Embed Size (px)
Citation preview

0 0 1 I .. principles and practice
The Structure and Function of Brown Adipose Tissue in the Neonate VIRGINIA DA VIS, RN, MSN
Presented is a discussion oJbrown adipose cell structure as compared to that of the white adipose cell, the role which brown adipose tissue ( B A T ) plays in thennoregulation, the physiologic mechanisms which control BA T heat generation, and the proposed biochemical processes which are responsible for heat production in the cell. Based on the physiology of BA T thnmogenesis, specijc nursing measures to minimize cold stimuli are recommended.
The newborn arrives in this world with special requirements for body temperature maintenance. Unlike the adult, the infant cannot rely on muscular activity such as shivering or moving to a warmer environment as a means of heat production or maintenance. The infant has a much larger body surface to mass ratio than an adult (0.066 m'/kg for a 3 kg infant' vs. 0.025 m'/kg for a 70 kg adult), which means that the infant will lose heat much more rapidly to the environment. The infant has less insulating fat than an adult (16% of body weight in a 3.5 kg infant vs. 20-30'36 in an adult'). The infant must rely, therefore, on increased metabolic processes (non-shivering thermogenesis) for the production of heat beyond basal production. One heat-producing process which is available to the neonate is the me- tabolism of brown adipose tissue, commonly referred to as BAT or brown fat. The contribution of BAT to the total heat produced during non-shivering thermogenesis is un- known, but it may be as high as 100% in the neonate.:'
Until the early 1960 '~~ the physio- logic role of BAT was uncertain. BAT was known to be present in some mammalian young (including human infants) and was linked to the arousal state of hibernating ani- mals. In 1963, Dawkins and Hull4
noted that there was an increase in the temperature of the skin over the intrascapular BAT in rabbits when they were cold-stressed or norepi- nephrine was given and that BAT had a higher oxygen consumption than white adipose tissue. From these observations, they proposed that BAT was a site of heat produc- tion.
Since that time, considerable sci- entific effort has been invested in de- scribing the role of BAT in non-shiv- ering thermogenesis. Most of the inquiry has been focused on the dif- ference between brown and white adipose tissue, the extent of the heat produced by BAT, the stimulus to BAT heat production, and the bio- chemical processes in BAT which are responsible for heat production. The exact role of BAT in non-shiver- ing thermogenesis is still unknown. The material presented in this ar- ticle is theoretical and so is subject to change as new information is gath- ered. The material does not include all principles of thermoregulation in the newborn but only those that are concerned with the relationship of BAT to heat production.
Development and Location of BAT BAT cells begin to differentiate in
the 26th-30th weeks of gestation.' They continue to increase in number and content until 2-3 weeks after
birth in the full-term infant. The premature infant does not have a full complement of BAT at birth, which seriously impairs the ability to create heat in cool environments. BAT is deposited in the midscapular region, the posterior neck, around the neck muscles and extending un- der the clavicles to the axillae, and around the trachea, esophagus, in- tercostal and mammary arteries, ab- dominal aorta, kidneys, and adrenal glands." (See Figure 1.) These areas of deposition serve a twofold pur- pose in maintaining the body tem- perature during cold exposure:
1. The BAT surrounding vital or- gans ensures the warmth necessary for optimal biochemical activities within the organs. At lower than op- timal temperatures the rate of enzy- matic reactions is slowed, leading to an accumulation of metabolic wastes.
2. The heat generated from the BAT surrounding arteries warms the blood before distribution to the pe- ripheral circulation.
Differences Between Brown and White Adipose Tissue
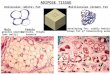
The structure of the BAT cell dif- fers from that of the white adipose cell in several ways (see Figure 2). The BAT cell contains multiple fat vacuoles, a central round nucleus, glycogen, and multiple large mito- chondria (the cellular power plants). The white adipose cell contains one large fat vacuole, a crescent-shaped nucleus located in the cell periphery, no glycogen, and few mitochondria.'
The structure of the BAT cell is well-suited to its function. The large number of small fat vacuoles pro- vides a large fat to cytoplasm inter- face which enhances the rapidity with which the s tored fat (tri- glycerides) can be utilized. The large number of mitochondria provide the energy, in the form of adenosine triphosphate (ATP), which is neces- sary for rapid metabolic turnover and heat production. The presence of glycogen in the cell provides a
November/Decernber 198OJOGN Nursinf 0090-03 1 1/80/ I I 17-0368)o 100
ready source of glucose for the in- creased energy needs and production of ATP.
Unlike white adipose tissue, BAT has a rich blood supply and venous drainage system. The blood not only carries the cellular nutrients and products of metabolism, but it is also the recipient and distributor of the heat generated by the cell. The venous blood returning to the heart from BAT is thus warmed before being pumped into the peripheral circulation. Brown adipose tissue de- rives its name from the reddish- brown color it becomes when de- pleted of fat. Most of the color comes from the hemoglobin of the surrounding capillaries.’
The intrascapular BAT, which does not surround any vital struc- tures, has a unique venous drainage system. Before its venous blood re- turns to the heart it passes into the venous plexuses of the vertebral col- umn. This route of drainage ensures a warm supply of blood to the spinal cord.
In contrast to white adipose tissue, BAT has an extensive nerve supply. Each BAT cell is cradled in a basket of axons which arise from the sym- pathetic nervous system. The nerve supply provides a major stimulus to metabolic action, which will be dis- cussed later.
Control of BAT The activation of BAT thermoge-
nesis begins with a cold stimulus to peripheral receptors located in the skin. Physiologists in the past have attributed the sensing of cold to spe- cialized nerve endings, but it is gen- erally thought now that cold recep- tion takes place in naked nerve endings.” Although the cold recep- tors are located throughout the skin of the infant, there is an abundance of them in the facial skin, which ren- ders the infant particularly sensitive to changes in temperature around the head.’
A cold stimulus to the peripheral receptors causes them to increase their rate of firing (impulse produc- tion). A sudden decrease in the tem- perature of the skin will elicit a brief burst of rapid firing, whereas a sus- tained lower-than-normal skin tem-
Figure 1. The distribution of BAT in the neonate.
perature will elicit an increased but steady rate of firing.”
The impulses of the skin receptors are transmittted to a neuron pool lo- cated in the posterior hypothalamus. The hypothalamus functions as the master control of thermoregulation. It is divided into two areas of ther- moregulatory control: anterior and posterior. The anterior area is tem- perat ure-sensit ive and controls the body’s heat-loss mechanisms. It is considered to be the site of the set- point or threshold temperature. The set-point temperature is defined as “ . . . the temperature at which ac- tivities of the various antirise and antidrop neurons are equivalent and opposite in effects.”“’ In other words, it is the hypothalamic (central) tem- perature at which there is no change in bodily activity directed specifi- cally towards the conservation, gen- eration, or dissipation of heat. The set-point varies between individuals b u t is normally near 37OC in adults.“ It may be lower in pre- mature in fan ts. ’’
The posterior area of the hypo- thalamus is not temperature-sensi-
BROWN ADIPOSE CELL
I/-+---=% ~~ -
ytoplaim
Capillary
at Vacuole
tive but does control the body’s responses to cold stimulation, in- cluding BAT thermogenesis. It is de- pendent on sensory input from other areas of the body for stimulation, whereas the anterior area uses infor- mation from peripheral and local temperature sensors.
The relationship between the an- terior and posterior areas of the hy- pothalamus remains in question. Benzinger theorizes that the anterior hypothalamus has an inhibitory ef- fect on the posterior portion if the central temperature is above set- point.’’ The cooling of peripheral re- ceptors, therefore, will not lead to BAT thermogenesis if the infant’s central temperature (measured at the tympanic membrane by Ben- zinger) is elevated above the set- point. Bligh, however, concludes from a review of experimental evi- dence that the sum of all cold and warm stimuli determines whether the anterior or posterior hypothala- mus will gain expression.“
If the posterior hypothalamus wins the “hypothalamic battle”, it will initiate BAT thermogenesis via
WHITE ADIPOSE CELL
Figure 2. The structures of the brown and white adipose cells.
Novernber/December 1980JOCN Nursing 369

SPINAL ICORO
Pituitary
Adrenal Medulla
Thyroid
Figure 3. The neural and endocrine pathways of BAT control. (TRF = thyrotropin releas- ing factor; TSH = thyroid stimulating hormone; NE = norepinephrine.)
the neural and endocrine pathways (see Figure 3). The sympathetic ner- vous system is stimulated to release norepinephrine (NE) at the nerve endings surrounding BAT and to re- lease catecholamines from the adre- nal medulla. Although BAT is not dependent on circulat ing cate- cholamines for activation," they will stimulate glycogenolysis, which pro- vides the glucose necessary for ther- mogenesis.
The posterior hypothalamus also controls thyroid activity via the pi- tuitary gland. The stimulation of the thyroid gland increases the produc- tion and release of thyroxine, which enhances the effects of NE on the BAT cell.
The Proposed Heat-Producing Cellular Mechanisms
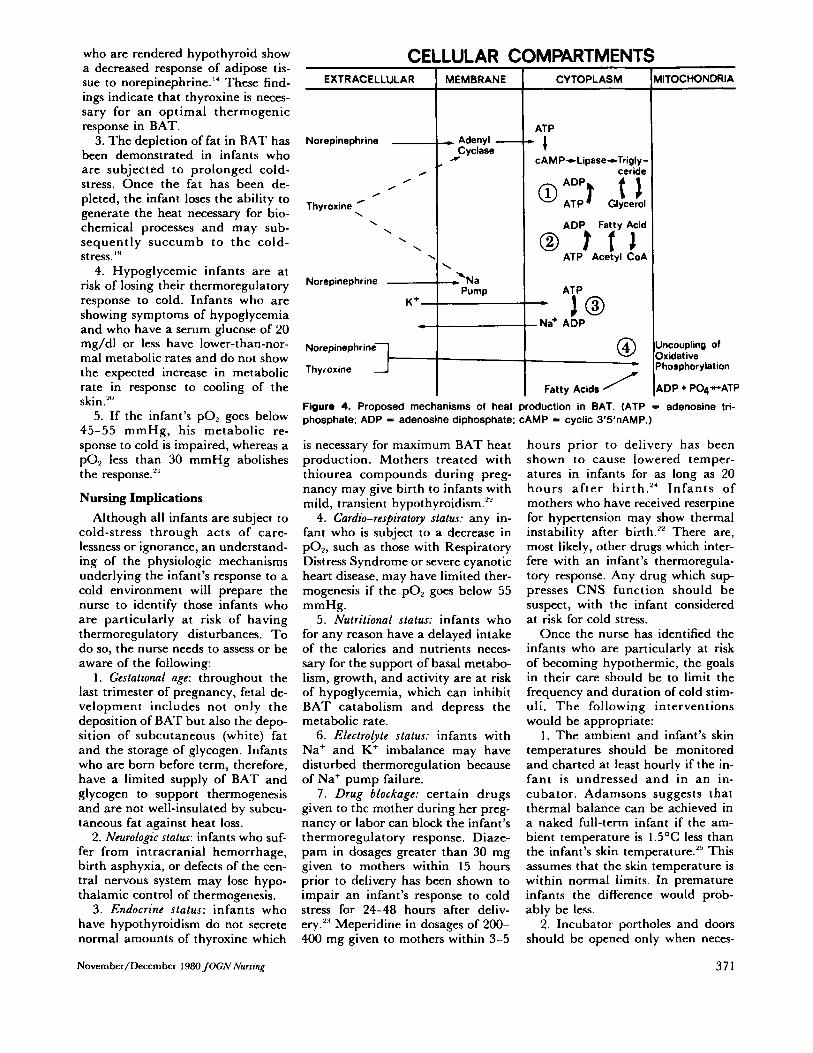
Four possible mechanisms of heat production in the BAT cell have been proposed" (see Figure 4). After NE has been released at the nerve ending, i t activates adenyl cyclase, a membrane-bound enzyme which catalyzes the conversion of ATP to cyclic 3',5'-AMP (adenosine mono- phosphate). ATP, a molecule con- sisting of ribose, adenine (a purine base), and three phosphates, plays a major role in supplying energy for biochemical reactions within the cell. Cyclic 3',5'-AMP, in turn, acti-
370
vates a lipase which catalyzes the hydrolysis of the stored triglycerides into glycerol and fatty acids. At this point, two heat-producing events may occur. The glycerol and fatty acids may be reconverted into tri- glycerides (Figure 4, Reaction 1) or the fatty acids may be oxidized to acetyl CoA and then resynthesized (Figure 4, Reaction 2). Thyroxine may enhance these mechanisms by increasing the amount of adenyl cy- clase in the cell membrane."
A third possible mechanism of heat production is the stimulation of the sodium pump by NE or thyrox- ine. It is thought that NE increases the permeability of the cell mem- brane so that there is an increased flow of Na' into the cell and K' out of the cell."' This disruption of the normal intracellular and extra- cellular ion concentration is cor- rected by the Na' pump. The acti- vation of the pump requires energy which is provided by ATP. The cleavage of the third phosphate bond of ATP releases energy, some of which drives the pump and some of which is lost as heat (Figure 4, Reaction 3). Thyroxine either di- rectly activates the pump or in- creases the number of pump sites within the cell membrane.15
A fourth possible mechanism of heat production is the uncoupling of
oxidative phosphorylation in the mitochondria (Figure 4, Reaction 4). Oxidative phosphoryla t ion is the process by which A T P is regener- ated. When ATP is used to supply the energy for chemical reactions such as the heat-producing reactions described above, i t is broken down into adenosine diphosphate (ADP) and a phosphate in the process. Un- der normal circumstances a portion of the energy provided by ATP is used to drive the chemical reaction while the remainder of it is lost as heat or is recycled through oxidative phosphorylation. During that proc- ess, the phosphate is rebonded to ADP forming ATP. This is an en- ergy-conserving process which re- quires oxygen for its completion. If substances such as NE, thyroxine, or fatty acids"' interfere with (un- couple) the oxidative phosphoryla- tion of ADP, the excess energy re- leased by ATP will be lost as heat.
The contribution of each of the mechanisms to the total heat pro- duced by the cell is unknown. There is evidence that the Na' pump may contribute 60% of the heat pro- duced."
Conditions Necessary for BAT Thennogenesis
In addition to NE, thyroxine, and triglycerides, glucose and oxygen are needed to provide continued energy to the cell. If any of these com- ponents are missing or available in reduced quantities, BAT will fail to produce heat. For example:
1. Infants who do not have intact central nervous systems will have re- duced or absent thermoregulatory responses. Cross, ct al.ln studied an anencephalic infant who had an in- tact peripheral nervous system and a full complement of BAT. When cold-stressed, the infant's temper- ature decreased without any evi- dence of increased heat production. On autopsy, the BAT remained fat- filled unlike the BAT of infants dying of cold-stress who have intact central nervous systems. .It can be concluded that any defect which in- terferes with hypothalamic control of body temperature (such as myelo- meningocele or intracranial hemor- rhage) could ultimately lead to the inhibition of BAT thermogenesis.
2. Laboratory studies of animals
November/December 1980 JOCN Nursing
who are rendered hypothyroid show a decreased response of adipose tis- sue to norepinephrine." These find- ings indicate that thyroxine is neces- sary for an optimal thermogenic response in BAT.
3. The depletion of fat in BAT has been demonstrated in infants who are subjected to prolonged cold- stress. Once the fat has been de- pleted, the infant loses the ability to generate the heat necessary for bio- chemical processes and may sub- sequently succumb to the cold- stress.'"
4. Hypoglycemic infants are at risk of losing their thermoregulatory response to cold. Infants who are showing symptoms of hypoglycemia and who have a serum glucose of 20 mg/dl or less have lower-than-nor- ma1 metabolic rates and do not show the expected increase in metabolic rate in response to cooling of the skin .20
5. If the infant's PO, goes below 45-55 mmHg, his metabolic re- sponse to cold is impaired, whereas a PO, less than 30 mmHg abolishes the response."
Nursing Implications Although all infants are subject to
cold-stress through acts of care- lessness or ignorance, an understand- ing of the physiologic mechanisms underlying the infant's response to a cold environment will prepare the nurse to identify those infants who are particularly at risk of having thermoregulatory disturbances. To do so, the nurse needs to assess or be aware of the following:
1 . Gestational age: throughout the last trimester of pregnancy, fetal de- velopment includes not only the deposition of BAT but also the depo- sition of subcutaneous (white) fat and the storage of glycogen. Infants who are born before term, therefore, have a limited supply of BAT and glycogen to support thermogenesis and are not well-insulated by subcu- taneous fat against heat loss.
2. Neurologic status: infants who suf- fer from intracranial hemorrhage, birth asphyxia, or defects of the cen- tral nervous system may lose hypo- thalamic control of thermogenesis.
3. Endocrine status: infants who have hypothyroidism do not secrete normal amounts of thyroxine which
Novem ber/Decem ber 1960JOGN Nursing
CE EXTRACELLULAR
Norepinephrine -
0 0
0 0
Thyroxine ' \ \ \ \ \
Norepinephrine - K+-
+
Norepinephrine
Thyroxine 3-
LULAR C MEMBRANE
* Adenyl - N Cyclase
\
2 N a Pump
CYTOPLASM
ATP
* I cAMP+Lipase+Trigly-
ceride
ADP Fatty Acid
ATP Acetyl CoA
ATP
- Na' ADP - i o
Fatty Acids /
VllTOCHONDRl A
Uncoupling of Oxidative Phosphorylation
ADP + POqvATP
Figure 4. Proposed mechanisms of heat production in BAT. (ATP = adenosine tri- phosphate; ADP = adenosine diphosphate; CAMP = cyclic 3'5'nAMP.)
is necessary for maximum BAT heat production. Mothers treated with thiourea compounds during preg- nancy may give birth to infants with mild, transient hypothyroidism."
4. Cardio-respiratory status: any in- fant who is subject to a decrease in pOz, such as those with Respiratory Distress Syndrome or severe cyanotic heart disease, may have limited ther- mogenesis if the PO, goes below 55 mmHg.
5. Nutritional status: infants who for any reason have a delayed intake of the calories and nutrients neces- sary for the support of basal metabo- lism, growth, and activity are at risk of hypoglycemia, which can inhibit BAT catabolism and depress the metabolic rate.
6. Electrolyte status: infants with Na' and K' imbalance may have disturbed thermoregulation because of Na' pump failure.
7. Drug bfockage: certain drugs given to thc mother during her preg- nancy or labor can block the infant's thermoregulatory response. Diaze- pam in dosages greater than 30 mg given to mothers within 15 hours prior to delivery has been shown to impair an infant's response to cold stress for 24-48 hours after deliv- ery.':' Meperidine in dosages of 200- 400 mg given to mothers within 3-5
hours prior to delivery has been shown to cause lowered temper- atures in infants for as long as 20 hours a f te r birth.'* Infants of mothers who have received reserpine for hypertension may show thermal instability after birth." There are, most likely, other drugs which inter- fere with an infant's thermoregula- tory response. Any drug which sup- presses CNS function should be suspect, with the infant considered at risk for cold stress.
Once the nurse has identified the infants who are particularly at risk of becoming hypothermic, the goals in their care should be to limit the frequency and duration of cold stim- uli. The following interventions would be appropriate:
1 . The ambient and infant's skin temperatures should be monitored and charted at least hourly if the in- fant is undressed and in an in- cubator. Adamsons suggests that thermal balance can be achieved in a naked full-term infant if the am- bient temperature is 1.5OC less than the infant's skin temperature.2S This assumes that the skin temperature is within normal limits. In premature infants the difference would prob- ably be less.
2. Incubator portholes and doors should be opened only when neces-
371

sary. This requires wise care plan- ning and vigilance on the part of the nurse. In studies cited by Ahlgren, the temperature in a n incubator will drop from 35OC to 25.5OC in ap- proximately 4 minutes when the side door is opened with a temperature regain time of approximately 20-30 minutes. T h e opening of two port- holes for 5 minutes in a room tem- perature of 21.3OC caused the in- cubator temperature to drop from 35OC to 32.4OC with a regain time of 12 minutes.”’ Since t h e thermo- regulatory response to cold occurs in minutes, the nurse should try to work only through the portholes and wa tch for a m b i e n t t e m p e r a t u r e changes while doing so.
3. Oxygen and humidity delivered by a head hood should be warmed to prevent a cold stimulus to the facial skin receptors. Head hood temper- ature should be monitored hourly.
4. T h e evaporation of liquids from the skin lowers the skin temperature, so will act as a cold stimulus. A draft increases the rate of evaporation. It is imperative, therefore, that the in- fant’s skin be kept dry a n d that bathing be done in draft-free envi- ronments.
5. Prolonged skin contact with cold surfaces such as x-ray tables or metal scale beds should be avoided. If contact is necessary, a warm cloth should be placed between the infant a n d the cold surface.
6. Infants nursed in radiant warm- ers a r e at risk of convective a n d evaporative heat loss especially if the bed is placed in a draft. T h e infant is also a t risk of losing heat to cooler objects placed in the bed such as head hoods or heat shields. There are several measures the caretaker can institute to minimize these risks: a) Pre-warm head hoods and heat shields before placing them near the infant. b) Place infants in radiant warmers away from air-conditioning ducts. c) Keep the protective side- guards on the bed u p at all times to decrease cross-current airflow.
7. Infants nursed in incubators who do not require close observation of respiratory movements should be dressed in a shirt and diaper. T h e addition of clothing decreases the number of calories needed for heat conservation, making them available for growth.
372
In addi t ion to the above pre- cautions, the infant’s blood glucose a n d gases should be monitored at frequent intervals until the values a re within acceptable limits, a n d the infant’s required and actual caloric intake should be calculated daily to determine discrepancies.
Acknowledgment T h e author wishes to thank Anna
Tichy, RN, PhD, for her support and guidance.
References 1. Widdowson EM: Growth and com-
position of the fetus and newborn, Biology of Gestation, Vol 1 I . Edited by NS Assali. New York, Academic Press, Inc, 1968
2. Altman PL, Dittmer DS (eds): Biol- ogy Data Book, Vol 111. Bethesda, Maryland, Federation of American Societies for Experimental Biology, 1974, p 2041
3. Barnard T: Brown adipose tissue as an effector of nonshivering ther- mogenesis. Experientia 33: 1124- 1126, 1977
4. Dawkins MJR, Hull D: Brown fat and the response of the new-born rabbit to cold. J Physiol 169:101P, 1963
5. Blumenfeld J, Behrman RE: Cir- culatory and thermal homeostasis in the fetus and neonate, Current Top- ics in Clinical Chemistry-The Neo- nate. Edited by DS Young and JM Hicks. New York, John Wiley & Sons, 1976, pp 8 1-88
6. Aherne W, Hull D: The site of heat production in the newborn infant. Proc R Soc Med 57: 1 172- 1 173, 1964
7. Smith RE, Horwitz BA: Brown fat and therrnogenesis. Physiol Rev 49:330-425, 1969
8. Bligh J: Temperature Regulation in Mammals and Other Vertebrates. Amsterdam, North Holland Pub- lishing Company, 1973
9. Mestyan J, et al.: The significance of facial skin temperature in the chem- ical heat regulation of premature in- fants. Biol Neonate 7:243-254, 1964
10. Bullard RW: Temperature regula- tion, Physiology. Edited by EE Sel- kurt. Boston, Little, Brown, and Company, 1971, p 662
11. Benzinger TH: Heat regulation: homeostasis of central temperature in man. Physiol Rev 49:671-759, 1969
12. Sinclair JC: Metabolic rate and tem- perature control in the newborn, Perinatal Medicine. Edited by JW Goodwin, JO Godden, and GW
Chance. Baltimore, Williams & Wil- kins Co, 1976
13. Seydoux J, Girardier L: Control of brown fat thermogenesis by the sym- pathetic nervous system. Experientai 33:1128-1130, 1977
14. Krishna G, et al.: Effects of thyroid hormones on adenyl cyclase in adi- pose tissue and on free fatty acid mobilization. Proc Natl Acad Sci USA 59884-889, 1968
15. Edelman IS: Thyroid thermogenesis. N Engl J Med 290:1303-1308, 1974
16. Himms-Hagen J: Cellular ther- mogenesis. Annu Rev Physiol 38: 315-351, 1976
17. Horwitz BA: Quabain-sensitive component of brown fat thermo- gensis. Am J Physiol 224:352-355, 1973
8. Cross KW, et al.: Therrnoregulation in an anencephalic infant as inferred from its metabolic rate under hypo- thermic and normal conditions. Clin Sci 31:449-460, 1966
9. Hull D: The structure and function of brown adipose tissue. Br Med Bull 2292-96, 1966
20. Scopes JW, Ahmed I: Minimal rates of oxygen consumption in sick and premature newborn infants. Arch Dis Child 41:407-416, 1966
21. Scopes JW: Metabolic rate and tem- perature control in the human baby. Br Med Bull 22:88-91, 1966
22. Stevenson RE: The Fetus and Newly Born Infant. St. Louis, CV Mosby Company, 1973
23. Cree JE, et al.: Diazepam in Labour: its metabolism and effect on clinical course and thermogenesis of the newborn. Br Med J 4:251-255, 1973
24. Adamsons K Jr, Towell ME: Ther- mal homeostasis in the fetus and newborn. Anesthesiology 4:53 1-548, 1965
25. Adamsons K: The role of thermal factors in feral and neonatal life. Pediatr Clin North Am 13(3):599- 619, 1966
26. Ahlgren EW: Environmental control of the neonate receiving intensive care. Int Anesthesiol Clin 12(4):173- 215, 1974
Address correspondence to Virginia Davis, RN, 1944 W. Albion Ave., Chi- cago, IL 60626. ~
Virginia Davis is an instructor in the Pediatric Nursing graduate program at the Universiv of Illinois at the Medical Center in Chicago, and has extensive experience in various areas of pe- diatric nursing. Ms. Davis hold a diploma from Methodist Hospital School of Nursing in Indianapolis, Indiana, and a BSN and MSN Jiom the University of Illinois at the Medical Center.
November/December I980 JOCN Nursing