Embed Size (px)
Citation preview

Hydrobiologia 269/270 : 427-436, 1993 .H. van Dam (ed.), Twelfth International Diatom Symposium .© 1993 Kluwer Academic Publishers . Printed in Belgium .
Introduction
Since European settlement in 1829, there has beenconsiderable decline of wetlands in Western Aus-tralia which are an important refuge for birds,especially during the summer months . WesternAustralia is the leading producer of titaniferousminerals like ilmenite, rutile, zircon and monazitein the world . Sand deposits have been mined inCapel, 200 km south of Perth (33 'S, 116 'E), inWestern Australia since 1956 (Fig . 1). The min-ing pits left after the extraction of mineral sandintercepting the water table were converted intoartificial wetlands with permanent waterbodies,between 1975 and 1979 . These wetlands occupy
427
The use of diatoms in monitoring the development of created wetlands ata sandmining site in Western Australia
Jacob JohnSchool of Environmental Biology, Curtin University of Technology, G.P.O. Box U1987, Perth, WA, 6001,Australia
Key words : acidophilous, alkaliphilous, bioindicators, circumneutral, diatoms, wetlands restoration,ecological engineering
Abstract
Former sand-mining pits at Capel, 200 km south of Perth in Western Australia, have been rehabilitatedinto artificial wetlands since 1975-1979 . A chain of fresh water lakes was created as a potential waterbirdrefuge and an area for passive recreation . Initially, the lakes had low pH, high ammonium, iron andmanganese levels and low phosphorus concentration . The lakes were characterised by low diversity ofdiatoms dominated by acidophilous species . Following an increase in pH in the effluent water dischargedinto the lake from the mining process plant and landscaping of the lakes since 1988, the diversity ofdiatoms gradually increased . The system is now dominated by periphytic diatom communities, prefer-ring high conductivity . There has been a marked transition in the diatom community from acidophilousto alkaliphilous species . Planktonic diatom blooms replaced dinoflagellate blooms . Concomitantly, therehas been a dramatic increase in the diversity of invertebrates and waterbirds in these lakes . The valueof diatoms in assessing the progressive development of created wetlands as self-sustaining ecosystemsat sand mines in Australia is discussed .
an area of 44 hectares, now known as the RGC(Renison Goldfields Consolidated) WetlandsCentre and comprises a chain of lakes mostlyinterconnected (Fig . 1). The chain of lakes re-ceives treated effluent water from the miningprocessing plant at one end ; there is an outletdischarge point at the other end .
RGC Mineral Sands Ltd in 1986 establishedthe wetlands centre with the objectives of devel-oping a self-sustaining ecosystem for the conser-vation of waterbirds and facilitating research intothe development of artificial wetlands . The term`created wetlands' is defined as artificial wetlandscreated from non-wetland sites to produce or re-place natural habitat according to the Environ-

428
mental Protection Agency (EPA) of U.S.A. Incontrast, `constructed wetlands' are constructedfor the sole purpose of waste water or storm watertreatment (Hammer, 1992). Following the abovedefinitions, the RGC wetlands are considered as`created wetlands' in this paper . As part of a mul-tidisciplinary research program, diatoms were in-vestigated in the lake system from 1987 to 1992 .
Surveys of waterbird usage between 1984 and86, showed that the system was not productiveenough to attract large numbers of birds, as in aneighbouring wetland (e .g. Higgins Lake, Fig. 1) .The pH of the lakes was mostly between 2 and4 and ammonium levels were very high and phos-phorus very low. The concentration of iron, man-ganese and sulphates was too high, favouring
metres
Fig . 1 . Location map of the study site . A channel carrying theeffluent water from the mine processing plant enters the sys-tem at Lake 2 and exits over a weir south-west of Lake 8 .Lakes 1, 6 and 8A are not directly connected to the chain oflakes .
binding of phosphates . The effluent treatment fa-cilities of the processing plant were upgraded in1988, resulting in the buffering of the effluent waterto a high pH level of above 8, and reducing con-siderably the iron and manganese content .
Data on diatoms as indicators of water qualityreflecting pH, salinity and organic pollution inEurope, America, South Africa and Japan havebeen available for a long time (e.g. Patrick, 1986 ;Schoeman, 1973 ; Round, 1986 ; 1990 ; Cox, 1991) .However, very little published information isavailable on diatoms as indicator species of de-veloping artificial wetlands . The aims of this paperare to examine the use of diatoms as bioindicatorsof the developing artificial wetland at the sand-mining sites at Capel, Western Australia andidentify indicator species of changing waterquality .
Study sites
The lakes sampled are indicated in Fig . 1 . Theeffluent water from the mining processing plant isdischarged into Lake 2 which is connected up toLake 8 by drainage channels. There is an outletchannel at the end of the chain. Lake 1, Lakes 6and 8A, are separated from the chain of lakes, butwere formed from extraction pits as other lakes .Lake 1 was established in 1975 and is the oldest .Lake 5 was constructed in 1976 and Pond 8 in1979 and the rest during the period in between .The lakes are well oxygenated with high light pen-etration . Only Lake 1 has a high percentage offringing vegetation, with considerably reducedopen water . Higgin's Lake, a natural lake (Fig . 1)proximal to the chain of the mine-site lakes is awell-known bird refuge . This lake was sampled toobtain data on the distribution of natural diatomflora of the local area for comparison .
The average annual rainfall of the region is 830mm and the climate is mediterranean character-ised by dry summers and wet winters with a tem-perature range of 20-36 ° C in the summer and8-15 °C in the winter . The lakes range in depthfrom 1 m to more than 6 m. The bottom of thelakes is mostly soft and boggy with fine sedi-

ments. The water level in the lakes reflects theseasonal fluctuation of the groundwater table .Since 1988, the shorelines were reshaped withshallow waterslopes, islands created and periph-eral vegetation planted .
Materials and methods
Benthic samples from the top 1-cm sediment fromthe shoreline to depths ranging from 10 cm to 150cm and epiphytic diatoms from the emergentmacrophytes were collected . Phytoplankton wassampled by use of a 10 pm-plankton net. Sampleswere collected from randomly selected sites inautumn 1987 and subsequently 3 to 4 times a yeartill 1992 autumn . pH and conductivity were mea-sured and the information on the water chemis-try was provided by the RGC Company .
Fresh benthic samples were examined to de-termine proportion of live and dead cells . Then aconstant volume of the samples were acid-cleanedand permanent slides made as described by John(1983). Light micrographs of species were takenby a Vanox photomicroscope and the diatomswere identified using Hustedt (1959), John (1983),Schoeman (1973), Patrick & Reimer (1966 &1975), and Krammer & Lange-Bertalot (1986) .Ecological pH preferences of identified specieswere compiled from an array of sources, in ad-dition to those mentioned above (Lowe, 1974 ;Gasse, 1986 ; Cassie, 1989 ; Foged, 1978) and fromunpublished work by the author . Permanent slidesof the benthic samples were quantitatively analy-sed .
Relative abundance of individual species ofdiatoms was determined by counting a minimumof 300 valves . The percentage of acidobiontic,acidophilous, alkaliphilous, alkalibiontic and cir-cumneutral diatoms in each lake was determined .Diversity of diatom communities of selected lakeswas analysed by Shannon-Weaver diversity index(H') (Abel 1988) .
S
H' _ - Y PP loglo (P),i = 1
where P; denotes proportion of the taxon i and sthe total number of taxa in the sample .
Based upon the analysis of the distribution ofspecies, they were identified as possible indica-tors of pH and conductivity, the two obvious keyenvironmental factors affecting their abundance .
Results
A list of the most common species with their al-lotted pH spectrum and their dominance and oc-currence from 1987 to 1992 is presented inTable 1 . Species richness in all the sand minewetlands increased from 19 in 1987 to 34 in1988-1990 and to 61 in 1992 . The species, char-acteristic of the lakes in 1987 under low pH areshown in Fig. 2A to J . Some of the species cur-rently dominant in the systems are shown in Fig . 2K to Z. The species identified as Cymbella minutashows high variation and is probably close to thevariety silesiaca (Bleish ex Rabenh .) Reimer. Thechange in species composition from 1987 to 1992according to their pH preference is presented inFig. 3 . The species richness in 1987 in Lakes 2 to8 was only 15, when the pH ranged from 2 to 4 .The least number of species (7) was in Lake 2 towhich the effluent water from the mining process-ing plant was directly discharged. Lake 8 whichis at the end of the chain had a species richnessof 14. The survey in 1987 also showed that Lake1, the oldest of all the artificial lakes (Fig. 1) whichwas isolated from the chain of lakes had a maxi-mum species richness of 19, with a pH of 5 .5-6.5 .The water chemistry of the chain of lakes is pre-sented in Table 2 .
The diversity index H' and species richness forselected Lakes (1, 2, 6, 8 and 8A) are given inTable 3 . The H' value of the benthic diatom com-munities in Lake 2 in 1987 when the pH was 2-4was very low i .e. 0.28 . Following an increase inpH of the effluent water, the diversity index valuerose to 0.52 in 1989 finally reaching the currentvalue of 0 .62. The same trend is seen in Lake 8 .
Although the three isolated lakes are formed asthe result of extraction of sands and located closeto the chain of lakes (Fig . 1) i .e. Lake 1, 8A and
429

430
Table] . Relative abundance of selected common species . *=(0.5-2.9%), **= (3-24%), ***=(>25%),
Species
Sand-mining wetlands
Higgin'sLake
1987
1988-90
1991-92
1987-92
AcidobionticBrachysira serians (Breb.) Round & MannFrustulia megaliesmontana Cholnoky
AcidophilousPinnularia divergentissima (Grun.) Cleve .Pinnularia gibba Ehrenb .Brachysira vitrea (Grun.) RossEunotia pectinalis (O .F . Muller) Rabenh .Navicula aff. festiva KasskeEunotia curvata (Katz .) LagerstedtNitzschia paleaeformis Hustedt
CircumneutralNavicula aff. cari Ehrenb .Pinnularia biceps Greg .Pinnularia braunii (Grun.) Cleve .Pinnularia subcapitata Greg .Pinnularia viridis (Nitzsch .) Ehrenb .Synedra rumpens Katz .
AlkaliphilousAchnanthes minutissima Katz .Anomoeoneis sphaerophora (Katz .) PfitzerCaloneis bacillum (Grun.) MereschkowskyChaetoceros muelleri LemmermannCyclotella meneghiniana Katz .Cymbella microcephala Grun .Cvmbella minuta Hilse ex Rabenh .Mastogloia elliptica (Ag .) Cleve .Mastogloia smithii ThwaitesNavicula cincta (Ehrenb.) Katz .Navicula cryptocephala Katz .Navicula cuspidata Katz.Navicula halophila (Grun.) Cleve .Navicula pupula Katz .Navicula rhynchocephala Katz .Navicula halophila (Grun.) Cleve.Nitzschia obtusa W. SmithStauroneis pachycephala Cleve .Thalassiosira weissflogii Hustedt .Cyclotella stelligera Cleve & Grun .Navicula salinarum Grun .
AlkalibionticCymbella pusilla Grun .Rhopalodia gibba (Ehrenb.) O . MullerRhopalodia novae-zelandiae Hustedt
• * * *• * * ****
*
*
*• * *•
* *•
* ****
**
*
•
* * *•
* * *•
**
****
*
*
***
*
*
*
***
******
***
*******
****
******
***
*
****
*
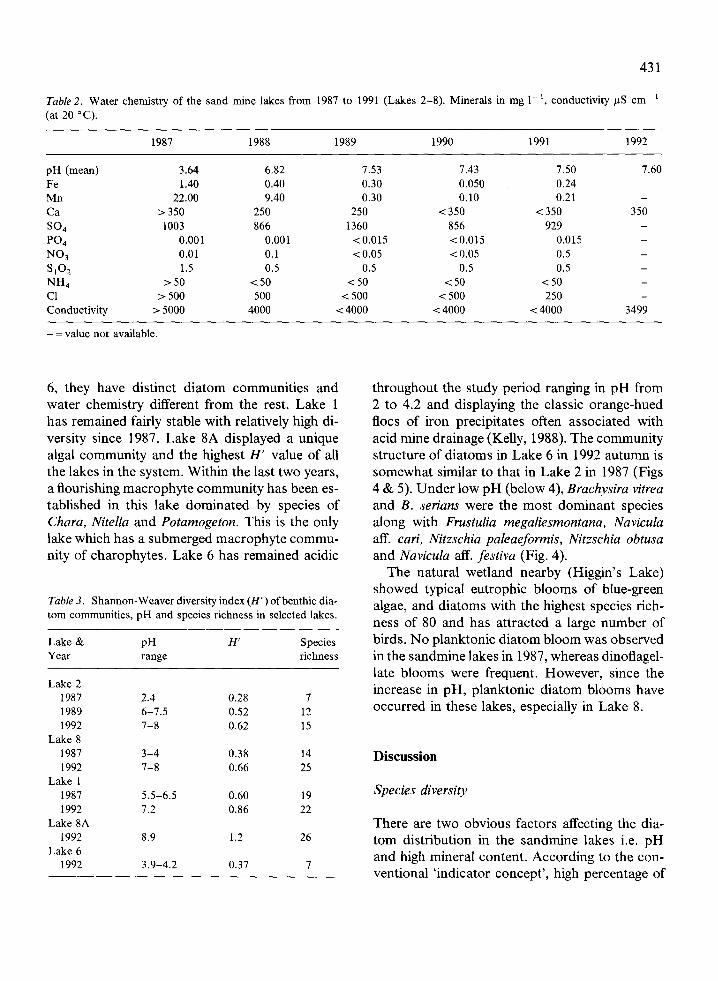
Table 2 . Water chemistry of the sand mine lakes from 1987 to 1991 (Lakes 2-8) . Minerals in mg 1 - ',conductivity µS cm -1(at 20 ° C).
- = value not available .
6, they have distinct diatom communities andwater chemistry different from the rest . Lake 1has remained fairly stable with relatively high di-versity since 1987 . Lake 8A displayed a uniquealgal community and the highest H' value of allthe lakes in the system. Within the last two years,a flourishing macrophyte community has been es-tablished in this lake dominated by species ofChara, Nitella and Potamogeton. This is the onlylake which has a submerged macrophyte commu-nity of charophytes . Lake 6 has remained acidic
Table 3 . Shannon-Weaver diversity index (H') of benthic dia-tom communities, pH and species richness in selected lakes .
43 1
throughout the study period ranging in pH from2 to 4.2 and displaying the classic orange-huedflocs of iron precipitates often associated withacid mine drainage (Kelly, 1988) . The communitystructure of diatoms in Lake 6 in 1992 autumn issomewhat similar to that in Lake 2 in 1987 (Figs4 & 5). Under low pH (below 4), Brachysira vitreaand B . serians were the most dominant speciesalong with Frustulia megaliesmontana, Naviculaaff. cari, Nitzschia paleaeformis, Nitzschia obtusaand Navicula aff. festiva (Fig. 4) .
The natural wetland nearby (Higgin's Lake)showed typical eutrophic blooms of blue-greenalgae, and diatoms with the highest species rich-ness of 80 and has attracted a large number ofbirds . No planktonic diatom bloom was observedin the sandmine lakes in 1987, whereas dinoflagel-late blooms were frequent . However, since theincrease in pH, planktonic diatom blooms haveoccurred in these lakes, especially in Lake 8 .
Discussion
Species diversity
There are two obvious factors affecting the dia-tom distribution in the sandmine lakes i .e . pHand high mineral content. According to the con-ventional `indicator concept', high percentage of
1987 1988 1989 1990 1991 1992
pH (mean) 3.64 6 .82 7 .53 7 .43 7 .50 7 .60Fe 1 .40 0 .40 0.30 0 .050 0 .24Mn 22.00 9 .40 0 .30 0.10 0 .21 -Ca > 350 250 250 <350 < 350 350SO 4 1003 866 1360 856 929P0 4 0.001 0.001 <0.015 <0.015 0.015NO3 0.01 0 .1 < 0 .05 < 0 .05 0 .5S,O2 1 .5 0 .5 0 .5 0 .5 0 .5NH4 >50 <50 <50 <50 <50Cl > 500 500 <500 < 500 250 -Conductivity > 5000 4000 < 4000 <4000 <4000 3499
Lake &Year
pHrange
H' Speciesrichness
Lake 21987 2 .4 0.28 71989 6-7 .5 0 .52 121992 7-8 0 .62 15
Lake 81987 3-4 0 .38 141992 7-8 0 .66 25
Lake 11987 5 .5-6 .5 0 .60 191992 7 .2 0 .86 22
Lake 8A1992 8 .9 1 .2 26
Lake 61992 3 .9-4 .2 0 .37 7

432

60-
so40
rnmCRd 20CL
10
0
30
11987
11988-90
1991-92
Year
u alp &albO cir•
acp & acb
Fig. 3 . Mean percentage of species according to the pH spec-trum 1987-1992 alp & alb = alkaliphilous and alkalibiontic,cir = circumneutral, acp & acb = acidophilous and acidobion-tic .
species in an environment is correlated to themost obvious environmental factors (Eloranta,1990). Any causality between species and theenvironmental factors has to be experimentallyverified (Cox, 1991). However, low diversity isoften associated with stressful conditions (Sulli-van, 1986) . Diversity index (H') therefore mightbe considered as a reasonable measure of thedevelopment of created wetland system as shownin this study .The H' value of the diatom community in Lake
2 for example was 0 .28 in 1987, when the pH was2-4, and the mineral content too high . With theincrease in pH and decrease in mineral content,H' has risen to 0 .62 in 1992 . The clear correla-tion between pH and species diversity is furtherexemplified by the diatom community in Lakes 1,6 and 8A all of which have been physically sepa-rated from the chain of sandmine lakes . Lake 1which is the oldest of all the `created lakes' witha pH of above 7 and a high coverage of macro-phytes Typha and Isolepis has a species richnessof 22 diatoms and H' value of 0 .86. Lake 8A with
00Cc0UNa
mNm
u
0.
0Z
Z
Z
in
z
43 3
Fig. 4 . Percentage species composition of benthic diatomcommunity in Lake 2, autumn, 1987 . By = Brachysira vitrea,Bs = Brachysira serians, Ne = Navicula aff. cari, Np = Nitzschiapaleaeformis, No = Nitzschia obtusa, Sr = Synedra rumpens,Nf=Navicula aff. festiva .
a pH up to 8 .9 and a wide variety of submergedmacrophytes had the lowest conductivity, but thehighest species richness and the highest H' value,whereas Lake 6, the only acidic lake remaining inthe system with a mean pH of 4 and the highestconductivity has the least species diversity remi-niscent of the condition of the system before 1988 .It is not surprising that the nearby natural lake(Higgin's Lake) has the highest species richnessand is highly productive attracting vast numbersof birds . It has been an alkaline eutrophic systemsimilar to most of the natural wetlands in West-ern Australia .
Community structure
A species list of the community with relativeabundance provides a vast amount of informa-tion compared with any single number such asdiversity index (Abel, 1989; Round, 1991) . Eachspecies can be a vital piece of information espe-cially with proven attributes of ecological prefer-ence. In 1987, the sandmine lakes were domi-
Fig . 2 . Light Micrographs . Scale = 10 pm . A. Brachysira vitrea, B . Brachysira serians, C . Pinnularia viridis, D . Frustulia mega-liesmontana, E. Navicula aff. festiva, F. Synedra rumpens, G. Nitzschia paleaeformis, H . Nitzschia obtusa, I . Eunotia curvata, J . Naviculaaff. cari, K . Achnanthes minutissima, L . Cymbella minuta (note variations in size and shape), M . Cyclotella meneghiniana, N .Cyclotella stelligera, 0 . Navicula halophila, P . Pinnularia subcapitata, Q . Navicula cuspidata, R. Caloneis bacillum, S . Stauroneispachycephala, T . Rhopalodia gibba, U . Rhopalodia novae-zelandiae, V. Anomoeoneis sphaerophora, W . Cymbella pusilla, X . Mastogloiasmithii, Y. Diploneis ovalis, Z . Amphora veneta .

434
Fig . 5 . Percentage species composition of benthic diatomcommunities in Lake 6, autumn 1992 . Bs = Brachysira serians,By = Brachysira vitrea, Fm = Frustulia megaliesmontana,Nc = Navicula aff. cari, Nf = Navicula aff. festiva, Np =Nitzschia paleaeformis, No = Nitzschia obtusa .
nated by acidobiontic and acidophilous species(Fig. 3) . After an increase of pH, since 1988, therehas been a dramatic increase of alkaliphilous andalkalibiontic species in the system, except for Lake6 which continued to harbour the remnantacidiobiontic/acidophilous flora .
Lake 6 is still dominated by the Brachysiraspecies and Frustulia megaliesmontana . pH pref-erence of Brachysira vitrea is presented as acido-philous and circumneutral in lake acidificationstudies in Wales (Battarbee et al., 1988) whereasFoged (1978) considered it as alkalibiontic .Brachysira vitrea is common in thermal springsunder low pH in New Zealand (Cassie, 1989) andin hot springs in Africa (Compere, 1986) and inHawaii (Fungladda et al., 1983) . Brachysira vitreahad been observed as dominant in the acidic lakeLoch Dee in Scotland after it was limed (Round,1990). Apart from pH, mineral content, especiallycalcium, might be implicated in the dominance ofthe above species . The calcium content of thesandmine lakes in this study is several times higherthan that in the acidic lakes of Wales and Scot-land (Table 2) .
Nitzschia obtusa is commonly found in thebrackish waters of the upper reaches of Austra-lian estuaries where salinity may range from 2 to20 (John, 1983) . The occurrence of this species inlarge numbers in the acidic waters in the sand-mine lakes could be attributed to high conductiv-ity and high mineral content . This is substantiated
by the fact that this species is still commonlyfound in most of the lakes where B. vitrea andB. serians have virtually disappeared . Nitzschiaobtusa is probably an acid tolerant species whichprefers high conductivity .
Shift in dominance of species
Changes in water quality are best shown by a shiftin the dominance of species (Van Dam, 1982 ;Sullivan, 1986 ; Round, 1990). This study pro-vides ample examples of shift in dominance . Fol-lowing a change in pH and water chemistry, thesystem displayed dominance of Cymbella minutaand Achnanthes minutissima replacing Brachysiraspecies . Round (1990) identified both Cymbellaminuta and Achnanthes minutissima as indicatormarker species appearing after liming in an acidi-fied Welsh lake. These two species have beenidentified as key species of change in pH of waterby many other workers .
No planktonic diatom blooms were observedin the sandmine lakes in 1987, whereas dinoflagel-late blooms were frequent . Since 1989, planktonicblooms of Cyclotella meneghiniana, Thalassiosiraweissflogii and Chaetoceros muelleri almost in syn-chrony with those in the natural wetland nearbyhave been observed . Chaetoceros muelleri bloomsare common in the alkaline lakes of America(Johansen & Rushforth, 1985) . Absence of plank-tonic diatom blooms in acidic lakes has been welldocumented (Round, 1986) . Currently Lake 6,which is acidic, is the only lake experiencing di-noflagellate blooms. Shift from dinoflagellateblooms to planktonic diatom blooms, thereforecan be considered as an indication of change fromlow pH to high .
Appearance of alkalibiontic species such asMastogloia elliptica andM. smithii, Cymbella pu-silla, Rhopalodia novae-zelandiae and R . gibba, asdominant forms in Lake 8A marks a shift to ahigh pH regime (pH above 8) . However, this isconfined only to Lake 8A . The latter two specieswere not observed in any other lakes . They occurmostly as epiphytes on Chara and Nitella whichare confined only to Lake 8A . The virtual disap-

pearance of many Pinnularia species except P.subcapitata and P. viridis can be again interpretedas the mark of change in water chemistry . P. viri-dis occurs in most of the alkaline lakes in West-ern Australia (personal observation) .
Parallel with the change in chemistry, thephysical features of the lakes also have beenchanging due to reshaping of the steep shorelineto slopes, creation of islands and general land-scaping and planting of macrophytes and periph-eral vegetation . The increasing dominance ofbenthos (periphyton and epilithon) in the systemis an indication of these changes . The system ispoor in soluble phosphorus and nitrogen(Table 2). The difference in species richness andbiomass observed between the sandmine systemand the nearby natural wetland highlights its poornutrient status .
Indicator species
Low pH due to mine drainage affect the waterchemistry differently from that due to peat drain-age and acid rains (Kelly, 1988) . High conduc-tivity is characteristic of acid mine drainage . Theconductivity of the sandmine lake system at Capelis above 4000 µ S cm -1 compared with < 100 p Scm -1 in most of the acid rain affected systems ofEurope. Where both acidity and high conductiv-ity are present, it is difficult to determine whichone has the greatest effect, and which speciesreflect specifically which factor . However, whenthe distribution of diatoms at low pH in 1987 andat high pH from 1988 to 1992 is analysed, Brachy-sira vitrea, Brachysira serians, Frustulia megalies-montana, Navicula aff. cari, Navicula aff. festiva,and a few Pinnularia spp . can be considered asindicators of low pH . At the other end of thepH spectrum, are Rhopalodia novae-zelandiae, R .gibba, Cymbella pusilla, Mastogloia spp . andChaetoceros muelleri . Apart from these indicatorspecies of extreme pH, the following can be rec-ognised as markers of transition from acidic toalkaline conditions : Achnanthes minutissima,Cymbella minuta and Pinnularia subcapitata . Thevast majority of other diatoms in this study
435
are common in the shallow lakes of WesternAustralia with a pH range of 7-8 and high con-ductivity. Accurate identification of species is piv-otal in their use as indicators. The taxonomy ofspecies identified as Navicula aff. cari and N. aff.festiva and Cymbella minuta in this study need tobe further investigated .
Diatom distribution has been documented asevidence for neutralization of acid-mine lakes(Brugam & Lusk, 1986). Diatom communities inLakes 1 and 8A in this study provide further evi-dence for natural acid neutralization . Isolationfrom the continuous discharge of acid mine efflu-ence appears to be a prerequisite for acid neu-tralization . Apart from the time required forweathering of sulphide minerals, benthic micro-bial communities may initiate changes in theirmicroenvironment leading to neutralization . Thecontinuing low pH in Lake 6 need to be furtherinvestigated .
As the diversity of diatom communities in-creased with the increase in pH in the mine lakesat Capel, the macroinvertebrate communities andthe birdlife also increased. In 1987, 27 species ofbirds were reported from the RGC wetland areas .In 1992, this number has increased to 104 (F .Doyle, pers . comm.). In many respects, the dia-tom species appeared to be more sensitive towater quality than the macroinvertebrates, manyof whose identification, even to generic level, re-mains problematic . The latter were more associ-ated with macrophytes and fringing vegetationcover than open water (D . Gale & D. Edward,pers. comm.). The use of diatoms in biomonitor-ing the development of created wetlands has yetto gain the proper recognition it deserves, as morewetlands are being created as part of restorationprograms throughout the world (Hammer, 1992) .Though most of the diatoms are universal in theirindicator value, local indicator species need to beestablished before they can be used for assessingwater quality in a given geographical area . Thisstudy has been a pioneering attempt to identifysuch indicator species in Australia, where manyexamples of ecological engineering in mined landscan be found .

436
Acknowledgements
I wish to thank Mr Stuart Helleren, Mr JustinLong and Miss Kylie Ashenbrenner for assistancewith field work involved in this project . Financialsupport offered by the RGC Company andencouragement by Mr Denis Brooks, Environ-mental Manager for the above Company aregratefully acknowledged .
References
Abel, P.O ., 1988. Water Pollution Biology. Ellis HarwoodLtd., New York, 231 pp.
Battarbee, R.W ., N .J . Anderson, P .G. Appleby, R .J . Flower,S .C . Fritz, E .Y. Haworth, S . Higgitt, V.J . Jones, A . Kre-iser, M.A.R. Munro, J . Natkanski, F . Oldfield, S .T . Patrick,N.J . Richardson, B . Rippey & A .C. Stevenson, 1988 . Lakeacidification in the United Kingdom 1800-1986 . Evidencefrom analysis of lake sediments . ENSIS Publishing, Lon-don, 68 pp .
Brugam, R.B. & M. Lusk, 1986 . Diatom evidence for neu-tralization in acid surface mine lakes . In J . Smol, R.W .Battarbee, R .B . Davis & J . Merilainen (eds), Diatoms andlake acidity, Dr W . Junk Publishers, Dordrecht : 115-129 .
Cassie, V ., 1989 . A contribution to the study of New Zealanddiatoms . Bibliotheca Diatomologica 17 : 1-226 .
Compere, P ., 1986. Diatoms in two hot springs in Zambia(Central Africa) . In F .E. Round (ed .), Proceedings of the9th International Diatom Symposium, Biopress, Bristol andO. Koeltz, Koenigstein : 29-39 .
Cox, E.J ., 1991 . What is the basis for using diatoms as moni-tors of river quality? In B.A. Whitton, E. Rott & G .Friedrich (eds), Use of algae for monitoring rivers . E . Rott,Innsbruck : 33-40 .
Eloranta, P., 1990 . Ecology of some acidophilic and acido-biontic diatoms in Finnish forest lakes . In Simola, H . (ed.),Proceedings of the Tenth International Diatom Sympo-sium. Koeltz, Scientific Books, Koenigstein, 491-498 .
Foged, N ., 1978 . Diatoms in Eastern Australia . BibliothecaPhycologica 41 : 1-232 .
Fungladda, N ., I . Kaczmarska, & S.R. Rushforth, 1983 .A contribution to the freshwater diatom flora of the Ha-waiian Islands . Bibliotheca Diatomologica 2 : 1-103 .
Gasse, F ., 1986 . East African diatoms and water pH . In J .P .Smol, R.W. Battarbee, R .B . Davis, and J . Merilainen (eds) .Diatoms and lake acidity . Dr W. Junk Publishers, Dor-drecht : 149-168 .
Hammer, D.A., 1992 . Creating freshwater wetlands . LewisPublishers Inc ., U.S.A . : 298 pp .
Hustedt, F ., 1959 . Die Kieselalgen Deutschlands, Osterreichsand der Schweiz. In Dr . L. Rabenhorst's Kryptogamenflora von Deutschlands, Osterreichs and der Schweiz 7(2) .Reprint 1977, Otto Koeltz, Koenigstein : 440-574 .
Johansen, J.R. & S .R. Rushforth, 1985 . A contribution to thetaxonomy of Chaetoceros muelleri Lemmermann (Bacillari-ophyceae) and related taxa . Phycologia 24 : 437-447 .
John, J., 1983 . Diatom flora of the Swan River estuary, West-ern Australia . Bibliotheca Phycologica 64 : 1-360 .
Kelly, M ., 1988. Mining and the freshwater environment .Elsevier Applied Science, London, 231 pp .
Krammer, K. & H. Lange-Bertalot, 1986 . Bacillariophyceae .Siisswasserflora von Mitteleuropa 2(1) . Gustav-FischerVerlag. Stuttgart, 876 pp .
Lowe, R.L ., 1974 . Environmental requirements and pollutiontolerance of freshwater diatoms . U.S . EPA, Cincinnati,Ohio, 334 pp .
Patrick, R . & C.W. Reimer, 1966-1975 . The diatoms of theUnited States . Monogr . Acad . Nat . Sc . Phila ., No . 13, Part1, 1-2 .
Patrick, R ., 1986 . Diatoms as indicators of changes in waterquality. In M . Ricard (ed .), Proceedings of the 8th Inter-national Diatom Symposium . Koeltz Scientific Books,Koenigstein: 759-766 .
Round, F.E ., 1986. Non-planktonic diatoms and acid lakes .In M. Ricard (ed .), Proceedings of the 8th InternationalDiatom Symposium . Koeltz Scientific Books, Koening-stein : 753-755 .
Round, F.E., 1990 . The effect of liming on the benthic diatompopulations in three upland Welsh lakes . Diatom Res . 5 :129-140 .
Schoeman, F.R ., 1973 . A systematical and ecological study ofthe diatom flora of Lesotho with special reference to thewater quality . National Institute of Water Research, SouthAfrica, 354 pp .
Sullivan, M.J ., 1986 . Mathematical expression of diatom re-sults : Are these pollution indices valid and useful? In M .Ricard Proceedings of the 8th International Diatom Sym-posium, Koeltz Scientific Books, Koenigstein : 772-775 .
Van Dam, H., 1982. On the use of measures of structure anddiversity in applied diatom ecology . Nova Hedwigia, Beih .73 : 97-115 .





























