. Jaillon, K. Marlarme, J. Weissen-bach, S. D. Ehrlich, and A. Sorokin. 2001. The complete genome sequenceof the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. GenomeRes. 11:731–753.
7. Boyd, D. A., D. G. Cvitkovitch, A. S. Bleiweis, M. Y. Kiriukhin, D. V.Debabov, F. C. Neuhaus, and I. R. Hamilton. 2000. Defects in D-alanyl-lipoteichoic acid synthesis in Streptococcus mutans results in acid sensitivity.J. Bacteriol. 182:6055–6065.
8. Brown, J. S., S. M. Gilliland, and D. W. Holden. 2001. A Streptococcuspneumoniae pathogenicity island encoding an ABC transporter involved iniron uptake and virulence. Mol. Microbiol. 40:572–585.
9. Campbell, E. A., S. Y. Choi, and H. R. Masure. 1998. A competence regulonin Streptococcus pneumoniae revealed by genomic analysis. Mol. Microbiol.27:929–939.
10. Cao, M., P. A. Kobel, M. M. Morshedi, M. F. W. Wu, C. Paddon, and J. D.Helmann. 2002. Defining the Bacillus subtilis W regulon: a comparativeanalysis of promoter consensus search run-off transcription macroarray anal-ysis (ROMA), and transcriptional profiling approaches. J. Mol. Biol. 316:443–457.
11. Claverys, J.-P., A. Dintilhac, E. V. Pestova, B. Martin, and D. A. Morrison.1995. Construction and evaluation of new drug-resistance cassettes for genedisruption mutagenesis in Streptococcus pneumoniae, using an ami test plat-form. Gene 164:123–128.
12. Degrassi, G., M. Kojic, G. Ljubijankic, and V. Venturi. 2000. The acetylxylan esterase of Bacillus pumilus belongs to a family of esterases with broadsubstrate specificity. Microbiology 146:1585–1591.
13. de Saizieu, A., C. Gardes, N. Flint, C. Wagner, M. Kamber, T. J. Mitchell, W.Keck, K. E. Amrein, and R. Lange. 2000. Microarray-based identification ofa novel Streptococcus pneumoniae regulon controlled by an autoinducedpeptide. J. Bacteriol. 182:4696–4703.
14. Echenique, J. R., S. Chapuy-Regaud, and M. C. Trombe. 2000. Competenceregulation by oxygen in Streptococcus pneumoniae: involvement of ciaRH andcomCDE. Mol. Microbiol. 36:688–696.
15. Filipe, S. R., and A. Tomasz. 2000. Inhibition of the expression of penicillin-resistance in Streptococcus pneumoniae by inactivation of cell wall muropep-tide branching genes. Proc. Natl. Acad. Sci. USA 97:4891–4896.
16. Gasc, A.-M., P. Giammarinaro, S. Richter, and M. Sicard. 1998. Organiza-tion around the dnaA gene of Streptococcus pneumoniae. Microbiology 144:129–138.
17. Giammarinaro, P., A. M. Sicard, and A.-M. Gasc. 1999. Genetic and phys-iological studies of the CiaH-CiaR two-component signal-transducing systeminvolved in cefotaxime resistance and competence of Streptococcus pneu-moniae. Microbiology 145:1859–1869.
18. Glauner, B., J.-V. Holtje, and U. Schwarz. 1988. The composition of themurein of Escherichia coli. J. Biol. Chem. 263:10088–10095.
19. Guenzi, E., A. M. Gasc, M. A. Sicard, and R. Hakenbeck. 1994. A two-component signal-transducing system is involved in competence and peni-cillin susceptibility in laboratory mutants of Streptococcus pneumoniae. Mol.Microbiol. 12:505–515.
20. Guenzi, E., and R. Hakenbeck. 1995. Genetic competence and susceptibilityto �-lactam antibiotics in Streptococcus pneumoniae R6 are linked via atwo-component signal-transducing system, p. 125–128. In J. J. Ferretti, M. S.Gilmore, T. R. Klaenhammer, and F. Brown (ed.), Genetics of Streptococci,Enterococci and Lactococci. S. Karger AG, Basel, Switzerland.
21. Hakenbeck, R., N. Balmelle, B. Weber, C. Gardes, W. Keck, and A. deSaizieu. 2001. Mosaic genes and mosaic chromosomes: intra- and interspe-cies variation of Streptococcus pneumoniae. Infect. Immun. 69:2477–2486.
22. Hakenbeck, R., T. Grebe, D. Zahner, and J. B. Stock. 1999. �-Lactamresistance in Streptococcus pneumoniae: penicillin-binding proteins and non-penicillin-binding proteins. Mol. Microbiol. 33:673–678.
23. Hakenbeck, R., A. Konig, I. Kern, M. van der Linden, W. Keck, D. Billot-Klein, R. Legrand, B. Schoot, and L. Gutmann. 1998. Acquisition of fivehigh-Mr penicillin-binding protein variants during transfer of high-level�-lactam resistance from Streptococcus mitis to Streptococcus pneumoniae. J.Bacteriol. 180:1831–1840.
24. Havarstein, L. S., G. Coomaraswamy, and D. A. Morrison. 1995. An un-modified heptadecapeptide pheromone induces competence for genetictransformation in Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA92:11140–11144.
25. Hoskins, J., W. E. Alborn, Jr., J. Arnold, L. C. Blaszczak, S. Burgett, B. S.DeHoff, S. T. Estrem, L. Fritz, D. J. Fu, W. Fuller, C. Geringer, R. Gilmour,J. S. Glass, H. Khoja, A. R. Kraft, R. E. Lagace, D. J. LeBlanc, L. N. Lee,E. J. Lefkowitz, J. Lu, P. Matsushima, S. M. McAhren, M. McHenney, K.McLeaster, C. W. Mundy, T. I. Nicas, F. H. Norris, M. O’Gara, R. B. Peery,G. T. Robertson, P. Rockey, P. M. Sun, M. E. Winkler, Y. Yang, M. Young-Bellido, G. Zhao, C. A. Zook, R. H. Baltz, S. R. Jaskunas, P. R. J. Rosteck,P. L. Skatrud, and J. I. Glass. 2001. Genome of the bacterium Streptococcuspneumoniae strain R6. J. Bacteriol. 183:5709–5717.
26. Lacks, S. A., and R. D. Hotchkiss. 1960. A study of the genetic material
VOL. 185, 2003 STREPTOCOCCUS PNEUMONIAE cia REGULON 69
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

determining an enzyme activity in pneumococcus. Biochim. Biophys. Acta39:508–517.
27. Lange, R., C. Wagner, A. de Saizieu, N. Flint, J. Molnos, M. Stieger, P.Caspers, M. Kamber, W. Keck, and K. E. Amrein. 1999. Domain organiza-tion and molecular characterization of 13 two-component systems identifiedby genome sequencing of Streptococcus pneumoniae. Gene 237:223–234.
28. Lau, G. W., S. Haataja, M. Lonetto, S. E. Kensit, A. Marra, A. P. Bryant, D.McDevitt, D. A. Morrison, and D. W. Holden. 2001. A functional genomicanalysis of type 3 Streptococcus pneumoniae virulence. Mol. Microbiol. 40:555–571.
29. LeBlanc, D. J., L. N. Lee, and J. M. Inamine. 1991. Cloning and nucleotidebase sequence analysis of a spectinomycin adenyltransferase AAD(9) deter-minant from Enterococcus faecalis. Antimicrob. Agents Chemother. 35:1804–1810.
30. Lee, M. S., and D. A. Morrison. 1999. Identification of a new regulator inStreptococcus pneumoniae linking quorum sensing to competence for genetictransformation. J. Bacteriol. 181:5004–5016.
31. Lockhart, D. J., H. Dong, M. C. Byrne, M. T. Follettie, M. V. Gallo, M. S.Chee, M. Mittmann, C. Wang, M. Kobayashi, H. Horton, and E. L. Brown.1996. Expression monitoring by hybridization to high-density oligonucleotidearrays. Nat. Biotechnol. 14:1675–1680.
32. Lortie, L. A., M. Pelletier, C. Vadeboncoeur, and M. Frenette. 2000. Thegene encoding IIAB(Man)L in Streptococcus salivarius is part of a tetracis-tronic operon encoding a phosphoenolpyruvate: mannose/glucose phospho-transferase system. Microbiology 146:677–685.
33. Maes, M., and S. Messens. 1992. Phenol as grinding material in RNApreparations. Nucleic Acids Res. 20:4374.
34. Martin, B., M. Prudhomme, G. Alloing, C. Granadel, and J.-P. Claverys.2001. Cross-regulation of competence pheromone production and export inthe early control of transformation in Streptococcus pneumoniae. Mol. Mi-crobiol. 38:876–878.
35. Martinez-Hackert, E., and A. M. Stock. 1997. Structural relationships in theOmpR family of winged-helix transcription factors. J. Mol. Biol. 269:301–312.
36. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
37. Mortier-Barriere, I., A. de Saizieu, J.-P. Claverys, and B. Martin. 1998.Competence-specific induction of recA is required for full recombinationproficiency during transformation in Streptococcus pneumoniae. Mol. Micro-biol. 21:159–170.
38. Murtif, V. L., C. R. Bahler, and D. Samols. 1985. Cloning and expression ofthe 1.3S biotin-containing subunit of transcarboxylase. Proc. Natl. Acad. Sci.USA 82:5617–5621.
39. Nieto, C., A. Puyet, and M. Espinosa. 2001. MalR-mediated regulation of theStreptococcus pneumoniae malMP operon at promoter PM. Influence of aproximal divergent promoter region and competition between MalR andRNA polymerase proteins. J. Biol. Chem. 276:14946–14954.
40. Parkinson, J. S. 1993. Signal transduction schemes of bacteria. Cell 73:857–871.
41. Pestova, E. V., L. S. Havarstein, and D. A. Morrison. 1996. Regulation ofcompetence for genetic transformation in Streptococcus pneumoniae by anauto-induced peptide pheromone and a two-component regulatory system.Mol. Microbiol. 21:853–862.
42. Pestova, E. V., and D. A. Morrison. 1998. Isolation and characterization ofthree Streptococcus pneumoniae transformation-specific loci by use of a lacZreporter insertion vector. J. Bacteriol. 180:2701–2710.
43. Peterson, S., R. T. Cline, H. Tettelin, V. Sharov, and D. A. Morrison. 2000.Gene expression analysis of the Streptococcus pneumoniae competence regu-lons by use of DNA microarrays. J. Bacteriol. 182:6192–6202.
44. Ramos, A., I. C. Boels, W. M. de Vos, and H. Santos. 2001. Relationshipbetween glycolysis and exopolysaccharide biosynthesis in Lactococcus lactis.Appl. Environ. Microbiol. 67:33–41.
45. Reichmann, P., and R. Hakenbeck. 2000. Allelic variation in a peptide-inducible two component system of Streptococcus pneumoniae. FEMS Mi-crobiol. Lett. 190:231–236.
46. Richter, S., W. R. Hess, M. Krause, and W. Messer. 1998. Unique organi-zation of the dnaA region from Prochlorococcus marinus CCMP1375, amarine cyanobacterium. Mol. Gen. Genet. 257:534–541.
47. Rimini, R., B. Jansson, G. Feger, T. C. Roberts, M. de Francesco, A. Gozzi,F. Faggioni, E. Domenici, D. M. Wallace, N. Frandsen, and A. Polissi. 2001.Global analysis of transcription kinetics during competence development inStreptococcus pneumoniae using high density DNA arrays. Mol. Microbiol.36:1279–1292.
48. Roth, A., and W. Messer. 1995. The DNA binding domain of the initiatorprotein DnaA. EMBO J. 14:2106–2111.
49. Sebert, M. E., L. M. Palmer, M. Rosenberg, and J. N. Weiser. 2002. Mi-croarray-based identification of htrA, a Streptococcus pneumoniae gene thatis regulated by the CiaRH two-component system and contributes to naso-pharyngeal colonization. Infect. Immun. 70:4059–4067.
50. Smith, M. D., and W. R. Guild. 1979. A plasmid in Streptococcus pneu-moniae. J. Bacteriol. 137:735–739.
51. Stock, J. B., M. G. Surette, M. Levit, and P. Park. 1995. Two-componentsignal transduction systems: structure-function relationships and mecha-nisms of catalysis, p. 25–51. In J. A. Hoch and T. J. Silhavy (ed.), Two-component signal transduction. American Society for Microbiology, Wash-ington, D.C.
52. Tettelin, H., K. E. Nelson, I. T. Paulsen, J. A. Eisen, T. D. Read, S. Peterson,J. Heidelberg, R. T. DeBoy, D. H. Haft, R. J. Dodson, A. S. Durkin, M.Gwinn, J. F. Kolonay, W. C. Nelson, J. D. Peterson, L. W. Umayam, O.White, S. L. Salzberg, M. R. Lewis, D. Radune, E. Holtzapple, H. Khouri,A. M. Wolf, T. R. Utterback, C. L. Hansen, L. A. McDonald, T. V. Feldblyum,S. Angiuoli, T. Dickinson, E. K. Hickey, I. R. Holt, B. J. Loftus, F. Yang,H. O. Smith, J. C. Vetern, B. A. Dougherty, D. A. Morrison, S. K. Holling-shead, and C. M. Fraser. 2001. Complete genome sequence of a virulentisolate of Streptococcus pneumoniae. Science 293:498–506.
53. Throup, J. P., K. K. Koretke, A. P. Bryant, K. A. Ingraham, A. F. Chalker,Y. Ge, A. Marra, N. G. Wallis, J. R. Brown, D. J. Holmes, M. Rosenberg, andK. R. Burnham. 2000. A genomic analysis of two-component signal trans-duction in Streptococcus pneumoniae. Mol. Microbiol. 35:566–576.
54. Warth, A. D., and J. L. Strominger. 1972. Structure of the peptidoglycanfrom spores of Bacillus subtilis. Biochemistry 11:1389–1396.
55. Weber, B., K. Ehlert, A. Diehl, P. Reichmann, H. Labischinski, and R.Hakenbeck. 2000. The fib locus in Streptococcus pneumoniae is required forpeptidoglycan crosslinking and PBP-mediated beta-lactam resistance. FEMSMicrobiol. Lett. 188:81–85.
56. Wodicka, L., H. Dong, M. Mittmann, M. H. Ho, and D. J. Lockhart. 1997.Genome-wide expression monitoring in Saccharomyces cerevisiae. Nat. Bio-technol. 15:1359–1367.
57. Young, R. A., and R. W. Davis. 1983. Yeast RNA polymerase II genes:isolation with antibody probes. Science 222:778.
58. Zahner, D., T. Grebe, E. Guenzi, J. Krau�, M. van der Linden, K. Terhune,J. B. Stock, and R. Hakenbeck. 1996. Resistance determinants for �-lactamantibiotics in laboratory mutants of Streptococcus pneumoniae that are in-volved in genetic competence. Microb. Drug Resist. 2:187–191.
59. Zahner, D., and R. Hakenbeck. 2000. The Streptococcus pneumoniae beta-galactosidase is a surface protein. J. Bacteriol. 182:5919–5921.
60. Zahner, D., K. Kaminski, M. van der Linden, T. Mascher, M. Merai, and R.Hakenbeck. 2001. The ciaR/ciaH regulatory network of Streptococcus pneu-moniae. J. Mol. Microbiol. Biotechnol. 4:211–216.
61. Zhang, J. R., I. Idanpaan-Heikkila, W. Fischer, and E. I. Tuomanen. 1999.Pneumococcal licD2 gene is involved in phosphorylcholine metabolism. Mol.Microbiol. 31:1477–1488.
70 MASCHER ET AL. J. BACTERIOL.
on February 6, 2018 by guest
http://jb.asm.org/
Dow
nloaded from