Embed Size (px)
Citation preview

TL'MOUR NECROSIS FACTOR ALPHA AND ULTRAVIOLET LIGHT ACTIVATION OF PROGRAMMED CELL DEATH BY APOPTOSIS IN D.
MELANOGASTER
Maurice Ennis
A thesis submitted in conformity with the requirements for the degree of Master of Science
Department of Medical Biophysics University of Toronto
O Copyright by Maurice Anthony Ennis (2001)

National L i i 1+1 dcumda Bibliotheque nationale du Canada
vwe Mm-
Our PI --
The author has granted a non- exclusive licence aUowiDg the National Li'brary of Canada to reproduce, loan, distri'bute or seil copies of this thesis in microfonn, paper or electronic formats.
The author retah ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fkom it may be printed or otherwise reproduced without the author's pamission.
L'auteur a accordé une licence non exclusive permettant à la B&lioth&que nationale du Canada de reprcduire, prêter, distribuer ou vendre des copies de cette thése sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thése ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisafim.

-
to W B and W C .
Tumow Necrosis Factor Alpha and Ultraviolet Light Activation of L L - z
Pmgrammed Ce11 Death by Apoptosis in D. Melanoguster; Master of Science,
ZOO1 Maurice Anthony Ennis, Department of Medical Biophysics, University
of Toronto
Molecular signals that stimulate ce11 growth and differentiation are an essential
part of Drosophila development from embryo to adult. Programmed celi death is
an equally important and regulated process. Grim, reaper, and hid of the H99
chromosomal region have been identified as activators of apoptosis and essential
requirements for all observed prograrnmed cell death. To date no system of
ligands and receptors which signal to activate cell death has been identified in
Drosophila. The C terminal signallhg portion of murine Tumor Necmsis Factor-
alpha (TNFa) under contrd of a heat shock promoter was successfully
introduced in to D rosuphila gemiline b y P elemen t mediated transformation.
Expression of TNFa demonstrated a limited ability to activate reaper expression
and apoptosis. W exposure by cornparison showed a dramatic increase of both
reaper expression and induction of apop tosis. Preliminary resulk demonstra te
that selective P element responses may be obsewed by exposing imagina1 discs

Table of Contents
................................................................................................ Abstract i ......................................................................... List of Tables and Figures iv
.............................................................................. List of Abbreviations v
1 . Introduction ................................. 1.1 Programmed Cell Death in Living Organisms 1
..................................................................... 1.1.1 Apoptosis 1 1.1.2 Normal and Pathological Contexts of PCD .......................... 2 1.1.3 Programmed Cell Death in C . elegans ................................. 2 1.1.4 Programmed Cell Death in D . melanogaster ........................ 4
............. 1.1.4.1 Apoptosis During Drosophila Development 6 1.1 .4.2 Apoptosis Linked to Segment Polarity Signaling ....... 7
1.2 Caspases-The Core of the Apoptotic Effector Machinery ................. 9 1.2.1 Caspase Function and Activation ...................................... 9
........................................................... 1.2.2 Caspase Targets 10 ........................................................ 1.2.3 Caspase Inhibitors 10
1.3 Signals Leading to Apop tosis ..................................................... -11 1.3.1 TNF Signaling-Receptor-linked Apoptosis ..................... -11
.......................................................... 1.3.2 NF43 Signaling 14 ........ 1.3.3 Activation of Apoptosis by UV and Ionizing Radiation 14
............................................................ 1.4 Statement of Objectives 16
2 . Materials & Methods ................................................................... 2.1 Molecular Biology 18
2.1.1 LB Medium (Luria-Bertani Medium) ................................. 18 ...................................................... 2.1.2 LB-ampicillin Plates -18
........................................................... 2.1.3 Competent Cells 18 ............................................. 2.1.4 Restriction Enzyme Digests 19 ........................................... 2.1.5 Agarose Gel Electrophoresis 19
2.1.6 Isolating and Purlfylng DNA Fragments ........................... 20 ..................................................................... 2.1.7 Ligation 20
............................................................ 2.1.8 Transformation 20 .................................... 2.1.9 Small-scale Plasmid Preparations 21
2.1.10 Large-scale Plasmid Preparations .................................. 21 2.1.1 1 Subcloning and Confirmation of hs mTNF ...................... 22
.............................................. 2.1.12 Embryo Protein Lysates 22 ................................................................ 2.1.13 SDS-PAGE 23
............................................... 2.2 Drosophila Handling Techniques 24 ...................................................... 2.2.1 Drosophila Culture 24
........................................................ 2.2.2 Apple Juice Plates 24 ......................................................................... 2.2.3 Yeast 24
...................................................... 2.2.4 Embryo Collections 25

.......................................................... 2.2.5 Embryo Fixation 25 ................................................................. 2.2.6 Heat Shock 25 2.2.7 UVB Exposure of Drosophila Imagina1 Discs and Staining for
.......................................................... galactosidase Activity 26 ........................................................... 2.2.8 W C Treatment 26
.................................................. 2.2.9 Cuticule Preparations -27 ............................................................. 2.2.10 Injection Mix 27
............................................. 2.2.11 Germline Transformation 28 ............................................... 2.2.12 Mapping and Balancing 28
........................ 2.2.13 Mobilization to the Second Chromosome 29 ............. 2.2.14 Combination with rpr Iacz to make hsTNF.rprlacz 30
............................................. 2.2.15 Marking Hg9 with yellow* 31 ........................... 2.2.16 Anti fl-galactosidase Antibody Staining 31
...................... 2.2.1 7 Al kaline Phosphatase Reac tion Developing 32 ................. 2.2.18 Acridine Orange Staining (Stellar lab protocol) 32
.............................................................. 2.2.19 Photography 33
3 . Results ....... 3.1 P Element-mediated Transformation of w"" by pCaSpeR.hsTNF 34
................................................ 3.2 Heat-Shock Expression of mTNF 36 ................................ 3.3 hsrpt, hsTNF and UV Induction of Ce11 Death 37
......................... 3.4 hsTNF vs . W C Activation of reaperlacz Expression 39 ....................................... 3.5 hsTNF vs . UVC Activation of Apoptosis 40
................................................. 3.6 WC Response in Hg9 Embryos 41 ............... 3.7 Response of Other P Elements to UVB and UVC Exposure 43
4 . Discussion .............................................................................. 4.1 Objectives 46 ................................................................. 4.2 Surnmary of Results 46 4.2.1 hsreaper. UVC. and hsTNF Cause Lethality in Drosophila
.......................................... ............... ......... Embryos .. .. -46 4.2.2 UVC but not hsTNF Activates Expression of renperlacz in ............................................................ Drosophiln Emb ryos -48 4.2.3 W C but not hsTNF Activates Apoptosis in Drosophila -- ........................................................................... Embryos 5~
........................ 4.3 Results in Context of Current Models and Research 51 ............................................ ............................. 4.4 Future Wotk .. 52
......................................................................................... References -55
............................................................................... Tables and Figures -65

List of Tables and Figures
Table 1 Plasmid stocks Table 2 Drosophila stocks Table 3 Drosophila embryo WC-induced lethality Table 4 WB response of several Drosophila P element lines Table 5 UVC response of several Drosophila P element lines
Figure 1 Schematic diagram of TNF/Fas and UV signalling leading to the activation of apop tosis Figure 2 çdiematic of pCaSper-hsmTNF plasrnid construct Figure 3 Expression of TNFa protein in Drosophila embryos after heat shock Figure 4 Average embryonic lethality after expresson of reaper, tnfa, or exposure to W C Figure 5 Ernbryonic reaper and TNFa expression and UVC exposure cuticle phenotypes F i g w 6 Anti-kgalactosidase staining in reaperlacz Drosoplrila embryos after hsTNF expression and UVC exposure Figure 7 Acridine Orange Staining in Drosophiln embryos after UVC exposure and hstnfexpression Figure 8 UVC cuticle phenotype in H99-deletion Drosophila embryos Figure 9 ~galactosidase staining of PS62 Drosophila larval imagina1 discs

List of Abbreviations
A 0 DD DED FADD FasL ICE IAP kD LB NF-KB PCD RF SDS TNF TNFR TRADD TRAF UVB W C
Acridine Orange Death Domain Death Effector Domain Fas-associated Death Domain Protein Fas Ligand Interleukin-1-p-converting Enzyme Inhibitor of Apoptosis Kilodal tons Luria-Bertani Medium Nuclear Factor kappa B Programmed Ce11 Deah Receptor Interacting Protein Sodium Dodecyl Sulphate Tumour Necrosis Factor Tumour Necrosis Factor Receptor TNF-associated death domain protein TNF receptor-associated factor Ultraviolet B Ultraviolet C

1. Introduction
1.1 PROGRAMMED CELL DEATH IN LIVING ORGANISMS
1.1.1 Apoptosis
With the use of electron and light microscopy, Kerr et. al observed ce11
death in a variety of contexts. These included malignant neoplasms, tumor
regression after some forms of therapy, rat liver after obstruction of portal blood
supply, and in the adrenal gland after withdrawal of adrenocorticotrophic
hormone (ACTH). In al1 these situations the same rnorphological features of
dying cells were observed. A ce11 destined to die would first shrink along with
its nucleus and become separated from surrounding ceils. The ce11 was then
observed to disintegrate into fragments, forming "apoptotic bodies".
Professional phagocytes or neighbouring cells would then consume the
condensed cell and apoptotic bodies. These were then digested by fusion with
lysosomes leading to the formation of electron-dense lysosomal residual bodies
(1).
These researchers realized that this must be the form of ce11 death
involved in development and other situations where a reduction of ce11 nurnbers
is required to counteract mitotic increase of cell numbers. The observation that
the program for apoptosis was intrinsic to al1 cells suggested a genetic basis.
Although various forms of ce11 death had been previously desaibed in different
situations (2)) the work of Kerr et al. was important in i d e n m g the common
features of these ce11 deaths and ascribing them to a common intrinsic cellular
mechanism. Henceforth, the term Programrned Ce11 Death (PCD) will refer to

the overail phenornenon of deleting cells in an ordered manner. The term
"apoptosis" will refer to the cellular mechanism by which this deletion is carried
out displaying the specific characteristics described by Kerr and co-workers in
1972 (1).
1.1.2 Nomal and pathological contexts of PCD
During development PCD plays a role in shaping organs, creating vesse1
lumen, deleting extra, unnecessary, or undifferentiated cells, and for atrophy of
vestigial structures (3). The role of PCD also continues in living organisms past
development. In the adult mammal PCD serves to maintain a balance in ce11
populations by counteracting growth and eliminating unnecessary cells. As with
other processes of biological organisms, the misregulation of PCD by apoptosis is
involved in several pathological states, including cancer, AIDS, stroke,
myopathies, and various neurodegenerative disorders (4,5).
One example of the misregulation of programmed ce11 death in human
pathology is the case of human follicular lymphomas involving the proto-
oncogene Bel-2. In this type of cancer, a t(14;lB) chromosomal translocation
juxtaposes Bcl-2, a 25 kD protein which inhibits apoptosis (6), with the
immunoglobulin (Ig) heavy chah locus (7). The ensuing expansion of B ce11
populations in hurnan follicular lymphomas is the result of extended B ce11
survival, not increased ce11 proliferation (8,9).
1.1.3 Pmgrammed Cell Death in C. elegans
Research of the organism Caenorhabditis elegrms has made signihcant
contributions to understanding the genetic basis of PCD. One significant
2

advantage to working with this organism is that al1 ce11 fates in the developing
organism have been mapped. This means that the ultimate fate of each ce11 that
arises during development is known allowing researchers to trace the
development of an individual cell through its entire developmental life span.
Along with the 959 somatic nuclei that develop during the formation of the adult
worm fonning the adult organism, 131 cells develop and then die by apoptosis
(O, 11) Thus, cell death has been recognized as a specific ce11 fate in the
development of C. elegans. By studying mutations that affect the normal
progression of this process, mutations in specific genes that alter the normal
pattern of ce11 death have been discovered.
The first mutations observed prevent the phagocytosis of dead cells (12)
whidi has been characterized as one of the final steps of apoptosis. Genetic
screens using these mutants, called Ced-1 and Ced-2 (cell death-1 and ce11 death-
2) identified other genes involved in the process of programmed ce11 death in C.
elegans. In CED-3 and CED-4 mutants, the cells normally destined to die fail to
do so resulting in extra cells in the adult organism. Mutations of the Ced-9 gene
in C. elegans fail to inhibit apoptotic ce11 death. As a result, more cells than
normal enter the ce11 death program resulting in fewer cells in the adult.
Genetic epistasis allowed researchers to put the genes into a functional
order. They obsewed that Ced-3 and Ced-4 act upstream of the phagocytosis
mutations to activate ce11 death in a ce11 autonomous fashion. Ced-9, a Bd-2
homologue, acts upstream of Ced-4 and Ced-3 to inhibit apoptosis. CED-3 has
been identified as a caspase homologue. As will be discussed below, these
proteins have been identified as the core of the apoptotic effector machinery. The
mammalian version of CED-4, apoptosis activating factor-1 (Apaf-1), also
3

activates downstream caspases and is inhibited by its interaction with Bcl-2 (13).
The work in genetic model systems such as C. elegans has been critical in
identifymg the proteins that work together to accomplish the function of
programmed ce11 death.
1.1.4 Programmed Ce11 Death in D. melanogaster
Another model system available for the study of PCD is Drosophila
rnelanogaster commonly known as the fruit fly. This organism is more complex
than C. elegans both in tems of the number and types of cells and life cycle.
Whereas the adult C. elegans develops a total of 1096 ceils, the Drosophila eye
alone consists of over 900 cells (14). The development of Drosophiln, also more
complex, progresses from the fertilized egg which is laid by the female through
three lawal stages before pupariation at 120 hours after egg laying. After 4-4.5
days of pupariation, the adult fly emerges (14). Though biologically more
complex, researchers still have the abiîity to shidy genetics in Drosophila as in C.
elegans. Genetic mutations are often expressed as recognizable phenotypes in the
developing and adult organism allowing researchers to keep track of genetic
changes. Research in Drosophila is also beneficial to humans because this
organism uses many similar cellular regulatory mechanisms including pathways
that regulate growth, differentiation, and ce11 death. These can be studied in this
organism and the information obtained applied to manunalian models.
In Drosoghila melanogaster, apoptotic cell death is obsewed to be under
the control of regulatory and effedor genes. The most important initiators of
apoptosis known to date are the genes of the H99 duomosomal region. White et
al. reported that embryonic double mutants for this deletion region block the
4

normal incidence of apoptosis in the developing embryos and prevent the
initiation of apoptosis in response to X-irradiation (15). In this research, White, et
al. also discovered that the expression of reaper, one of the genes located in this
region, is followed by apoptosis 1 to 2 hours later. Thus they concluded that this
region, corresponding to cytological chromosomal band 75C1,2 is required for
apoptosis in Drosophila.
The genes that have been identified in this region, @, reaper, and hid,
have no known mammalian homologs. The 65 amino acid sequence of REAPER,
however, does share homology to cytoplasmic regions of the Tumor Necrosis
Factor Receptor 1 and Fas Ligand receptors (discussed below) called death
domains. These regions are proposed to mediate protein-protein interactions
that mediate ce11 death signaling. Although REAPER does share the ability of
death domains to self-aggregate, the functional significance of this homology is
questionable. Mutations that cause loss-of-function in mammalian death
domains have no effect on REAPER-mediated activation of apoptosis (16).
Reaper plays an important role in initiating programmed cell death by
apoptosis. It is suggested that reaper serves to integrate a variety of signals
leading to apoptosis including ceU-ceU interactions, lineage signals, x-rays, and
inhibition of differentiation (15). As with the mammalian models, apoptosis by
reaper requires function of caspases. The Drosophila caspase, drICE, is activated
by proteolytic processing in response to apoptosis initiated by reaper over
expression and is expressed at stages where PCD c m be induced in Drosophila
(17). Thus, drICE is implicated as a caspase involved in apoptosis in Drosophila
and suggests that the classical apoptotic pathway identified in mammals and C.
e l e g ~ may exist in Drosophila.
5

In Drosophila where the H99 chromosomal region is deleted removing
grim, reaper, and hi#, low doses of X-rays in the range of 500 rads fail to activate
celi death. However, higher doses are still able to activate apoptotic ce11 death.
The response of embryos with deleted Hg9 regions to X-ray exposure may be
attributed to the presence or absence of reaper. With the loss of reaper, ce11 death
during embryogenesis and ce11 death induction by low doses of X-rays is
inhibited. High doses of X-rays are still able to initiate apoptosis (15). These
results suggest that these genes function somewhere upstream in the apoptotic
pathway as initiators. The loss of these genes alone still leaves the effector
mechanism(s) of apoptosis intact and able to carry out cellular suicide upon
ectopic activation.
Though pim, reaper, and hid have no known mammalian homologs, they
do, however, share between them a homologous amino-terminal 15 amino acid
sequence. This region is proposed to mediate inhibitory regulation of their
activity by interacting with BIR domains of Drosophiln Inhibitor of Apoptosis
(DIAPI) protein (18). Expression of DIAPI, a protein homologous to baculovirus
IAPs, in Drosophila is able to counteract the small-eye phenotype induced by the
expression of reaper in the eye via pGMRreaper (18).
1.1.4.1 Apoptosis dunng Dmsophila development
The vital dyes, acridine orange (AO) and nile blue (NB) are able to
selectively stah apoptotic cells in developing Drosophila embryos (19). The
stained ce& observed during Drosophila embryogenesis displayed the same
morphological features of apoptotic ce11 death-separation, nudear and
cytoplasrnic condensation, fragmentation, and phagocytosis-as described by
6

Kerr, WyUe, and Currie in 1972 (1). Cells displaying features of necrosis are not
stained by these dyes suggesting that molecular feahves specific to apoptosis are
responsible for the selective staining. Work by Delic and others suggests that it is
the intercalation of acridine orange with duplex DNA that is responsible for the
fluorescence observed in apoptotic cells (20). Using A 0 staining, Abrams
observed that normal programmed cell death begins at stage 11 (approximately 7
hours of development) of embryogenesis after which it is observed extensively
throughout the embryo (19). There is a reproducible pattern of cell death
obsewed, though not with the predictability of the ce11 deaths observed in C.
elega~ls.
The genetic mutations polyhomeotic and cnîmb cause degeneration of the
ventral epidermis and epithelial tissue respectively in the developing Drosophila
embryo. A 0 staining in these tissues accurately reflects the increase in apoptosis
induced by these mutations. Exposure to high doses of X-rays in the range of
4000 rads induces a rapid increase in AOpositive cells. Lower doses of X-
irradiation did not show the same rapid increase of cell death. A 0 staining only
revealed an increase in apoptosis when the staining was done 7 hours after
exposure. This research establishes the use of acridine orange as a tool for
apoptotic staining induced by various methods.
1.1.4.2 Apoptosis Linked to Segment Polarity Signaling
Drosophila development from embryo to larva depends extensively on
the h c t i o n of molecular signals to develop anterior-posterior and dorso-ventral
axes. Based on these boundaries, other molecular signals determine cell fates of

different regions of the embryo, dividing it hto segments that ultimately lead to
the various structures of the adult organism.
The segment polarity genes are a specific group of genes that function to
detennine the ce11 fates. Their ability to regulate cell death was studied using
tirne-lapse fluorescence microscopy to follow apoptosis throughout embryonic
development (21). Co-staining for segment polarity genes and AOpositive cells
demonstrates that the segment polarity genes provide survival signals for
specific rows of epidermal cells. Mutants of segment polarity genes result in
increased amounts of cell death in epidermal cells. Using a temperature-
sensitive mutant allele of wingless, it was shown that the loss of wingless
signaling during the fate specification phase of epidermal development results in
increased apoptosis in the anterior stripe of cells in each segment
border-ultirna tely giving rise to the wingless cu ticle phenotype. These results
suggest that wingless signaling mediates the survival of wingless-responsive cells
in the anterior half of the developing segment. Conversely nkd mutations show
increased ce11 death in the posterior, EN-expressing domain of the segment with
no increase of ce11 death in the anterior portion of the segment as with wg
mutations. This work establishes the link between signaling leading to fate
specification and apoptosis and underlines the importance of the regulated
process of ce11 death in the normal development of Drosophila and other living
organisms.

1.2 CASPASES-THE CORE OF THE APOPTOTIC EFEECTOR MACHINERY
1.2.1 Caspase Function and Activation
The understanding of apoptosis has developed largely as the key
enzyrnatic players in the mechanism of apoptosis became known. At the core of
the effector mechanism of apoptosis are the members of the caspases family of
proteolytic enzymes. The molecular basis of many hallmark apoptotic features
has been traced to the action of caspases. This makes the regdation of their
function and proteolytic targets of central importance to understanding PCD in
biological systems.
Caspases are proteolytic enzymes that require an aspartate amino acid
residue at the target cleavage site and are related to interleukin-1-kconverting
enzyme (ICE) which is involved in proteolytic processing of human Interleukin-1
(22). The active form of human ICE consists of pl0 (10 kDa) and C-terminal
pz0 (20 kDa) subunits which are processed from the p45 (45 kDa) cytosolic
precursor protein (23). ICE has autoprocessing ability (23) and exists as a
tetramer (plO/p20)2 which requires further oligomerization for its
autoprocessing ac tivity (24).
In vivo, caspases are proposed to act in a cascade of self-activation that
requires a critical number of initiator caspase protehs be assembled locally
before activation of downstream executioner caspases (5). This is the 'proximity
model' of caspase activation. This suggests that caspases, and thus the process of
apoptosis, may be activated by processes that cause the association of a
significant number of cytosolic precaspase pretursor molecules.

l.2.2 Caspase Targets
Once activated, the caspase proteolytic enzymes accomplish their
"executioner" function by cleaving target macromolecules. Among the sub-
cellular targets are the DNA assembly proteins leading to the appearance of
characteristic DNA ladder (25). One of the early events of apoptosis is the
poly(ADP-ribosy1)ation of nuclear proteins by the enzyme poly(ADP-ribose)
polymerase (PARP). This activity is required for the progression of apoptosis
and the obsewed morphological changes (26). PARP is a caspase-3 target and is
inactivated by cleavage during early apoptosis after its initial burst of activity
(27). The PITSLRE protein kinase, related to p35cdc2 kinase, is also a target of
caspase activity during apoptosis. In contrast to PARP, this protein is processed
and activated by caspase activity possibly leading to further apoptotic signaling
(28). Other targets for proteolytic breakdown include proteins involved in DNA
repair, signal transduction, cellular cytoskeleton, and control of the cell cycle
(29)-
1.2.3 Caspase Inhibitors
Cellular suicide is one of the tactics used by multiceilular organisms to
defend against viral infection. To counteract this, the baculovirus genome
contains a caspase-based anti-apoptosis defense mechanism that prevents the
death of the infected ce11 allowing the spread of the virus to uninfected cells. The .
baculovirus protein p35, produced during the early stages of infection, acts as a
cornpetitive inhibitor of ICE fonning a stable ICE-p35 complex inhibithg even
adivated caspase molecules (30). p35 acts in C. elegans (31,32), Drosophila (18,
33), as well as marnmalian systems to prevent caspaseinduced ceil death (34).
10

The bacdovirus Inhibitor of Apoptoôis (LW) genes provide another
mechanism to inhibit apoptosis. IAP genes block ce11 death by inhibiting
ICE/CED3 proteases. Unlike p35, however, IAPs are not able to inhibit
activated caspases. IAPs are able to inhibit caspase activation suggesting that
they work in a distinct mechanism from p35 to inhibit caspase-mediated
apoptosis (30). IAPs contain two functional domains; C-terminal RING-finger
motifs which mediate protein interaction and BIRs (Baculovirus Inhibitory
Repeats) found in the N-terminal region. It is the BIR domain that mediates
prevention of both normally occurring and ectopic ce11 deaths. CrmA, a cowpox
virus protein has also been demonstrated to inhibit caspase-induced ce11 death
(35).
1.3 SIGNALS LEADING TO APOPTOSIS
1.3.1 TNF Signaling-Receptor-linked Apoptosis
As in many regulated processes, the activation of the apoptotic
mechanism is regulated by both intemal and extemal molecular signals.
External signals capable of activating or inhibiting cellular apoptosis are
important mechanisms for organisms to regulate c d populations in various
tissues. In marnmals, the receptors TNF-RI and FasL are involved in
transmitting extracellular signals leading to the initiation of apoptosis.
Specifically, it is the cytoplasmic death domain, an 80-amino acid sequence of
homology that mediates the protein-protein interactions leading to apoptotic ce11
death (36,37).
The biological responses of TNF signahg in cells of the hematopoietic
system indude ceil death (apoptotic or necrotic) or proliferation and depend on
11

cell type, differentiation state, transfomation status, and the presence of other
stimuli (38,39). Among their many functions, TNF family members serve to
remove T and B cells after proliferation in response to infections and TNF-a is
required for normal elhination of potentially autoreactive peripheral T cells and
contributes to T cell-mediated cytotoxicity (40). TNF-related ligands and
receptors ensure response is initiated at correct tirnes, places and involves correct
ce11 types. To carry out these functions, TNF-related ligands and receptors are
expressed on activated macrophages and T cells. T ce11 responses are largely
dependent on antigen stimulation and subsequent cell-ce11 interaction (39).
The TNF ligand family includes the following members: TNF, LT-a, Fas
ligand (CD95L), OX40L, CD40L, CD27L, CD3OL, 4-lBBL, and LTP,
Apo3L/TWEAK, AP02L/TRAIL and RANICL (39). Two of the ligand family
members, TNF and Fas ligand (FasL) can function as soluble ligands while the
others are mostly membrane-bound. TNFa is released from its membrane forms
by metalloprotease proteolytic processing (41,42). These ligands are type II
membrane proteins with their C termini facing the extraceilular space.
Approximately 150 amino acids of the C terminal region, albeit with low (20025%
at the protein level) homology, defines the ligand family and is responsible for
TNF signahg activity.
The receptors are typically type 1 membrane proteins with the region of
homology residing in the ligand-binding regions. These regions contain
cysteine-rich domains (CRDs) which are defined by the presence of 6 cysteines
separated by stretches of 40 amino acids. The TNF receptor (TNFR) family
includes TNF-RI (p55), TNF-RII (p75), TNF-RLII (TNF-RP), 0x40, &lSB, CD40,

CDN, CD27, the poxvinis gene prducts PV-T2 and PV-A53R, and p75 NGFR,
CARI, DR3 (also known as AP03, WSL-1, TRAMP, LARD), DR4, and DR5 (also
known as AP02, TRAIT.,-R2, TFüCK 2 OR KZLLER) (39). The receptors are
usually trimeric or multimeric complexes when functional. htracyçteine
disulfide bonds formed between the CRDs of the subunits stabilize these
complexes. Most receptors also exist in a soluble form that is achieved by
prote01 ytic cleavage.
The basic mode1 of Fas and TNF signaling leading to apoptosis begins
with the engagement of the trimeric ligand to the receptor (figure 1A). In Fas
signaling, this brings the intracellular death domains of Fas together causing the
recruitment of the death domain in FADD/MORTl. Death dcinains are
approximately 80 amino acid motifs that are able to bind to each other thereby
mediating protein-protein interactions. The cytoplasmic portion of Fas also has
another protein interaction domain called the Death Effector Domain (DED).
This motif causes the association of caspase 8 via its own DED to the receptor
complex. The recruitment of caspase 8 to the receptor complex leads to the
activation of the caspase cascade. As mentioned above it is the action of these
proteolytic enzymes that is ultimately responsible for many of the molecular
features of apoptosis including the degradation of dvomosomal DNA. In this
manner, signaling by Fas and FasL leads to the activation of caspases and the
ordered breakdown of the cell.
TNF signaling leading to apoptosis proceeds in a similar fashion to FasL.
TNF signaling begins with the engagement of the trimeric ligand to the receptor.
In the cytoplasm, the oligomerized death domains of the receptor induce the
association of TRADD (TNFR-associated death domain prottin) (43) which
13

reCIUits FADD / MORT1 (Fas-associated death domain-containhg pro tein) (44)
again via death domain interactions. Similar to Fas signaling, the recruitment of
FADD leads to the activation of the caspase cascade and apoptosis. The
recruitment of TRADD also leads to the recmitrnent of TRAF proteins and the
kinase RIP, which also contains a death domain. The latter interactions lead to
the activation of NF-KB (45,46).
1.3.2 NF-KB signaling in apoptosis
Nuclear factor K B (NF-KB) is a heterodimer transcription factor made up
of 65 kD and 50 kD protein subunits. The protein binding and DNA recognition
sequences of this transcription factor reside in an N-terminus region of
homology. Subcellular localization studies show NF-KB resides in the cellular
cytoplasm but, in response to extracellular signals, translocates to the nucleus
where it mediates the transcription of genes by binding to specific DNA
sequences.
The activation of NF-KB by TNF is a suwival signal that hc t ions to
antagonize TNF-induced apoptosis (47). TNF signals result in the
phosphorylation of the inhibitory protein, 1-ICB leading to itsdegradation by the
26Sproteosorne pathway. This releases the inhibition of NF-KB and allows this
transcription factor to activate its target genes by translocation to the nucleus.
1.33 Activation of Apoptosis by W and Ionizing Radiation
Another method of activating cell death in living organisms is exposure to
various environmental agents. These can be diemical or in the form of radiation.

Of these, ionizing radiation in the fom of x-rays andior ultraviolet radiation is
of particular significance to humans. Human exposure to x-rays most often
cornes in the form of diagnostic x-ray machines and to ultraviolet radiation from
exposure to the sun.
Current models of the UV response in mammals, shown in figure lB,
suggest either a DNA damage-dependent (48) or DNA damage-independent
mechanism (49). DNA damage-independent mechanisms may involve the action
of receptor tyrosine kinases (RTKs) (50). In this case, UV stimulation leads to the
activation of RTKs and consequent phosphorylation-mediated signaling events.
In one mode1 of UV-induced activation of RTKs, UV energy induces the
formation of disulfide bonds between receptor molecules, mimicking ligand-
induced oligomerization (51,52). In mammals the JNK/SAPK pathway is
responsible for integrating environmental stress into a response of adaptation
(growth arrest or immune response) or initiation of apoptotic ce11 death (53).
Thus, the Drosophila ortholog of the JNK/SAPK pathway (54) may be a potential
area of research for studying UVC (peak emission 254nm) responses in
Drosophila. During development, the Drosophila pathway plays a role in dorsal
closure-the morphogenetic movement of lateral epithelial cells to the midline
and the covering of the developing Drosophila embryo (54).
h a DNA damagedependent mechanism apoptosis would be activated
via a DNA monitor such as mammalian p53 in response to UV-induced DNA
damage. p53 is a widely studied protein involved in mammalian responses to
stress stimuli causing DNA damage (review in (55)). The loss or mutation of p53
in a large number of human cancers highlights the role of p53 in inducing cell
cycle arrest or initiating apoptosis in response to DNA damage. Tumor cells that
15

have lost the function of p53 exhibit a selective growth advantage since the
negative growth regulation conferred by p53 is also lost. p53 protein is stabilized
in response to UV stimulation (56) and has the abiiity to initiate apoptosis when
DNA damage cannot be repaired. Pro-apoptotic genes that are transcriptionally
activated by p53 include Bax (57) and Fas (58). Recently, a Drosophila p53
homologue has been identified and implicated in the activation of radiation-
induced apoptosis (59,60). Work by Nordstrom and Abrams demonstrates that
reaper is a transcription target of Drosophila p53 (60) and establishes a critical
link in the observations of White and Abrams (15) where reaper is required for
the activation of radiation-induced apoptosis.
1.4 Statement of objectives
In this research we examined two forms of stress stimuli and their effect of
activating programmed ce11 death in Drosophila. The first is a molecular signal,
TNF-a. nie mouse version of the signaling portion of TNF was introduced into
the Drosophila genome using P element-mediated transformation creating a line
that is able to express the TNF-a polypeptide in a controlied fashion. The
objective was to detennine if this type of signaling exists in Drosophila and leads
to apoptotic celi death. The second stimulus used is ultraviolet radiation (UV) to
which Drosophila embryos or lawae were exposed. Research to date on the effect
of UV on Drosophila has focused on overall lethality (61) and initiation of DNA
repair mechanisms (62,63). UV activation of apoptosis has not been described in
Drosophila. Other forms of radiation such as X-rays, however, have been

obsenred to activate apoptosis in this organism (15). Our objective is to
determine the ability of W radiation to activate apoptotic ce11 death and attempt
to determine specific genes that are transcriptionally activated during the
process.
%me of the core elements of the known apoptotic pathway, such as
cytoplasmic initiators (reaper (IS)), inhibitors ( D M (18)), effectors (caspases (5,
17)), and targets (PARP (64)) have already been identified in Drosophila.
Overall, we would like to establish a system where a known mammalian initiator
of ce11 death, specifically TNF-a and/or U V , is able to activate apoptotic ce11
death in Drosophila. Once established, this would create the basis for a genetic
screen of random P elements in search of novel genes involved in the process.
This would be accomplished by taking advantage of the available P element
library in Drosophila that represents a significant proportion of the genes that are
critical for Drosophila development and sumival (65).

2.1 MOLECULAR BIOLOGY
2.1.1 LB MEDIUM (LURIA-BERTANI MEDIUM)
LB medium was made up of 10 g bacto-tryptone, 5 g bacto-yeast extract, and 10 g
NaCl per litre of distilled water. The solutes were dissolved and the solution
autoclaved. When required, ampicillin was added just before use to a final
concentration of 50 pg/mL from a 50 mg/rnL stock.
2.1.2 LB-ampicillin plates
For LB-ampicillin (LBamp) plates, 15 g of bacto-agar is added per litre of LB
medium prior to autoclaving. After autoclaving, the solution was stirred as it
was allowed to cool. When the solution was only warm to the touch, ampicillin
was added to a final concentration of 50 pg/mL from a 50 mg/mL stock. The
LBamp medium was then poured into 100 mm X 15 mm disposable sterilized
plates (Falcon). The medium was allowed to solidify at room temperature
overnight and the plates then stored at 4 OC.
2.1.3 Competent cells
A single colony of XL-1 Blue E. coli bacteria was used to inoculate 50 mL LB
medium and incubated with shaking at 37 O C overnight. 2 mL of the ovemight
culture was used to inoculate 200 mL of pre-warmed LB and incubated in a 37 "C
shaker until an absorbance (A) at 600 nm of about 0.35-0.4 was reached. The
culture was split into 4 X 50 mL aliquots in pre-chilled Falcon tubec and left on
ice for 10 minutes. The 50 mL cultures were then centrifuged at 2700 rpm at 4 "C
18

(1 600 X g) for 7 minutes Afkr removing the supematant the pellets were gently
resuspended in 10 mL ice cold CaCI, solution made up of 60 mM CaCl,, 15%
glycerol, 10 mM Pipes pH 7.0 (Sigma) and left on ice for 30 minutes. The cells
were once again centrifuged for 5 minutes at 2 300 rpm at 4 OC. The supematants
were removed and the pellets resuspended in 2 mL ice cold CaCI, solution.
Finally, the cells were distributed in 100 PL aliquots into sterile pre-chilled
microfuge tubes, frozen on dry ice, and stored at -70 OC.
2.1.4 Restriction enzyme digests
Restriction enzyme digests were typically carried out in 20 pl total volume using
1 pl of enzyme (New England Biolabs), the recommended buffer, and optimal
temperature for approximately 2 hours. For digests where the DNA would not
cut, spermidine was added to 4 rnM final concentration.
2.1.5 Agarose gel electrophoresis
DNA samples were prepared by combining 1 pl agarose-gel loading dye (0.25%
bromophenol blue, 0.25 % xylene cyan01 FF, 15% Ficoll in H,O), restriction
enzyme digest, and water to a total volume of 10 pl. DNA was resolvd by
electrophoresis on 1% agarose gels in TAE (0.04 M Tris-acetate, 0.001 M EDTA
p H 8.0) using the 1 kB (Gibco) marker for size comparison. The bands of DNA
were visualized by including ethidium bromide in the gel and using an
ultraviolet light-box.

2.l.6 Isolathg and purifyine DNA fragments
DNA bands were isolated using the GenecleanB DNA isolation kit according to
the manufacturer's protocol. DNA was eluted into 20 pl of distilled water.
2.1.7 Ligation
Ligations of DNA fragments were done in 10 pl volumes using T4 DNA ligase
(New England Biolabs) and the supplied buffer. Approxirnately 5:1 molar ratios
of insert to vector DNA were used to optirnize ligation effeciency. A vector-
alone control was set up in parauel with the vector-plus-insert ligation.
2J.8 Transformation
1 / IOh volume of the ligation reaction was added to 100 111 of competent cells
freshly thawed on ice. After a 30 minute incubation on ice, the cells were heat
shocked for 90 seconds at 42 OC. 400 pl of LB were added and the cells were then
incubated at 37 O C for 45 minutes. 200 pl of the transformation mixture was
plated on LBamp plates which were then incubated at 37 O C overnight. Colonies
were isolated from the vector-plus-insert ligation/ transformation when the
number of colonies outnumbered the number of colonies from the vector-alone
ligation/transformation by at least two thes . A minimum of 10 unique colonies
were used to inoculate 5 ml of Lbamp liquid medium each for an overnight
incubation in a 37 OC shaker.

U 9 Small-scaie plasmid prepaxations
1.5 ml of the 5 ml overnight incubation was transferred to a microfuge tube while
the rest of the culture was stored at 4 O C . After centrifugation in a microfuge at
14 000 g for one minute, the supematant was removed and the pellet
resuspended by vortexing in 100 pl Solution 1 (50 mM glucose, 25 rnM Tris-HCl
pH 8.0,10 mM EDTA p H 8.0). 200 ~1 of freshly prepared Solution II (0.2 N
NaOH, 1% S E ) was added to the microfuge tubes which were gently inverted
repeatedly to ensure complete mixing. 150 pl of Solution III (3 M potassium
acetate, 11.5% v/v glacial acetic acid) was added and mixed by inverting and
gently shaking the microfuge tube. The tubes were then centrifuged for one
minute at 14 OOOg and 400 pl of the resulting supematant removed to a new
microfuge tube. The solution was subjected to one phenol/diloroform extraction
and the DNA precipitated for up to 1 hour at room temperature with 2 volumes
of 95% ethanol and sodium acetate, pH 5.2, to a final concentration of 300 mM.
The DNA was pelleted by centrifugation at 14 OOOg, washed with 70% ethanol,
and resuspended in 50 pl distilled water. Typically 5 pl of plasmid DNA was
used in diagnostic restriction enzyme digests which included 1 pl Rnase
(Boehringer-Mannheim). Ovemight cultures which gave plasmici DNA
preparations with the correct insert and orientation were used to inoculate 100
mL LBamp media for large-scale plasmid preparations.
2.1.10 Large-scaie plasmid preparations
The 100 mL ovemight cultures were divided into hvo 50 mL Falcon tubes and
centrifuged at 3 500 rpm. Large-scale plasmid DNA preparations were made

using the QIAC=EN@ Maxi kit and prabcol, After DNA precipitation with
isopropanol and centrifugation the pellet was resuspended in 400 pL distilled
water and treated with 2 pL Rnase at 37 OC for 1 hour. One phenol/chloroform
extraction followed and then the DNA was precipitated by adding 80 PL 1.5 M
sodium acetate, pH 5.2 and 800 PL 95% ethanol and incubating at room
temperature for 5 minutes. A 5 minute centrifugation at maximum rpm served
to pellet the DNA which was washed with 70% ethanol and resuspended in 75
PL distilled water. An A 260 nm reading of a 1:100 dilution was taken to
determine the amount and concentration of the purified plasmid DNA.
2.1.11 Subcloning and confimation of hs mTNF
The mTNF insert was removed from pBluescript (Wen Chen, Amgen Research
Institute) with the NotI and EcoRV restriction enzymes (New England Biolabs)
and ligated into pCaSpeR-hs via the NotI and StuI sites. After transfomihg XL1-
Blue competent cells, minipreps were made from 10 of the resulting colonies on
the vector-plus-insert plate. Miniprep DNA was checked with a NotI/BamHI
double digest to confirm the presence of the mTNF insert in the pCaSpeR-hs
vedor. A largescale DNA preparation was made from the miniprep which
showed the approximately 700 bp band after agarose gel electrophoresis.
2.1.12 Embryo protein lysates
Dtosophila embryos were collected, dechorionated, frozen in an ethanol/dry ice
bath and stored at -70 O C . To prepare the samples, between 200 and 300 PL
Gentle Soft Buffer (10 mM NaCl, 20 m M Pipes pH 7.0.0.596 IGEPAL CA-630

(Sigma) ,0.05Ye Rmercaptoethanol, 5 mM EDTA, 50 mM NaF, 5 pg/mL
leupeptin, 1 rnM benzamidine, 100 pM NaVO,) was added to the frozen embryos
which were pulverized with a pestle. This was followed by repeated pipetting
with a 21g needle and 3 mL syringe and boiling for 10 minutes. The sample was
then centrifuged for 5 minutes in a microfuge (Ependorf) at maximum rpm and
the supernatant transferred to a new microfuge tube. Protein concentration and
amount was determined by use of the Bradford assay (Biorad). Before loading
ont0 SDSpolyacrylamide gels, an equal volume of 2X sample loading buffer
(0.125 M Tris-Cl pH 6.8,4% SDS, 20% glycerol, 10% B-mercaptoethanol, 0.01%
bromophenol blue) was added and the samples boiled once again for 5 minutes.
2.1.13 SDS-PAGE
Protein samples were resolved on 12.5% SDSpolyacrylamide gels according to
standard techniques(&) using the Bio-Rad@ mini-gel resolving and transfer
apparatus. New England Biolabs protein molecular weight marker was included
for size cornparison. Resolved proteins were transferred to polyvinylidine
fluoride filters and incubated ovemight in Bovine Lacto Transfer Technique
Optimizer (BLOTTOg. 4% skim milk,lX PB, 0.05% tween-20). The membrane
was then probed with the anti mouse TNF polyclonal antibody (Genzyme) at a
1:1000 dilution in BLOTT0:PBS (2% rnilk, 1X Phosphate Buffered Saline (PBS),
0.025% tween-20) for 3-4 hours at room temperature. The membrane was
washed in BL0O:PBS 6 times for approximately 3-5 minutes each wash
before incubation with the HRP-conjugated anti rabbit IgG secondary antibody
(Amersham). û-10 other washes of 2-5 minutes preceded the development of the

HRP reaction. Once developed according ta the manufactuw"~ protocol, the
membrane was wrapped in Saran wrap and exposed to film (Kodak X-OmatTM
Blue ml). The film was then developed under dark room conditions.
2.2 DROSOPHILA HANDLING TECHNIQUES
2.2.1 Drosophila culture
Drosophiln culhws (table 2) were raised on media containing 21.74 g yeast, 65.22
g commeal, 12.83 g sugar, 9.35 g agar, 21.73 mL molases, 13.06 rnL tegosept
solution, 2 mL propionic acid, and 0.1 g ampicillin per litre. Cultures were
maintained in either vials (Applied Scientific) or botties (Applied Scientific) at
room temperature (19-22 O C ) or 25 OC.
2.2.2 Apple juice plates
Apple juice plates (3.4% agar, 50% apple juice) were made by adding 34 grams of
agar to 500 mL distilled water. This was brought to a boil with stirring. 500 mL
of apple juice was then added slowly, brought to a boil, and then allowed to boil
for 5 minutes. The solution was then removed from the hot plate and allowed to
cool to 80 OC. 7 mL of 10% tegosept was then added before filtering through
cheesecloth. The solution was then poured into 60 mm X 15 mm disposable
sterilized plates (Falcon).
2.2.3 Yeast
Yeast paste was made by combining dried baker's yeast (SIGMA) with distilled
water and mixing to desired consistency.

2.2.4 Embryo collections
Drosophila embryos were collected on yeasted apple juice/agar plates for
specified periods of tirne at 25 O C . The embryos were then swept into collection
baskets made from the top half of 15 mL Falcon tubes with a hole created in the
cap. A small square of nylon mesh was used to capture the embryos while
allowing liquid to drain away. After the embryos were collected the chorion was
removed by holding the collection basket in 50% bleach for 1 minute. The
embryos were then rinsed copiously with water.
2.2.5 Embryo fixation
After collection, embryos were transferred to a solution containing 680 pl of 37%
fomaldehyde in 4.3 ml 1 X PBS layered with 5 ml heptane in a glass scintillation
vial. Embryos were held in this solution for 20 minutes and then transferred to
microfuge tubes with 500 pl methanol and 500 pl heptane. The embryos were
then briefly vortexed and the heptane removed. The embryos were washed
approximately 6 times in methanol and then stored at -20 O C .
2.2.6 Heat shock
Drosophila embryos were held for 8 minutes in a 37 "C water bath after collection
as described above (2.2.4) without dechorionation. The embryos were then
rehuned to the apple juice/agar plates on the wire mesh and aged for 1 hour.
The nylon mesh containing the embryos was then returned to the collection
basket, then dechorionated and fked as described above.

2.2.7 WB exposure of Drosophila imaginal discs and staining for
gdactosidase ac tivity
First-to-second instar Drosophila larvae were exposed to an exposure 90J/m2
from a UVB (270-330 nm) source (Hill Lab, OCI), then held in dark conditions on
yeasted apple juice plates for 24 hours. Using forceps, the lamal heads were
then isolated and inverted in ice cold PBS (1401nM NaCl, 25mM sodium
phosphate buffer, pH 7.2) exposing the imagina1 discs. The inverted heads were
h e d for lOmin at room temperature with freshly prepared 0.75% glutaraldehyde
in PBS, washed 3 times with PBT (0.05% TritonX-100 in PBS), then stained in
0.2% X-gal in staining solution (10mM sodium phosphate, pH7.2150mM NaCl,
1m.M MgCl,, 5mM potassium femcyanide, 5mM potassium ferrocyanide),
freshly prepared from an 8% X-gal stock solution in DMSO kept at -20 OC (67).
Staining was done at 37 OC in the dark for 6 to 12 hours and then was terminated
with 3 washes in PBT. The tissues were then allowed to sit in mountant (70%
glycerol buffered with 0.03M Tris, pH9.0) overnight at 4 OC. The imaginal discs
were isolated in mountant and placed on a microscope slide then flattened with a
cover slip before being observed with a compound microscope.
2.2.8 UVC Treatment
Drosophila embryos were coliected for specified amounts of tirne on apple
juice/agar plates and then given a 30 second W C exposure (254 n m peak
emission) in a Stratagene Stratalinker.

2*2*9 Cuticule preparations
Embryos were collected and treated as indicated and then plated individually in
groups of 100 on apple juice/agar plates. The plates were then kept at room
temperature or 25 O C for 36 hours. The unhatched brown and white embryos
were counted to calculate the lethality of the treatment
(lethality=#brown/ (#plated - #white)). The embryos were then transferred to
nylon mesh and placed in collection baskets for dechorionation as described
above. The embryos were then placed in Hoyer's solution on a microscope slide,
the coverslip put in place, and the slide placed on a 60 O C slide warmer overnight
for the embryos to clear, revealing the cuticle.
2.2.10 Injection mlx
In a 1.5 mL microfuge tube, 10 pg of pCaSpeR-hs mTNF plasmid DNA was
combined with 2 pg phsn helper plasmid DNA and distilled water was added to
a final volume of 400 PL. Two phenol/chloroform extractions were then
performed. The DNA was then precipitated with 800 PL 95% ethanol and 80 PL
1.5 M sodium acetate pH 5.2 (300 mM final concentration) for 1 hour at room
temperature or overnight at -20 OC. The DNA pellet was obtained by
centrifugation for 10 minutes at 14 OOOg in a miaofuge. The supernatant was
then removed. Following a wash with 70% ethanol, the DNA was resuspended
in injection buffer (5mM KCl, 0.1 mM NaPO, pH 6.8) and 0.5 pl green food dye
added for visualization in embryos.

2.2.11 Germline Transformation
ûrosophila embryos of the line w ' ~ ' ~ were collected for one hour, dechorionated
and transferred to a pad of agarose containing green food dye. The embryos
were lined up in groups of 20-30 with their posterior ends at the edge of the pad.
The row of embryos was then hansferred to a thin strip of double-sided tape on a
microscope slide. The embryos on the slide were then desiccated in a large petri
dish containing desiccant. Desiccation times were empirically determined at
each injection session. After desiccation the embryos were covered with a layer
of heavy (Series 700) Halocarbon@ oil. With the microscope slide on the micro-
injection apparatus, small amounts of the injection mix were deposited into the
posterior ends of the developing embryos in the area of the developing pole cells.
After injection, the embryos were covered with light (Series 56) Halocarbon@ oil
held in place by a petroleum jelly moat. The slides were placed in a humidifying
chamber made of a large petri dish and moist filter paper and the embryos
allowed to develop for approximately 36 hours at room temperature. At this
t h e , lawae that have survived the injection are placed in vials and allowed to
develop to adulthood. The surviving adults were crossed to w1'18 fies. Progeny
with red (could be pale yellow, dark yellow, light orange, or dark orange) eyes
represent successful germline pslement transformants. These were crossed to
wl"* to propagate the transformant line.
2.2.12 Mapping and Balancing
P[w+l males were crossed to wll18;L/CyO;Kiftnu20/TM3 virgin females and
male w;P[w+]/CyO;TM3 progeny were isolated. The observation that red-eyed
males were isolated indicated that the insert was not likely on the X
28

chromosome. To balance the iine, these males were aossed to sibling vkgin
females of the same genotype. To determine the location of the insert on either
the second or third chromosome these males were crossed to w~''~;L/C~O;K~
~ m 2 0 / T M 3 virgin females. Second chromosome inserts do not show
P[w+],L/CyO phenotypes while third chromosome inserts do not show
Plw+],Ki/TM3 phenotypes resulting from this cross. n, results of the p[w+~
insert crossed with w;L/CyO;Ki/TM3 indicate that the pCaSpeR-hsTNF P element
had inserted into the third chromosome. As w;L/CyO;w+/ïM3 siblings were
crossed, the loss of the stubble phenotype indicated that the w+ third
chromosome did not require the TM3 balancer to maintain the viability of the
line. Flies that had lost the balancer chromosome (w;L/CyO,w+/b+) also had
distinctly red eyes whereas flies with the balancer had orange eyes
(w;L/CyO,w+/TM3). The red-eyed aies correspond to a double dose of the w+
plasmid. Therefore, the insertion of the transforming plasmid into the third
chromosome did not disrupt the function of essential genes and is cowidered
non-lethal,
2.2.13 Mobilization to the Second Chromosome
Five vials of ~ ~ ~ ~ ~ ; / C y 0 C S p e R - s mTNF virgin females crossed to wu'*; r y Sb e
P M 3 ; r y]99B/TM6 Ubx e males were established. From this cross, mottled-eyed
(orange/red and white), stubble males were isolated representing the genotype
p h CaSpeR rnTNF/r y Sb e P[d 2-3;r yJ99B at their third chromosomes. Each male
was then crossed to w"'~ virgin females. From this cross, Sb' (long brides) males
lacking the A2-3 chromosome and having one pCaSpeR-hs mTNF third

chromosome were isolated. Flies from this cross with darker red eyes should
have another copy of the phs CaSpeR mTNF insert on either the same (ID) or
another chromosome. At least 45 males with dark red and 10 males with orange
eyes were isolated and crossed individually to W ~ ~ ~ * ; L / C ~ O ; K ~ Z ~ ~ ~ M ~ virgin
females to determine the position of the potentially new inserts. From this cross,
W1118, , pCaSpeR-hs rnTNF/CyO;TM3 males were picked and then crossed to
W~~'~;UC~O;K~~~"ZO/TM~ virgin females to establish a stable line of genotype
~sTNF; Kiflzw2'/TM3* The rnobilization protocol resulted in approximately 60
iines with potential P element mobilizations. To test for the position of the insert,
males from each of the 60 Iines were crossed to w;L/CyO;Ki/7'M3 virgin females. 4
out of the 60 su& crosses gave results indicating the presence of the insert on the
second but not on the third chromosome. Matings of sibling w;w+/CyO;Ki/TM3
flies were used to establish the line designated hsTNF (II). In subsequent
generations of sibling crosses, the second chromosome balancer was lost
resulting in a distindly red-eyed w,w+;Ki/TM3 line. This suggests that the
mobilization of the insert to the second chromosome, as with the original
insertion to the third, did not disrupt the function of any essential genes and is
there fore non-lethal.
2.2.14 Combination with reaperlacz to make hsm~'eaper1acz
Virgin ~ 1 1 1 8 ; pCaSpeR-hs mTNF; Ki ftzw2O/TM3 females were crossed with
wl 1 lB;L/CyO~enperlacz males. Virgin femaies and sibling brothers of the
genotype w1118; pCaSpeR-hs rnTNF/CyO;reaperlazz/TM3 resulting from this

cross were then mated. Virgin female and male progeny which lacked both
balancer chromosome phenotypes representing the genotype ~ 1 1 1 8 ; pCaSpeR-
hsmTNF;teaperlacZ were crossed to establish the line.
2.2.15 Marking Hg9 with yellow+
y w prdGALQ;Ly/TM3,y+ e Ser virgin fernales were mated to Df (3L)H99,kni [ri-
l]p[p] /TM3,Sb[l] males. y w prdGAL4;H99 /TM3,y+ e Ser male progeny were
then crossed to y w prdGAL4;Ly/TM3,y+ e Ser virgin fernales. From this cross,
the y w prdGAL4;H99/TM3,y+ e Ser virgins were crossed to their male siblings
of the same genotype to establish a line of genotype y w prdGAL4;H99/TMEty+
e Ser.
2.2.16 Anti kgalactosidase Antibody Staining
Fixed embryos were transferred to a 0.5 mL microfuge tube, the methanol
removed, and then 250 pl of methanol and 250 p1 of TBST (150 mM NaCl, 50 m M
Tris pH 7.6,0.1% Triton X-100) added. After inverting the microfuge tube to mix,
25û p1 of the solution was removed and another 250 pl TBST added and mixeci.
Ail of the TBST/methanol was then removed and the embryos were washed 3X
with TBST. The embryos were incubated at rmm temperature on a rotisserie for
1 hour and then washed again for 3X with TBST. The embryos were incubated
overnight at 4 O C with the mouse anti galad do sida se antibody at 1:5000
dilution in TBST. The embryos were then rinsed 2-3 ümes with TBST, washed
for 1 hour at room temperature on a rotissene, and rinsed another 2-3 t h e s with
TBST before the addition of the biotinylated anti-mouse IgG secondary antibody.

This incubation was carried out for 2 hours at room temperature on a rotisserie at
1:300 dilution in TBST. Approxirnately 5X 15 minute washes were then done at
room temperature on a rotisserie before incubation with streptavidin-alkaline
phosphatase at 1:300 dilution for 45 minutes. The embryos were then washed as
after the secondary incubation. The alkaline phosphatase reaction was
developed as described below .
2.2.17 Alkaline Phosphatase Reaction Developing
After washing, the ernbryos were washed 3X with alkaline phosphatase (AP)
staining buffer (100 m M NaCl, 50 mM MgCl,, 100 mM Tris-HCl pH 9.5,0,1%
Triton X-100). 400 pl of AP stauiing buffer was added and then 1.8 pl nitroblue
tetrazolium (NBT), and 1.4 pl bromo-chloro-indoyl phosphate (BCIP). Sample
embryos were removed to gauge the progression of the reaction under a
microscope. Once the reaction was developed, the embryos were washed 8-10
tirnes with PBTR and covered with 100 fl of glycerol montant (70% glycerol,
30% TnsCl pH 7.5).
2.2.18 Acridine orange staining (Stellar lab protocol)
Drosophila embryos were dechorionated in 50% bleach and rinsed thorouglùy
with distilled water. The embryos were then transferred to a scintillation via1
containing 5 ml of 5pg/ml acridine orange in 0.1M phosphate buffer, p H 7.2
(made by rnixing 3.6 ml 0.1M Na,HPO, and 1.4 ml NaH2P04) layered with 5 ml
heptane. The embryos were shaken for 3 minutes by hand. The embryos were
then transferred to 1.5 ml rnicrohge tubes. After removing both upper and

lower phases, hesh heptane was added. The embryos were then transferred to a
slide and the heptane allowed to evaporate. Halocarbon oil was used to cover
the embryos which were then covered by a cover slip. The embryos were then
viewed and photographed within 30 minutes using the rhodamine filter.
2.2.19 Photography
A Leica DMLB microscope was used for photography of antibodystained
Drosophila embryos. For black and white photography, Kodak TMAX 100 ASA
black and white film was used along with a red (25A) filter to improve contrast.
Kodak Ektachrome 64T colour reversa1 slide film was used for colour
photography.

3.1 P element-mediated Transformation of wl"' by pCaSpeR-hsTNF
The study of mammalian signaling pathways such as Ras and Wnt/wg
has benefited from the study of their homologous pathways in Drosophila. Two
important advantages to being able to study signaling pathways in Drosophiln are
that: i) the functional order of pathway components can be determined by
genetic epistasis studies and; ii) new members of the pathway cm be identified
by conducting screens for Drosophila mutants that genetically interact with a
given gene or pathway (68,69).
To date, Drosophila homologues of TNF-family ligands or receptors have
not been identified. Thus, the TNF signaling pathway has not benefited from the
advantages of being shidied in Drosophila. We wanted to determine whether
TNFa ligand has any signaling activity in Drosophila. If so, the TNF signaling
pathway could be genetically dissected in this organism. The currently known
Drosophila regulators and effectors of celi death could then be placed in this
larger signaling context. hhacellular activators of apoptotic ce11 death namely
grim, reaper, and hid are already known in Drosophila. There is, however, no
known extracellular system of ligands and receptors in Drosophila that functions
to activate apop totic ce11 death.
One of the key players in the current mode1 of apoptotic cell death in
Drosophila is the gene reaper. The 65 amino acid protein product of the reaper
gene has limited homology to the death domains of the TNF and Fas receptors.
The functional significance of this homology, however, has been brought into

question (16,70). Mutations of certain TNF receptor and Fas death domain
amino aads cause these motifs to lose their protein-binding ability. This prevents
the subsequent activation of the downstream apoptotic pathway. Mutations of
the equivalent amino acids in reuper, however, do not diminish reaper's ability to
activate cell death as with the death domains of TNFRI and Fas. In spite of
reaper's limited and functionally questionable homology, it does share the
marnmalian death domain's ability to self-aggregate (71). This suggests that a
Drosophila prototype of the ce11 death signaling mechanism found in higher
mammals, including ligands and receptors, may exist.
To begin to address the issue of TNFa signaling in Drosophila P element-
mediated germline transformation was used to introduce the C-terminal
signaling portion of TNFa into the Drosophila genome. The C-terminal region
used here mimics the normal processirtg of TNFix that occua to create the active
signaling protein. The P element vector, pCaSpeR-hs (Genebank AC U59056),
permits genes to be introduced into the Drosophila genome (72) and selectively
expressed under the control of the hsp70 promoter by inueasing the incubation
temperature to 37 OC. Germline transformation is facilitated by injecting the
vector, dong with the helper plasmid, phsz (73). into the pole cells of w""
embryos approxirnately one h o u into development. This permits the
incorporation of the injeded DNA into the chromosomes of developing pole cells
and into the Drosophila gemiline.
Figure 28 shows the predicted expressed amino acid sequence of the sub-
cloned mouse TNFa vs. the M-length protein. The full-length protein contains
N-terminal sequences that anchor the translated protein to the cell membrane.

Post-translational processing by proteolysis results in the release of the soluble C
terminal signalhg portion of the TNFa protein (74). In the mouse TNFa clone
used here, the transmembrane and cleavage sites are omitted. The result is that
only the signaling-capable, soluble C-terminal region of the protein is expressed.
The presence of the plasmid in the Drosophila germline was confirmed by
observing the red eye phenotype in adult flies conferred by the white gene of the
pCaSpeR-hs plasmid. Subsequent crosses to wlll* and then siblings of the
resulting generations were used to establish the transformant line. Mapping
studies showed that the initial P element insertion occurred as a non-lethal
insertion on the third chromosome.
3.2 Heat-Shock Expression of mTNF
To veriQ that the mouse TNFa protein is translated, hsTNF embryos were
collected and subjected to an 8-minute heat shock at 37 O C . Protein extracts were
made and analyzed for the presence of the TNF protein. The use of an anti-
mouse TNFa antibody (Genzyme) revealed the presence of an approximately 20
kDa band after heat shock (Figure 3.) which is not present in w"'~ or untreated
hsTNF embryos. The predicted molecular weight of the soluble TNFa deavage
product is approximately 17 kD (74). The TNFa insert used here contains one
potential N-linked glycosylation site with the sequence Asn-Ser-Ser (Figure 28,
underlined). An apparent size shift of 3 kD is within the anticipated range of one
N-glycosylation modification (75). Another study with FasL expression in Cos
cells also showed a size discrepancy from the predicted amino acid sequence.
M e n expressed as a soluble protein in Cos ceus, FasL appears as 25 and 23 kD

bands when the predicted size is only 17 kD. The size discrepancy in this case
has been accounted for as glycosylation of the FasL protein (76).
Expression of the 20 kD TNF band is observed 30 minutes and 120
minutes after heat shock. Thus, when embryos of the hsTNF transformant line
are subjected to a heat shock, a protein in the predicted size range of the TNFa C-
terminal region is expressed. This protein reacts irnmunologically with an anti-
mouse TNFa polyclonal antibody.
3.3 hsreaper, hsTNF and UV Induction of Ce11 Death
Reaper is established as being able to activate cell death in Drosophila. in
mammals, TNF expression and exposure to UV radiation also have the ability to
activate ce11 death. In the next area of study, we attempt to establish a
comparable system of TNF and/or W activation of ce11 death in Drosophila
using the activity of heat shock induced reaper as the benchmark. The factors
used to initially characterize the effect of these three factors, were embryonic
lethality and cuticle phenotypes (Figures 4 and 5). The use of embryonic lethality
allows for a statistical cornparison of the ability of hsreaper, hsTNF, and UV
exposure to kill embryos while examination of resulting cuticle phenotypes
pennits the qualitative analysis of the effect of these treatments on the
developing embryos. To calculate the lethality, 200 treated embryos are placed
on a grid and given the opportunity to develop to lawal stages. The embryos
that failed to develop remained in their place on the grid and were counted.
The average lethality induced in h s ~ ; r p r l a c z embryos after heat shodc
was essentially identical to that induced in heat shock treated hsreaper embryos

(Figure 4) at 14% (0.14) compared to 13% (0.13) for hsreaper. The lethality
observed in untreated wl'" embryos is 5% (0.05). Thus, expression of the TNFa
protein in Drosophila is able to cause a similar lethality to expression of the
proapoptotic Drosophila gene, reaper. Using the same method, it was determined
that w"" embryos subjected to UVC treatment suffered much greater lethality
than both untreated w"'~ and heat shock treated hsreaper and hsTNF.
Calculations averaged 93% (0.93) for the treatment used here.
Next, the cuticle phenotypes resulting from expression of reaper or TNF, or
exposure to UVC were compared (Figure 5). Drosophila larval cuticle is secreted
by the epidermis of the developing larvae with a pattern of features that includes
segment boundaries, hairs, denticles and sensory organs. The epidermis in hm
is derived from the ectoderm of the developing embryo. Molecular signaling
events determine specific anterior/posterior and dorsal/ventral patterns of the
developing embryo including, the cuticle. Mutations in pattern formation
manifest themselves as changes in the normal development of cuticle. For this
reason, the lama1 cuticle has been used as a target for phenotypic screens for
mutations that affect pattern formation, establishing the relationship between the
cuticle phenotype and the signaling processes leading to its development (77).
The establishment of the cuticle phenotype as a method of determining the
qualitative effects of genetic changes in Drosophila allows for the use of this
method to determine the effect of hsreaper, hsTNF, and UV exposure on
Drosophila embryo development in this study.
Mutations of genes in the Hg9 genetic region, grim, reaper and hid resulting
in the loss of normal cell death activation, results in extra cells in the embryo (15,
78,79,80). Therefore it is reasonable to exped that ectopic activation of cell
38

death in Drosophila embryos would result in a cuticle phenotype reflecting a
reduction of ceIl numbers. Figure 5 shows that reaper and TNFa expression and
W C exposure are al1 capable of inducing similar cuticle phenotypes in
developing embryos. In al1 cases, the embryos were treated after 0-4 hour
collections and allowed to age for 2 days. The most comrnon features of these
treatments were the complete loss of anterior and dorsal structures of the
embryo. The number of denticle belts was generally maintained but was
condensed along the remaining ventral regions of the embryo.
These results show that, quantitatively, the W C exposure protocol used
here had a much p a t e r effect on Drosophila embryo lethality than both hsreaper
and hsTNF. Qualitatively, however, the effect was similar to deletions of anterior
and dorsal structures of the embryo observed as the predominant morphological
changes relative to the wildtype embryos.
3.4 hsTNF vs. UVC Activation of reaperlacz Expression
The expression of renper predicts ce11 death in Drosophila (81) making it a
likely target for potential hsTNF or W C ce11 death activating activity. To
determine the ability of these two stimuli to transcriptionally activate reaper and
facilitate comparison, the TNFa insert on the second chromosome (hsTNF II) was
genetically combined with the renperlncz reporter construct located on the third
chromosome (table 2). The resulting line, hsTNF;reaperlacZ was then subjected to
either a heat shock or W C treatment. The embryonic expression pattern of the
reaper promoter was then determined by anti-Bgalactosidase antibody staining.

In untreated embryos, reaper expression is first observed in the head and
tail regions as the germ band extends towards the anterior of the embryo in stage
9 (Figure 6A). By stage 10, expression is obsewed in clusters in each segment of
the developing embryo (Figure 68 and C). UVC treatment of hsTNF;renperlacz
embryos was able to activate an early expression of reaperlacz when it is norrnally
not expressed (not shown). UVC treatment was also able to induce ectopic
expression at later stages when reaper is expressed in the developing segments
(Figure 6G-1). Heat shock treatment of hs TNF;reaperlacz embryos, which drives
the expression of the TNF insert, did have some effect of expanding the areas of
existing reaper expression indicated by the lac2 reporter constnict (Figure 6D-F).
The level of reaper expression observed in Figure 6 for hsTZVF expression (D-F)
appears sirnilar to that for UVC exposure ((2-1). Upon closer examination the
observation of embryos with such significant P-galactosidase staining was much
less frequent than with W C exposure. UVC exposure was able to consistently
and dramatically increase the level of P-galactosidase staining in
hsTNF;reaperlacZ embryos. This is in line with other observations where ionizing
radiation has also been shown to activate reaper expression in Drosophila (15,60).
3.5 hsï7Vï vs. UVC Activation of Apoptosis
Instead of examining an indirect measure of cell death activation, we
wanted to detennine the ability of TNF-a and W C to directly activate apoptosis
in Drosophila embryos. Aaidine orange (AO) can be used to selectively stain
apoptotic cells in developing Drosophila embryos (19). This is based on the
inaeased permeability of the ceii membrane in cells undergohg apoptosis.

Here, the identical method of A 0 staining was used to compare the ability of
hsTNF versus UVC to activate apoptosis.
In these experiments, 0-4 hour w"'~ and hsTNF Drosophiln embryos were
collected. w"'~ embryos were then either left untreated or subjected to a UVC
treatment while hsTNF embryos were given an &minute 37 O C heat shock
treatment. The embryos were then aged for another 2 hours before being stained
with the dye.
Untreated w"'~ embryos at this stage (pre stage 11) show no A 0 staining
under these conditions (Figure 7A). This is consistent with the Abram's
observation that apoptotic ce11 death in Drosophila embryonic development does
not normally begin until stage 11 or approximately 7 hours of development (19).
In the case of the UVC-treated embryos, however, signhcant A 0 staining was
observed (Figure 78). A 0 staining after expression of TNFa by heat shock was
also observed (Figure 7C) though not as consistently or dramatically as with
UVC treatment. Under these conditions UVC, and to a limited extent, hsTNF
expression was able to activate programmed cell death by apoptosis in Drosophil~
embryos prior to its normal occurrence.
3.6 W C Response in Hg9 Embryos
The genes of the Hg9 region on the third chromosome, p i m , reaper, and
hid, are implicated in virtually ali cell death in the Drosophila embryo (79). To
detennine if these genes are required for the apoptotic cell death observed as a
result of W C exposure, the experimental procedure required a method of
visually distinguishing embryos that had completely lost the if99 deletion region
versus those that stiil had the balancer. This would allow for a visual
41

cornparison of the resulting cuticle phenotype and a detemiination of the
phenotypic effect of deletion of the H99 chromosomal region.
For this experiment, the Hg9 deletion chromosome from the line H99,kni
[ri4 ]p[p]/TM3,Sb[l 1 was gene ticall y combhed over the marker, yel low-plus, in a
yellow background (from the line y w prd GAL4;Ly/TM3,y+ e Ser) as described in
the Materials and Methods. This marker was used to determine the presence or
absence of Hg9 on the third chromosome (82). The resulting line is y w
prdCAL4;H99/TM3,y+ e Ser designated y;H99@. Yellow embryos of this line
indicate that both third chromosomes have the Hg9 deletion while non-yellow
embryos have at least one wildtype third chromosome.
Embryos resulting from y;H99&+ sibling crosses were exposed to U'V'C.
Cuticle preparations were then performed to determine their phenotype. Yellow
versus non-yellow colour determination was made under bright-field
microscopy. Both non-yellow (at least one wildtype third chromosome) and
yellow (both copies of Hg9 region lost) embryos showed the same phenotype
(Figure SA and B respeaively). The loss of dorsal and anterior regions with the
condensation of the denticle belh along the remaining ventral regions was
observed as in wildtype WC-treated embryos.
The sarne phenotype was also obsewed in untreated, yellow embryos (not
shown). These embryos harbour a complete loss of the Hg9 region. This
phenotype may therefore result from any aberration of cell death during
embryonic development.
In light of these results, the significance of WC activation of reaper cornes
into question. Other studies have shown that loss of reaper blocks apoptosis
induction by low doses of X-irradiation. Higher doses, however, were able to
42

induce apoptosis in the same mutant embryos (15). The dose of UVC used here
may represent a relatively high dose considering that the loss of reaper had no
apparent effect, at least on the cuticle phenotype generated by W C exposure
(table 3). The Hg9 experirnent suggests that reaper, along with grim and hid, are
not required for the ce11 death phenotype induced by UVC exposure at this dose.
If, in fact, grim, reuper, and hid, are not required for the ce11 dea th observed
by high UVC doses, there may be other pathways leading to the activation of
apoptotic ce11 death.
3.7 Response of Other P Elements to UVB and W C Exposure
In this shidy, the search for pathways outside of the genes of the H99
region that lead to the activation of apoptosis in response to UV exposure began
with the use of Drosophila P elements. The Spradling P element collection is a
large collection of unique P element insertions in genes throughout the Drosophila
genome (65). It has been used extensively in Drosophila research for identifying
novel components of signaling pathways. Most commoniy, the P element
collection is used to screen for suppressors and/or enhancers of phenotypes
induced by characterized genetic mutations. P elements that are observed to
alter the severity of the phenotype are irnplicated as part of the mechanism that
created the original phenotype. In this study, the expression of P elements in
response to W was determined. The Spradling collection of P-elements
represents 25% of the genes required for Drosophila viability (83). This means
that a signhcant portion of the important genes in Drosophila can be screened
for their interaction with UV.

In Our initial attempt to screen P elements for responsiveness to UV,
Drosophila larval imaginal discs were exposed to a UVB (270-330 nm) source. The
larval imaginal discs are self-contained sacs of epithelial cells that develop into
the adult legs, wings, and other various structures (84). During their
development they contain cells representing al1 stages of the ce11 cycle. The
responsiveness of each P element Line to UVB was determined by virtue of the @
galactosidase gene associated with the P elements (67,85). First to second instar
Drosophila lawae were exposed to approximately 90 J/m2 of UVB. After a 24-hr
recovery period under dark conditions, the lama1 heads were isolated and
inverted, exposing their imaginal discs. The inverted heads were fiied and then
stained for fbgalactosidase activity. Figure 9 shows a cornparisor, of staining in
imaginal discs of the P element line 862 without (A) and after (B) WB exposure.
In this P element line, it appears that exposure to W B under these conditions is
able to widen the area of expression of the P element.
Table 4 is a surnmary of the initial qualitative results with a number of P
element lines whose imaginal discs have been exposed to UVB. For each
cornparison, the imagina1 discs from 3-5 lawae are compared for &galactosidase
staining before and after exposure to the W B source. Of the 68 lines tested in
this manner, four produced significant increases in bgalactosidase staining after
UVB exposure (P862, P1244, P1341, and P1587) while two showed a significant
decrease (Pl062 and P1542). These initial results suggest that P elements may be
used to identify selective transcriptional responses in Drosophila. These
responses can be either for activation or repression of transcription. Subsequent

exposures of these P elements did not always produce the same result of
increased X-gal staining.
A preliminary screen was also conducted in Drosophila embryos for
responsiveness to UVC (254 nm). This was done in attempt to create another
experirnental protocol whereby the ability of UV exposure to effect gene
expression and regulation in Drosophila could be determined. Here, P element
embryos were collected, exposed to UVC, and then stained with anti-b
galactosidase antibody. The effect on the Pgalactosidase expression, and
therefore the associated P element, was thus determined. Table 5 shows the
initial qualitative effects of W C on the observed anti-Bgalactosidase staining in
a few selected Drosophila P element embryos. Pl642, a P element insertion in the
hid gene, shows no response to W C under these conditions. Hid is one of the
genes in the Hg9 deletion also found to be a significant player in Drosophila
prograxmned ce11 death. Other studies using ionizing radiation have also shown
that hid did not respond under conditions of environmental stress where reaper
did (19).
None of the P-element lines tested here showed a significant
transcriptional response. However, the positive response of the reaperlacz
reporter consmict along with the UVB results in imaginal discs suggests that
other responses may be observed. Finding the ideal conditions and use of a
wider selection on P elements may be the key.

4. Discussion
4.1 OBJECTIVES
This work describes the search for a system of activating ce11 death in
Drosophila that will allow for further study into the mechanism of the activation
of apoptosis. To date, only a relatively few genes have been identified in
Drosophila as being involved in the activation and regdation of ce11 death. The
availability of Drosophila as a genetic tool for determining functional pathways
and identifying novel pathway members suggests that this organism could make
great contributions to the study of programmed ce11 death by apoptosis.
One of the tools available in Drosophila is the use of P elements that can be
used to discover novel elements of known molecular pathways. In these
experiments, two stimuli, TNFa expression and UV exposure are used in attempt
to create ce11 death phenotypes in Drosophila. After establishing a ce11 death
phenotype caused by one or both of these stimuli, the aim of this line of
investigation is to establish a P element screen to search for novel genes and
pathways involved in apop tosis in Drosophiln.
4.2 SUlMMARY OF RESULTS
4.2.1 hsreaper, UVC, and h s M cause lethality in Drosophila embryos
Lethality experiments show that hsreaper and hsTNF show similar lethality
after heat shock. hsreaper is used here for comparison because the ability of
reaper to activate programmed ceil death has already been detennined. The
observation that both hsreaper and hsTNF cause similar levels of lethality in

-- Drosophila embryos suggests that expression of hsTNF may act in a non-specific
manner.
W C treatment of w"'' embryos is signihcantly higher at 95% (Figure 4).
The dose of radiation received by the embryos here is in the range of 200-400
J/m2. This corresponds to approxirnately 30 seconds of exposure in the
StratalinkerTM used in this study. An early study by Donini and Boyd studyhg
embryo survival in response to UV exposure showed that doses in the range of
40-50 J/m2 are able to cause 90-957'0 lethality (61). In their work, sumival of
embryos to adulthood was measured. This test is more stringent than that used
in this study where lethality was determined after only sumival to hatching. The
level of exposure used here results in essentially quantitative lethality of the
exposed Drosophila embryos.
An examination of the cuticle phenotype shows that al1 three effectors,
hsrenpet, hsTNF, and UVC, cause sirnilar loss of dorsal and anterior structures.
Also, the denticle belts and naked segments are condensed along the remaining
ventral regions of the embryos. This phenotype is not a ventralization of the
embryo but demonstrates a complete lossof the dorsal and anterior structures.
This phenotype shows similar features as basket (54,86). Cuticle phenotypes in
this and other mutants of the Drosophila JNK pathway result from the failure of
dorsal closure. In untreated H99-deletion embryos, the loss of anterior and
dorsal features is also observed. This may suggest that any disruption of the
process, whether loss of regulators or activation of cell death, may be responsible
for the observed phenotype.
Work done by Herrlich et al. (87) suggests that cells are able to initiate a
molecular response to cellular damage caused by WB doses of up to 5 kJ/m2.
47

This suggests that these doses of radiation are able to act as a molecular signal
activating signaling transduction pathways and affecting protein translation or
post-translational processing. Since the embryos are not physically destroyed by
W C and no heat is generated during the exposure, W C may also be generating
a molecular response leading to the observed lethality. The lethality and cuticle
phenotype results suggest that W C exposure is a strong inducer of lethality in
Drosophila embryos. The ability of this agent to specifically activate
programrned ce11 death by apoptosis is the next point of cowideration in this
study.
Expression of rcaper is already established as being able to induce
apoptotic ce11 death in Drosophila. This allows its use here as a control for hsTNF
expression and UV exposure. The similar behaviour of hsTNF to hsreaper in
lethality and cuticle phenotype only suggests that further investigation of hsTNF
may be watranted.
4.2.2 W C but not hsTNF Activates Expression of reaperlacz in Drosophila
Embryos
Previous studies have shown that reaper is not only able to activate
apoptotic cell death but acts as a marker of cell death. Expression of reaper often
precedes and predicts ce11 death by various stimuli (81). Since reaper expression
has been shown to be a marker of programmed ceil deaths in Drosophila, the
ability of TNFa and UVC to activate reuper expression was then assessed. For
this determination the line, hsTNF;reaperlacz, wwas used to test the response of
reaper to both TNFa expression and WC exposure. This Drosophila line

-- = - contains a genetic insert that uses the reaper promoter element to drive the
expression of the ~Galactosidase gene from the yeast Lac operon. In this
experiment, PGalactosidase expression is driven by the reaper promoter and
monitored by anti-B-galactosidase antibody staining. The resulting expression
pattern represents the expression pattern of the reaper promoter
As shown in Figure 6A-C, reaper expression begins at approximately stage
9 and increases through stage 10. W C exposure is able to expand regions of
reaper expression and induce ectopic expression in the head and between
segments (Figure 6G-1). When the same line, hsTNF;renperlacz was subjected to a
37 O C heat shock, some expansion of reaper expression also seen (Figure 6D-F).
TNFa expression, however, did not cause the same level of ectopic expression.
Reaper has already been determined to be an integrator of various types of
stimuli, both developmental and extemal, to ce11 death signals (John Abrarns,
Ontario Cancer Institute persona1 communication). Here, it is shown that W C is
a strong activator of reuper expression. The UVC activation of the gene reaper
represents one specific molecular response to an extemal environmental signal.
Thus reaper represents a potential mediator of c d death activation by UVC.
When we examined WC-induced cutide phenotypes in the absence of reaper
and the other genes of the Hg9 chromosomal deletion, grim, and hid, we found no
change in the observed phenotype (Figure 8). Both untreated and UVC-treated
H99-deletion embryos showed similar phenotype as WC-treated w"'' embryos.
This phenotype may therefore result from any disruption of the ce11 death
program during embryo development.

4.2.3 UVC but not h s m Activates Apoptosis in Drosophila Embryos
One of the hallmark features of apoptotic ce11 death is the breakdown of
the ce11 membrane. This feature allows the fluorescent dye acridine orange to be
incorporated into dying cells making it an excellent marker for apoptosis (19).
Figure 7 shows that UVC treatment is able to activate apoptosis as indicated by
the increase in acridine staining over the untreated wildtype embryos. The
embryos here are 2-6 hours into development and represent stages where
programmed ce11 death is normally not obsenred. During normal development
programmed ce11 death is not observed in Drosophila embryos until stage 11 or at
least 7 hours of development (19). The observation of acridine orange staining in
embryos prior to stage 11 of development indicates that ai least some portion of
the apoptotic machinery is in place prior to the appearance of normal ce11 death
in embryos. This machinery c m apparently be activated by exposure to the dose
W C used here. This supports the notion that UVC interacts with genetic
components of biological systems to activate ce11 death. In some cases (Figure
7C) TNFa expression increased the number of cells that were able to take up the
A 0 stain. However, the increase in acridine orange staining was not as
sigruficant or as consistent as in the case with W C exposure.
Taken together, however, these results suggest that expression of hsTNF
may cause lethality and cuticle phenotype by a non-specific mechanism. The
expressed protein may not adopt the appropriate tertiary or quatemary
structures. Also, there may simply be no TNF receptor expressed in Drosophila
embryos to respond to the TNF ligand expressed in these experiments. Thus far,
searches of the available Drosophila genome sequence have not revealed any

suitable TNF recep tor sequence. The lethality observed a fier TNFa expression
may simply be due to non-specific toxic effects.
4.3 Results in the context of cumnt Research
An important finding in this study is that the pro-apoptotic gene, reaper, is
transcriptionally activated in response to UVC exposure even prior to the normal
appearance of programmed ce11 death (Figure 6G-1). This finding corroborates
similar results using X-irradiation as the stimulus (15) and further establishes
reaper as an integrator of external signals to apoptotic ce11 death. As discussed in
the introduction, researchers have discovered a Drosophila homologue of
mammalian p53 capable of transcriptionally activating the pro-apoptotic gene
reaper (59,60,88). In light of this result, the obsewed transcriptional activation of
reaper and activation of apoptosis by UVC (Figure 6) may be mediated in part by
the activity of Drosophila p53. The activity of p53 in responding to radiation-
induced DNA damage by transcriptionally activating reaper creates the link
between the external action of UVC or X-irradiation and the intemal activation of
apoptotic cell death. The next likely step in this research is to determine the
activation statu of p53 and its ability to activate reaper in response to WC.
The existence of Drosophila p53 also serves as a possible explanation for
the observation by White et RI that higher doses of radiation can bypass the
requirement of reaper (15). In this study, loss of the H99 region containhg the
apoptotic regulatory elements grim, reaper, and hid, resulted in no change in the
obse~ed embryonic cuticle phenotype (Figure 8). The observation of apoptotic
responses in the absence of reaper may be a result of p53 transcriptional
activation of other pro-apoptotic targets. Further research into radiation-induced
51

apoptosis in Drosophila is likely to focus on pathways involving p53 andfor
reaper activation and other pro-apoptotic targets of p53 in Drosophiln.
m e r points of research may include distinguishing between DNA
damagedependent and damage-independent mechanisms of ce11 death
activation. Here, the use of repair-deficient Drosophila mutants may be of
assistance to compare the molecular outcome exposure to radiation.
Based on comparisons to other work, the W C radiation protocol used
here likely falls into the category of high dose response. h other studies, high
doses of radiation have been shown to bypass the requirement for protein
synthesis and in Drosuphila, the requirement for the regulatory genes of the H99
region. The question of exactly how the apoptotic effector machinery is activated
in situations of high radiation doses is another possible area to explore in further
research.
4.4 FUTURE WORK
Future work stemming from this research wili likely focus on the
activation of reaper by UVC radiation. Recently, researchers have published
reports of a Drosophila p53 and p53 binding sites in the reaper promoter. These
hdings establish a possible a link between the UVC exposure and activation of
reaper transcription and apoptosis observed in the preceding experiments. A
simple explanation may be that UVC activation of apoptosis occurs via the DNA-
damage dependent activation of p53 and subsequent transcriptional activation of
reaper. However, since high doses of radiation are able to bypass the
requirement for protein synthesis and Drosophila regulatory elements, there are
quite likely other pathways involved.
52

Present models of W activation of apoptosis identify p53, bcl-2, bax ,
Apaf 1, caspases, and NF-KB as pathway members. With the recent discoveries
of p53 (59,60), bcl-2/bax (89,90,91,92), and Apafl (93,94) Drosophila homologs,
the known pathways leading to apoptosis in this organism have become more
reflective of what is known in manunals. The ability of these genes to respond to
UV and the level of apoptosis observed in genetic mutants are early experiments
to be conducted with these genes. Genetic combinations of mutants of these
genes with the Hg9 deletion and other existing apoptosis mutants will help to
determine if these newly discovered Drosophila homologs interact with the
apoptotic regulators already known.
Although experiments with mouse TNF in Drosophila failed to produce
conclusive results in the activation of apoptotic ce11 death, the TNF model of
signaling in the cytoplasm may still have application in understanding the UV
response. The basic model of TM: signaling suggests that protein-protein
interactions lead to the aggregation and self-activation of caspases requiring no
new protein synthesis. High doses of radiation, and specifically U V , may bypass
the requirement for regdatory elements by promoting the aggregation of the
caspases themselves. In the model of Ligand-independent activation of receptor
tyrosine kinases, exposure to UV radiation is able to fuse receptors together by
forming stable disulfide bonds. This leads to the association of their cytoplasmic
domains and autophosphorylation leading to activation of the signaling pathway
(51,52). In vitro studies of caspase physical structure could be used to determine
if UV is in fact able to ma te stable bonds between individual caspase protein
molecules. If stable bonds are obsewed between specific arnino acids point
mutations could be introduced to replace these critical amino acids. The
53

genetically altered caspase protein couid be expressed in Drosophila caspase-nul1
mutants to determine the UV response in vivo.
Research in the genetically simpler and more easily accessible organism
Drosophila will be quite usefd to understanding how each of these plays a role in
radiation-induced apoptosis. This research may lead to a better understanding
of situations where radiation therapy been ineffective and help create new
criteria for determining methods of treatment. Understanding radiation-induced
apoptosis may make existing radiation treatments more efficient and help to
develop novel therapy strategies that take advantage of radiation-induced
apoptosis to eliminate potentially harmful cancer cells (95).

REFERENCES
1. J. F. Kerr, A. H. Wyllie, A. R. Currie, Br 1 Cancer 26,239-57
(1972).Apoptosis: a basic biological phenomenon with wide-ranging implications
in tissue kinetics
2. G. Majno, 1. Joris, Am J Pathol146,3-15 (1995).Apoptosis, oncosis, and
necrosis. An overview of ce11 death [see comments]
3. M. D. Jacobson, M. Weil, M. C. Raff, Cell88,347-54 (1997).Programmed
cell death in animal development
4. C. B. Thompson, Science 267,1456-62 (1995).Apoptosis in the pathogenesis
and treatment of disease
5. A. Bergmann, J. Agapite, H. Steller, Oncogene 17,3215-23
(1998).Mechanisms and control of programrned ce11 death in invertebrates
6. D. Hockenbery, G. Nunez, C. Milliman, R. D. Schreiber, S. J. Korsmeyer,
Nature 348,334-6 (1990).Bcl-2 is an inner mitodiondrial membrane protein that
blocks programmed ce11 death
7. Y. Tsujimoto, C. M. Croce, Proc Nat1 Acad Sci U S A 83,52148
(1986).Anaiysis of the structure, transcripts, and protein products of bcl-2, the
gene involved in human follicular lymphoma
8. T. J. McDomell, et al., Mol C d Bi01 10,1901-7 (1990).Deregulated Bcl-2-
immunoglobulin transgene expands a resthg but responsive immunoglobulin M
and Dsxpressing B-ce11 population
9. T. J. McDonnell, et al., Cell57,79-88 (1989).bcl-2-immunoglobulin
transgenic mice demonstrate extended B cell survival and foliicular
lymphoproliferation
55

10. J. E. Sulston, H. R HoMtz, Dm Bi01 56,110-56 (1977).Postsrnbryonic ce11
lineages of the nematode, Caenorhabditis elegans
11. H. R. HoMtz, P. W. Sternberg, 1. S. Greenwald, W. Fixsen, H. M. Ellis,
Cold Spring Harb Symp Quant Bi01 48,453-63 (1983).Mutations that affect neural
ce11 lineages and ce11 fates diiring the development of the nematode
Caenorhabditis elegans
12. E. M. Hedgecock, J. E. Sulston, J. N. Thomson, Science 220,1277-9
(1983).Mutations affecting programmed cell deaths in the nematode
Caenorhabditis elegans
13. Y. Hu, M. A. Benedict, D. Wu, N. Inohara, G. Nunez, Proc Nat1 Acad Sci U
S A 95,4386-91 (1998).Bcl-XL interacts with Apaf-1 and inhibits Apaf-1-
dependent caspase-9 activation
14. M. Ashbumer, Drosophila-A Laboratoy Manual (Cold Spring Harbor
Laboratory Press, 1989).
15. K. White, et al., Science 264,677-83 (1994).Genetic control of programmed
ce11 death in Drosophila [see comments]
16. P. Chen, P. Lee, L. Otto, J. Abrams, 1 Bi01 Chem 271,25735-7
(1996).Apoptotic activity of REAPER is distinct from signaling by the hunor
necrosis factor receptor 1 death domain
17. A. G. Fraser, G. 1. Evan, Embo J16,280513 (1997).Identification of a
Drosophila melanogaster ICE/CED-Srelated protease, drICE
18. B. A. Hay, D. A. Wassarman, G. M. Rubin, Ce11 83,1253-62
(1995).Drosophila homologs of baculovinis inhibitor of apoptosis proteins
function to block celi death

19. J. M. Abrams, K. White, L. 1. Fessler, H. Steller, Development 117,29-43
(1993).Programmed cell death during Drosophila embryogenesis
20. J. Delic, J. Coppey, H. Magdelenat, M. Coppey-Moisan, Exp Ce11 Res 194,
147-53 (1991).Impossibility of acridine orange intercalation in nuclear DNA of the
living ce11
21. T. M. Pazdera, P. Janardhan, J. S. Minden, Dewlopment 125,3427-36
(1998).Pattemed epidermal ce11 death in wild-type and segment polarity mutant
Drosophila embryos
22. H. R. Stennicke, G. S. Salvesen, Ce11 Death Differ 6,1054-9 (1999).Catalytic
properties of the caspases
23. T. T. Yamin, J. M. Ayala, D. K. Miller, 1 Biol Chem 271,13273-82
(1996).Activation of the native 45-kDa precursor form of interleukin-l-
converting enzyme
24. Y. Gu, et al., Embo 1 14,1923-31 (1995).Interleukin-1 beta converting
enzyme requires oligomerization for activity of processed forms in vivo
25. H. Sakahira, M. Enari, S. Nagata, Nature 391,969 (1998j.Cleavage of CAD
inhibitor in CAD activation and DNA degradation during apoptosis [see
comrnents]
26. C. M. Simbulan-Rosenthal, D. S. Rosenthal, S. Iyer, A. H. Boulares, M. E.
Srnulson, 1 Biol Chem 273,13703-12 (1998).Transient poly(ADP-ribosy1)ation of . nuclear proteins and role of poly(ADP-ribose) polymerase in the early stages of
apoptosis
27. A. H. Boulares, et al., 1 Biol Chem 274,22932-40 (1999).Role of poly(ADP-
ribose) polymerase (PARP) cleavage in apoptosis. Caspase bresistant PARP
mutant increases rates of apoptosis in transfected cells
57

28. J. M. Lahti, J. Xiang, L. S. Heath, D. Campana, V. J. Kidd, Mol Ce11 Bi01 15,
1-1 1 (1995).PITSLRE protein kinase activity is associated with apoptosis
29. A. Fraser, G. Evan, Cell85,781-4 (1996).A license to kill
30. G. A. Manji, R. R. Hozak, D. J. LaCount, P. D. Friesen, 1 Virol71,4509-16
(1997).Baculovirus inhibitor of apoptosis functions at or upstream of the
apoptotic suppressor P35 to prevent programmed ce11 death
31. A. Sugirnoto, P. D. Friesen, J. H. Rothman, Embo 113,2023-8
(1994).Baculovirus p35 prevents developmentally programmed ce11 death and
rescues a ced-9 mutant in the nematode Caenorhabditis elegans
32. D. Xue, H. R. Horvitz, Nature 377,248-51 (1995).lnhibition of the
Caenorhabditis elegans cell-death protease CED3 by a CEP3 cleavage site in
baculovirus p35 protein
33. B. A. Hay, T. Wolff, G. M. Rubin, Development 120,2121-9
(1994).Expression of baculovirus P35 prevents ce11 death in Drosophila
34. D. R. Beidler, M. Tewari, P. D. Friesen, G. Poirier, V. M. Dixit, 1 Biol Chem
270,16526-8 (1995).The baculovirus p35 protein inhibits Fas- and tumor necrosis
factor- induced apoptosis
35. M. Tewari, W. G. Telford, R. A. Miller, V. M. Dixit, Bi01 Chem 270,22705-
8. (1995).CrmA, a poxvirus-encoded serpin, inhibits cytotoxic T-lymphocyte-
mediated apop tosis
36. L. A. Tartaglia, T. M. Ayres, G. H. Wong, D. V. Goeddel, Ce11 74,845-53
(1993).A novel domain within the 55 kd TNF receptor signals cell death
37. N. Itoh, S. Nagata, \ Biol Chem 268,10932-7 (1993)A novel protein domain
required for apoptosis. Mutational analysis of human Fas antigen

38. C. A. Smith, T. Farrah, R G. Goodwin, Cell76,959-62 (1994).The TNF
receptor superfamily of cellular and viral proteins: activation, costimulation, and
death
39. S. J. Baker, E. P. Reddy, Oncogene 17,3261-70 (1998).Modulation of life and
death by the TNF receptor superfamily
40. L. H. Boise, C. B. Thompson, Science 274,674 (1996).Hierardiical control
of lymphocyte swiva l
41. S. Chandler, et al., J NeuroimmunoI 72,15561 (1997).Matrix
metalloproteinases, tumor necrosis factor and multiple sclerosis: an overview
42. C. P. Blobel, Ce11 90,589-92 (1997).Metalloprotease-disintegins: links to
ce11 adhesion and cleavage of TNF alpha and Notch
43. H. Hsu, H. B. Shu, M. G. Pan, D. V. Goeddel, Ce11 84,299-308
(1996).TRADD-TRAF2 and TRADD-FADD interactions define two distinct TNF
receptor 1 signal transduction pathways
44. A. M. Chinnaiyan, K. O'Rourke, M. Tewari, V. M. Dixit, Ce11 81,505-12
(1995).FADD, a novel death domain-containing protein, interacts with the death
domain of Fas and initiates apoptosis
45. Z. G. Liu, H. Hsu, D. V. Goeddel, M. Karin, Ce11 87,565-76
(1996).Dissection of TNF receptor 1 effector functions: JNK activation is not
linked to apoptosis while NF-kappaB activation prevents ce11 death
46. S. Schutze, K. Wiegmann, T. Machleidt, M. Kronke, Immunobiology 193,
193-203 (1995).TNF-induced activation of NF-kappa B
47. D. J. Van Antwerp, S. J. Martin, 1. M. Verma, D. R. Green, Trends Ce11 Biol
8,107-1 1 (1998).Inhibition of TNF-induced apoptosis b y NF-kappa B

48. J. F. Ward, Radint Res 138, S85-8 (1994).DNA damage as the cause of
ionizing radiation-induced gene activation
49. Y. Devary, R. A. Gottlieb, T. Smeal, M. Karin, Cell 71,1081-91 (1992).The
mammalian ultraviolet response is triggered by activation of Src tyrosine kinases
50. C. Sachsenmaier, et al., Ce11 78,963-72 (1994).Involvement of growth factor
receptors in the rnammalian UVC response
51. M. Kato, et al., Mol Bi01 Cell 11,93- 101. (2000).Ultraviolet light induces
redox reaction-mediated dimerkation and superactivation of oncogenic Ret
tyrosine kinases
52. M. Kato, et al., Antioxid Redox Signal 2, $41-9. (2000).Molecular mechanism
of activation and superactivation of Ret tyrosine kinases by ultraviolet light
irradiation
53. J. R. Woodgett, J. Avruch, J. M. Kyriakis, Clin Exp Pharmacol Physiol22,
281-3 (1995).Regulation of nuclear transcription factors by stress signals
54. H. K. Sluss, Z. Han, T. Barrett, R J. Davis, Y. T. Ip, Genes Dev 10,2745-58
(1996).A JNK signal transduction pathway that mediates morphogenesis and an
immune response in Drosophila
55. L. J. Ko, C. Prives, Genes Dev 10,1054-72 (1996).p53: puzzle and paradigrn
56. W. Maltzman, L. Czyzyk, Mol Ce11 Biol 4,1689-94 (1984) .UV irradiation
stimulates levels of p53 cellular hunor antigen in nontransformed mouse cells
57. T. Miyashita, J. C. Reed, Cell80,293-9 (1995).Tumor suppressor p53 is a
direct transcriptional activator of the human bax gene
58. L. B. Owen-Schaub, et al., Mol Ce12 Biol15,3032-40 (1995).Wild-type
human p53 and a tempera-sensitive mutant induce Fas/APO-1 expression

damage response element at the reaper locus
60. W. Nordstrom, J. M. Abrams, Cell Death Differ 7,1035-1038
(2000).Guardian ancestry: fly p53 and damage-inducible apoptosis
61. S. D. Donini, J. B. Boyd, Mutat Res 44,5362 (1977)Sensitivity of wild type
and recombination-deficient strains of Drosophila melanogaster to ultraviolet
radiation
62. T. M. Koval, Experientia 43,445-6 (1987).Photoreactivation of UV damage
in cultured Drosophila cells
63. J. G. de Cock, et al., Nucleic Acids Res 20,4789-93 (1992).Repair of UV-
induced (64)photoproducts measured in individual genes in the Drosophila
embryonic Kc ce11 Iine
64. P. Poltronieri, et al., Biochem Ce11 Bi01 75,445-9 (1997).PARP cleavage in the
apoptotic pathway in S2 cells from Drosophila melanogaster
65. L. Cooley, R. Kelley, A. Spradling, Science 239,1121-8 (1988).Insertional
mutagenesis of the Drosophila genome with single P elements
66. E. F. F. J. Sambrook, T. Maniatis, Molecular Cloning, A kiboratonj Manital. C.
Nolan, Ed. (Cold S p ~ g Harbor Laboratory Press, ed. 2nd Edition, 1989).
67. H. J. Bellen, et al., Genes Dm 3,1288-300 (1989).P-element-mediated
enhancer detection: a versatile method to study development in Drosophila
68. S. Greaves, B. Sanson, P. White, J. P. Vincent, Genetics 153,1753-66
(1999).A screen for i d e n m g genes interacting with amadillo, the drosophila
homolog of beta-catenin [In Process Citation]
69. F. D. Karim, et al., Genetics 143,315-29 (1996) .A screen for genes that
function downstream of Ras1 during Drosophila eye development
61

70. D. Vucic, S. Seshagiri, L. K. Miller, Mol Ce11 Bi01 17,667-76
(1997).Characterization of reaper- and FADD-induced apoptosis in a
lepidopteran ce11 line
71. K. McCail, H. Steller, Trends Genet 13,222-6 (1997).Facing death in the fly:
genetic analysis of apoptosis in Drosophila
72. A. C. Spradling, G. M. Rubin, Science 218,341-7 (1982).Transposition of
cloned P elements into Drosophila germ line chromosomes
73. H. Steller, V. Pirrotta, Mol Ce11 Bi01 6,1640-9 (1986)P transposons
controlled by the heat shock promoter
74. C. Perez, et al., Ce11 63,251-8 (1990).A nonsecretable ce11 surface mutant of
tumor necrosis factor (TNF) kills by cell-to-ce11 contact
75. K. Katanosaka, F. Tokunaga, S. Kawamura, K. Ozaki, FEBS Lett 424,149-
54 (1998).N-linked glycosylation of Drosophila rhodopsin occurs exclusively in
the amino-terminal domain and functions in rhodopsin maturation
76. M. Tanaka, T. Suda, T. Takahashi, S. Nagata, Embo 1 14,1129-35
(1995).Expression of the functional soluble form of human fas ligand in activated
lymphocytes
77. A. Martinez-Arias, in The Development of Drosophila melanogaster M . Bate,
A. Marünez-Arias, Eds. (Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, New York, 1993) pp. 517608.
78. P. Chen, W. Nordstrom, B. Gish, J. M. Abrams, Genes Dev 10,1773-82
(1996).grim, a novel cell death gene in Drosophila
79. M. E. Grether, J. M. Abrams, J. Agapite, K. White, H. Steller, Genes Dm 9,
1694-708 (1995).The head involution defective gene of Drosophila melanogaster
functions in programmed cell death
62

80. M. J. Sonnenfeld, J. R. Jacobs, Development 121,569-78 (1995).Apoptosis of
the midline glia during Drosophila embryogenesis: a correlation with axon
contact
81. K. White, E. Tahaoglu, H. Steller, Science 271,805-7 (1996)Cell killing by
the Drosophila gene reaper
82. R. C. Binari, et al., Development 124,2623-32. (1997).Genetic evidence that
heparin-like glycosaminoglycans are involved in wingless signaling
83. A. C. Spradling, et al., Genetics 153,135-77 (1999).The Berkeley Drosophila
Genome Project gene disruption project: Single P-element insertions mutating
25% of vital Drosophila genes
84. S. M. Cohen, in Development of Drosophiln M . B. A. Martinez-Arias, Ed.
(Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1993).
85. C. Wilson, et al., Genes Da, 3,1301-13 (1989) .P-element-mediated enhancer
detection: an efficient method for isolating and characterizhg developmentally
regulated genes in Drosophila
86. J. R. Riesgo-Escovar, M. Jenni, A. Fritz, E. Hafen, Genes Dm 10,2759-68
(1996).The Drosophila Jun-N-terminal kinase is required for ce11 morphogenesis
but not for DJun-dependent ce11 fate specification in the eye
87. P. Herrlich, et al., C R Acnd Sci 111 322,121-5 (1999).Radiation-hduced
signal transduction. Mecha~sms and consequences
88. M. Ohann, et al., Ce11 101,91-101 (2ûûû).Drosophila p53 is a structural
and functional homolog of the -or suppressor p53
89. C. B. Brachmm, O. W. Jassim, B. D. Wachsmuth, R. L. Cagan, Curr Bi01
10,547-50. (ZûûO).The Drosophila bcl-2 family member dBorg-1 functions in the
apop totic response to UV-irradiation
63

90. H. Zhang, et al., J Bi01 Chem 275,27303-6. (2000).Drosophila pro-apoptotic
Bd-2/Bax homologue reveals evolutionary conservation of ce11 death
mechanisms
91. T.Igaki,etal.,ProcNatlAcndSciUSA97,662-7.(2000).Drob-l,a
Drosophila mernber of the Bcl-2/CED-9 family that promotes ce11 death
92. P. A. Colussi, et al., J Ce11 Bi01 148,70344. (2000).Debcl, a proapoptotic Bcl-
2 homologue, is a component of the Drosophila melanogaster ce11 death
machinery
93. L. Zhou, 2. Song, J. Tittel, H. Steller, Mol Ce11 4,745-55. (1999).HAC-1, a
Drosophila homolog of APAF-1 and CED-4 functions in developmental and
radiation-induced apoptosis
94. A. Rodriguez, et al., Nat Ce11 Biol 1,272-9. (1999).Dark is a Drosophila
homologue of Apaf-l/CED-4 and functions in an evolutionarily conserved death
pathway
95. C. A. Schmitt, S. W. Lowe, J Pathol187,127-37. (1999).Apoptosis and
therap y

1 Plasmid I Source 1 phsrr 1 Perrimon Lab
Car1 S. Thummel and V. Pirrotta, Genebank AC US9056
Table 1. Plasmid stocks.
pBluescript mTNF phsCaSpeR mTNF
Wen Chen, Amgen Research Institute Maurice Ennis, Ontario Cancer Institute
Stock W~~~~
wfl1$UCyO; Ki flzw20flM3 yw;hsRpr[w+l46A Sp/CyO;rpr lacz Df (3L)H99, kni [ri- l]p[p]/TM3,,SblI ] (#15 76) y w prd GALA; Ly/TM3.y+ e Ser wll18. , r y Sb e PL&-3;r y]99B/TM6 Ubx e ~"'~;UCyO;phsC<iSpeR rn TNF ~'"~phsCaSpeR mTNF;KifkzWto/rM3
1 Pl642 hid 1 Bloomington Stock Center
Source Bloomington Stock Center Amen Manoukian, Ontario Cancer Institute Kristin White, MGH John Abram, University of Texas Bloomington Stock Center Amen Manoukian, Ontario Cancer Institute Armen Manoukian, Ontario Cancer Institute Maurice Ennis, Ontario Cancer Institute
. Maurice Ennis, Ontario Cancer Institute W ~ ~ ~ & , phsCaSpeR mTNF;rpr lacz y w prd GALrl;H99/TM3,y+ e Ser
Table 2. Drosophila Stocks.
Maurice Ennis, Ontario Cancer Institute Maurice Ennis, Ontario Cancer Institute

Total #
1 30 sec 1 200 1 '%O 1 infertile
1 min
Wbrown ( lethality
200
Table 3. Observed infertility and lethality of w"18 embryos exposed to UVC. 0-4 hour embryos are collected and exposed to a UVC source for the time indicated. A fixed number of embryos is plated on an agar petri dish in a grid and allowed to age for approximately 36 hours. Of the embryos that fail to develop, the number of white and brown embryos are counted. % infertility is calculated as 100 x (#white/total). The number of white embryos is not considered for the lethality calculation (lethality = # brown/ (total - #white)).

Relat;?p Se- ' P ekment line
Control 1 UVB Relative Staininp
Pekmenf &e '
Control 1 W B
Table 4. Relative &galactosidase staining in Drosophila imagina1 discs before and after exposure to WB. P element lines that showed a significant change in staining level or pattern are indicated in bold. Cornparisons are based on imagina1 discs recovered from 3-5 lawae

1 1642 1 hid
L 1193 1525 1568 1616 1627
Table 5. Observed response of various P elements to UVC exposure. Results of anti B-galactosidase staining on 1-5 hour embryos without or with UVC treatment. Gastrulating embryos are scored for expression of kgalactosidase indicated by antibody staining. Minus signs represent no observed expression while plus signs represent relative levels of observed expression.
pp2A string twins cyclin A Dakt1
+ ++ + + +
+ ++ + + +
\

Figure 1. Schematic diagram showing (A) TNF and (B) UV molecular signaling leading to the activation of apoptosis in mammals. h w h e a d s indicate activating interactions and bar ends indicate inhibitory interactions. C. elegans orth01ogs of speafic components are indicated in brackets. D. melanoguster activators and inhibitors are shown in blue boxes. See introduction text for details.

Figure 2. (A) Schematic representation of the hsTNF construct used to make hsTNF transformant line. The mouse TNFa hsert used here represents bases of the full length mouse TNFa cDNA.
pCaSpeR- Drosophiln the 3' 468 The TNF
polypeptide is translated under control of the upstream hsp70 promoter of the pCaÇpeR-hs vector. (8) Single letter amino acid sequence of the mouse TNFa protein. The polypeptide created by pCaSpeR-hsTNF is indicated in bold.

Figure 3. Expression of TNFa protein after heat shock. Western blotting with anti-mTNFa antibody on embryo protein extracts. 0-4 hour embryos were collected and treated as indicated in materials and methods.
hs TNF
w1118
non-hs non-hs shock shock
30 min after heat
120 min after heat

w 1 1 18 ut hsrpr hs hsTNF hs wl 1 18 UVC
B 01080phh I l i n Treitmint Ii Plited # Brown r W h b Lithrllty Avg. IolhdityStmndird Doviition w l l l 8 untrealed 200 7 22 0.039 0.047 0.014
200 7 10 0.039 200 10 4 0 0.063
hsrpr heatshock 200 1 0 9 3 0.168 0.13 0.052
200 2 3 30 0.1 35 200 11 4 0 0.069
;rprIacz heatshock 200 18 7 5 0.144 0.14 0.0058 200 2 O 57 0.14 200 17 69 0.13
Figure 4. (A) Average embryonic lethality after expression of reaper, TNFa, or exposure to UVC. Embryos are treated after 0-4 hour collections and plated in groups of 200. The average of three separate trials is represented by the columns with the error bars representing the sample standard deviation. (B) Table of actual results. Lethality = #Brown/(# Plat ed - # White).

Figure 5. Embryonic reaper and TNFa expression and W C exposure cause similar cuticle phenotypes. 0-4 hr embryo collections were treated as follows: (A) wn18, untreated; (8) hsreaper, heat shock; (C) hsTNF;reaperlacz, heat shock; and (D) w1I1$, UVC.

Figure 6. Stage 9-10 hsTNF;reaperlacz embryos stained with anti-fl galactosidase antibody to indicate reaper expression pattern. A-C are untreated and show a progression of development from stage 9 to stage 10. D-F are heat shock treated embryos, driving TNF expression displaying some early and ectopic expression of reaper. G-I are W C treated, showing early and significant ectopic expression of reaper staining .

Figure 7. Addine orange stain showing apoptotic ce11 death in Drosophila embryos at 2-6 hours of development. Cell death is visuaüzed b the appearance of red, fluorescent spots in embryos stained w d acridine orange (arrowheads) and viewed under microscope with a rhodamine filter. A and B are kllB embryos which are untreated or W C treated respectively. C represents hsTNF embryos treated with heat shock.

Figure 8. W C cuticle phenotype in H99 embryos. Cuticle preparations made of y;H99/y+ embryos exposed to W C . Embryo colour was determined empiricaily by visualization under the microscope before being photographed. Yellow+ embryos (white in colour) (A) show similar WC-induced phenotype as in WC-treated wildtype embryos. Yellow embryos (yellow in colour) (B), representing H99 /Hg9 genotype, also showed similar phenotype.

Figure 9. &$alactosidase staining of Drosophila larval ha@ discp of P element line 862. Third instar l w a l di- are shown without (A) or after (B) UVB expure. Blue staining shows expandecl region of expression (B) in haguial disc exposed to UVB radiation.



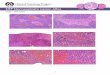






![[Nego] Digests](https://img.pdfslide.net/doc/110x75/5456d645b1af9fda448b47d1/nego-digests-55844f0cb5a76.jpg)


















