Embed Size (px)
Citation preview

J. therm. Biol. Vol. 15, No. 3/4, pp. 291-300, 1990 0306-4565/90 $3.00 + 0.00 Printed in Great Britain. All fights reserved Copyright © 1990 Pergamon Press pie
THERMAL PARAMETERS OF FOUR SPECIES OF GERBILLURUS
COLLEEN T. DOWNS and M. R. PERRXN Department of Zoology and Entomology, University of Natal, Box 375, Pietermaritzburg, R.S.A.
(Received 5 August 1989; accepted in revised form 2 December 1989)
Abstract--1. Thermal parameters of the four Gerbillurus species measured in the laboratory were examined in relation to their micro-environments in a xeric habitat.
2. Basal metabolic rates (BMR) were lower than predicted, while thermoneutral zones (TNZ) were narrow and exceeded burrow temperatures.
3. Body temperatures (Tb) were regulated over a range of ambient temperatures (Ta). Evaporative water loss was used as a short-term cooling mechanism to reduce hyperthermia above the TNZ.
4. Conductance was low below the TNZ to reduce heat loss. 5. Adaptation to low temperatures is important for gerbils when active at night. 6. The adaptive significance of the thermal biology of Gerbillurus is discussed in relation to phylogeny,
distribution, food availability and nocturnal activity.
Key Word Index: Gerbillurus; thermal preferences; temperature regulation; oxygen consumption; conduc- tance; evaporative water loss; behavioural avoidance
INTRODUCTION
TO interpret the adaptive physiology and ecology of desert rodents in an environment characterized by temperature extremes and water scarcity, various parameters of water usage, energy metabolism and thermoregulation must be investigated (MacMillen and Lee, 1970). The thermal biology of four Gerbillu- rus species of the arid regions of southern Africa is presented here as part of a comprehensive study of their ecophysiology.
Granivorous desert rodents including species of the Heteromyidae (McNab, 1979; Hinds and MacMillen, 1985), and Muridae (MacMillen et al., 1972; Shkol- nik and Borut, 1969) have lower basal metabolic rates (BM R) than predicted. Energy needs and overheating are reduced, and consequently water intake and evaporative water loss are reduced (McNab, 1979). However, other desert rodents including Leggadina (MacMillen and Lee, 1970) have normal or high BMR relative to predicted values. Two hypotheses have been proposed for the evolution of different levels of energy use among mammals (Thompson, 1988). Energy expenditure and thermoregulation are either phylogenetic in origin i.e. evolving at the time of the speciation event--the complete allopatric model; or are determined by reinforcement (ecolo- gical factors and competition) after the speciation event--the partial allopatric model. However, it is difficult to separate these alternatives.
Desert rodents that respond behaviourally to extreme climates through the exploitation of mesic micro-environments and nocturnalism, uncouple thermoregulation from evaporative cooling (Bartholomew and Dawson, 1974). Behavioural therrnoregulation infers that balance of water and energy budgets becomes the principal determinant of a desert existence for rodents. Water and energy
budgets of granivorous desert rodents are balanced either by reduction of BMR or increase in urinary concentrating abilities, or by a combination of both (McNab, 1979). Trends in BMR reflect differences in food selection, as a reduction in BMR is related to the availability and/or predictability of food resources. Water content of food and urine concentration also affect BMR (McNab, 1979).
Hayward (1965) suggested that plasticity in BMR had an insignificant role in climatic adaptation, distri- bution and speciation. He suggested that insulation and metabolic capacity at low temperatures and behavioural response to temperature were important, and were modified by utilization of a favourable microenvironment.
The closely related Gerbillurus species (Schlitter et al., 1984; Qumsiyeh et al., 1987), G. paeba paeba, G. tytonis, G. setzeri and G. vallinus vallinus that inhabit arid areas of southern Africa (Downs and Perrin, 1989a), are omnivorous and opportunistic feeders (Boyer, 1988; Downs and Perrin, 1989b), with efficient urinary concentrating abilities (Downs and Perrin, 1989c). Consequently the objectives of this study were (1) to estimate BMR of each species and relate them to water and energy budgets and/or phylogeny; (2) to define the zones of thermoneutrality (TNZ) and thermal preferences and correlate them with microenvironment and (3) to quantify the degree of temperature regulation above and below the TNZ, concurrently with conductance, evaporative water and heat production.
MATERIALS AND METHODS
Collection and maintenance of animals
Gerbillurus species were collected at various sites in SWA/Namibia and the Northern Cape, and kept in
291

292 COLLEEN T. DOWNS and M. R. PERRIN
the laboratory as described previously (Downs and Perrin, 1989b).
Thermal preferences
A box ( l .50m × 0.03m x 0.03 m) heated at one end, in a constant environment (CE) room of 15°C ( _ I°C) had a temperature gradient of 20-45°C along its length. The base of the box was covered with sand. Artificial burrows (glass bottles, 300 ml) were placed at intervals along the thermocline, beneath tempera- ture recording ports. Experiments were conducted during the gerbils inactive phase. Individuals were placed midway along temperature gradient to explore for an hour. Position in the thermocline, posture and activity of each animal was noted hourly from 0900 to 1700 h. Control experiments were done at a spa- tially regular ambient temperature of 25'~C to deter- mine if there were any behavioural preferences for particular positions in the tank.
Heating curves
An individual of each species was euthanased, skinned and used as the cast for a hollow brass-alloy core for a taxidermic model (Bakken, 1976). Com- pleted taxidermic models were frozen to a tempera- ture of - 18°C (+ 2°C). Each model was removed to a CE room (30 + I°C) where a copper-constan ther- mocouple connected to a digital display thermometer (Digitron Instrumentation, Model 1408K) was in- serted into the body of the model through the buccal opening. Model body temperatures were recorded 1 min after removal from the freezer, every minute until model body temperature reached ambient tem- perature.
Thermal parameters
Effects of ambient temperature (T,) on body tem- perature (Tb), oxygen consumption (Vo2), evapor- ative water loss (EWL) and thermal conductance of adult Gerbillurus species were determined. Individuals were weighed and then acclimated for a minimum of 3 h at the experimental temperature in a CE room (T,_+ 2°C, r.h. 50% + 5%), however at 5, 10 and 40°C, acclimation period was reduced to 1 h. Experi- mentation was done during the day when the gerbils were inactive, non-fasting and post-absorptive.
A copper-constan thermocouple connected to a digital display thermometor measured temperature of the respirometer chamber and rectal Tb of the gerbils. The probe was inserted _+7.5mm into the rectum. Oxygen consumption and evaporative water loss were measured simultaneously at each temperature using an open-circuit airflow system (Depocas and Hart, 1957). Air was dried over silica-gel before flowing into a round glass respirometer chamber (210ml) and again before entering the oxygen analyser (Amtek S-3A/I). Flow rates (8.0-8.5mm Hg con- verted to ml/min Air 760 mm Hg) were controlled and measured using a mercury flowmeter (Tube RSI, Float Hi-Den Air, Glass Precision Engineering). Carbon dioxide was removed from the air before entering the 02 analyser over a column of soda lime. A data logger (chart recorder, Model RYT) running at 2 mm min t recorded Vo, continuously. Initially oxygen consumption was measured by a Beckman OM 14 oxygen analyser but it overestimated readings
by a fixed factor at each temperature. A correction factor, determined by running standard air samples through both types of oxygen analysers under set conditions, was established.
After a standard run (30 rain) to determine average percent oxygen influx, each S was placed in the respirometer chamber for a further 30min. Mean highest percent oxygen recorded for 5 min intervals was calculated as efflux. Influx and efflux were corrected to STP, for flow rate and used to calculate Vo2 using the equations of Hill (1972).
Mass increase of the silica drying column (measured to 0.1 mg) used to dry excurrent air, was used to calculate residual water content of incurrent air during calibration, and evaporative water loss of each gerbil while in the chamber. Measurements were discarded if animals urinated in the respirometer chamber.
Dry thermal conductance (Cd), the rate of dry heat transfer per unit area to/from an animal per C temperature difference between the animal and its environment, excluding heat lost through evaporative water loss, was calculated according to Dawson and Schmidt-Nielsen (1966).
At T,s below the TNZ, rate of metabolism of a homeotherm is proportional to the temperature differential between the body and the environment. Minimal thermal conductance (Cm~n), including EWL, measures ease of heat loss when metabolic rate approximates resting metabolism, and was calculated using the formula of Scholander et al. (1950).
Behavioural thermoregulation
Postures of animals were noted during each experiment, together with evidence of salivation, vasodilation or shivering.
Statistics
Student's t-tests and l-way analysis of variance were done using Minitab5 and Statgraphics packages with 0.05 as the level of significance. Regression equations were calculated by the method of least squares using all individual data points.
RESULTS
Thermal preferences
Gerbils moved randomly within the thermocline for approx. 1 h before selecting a temperature where they rested. Figure 1 shows the frequency distribution of gerbils under experimental (thermal gradient) and control conditions (no thermal gradient). With the exception of G. v. vallinus, preferred temperatures approximated to 28°C while this species had a prefer- ence for corners irrespective of temperature gradient. At 25°C, controls distributed themselves randomly, or selected corners.
Thermal behaviour
At 10°C animals occasionally shivered, while at T~s below 20~C all individuals lay in a crouched position. Above 20°C, animals lay stretched out, but above 35°C each species began to salivate, wetting the neck region. Above 35°C, all animals showed vasodilation especially in the toes, ears and tails and the skin was visible as a result of piloerection. Sand-bathing

3,5
3 2 o v
29
P 26 o. E 23
2O
Thermal parameters of four species of Gerbillurus
(1o) IIIIIIII l I (lb) IIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
I 23 I z°] I I I I , J I :
4 3 2 1 0 1 2 3 4
(20)
1111111111111111111111111111111111111 I , l l l , i l
I I I I I I I I I I I I I I I I I I I I I I I I I , , , , I I
5 4 3 2 1 0 1 2 3
293
(2b)
3 5
32
29
26
E 23
20
6
- (50) I IIIIIIIIIIIIIIIIIIII
IIIIIIIIIIII111111111111111111111
I I 4 2 0
Frequency
(3b)
I • 2 4
35 (40)
32
29
26
23
2O
6
IIIIIIIIIIIII I IIII1!1111111111111,1111111111111, I L
4 2 0 2
Frequency
Fig. 1. The distribution of four Gerbillurus species in (a) a temperature gradient and (b) no temperature gradient (T a = 25°C) for (1) G. tytonis, (2) G. p. paeba, (3) G. setzeri and (4) G. v. vallinus.
1 (,b)
behaviour was displayed when the animals were returned to their vivaria after experimentation at Tas above 35°C.
Oxygen consumption
Figure 2 summarizes changes in specific Vo2 with change in Ta. Best-fit curves were determined for change in Vo~ with T~. Oxygen consumption increased below the TNZ in response to cold. Regression curves were used to determine the limits of the TNZ for each species by assuming the slope in the TNZ approached zero. Segmented line ana- lysis was done to show that slopes and transition periods between slopes were not significantly different to those obtained using linear regression analysis. Limits of the TNZ, BMR, and mean masses of the four species are shown in Table 1. BMR values expressed as a percentage of predicted values (calculated using the Kleiber (1975) relationship M = 3 . 8 W -°25) and lower critical temperatures calculated according to Newton's Law (Lc= Tb- -3 .8W °23, Hart, 1971) are also shown in Table 1.
To determine whether differences in metabolic response to cold temperatures were a function of body mass, slopes of regression equations below the TNZ were plotted against body mass. There was no significant correlation (r2 --- 0.07) specifying that species differences in heat production are not related to decreasing ratio of surface to body mass with decrease in body mass.
Dry thermal conductance (Ca)
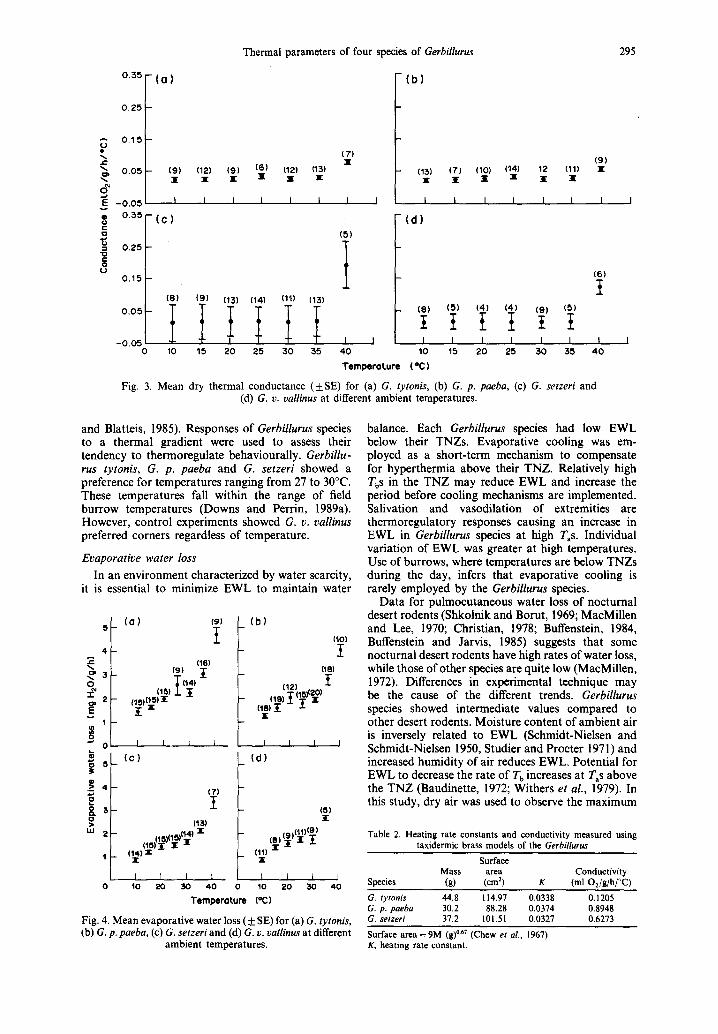
Figure 3 illustrates the change in Cd with tem- perature. Gerbillurus tytonis and G. p. paeba had a small but significantly higher conductance than the other two species at 25°C. The Cd of all four species
was low below the TNZ. Above 35°C the Cd of G. setzeri and G. v. vallinus increased sharply and were two to three-fold higher than G. p. paeba and G. tytonis.
Minimal "wet" conductance
The "critical gradient" (the difference between the critical temperature and Tb) is a measure of the insulative properties (Scholander et aL, 1950) and was calculated for each species. Results are shown in Table I together with predicted Cmi n (Herried and Kessel, 1967) values for 30°C. Cmi, values were calculated for other temperatures and are shown in Fig. 4.
Heating rates were measured using a taxidermic model placed in a constant temperature. Logarithmic temperature differential was plotted against time to obtain rate of conductance. Table 2 includes rate constants and calculated conductances of the taxider- mic models using the formula of Bakken (1976). Heat capacitance of models were computed using the mass of the model and the specific heat capacity of copper (Kane and Sternheim, 1978) and converted to J/g/°C.
Body temperature
Changes in T b with T a for the four Gerbillurus species are shown in Fig. 5. Below 35°C, Tb of each species was not significantly different at each exper- imental temperature. Above 35°C, all four species became hyperthermic with Tb significantly higher than values at lower temperatures. Exposure to Tas/> 40°C for periods > 1 h caused death in all four species. To show that Tb was independent of T,, the Tb predicted from the regression equation and the mean measured values below the TNZ are shown in Table 3.

294
£:
E
E
E g x O
3 5
3 0
2 5
2 . 0
1 . 5
1 . 0
0 . 5
0 3 . 5
3 . 0
2 . 5
2 . 0
1 . 5
1 . 0
0 . 5
COLLEEN T. DOWNS and M. R. PEPJUN
( a ) (14) E (151
E
(15) ( 9 1
113) ( 1 6 ) lr
( 9 )
E
_ y = 4 . 2 3 5 - 0 . 0 9 8 x r 2 = 0 . 7 6 d.f.= 6 4
y = - 2 . 6 7 6 + 0 . 1 0 7 x r 2 = 0 . 5 4 d . f .=23
I I [ I [ [ I I I
( c )
(11)
E (13)
! (13)
(13)
(8) (13)
E (13) Z
y = 3 . 9 2 4 - 0 . 0 9 7 x r 2 = 0 . 8 5 d.f. = 61
y = - 1 . 2 5 2 + 0 . 0 5 9 x r z = 0 . 4 9 d,f, = 19
J I I I I J ] I I .'5 10 15 2 0 2 5 50 35 4 0 4 5 0
( b ) (17)
t (19)
(17) [
(17) ¥
(18) ~- (16)
(10)
_ y = 4 . 2 9 0 - 0 . 1 0 1 x r e = 0 . 7 7 df. = 8 6
y = - 2 . 4 8 2 + 0 . 1 0 0 x r e = 0 . 3 9 d.f. = 2 4
I I I I I I I I I
-(d)
( 9 )
( 8 )
~" (81 E
(10)
(10) ~- (11)
l r
18)
t
y = 3 . 4 4 5 - 0 . 0 7 7 e r e = 0 . 8 3 dr. = 4 8
y = - 4 . 6 4 0 + 0 , 1 5 8 r 2 = 0 . 7 2 d.f. = 17
I I I I I I I I I 5 10 15 2 0 25 3 0 3 5 4 0 4 5
T e r n p e r o t u r e ( * C )
Fig. 2. M e a n rate o f oxygen c o n s u m p t i o n (_+SE) for (a) G. tytonis, (b) G. p. paeba, (c) G. setzeri and (d) G. v. vallinus at different amb ien t t empera tures .
Evaporative water loss
Pulmocutaneous water loss of each Gerbillurus species was low and not significantly different among Tas below 35°C (Fig. 6). However, EWL increased sharply from 35°C when animals were hyperthermic and began to salivate. To show the importance of EWL in increasing heat loss, the ratio of EWL: Vo2 was calculated for each experimental T,. EWL: Vo2 did not differ significantly between /'as below 30°C among the four Gerbillurus species. However, it was significantly different between 10 and 30°C. All four species showed a sharp increase in EWL: Vo2 above 30°C.
DISCUSSION
Thermal preferences
Diurnally fossorial habits and nocturnal activity of small desert rodents prevent hyperthermia and allow therrnoregulation to be uncoupled from obligatory evaporative water loss. Gerbillurus species exemplify behaviourai thermoregulation, taking advantage of a stable burrow micro-environment secluded from the extreme and harsh macroenvironment (Downs and Perrin, 1989a).
Monitoring of preferred T~ is a common means of assessing behavioural thermoregulation (Laughter
Table 1. Thermal parameters of four Gerbillurus species
Species G. tytonis G. p. paeba G. v. vallinus G. setzeri
n t6 16 11 13 Mass (g) 29.9 4- 4.76 33.9 4- 6.93 38.8 4- 6.37 46. I 4- 10.32
TNZ 32.4 - 34.9 32.3 - 35.1 33.1 - 35.0 32.2 + 34.8 T~¢C C) 28.6 30.2 28.6 28.4 BMR 1.060 + 0.048 1.027 4- 0.086 0.896 4- 0.039 0.803 4- 0.033 BMR a 1.468 4- 0.015 1.408 4- 0.037 1.371 4- 0.017 1.323 4- 0.024
% 72.1 75.0 65.25 60.63 Cm, . 0.1680 4- 0.0072 0.1367 4- 0.0104 0.1300 4- 0.0089 0.1525 4- 0.0194 Cmin b 0.1826 4- 0.004l 0.1678 4- 0.0059 0.1591 4- 0.0044 0.1469 4- 0.0051
% 92.3 81.9 81.5 90.2
TNZ, thermoneutral zone. T~¢, predicted lower critical temperature (Bartholomew, 1977). BMR, basal metabolic rate (mean + SEM, ml O:/g/h). BMR% calculated using the formula of Kleiber (1975). Cm~ ., minimum conductance (mean 4- SEM, ml O2/g/hpC). Cml, ~, calculated using the formula of Herried and Kessel (1967).
0.35
0.25
0.1,5
" 0.05
E -0 .05 ~ 0.35
o
0.25 "o g
0.1,5
0.05
-0.05 0
Thermal parameters of four species of Gerbillurus
to)
(9) (12) (9) (6) (12) "It • ][ ~ •
I I I I I
(c)
( 8 ) ( 9 ) (13) (14) (11) (13)
10 15 20 25 30 35
(7)
(13)
I I
- ( b )
i
J - (13)
x
I i
(d) (5)
t - - (81 151
t 1[ I I I I
40 10 15
(7) (10) (14) 12 (11) I ] [ • ][ ][
(9) K
295
I I I I I ! I
(6)
t (4) (4) (9) (5)
[ t t t I I I I I
20 25 30 35 40
T e m p e r o t . u r e ( * C )
Fig. 3. Mean dry thermal conductance (+S E ) for (a) G. tytonis, (b) G. p. paeba, (c) G. setzeri and (d) G. v. vallinus at different ambient temperatures.
and Blatteis, 1985). Responses of Gerbillurus species to a thermal gradient were used to assess their tendency to thermoregulate behaviourally. Gerbillu- rus tytonis, G. p. paeba and G. setzeri showed a preference for temperatures ranging from 27 to 30°C. These temperatures fall within the range of field burrow temperatures (Downs and Perrin, 1989a). However, control experiments showed G. v. vallinus preferred corners regardless of temperature.
Evaporat ive water loss
In a n e n v i r o n m e n t c h a r a c t e r i z e d by w a t e r scarc i ty , it is e s sen t i a l to m i n i m i z e E W L to m a i n t a i n wa t e r
4
o
2 E
1
~ 4 P g 3 o >
w 2
( o ) {9)
t 116)
(9 )
(1~) t (1.~) ( 1 5 ) 1 1 5 ) Z i x
(b) 110)
t 1181
(19) "~" (18) l r
.., i I I I
( d )
I I i i
( e )
(7)
t 113)
1 1 5 ) I I 114) l r l r
I I i I I 10 20 30 40 0 10
Temperoture (°C)
( 5 ) I
(9)111)(9) (8) I T t
(11) "It "x
I i I 20 30 40
Fig. 4. Mean evaporative water loss ( + SE) for (a) G. tytonis, (b) G. p. paeba, (c) G. setzeri and (d) G. v. vallinus at different
ambient temperatures.
balance. Each Gerbillurus species had low EWL below their TNZs. Evaporative cooling was em- ployed as a short-term mechanism to compensate for hyperthermia above their TNZ. Relatively high TbS in the TNZ may reduce EWL and increase the period before cooling mechanisms are implemented. Salivation and vasodilation of extremities are thermoregulatory responses causing an increase in EWL in Gerbillurus species at high Tas. Individual variation of EWL was greater at high temperatures. Use of burrows, where temperatures are below TNZs during the day, infers that evaporative cooling is rarely employed by the Gerbillurus species.
Data for pulmocutaneous water loss of nocturnal desert rodents (Shkolnik and Borut, 1969; MacMiilen and Lee, 1970; Christian, 1978; Buffenstein, 1984, Buffenstein and Jarvis, 1985) suggests that some nocturnal desert rodents have high rates of water loss, while those of other species are quite low (MacMillen, 1972). Differences in experimental technique may be the cause of the different trends. Gerbillurus species showed intermediate values compared to other desert rodents. Moisture content of ambient air is inversely related to EWL (Schmidt-Nielsen and Schmidt-Nielsen 1950, Studier and Procter 1971) and increased humidity of air reduces EWL. Potential for EWL to decrease the rate of T b increases at Tas above the TNZ (Baudinette, 1972; Withers et al., 1979). In this study, dry air was used to observe the maximum
Table 2. Heating rate constants and conductivity measured using taxidermic brass models of the Gerbillurus
Surface Mass area Conductivity
Species (g) (cm 2) K (ml O2/g/h/°C)
G. tytonis 44.8 114.97 0.0338 0.1205 G. p. paeba 30,2 88.28 0.0374 0.8948 G. setzeri 37.2 101.51 0.0327 0.6273
Surface area = 9M (g)O,67 (Chew et al., 1967) K, heating rate constant.

296 COLLEEN T. DOWNS and M. R. PERRIN
0.85
0.68
0 .51
0 . 3 4 0
f 17
~ 0 ~ 0 . 8 5 E
~ 0 . 6 8 E
0.51
0 . 3 4
0 . 1 7
(a)
I
(c)
(117 12 (11] =. I- T
I I I
( 6 ) (15 ) 14 I
I 1 I
114) (7) (161 (161 (12) = i" = •
I l I 1 I
(11:2) (13711t. (11:1)
I I I 2 5 3 0 3 5
( b )
16)
t
I I
(d)
(61
t
I I 40 0 5
(9)
112) I
I I (8)
t
(81x (91 (1.~1_ 177. (61. 141. (61. (97. (91z I [ J I [ ] J I I I I
0 5 10 15 20 10 15 20 25 :50 35 40
T e m p e r 0 t u r e (*C)
Fig. 5. Mean C~i . values (_+SE) for (a) G. tytonis, (b) G. p. paeba, (c) G. setzeri and (d) G. ~. vallinus at different ambient temperatures.
use of evaporative cooling but gives a limited under- standing of the mechanism of evaporative water loss. Christian (1978) found EWL correlated positively and significantly with humidity at 23°C for G. p. paeba. Preliminary studies of burrow systems show that humidities are high in the morning decreasing sharply through the day (Downs and Perrin, 1989a). However, burrow temperatures are below Tas where evaporative cooling is employed (Downs and Perrin, 1989a) enhancing the economic use of water.
Body temperature
Endothermic, nocturnal desert rodents (when active or resting), have body temperatures ranging between 36 and 39°C that invariably exceed the T~s to which they are exposed (MacMillen, 1972). Body- temperature range of each Gerbillurus species was broad but Tb was always greater than Ta, but below 25°C, Tb was independent of T, indicating good temperature regulation.
Previously G. p. paeba has been shown to be thermolabile, especially below 25°C (Nel and Rauten- bach, 1977; Buffenstein, 1984) suggesting decreased precision in temperature control. Results of this study indicate a certain degree of thermolability within a range that allows maintenance of endothermy. A decrease in body temperature of only a few degrees during its inactive phase saves considerable energy for a small animal whose energy sources are limited, but not as great as those resulting from torpor. Gerbillurus species showed good regulatory ability by
Table 3. Body temperature of four Gerbillurus species Species Tb" T b 25'C T b 30C G. O'tonis 43.21 34.9 + 1.59 36.9 _+_ 1.02
( ]0 ) (14) G. p. paeba 42.48 36.2 + 1.48 38.7 _+ 1.19
(18) (14) G. r. vallinus 44.74 38.0 + 1.43 37.4 + 1.29
(6) (70) G. setzeri 40.45 36.1 + 1.98 37.6 i 1.06
(75) (12) Tb ~, predicted from regression equations.
increased Tb during cold stress. Microtine rodents (Hart, 1971) similarly increase 7"8 during cold stress. Gerbillurus species have to contend with low surface Tas while active at night. However, heat loss would be offset by increased metabolism resulting from exercise and the uptake of food (Collins and Bradshaw, 1973).
Gerbillurus species increased body temperature in the upper TNZ before energy or evaporative cooling mechanisms were employed. This trend frequently occurs in nocturnal rodents (Shkolnik and Borut, 1969; MacMillen and Lee, 1972; Haim, 1984) where tolerance to hyperthermia increase before costly cool- ing mechanisms are employed and allows a greater dependence on physical mechanisms. Yet this in-
4 4
4 2
40
38
3 s o v
3 4 3 2 3 2 o 4 4
42
~" 4 o ID
38
36
34
(a) (8) v
114} ]¢
112) 11.221 "-i- (12) 114)
t _T. ! (lO) "¢ [
I [ I I I I I I I
( c ) (8) I
(13) !
(11) (14} !11211 1~1
];" ~1151 R-
Z
(9) (b ) x
1151 1"
1141 115)17) i
I 711~1 ± t1' '
I i I I I I I I
(d ) (s) t
(10) ar
( 6 ) ( 4 ) ~ 110)
I [ l l l l l l 3 2 [ I I I I I I I I 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0
Temperoture (%)
Fig. 6. Mean body temperature (+SE) for (a) G. tytonis, (b) G. p. paeba, (c) G. setzeri and (d) G. v. vallinus at different
ambient temperatures.

Thermal parameters of four species of Gerbillurus 297
creased Tb in the TNZ was also a direct consequence of the proportional increase in Tb with ambient temperature from 25°C for Gerbillurus species, and probably represents the threshold above which evaporative cooling is essential.
Despite tolerance to hyperthermia, each Ger- billurus species used specific physiological and/or behavioural mechanisms in extreme heat. Duration of high temperature exposure is important in toler- ance, since the Gerbillurus species could not tolerate high temperatures for periods > 1 h, emphasizing the importance of behavioural avoidance of these temperatures.
Thermal conductance
Tb is an interaction of metabolism and conductance and not in itself an independent character (McNab, 1966) but low Cmi ~ values indicate efficient use of physical thermoregulation (Aschoff, 1981). However, high Cmi n values can be advantageous if they reduce heat storage and prevent thermal death (Buffenstein, 1984). Decreased values reflect very good insulation which reduces the rate of heat loss at low Tas while limiting physical heat gain in the TNZ. Calculated conductances of taxidermic models are similar to Cmi a
values at 40°C, suggesting that the gerbils lower conductance physiologically and behaviourally at lower ambient temperatures. Variation in body size among species may represent the optimum size for the control of heating and cooling rates.
Use of power equations for the description of aliometric relationships between heat exchange and body size have been reviewed and criticized exten- sively (Turner, 1988). Use of equations derived from first principles of heat transfer are statistically and biologically more correct, particularly for interpret- ing the thermal energetics of animals differing in body size. There was no linear relationship between C~n and logarithmic body mass for the Gerbillurus species.
Extensive and dogmatic use of power equations makes comparative analysis of adaptations in ther- mal energetics difficult. Lower than predicted rates using power equations of conductance have been found for cricetids (Bradley and Deavers 1980), the heteromyids (Carpenter, 1966; McNab, 1980) and G. pusillus (Buffenstein and Jarvis, 1985) while higher values are found in Skeetamys calurus (Haim and Borut, 1986). Higher-than-predicted values of ther- mal conductance are important for heat dissipation in a habitat where water is limited (Haim and Borut, 1986). Also, at lower Tas survival would be dependent on increased heat capacity rather than reduced con- ductance. Gerbillurus setzeri, G. v. vallinus, G. p. paeba and G. tytonis had Cmi n values 109.2; 85.1; 78.5 and 96.0% respectively of predicted values using Herried and Kessels' equation (1967).
Dry thermal conductance
Cd is affected by a highly complex interaction of environmental factors with physiological and anatomical features such as pilosity, piloerection, subcutaneous fat deposition and vasomotor changes (Herreid and Kessel, 1967). Dry thermal conductance is indirectly measured from T a - Tb, BMR and the
rate of EWL. Below 30°C, Cd is consistently low for each Gerbillurus species with Vo2 differences being balanced by differences in T a and T b. At high tem- peratures Cd increases sharply representing a decrease in insulation to facilitate heat loss. Gerbillurus setzeri and G. v. vallinus show a sharper increase in Cd at temperatures above TNZ compared to the two dune species. McNab (1966) suggested that the reduction of BMR and increase in Cd in fossorial rodents was directly related to mean burrow temperatures to reduce the probability of overheating in an environ- ment where EWL is reduced. For G. setzeri and G. v. vallinus a lower rate of increase in Vo2 and a sharp increase in Cd at temperatures above the TNZ may reflect an avoidance of EWL to reduce overheating. Although heating rate constants of Gerbillurus species are similar, the two gravel-plain species are larger and have thicker pelages (Schlitter et al., 1984, personal observation) facilitating a greater change in Cd. Lower conductance may represent an adaptation to prevent excessive heat loss in fossorial rodents (Gettinger, 1975). Below 30°C all four species of Gerbillurus have low Cd values, particularly the two larger, gravel plain species. This reduces heat loss when exposed to low /'as at night and enhances temperature regulation.
Oxygen consumption
Comparison of change in specific Vo~ with change in Ta showed the typical endothermic response for each Gerbillurus species, i.e. Vo2 decreasing linearly to the lower limit of the TNZ and then increasing above the upper critical temperature limit, with increase in Ta. When the logarithm of Vo2min (BMR) was plotted against the logarithm of body mass for the four species, there was no significant linear relationship (d.f. = 55, t2= 0.013) despite smaller species having greater Vo2.
The relationship between body size and BMR suggests that similarities and differences in BMR of the four species are related to phylogeny, size and habitat. Gerbillurus tytonis and G. p. paeba are closely related as are G. setzeri and G. v. vallinus (Qumsiyeh et al., 1987). The former two species are smaller than the latter two species. In addition G. tytonis and G. p. paeba occur in sandy substrates while the other two species have a preference for gravel plains (Downs and Perrin, 1989a). Differences in food availability and quality (in terms of water and energy content) between gravel and sandy habitats may be important determinants of size and BMR.
Desert mammals usually exhibit a gradient below the 0.25 slope of the typical mammalian BMR vs body mass curve, as the BMR values are significantly lower than predicted by Kleiber's (1975) equation (Yousef and Johnson, 1975). Observed BMR values of each Gerbillurus species were lower than the pre- dicted values on the basis of body mass, while in the laboratory, DEE values obtained during feeding trials were higher than expected for body mass (Downs and Perrin, 1989b).
Previously, lower relative metabolic rates of desert mammals have been related to scarce and/or unpre- dictable food resources (Yousef and Johnson, 1975; McNab, 1979, Buffenstein, 1985; Haim and Borut,

298 COLLEEN T. DOWNS and M. R. PERRIN
1986). Reduced BMR lowers energy needs and con- serves water. However, those desert rodents with efficient urinary concentrating abilities or without torpor have BMR values closer to predicted values (McNab, 1979). No evidence of torpor was observed in any Gerbillurus species (personal observation). The two levels of modifying water loss, reduced BMR and efficient urinary concentrating abilities are generally combined, but ultimately they are related to food selection (McNab, 1979).
For Gerbillurus species, the limiting factors of an arid environment are food and water availability because of behavioural temperature avoidance. The gerbils rely on metabolic and preformed water (Downs and Perrin, 1989b). The gravel-plain species, G. setzeri and G. v. vallinus have BMR values 35% lower than predicted compared to the other two species which are 25% lower using Kleiber's equation (1975). The larger body size of G. setzeri and G. i~. vallinus reduces the differences between total water loss and metabolic production, thereby reducing the role of metabolism in water budgets. The lowered BMR of G. setzeri together with their efficient urinary concentrating ability (Downs and Perrin, 1989d) sug- gest selection to reduce water loss.
Comparison of BMR with other species is only possible if the experimental conditions are well defined and equivalent. G. p. paeba values of BMR (Buffenstein, 1984) are lower than values determined in this study, however the slope and intercept of the regression equation below the thermoneutral zone are similar.
The TNZ is the range of temperatures where heat balance is maintained by changes in total conduc- tance. The TNZ of G. setzeri, G. r. vallinus and G. tytonis were similar, overlapping and narrow. G. v. vallinus had a higher lower critical temperature L~ and a narrow TNZ. Desert rodents have narrow TNZs particularly if high temperatures are avoided behaviourally (Hart, 1971). The TNZ range for each gerbil species was higher than the maximum temperatures measured in their respective burrow micro-environments (Downs and Perrin, 1989a). Environmental temperatures usually encountered are lower than the TNZ so metabolic rates are usually elevated above basal rates. This relationship between metabolic rate and encountered temperatures would contribute towards the selection for lowered BMRs so that metabolic rates at burrow temperatures would be relatively low. For Gerbillurus species, metabolic rates at burrow temperatures (28-30C) are similar to predicted BMR values (Kleiber, 1975) suggesting that the high and narrow TNZs and lowered BMR values are genotypic adaptations to lower energy require- ments in response to the xeric environment where food availability is unpredictable.
The phenotypic response is a consequence of the micro-environment where temperatures lower than the TNZ are experienced and metabolic rates are approximately equal to predicted values for body size. Changes in insulation would decrease the slope of metabolic rate versus ambient temperature, lower- ing energy requirements as temperature decreases (Cossins and Bowler, 1987). A combination of low- ered BMR and increase in insulation would be opti- mal. Slopes of the curves below the TNZ were similar
for the different Gerbillurus species because of lower and similar insulative capacities. Increased heat pro- duction below the TNZ is also a temperature adapta- tion to maintain Tb. The ability to maintain Th at low temperatures would be important to the Gerbillurus species when active on the surface during the cool nights typical of the Namib (Downs and Perrin, 1989a).
The critical temperature of a homeotherm is defined as the temperature at which physical regu- lation is insufficient to maintain T b and chemical heat production is induced (Hayward, 1965). Pre- dicted lower critical temperatures (L~) (Hart, 1971) for each species were lower than measured values. Haim and Borut (1986) suggest that these differences may point to an adaptation to a hot and arid environment.
Thermoregulatorv abilio"
The ratio, percentage of expected I~o:: percentage of expected Cm~n, may be used as an index of the maintenance of the relative degree of endothermy at the lower limit of thermoneutrality (Bradley et al., 1975; McNab, 1980; Buffenstein, 1984). A ratio of one or above indicates a well developed and main- tained endothermy. Ratios obtained for G. tytonis, G. p. paeba, G. v. vallinus and G. setzeri are 0.76; 0.92: 0.77 and 0.56 respectively. Precision of endothermy appears to be far greater in G. p. paeba than in the other three Gerbillurus species, particularly G. setzeri. The complex interaction between metabolism, con- ductance and thermoregulation for each Gerbillurus species were further evaluated using the "'thermo- regulatory index" model of Tomasi (1985) for evalu- ating relative thermoregulatory ability. Gerbillurus O'tonis, G. p. paeba, G. t,. vallinus and G. setzeri had values of 14.27; 12.78; 21.77 and 19.58 respectively. The latter two species appear to have greater thermo- regulatory abilities which contrasts with the former index of thermoregulatory ability. These two gravel- plain species, by virtue of their size and reduced BMR, have a thermoregulatory advantage. For Ger- billurus species generally insulation and metabolic capacity at temperatures below the TNZ are ecologi- cally important determinants of thermoregulation. Furthermore BMR is an intrinsic factor affecting thermoregulation. BMR is in turn affected by phylo- genetically inherited thresholds and modified by tem- perature and energy availability. Increased body size attenuates BMR further. Consequently it would ap- pear that the larger species that occur on the gravel plains are adapted to colder temperatures and more erratic food supply by their thermoregulatory charac- teristics.
Analysis of the thermal biology ot these four Gerbillurus species suggests that water conservation is a more limiting factor determining their survival than temperature regulation considering their ability to avoid high temperatures and exploit a mesic micro- environment. Secondly differences among the four Gerbillurus species in their ability to cope with cold temperatures and the capacity to generate heat using non-shivering thermogenesis (NST) would be import- ant to their use of the nocturnal niche. At low temperatures a rapid increase in heat production is necessary to compensate for heat loss.

Thermal parameters of four species of Gerbillurus 299
SUMMARY
The thermal physiology of Gerbillurus spp must be examined in relation to their micro-environment. Fossorial and nocturnal habits of these xeric rodents enables avoidance of stressful temperatures. Basal metabolic rates (BMR), thermal conductances and evaporative water losses were low in the thermo- neutral zones (TNZ), particularly for the larger two species G. setzeri and G. v. vallinus which occur on gravel plains. The TNZs of all four species studied are narrow and occur at high temperatures. The significance o f the thermal parameters are discussed in terms of Gerbillurus phylogeny, distribution, noc- turnal activity and food availability in xeric regions of southern Africa.
Acknowledgements--This work was supported by the Uni- versity of Natal and the FRD of the CSIR. The Department of Nature Conservation, SWA/Namibia is thanked for permission to trap animals.
REFERENCES
Aschoff J. (1981) Thermal conductance in mammals and birds: its dependance on body size and circadian phase. Comp. Biochem. Physiol. 69A, 611-619.
Bakken G. S. (1976) A heat transfer analysis of animals: unifying concepts and the application of metabolism chamber data to field ecology. J. theor. Biol. 60, 337-384.
Bartholomew G. W. and Dawson W. R. (1974) Tempera- ture regulation in desert mammals. In Desert Biology (Edited by Brown G. W.), Vol. 1, pp. 396-421. Academic Press, New York.
Baudinette R. V. (1972) Energy metabolism and evaporative water loss in the Californian ground squirrel. 3. comp. Physiol. 81, 57-72.
Boyer D. C. (1988) Effects of rodents on plant recruitment and production in the dune area of the Namib Desert. M.Sc. thesis, University of Natal, Pietermaritzburg.
Bradley S. R. and Deavers D. R. (1980) A re-examination of the relationship between thermal conductance and body weight in mammals. Comp. Biochem. Physiol. 65A, 465-476.
Bradley W. G., Yousef M. K. and Scott I. M. (1975) Physiological studies of the rock pocket mouse Per- ognathus intermedius. Comp. Biochem. Physiol. 50A, 331-337.
Buffenstein R. (1984) The importance of microhabitat in thermoregulation and thermal conductance in two Namib rodents--a crevice dweller, Aethomys namaquensis, and a burrow dweller, Gerbillurus paeba. J. therm. Biol. 9, 235 -241.
Buffenstein R. and Jarvis J. V. M. (1985) Thermoregulation and metabolism in the smallest African gerbil, Gerbillus pusillus. J. Zool. (Lond.) 205, 107-122.
Carpenter R. E. (1966) A comparison of the thermo- regulation and water metabolism in the kangaroo rats Dipodomys agilis and D. merriami. Univ. Calif. Publ. Zool. 78, 1-36.
Christian D, P. (1978) Effects of humidity and body size on evaporative water loss in three desert rodents. Comp. Biochem. Physiol. 60A, 425-430.
Collins B. G. and Bradshaw S. D. (1973) Studies on the metabolism, thermoregulation and evaporative water losses of two species of Australian rats, Rattus villosis- simus and Rattus rattus. Physiol. Zool. 46, 1-21.
Cossins A. R. and Bowler K. (1987) Temperature Biology of Animals. Chapman & Hall, London.
Dawson T. J. and Schmidt-Nielsen K. (1966) Effect of thermal conductance on water economy in the antelope jack rabbit. J. cell. Physiol. 67, 463-472.
Depocas F. and Hart J. S. (1957) Use of the pauling oxygen analyser for measurement of oxygen consumption of animals in open-circuit systems and short lag, closed- circuit apparatus. J. appl. Physiol. 10, 388-392.
Downs C. T. and Perrin M. R. (1989a) The macro- and micro-environments of four species of Gerbillurus. Cimbebasia 11, 41-54.
Downs C. T. and Perrin M. R. (1989b) The effect of diet on the energy and water turnover rates of four Gerbillurus species in the laboratory. J. Zool. (Lond.). In press.
Downs C. T. and Perrin M. R. (1989d) Urinary con- centrating abilities of four Gerbillurus species. J. Arid Envir. In press.
Gettinger R. D. (1975) Metabolism and thermoregulation of a fossorial rodent, the northern pocket gopher (Thomomys talpoides). Physiol. Zool. 48, 311-322.
Goyal S. P., Ghosh P. K. and Prakash I. K. (1982) Energetic adaptation in the Indian desert gerbil Meriones hurriannae (Jerdon). J. Arid Envir. 5, 69-75.
Haim A. (1984) Adaptive variation in heat production within gerbils (genus Gerbillus) from different habitats. Oecologica (Berl.) 61, 49-52.
Haim A. and Borut A. (1981) Heat production and dissi- pation in golden spiny mice, Acomys russatus from two extreme habitats. J. comp. Physiol. 142, 445-450.
Haim A. and Borut A. (1986) Reduced heat production in the bushy-tailed gerbil Sekeetamys calurus as an adap- tation to arid environments. Mammalia 50, 27-34.
Hart J. S. (1971) Rodents. In Comparative Physiology of Thermoregulation (Edited by Whittow G. C.), Vol. I, pp. 2-149. Academic Press, New York.
Hayward J. S. (1965) Metabolic rate and its tempera- ture adaptive significance in six geographic races of Peromyscus. Can. J. Zool. 43, 309-323.
Herded C. F. and Kessel B. (1967) Thermal conductance in birds and mammals. Comp. Biochem. Physiol. 21, 405-414.
Hill R. W. (1972) Determination of oxygen consumption by use of the paramagnetic oxygen analyser. J. appl. Physiol, 33, 261-263.
Hinds D. S. and MacMillen R. E. (1985) The scaling of energy metabolism and evaporative water loss in Heteromyid rodents. Physiol. Zool. 58, 282-298.
Kane J. W. and Sternheim M. M. (1978) Physics. Wiley, New York.
Kleiber M. (1975) The Fire of Life: an Introduction to Animal Energetics, 2nd edition. Krieger, New York.
Laughter J. S. and Blatteis C. M. (1985) A system for the study of behavioral thermoregulation of small animals. Physiol. Behav. 35, 993-997.
MacMillen R. E. (1972) Water economy of nocturnal desert rodents. Syrup. Zool. Soc. Lond. 31, 147-174.
MacMillen R. E. and Lee A. K. (1970) Energy metabolism and pulmocutaneous water loss of Australian hopping mice. Comp. Biochem. Physiol. 35, 355-369.
MacMillen R. E., Baudinette R. V. and Lee A. K. (1972) Water economy and energy metabolism of the sandy inland mouse, Leggadina hermansburgensis. J. Mammal. 53, 529-539.
McNab B. K. (1966) The metabolism of fossorial rodents: a study in convergence. Ecology 47, 712-732.
McNab B. K. (1979) Climatic adaptation in the energetics of heteromyid rodents. Comp. Biochem. Physiol. 62A, 813-820.
McNab B. K. (1980) On estimating thermal conductance in endotherms. Physiol. Zool. 53, 145-156.
Nel J. A. J. and Rautenbach I. L. (1977) Body temperatures of some Kalahari rodents. (Mammalia: Muridae, Criceti- dae). Ann. Trans. Mus. 30, 207-209.
Perrin M. R. and Campbell B. S. (1981) Some aspects of thermoregulation in three species of southern African tortoise. S. Afr. J. Zool. 16, 35-43.

300 COLLEEN T. DOWNS and M. R. PERRIN
Qumsiyeh M. B., Hamilton M. J. and Schlitter D. A. (1987) Problems in using Robertsonian rearrangements in determining monophyly: examples from the genera Tatera and Gerbillurus. Cytogenet. Cell. Genet. 44, 198-208.
Schlitter D. A., Rautenbach I. L. and Coetzee C. G. (1984) Karyotypes of southern African gerbils, genus Gerbillurus Shortridge, 1942 (Rodentia: Cricetidae). Ann. Cam. Mus. 53, 549-557.
Schmidt-Nielsen B. and Schmidt-Nielsen K. (1950) Pul- monary water loss in desert rodents. Am. J. Physiol. 162, 31 36.
Scholander P. F., Hock R., Waiters V. and Irving L. (1950) Adaptation to cold in artic and tropical mammals and birds in relation to body temperature, insulation and basal metabolic rate. Biol. Bull. 99, 259-271.
Shkolnik A. and Borut A. (1969) Temperature and water relations in two species of spiny mouse (Acomys). J. Mammal. 50, 245-255.
Studier E. H. and Proctor J. W. (1971) Respiratory gases in the burrows of Spermophilus tridecemlineatus. J. Mammal. 52, 631-633.
Thompson S. D. (1988) Thermoregulation in the water opossum (Chironectes minimus): an exception that proves a rule. Physiol. Zool. 61, 450-460.
Tomasi T. E. (1985) Basal metabolic rates and thermo- regulatory abilities in four small mammals. Can. J. Zool. 63, 2534-2537.
Turner J. S. (1988) Body size and thermal energetics: how should thermal conductance scale? J. therm. Biol. 13, 103-117.
Withers P. C., Lee A. K. and Martin R. W. (1979) Metabolism, respiration, and evaporative water loss in the Australian hopping-mouse, Notomys alexis (Rodentia: Muridae). Aust. Zool. 27, 195-204.
Yousef M. K. and Johnson H. D. (1975) Thyroid activity in desert rodents: a mechanism for lower metabolic rate. Am. J. Physiol. 229, 727 731.





























