Embed Size (px)
Citation preview

ARTICLE IN PRESS
0277-3791/$ - se
doi:10.1016/j.qu
�Tel.: +1 517
E-mail addr
Quaternary Science Reviews 25 (2006) 263–281
The timing and nature of Late Quaternary vegetation changesin the northern Great Plains, USA and Canada:
a re-assessment of the spruce phase
Catherine H. Yansa�
Department of Geography, Michigan State University, East Lansing, MI 48824, USA
Received 17 September 2004; accepted 21 February 2005
Abstract
This paper revises the chronology for the northward migration of Picea glauca (white spruce) across the northern Great Plains,
following the recession of the Laurentide Ice Sheet, and reinterprets the species composition and structure of the late-glacial
vegetation on the basis of pollen and plant-macrofossil analysis. The timing of spruce migration is based on 26 14C ages obtained
from Picea macrofossils. The date for the appearance of white spruce in southern South Dakota, USA, remains unchanged, 12,60014C yrBP (ca 15,000 cal yr BP), but its arrival in southern Saskatchewan, Canada, by 10,300 14C yr BP (ca 12,100 cal yr BP) is about
1500 years later than previously estimated based on an organic sediment date. Picea glauca thus migrated northwards at an average
rate of 0.38 km/14C year (0.30 km/calendar year), significantly slower than the previously published rate of 2 km/14C year. White
spruce trees probably inhabited lake shorelines, whereas prairie, parkland, and boreal plants occupied both lowlands and uplands,
forming an open white spruce parkland. This interpretation differs from a previous reconstruction of a boreal-type spruce forest and
thus offers another paleoclimatic interpretation. Precipitation was probably low and summer temperatures relatively mild, averaging
about 19 1C.
r 2005 Elsevier Ltd. All rights reserved.
1. Introduction
Picea glauca (white spruce) is today an importantconstituent of the boreal forest biome in northern NorthAmerica. The southern range limit of this taxon atpresent follows the mean summer isotherm of 18 1C inCanada (Fig. 1), where the precipitation is greater than39 cm/year (Ritchie and Harrison, 1993). White sprucemay also be found at lower latitudes in high-elevationsettings, which have cool and humid climates similar tothat of the boreal forest. Numerous reports of sprucefossil deposits throughout North America indicate thatPicea glauca occupied areas farther south during full-glacial Wisconsinan time (21,000–14,000 14C yrBP,equivalent to 24,900–16,800 cal yrBP) (e.g., Webb, 1987;Jacobson et al., 1987). Populations close beyond the ice
e front matter r 2005 Elsevier Ltd. All rights reserved.
ascirev.2005.02.008
353 3910; fax: +1 517 432 1671.
ess: [email protected].
front were the probable seed sources for the spruce thatcolonized recently deglaciated terrain, following thenorthward recession of the Laurentide Ice Sheet (Bakerand Waln, 1985; Wright, 1987). The late-glacial north-ward migration of white spruce to form what laterbecame the boreal forest biome in Canada has been atopic of research by paleoecologists for decades (e.g.,Wright, 1968; Ritchie and MacDonald, 1986; Ritchie,1987; McLeod and MacDonald, 1997; MacDonald,2002).
The last survey of Late-Pleistocene white sprucemigration across the northern Great Plains was pro-vided by Ritchie and MacDonald (1986), who summar-ized previously published 14C chronologies. Accordingto their scenario, white spruce colonized southernmostSouth Dakota, USA, by 12,600 14C yrBP (ca 15,000cal yr BP) (Watts and Wright, 1966), arrived in southernSaskatchewan, Canada, by about 11,700 14C yr BP(ca 13,600 cal yr BP) (Ritchie and DeVries, 1964), and

ARTICLE IN PRESS
14,15
MONTANADAKOTANORTH
Missouri Coteau
Prairie Coteau
SWPicea glauca (modern range)
BH
BH Black Hills
CH Cypress Hills
SW Spruce Woods
CH
MINNESOTA
DAKOTASOUTH
MANITOBA
SASKATCHEWAN
ALBERTA
Missouri
River
Areashown
km
0 100 200
Lake Agassiz(maximum extent)
2
34
5
7
9
71
91
1110
0
12
16
18
1
6
8
13
2
21 222
Fig. 1. Locations of Late Pleistocene spruce fossil localities on the northern Great Plains in relation to physiography and the modern distributions of
Picea glauca in the boreal forest, Black Hills, Cypress Hills, and Spruce Woods. See Table 1 for information about these sites (#1–22).
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281264
reached what is today the southern limit of the borealforest in Manitoba, Canada, by 11,500 14C yr BP(ca 13,600 cal yr BP) (Ritchie, 1969). Ritchie and Mac-Donald’s (1986) chronology, which is based on 14C agesobtained from terrestrial plant macrofossils as well asorganic sediments, has been perpetuated in the literature(e.g., Baker and Waln, 1985; Ritchie, 1987; Barnoskyet al., 1987; Grimm, 2001; MacDonald, 2002; Webbet al., 2004). Organic sediments are well known toprovide erroneously older 14C ages (e.g., MacDonaldet al., 1991), so the chronology presented in this paper isbased solely on 14C ages obtained from terrestrial plantmacrofossils. This revised chronology dates the arrivalof white spruce in southern Saskatchewan about 1500calendar years later than previously thought. This paperalso provides a more realistic estimate of the possiblespeed of future white spruce migration northward as aconsequence of global warming, baring habitat frag-mentation and other anthropogenic factors.
This paper also presents the results of recentinvestigations of several pollen and plant-macrofossillocalities in North Dakota, USA, and southern Sas-katchewan, Canada, which provide a re-assessment ofthe species composition and structure of this anomalousLate-Pleistocene vegetation. Also used in this survey arepollen data published by other researchers and 14Cchronologies (based on terrestrial macrofossils) reportedfor late-glacial spruce fossil sites in the glaciatedportions of South Dakota and North Dakota as wellas southern Saskatchewan and southwestern Manitoba.This paleovegetation reconstruction has implications for
other regions of North America and Europe that havesimilar histories of deglaciation and Picea colonization.
2. Study area
The study area is that portion of the northern GreatPlains covered by the Laurentide Ice Sheet during thelast (late-Wisconsinan) glaciation, which can be roughlydefined as the area north and east of the Missouri River(Fig. 1). The topography of the study area is level torolling. The Missouri Coteau (700–800masl) and PrairieCoteau (530–625masl) uplands are hilly (hummockymoraine) with numerous kettle lakes and ponds (Fig. 1)(Clayton and Moran, 1982). To the east and northeastof these uplands, respectively, lies the Glaciated TillPlain (about 330–600masl), which has large shallowlacustrine basins that were once glacial lakes drained bymeltwater channels (Holliday et al., 2002). Farther eastis located the Red River Valley Lowland (about240–320masl), which was formerly occupied by glacialLake Agassiz (Fig. 1; Teller, 2004).
Plant and animal fossils of Late-Pleistocene andHolocene age are commonly found in these kettle lakesand other wetland sites. The regional water level wasvery high during deglaciation due to the melting ofglacial ice, but it lowered as a result of severe Holocenedroughts (Yansa, 1998; Grimm, 2001). Significant eolianand colluvial deposition in lakes and ponds during theHolocene partly in filled these basins, thereby reducingtheir water volumes (Yansa and Basinger, 1999; Grimm,

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 265
2001). Consequently, only a few lakes on the northernGreat Plains are deep enough to bear complete fossilsequences and the rest have become ephemeral wetlandsor saline playas (Barnosky et al., 1987; Grimm, 2001).Yansa (1998) demonstrated that some shallower lakeand pond basins contain well-preserved fossils in anoxicsediments below the water table, providing valuable,although truncated, fossil records dating from the Late-Pleistocene to the mid-Holocene. Intact fossil siteslocated farther south in the unglaciated portion of theGreat Plains are even rarer, because of the prevailingarid climatic regime throughout much of the Holocene.
The modern climate of the northern Great Plains ischaracterized by spatial and temporal variability inprecipitation and temperature, which affect not onlylake levels and chemistries but also vegetation. MeanJuly temperatures range from 24 1C in South Dakota to19 1C in southern Saskatchewan, with the north–southtemperature gradient being greater during the coldwinters (Owenby and Ezell, 1992; Environment Canada,1993). Precipitation grades from about 49 cm in easternNorth Dakota to 30 cm in eastern Montana because ofreduced westward penetration of the humid Gulf ofMexico air mass and a rainshadow effect of the RockyMountains (Bryson et al., 1970; Owenby and Ezell,1992).
This precipitation gradient is reflected in the nativegrassland communities of the northern Great Plains(which have been largely altered by agriculture andlivestock grazing), which grade east-to-west from tall-grass prairie to short-grass prairie. This paper focuseson the area characterized by mixed-grass prairie in theeastern Dakotas and southern Saskatchewan, which iscomposed of a mixture of tall (C3) and short (C4) grassesand forbs. Prairie marsh plants inhabit the numerousshallow wetlands and are adapted to fluctuating waterlevels (Kantrud, 1989). Moist soils along rivers andaround perennial (deep) lakes within the northern GreatPlains support Populus tremuloides (aspen poplar),Quercus macrocarpa (bur oak), and other tree and shrubtaxa more common in the forests to the east.
Stands of Populus and Betula (birch) are moreabundant farther north in the aspen parkland ofsouthern Manitoba, central Saskatchewan, and south-eastern Alberta, where the climate is slightly morehumid and cooler. The aspen parkland belt forms atransitional ecozone between the grassland and southernboreal forest. The tree flora of the southern portion ofthe boreal forest is composed of Picea glauca (whitespruce), Picea mariana (black spruce), Pinus banksiana
(jack pine), Abies balsamea (balsam fir), Larix laricina
(tamarack), Populus balsamifera (balsam poplar), Popu-
lus tremuloides (aspen poplar), and Betula papyrifera
(paper birch). The northern range limit of Fraxinus nigra
(black ash) occurs at the southeastern edge of the borealforest.
The absence of native white spruce stands on thenorthern Great Plains since the Late-Pleistocene hasbeen attributed to insufficient soil moisture and tosummer warmth, both of which inhibit white spruceseedling establishment and growth (Ritchie and Mac-Donald, 1986; MacDonald and Case, 2000). Today, therange of Picea glauca is restricted to the boreal forestand higher elevations of the Black Hills of SouthDakota (41250masl) and Cypress Hills of southernSaskatchewan and Alberta (41200masl) where theclimate is cool and moist compared to the adjacentplains (Fig. 1). One exception is the anomalous ‘‘SpruceWoods’’ population in southeastern Manitoba, wherestands of Picea glauca occupy a deltaic deposit of sand(along the paleoshoreline of glacial Lake Agassiz) withinthe aspen parkland, about 110 km south of the borealforest (Fig. 1).
3. Study design
3.1. Radiocarbon chronologies
The chronologies for several previous paleovegetationreconstructions, including an earlier assessment of thepostglacial spread of white spruce in northern NorthAmerica (i.e., Ritchie and MacDonald, 1986), aresuspected here to be too old, because they are basedon the 14C dating of organic sediments containingradioactively dead carbon. Other researchers havediscussed the problems associated with dating suchsediments (see Barnosky et al., 1987; MacDonald et al.,1991; Grimm and Jacobson, 2004). Radiocarbon agesobtained from organic sediments (including gyttja andmarl) differ from those acquired from the dating ofterrestrial plant macrofossils significantly, 1000–800014C years for paired samples from the same stratigraphiclevels (e.g., Clayton and Moran, 1982; MacDonald etal., 1991; Grimm, 2001).
Consequently, the chronology used in this paper isbased on the 14C dating of wood and spruce needles(Table 1). Several of these ages occur at 14C plateaus,which provide multiple possible calendar ‘‘dates’’ for asingle 14C age (Lowell and Teller, 1994). 14C ages areincluded in this paper along with calibrated datesobtained from the INTCAL98 dataset (Stuiver et al.,1998), so that the former may be recalibrated in thefuture when the calibration curve is further improved.
3.2. Fossil data sources
The dataset assembled here includes pollen percen-tages and plant-macrofossil abundances paired withreliable chronologies, which together document the firstarrival and spread of spruce across the northern GreatPlains of the USA and Canada. The ‘spruce rise,’ a
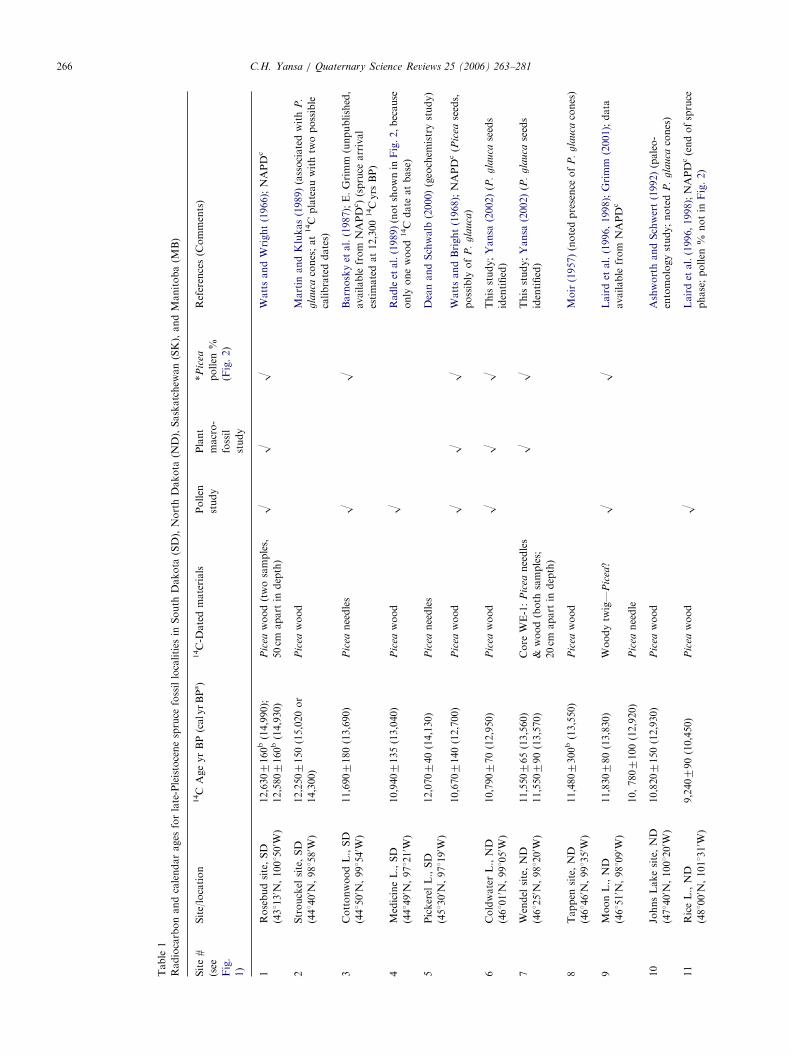
ARTICLE IN PRESS
Table
1
Radiocarbonandcalendarages
forlate-Pleistocenespruce
fossillocalities
inSouth
Dakota
(SD),NorthDakota
(ND),Saskatchew
an(SK),andManitoba(M
B)
Site#
(see
Fig.
1)
Site/location
14C
AgeyrBP(calyrBPa)
14C-D
atedmaterials
Pollen
study
Plant
macro-
fossil
study
*P
icea
pollen
%
(Fig.2)
References(C
omments)
1Rosebudsite,SD
(431130 N
,1001500 W
)
12,6307160b(14,990);
Pic
eawood(twosamples,
50cm
apart
indepth)
OO
OWattsandWright(1966);NAPD
c
12,5807160b(14,930)
2Strouckel
site,SD
(441400 N
,981580 W
)
12,2507150(15,020or
14,300)
Pic
eawood
Martin
andKlukas(1989)(associatedwith
P.
gla
uca
cones;at14C
plateauwithtw
opossible
calibrateddates)
3CottonwoodL.,SD
(441500 N
,991540 W
)
11,6907180(13,690)
Pic
eaneedles
OO
Barnoskyet
al.(1987);E.Grimm
(unpublished,
available
from
NAPD
c)(spruce
arrival
estimatedat12,300
14Cyrs
BP)
4MedicineL.,SD
(441490 N
,971210 W
)
10,9407135(13,040)
Pic
eawood
ORadleet
al.(1989)(notshownin
Fig.2,because
only
onewood
14C
date
atbase)
5Pickerel
L.,SD
(451300 N
,971190 W
)
12,070740(14,130)
Pic
eaneedles
DeanandSchwalb
(2000)(geochem
istrystudy)
10,6707140(12,700)
Pic
eawood
OO
OWattsandBright(1968);NAPD
c(P
icea
seeds,
possibly
of
P.
gla
uca)
6ColdwaterL.,ND
(461010 N
,991050 W
)
10,790770(12,950)
Pic
eawood
OO
OThisstudy;Yansa
(2002)(P
.g
lau
caseeds
identified)
7Wendel
site,ND
(461250 N
,981200 W
)
11,550765(13,560)
11,550790(13,570)
Core
WE-1:
Pic
eaneedles
&wood(both
samples;
20cm
apart
indepth)
OO
Thisstudy;Yansa
(2002)(P
.g
lau
caseeds
identified)
8Tappen
site,ND
(461460 N
,991350 W
)
11,4807300b(13,550)
Pic
eawood
Moir(1957)(notedpresence
of
P.
gla
uca
cones)
9MoonL.,ND
(461510 N
,981090 W
)
11,830780(13,830)
Woodytw
ig—
Pic
ea?
OO
Lairdet
al.(1996,1998);Grimm
(2001);data
available
from
NAPD
c
10,7807100(12,920)
Pic
eaneedle
10
JohnsLakesite,ND
(471400 N
,1001200 W
)
10,8207150(12,930)
Pic
eawood
AshworthandSchwert(1992)(paleo-
entomologystudy;noted
P.
gla
uca
cones)
11
RiceL.,ND
(481000 N
,1011310 W
)
9,240790(10,450)
Pic
eawood
OLairdet
al.(1996,1998);NAPD
c(endofspruce
phase;pollen
%notin
Fig.2)
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281266
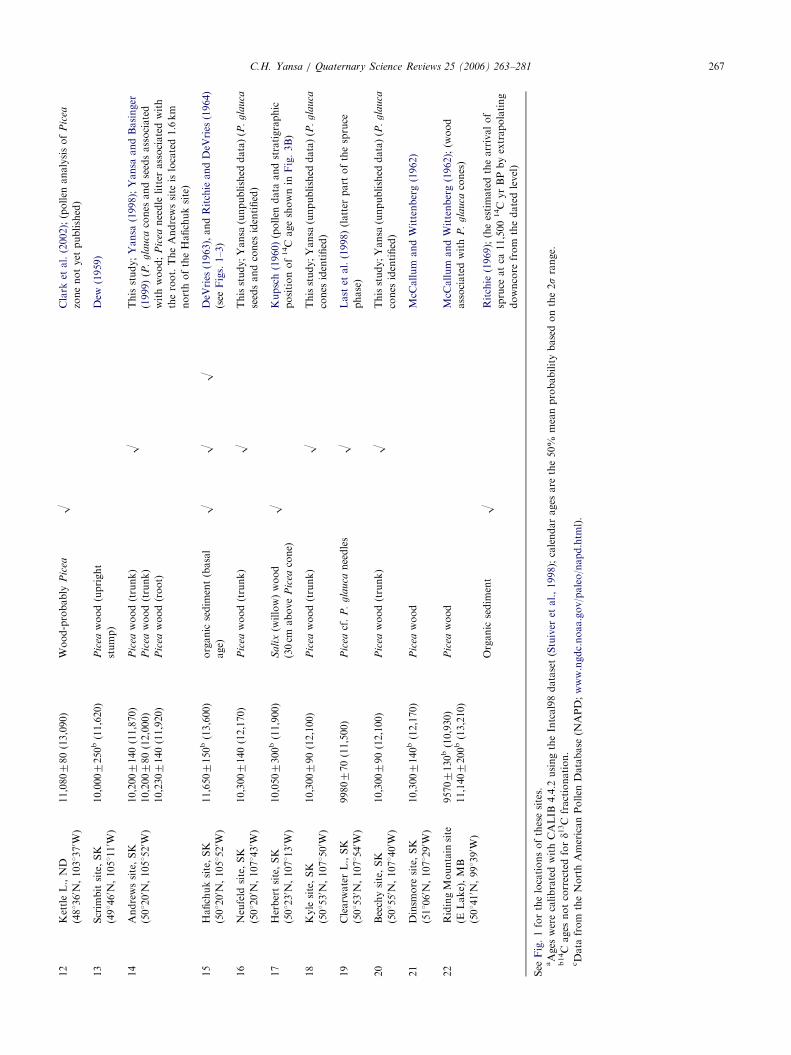
ARTICLE IN PRESS12
KettleL.,ND
(481360 N
,1031370 W
)
11,080780(13,090)
Wood-probably
Pic
eaO
Clark
etal.(2002);(pollen
analysisof
Pic
ea
zonenotyet
published)
13
Scrim
bitsite,SK
(491460 N
,1051110 W
)
10,0007250b(11,620)
Pic
eawood(upright
stump)
Dew
(1959)
14
Andrewssite,SK
(501200 N
,1051520 W
)
10,2007140(11,870)
10,200780(12,000)
10,2307140(11,920)
Pic
eawood(trunk)
OThisstudy;Yansa
(1998);Yansa
andBasinger
(1999)(P
.g
lau
cacones
andseedsassociated
withwood;
Pic
eaneedle
litter
associatedwith
theroot.TheAndrewssite
islocated1.6km
northoftheHafichuksite)
Pic
eawood(trunk)
Pic
eawood(root)
15
Hafichuksite,SK
(501200 N
,1051520 W
)
11,6507150b(13,600)
organic
sedim
ent(basal
age)
OO
ODeV
ries
(1963),andRitchieandDeV
ries
(1964)
(see
Figs.1–3)
16
Neufeld
site,SK
(501200 N
,1071430 W
)
10,3007140(12,170)
Pic
eawood(trunk)
OThisstudy;Yansa
(unpublished
data)(P
.g
lau
ca
seedsandcones
identified)
17
Herbertsite,SK
(501230 N
,1071130 W
)
10,0507300b(11,900)
Sa
lix(w
illow)wood
(30cm
above
Pic
eacone)
OKupsch(1960)(pollen
data
andstratigraphic
positionof14C
ageshownin
Fig.3B)
18
Kyle
site,SK
(501530 N
,1071500 W
)
10,300790(12,100)
Pic
eawood(trunk)
OThisstudy;Yansa
(unpublished
data)(P
.g
lau
ca
cones
identified)
19
Clearw
aterL.,SK
(501530 N
,1071540 W
)
9980770(11,500)
Pic
eacf.
P.
gla
uca
needles
OLast
etal.(1998)(latter
part
ofthespruce
phase)
20
Beechysite,SK
(501550 N
,1071400 W
)
10,300790(12,100)
Pic
eawood(trunk)
OThisstudy;Yansa
(unpublished
data)(P
.g
lau
ca
cones
identified)
21
Dinsm
ore
site,SK
(511060 N
,1071290 W
)
10,3007140b(12,170)
Pic
eawood
McC
allum
andWittenberg(1962)
22
RidingMountain
site
(ELake),MB
(501410 N
,991390 W
)
95707130b(10,930)
11,1407200b(13,210)
Pic
eawood
McC
allum
andWittenberg(1962);(w
ood
associatedwith
P.
gla
uca
cones)
Organic
sedim
ent
ORitchie
(1969);(heestimatedthearrivalof
spruce
atca
11,500
14C
yrBPbyextrapolating
downcore
from
thedatedlevel)
See
Fig.1forthelocationsofthesesites.
aAges
werecalibratedwithCALIB
4.4.2
usingtheIntcal98dataset(Stuiver
etal.,1998);calendarages
are
the50%
meanprobabilitybasedonthe2srange.
b14C
ages
notcorrectedford1
3C
fractionation.
cData
from
theNorthAmericanPollen
Database
(NAPD;www.ngdc.noaa.gov/paleo/napd.htm
l).
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 267
ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281268
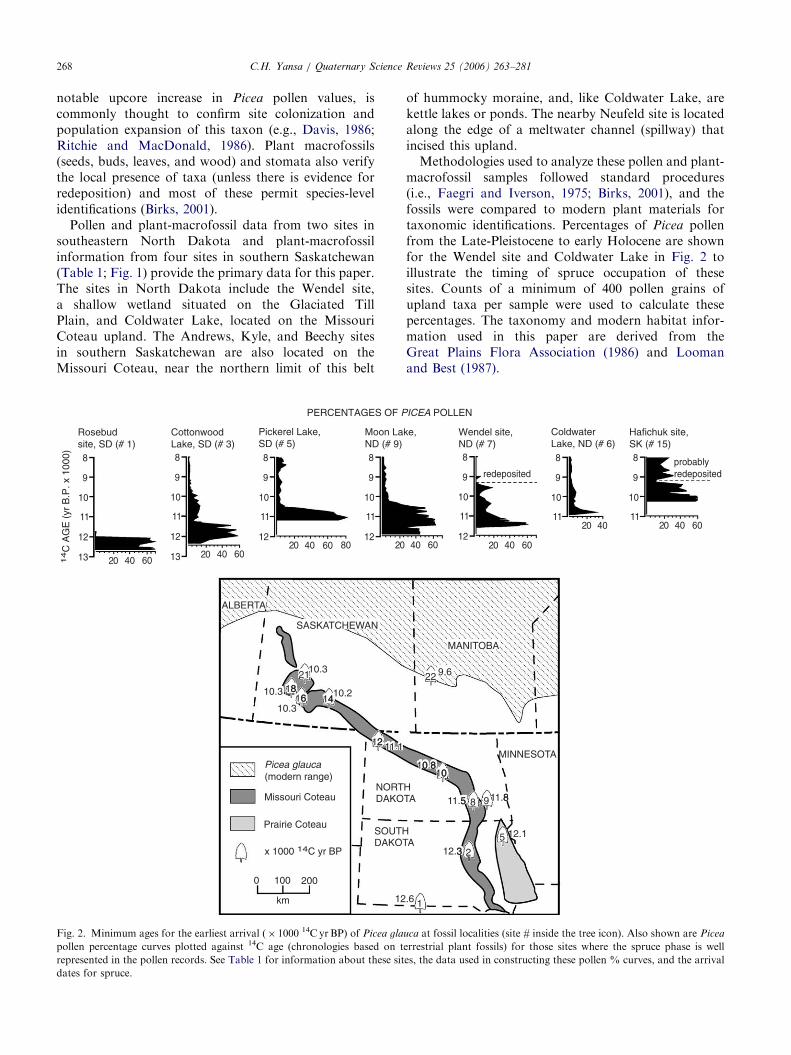
notable upcore increase in Picea pollen values, iscommonly thought to confirm site colonization andpopulation expansion of this taxon (e.g., Davis, 1986;Ritchie and MacDonald, 1986). Plant macrofossils(seeds, buds, leaves, and wood) and stomata also verifythe local presence of taxa (unless there is evidence forredeposition) and most of these permit species-levelidentifications (Birks, 2001).
Pollen and plant-macrofossil data from two sites insoutheastern North Dakota and plant-macrofossilinformation from four sites in southern Saskatchewan(Table 1; Fig. 1) provide the primary data for this paper.The sites in North Dakota include the Wendel site,a shallow wetland situated on the Glaciated TillPlain, and Coldwater Lake, located on the MissouriCoteau upland. The Andrews, Kyle, and Beechy sitesin southern Saskatchewan are also located on theMissouri Coteau, near the northern limit of this belt
PERCENTAGES OF P
14
C A
GE
(yr
B.P
. x 1
000)
10.2
DAKONORT
DAKOTSOUTH
10.3
12
11 1
10.3
10.3
20 40 60
12
11
10
9
8
13
Rosebudsite, SD (# 1)
CottonwoodLake, SD (# 3)
20 40 60
12
11
10
9
8
13
Pickerel Lake,SD (# 5)
20 40 60 8012
11
10
9
8
Moon LaND (# 9)
2012
11
10
9
8
ALBERTA
SASKATCHEWAN
km
0 100 200
Missouri Coteau
Prairie Coteau
Picea glauca (modern range)
122
x 1000 14C yr BP
1441618
68
21
Fig. 2. Minimum ages for the earliest arrival (� 1000 14C yrBP) of Picea gla
pollen percentage curves plotted against 14C age (chronologies based on t
represented in the pollen records. See Table 1 for information about these sit
dates for spruce.
of hummocky moraine, and, like Coldwater Lake, arekettle lakes or ponds. The nearby Neufeld site is locatedalong the edge of a meltwater channel (spillway) thatincised this upland.
Methodologies used to analyze these pollen and plant-macrofossil samples followed standard procedures(i.e., Faegri and Iverson, 1975; Birks, 2001), and thefossils were compared to modern plant materials fortaxonomic identifications. Percentages of Picea pollenfrom the Late-Pleistocene to early Holocene are shownfor the Wendel site and Coldwater Lake in Fig. 2 toillustrate the timing of spruce occupation of thesesites. Counts of a minimum of 400 pollen grains ofupland taxa per sample were used to calculate thesepercentages. The taxonomy and modern habitat infor-mation used in this paper are derived from theGreat Plains Flora Association (1986) and Loomanand Best (1987).
ICEA POLLEN
TAH
MINNESOTA
A
.6
12.33
12.1
11.811.55
10.80 8
ke,
40 60
Wendel site,ND (# 7)
20 40 6012
11
10
9
8
ColdwaterLake, ND (# 6)
20 4011
10
9
8
20 40 6011
10
9
8
Hafichuk site,SK (# 15)
probably redepositedredeposited
MANITOBA
9.6
2
5
9 88
100
22
1
uca at fossil localities (site # inside the tree icon). Also shown are Picea
errestrial plant fossils) for those sites where the spruce phase is well
es, the data used in constructing these pollen % curves, and the arrival
ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 269
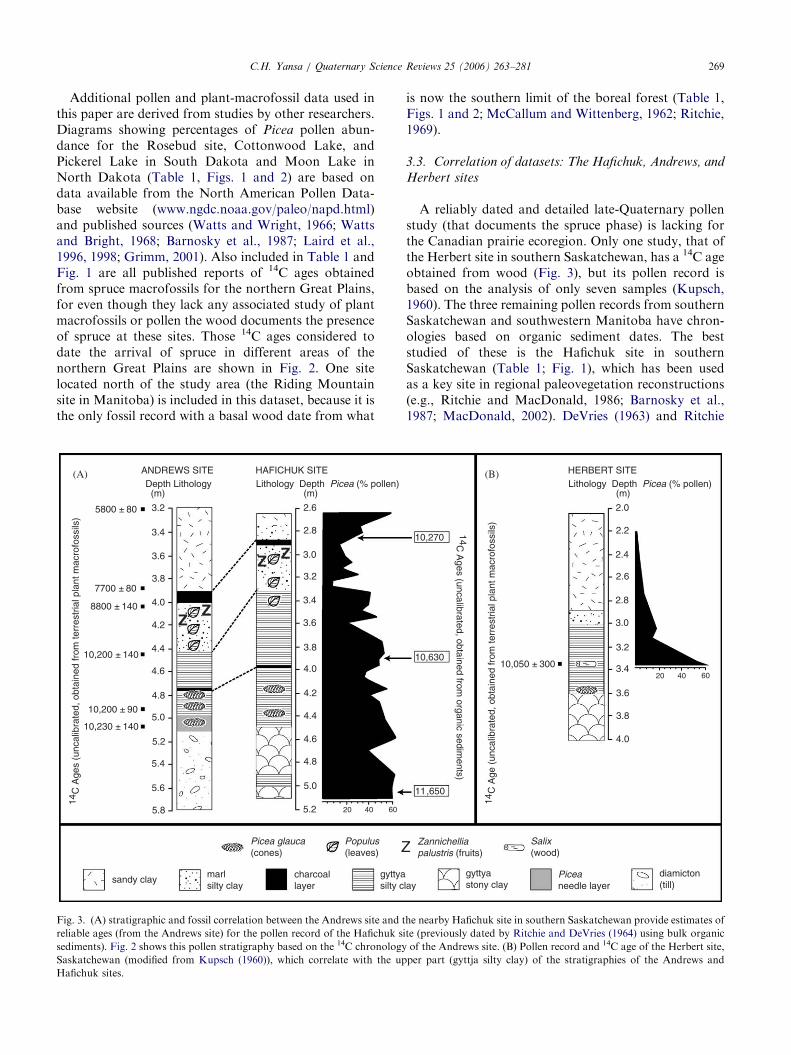
Additional pollen and plant-macrofossil data used inthis paper are derived from studies by other researchers.Diagrams showing percentages of Picea pollen abun-dance for the Rosebud site, Cottonwood Lake, andPickerel Lake in South Dakota and Moon Lake inNorth Dakota (Table 1, Figs. 1 and 2) are based ondata available from the North American Pollen Data-base website (www.ngdc.noaa.gov/paleo/napd.html)and published sources (Watts and Wright, 1966; Wattsand Bright, 1968; Barnosky et al., 1987; Laird et al.,1996, 1998; Grimm, 2001). Also included in Table 1 andFig. 1 are all published reports of 14C ages obtainedfrom spruce macrofossils for the northern Great Plains,for even though they lack any associated study of plantmacrofossils or pollen the wood documents the presenceof spruce at these sites. Those 14C ages considered todate the arrival of spruce in different areas of thenorthern Great Plains are shown in Fig. 2. One sitelocated north of the study area (the Riding Mountainsite in Manitoba) is included in this dataset, because it isthe only fossil record with a basal wood date from what
2.6
2.8
3.0
3.2
3.4
3.6
3.8
4.0
4.2
4.4
4.6
4.8
5.0
5.2
HAFICHUK SITE
3.2
3.6
3.4
3.8
4.0
4.2
4.4
4.6
4.8
5.0
5.2
5.4
5.6
5.8
5800 ± 80
7700 ± 80
8800 ± 140
10,200 ± 140
10,200 ± 90
10,230 ± 140
14C
Age
s (u
ncal
ibra
ted,
obt
aine
d fr
om te
rres
tria
l pla
nt m
acro
foss
ils)
Lithology Depth Picea (% pollen) (m)
Depth Lithology (m)
ANDREWS SITE
20 40 60
ZZ
Z Z
charcoallayer
sandy clay marlsilty clay
gyttyasilty c
(A)
Populus (leaves) ZPicea glauca
(cones)
Fig. 3. (A) stratigraphic and fossil correlation between the Andrews site and t
reliable ages (from the Andrews site) for the pollen record of the Hafichuk si
sediments). Fig. 2 shows this pollen stratigraphy based on the 14C chronology
Saskatchewan (modified from Kupsch (1960)), which correlate with the up
Hafichuk sites.
is now the southern limit of the boreal forest (Table 1,Figs. 1 and 2; McCallum and Wittenberg, 1962; Ritchie,1969).
3.3. Correlation of datasets: The Hafichuk, Andrews, and
Herbert sites
A reliably dated and detailed late-Quaternary pollenstudy (that documents the spruce phase) is lacking forthe Canadian prairie ecoregion. Only one study, that ofthe Herbert site in southern Saskatchewan, has a 14C ageobtained from wood (Fig. 3), but its pollen record isbased on the analysis of only seven samples (Kupsch,1960). The three remaining pollen records from southernSaskatchewan and southwestern Manitoba have chron-ologies based on organic sediment dates. The beststudied of these is the Hafichuk site in southernSaskatchewan (Table 1; Fig. 1), which has been usedas a key site in regional paleovegetation reconstructions(e.g., Ritchie and MacDonald, 1986; Barnosky et al.,1987; MacDonald, 2002). DeVries (1963) and Ritchie
10,270
10,630
11,650
14C A
ges (uncalibrated, obtained from organic sedim
ents)
diamicton(till)lay
Piceaneedle layer
gyttyastony clay
(B)
4.0
2.0
2.2
2.4
2.6
2.8
3.0
3.2
3.4
3.6
3.8
20 40 6010,050 ± 300
HERBERT SITELithology Depth Picea (% pollen) (m)
14C
Age
(un
calib
rate
d, o
btai
ned
from
terr
estr
ial p
lant
mac
rofo
ssils
)
Zannichellia palustris (fruits)
Salix (wood)
he nearby Hafichuk site in southern Saskatchewan provide estimates of
te (previously dated by Ritchie and DeVries (1964) using bulk organic
of the Andrews site. (B) Pollen record and 14C age of the Herbert site,
per part (gyttja silty clay) of the stratigraphies of the Andrews and

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281270
and DeVries (1964) interpreted the pollen, plantmacrofossils, and an organic-sediment date from theHafichuk site as indicating the establishment of aboreal-type spruce forest at 11,6507150 14C yrBP (ca13,600 cal yr BP). In contrast, 10 14C ages obtained fromwood and needles from eight nearby sites in southernSaskatchewan (Table 1; Fig. 2) indicate that insteadwhite spruce arrived in southern Saskatchewan by10,300–10,200 14C yrBP (12,100–11,870 cal yr BP) andpersisted until about 9980 14C yrBP (11,500 cal yr BP).
A revised chronology for the pollen record of theHafichuk site, shown in Fig. 2, is presented here basedon lithostratigraphic and fossil correlation with thereliably dated macrofossil record of the nearby Andrewssite (Fig. 3A) for which an available pollen record islacking. Correlation between these sites is robust, forthey are located only 1.6 km apart, share identicaltopographic settings (kettle ponds within the same trackof hummocky moraine), possess similar plant-macro-fossil records, and share several distinctive marker beds(charcoal layers, lithology, fossil beds, etc.) (Fig. 3A).The detailed descriptions by DeVries (1963) and Ritchieand DeVries (1964) of the sediment (MunsellTM color,texture, % organic carbon content) and fossils at 10-cmintervals for the Hafichuk site facilitated this correla-tion. Picea cones and Populus (poplar) leaves wereespecially useful for this correlation, as was the presenceof fruits of Zannichellia palustris (horned pondweed),which indicates increased salinity within these closedbasins (Kantrud, 1989; Yansa, 1998).
High Picea pollen values and abundant needles above2.9m at the Hafichuk site (Ritchie and DeVries, 1964)correlate with the mid-Holocene-age section of theAndrews site (Fig. 3A). These fossils were probablyeroded and redeposited during rainstorms followingdroughts, as have been interpreted for other sprucefossils recovered from mid-Holocene age sediments(Laird et al., 1998; Yansa, 2002). The stratigraphicsection sampled at the Hafichuk site is less complete(5.1m) than that of the Andrews site which extends intodiamicton (5.8m), so exact basal correlation betweenthese two sections is not possible. However, the bottomof the Hafichuk pollen record is unlikely to predate thebasal age of the Andrews site substantially, given theuniform geological setting and lithostratigraphy of thesenearby kettle sites.
This correlation (Fig. 3A) is further strengthened bycomparison with the pollen record of the Herbert site(Fig. 3B), which has one 14C age of 10,0507300 yr BP(11,900 cal yr BP) obtained from Salix (willow) woodpositioned 0.3m above a Picea glauca (white spruce)cone (Kupsch, 1960). The Herbert site pollen record,lithostratigraphy, and 14C date represent the latter partof the spruce phase (Fig. 3B), which roughly correlateswith the upper part of the gyttja silty clay units of boththe Hafichuk and Andrews sites (Fig. 3A). Peak values
of about 60% at the bases of both the Hafichuk andHerbert pollen records when correlated with thechronology of the Andrews site indicate that Picea hadarrived at these sites before 10,230 14C yrBP. AtCottonwood Lake, South Dakota, Picea pollen valuesincreased from about 15% (basal sample) to 60% over300 14C years (Fig. 2), which suggests that sprucearrived in southern Saskatchewan (Kyle, Beechy andNeufeld sites) several centuries before 10,300 14C yr BP(12,100 cal yr BP; Table 1). However, spruce did notappear in southern Saskatchewan as early as11,6507150 14C yrBP, as proposed by Ritchie andDeVries (1964) based on an organic sediment date, sincethis age predates spruce colonization of North Dakota(Fig. 2) and the currently accepted chronology forthe deglaciation of southern Saskatchewan by about11,300 14C yr BP (ca 13,250 cal yr BP) (Clayton andMoran, 1982).
4. White spruce migration, colonization, and extirpation
4.1. Deglaciation and soil development
The Late-Pleistocene was a time of rapid environ-mental changes on the northern Great Plains. The finalmelting of the Laurentide Ice Sheet created an unstablewater-saturated landscape. One would expect tundraherbs and shrubs to be the first vascular plants to havecolonized this recently deglaciated terrain, since ecolo-gical studies of modern glacial moraines, such as thosein the Yukon, have documented this stage in primaryplant succession (Birks, 1980). A pioneering tundra-likevegetation dating to the late-glacial has been reportedfrom pollen and plant-macrofossil studies conducted incentral Minnesota and eastward (e.g., Birks, 1976;Maher et al., 1998). In contrast, there is no evidencefor such a vegetation in the fossil records of the northernGreat Plains, and instead the earliest plant communityreconstructed was composed of white spruce trees,deciduous shrubs, and herbs.
A plausible explanation for the absence of tundrafossils is that during the early phases of deglaciationthere was a lack of stable depositional sites to preservethe remains of this pioneering vegetation (Florin andWright, 1969; Maher et al., 1998). Meltwater drainageon the Glaciated Till Plain and in the Lake Agassizbasin, as well as solifluction associated with the meltingof stagnant ice blocks buried under till on the Missouriand Prairie coteaus might not have been conducive forpreserving plant materials long after ice retreat (Wright,1976; Clayton and Moran, 1982). This tundra phasemay have been short-lived, since ecological studies ofplant colonization of newly formed moraines in theYukon indicate that succession from tundra to spruceforest can occur as quickly as 120 years (Birks, 1980).

ARTICLE IN PRESS
bedrock
tilltill
stagnant ice stagnant icelake sediment
Fig. 4. Schematic depiction of the open white spruce parkland which existed on the northern Great Plains during the late-glacial (adapted from
Tuthill et al., 1964). Picea glauca probably occupied the shores of lakes and ponds and the uplands were covered by shrubs and herbs. These plants
grew in till overlying buried stagnant ice, which slowly melted to form these waterbodies (not drawn to scale).
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 271
Fossil and lithostratigraphic data from the northernGreat Plains suggest that white spruce trees and otherplants became established in till overlying buriedstagnant ice on the Missouri and Prairie coteaus priorto and during the formation of lakes and ponds (Fig. 4).Wright (1976) described how this buried ice tookthousands of years to melt, during which time plantdetritus accumulated in temporary supraglacial poolsand were redeposited with the gradual downwasting ofthe ice beneath. Only when all of the ice melted didstable lake basins form, preserving spruce ‘‘trash layers’’at the bottom of many of these basins. In the case of theAndrews site in southern Saskatchewan, however, 10-cmPicea litter layer penetrated by in situ spruce roots waslaid down intact and flooded (Yansa, 1998).
The timing of basin establishment, which determinedthe starting point for preservation of plant remains,varied across the northern Great Plains, thereby makingthe exact arrival of spruce and other plants uncertain(Clayton and Moran, 1982; Yansa, 1998, 2002). Conse-quently, some sites within the region have morecomplete records of the spruce phase (e.g., Moon Lake)than others (e.g., Coldwater and Rice Lakes), and the14C dates for the spruce at all of these sites should beconsidered to be minimum ages (Table 1; Figs. 1 and 2).
The extent of soil development prior to spruceestablishment is unknown. Picea, however, is betterable to germinate and grow in mineral soils than mostother trees, because its roots can uptake nutrients fromnitrogen-fixing ectomychorrhizal fungi (Miller et al.,1994; MacDonald, 2002). Co-occurrences of fossilmychorrhizae and Picea root fragments from Late-Pleistocene sediments of the Andrews and Wendel sites(Yansa, unpublished data) indicate that this symbioticrelationship was also important during spruce coloniza-tion of the northern Great Plains.
4.2. Late-glacial climate
The changing range expansion of spruce on thenorthern Great Plains occurred during the transitionfrom glacial to interglacial climates. The cool climateduring deglaciation was offset by greater (�7%)summer insolation than modern, as determined fromastronomical calculations (e.g., Kutzbach and Webb,1991; Bartlein et al., 1998). Ritchie and MacDonald(1986) proposed that these relatively warm summerswould have aided in the seed germination and saplinggrowth of spruce. However, photosynthesis and hencegrowth rates of these plants were probably slowed by thelow partial pressure of carbon dioxide in the atmo-sphere, compared to modern conditions (Cowling andSykes, 1999).
The late-glacial climate was probably not uniformacross the northern Great Plains and neighboringregions. Rather, the northeastward retreat of theLaurentide Ice Sheet probably caused steep east–westand north–south thermal gradients across north-centralNorth America (Barnosky et al., 1987; Webb et al.,1987; Bartlein et al., 1998). The eastern part of thisregion, closer to the retreating glacier, probablyexperienced cooling both from albedo and from colddry winds generated from a persistent high-pressure cellcentered over the ice sheet (Barnosky et al., 1987;Kutzbach and Webb, 1991). This anticyclonic flow mayhave reduced the penetration of moist Gulf airmass intonorth-central North America from about 14,000 to10,000 14C yr BP (ca 16,800–11,600 cal yr BP) (Mac-Donald and Case, 2000; Hostetler et al., 2000). Theorientation of sand spits along the shore of a glacial lakein northern Michigan confirm the prevalence of east andsoutheast winds between 11,800 and 10,500 14C yr BP(ca 13,900–12,500 cal yr BP) just south of the Laurentide

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281272
Ice Sheet (Krist and Schaetzl, 2001). This anticycloniccirculation (with outward air flow in all directions)explains how the winged seeds of Picea glauca and otherplants were transported northwards into deglaciatedterrain (Ritchie and MacDonald, 1986). The remainderof the flora spread their seeds readily by interconnectingwaterways and by water fowl transporting seeds on theirmuddy feet.
Pollen data also support these paleoclimate inter-pretations. At the western limit of the northern GreatPlains, prairie colonized northwestern Montana soonafter deglaciation, indicating the presence of warmerand drier conditions away from the ice front (Barnoskyet al., 1987; Barnosky, 1989). Similarly, drought-tolerant Populus (poplar) has been identified as theearliest arboreal colonist in southeastern Alberta to thenorth (MacDonald and Case, 2000; Beaudoin andOetelaar, 2003). Thus, on the plains of Montana andsouthern Alberta, areas deglaciated first, the Late-Pleistocene vegetation directly responded to increasedsummer insolation shortly after deglaciation, comparedto areas farther east (Barnosky, 1989; MacDonald andCase, 2000). Picea never colonized these areas duringthe Late-Pleistocene, for evidence presented here putsthe western limit of this taxon on the Missouri Coteau.Spruce colonization of the eastern portion of northernGreat Plains, in other words, was a product of aglacially influenced climate, and the pace of its northernmigration was controlled primarily by the speed of ice-margin recession, the development of a suitable climate,and the rate of seed dispersal.
4.3. White spruce migration chronology
After deglaciation, Picea glauca migrated time-trans-gressively by moving northwards across the northernGreat Plains over 800 km from the South Dakota-Nebraska border to reach its present range in thesouthern boreal forest of Manitoba and Saskatchewan(Table 1, Figs. 1 and 2). Spruce occupied the areaimmediately south of the late-Wisconsinan ice limit, butthe extirpation of this taxon in Iowa, Kansas, andNebraska is poorly dated, occurring sometime duringthe late-glacial (Van Zant, 1979; Baker and Waln, 1985).Picea colonized the Rosebud site situated on the SouthDakota-Nebraska border (Table 1, Figs. 1 and 2),immediately south of the late-Wisconsinan glacial limit,by 12,600 14C yrBP (14,990 cal yr BP) (Watts andWright, 1966; Wright, 1992).
There are several spruce fossil localities within theglaciated portion of the northern Great Plains (sites areshown in Table 1 and Fig. 1 and dates for sprucecolonization are also depicted in Fig. 2). Plant macro-fossils verify the local occurrence of white spruce on theMissouri Coteau (Strouckel site) in northern SouthDakota at 12,250 14C yrBP (which has two possible
calendar ages of 15,020 and 14,300 yr BP, because of a14C plateau) (Martin and Klukas, 1989). Picea, probablyPicea glauca, inhabited the nearby Cottonwood Lakearea at about 12,300 14C yrBP (ca 14,300 cal yr BP)(Barnosky et al., 1987; Grimm, 2001). White sprucearrived on the Prairie Coteau upland in northeasternSouth Dakota (Pickerel Lake) by 12,070 14C yr BP(14,130 cal yr BP) (Dean and Schwalb, 2000). Picea alsodominated the earliest part of the pollen record ofnearby Medicine Lake (Radle et al., 1989), which has abasal wood date of 10,940 14C yrBP (13,040 cal yr BP).
Several localities contain spruce fossils in the MissouriCoteau of North Dakota. Picea occupied ColdwaterLake on the North Dakota–South Dakota border priorto 10,790 14C yrBP (12,950 cal yr BP), since only the tailend of the peak of spruce pollen is recorded at this site(Yansa, 2002). Picea must have colonized this part ofthe Missouri Coteau earlier, for farther north at theTappen site in central North Dakota an excavatedspruce stump dating to 11,480 14C yrBP (ca 13,550cal yr BP) was found in growth position and associatedwith cones of Picea glauca (Moir, 1957). Sprucecolonized the Johns Lake site in central North Dakotaby 10,820 14C yrBP (cal 12,930 yr BP), based on theidentification of Picea wood associated with conifer-bark beetle fossils (Ashworth and Schwert, 1992).Farther north, spruce occupied the shores of KettleLake by 11,080 14C yrBP (13,064 cal yr BP), and sprucepersisted at Rice Lake until at least 9240 14C yr BP(10,450 cal yr BP) (Laird et al., 1996, 1998; Grimm, 2001;Clark et al., 2002).
On the Glaciated Till Plain of North Dakota, east ofthe Missouri Coteau, spruce became established at theWendel site by 11,550 14C yr BP (ca 13,560 cal yr BP)(Yansa, 2002). Also in this area, spruce occupied thevicinity of Moon Lake by 11,830 14C yrBP (ca13,830 cal yr BP) (Laird et al., 1996, 1998). Farther east,meanwhile, glacial Lake Agassiz drained southward intothe Mississippi system (Teller, 2004), thereby restrictingthe northwestward migration of plants from the Mid-west. During a temporary low-water stand of this lake(caused by a drainage diversion), scattered standsof spruce and deciduous hardwoods colonized theswampy lake plain from 10,230 to 9900 14C yr BP(ca 11,960–11,320 cal yr BP) in the vicinity of Fargo,North Dakota (site not shown in Fig. 1; Yansa andAshworth, 2005).
In the Canadian portion of the northern Great Plains,Late-Pleistocene spruce fossil localities are restricted tothe Missouri Coteau, except for the Neufeld site (Table1, Figs. 1 and 2). Because the Neufeld site is located onthe edge of a meltwater channel that incised theMissouri Coteau, the wood was derived either fromthe coteau (where it was redeposited down slope) or thewood was rafted to the site by meltwater. Uprightspruce stumps recovered from the Scrimbit site (Dew,

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 273
1959) and the Andrews site (Yansa, 1998; Yansa andBasinger, 1999) provide conclusive evidence for treegrowth in situ at these sites. Radiocarbon ages from alleight spruce localities in southern Saskatchewan rangebetween 10,300 and 9980 14C yrBP (ca 12,100–11,500cal yr BP) (Table 1). The clustering of these ages doesnot necessarily indicate that the spread of spruce wasmore rapid in Saskatchewan than earlier in the Dakotas,since these ages occur at a well-documented 14C plateau(Lowell and Teller, 1994; Stuiver et al., 1998).
The plant-macrofossil records of the Andrews site(Yansa, 1998; Yansa and Basinger, 1999) and nearbyClearwater Lake (Last et al., 1998) and the pollen recordof the Herbert site (Fig. 3B; Kupsch, 1960) indicate thatspruce persisted in southern Saskatchewan until about9980 14C yrBP (ca 11,500 cal yr BP). A stump of spruceassociated with other macrofossils of Picea glauca froma site within the Riding Mountain National Park,Manitoba, dates to 95707130 14C yrBP (ca 10,930cal yr BP) (Table 1; McCallum and Wittenberg, 1962).This site, unlike the others mentioned, is located todaywithin the southernmost range limit of white spruce inthe boreal forest (Figs. 1 and 2). This basal wood date ofca 9600 14C yr BP (ca 10,950 cal yr BP) for the RidingMountain site (McCallum and Wittenberg, 1962)provides a minimum age for the initial formation ofwhat later became the southern boreal forest, because ofuncertainty whether the wood came from the earliestcolonists. However, this age is closer to the actual timingof spruce arrival than the age of ca 11,500 14C yr BP(ca 13,500 cal yr BP; Table 1) proposed by Ritchie (1969,1987) based on the dating of organic sediment from theRiding Mountain site (for the reasons discussed inSection 3.1). This age of 11,500 14C yrBP, furthermore,predates the accepted timing of deglaciation in southernManitoba (Clayton and Moran, 1982).
4.4. Estimated migration rate for white spruce
An average migration rate in 14C (and calendar) years(Table 2) was calculated based on the results of thisstudy (Table 1). The dates used in these calculations areminimum ages, since the wood samples used to obtainthese 14C ages may not have been from the first treeswhich colonized these sites. Another assumption is that
Table 2
Migration rate for Picea in both 14C and calendar years per kilometer, ass
Dakota, northwest to the Andrews site, Saskatchewan (based on data in Ta
Distance (km) between
(direction from the Ros
Rosebud site, SD to Andrews site, SKa 920 (northwest, �51 lon
aThe oldest ages were used for the Rosebud site (12,630 14C yrBP, equ
equivalent to 11,920 cal yr BP).
the spread of white spruce was more-or-less northwardsfrom populations south of the Laurentide Ice Sheet (i.e.,the Rosebud site area). This assumption is supported bygeomorphic evidence indicating that routes from theeast and southeast were blocked by glacial LakesDakota and Agassiz and associated meltwater streamsfrom about 12,000 to 10,900 14C yrBP (ca 14,000–13,000 cal yr BP) (Clayton and Moran, 1982; Teller,2004).
With these caveats in mind, spruce migrated from theSouth Dakota–Nebraska border (Rosebud site) north-west along the Missouri Coteau upland to reachsouthern Saskatchewan (Andrews site) at an averagerate of 0.38 km/14C year (0.30 km/calendar year)(Table 2). This rate is far less than the rate of2 km/14C year or about 1 km/calendar year previouslycalculated for the northward spread of white spruce innorth-central North America (Ritchie and MacDonald,1986; Bartlein et al., 1995). However, the rate presentedin this paper agrees with those obtained for theHolocene range expansions of spruce (0.25 km/14C year)and other coeval trees (0.1–0.5 km/cal yr) in easternNorth America (Davis, 1981; Grimm and Jacobson,2004), and is similar to the late-glacial migration rateof another species of Picea in Europe (Huntley andBirks, 1983).
The northward spread of white spruce on the north-ern Great Plains during the late-glacial was controllednot only by climate change, but also by the timing ofdeglaciation and the local availability of seeds. Conse-quently, it took about 3000 calendar years for late-glacial white spruce to spread 900 km. Even after takinginto account periodic lags in spruce migration, becauseof the influences of non-climate factors, this late-glacialmigration rate is still far slower than the predicted600–700 km northward advance of the boreal forestbiome over the next 50 years as a result of globalwarming (Kojima, 1994). The boreal forest biome willundoubtedly shift northwards in the future, but thismigration will probably lag behind warming tempera-tures, as in the past. Picea glauca will probably becomelocally extinct along its southernmost range, wheredeciduous trees will become more common, agreeingwith the predictions of other paleoecologists (e.g., Daviset al., 2000; MacDonald, 2002).
uming a constant speed of movement from the Rosebud site, South
ble 1)
sites
ebud site)
Picea migration rate
(km/14C yr)
Picea migration rate
(km/cal yr)
gitude) 0.38 0.30
ivalent to 14,990 cal yr BP) and the Andrews site (10,230 14C yrBP,

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281274
4.5. Open white spruce parkland
The late-glacial vegetation and climate interpretationspresented here for the northern Great Plains differ fromthose of other pollen and plant-macrofossil researchers.Some have described the species composition andstructure of this vegetation as being like the borealforest (Ritchie and DeVries, 1964; Watts and Bright,1968; Ritchie, 1987; Radle et al., 1989; Last et al., 1998).Other researchers have shied away from the term‘‘boreal forest,’’ because they recognized that thisLate-Pleistocene vegetation on the northern GreatPlains lacked Pinus banksiana (jack pine) and otherspecies which constitute this forest today, and insteadreconstructed a ‘‘spruce forest’’ for the northern GreatPlains (Watts and Wright, 1966; Laird et al., 1996, 1998;Grimm, 2001).
Ritchie and DeVries (1964) and Ritchie (1969, 1987)further considered this forest to have been closedcanopy, and to have included both Picea mariana (blackspruce) and Picea glauca. Their identification of Picea
mariana was based on the morphology of needles, butthis is considered by Baker and Drake (1994) to not be areliable character. All female cones and seeds of sprucerecovered from several sites on the northern GreatPlains have been identified as those of Picea glauca
(Table 1). Furthermore, Yansa and Basinger (1999)confirmed that white spruce was the only conifer at theAndrews site by stomatal analysis of several needlesfrom a buried litter layer. Differentiating between thesespecies has paleoenvironmental implications, for todaythese taxa typically inhabit different habitats. Picea
glauca prefers moderate to well-drained soils, whereasPicea mariana is more tolerant of both wet and acidicsoil conditions, such as in bogs (Ritchie, 1987; McLeodand MacDonald, 1997).
This paper reconstructs an open white spruce park-land (Fig. 4), rather than a spruce forest, based onseveral lines of evidence. First, the spatial coverage andabundance of white spruce in the Late-Pleistocenevegetation of the northern Great Plains were previouslyoverestimated. Typical pollen spruce maxima of40–60% for Late-Pleistocene sites within the region(see the pollen % curves in Fig. 2), should not becompared to the pollen spectra of modern forests, sincePicea glauca was the dominant, if not the sole, treeduring the late-glacial. Moreover, some of these sprucepollen grains were probably redeposited with otherdetritus as ‘‘trash layers’’ in kettle basins (described inSection 4.1), thereby inflating Picea values compared tothe regional pollen rain.
Associated with spruce pollen peaks are low values ofPinus (pine; o10%), which have been interpreted asderived from long-distance dispersal by this prolificpollen producer (Watts and Wright, 1966; Whitlocket al., 1993). Pollen values of Larix (tamarack) are low
(1–2%) and macrofossils of this taxon have only beenreported at a few sites (e.g., Watts and Bright, 1968).Pollen percentages of Fraxinus nigra-type (black ash) aremoderate (2–5%) for this species and indicate that blackash may have arrived sometime during the spruce phase,although macrofossils of this taxon have not yet beenrecovered (Yansa, 2002). The stratigraphic positions ofmacrofossils and pollen of Populus spp. (poplar) andBetula spp. (birch) in relation to those of spruce atseveral sites in the study area suggest that thesedeciduous trees arrived near the end of the spruce phase(Laird et al., 1998; Yansa, 1998).
Late-glacial white spruce dominated the arborealpollen spectra and swamped the pollen signal ofherbaceous plants, as observed today along forest-prairie ecotones (Baker and Drake, 1994). Non-arborealpollen (NAP) values of 20–40%, largely consisting ofArtemisia (sage), Poaceae (grass), Cyperaceae (sedgefamily) and, to a lesser extent, Ambrosia-type (ragweed),are significant for these late-glacial fossil sites. Thesevalues are comparable to those of the present-daygrassland and aspen parkland (Baker and Drake, 1994).
Second, macrofossils of the herbs and shrubs asso-ciated with those of white spruce from sites on thenorthern Great Plains have been previously under-appreciated or misinterpreted. Some of the plantmacrofossils identified at the Hafichuk site (DeVries,1963; Ritchie and DeVries, 1964), Pickerel Lake (Wattsand Bright, 1968), and Clearwater Lake (Last et al.,1998) were interpreted as boreal taxa. Upon closerexamination of the modern geographic ranges of thesetaxa, most of these ‘‘boreal’’ plants, such as the shrubArctostaphylos uva-ursi (bearberry), occupy not only theboreal forest but also the aspen parkland and mixed-grass prairie (see Table 3 for a complete list of speciesassociated with white spruce). In fact, the majority ofspecies identified from plant macrofossils at theselate-glacial sites today inhabit all three ecozones. Thisfossil assemblage includes only a handful of speciesrestricted today to the boreal forest and an equalnumber of species presently limited to the grasslandecozone (Table 3). This interpretation is supported byFredlund and Tieszen’s (1997) study of phytoliths andstable carbon isotopes in South Dakota, which recon-structed that a modern-like mixed-grass prairie becameestablished on the lower-elevation slopes of the BlackHills by 10,500 14C yrBP (ca 12,500 cal yr BP).
Third, additional data supporting this reconstructionof an open spruce parkland on the northern GreatPlains are provided by the limited abundance of buriedspruce logs, as well as coeval paleosols and mammothfossils. Excavations of entire kettle basins (�30mdiameter and �5m deep) at the Andrews, Kyle, andBeechy sites in Saskatchewan (for livestock-wateringholes) exposed about 3–5 logs or rooted stumps in eachbasin, suggesting that a few white spruce trees occupied

ARTICLE IN PRESS
Table 3
Presence (*) of taxa associated with white spruce at all plant-macrofossil localities investigated within the region
Macrofossil taxa Range Site (#)
1 5 6 7 14 15 16 18 19 20
Trees:
Betula sp. (birch) G, A, B *
Betula cf. B. occidentalis (river birch; water birch) G, A, B * * *
B. papyrifera (paper birch) G, A, B *
Larix laricina (tamarack) B *
Picea sp. (spruce) B * * * * * * * * * *
P. glauca (white spruce) A, B * * * * * * *
P. mariana (black spruce) B *
Populus tremuloides (aspen poplar; quaking aspen) G, A, B * * * *
Shrubs:
Andromeda polifolia (bog rosemary) B *
Arctostaphylos uva-ursi (bearberry) G, A, B * * *
Cornus canadensis (bunchberry) G, A, B *
C. stolonifera (C. alba) (red-osier dogwood) G, A *
Juniperus communis (low juniper; common juniper) G, A, B * *
Rubus idaeus (red raspberry) G, A, B * * * * * *
R. pubescens-type (dewberry; creeping blackberry) G, A, B * *
Salix sp. (willow) G, A, B *
Shepherdia canadensis (buffaloberry; rabbitberry) G, A, B * * *
Mudflat and Upland Herbs:
Aralia nudicaulis (wild sarsaparilla) G, A, B *
Aster sp. (aster) G, A, B * * *
Aster cf. A. novae-angliae (New England aster) G, A, B *
Asteraceae (aster family) G, A, B *
Brassicaceae (Cruciferae) (mustard family) G, A, B * *
Caltha palustris (marsh marigold) G, A, B *
Chenopodium berlandieri (pitseed goosefoot) G * * * * *
C. capitatum (strawberry blite) G, A, B *
C. rubrum (red goosefoot; alkali blite) G, A * * * * *
C. salinum (saline goosefoot) G, A * * * * * *
Cirsium sp. (thistle) G, A, B *
Cirsium cf. C. muticum (swamp thistle) G, A, B *
Epilobium sp. (willowherb) G, A, B *
Epilobium cf. E. ciliatum (northern fireweed) G, A, B *
Erigeron sp. (daisy fleabane) G, A, B *
E. philadelphicus (Philadelphia fleabane) G, A, B *
Fragaria vesca (American wild strawberry) G, A, B *
F. virginiana (wild strawberry) G, A, B * * * *
Geranium cf. G. bicknellii (Bicknell’s cranes-bill) G, A, B *
Lycopus americanus (water hoarhound) G, A, B * * * * * * *
Mentha arvensis (wild mint) G, A, B * * * * *
Poaceae (grass family) G, A, B * *
Polygonum sp. (smartweed) G, A, B *
Polygonum amphibium (water smartweed) G, A, B * *
P. lapathifolium (pale smartweed) G, A, B * *
Potentilla anserina (silverweed) G, A, B *
Potentilla gracilis (graceful cinquefoil) G, A, B *
Potentilla millegrana (P. rivalis) (brook cinquefoil) G *
Potentilla norvegica (rough cinquefoil) G, A, B * * * * * *
Potentilla palustris (marsh cinquefoil) A, B * *
Ranunculus gmelinii (small yellow buttercup) G *
Ranunculus pennsylvanicus (bristly buttercup) G, A, B *
Ranunculus sceleratus (ditch buttercup) G, A, B * * * * *
Ranunculus section Batrachium (buttercup) G, A, B * * *
Rorippa sp. (yellow cress) G, A, B *
Rorippa cf. R. truncata (yellow cress) G, A, B * *
Rumex maritimus (golden dock) G, A, B * * * * * * * *
Solidago sp. (goldenrod) G, A, B *
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 275

ARTICLE IN PRESS
Table 3 (continued )
Macrofossil taxa Range Site (#)
1 5 6 7 14 15 16 18 19 20
Stachys sp. (hedge nettle) G, A, B * *
Stellaria crassifolia (fleshy stitchwort) G, A, B *
Triglochin maritima (seaside arrow-grass) G, A, B * * *
Urtica gracilis-type (U. dioica) (stinging nettle) G, A, B * *
Viola sp. (violet) G, A, B * *
Aquatic-Emergents:
Alisma– planto-aquatica (water-plantain) G, A, B * *
Carex sp. (sedge) G, A, B * *
Carex aquatilis (water sedge) G, A, B * * *
Carex cf. C. atherodes (awned sedge) G, A, B * * * *
Carex cf. C. aurea (golden sedge) G, A, B *
Carex cf. C. lenticularis (lens-fruited sedge) B *
Carex cf. C. rostrata (beaked sedge) G, A, B * * * * * *
Carex cf. C. sychnocephala (long-beaked sedge) G, A, B * * * *
Carex retrosa (turned sedge) G, A, B *
Carex tenera (slender sedge) G *
Carex vulpinoidea (fox sedge) G, A, B *
Eleocharis sp. (spike-rush) G, A, B *
Juncus sp. (rush) G, A, B *
Sagittaria latifolia (broad-leaved arrowhead) G, A *
Sagittaria rigida (sessile-fruit arrowhead) G *
Scirpus americanus (three-square bulrush) G, A * * * * * *
S. cyperinus (wool-grass) G, A, B *
S. nevadensis (Nevada bulrush) G, A *
S. paludosus (prairie bulrush) G, A, B *
S. validus (common bulrush) G, A, B * * * * * * * *
S. cf. S. atrovirens (green bulrush) G, A *
Sparganium sp. (bur-reed) G, A, B * *
Sparganium chlorocarpum (stemless bur-reed) G, A, B * *
Subularia aquatica (water awl-wort) B * *
Typha latifolia (broad-leaved cattail) G, A, B * * * * * * * *
Aquatics:
Hippuris vulgaris (mare’s tail) G, A, B * * * *
Lemna minor (lesser duckweed) G, A, B *
Lemna trisulca (ivy-leaved duckweed) G, A, B * * * *
Myriophyllum sp. (water-milfoil) G, A, B * * * *
M. verticulatum (whorled water-milfoil) G, A, B * * * *
Najas flexilis (slender naiad) G, A, B * * * *
Potamogeton filiformis (slender pondweed) G, A, B * * * * * *
P. gramineus (variable pondweed) G, A, B *
P. natans (floating-leaved pondweed) G, A, B *
P. obtusifolius (blunt-leaved pondweed) A, B * *
P. pectinatus (sago pondweed) G, A, B * * * * *
P. pusillus (P. berchtoldii) (small pondweed) G, A, B *
P. richardsonii (Richardson’s pondweed) G, A, B * *
P. strictifolius (straight-leaved pondweed) G
P. vaginatus (sheathed pondweed) G, A, B * * *
P. vaseyi (Vasey’s pondweed) G, A, B *
P. cf. P. foliosus (leafy pondweed) G, A, B *
Utricularia vulgaris-type (greater bladderwort) G, A, B *
Wolffia arrhiza (water-meal) G, A, B * *
Non-Vascular Plants:
Chara sp. (green alga) G, A, B * * * * * * *
Drepanocladus sp. (sickle-branch moss) G, A, B * * * * * *
Equisetum sp. (horsetail) G, A, B * * *
Included are the geographic ranges where these taxa currently exist: grassland (G); aspen parkland (A); and boreal forest (B). See Table 1 and Fig. 1
for the names and locations of these sites.
C.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281276

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 277
the shorelines of each of these ponds. This paleovegeta-tion interpretation is supported by the lack of spodichorizon development in the sediments underlying spruce-needle layers at the Andrews, Kyle, and Beechy sites(Yansa, 1998; unpublished data) and in the buried soilsof eastern North Dakota (Hopkins and Running, 2000).Moreover, the recovery of Mammuthus primigenius
(woolly mammoth) fossils on the northern Great Plains,some dating to about 11,500 14C yrBP (Harington andAshworth, 1986), have an autecology that agrees withthe interpretation presented here for uplands coveredprimarily by herbaceous plants.
The vegetation that colonized the northern GreatPlains after deglaciation was composed of an unusualassemblage of prairie, aspen parkland, and borealplants, forming a spruce parkland environment. Thisparkland interpretation was originally proposed for theTappen site (Moir, 1957) and a mollusk site (Tuthillet al., 1964) in North Dakota and for the Herbert site inSaskatchewan (Kupsch, 1960), but it was supplanted bythe more popular reconstruction of a boreal forest byRitchie (1969, 1987) and others. This paper supportsthe spruce parkland interpretation, which is depictedin Fig. 4.
This paper is also in agreement with the interpretationof an anomalous parkland of Picea, Fraxinus nigra
(black ash), Artemisia, and Cyperaceae occupying theeastern USA during the late-glacial (Webb et al., 1983,2004; Webb, 1987; Jacobson et al., 1987; Williams et al.,2001). Their reconstruction of scattered spruce treessurrounding numerous sedge wetlands is identical tothat presented in this paper for the northern GreatPlains during the same time period, the only exceptionbeing that Fraxinus nigra pollen values are higher forsites in the eastern USA. Furthermore, both of theseinterpretations differ from the ‘‘classic’’ interpretationof a spruce forest comprising the late-glacial vegetationof the eastern USA (e.g., Wright, 1968; Davis, 1981;Delcourt and Delcourt, 1991; Whitlock et al., 1993), andthe northern Great Plains (e.g., Ritchie, 1969, 1987;Laird et al., 1996, 1998; Grimm, 2001).
The absence of a suite of plants now restricted to theboreal forest in the fossil records of the northern GreatPlains (Table 3) indicates that the climate was not socold as proposed in an earlier interpretation of a borealforest (Ritchie and DeVries, 1964; Ritchie, 1987;MacDonald, 2002). Similarly, Grimm and Jacobson(2004) proposed that during the late-glacial spruce phasein the Midwest the regional climate was somewhatwarmer than that of the boreal forest today. There is noevidence for Younger Dryas cooling in the fossil recordsof the northern Great Plains, although such coolinghas been reported in the eastern USA (e.g., Shumanet al., 2002).
An exact modern analog is lacking for the open whitespruce parkland that once existed on the northern Great
Plains. The closest facsimile, however, is the vegetationof the Spruce Woods Provincial Park of southernManitoba, where pockets of Picea glauca occupy low-lying locations surrounded by upland prairie within theaspen parkland belt. Picea glauca is more droughttolerant than Picea mariana (black spruce) and Pinus
banksiana (jack pine) (Ritchie, 1987), which may explainwhy the former, rather than one of the latter, colonizedthe northern Great Plains under a low precipitationregime (as discussed above in Section 4.2). White sprucetrees may have supplemented their moisture require-ments from the moist lakeshore soils during the late-glacial (Fig. 4).
The modern climate of the Spruce Woods and aspenparkland of southern Manitoba and south-centralSaskatchewan (Shay, 1999) provides the best approx-imation for the late-glacial climate of the Dakotasand Canadian prairies based on the fossils identified(Table 3) and their relative abundances. The meansummer temperature during the late-glacial in thisregion was probably about 19 1C, a degree warmer thanthat of the southern limit of the boreal forest today. Thispaleotemperature reconstruction is also 3 1C and 5 1Ccooler than present-day mean July temperatures inNorth Dakota and South Dakota, respectively (Owenbyand Ezell, 1992).
4.6. White spruce extirpation and plant-community
succession
The late-Quaternary fossil records of the northernGreat Plains suggest that during the Pleistocene–Holo-cene transition Picea glauca was replaced by deciduoushardwoods, primarily Ulmus (elm) and Quercus (oak) inthe Dakotas, and Populus (poplar) and Betula (birch) inSaskatchewan (Grimm, 2001; Yansa, 2002). The ex-tirpation of spruce was more-or-less time-transgressive,occurring at: 11,200 14C yr BP (ca 13,200 cal yr BP) innorthern South Dakota (Cottonwood Lake; Barnoskyet al., 1987); 10,600 14C yrBP (ca 12,750 cal yr BP) insouthernmost North Dakota (Coldwater Lake; Yansa,2002); 10,300 14C yrBP (ca 12,100 cal yr BP) in south-eastern North Dakota (Moon Lake; Laird et al., 1996,1998); and about 9980 14C yrBP (ca 11,500 cal yr BP) insouthern Saskatchewan (Clearwater Lake; Last et al.,1998).
The shift from spruce parkland to deciduous parklandon the northern Great Plains occurred at about the sametime as the transition from spruce parkland (or forest) topine woodland in neighboring Minnesota to the east(Shay, 1967; Whitlock et al., 1993; Williams et al., 2001).These regional vegetation changes have been attributedto greater summer warmth and reduced soil moisture,which would have desiccated white spruce seedlingsand stressed older trees (e.g., Bartlein et al., 1995;Grimm and Jacobson, 2004). Summer droughts became

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281278
common during the early and mid-Holocene in north-central North America. They were probably caused byzonal westerly (Pacific) airflow blocking the penetrationof moist Gulf air (Kutzbach and Webb, 1991; Dean andSchwalb, 2000).
Besides climate, the drainage and evaporation ofmeltwater on the landscape during the late-glacial andearly-postglacial would also have been responsible forthe drying of soils within the region. On the broaderscale, the regional water table lowered, affecting thevegetation. On the local scale, basins within an areareceived variable groundwater inflow depending upon(1) their landscape positions relative to the water table(Smith et al., 1997), and (2) volumes of stagnant iceblocks buried under local tills, the meltwater of whichseeped into overlying basins over 1000–3000 years(Wright, 1976; Clayton and Moran, 1982). Yansa(2002) referred to this as the ‘‘residual meltwater effect,’’because it probably supported trees around waterbodiesindependently of the climatic regime for a time.
These changes in climate and hydrology during thePleistocene–Holocene transition would have favoredgreater fire frequency. Charred Picea logs and needles atthe Andrews site (Yansa, 1998), and reports of abundantcharcoal associated with declining spruce pollen valuesat several other sites (Watts and Bright, 1968; Yansa,2002; Clark et al., 2002) suggest that fire probablycontributed to the demise of spruce trees at these fossillocalities. Species of Populus and Betula colonized theseburned areas, as seen today after fires and otherdisturbances in the boreal forest and aspen parkland.
Although these fires reflected a regional drying trendduring the transition from the late-glacial to early-postglacial, they were probably localized. Spruce per-sisted at Rice Lake, for example, until shortly after 924014C yrBP (ca 10,450 cal yr BP) (Laird et al., 1996, 1998),long after it disappeared from nearby sites (Table 1).The timing of Picea glauca extirpation varies from site-to-site because these trees are long-lived (up to 500years) in the absence of local fires and other distur-bances (Szeicz and MacDonald, 1994). White sprucetrees once established can also tolerate warm summers(418 1C mean July temperatures) so long as precipita-tion or soil moisture is high (Webb et al., 1993). Grimmand Jacobson (2004) suggested that a moist climate andlack of fires favored Picea during the late-glacial.Alternatively, I contend that it was not high precipita-tion but a meltwater-saturated landscape that inhibitedfires and prolonged spruce growth during this period.Only when the landscape began to dry out in the early-Holocene under a warm-dry climatic regime did firesbecame common and deciduous trees appear. Thearrival of deciduous trees has also been correlated withlowering of the local water tables and higher lakesalinities in the Dakotas and Saskatchewan (e.g., Last etal., 1998; Dean and Schwalb, 2000).
White spruce therefore became extinct on the north-ern Great Plains for a variety of reasons, many of whichare inter-related. These causal agents probably includeddesiccation, fire, and being outcompeted by deciduoustrees for limited resources. The prairie grasses and forbsthat had occupied the drier soils during the spruce phaseprobably expanded in their coverage during this regionalwarming and drying trend. This aridity continued, andabout a millennium later the deciduous trees disap-peared and prairie expanded to form a widespreadgrassland, as seen today in this region (Laird et al., 1996,1998; Yansa, 1998, 2002; Grimm, 2001).
5. Summary
This paper reassesses the timing and communitycomposition of the white spruce vegetation thatcolonized and spread across the northern Great Plainswith the recession of the Laurentide Ice Sheet. Availablefossil data indicate that white spruce was the mainconstituent of this vegetation along with prairie, park-land, and boreal shrubs and herbs and that a precedingtundra-like vegetation was absent. This paper confirmsthe results of other pollen researchers that the north-ward spread of white spruce was time-transgressive butoffers a more accurate chronology for this migrationbased on the 14C dating of terrestrial plant macrofossils.Previous chronologies had integrated 14C ages obtainedfrom spruce wood with those ages obtained fromorganic sediments (gyttja).
White spruce existed on the South Dakota-Nebraskaborder, immediately south of the Late-Wisconsinanglacial limit, at 12,600 14C yrBP (ca 15,000 cal yr BP).The onset of Picea glauca migration into glaciated SouthDakota is poorly dated. However, by 12,250 14C yr BP(15,020–14,300 cal yr BP) white spruce had reachednorthern South Dakota, by 10,820 14C yrBP (12,930 calyr BP) had arrived in central North Dakota, by 10,30014C yrBP (12,100 cal yr BP) had migrated to southernSaskatchewan, and sometime before 9570 14C yr BP (ca10,930 cal yr BP) reached what is now the southernboreal forest in southern Manitoba. White sprucemigrated northwest along the Missouri Coteau uplandinto southern Saskatchewan at an estimated rate of0.38 km/14C year (0.30 km/calendar year). This rate issubstantially slower than the previously published rateof 2 km/14C year (1 km/calendar year) for the postglacialspread of white spruce in the study area, but iscomparable to published rates for Holocene treemigration in eastern North America.
In addition, this paper proposes that this vegetationwas an open white spruce parkland, rather than a spruceforest or boreal forest, as previously interpreted by otherresearchers. Evidence supporting a more open characterto this late-glacial vegetation includes: (1) Picea pollen

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 279
percentages being inflated, compared to modern pollenspectra, due to the low species diversity of this unusuallate-glacial vegetation and redeposition of spruce fossilsduring the formation of kettle basins; (2) absence of a‘‘boreal’’ macrofossil flora and the presence of severalgrassland and aspen parkland plants; (3) spruce logs andtrunks exposed during excavations of kettle pondsbeing few in number; (4) the lack of buried Spodosols,which would have formed under coniferous forestcover; and (5) localities of grazing woolly mammoth ofages comparable to those of white spruce. An openwhite spruce parkland was previously interpreted bya few other authors decades ago, but this paper isthe first to reconstruct this vegetation using detailedplant-fossil analysis. Moreover, this paleovegetationreconstruction concurs with an interpretation recentlyproposed by other pollen researchers for a spruce-sedgeparkland existing in the eastern USA during thesame time.
This interpretation of a spruce parkland on thenorthern Great Plains agrees with paleoclimate modelsimulations for reduced precipitation during the late-glacial. White spruce trees today inhabit well-drainedsoils, but during the late-glacial they apparently wererestricted to the shorelines of lakes and ponds where soilmoisture may have offset the limited precipitationreceived. The association of prairie and parkland shrubsand herbs with spruce trees and a few boreal plants alsosuggests that the temperatures were not so cold asproposed earlier for a boreal forest, but were still coolerthan present on the northern Great Plains, as expectedfor a glacially influenced climate. Mean summertemperatures on the northern Great Plains during thelate-glacial were probably about 19 1C, like that of apocket of white spruce parkland which exists todaywithin the aspen parkland ecozone of southern Mani-toba. Continued warming during the early-Holoceneresulted in the extirpation of white spruce in this region,starting before 11,200 14C yrBP (ca 13,200 cal yr BP) inSouth Dakota and ending at about 9980 14C yrBP (ca11,500 cal yr BP) in southern Saskatchewan. This sprucevegetation was replaced by a parkland of deciduoustrees and prairie, which about a millennium later wassupplanted by extensive grassland that comprises thenative vegetation of this region today.
Acknowledgments
Reviews of this paper by Tom Webb III and HerbWright Jr. have been very helpful in improving the finalproduct. Earlier versions of this paper have benefitedfrom the comments of Vance Holliday, Jim Knox, LeeClayton, Ben LePage, and Jim Basinger. I also thankEric Grimm for providing the sediment cores fromColdwater Lake and Pietra Mueller for conducting
pollen counts on some of the samples from these cores.As well, I acknowledge Ed Murphy for coring theWendel site and Dennis Wendel for allowing us access tothis site. The Palliser Triangle Global Change Project(Geological Survey of Canada) provided funds to collectsamples from the Andrews, Beechy, Kyle, and Neufeldsites and to obtain 14C ages for fossils from these sites.The 14C ages for the Wendel site and some of thoseobtained for Coldwater Lake were provided free by theUniversity of Arizona AMS 14C Laboratory.
References
Ashworth, A.C., Schwert, D.P., 1992. The Johns Lake site: A late
Quaternary fossil beetle (Coleoptera) assemblage from the Mis-
souri Coteau of North Dakota. North Dakota Geological Survey
Miscellaneous Series No. 76, pp. 257–265.
Baker, R.G., Drake, P., 1994. Holocene history of prairie in
midwestern United States: pollen versus plant macrofossils.
Ecoscience 1, 333–339.
Baker, R.G., Waln, K.A., 1985. Quaternary pollen records from the
Great Plains and central United States. In: Bryant, Jr., V.M.,
Holloway, R.C. (Eds.), Pollen Records of Late-Quaternary North
American Sediments. American Association of Stratigraphic
Palynologists Foundation, Texas, USA, pp. 191–203.
Barnosky, C.W., 1989. Postglacial vegetation and climate in the
northwestern Great Plains of Montana. Quaternary Research 31,
57–73.
Barnosky, C.W., Grimm, E.C., Wright Jr., H.E., 1987. Towards a
postglacial history of the northern Great Plains: a review of the
paleoecologic problems. Annals of Carnegie Museum 56 (article
16), 259–273.
Bartlein, P.J., Edwards, M.E., Shafer, S.L., Barker Jr., E.D., 1995.
Calibration of radiocarbon ages and the interpretation of
paleoenvironmental records. Quaternary Research 44, 417–424.
Bartlein, P.J., Anderson, K.H., Anderson, P.M., Edwards, M.E.,
Mock, C.J., Thompson, R.S., Webb, R.S., Webb III, T., Whitlock,
C., 1998. Paleoclimate simulations for North America over the past
21,000 years: Features of the simulated climate and comparisons
with paleoenvironmental data. Quaternary Science Reviews 17,
549–585.
Beaudoin, A.B., Oetelaar, G.A., 2003. The changing ecophysical
landscape of southern Alberta during the Late Pleistocene and
early Holocene. Plains Anthropologist 48, 187–207.
Birks, H.J.B., 1980. The present flora and vegetation of the moraines
of the Klutan Glacier, Yukon Territory, Canada: a study in plant
succession. Quaternary Research 4, 60–86.
Birks, H.H., 2001. Plant macrofossils. In: Smol, J.P., Birks, H.J.B.,
Last, W.M. (Eds.), Tracking Environmental Change Using Lake
Sediments: Volume 3, Terrestrial, Algal, and Siliceous Indicators.
Kluwer Academic Press, Dordrecht, The Netherlands, pp. 49–74
chapter 4.
Birks, H.J.B., 1976. Late-Wisconsinan vegetational history at Wolf
Creek, central Minnesota. Ecological Monographs 46, 395–429.
Bryson, R.A., Baerreis, D.A., Wendland, W.M., 1970. The character
of late-glacial and post-glacial climatic changes. In: Dort, Jr., W.,
Jones, Jr., J.K. (Eds.), Pleistocene and Recent Environments of the
central Great Plains. Department of Geology, University of Kansas
Special Publication 3, pp. 53–74.
Clark, J.S., Grimm, E.C., Donovan, J.J., Fritz, S.C., Engstrom, D.R.,
Almendinger, J.E., 2002. Drought cycles and landscape responses
to past aridity on prairies of the northern Great Plains, USA.
Ecology 83, 595–601.

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281280
Clayton, L., Moran, S.R., 1982. Chronology of late-Wisconsinan
glaciation in middle North America. Quaternary Science Reviews
1, 55–82.
Cowling, S.A., Sykes, M.T., 1999. Physiological significance of low
atmospheric CO2 for plant–climate interactions. Quaternary
Research 52, 237–242.
Davis, M.B., 1981. Quaternary history and the stability of deciduous
forests. In: West, D.C., Shugart, H.H., Botkin, D.B. (Eds.), Forest
Succession. Springer, New York, USA, pp. 132–177.
Davis, M.B., 1986. Climatic instability, time lags, and community
disequilibrium. In: Diamond, J., Case, T. (Eds.), Community
Ecology. Harper and Row, New York, USA, pp. 269–284.
Davis, M., Douglas, C., Calcote, R., Cole, K.L., Winkler, M.G.,
Flakne, R., 2000. Holocene climate in the western Great Lakes
national parks and lakeshores: implications for future climate
change. Conservation Biology 14, 968–983.
Dean, W.E., Schwalb, A., 2000. Holocene environmental and climatic
change in the northern Great Plains as recorded in the geochem-
istry of sediments in Pickerel Lake, South Dakota. Quaternary
International 67, 5–20.
Delcourt, H.R., Delcourt, P.A., 1991. Quaternary Ecology: A
Paleoecological Perspective. Chapman & Hall, London, UK.
DeVries, B., 1963. An Investigation of a Late-Glacial Deposit from the
Missouri Coteau in Saskatchewan. M.S. Thesis, Department of
Biology, University of Manitoba, Winnipeg, Canada.
Dew, J., 1959. Post-glacial forest at Kayville uncovered. The Blue Jay
(Saskatchewan Natural History Newsletter) 17, 20–21.
Environment Canada, 1993. Climatic Normals, 1961–1990: Prairie
Provinces. Supply and Services Canada, Ottawa, Ont, Canada.
Faegri, K., Iverson, J., 1975. Textbook of Pollen Analysis. Hafner
Press, New York, USA.
Florin, M.B., Wright Jr., H.E., 1969. Diatom evidence for the
persistence of stagnant glacial ice in Minnesota. Geological Society
of America Bulletin 80, 695–704.
Fredlund, G.G., Tieszen, L.L., 1997. Phytolith and carbon isotope
evidence for Late Quaternary vegetation and climate change in the
southern Black Hills, South Dakota. Quaternary Research 47,
206–217.
Great Plains Flora Association, 1986. Flora of the Great Plains.
University of Kansas Press, Lawrence, Kansas, USA.
Grimm, E.C., 2001. Trends and paleoecological problems in the
vegetation and climate history of the northern Great Plains, USA.
Proceedings of the Royal Irish Academy 101B (1–2), 47–64.
Grimm, E.C., Jacobson Jr., G.L., 2004. Late-Quaternary vegetation
history of the eastern United States. In: Gillespie, A.R., Porter,
S.C., Atwater, B.F. (Eds.), The Quaternary Period in the United
States. Elsevier, Amsterdam, The Netherlands, pp. 381–402.
Harington, C.R., Ashworth, A.C., 1986. A mammoth (Mammuthus
primigenius) tooth from late Wisconsin deposits near Embden,
North Dakota, and comments on the distribution of woolly
mammoths south of the Wisconsin ice sheets. Canadian Journal of
Earth Sciences 23, 909–918.
Holliday, V.T., Knox, J.C., Running IV, G.L., Mandel, R.D., Ferring,
C.R., 2002. The Central Lowlands and Great Plains. In: Orme,
A.R. (Ed.), The Physical Geography of North America. Oxford
University Press, New York, pp. 335–362.
Hopkins, D.G., Running, G.L., 2000. Soils, dunes, and prairie
vegetation: Lessons from the sandhills of North Dakota. Prairie
Forum 25, 45–64.
Hostetler, S.W., Bartlein, P.J., Clark, P.U., Small, E.E.,
Solomon, A.M., 2000. Simulated influences of Lake Agassiz on
the climate of central North America 11,000 years ago. Nature 405,
334–337.
Huntley, B., Birks, H.J.B., 1983. An Atlas of Past and Present Pollen
Maps for Europe, 0–13000 B.P. Cambridge University Press,
Cambridge, UK.
Kantrud, H.A., 1989. Vegetation of wetlands of the prairie pothole
region. In: van der Valk, A. (Ed.), Northern Prairie Wetlands. Iowa
State Press, Iowa, USA, pp. 132–187.
Kojima, S., 1994. Effects of global climatic warming on the boreal
forest. Journal of Plant Research 107, 91–97.
Krist Jr., K., Schaetzl, R.J., 2001. Paleowind (11,000 BP) directions
derived from lake spits in northern Michigan. Geomorphology 38,
1–18.
Kupsch, W.O., 1960. Radiocarbon-dated organic sediment near
Herbert, Saskatchewan. American Journal of Science 258, 282–292.
Kutzbach, J.E., Webb III, T., 1991. Late Quaternary climatic and
vegetational change in eastern North America: concepts, models,
and data. In: Shane, L.C.K., Cushing, E.J. (Eds.), Quater-
nary Landscapes. University of Minnesota Press, Minneapolis,
Minnesota, USA, pp. 175–217.
Jacobson Jr., G.L., Webb III, T., Grimm, E.C., 1987. Patterns and
rates of vegetation change during the deglaciation of eastern North
America. In: Ruddiman, W.F., Wright, Jr., W.E. (Eds.), North
American and Adjacent Oceans during the Last Deglaciation, The
Geology of North America, v. K-3. Geological Society of America,
Boulder, Colorado, USA.
Laird, K.R., Fritz, S.C., Grimm, E.C., Mueller, P.G., 1996. Century-
scale paleoclimatic reconstruction from Moon Lake, North
Dakota, a closed-basin lake in the northern Great Plains.
Limnology and Oceanography 41, 890–902.
Laird, K.R., Fritz, S.C., Cumming, B.F., Grimm, E.C., 1998. Early
Holocene limnological and climatic variability in the Northern
Great Plains. The Holocene 8, 275–285.
Last, W.M., Vance, R.E., Wilson, S., Smol, J.P., 1998. A multi-proxy
limnologic record of rapid early Holocene hydrologic change on
the northern Great Plains, southwestern Saskatchewan, Canada.
The Holocene 8, 503–520.
Looman, J., Best, K.F., 1987. Budd’s Flora of the Canadian Prairie
Provinces. Minister of Supply and Services, Agriculture Canada
Research Branch Publication 1662, Ottawa, Ont, Canada.
Lowell, T.V., Teller, J.T., 1994. Radiocarbon vs. calendar ages of
major late-glacial hydrological events in North America. Quatern-
ary Science Reviews 13, 801–803.
MacDonald, G.M., 2002. The Boreal Forest. In: Orme, A.R. (Ed.),
The Physical Geography of North America. Oxford University
Press, New York, USA, pp. 270–290.
MacDonald, G.M., Case, R.A., 2000. Biological evidence of multiple
temporal and spatial scales of hydrological variation in the western
interior of Canada. Quaternary International 67, 133–142.
MacDonald, G.M., Beukens, R.P., Kieser, W.E., 1991. Radiocarbon
dating of limnic sediments: a comparative analysis and discussion.
Ecology 72, 1150–1155.
Maher Jr., L.J., Miller, N.G., Baker, R.G., Curry, B.B., Mickelson,
D.M., 1998. Paleobiology of the sand beneath the Valders
diamicton at Valders, Wisconsin. Quaternary Research 49,
208–221.
Martin, J.E., Klukas, R.W., 1989. New locality for Late Pleistocene
fossils from Edmunds County, South Dakota. Proceedings of
South Dakota Academy of Science 68, 57–58.
McCallum, K.J., Wittenberg, J., 1962. University of Saskatchewan
radiocarbon dates III. Radiocarbon 4, 71–80.
McLeod, T.K., MacDonald, G.M., 1997. Postglacial range expansion
and population growth of Picea mariana, Picea glauca and Pinus
banksiana in the western interior of Canada. Journal of Biogeo-
graphy 24, 865–881.
Miller, S.L., Torres, P., McClean, T.M., 1994. Persistence of
basidiospores and sclerotia of ectomcorrhizal fungi and Morchella
in soil. Mycologia 86, 89–95.
Moir, D.R., 1957. An occurrence of buried coniferous wood in the
Altamont Moraine in North Dakota. Proceedings of North
Dakota Academy of Science 11, 31–36.

ARTICLE IN PRESSC.H. Yansa / Quaternary Science Reviews 25 (2006) 263–281 281
Owenby, J.R., Ezell, D.S., 1992. Monthly Station Normals of
Temperature, Precipitation, and Heating and Cooling Degree
Days 1961–1990: North Dakota. National Oceanic and Atmo-
spheric Administration, Climatography of the United States
No. 81.
Radle, N., Keister, C.M., Battarbee, R.W., 1989. Diatom, pollen, and
geochemical evidence for the palaeosalinity of Medicine Lake, S.
Dakota, during the Late Wisconsin and early Holocene. Journal of
Paleolimnology 2, 159–172.
Ritchie, J.C., 1969. Absolute pollen frequencies and carbon-14 age of a
section of Holocene lake sediment from the Riding Mountain area
of Manitoba. Canadian Journal of Botany 47, 1345–1349.
Ritchie, J.C., 1987. Postglacial Vegetation of Canada. Cambridge
University Press Cambridge, Cambridge, UK.
Ritchie, J.C., DeVries, B., 1964. Contributions to the Holocene
paleoecology of West Central Canada. II. A late-glacial deposit
from the Missouri Coteau. Canadian Journal of Botany 42,
677–692.
Ritchie, J.C., Harrison, S.P., 1993. Vegetation, lake levels, and climate
in western Canada during the Holocene. In: Wright, Jr., H.E.,
Kutzbach, J.E., Webb, III, T., Ruddiman, R.F., Street-Perrott,
F.A., Bartlein, P.J. (Eds.), Global Climates Since the Last Glacial
Maximum. University of Minnesota Press, Minneapolis, USA,
pp. 401–414.
Ritchie, J.C., MacDonald, G.M., 1986. The patterns of post-
glacial spread of white spruce. Journal of Biogeography 13,
527–540.
Shay, T.C., 1967. Vegetation history of the southern Lake Agassiz
basin during the past 12,000 years. In: Mayer-Oakes, W.J. (Ed.),
Life, Land and Water. University of Manitoba Press, Winnipeg,
Manitoba, Canada, pp. 231–252.
Shay, J.M., 1999. Annotated vascular plant species list for the Delta
Marsh, Manitoba and surrounding area. University of Manitoba
Field State (Delta Marsh) Occasional Publication No. 2, Winnipeg,
Manitoba, Canada, pp. 1–52.
Shuman, B., Webb III, T., Bartlein, P., Williams, J.W., 2002. The
anatomy of a climatic oscillation: vegetation change in eastern
North America during the Younger Dryas chronozone. Quatern-
ary Science Reviews 21, 1777–1791.
Smith, A.J., Donovan, J.J., Ito, E., Engstrom, D.R., 1997. Ground-
water processes controlling a prairie lake’s response to middle
Holocene drought. Geology 25, 391–394.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen,
K.A., Kromer, B., McCormac, G., van der Plicht, J., Spurk, M.,
1998. INTCAL98 radiocarbon age calibration, 24,000-0 cal BP.
Radiocarbon 40, 1041–1083.
Szeicz, J.M., MacDonald, G.M., 1994. Age-dependent tree-ring
growth responses of subarctic white spruce to climate. Canadian
Journal of Forestry 24, 120–132.
Teller, J.T., 2004. Controls, history, outbursts, and impact of large
late-Quaternary proglacial lakes in North America. In: Gillespie,
A.R., Porter, S.C., Atwater, B.F. (Eds.), The Quaternary Period in
the United States. Elsevier, Amsterdam, The Netherlands,
pp. 45–61.
Tuthill, S.L., Clayton, L., Laird, W.M., 1964. A comparison of a fossil
Pleistocene molluscan fauna from North Dakota with a recent
molluscan fauna from Minnesota. American Midland Naturalist
71, 344–362.
Van Zant, K., 1979. Late glacial and postglacial pollen and plant
macrofossils from Lake West Okoboji, northwesterm Iowa.
Quaternary Research 12, 358–380.
Watts, W.A., Bright, R.C., 1968. Pollen, seed, and mollusk analysis of
a sediment core from Pickerel Lake, northeastern South Dakota.
Geological Society of America Bulletin 79, 855–876.
Watts, W.A., Wright Jr., H.E., 1966. Late-Wisconsin pollen and seed
analysis from the Nebraska Sandhills. Ecology 47, 202–210.
Webb III, T., 1987. The appearance and disappearance of major
vegetation assemblages: Long-term vegetational dynamics in east-
ern North America. Vegetatio 69, 177–187.
Webb III, T., Cushing, E.J., Wright Jr., H.E., 1983. Holocene changes
in the vegetation of the Midwest. In: Wright, Jr., H.E. (Ed.),
Late-Quaternary Environments of the United States. Volume 2:
The Holocene. University of Minnesota Press, Minneapolis,
pp. 142–165.
Webb III, T., Bartlein, P.J., Kutzbach, J.E., 1987. Climatic change in
eastern North America during the past 18,000 years; Comparisons
of pollen data with model results. In: Ruddiman, W.F., Wright, Jr.,
W.E. (Eds.), North American and Adjacent Oceans during the Last
Deglaciation, The Geology of North America, v. K-3. Geological
Society of America, Boulder, Colorado, USA, pp. 447–462.
Webb III, T., Bartlein, P.J., Harrison, S.P., Anderson, K.H., 1993.
Vegetation, lake levels, and climate in eastern North America for
the past 18,000 years. In: Wright, Jr., H.E., Kutzbach, J.E., Webb,
III, T., Ruddiman, R.F., Street-Perrott, F.A., Bartlein, P.J. (Eds.),
Global Climates Since the Last Glacial Maximum. University of
Minnesota Press, Minneapolis, Minnesota, USA, pp. 415–467.
Webb III, T., Shuman, B., Williams, J.W., 2004. Climatically forced
vegetation dynamics in eastern North America during the late
Quaternary Period. In: Gillespie, A.R., Porter, S.C., Atwater, B.F.
(Eds.), The Quaternary Period in the United States. Elsevier,
Amsterdam, The Netherlands, pp. 459–478.
Whitlock, C.W., Bartlein, P.J., Watts, W.A., 1993. Vegetation history
of Elk Lake. In: Bradbury, J.P., Dean, W.E. (Eds.), Elk Lake,
Minnesota: Evidence for Rapid Climate Change in the North-
Central United States. Geological Society of America Special Paper
276, pp. 251–274.
Williams, J.W., Shuman, B.N., Webb III, T., 2001. Dissimilarity
analyses of Late Pleistocene and Holocene vegetation and climate
in eastern North America. Ecology 82, 3346–3362.
Wright Jr., H.E., 1968. The roles of pine and spruce in the forest
history of Minnesota and adjacent areas. Ecology 49, 937–955.
Wright Jr., H.E., 1976. Ice retreat and revegetation in the western
Great Lakes area. In: Mahaney, W.C. (Ed.), Quaternary Strati-
graphy of North America. Dowden, Hutchingson and Ross,
Stroudsburg, Pennsylvania, pp. 119–132.
Wright Jr., H.E., 1987. Synthesis; The land south of the ice sheets. In:
Ruddiman, W.F., Wright, Jr., W.E. (Eds.), North American and
Adjacent Oceans during the Last Deglaciation, The Geology of
North America, v. K-3. Geological Society of America, Boulder,
Colorado, USA, pp. 479–488.
Wright Jr., H.E., 1992. Patterns of Holocene climatic change in the
Midwestern United States. Quaternary Research 38, 129–134.
Yansa, C.H., 1998. Holocene paleovegetation and paleohydrology of a
prairie pothole in southern Saskatchewan, Canada. Journal of
Paleolimnology 19, 429–441.
Yansa C.H., 2002. Spatial and Temporal Vegetation Changes
in the Northeastern Great Plains During the Late Pleistocene and
Early Holocene. Ph.D. Dissertation, University of Wisconsin-
Madison, USA.
Yansa, C.H., Ashworth, A.C., 2005. Late Pleistocene palaeoenviron-
ment of the southern Lake Agassiz basin, USA. Journal of
Quaternary Science 20, 255–267.
Yansa, C.H., Basinger, J.F., 1999. A postglacial plant macrofossil
record of vegetation and climate change in southern Saskatchewan.
In: Lemmen, D.S., Vance, R.E. (Eds.), Holocene Climate and
Environmental Change in the Palliser Triangle, Southern Canadian
Prairies, vol. 534. Geological Survey of Canada Bulletin,
pp. 139–172.





























