Embed Size (px)
Citation preview

Report of the
Tomato Genetics Cooperative
Volume 60 December 2010

2
Report
of the
Tomato Genetics Cooperative
Number 60- December 2010 University of Florida
Gulf Coast Research and Education Center
14625 County Road 672
Wimauma, FL 33598 USA
Foreword
The Tomato Genetics Cooperative, initiated in 1951, is a group of researchers who share and interest in tomato genetics, and who have organized informally for the purpose of exchanging information, germplasm, and genetic stocks. The Report of the Tomato Genetics Cooperative is published annually and contains reports of work in progress by members, announcements and updates on linkage maps and materials available. The research reports include work on diverse topics such as new traits or mutants isolated, new cultivars or germplasm developed, interspecific transfer of traits, studies of gene function or control or tissue culture. Relevant work on the Solanaceous species is encouraged as well. Paid memberships currently stand at approximately 94 (includes those paid in 2009 and beyond) from 16 countries. Cover: Design by Dolly Cummings. Bacterial wilt incited by Ralstonia solanacearum is a serious threat to tomato production in many humid tropical production regions. Breeding for resistance has been a challenge due to multiple strains of the bacterium, variable environmental effects on disease expression, largely unknown genetics for resistance, and linkage of resistance to small fruit size. Locating molecular markers tightly linked to resistance genes should be a boon to future breeding efforts and with the tomato genome now sequenced this research may advance much more rapidly than in the past. This years‟ feature article explores the major sources (“roots”) of bacterial wilt resistance and thereby sheds light on the relationships of genotypes that could be used in studies to locate molecular markers. Precision in the exact identification of sources is hampered by a lack of information or by conflicting information. Unfortunately some of the old sources are no longer available. If you read the article and have information that would help us improve the article please contact us. We live in an electronic age and if we get better information we will put it in the article and future versions may be better than the one published in December of 2010. If you have seed of a rare source line send it to us and we will see that it gets in a gene bank. Thanks for your help.

3
Table of Contents
Foreword 2
Announcements 4
Feature Article
Tomato resistance to bacterial wilt caused by
Ralstonia solanaearum E.F. Smith: ancestry and peculiarities
Daunay M.C., Laterrot H., Scott J.W., Hanson P., Wang J.-F 6
Fig 1: Origins Tomato Bacterial Wilt material 20
Table 1: Summing up of the phenotype of some breeding lines 21
Fig 2-12: Pedigree Montage 30
Research Reports
Preliminary Observations on the Effectiveness of Five
Introgressions for Resistance to Begomoviruses in Tomatoes
Luis Mejía, Rudy E. Teni, Brenda E. García, Ana Cristina Fulladolsa,
and Luis Méndez; Sergio Melgar, and Douglas P. Maxwell 41
Preliminary report on association of ‘Candidatus Liberibacter
solanacearum’ with field grown tomatoes in Guatemala
Luis Mejía, Amilcar Sánchez, and Luis Méndez; D. P. Maxwell;
R. L. Gilberston; V.V. Rivera and G.A. Secor 54
Study of epidermal cell size of petals and stamens in tomato
species and hybrids using confocal laser-scanning microscopy
Christopher Lofty, Julian Smith, Pravda Stoeva-Popova 58
Stock Lists 66
Membership List 101
Author Index 107

4
Announcements From the editor:
Help I‟ve fallen behind and I can‟t catch up! The 2010 TGC is late as a result but it is still 2010 so it could be worse- my apologies for the delay. This is our first year of our “electronic only” (see below) format and we are determining how to proceed. No dues were requested from the members in 2010 as most (but not all) costs are associated with printing and mailing the report. On the web we will post only the Table of Contents for a year and will mail a link to an electronic version of Volume 60 to members who paid in 2009. We can also send a printed version to those who want one and will pay to have one sent. Members will receive an email about this option. The cost would be $20US for domestic members and $25US for foreign members. Make checks payable to The University of Florida from a US bank or a bank with a US affiliation. Sorry no credit cards can be used. If you do not have easy access to a bank with a US affiliation we can accept cash in US dollars. For those who only want the electronic version we will ask for dues of $10 per year starting in 2011. Members will receive an email about this in spring 2011 but send in your dues at any time, either for electronic only or for electronic and printed versions as per the prices stated above.
I have not been happy with the key word search of the TGC Reports available on our website: (http://tgc.ifas.ufl.edu/) as it picks up words in areas outside of the reports such as from the Table of Contents and thus is somewhat messy. We have discussed a way to fix this and hope to have it fixed in 2011.
You can see that there are only 3 research reports in this volume. This epitomizes the trend we have been seeing over the last several years as researchers are not sending in reports. Perhaps this year‟s dearth of reports is due in part to the change to an electronic only format. However, I do see a place for the TGC here in the 21st century and plan to keep moving forward. I hope you will help by retaining your membership or becoming a member if you are not presently one and by sending in reports, varietal pedigrees etc.
Last but certainly not least, my heartfelt thanks to Dolly Cummings who keeps TGC business in order around here. Thanks to Dolly and Christine Cooley who work on the website updates.
My contact information: Jay W. Scott, Ph.D. Gulf Coast Research & Education Center 14625 CR 672 Wimauma, FL 33598 USA Phone: 813-633-4135; Fax: 813-634-0001 Email: [email protected] Jay W. Scott Managing Editor

5
Upcoming meetings: February 17-19, 2011, Sol-Conference 2011 Chiangmai, Thailand http://www.sol-symposium2011.com/abstra.aspx March 20-23, 2011 43rd Tomato Breeders Roundtable * El Cid Castilla Beach Resort Hotel, Mazatlan, Sinaloa,, Mexico. http://tgc.ifas.ufl.edu/2011TBR.htm April 11-14 2011 XVIIth EUCARPIA Meeting - Section Vegetables - Working Group Tomato, Málaga, Spain http://www.eucarpiatomato2011.org October 16-20, 2011 SOL & ICuGl Joint Conference 2011 Tsukuba International Congress Center (EPOCHAL), Tsukuba, Japan http://www.sol2011.jp * Recently cancelled-might be held in conjunction with the Tomato Disease Workshop in October at Cornell University, date not yet set nor is it official yet so check for updates on the TGC website at http://tgc.ifas.ufl.edu .

6
FEATURE ARTICLE TGC REPORT VOLUME 60, 2010 Tomato resistance to bacterial wilt caused by Ralstonia solanaearum E.F. Smith: ancestry and peculiarities
Daunay M.C. (1), Laterrot H. (1bis), Scott J.W. (3), Hanson P. (4), Wang J.-F (4).
(1) INRA, UR 1052, Montfavet, France, (1bis) retiree of INRA, UR 1052 (2) CIRAD, Pôle 3P, Saint Pierre, Réunion Island, France (3) University of Florida, Gulf Coast Research & Education Center, Wimauma,
Florida, USA (4) AVRDC-The World Vegetable Center, Tainan, Taiwan
Summary Several national tomato breeding projects began work on developing varieties
resistant to bacterial wilt over 60 years ago and several varieties created in the 1950s, 1960s, 1970s and later on are still found as reference varieties in many recent publications dealing with the genetics of resistance. From the beginning there were many exchanges of resistant material between the breeding programs that are difficult to retrace because published information is scarce. As a consequence the source(s) of resistance of the reference varieties, and the relationships between these varieties are often unclear. This paper provides a synthesis of the relationships between the breeding carried out in Puerto Rico, the USA (North Carolina, Hawaii, Florida), Japan, the Philippines, the French West Indies, and Taiwan, the main sources of resistance that they used, as well as the parentage between the lines they created. The limits of the reliability of our results are explained. The information on the resistance of many bacterial wilt resistant lines to other vascular diseases is also summarized together with some other peculiarities, in order to provide a synthesis useful for breeding bacterial wilt resistant tomatoes and for further genetic studies of the resistance patterns.
Introduction Bacterial wilt is caused by the pathogen formerly known as Pseudomonas
solanacearum, transitorily renamed Burkholderia solanacearum (Yabuuchi et al., 1992) and presently accepted as Ralstonia solanacearum (Yabuuchi et al., 1995; Vaneechoutte et al., 2004). Developing varieties with resistance has challenged tomato breeders for over 60 years for several reasons. Strong interactions are observed between resistance, environmental conditions and strains (e.g. Kelman, 1953; Acosta, 1963; Krausz & Thurston, 1975; Messiaen, 1989; Peter et al., 1993; Prior et al., 1994; Hanson et al., 1996; Jaunet & Wang, 1999; Balatero et al., 2005; Hai et al., 2008). Further, several defaults are often associated to bacterial wilt resistance, such as small fruit size (Acosta et al., 1964; Gilbert & Tanaka, 1965; Scott et al., 2005), bitterness due to high tomatine content (Borchers & Nevin, 1954; Mohanakumaran et al., 1967; Digat & Derieux, 1968; Messiaen, et al., 1978), green gel around the seeds and epidermis cracking (Acosta, 1963; Cordeil & Digat, 1967; Daly, 1976). The difficulty in developing resistance with adaptation to heat and with good horticultural features, in particular fruit firmness and large size, has been reported by many authors (e.g. Kaan et al., 1969;

7
Opena et al., 1989; Celine et al., 2003; Scott, 1997; Scott et al., 2003). It is also very difficult to combine bacterial wilt resistance to resistance to root knot nematode (Messiaen et al., 1978; Prior et al., 1994; Deberdt et al., 1999 a & b).
Kelman (1953) and Acosta (1963) reported the first surveys of the early screening trials carried out during the first half of the 20th century, in the USA and other countries, with hundreds of tomato varieties. The very limited success obtained at that time outlined the difficulty to identify highly resistant material. The breeding for resistance took on new momentum after World War II. Sources of high levels of resistance were identified and used for breeding. The material most widely used nowadays as; controls, key source(s) of resistance for ongoing breeding programs and/or for genetic studies was created in the span of time running from the 1950s and 1970s. Our main objective was to draw a worldwide historical outline of the major breeding inputs of this period, in order to get a global picture of the key original resistance sources used, of the major breeding lines obtained, and of their relationships. Indeed, as information on these topics is scarce, scattered and confusing, it is useful to sum it up for the sake of present and future research on tomato resistance to bacterial wilt. The major programs were carried out (1) in Puerto Rico, (2) in US Universities (North Carolina, Hawaii, Florida), (3) in the Horticulture Research Station1, Japan, (4) in the University of Philippines College of Agriculture, (5) in the French public institutes INRA2 in Guadeloupe and IRAT3 in Guadeloupe and Martinique (French West Indies), and later on (6) in AVRDC4, Taiwan.
1. Primary historical breeding programs, their sources of resistance and germplasm flow between programs
The main accessions used and created in the major research and breeding programs of the USA, Japan, Philippines, French West Indies and Taiwan, as well as their relationships, are outlined in Fig. 1. For the sake of clarity the accessions listed are limited to the major ones, i.e. those which are most frequently mentioned in the literature for their high level of resistance in worldwide trials, or for which the sources of resistance have been published. We also took care to mention enough accession names for the reader to obtain the encoding system used by the different programs. The arrows linking two accession names indicate their parental link. The arrows starting from the frame of a given program indicate the use of material of this program in another program or for creating a given breeding line. The number(s) in brackets indicate the literature reference where the information about the relationships between accessions and programs is provided. For the convenience of the reader, we summed up in Table 1
1 The name and locations of this institute changed along the time: 1921-1950: Horticulture Research
Station (Okitsu, Shizuoka); 1950-1961: Department of Horticulture, National Institute of Agricultural Sciences (Hiratsuka, Kanagawa); 1961-1973: Horticulture Research Station (Hiratsuka, Kanagawa); 1973-1986: Vegetable and Ornamental Crops Research Station (Tsu, Mie); 1986-2001: National Research Institute of Vegetables, Ornamental plants and Tea (NIVOT) (Tsu, Mie); 2001-present: National Institute of Vegetable and Tea Science (NIVTS) (Tsu, Mie). Dr H. Fukuoka, NIVTS, pers. com. 2 Institut National de la Recherche Agronomique.
3 Institut de Recherche en Agronomie Tropicale (now part of CIRAD, Centre de Coopération
Internationale en Recherche Agronomique pour le Développement). 4 The Asian Vegetables Research and Development Centre (now AVRDC-The World Vegetable Center).

8
available information on the phenotype of the main resistant accessions lines displayed in Figure 1 and issued from the main historical breeding programs. Puerto Rico
Information about tomato breeding for bacterial wilt resistance in the University of Puerto Rico is very scarce and dispersed in annual reports of the Puerto Rico University Agricultural Experiment Station. For instance, Cook (1934, 1935), Roque (1935) and Theis (1950) mention partial resistance of some native material and its use in breeding. Warmke & Cruzado (1949) experimented with local selections from hybrids between native and imported tomato varieties, some of which showing some resistance to bacterial wilt and out-yielding the controls. Azzam (1964) reported the existence of resistance in S. pimpinellifolium as well as the development of breeding lines with some degree of resistance but with unacceptable fruit quality.
At the beginning of the 1960s, at Rio Piedras station, a double cross involving [„Platillo‟,( a native variety) X a S. pimpinellifolium, (of unknown identity)] X [a tomato, (of unknown identity) X „Platillo‟] was made by H. Azzam (Digat & Derieux, 1968; Daly, 1976) and its progenies were used by IRAT in the 1960s and 1970s [see below]. The literature also mentions „Beltville 3814 (=T414)‟ which was said to be from Puerto Rico and was used in North Carolina breeding effort [See below].
North Carolina The search and breeding of bacterial wilt resistant tomatoes began long ago in the
USA, at the turn of the 19th and 20th century in the agricultural experiment stations of several States including North Carolina, Florida, Alabama, and Mississipi (Kelman, 1953). After a lapse of years, efforts were resumed at the North Carolina Experiment Station in 1936 (Schmidt, 1936, 1937) and involved many horticultural scientists such as W.S. Barham, F.D. Cochran, M.E. Gardner, W.R. Henderson, J.S. Weaver, and plant pathologists such as D.E. Ellis, S.F. Jenkins, A. Kelman and N.N. Winstead (Henderson & Jenkins, 1972b). Warmke & Cruzado (1949) as well as Walter (1967) mention the existence of a US Southern Tomato Exchange Program (STEP) that was put into operation in 1946 (Yarnell, 1948), and was complemented with the National Screening Program for evaluation of PI (Plant Introduction) accessions of Lycopersicon for disease resistance. These programs are probably at the origin of the complex relationships between the breeding research carried out in North Carolina, Hawaii, Florida and Puerto Rico for bacterial wilt resistance.
The two widely mentioned sources of resistance of North Carolina breeding material are S. lycopersicum var. cerasiforme „PI 129080‟ (=T 702) from Colombia (initially classified as L. pimpinellifolium -Henderson & Jenkins, 1972b) and (ii) S. lycopersicum var. pyriforme „Beltsville No. 3814‟ (=T 414) (Henderson & Jenkins, 1972a & b; Laterrot et al., 1978; Hanson et al., 1998). „Beltsville No. 3814‟ also named „P.I. No. 3814‟ by Kelman (1953) originated in Puerto Rico according to this author and others (Winstead & Kelman, 1952; Henderson & Jenkins, 1972a & b) without further details. Thurston (1976) also said that it is a selection from Puerto Rico. However the name of this line and the fact that it has a PI number both suggest that „Beltsville No. 3814‟ was associated with the USDA, Beltsville (Maryland). Perhaps USDA researchers were collaborating with Puerto Rican researchers. „T414‟ displayed, as other bacterial wilt

9
resistant lines, a specific bitter taste with lasting burning sensation, and it was used by Borchers & Nevin (1954) for setting up a quantitative chemical test of the alkaloids responsible of this taste. Another source of resistance, S. lycopersicum „Mulua‟ from Guatemala, is mentioned by Winstead & Kelman (1952) and by Suzuki et al. (1964) [who refer to Winstead & Kelman (1952) as well as to a personal communication of D.E. Ellis]. According to Winstead & Kelman (1952), „Mulua‟ yielded resistant breeding material after an initial cross with „Rutgers‟. Suzuki et al. (1964, p.99) still referring to Winstead & Kelman (1952), mention also a „T-141‟ from Puerto Rico as another source of resistance used in North Carolina, though these latter authors mention „T414‟ and not „T-141‟. The identity of „T-141‟ is henceforth doubtful, either a mistyping of „T414‟ by Suzuki et al. (1964) or another line not reported in any other source we found.
To sum up, the cherry tomato „PI 129080‟ (=T 702) from Colombia, the pear shaped tomato „Beltsville No. 3814‟ (=T 414) and the tomato „Mulua‟ from Guatemala, have been included in NC breeding programs as genitors of bacterial wilt resistance.
„Venus‟ and „Saturn‟, released in the early 1970s (Henderson & Jenkins, 1972a & b) are the best known commercial varieties with resistance to bacterial wilt issued from the North Carolina State program. Their pedigrees are provided in Fig. 2, and their phylogenic relationship with two other NC lines, „MR4‟ and „NC 72 TR 4-4‟, is provided in Fig.3.
Hawaii D.C. McGuire, J.C. Gilbert and J.C. Acosta (breeders) as well as I.W. Buddenhagen
(pathologist) are among the scientists having worked on tomato resistance to bacterial wilt in the course of the 1950s and 1960s. Breeding for resistance in commercial type tomatoes was confined first (prior to 1955) to crossing root knot nematode resistant Hawaii lines and North Carolina bacterial wilt tolerant lines (Acosta et al., 1963, 1964). Acosta (1963) indicated that several North Carolina lines which had been bred for bacterial wilt resistance, proved to be intermediate in wilt susceptibility under Hawaiian conditions. In 1953 a new source of resistance, S. pimpinellifolium „PI 127805A‟, originating from Peru, was “obtained” [sic]5 and field selected in Hawaii through 9 generations (Acosta et al., 1964). This accession would be at the origin of the line „5808-2‟ (Mohanakumaran et al., 1967). Acosta (1963) writing the name as „HES 5808-2‟, mentions that it is an inbred line of L. pimpinellifolium obtained by D.C. McGuire, but he does not refer to any PI number. The commercial variety „Kewalo‟ (Fig. 4), developed by Gilbert et al. (1974), recombines the resistance to bacterial wilt originating from „PI 127805A‟ with root-knot nematode resistance(gene Mi) and other useful traits from „Anahu‟, a local tomato, and its derivative „Kalohi‟.
„Hawaii 7996‟ (H7996), „Hawaii 7997‟ (H7997) and „Hawaii 7998‟ (H7998) were later bred by J.C. Gilbert in the 1970s (Scott et al., 2005) and it has been reported in the literature (Hanson et al., 1998; Balatero & Hautea, 2001; Scott et al., 2005) that these lines had resistance derived from PI 127805A. However, correspondence dated October 1983 from J. Tanaka (Assistant Horticulturist at Hawaii University) to J. Scott indicates that the 3 mentioned Hawaiian lines and five others („H7975‟, „H7976‟, „H7981‟, „H7982‟, „H7983‟) are sister lines selected from a initially highly variable
5 The word “obtained” in this sentence is quite imprecise, since it might mean “received from someone” or
“obtained by breeding”.

10
accession named „HSBW‟, which acronym might mean „Hot Set Bacterial Wilt‟. This enigmatic accession, delivered to J.C. Gilbert via an unidentified way and planted first in 1973, displayed a high level of bacterial wilt resistance in hot tropical areas. An earlier correspondence, dated March 1978, from J.C. Gilbert to H. Laterrot (INRA) indicates that „Hawaii 7996‟ could be cited as having been selected at the University of Hawaii for bacterial wilt resistance, and as having its origin somewhere in the Philippines. J.C. Gilbert admitted in this letter that no publications had been specifically written for this line, and that he was not fully satisfied with it because of its flavour due to alkaloid residues in the ripe fruit. He recommended to using it as a rootstock or as a parent to be crossed with another parent of good flavour and some bacterial wilt resistance, for making F1s. Laterrot et al. (1978) also mentioned as a personal communication of Gilbert, a Philippine source at the origin of „Hawaii 7996‟. H. Laterrot recorded in his handwritten notes, based on Kaan personal communication, that „Hawaii 7996‟ was a selection made in „Hotset X Philippine tomato‟ and that it had very small fruits and a determinate growth habit. Given the absence of further written details, it is probably impossible to unravel further the pedigrees of „HSBW‟ and „Hawaii 7996‟.
In conclusion, a close look at the dispersed information relative to the breeding programs of Hawaii University indicates that several sources of resistance have been used successively there, first North Carolina material, then „PI 127805A‟ and lastly Philippines material. It is possible (or probable), that J.C. Gilbert recombined these sources along the time in his breeding material, in one way or another. Breeding is an art as much as a science, and the exact pedigree of the most famous bacterial wilt resistant line „Hawaii 7996‟ will probably remain the secret of the breeding genius of the late J.C. Gilbert.
Florida According to Sonoda et al. (1979) the first attempts to search for bacterial wilt
resistance in Florida started over one century ago, but the breeding efforts took a real momentum in the late 1970s. The original sources of resistance used were „Hawaii 7997‟, S. lycopersicum var. cerasiforme „CRA 66‟ and S. lycopersicum „PI 126408‟. The latter is one of the 28 PI accessions determined as resistant to bacterial wilt out of 909 accessions tested (Barham & Ellis, 1951). However most of the material derived in Florida utilized „Hawaii 7997‟. Attempts to pyramid resistance genes in the early 1980‟s were not successful as there were no molecular markers to identify the genes in resistant plants. No lines were developed that had resistance greater than any of the sources, so there was no evidence that improvements were made under Florida conditions. Over the years it was evident that it was difficult to attain large fruit with high resistance levels. In 1995 „Neptune‟ (Fig.5), a line with larger fruit size than „Hawaii 7997‟, was released, but its resistance level was less than that of „Hawaii 7997‟ (Scott et al., 1995a), and when tested in the world wide test (as Fla. 7421) „Neptune‟ also displayed a much narrower spectrum of resistance (Wang et al., 1998). Breeding efforts then focused on taking lines like „Neptune‟ and crossing them back to „Hawaii 7997‟ to attain large fruited lines with high resistance levels. At first, new lines were developed with moderate fruit size and high resistance. The next crosses were with very large fruited susceptible inbreds. From this work the sister lines „Fla. 8109‟ and „Fla. 8109B‟ were developed (Scott et al., 2003) and further crossing provided new inbreds with high

11
resistance and very large fruit (Scott et al., 2009). Hypothetically, „Fla. 8109‟ and the lines developed thereafter contain a resistance gene, missing in „Neptune‟, that was unlinked from a gene preventing large fruit formation, but this has not been elucidated yet.
Japan In Japan, development of bacterial wilt resistance in tomato (and eggplant) started
as early as 1951 and was based on the use of North Carolina lines. „OTB-1‟ and „OTB-2‟ are self pollinated offspring obtained in the 1950s respectively from „NC1953-60N‟ and „NC1953-64N‟ (Suzuki et al., 1964). According to later reports and papers published [in Japanese] by the Ministry of Agriculture and Forestry (H. Fukuoka6, pers. com.) „OTB-2‟ was segregating for several traits including bacterial wilt resistance (but was fixed for Fusarium wilt resistance) and was submitted to further screening for bacterial wilt resistance and further selfing. In 1969 „BF-Okitsu 101‟ was obtained7 from this process. „OTB1‟ and „OTB2‟ were described in 1964 by IRAT (French West Indies) as possessing the traits of the Puerto Rican S. pimpinellifolium, but with markedly bigger fruits and exceptional fruit productivity, together with a good behaviour towards viruses.
Philippines In Philippines, breeding was established as early as 1954 by T.L. York and J.R.
Deanon who evaluated local and foreign accessions with known resistance to bacterial wilt (Deanon, 1988). The exact origin of the resistance source(s) used in the Philippines breeding scheme is not found in the literature and hence remains confusing (Deanon, 1988; Wang et al., 1998). Empig et al. (1962) report some resistance in Philippine native material, such as „Los Baños native‟ which has possibly been used in the local breeding research. J. Acosta8 conducted research on the inheritance of tomato bacterial wilt resistance at the University of Hawaii (Acosta et al., 1964) and one can hypothesize (i) that he took Philippine material to Hawaii9 and conversely (ii) that he brought material back home (to the Philippines). North Carolina material entered Philippines breeding program according to Mew & Ho (1977). These authors, on the basis of a personal communication of J.R. Deanon, indicate that „Venus‟ (North Carolina line) entered the pedigree of the Philippine line „UPCA1169‟, together with a „CA64-1169‟ of un-mentioned origin (Figure 1). „UPCA1169‟ is itself at the origin of other Philippines material such as „VC8-1-2‟ and „VC9-1‟ (Mew & Ho, 1977; Wang et al., 1998; Scott et al., 2005). The origin(s) of the resistance of other valuable Philippine material, such as „TML46‟, „TML114‟, „R3034‟, or „HSBW‟ mentioned in the Hawaii section is not known.
6 Dr H. Fukuoka, National Institute of Vegetable and Tea Science, Kusawa 360, Ano, Tsu, Mie 514-2392,
Japan. 7 „BF-Okitsu 101‟ was obtained by A. Kotani, T. Kuriyama, H. Shimada-Mochizuki, S. Sakuma, and I.
Suzuki (H. Fukuoka, pers. com.). 8 According to http://www.tofil.ph/awardee_profile.php?id=78, J Acosta won a Rockefeller scholarship to
the University of Hawaii in 1958. This is also indicated in Acosta (1963). 9 This hypothesis is consistent with the information of Gilbert to Laterrot (Laterrot et al., 1978) that a
Philippine source had been used in Hawaii and is at the origin of „Hawaii 7996‟.

12
French West Indies (INRA & IRAT) and Burkina Faso (IRAT) In the French West Indies, the research on tomato bacterial wilt resistance started in
1963 at IRAT, and in 1964 at INRA, with some collaboration between the two institutes. According to Cordeil & Digat (1967) a collection from Rio Pedras station of the
University of Puerto Rico was introduced at IRAT Guadeloupe beginning of 1964. These authors mention a variable tolerance to bacterial wilt of some lines such as „199 PR‟, „Platillo 78‟ and L. pimpinellifolium under local conditions. „199 UPR‟, a derivative from the double cross made at Puerto Rico University [„Platillo‟ X S. pimpinellifolium] X [a tomato of unknown identity X „Platillo‟] -see above-, was chosen for its good tolerance to bacterial wilt (Daly, 1976). After pedigree selection, the F8 lines „199 UPR -39.15‟ and „199 UPR -39.16‟ were obtained by Daly (1976). These lines were described as having small watery and not fleshy fruits, with greenish gel around the seeds, and displaying many concentric cracks (IRAT, 1965; Cordeil & Digat, 1967). Both were crossed with „Floralou‟, a variety of good quantitative and qualitative yield in the conditions of the French West Indies (Cordeil & Digat, 1967; Digat & Derieux, 1968; Daly, 1976). After pedigree selection, the lines „IRAT L3‟ (Daly, 1976; Laterrot et al., 1978) -Fig.6- and „Farako-Ba‟ (D‟Arondel de Haye, unpubl., IRAT, 1974, 1975; Laterrot et al., 1978; Rouamba et al., 1988) were respectively obtained in Martinique and Burkina Faso.
The INRA program in Guadeloupe was based on the use of „CRA66‟. The origin of this line is still controversial. It is given by Digat & Derieux (1968) and Anaïs (pers. com.) as one of the many small fruited tomato ecotypes grown in Guadeloupe at that time, and known there as „tomadoses‟. Digat & Derieux described „CRA66‟ as a vigorous line bearing small, pink, and bitter fruits with resistance to bacterial wilt. However another origin of CRA66 is suggested by Kaan (pers. com.) as being „OTB2‟, because the phenotype of „CRA66‟ is very different from the phenotype of the tomadoses: the plant is more vigorous, the leaves have a spreading leaf growth habit, the flowers display an exerted style, the fruits are fasciated and although of pink colour, they have a larger size than tomadoses fruits. There is a green gel inside the fruit, the taste of which is more acrid and extremely bitter, and its bacterial wilt resistance level is higher. However, Suzuki et al. (1964) reported „OTB1‟ was pink fruited, whereas they described „OTB2‟ as red fruited (but segregating for several traits). As IRAT experimented in Guadeloupe „OTB1‟, „OTB2‟ and many other bacterial wilt resistant lines in the mid- 1960s (IRAT 1964, 1965), it is plausible that valuable material „reached‟ INRA by some path and perhaps under a distorted identity for some reason. If „CRA66‟ = „OTB2‟, then all the French West Indies material would derive indirectly from North Carolina material -see Fig. 1-. Comparison of molecular fingerprinting of „OTB2‟ and „CRA66‟ is necessary for elucidating the identity of these two lines.
Crosses started in the 1960s between „CRA66‟ and the susceptible commercial type „Floradel‟ resulted in the varieties „Cranita‟ (Fig.7), „CRA74‟ & „Carette‟ (Fig. 8 & 9), and „Caraibo‟ (= „Caraibe‟) -Fig. 9- that were respectively released in 1971, 1973, 1975, and 1980 (Anaïs, 1986, Anaïs 1997). „CRA84-26-3‟ and „Caravel‟ are offspring of the cross [„Caraibo‟ X „HC8‟] where „HC8‟ is a heat-tolerant and bacterial wilt resistant line derived from the cross „Hawaii 7996‟ x „Campbell 28‟. Later on „Caraibo‟, „HC8‟ and „Caravel‟ were used as genitors in a recurrent selection for recombining their bacterial wilt

13
resistance and agro-climatic adaptation to French West Indies conditions, together with resistance to Begomoviruses (Ano et al., 2002; Ano et al., 2004).
Taiwan AVRDC started tomato breeding in 1972, and from 1973-1980 emphasized the
development of breeding lines with heat-tolerance and bacterial wilt resistance (Opena et al., 1989). Sources of BW resistance frequently used as parents in AVRDC breeding included varieties „Venus‟ and „Saturn‟ from North Carolina State University and lines „VC 11-3-1-8‟, „VC 8-1-2-7‟, „VC 48-1‟ from the University of the Philippines. Most AVRDC bacterial wilt resistant lines developed in the 1970‟s such as „CL8d-0-7-1‟ (derived from „VC11-1-2-1B‟ x „Venus‟), „CL9-0-0-1-3‟ (derived from „VC11-1-2-1B‟ x „Saturn‟), and „CL11d-0-2-1‟ (derived from „VC9-1-2-9B‟ x „Venus‟) were bred from crosses of these two sources. Many AVRDC lines developed in the late 1970‟s and early 1980‟s such as „CL1131‟, „CL5915‟ (Fig.10), and „CLN65‟ (Fig. 11) arose from complex crosses involving North Carolina, Philippines, or AVRDC lines with BW resistance derived from the above sources. High levels of BW resistance were detected in „L285‟, a small-fruited S. lycopersicum germplasm accession from Taiwan (Chang #1) but this source was not used at AVRDC in breeding because it was thought that its bacterial wilt resistance and small fruit size were closely associated (Opena et al., 1992). In 1985 AVRDC received resistant lines developed in Guadeloupe, including „CRA84-58-1‟ and „CRA84-26-3‟, that combined BW resistance and large fruit size. CRA lines were crossed to heat tolerant and BW resistant AVRDC lines which led to the development of AVRDC lines „CLN1462‟, „CLN1463‟ (Fig.12), „CLN1621‟, „CLN2026‟, and many others.
2. Limit of the reliability of the survey: insufficient accuracy of the information The information found in the literature is often vague and sometimes inconsistent
between publications. The absence of published pedigrees for many of the important varieties resistant to bacterial wilt is a real impediment for ascertaining the original sources of their resistance. Henceforth, Figure 1 is the result of our interpretation of sometimes blurred information as exemplified below.
Names of the accessions Depending on the publications, the accessions used in the various trials or breeding
programs are not exactly named the same way. This is due in some cases to probable renaming such as for „199 PR‟ (Cordeil & Digat, 1967), which is also found as “199” (Digat & Derieux, 1968), „199 UPR‟ (Daly, 1976), and „UPR 199‟ (Kaan et al., 1969). The same situation is encountered for „P.I. No. 3814‟ (Kelman, 1953), also found as „Beltsville #3814‟ (Henderson & Jenkins, 1972a; Hanson et al., 1998), „Beltsville No. 3814‟ (Henderson & Jenkins, 1972b), and „Beltsville 3814‟ (Laterrot et al., 1978). However other variations of names such as for „H7997‟ also found as „H7997S‟ and „H7997L‟, „H7998‟ also found as „H7998S‟ and „H7998M‟, or „CRA66‟ found as „CRA66P‟ and „CRA66S‟ in Wang et al. (1998) and Scott et al. (2005) do not mean further selections but encode only the name of the person who provided the seeds used in the trials. These latter authors mention „BF-Okitsu‟, otherwise found as „BF-Okitsu 101‟ in Laterrot (1999).

14
Despite the word “Okitsu” (location in Japan where breeding was carried out, H. Fukuoka, pers. com.) is present in the names of bacterial wilt resistant „BF-Okitsu 101‟ and of bacterial canker resistant „Okitsu Sozai n°110 (Kuriyama & Kuniyasu, 1974), these accessions should not be confused with each other.
Contradictory data found in the literature Apart from the controversy concerning the identity of „CRA66‟, other contradictory
information is found in the literature. Particularly in the case of „Hawaii 7996‟and the other Hawaii 79## accessions, resistance was said to originate from a Philippines accession according to several personal communications between breeders in the 1970s, but according to Acosta et al. (1964) „PI 127805A‟ and North Carolina material are at the origin of the resistance to bacterial wilt of Hawaiian material. It is not possible to reconcile these conflicting statements other than to say that the various sources of resistance were introduced to the Hawaiian program over time. Therefore we left all options on Figure 1.
Unclear origin of some accessions We found no original information on the origin or pedigree of „Beltsville 3814‟ and
further search of 1960s and 1970s publications of Beltsville USDA research station is needed. Another case concerns the unclear relationship between the Peruvian S. pimpinellifolium „PI 127805‟ collected in 1938 and maintained by the USDA Northeast Regional PI Station and „PI 127805A‟ obtained in 1953 and field selected for resistance through 9 generations in Hawaii according to Acosta et al. (1964). Furthermore, „5808-2‟ was derived from „PI 127805A‟ according to Mohanakumaran et al. (1967) but is given as an inbred line of an anonymous L. pimpinellifolium according to Acosta (1963).
Likeness between accessions Phenotypically „BF-Okitsu‟ is very close to „Hawaii 7998‟ (J. Scott and J. Wang, pers.
obs.), though the published information (Fig.1) does not indicate a closer relationship than the presence of North Carolina material in both their pedigrees. Comparison of their molecular fingerprinting would be worthwhile to clarify their genetic relationship.
3. Sources of resistance and inheritance patterns The global survey of the major breeding research for tomato bacterial wilt resistance
points out that the main sources of resistance used worldwide are perhaps only half a dozen accessions of S. pimpinellifolium („PI 127805A‟), S. lycopersicum var. cerasiforme („PI 129080‟), S. lycopersicum var. pyriforme („Beltsville 3814‟), a progeny from a cross between a S. pimpinellifolium and S. lycopersicum („199 UPR‟), S. lycopersicum („Mulua‟) and the enigmatic Philippine accession used in Hawaii in the 1970s. This number could be extended to seven accessions if one adds „CRA66‟ by assuming it is a Guadeloupe tomadose and not a progeny from „OTB2‟. Whether there are 6 or 7 main sources of resistance, the genetic basis of tomato resistance mechanisms used worldwide for breeding is quite narrow. North Carolina material has been integrated in most of the other breeding programs, in particular those of Hawaii, Japan, Philippines and Taiwan. By combining NC material, or not, with other sources of
10 „Okitsu Sozai n°1‟ resistance to Clavibacter michiganensis (formerly named Corynebacterium michiganense) originates from S. habrochaites (L. hirsutum var.
glabratum) „PI 134418‟.

15
resistance, and breeding in geographical areas where different strains of bacterial wilt are prevalent, the breeders exploited the genetic potentialities at their disposal, and created material resisting a wide range of bacterial wilt strains as exemplified by the results obtained in the worldwide trial carried out by Wang et al. (1998). Indeed, the top nine resistant accessions which had high levels of resistance in almost all 12 locations tested (>90% survival on average) were developed in Hawaii („H7996‟, „H7997 S and L‟, „H7998 S and M‟), Philippines („TML46‟ and „TML114‟, „R3034‟), and Japan („BF-Okitsu‟).
Other sources of resistance in wild tomatoes have been described sporadically in
the literature in accessions of the same species (S. pimpinellifolium, cherry and pear S. lycopersicum) as well as in other wild relatives of tomato (Laterrot & Kaan, 1977; Jaworski et al., 1987; Anaïs, 1997; Mohamed et al., 1997; Carmeille et al. 2006b; Hai et al., 2008). From these results, it seems that resistance to bacterial wilt is not that frequent in tomato germplasm. The high genetic diversity displayed by Ralstonia solanacearum complex (Fegan & Prior, 2005) and the strong interactions between strains and resistant material (Lebeau et al., 2011) suggest that various resistance mechanisms, including strain specific ones, exist in tomato resistant germplasm. Therefore, breeders have some opportunities at their disposal to enlarge the relatively narrow range of resistance sources primarily used so far, and to continue accumulating different mechanisms of resistance in tomato genotypes to obtain better stability of resistance in different environments. However, they might be limited by the fact that some bacterial strains are not controlled by any resistant accession (see section 5. below).
Inheritance studies have focused mostly on F2, F3 and RILs progenies of „Hawaii
7996‟ crossed with the susceptible „WVa700‟ (S. pimpinellifolium). Several QTLs of resistance have been identified, including a major QTL on chromosome 6 effective towards „GMI8217‟, an isolate of race 1 biovar 1 (Thoquet et al. 1996a & b); of „Pss4‟, an isolate of race 1, biovar 3, phylotype 1 (Wang et al., 1998); and „JT516‟, an isolate representative of race 3-phylotype II (Carmeille et al., 2006a). Wang et al. (2000) identified another major QTL of resistance of „Hawaii 7996‟ effective towards „Pss4‟ and located on chromosome 12. Several minor QTLs located on chromosomes 3, 4, 8, some of which having a season dependent expression (Carmeille et al., 2006a) have also been identified. Mejia et al (2009) confirmed the QTLs on chromosome 6 and 12 were associated with resistance in „Hawaii 7996‟ observed in Guatemala field evaluation against local phylotype I strains. Work is ongoing at AVRDC for adding markers to the QTLs regions of „Hawaii 7996‟ associated to resistance to bacterial strains belonging to phylotype I, in order to develop tools for marker assisted selection.
QTLs of resistance of the resistant line „L285‟ effective towards „UW364‟ an isolate of race 1, biovar 4, have also been located on chromosomes 6, as well as on chromosomes 7 and 10 (Danesh et al., 1994). Pattern of resistance derived from CRA66 has been described as polygenic (Prior et al., 1994) but no molecular data are available for this source.

16
4. Resistance to bacterial wilt is sometimes associated to resistance to other
bacterial and fungal pathogens Kaan & Laterrot (1977) were the first to mention a quantitative resistance to
Fusarium wilt (Fusarium oxysporum f.sp. lycopersici) race 2 in lines bred for bacterial wilt resistance in Puerto Rico, North Carolina, and the French West Indies and they suggested this relation to be more likely of a pleiotropic nature than being due to a genetic linkage. Further, Laterrot & Kaan (1978) as well as Laterrot et al. (1978) have pointed out the frequent association of both these partial resistances with the partial resistance to a third vascular disease, i.e. bacterial canker caused by Clavibacter michiganensis. These authors exemplified this relation between the resistance to the three diseases on a set of varieties bred for bacterial wilt in North Carolina („NC 72 TR 4-4‟, „MR4‟, „Venus‟ and „Saturn‟), in the French West Indies („Carette‟, „53 RC‟, „IRAT L3‟), in Burkina Faso („Farako-Ba‟), and in Hawaii („Hawaii 7996‟). They checked that the resistance to Fusarium wilt race 2 observed was not due to the gene I-2. „Kewalo‟ was the only variety resistant to bacterial wilt that they found susceptible to bacterial canker, and of a low level of resistance to Fusarium wilt race 2. This exception suggests that the mechanisms controlling the resistance to bacterial wilt can be dissociated, in some genotypes, from those involved in the resistance to the two other vascular diseases. Unfortunately Laterrot & Kaan (1978) and Laterrot et al. (1978) did not test bacterial wilt resistant material created in the Philippines and Taiwan, and the general picture of the relationships between the resistance to bacterial wilt and the two other vascular diseases is incomplete.
The reciprocal relationship between the resistance to bacterial canker and the two other diseases is verified in some cases, as for instance for „Okitsu Sozai n°1‟ whose resistance to bacterial canker originates from S. hirsutum var. glabratum PI 134418. This line is also partially resistant to Fusarium wilt race 2 (Laterrot, unpub. results) as well as to some strains of Ralstonia solanacearum (Lebeau et al., 2011). But „Plovdiv 8-12‟, the resistance of which to bacterial canker originates from a S. pimpinellifolium, is susceptible to Fusarium wilt race 2 (Laterrot et al. 1978); its behaviour towards bacterial wilt is unknown.
The bacterial wilt resistant „Hawaii 7998‟ was resistant to a race of bacterial spot
later confirmed to be race T1 (Scott & Jones 1986). Later, Scott et al (1995b) discovered „Hawaii 7981‟, a line susceptible to race T1, was resistant to bacterial spot race T3 while „Hawaii 7998‟ was susceptible. Thus, bacterial spot resistance has been found in bacterial wilt resistant lines from Hawaii. For race T1, resistance to bacterial wilt was not correlated with bacterial spot resistance in an F2 population suggesting separate genes were responsible for resistance to each disease (Scott et al., 1988). Resistance to bacterial spot race T4 has been seen in some bacterial wilt resistant breeding lines derived from „Hawaii 7997‟ even though this line is not resistant to race T4 (Scott et al., 2010). The genetic control of this response is not known but again illustrates the association of bacterial wilt resistance with resistance to another disease. A further example is that „Hawaii 7998‟ had resistance to bacterial canker (Panthee and Gardner, 2010) especially the foliar phase. These examples and those mentioned before indicate the existence of frequent associations between the resistance to some

17
bacteria (vascular or not) and even fungi (vascular) within single tomato genotypes. Hence, Scott (1997) advised that when searching for resistance to a given bacterial pathogen, breeders should not overlook genotypes resistant to other bacteria. In this regard, resistance genes can emerge by combining genotypes with some reported bacterial resistance even when one of the parent lines does not show resistance to a particular disease/race. For instance, Hutton et al. (2010) reported that an allele from „Hawaii 7998‟ was associated with resistance to bacterial spot race T4 even though this line was susceptible to race T4, suggesting epistasis with genes from the other parent that was T4 resistant. Another possibility is that the pleiotropic nature of resistance in tomato against several diseases could be associated with higher level of expression of systemic acquired resistance (SAR). Lin et al (2004) found over-expression of Arabidopsis NPR1 (non-expresser of PR genes) gene in a susceptible tomato line could enhance resistance to both Fusarium wilt race 2, bacterial wilt, as well as bacterial spot and gray leaf spot. In this study, they did not test the enhanced resistance against the bacterial canker pathogen.
Lastly, Rouamba et al. (1988) tested material resistant to bacterial wilt towards a
fourth vascular disease, the Verticillium wilt caused by Verticillium race 2, but they found only a loose relationship between both resistances since only two („IRAT L3‟ and „Farako-Ba‟) out of ten lines tested, were resistant to both diseases.
5. Grafting of susceptible cultivars on resistant rootstocks, an alternative to resistant cultivars
Given the difficulty to create highly resistant lines with good commercial quality, grafting susceptible scions on resistant rootstocks remains an alternative to the cultivation of resistant cultivars. As early as 1969, Gilbert and Chin pointed out that highly resistant tomato lines with poor fruit quality, could be efficiently used as rootstocks on which susceptible scions of good fruit quality could be grafted. These authors reported the bacterial wilt resistance of the root system as being effective even when completely susceptible scions are used. This technique is still used nowadays (Cardoso et al., 2006; Wang et al., 2009), though the protection provided by the rootstock is sometimes incomplete (Nakaho et al., 2004). Indeed, tomato resistant material harbours the bacteria symptomlessly and the resistance is associated with the ability of the plant to restrict bacteria invasiveness (Grimault et al., 1993). The absence of incompatible interactions in tomato resistant lines (no symptoms, no latent infection) has been confirmed by Lebeau et al. (2011) by testing a core collection of bacterial wilt resistant accessions with a core collection of bacterial strains.
Eggplant is an alternative rootstock for cultivating susceptible tomatoes in contaminated conditions. It was shown to provide a better protection than tomato rootstocks (AVRDC, 1998). This result was confirmed and extended by Lebeau et al. (2011) who found that apart from common cases of latent infection for some eggplant accessions and all tomato accessions, there also exist incompatible interactions between some eggplant resistant lines and some bacterial strains. Further, some eggplant lines control bacterial strains that are not controlled by any of the tomato resistant lines tested so far, as exemplified by Carmeille et al. (2006b), Wicker et al. (2007) and Lebeau et al. (2011).

18
Conclusion The earliest breeding efforts for tomato resistance to bacterial wilt started in Puerto
Rico and two American States (North Carolina, Hawaii) 80 years ago followed by programs in the 1950s in Japan and the Philippines. The programs carried out by French research institutes in the Caribbean (and Burkina Faso) started during the course of the 1960s, whereas AVRDC started at the beginning of the 1970s. There were many exchanges of material between the breeding programs for bacterial wilt resistance carried out in several US States, and between them and the Philippines. AVRDC is using and recombining now the resistances bred in the USA, the Philippines and the French West Indies. The breeding material created in Japan was mostly used locally, and perhaps also in Guadeloupe, if one assumes that the Japanese line „OTB2‟ is equivalent to „CRA66‟, which is not certain.
Our attempt to draw a general picture of the main sources of resistance to bacterial wilt, of the main breeding programs for this resistance, of their most frequently mentioned resistant varieties, and of the relationships between varieties, is based on a careful work of assembling bits and pieces dispersed in many publications. Given the unavailability of complete information, it is not now possible to come up with a better picture than the one we present here, though more information can emerge out of archives of the scientists, Universities and research institutes involved. Henceforth the synthesis provided here displays the most probable general picture, but it may include some mistakes in the absence of further information.
We added to this survey complementary information on some peculiarities of tomato bacterial wilt resistance. The frequent association of bacterial wilt resistance with resistance to other bacterial or fungal diseases should be of strong interest for breeding and/or research on its genetic basis. Breeding over many decades succeeded in eliminating a number of undesirable traits initially associated to high level of bacterial wilt resistance, but breeding efforts are still ongoing for obtaining large fruit size in resistant material. The adaptation of the breeding material to hot environmental conditions is often mentioned as necessary for obtaining breeding lines which are resistant and agronomically acceptable.
On the whole, we hope this paper to be useful for further research using the plant material mentioned, in particular for comparative genetic studies and breeding concerning the resistance of tomato to bacterial wilt and to other vascular diseases. Acknowledgments The authors are very grateful to Ph. Prior and E. Wicker (phyto-bacteriologists at CIRAD La Réunion Island, Mascarenes) for having motivated the authors to write this historical review, to F. Kaan and G. Anaïs (retirees of INRA Guadeloupe) for having provided precious complements of information, to H. Fukuoka (National Institute of Vegetables & Tea Science, Tsu, Mie, Japan) for having fully clarified the relationships between former Japanese scientists, former Japanese Institutes and tomato lines and for having translated key passages of Suzuki et al. (1964). We thank also the consortium of private companies (Vilmorin, Gautier Semences, DeRuiter Seeds, Enza Zaden, Nunhems, Rijk Zwaan) who financed (2007-2010) research based on the present review. Last but not least, we acknowledge Ch. Olivier (librarian of INRA GAFL, Montfavet, France), M.L.

19
Abinne (librarian at INRA-CRAAG, Guadeloupe),and the librarians at North Carolina State University, the University of Hawaii, AVRDC, and the University of Puerto Rico along with Linda Wessel-Beaver for having provided reprints of numerous archives. Acronyms found in some names of tomato lines or in related literature UPR: University of Puerto Rico NCSU: North Carolina State University UPLB = UPCA: University of Philippines Los Banos = University of Philippines College of Agriculture CRA: Centre de Recherche Agronomique des Antilles (INRA)

20
Figure 1: General scheme of the relationships between worldwide programs and lines. Notations in color correspond to Literature Cited starting on page 23.

21
Table 1. Summing up of the phenotype of some breeding lines created or used in the breeding programmes of North Carolina, Hawaii, Florida, Japan, Philippines, French West Indies, and Taiwan.
origin
line or accession name growth habit fruit shape fruit size
fruit colour source
North Carolina University NC 72 TR 4-4 indeterminate
slightly flattened 60-100 g red
Laterrot et al. (1978); INRA germplasm database
North Carolina University MR4 indeterminate
Laterrot et al. (1978)
North Carolina University NC1953-60N 6,5 g
Suzuki et al. (1964)
North Carolina University NC19/53-64N 7,6 g
Suzuki et al. (1964)
North Carolina University Saturn indeterminate deep globe 100-140 g red
Kaan et al. (1975); INRA germplasm database; Henderson & Jenkins (1972)
North Carolina University Venus indeterminate slightly oblate > 180 g red
Laterrot et al. (1978); INRA germplasm database; Henderson & Jenkins (1972)
Hawaii University HES 5808-2 indeterminate 15g
Hawaii University H7996 determinate small oblate 20-60g red
INRA germplasm database, Scott. pers. com.
Hawaii University H7997 indeterminate small oblate 30-80 g red
Wang et al. (1998), Scott et al. (2005), Scott. pers. com.
Hawaii University H7998 indeterminate small oblate 30 g red
Wang et al. (1998), Scott et al. (2005), Scott. pers. com.
Hawaii University Kewalo determinate flattened 140-180 g red
INRA germplasm database
University of Florida Neptune determinate 123-136g red
Scott et al. (1995a)
Hort. Res. Sta.,Japan BF-Okitsu indeterminate 15-20 g red
Wang et al. (1998)
Hort. Res. Sta.,Japan OTB1 13,8 g pink
Suzuki et al. (1964)
Hort. Res. Sta.,Japan OTB2 30,6 g red
Suzuki et al. (1964)

22
Philippines University UPCA1169 determinate 20-30 g Source ?
Philippines University CA-64-1169
Philippines University VC8-1-2
Philippines University VC9-1
Philippines University TML46 determinate oblate/oblong 30 g red/pink
Wang et al. (1998)
Philippines University TML114 determinate oblate/oblong 40 g red/pink
Wang et al. (1998)
Philippines University R3034
semi determinate deep oblate 30-60 g red
Wang et al. (1998)
INRA, Guadeloupe CRA66 indeterminate
slightly flattened 20-60 g pink
INRA germplasm database. This line is recorded as red fruited in Wang et al. (1998); Scott et al. (2005)
INRA, Guadeloupe Cranita indeterminate pink
Messiaen et al.(1978); Laterrot
INRA, Guadeloupe CRA74 indeterminate wide, deep medium
Kaan et al. (1975)
INRA, Guadeloupe Carette indeterminate
slightly flattened 100-140 g red
Laterrot et al. (1978); INRA germplasm database
INRA, Guadeloupe
Caraibo = Caraibe determinate
flattened / oblate
140-180 g / 150 g red
Anais (1986); Ano et al.(2004); INRA germplasm database
INRA, Guadeloupe Caravel determinate oblate 150-300 g red
Wang et al. (1998)
INRA, Guadeloupe CRA84-26-3 determinate
Hanson et al. (1996)
IRAT (Guadeloupe & Martinique) IRAT L3 indeterminate round 45 g red
(Laterrot et al., 1978); INRA germplasm database; Denoyés (1988)
IRAT (Burkina Faso) Farako-Ba indeterminate round 140-180 g red
INRA germplasm database
AVRDC, Taiwan CL5915 determinate oblong 50 g red
Wang et al. (1998)
AVRDC, Taiwan CLN65 determinate oblate 70 g red
Wang et al. (1998)
AVRDC, Taiwan CLN1463 indeterminate globe 150-200 g red
Wang et al. (1998)
AVRDC, Taiwan L285 indeterminate plum 30 g red
Wang et al. (1998)

23
Literature cited
1. Acosta J.C., 1963. Genetic analysis of bacterial wilt resistance and certain other characters in a tomato cross Lycopersicon esculentum Mill. and L. pimpinellifolium Mill. PhD Thesis, University of Hawaii (Agriculture, Plant Pathology). On line accessible at: http://scholarspace.manoa.hawaii.edu/bitstream/10125/11673/2/uhm_phd_6402645_r.pdf
2. Acosta, J., Gilbert, J., and Quinon, V.L., 1964. Heritability of bacterial wilt resistance in tomato. Proceedings of the American Society for Horticultural Science 84:455-461.
3. Anaïs G., 1986. Utilisation de la résistance variétale dans la lutte contre le flétrissement bactérien de la tomate Pseudomonas solanacearum E.F. Smith. Bull. Tech. D‟Information 409/411 : 449-452.
4. Anaïs G., 1997. La tomate. Pp 591-605 In : L‟Amélioration des Plantes tropicales. A. Charrier, M. Jacquot, S. Hamon, D. Nicolas (Sc. Eds), Ed. CIRAD, ORSTOM..
5. Anais G., pers. com. 2010. 6. Ano G., Anaïs G., Marival P., 2002. Création de variétés de tomate résistantes à
Ralstonia solanacearum et PYMV, adaptées aux régions tropicales. Proceedings of the XXXVIIIth Meeting of the Carribean Food Crops Society, Martinique : 234-238.
7. Ano G., Anaïs G., Marival P., Chidiac A., 2004. L‟amélioration de la tomate pour les régions tropicales de plaine: travaux en Guadeloupe. Les familles CRAPY associent les résistances aux Bégomovirus PYMV et TYLCV à la résistance au Ralstonia solanacearum (race 1). Phytoma, la défense des végétaux 573 : 23-25.
8. AVRDC Report 1998. Asian Vegetables Research and Development Center, Taiwan.
9. Azzam H., 1964. Tomato breeding for the tropics. Proc. of the 2nd Annual Meeting of the Caribbean Food Crops Society, Bridgetown, Barbados 2: 56-59.
10. Balatero, C. H., and Hautea, D. M. 2001. Identification of AFLP and RGA markers associated with bacterial wilt resistance QTL derived from tomato Lycopersicon pimpinellifolium. p.225-243 in Solanaceae V: advances in taxonomy and utilization, edited by R. G. van den Berg, G. W. M. Barendse, G. M. van der Weerden and C. Mariani. Nijmegen University Press.
11. Balatero C.H., Hautea D.M., Narciso J.O., Hanson P.M., 2005. QTL mapping for bacterial wilt resistance in Hawaii 7996 using AFLP, RGA and SSR markers. Pp 301-308 In: Bacterial wilt: the disease and the Ralstonia solanacearum complex. C. Allen, P. Prior, A.C. Hayward (Sc. Eds), American Phytopathological Society (APS), St Paul, USA.
12. Barham W.S. Ellis D.E., 1951. Sources of resistance to late blight and bacterial wilt. Rept. Tomato Genet. Coop. 1: 2.
13. Borchers E.A.,Nevin C.S., 1954. Quantitative estimation of a bitter principle in tomato. Proc. Amer. Soc. Hort. Sci. 63:420-426.
14. Cardoso, S. C.; Soares, A. C. F.; Brito, A. dos S.; Carvalho, L. A. de; Ledo, C. A. da S., 2006. Potential of Hawaii 7996 hybrid as rootstock for tomato cultivars. Bragantia 65(1): 89-96.

24
15. Carmeille, A., Caranta, C., Dintinger, J., Prior, P., Luisetti, J., and Besse, P. 2006a. Identification of QTLs for Ralstonia solanacearum race 3-phylotype II resistance in tomato. Theoretical and Applied Genetics 113 (1):110-121.
16. Carmeille A., Prior P., Kodja H., Chiroleu F., Luisetti J., Besse P., 2006b. Evaluation of resistance to race 3, Biovar 2 of Ralstonia solanacearum in tomato germplasm. J. Phytopathology 154: 398-402.
17. Celine V.A., Chandrmony D., Gokulapalan C., Rajamony L., 2003. Heat tolerance and bacterial wilt resistance of tomato genotypes in the humid tropics of Kerala. Rept. Tomato Genet. Coop. 53: 11:13.
18. Cook M.T., 1934. Annual report of the division of botany and plant pathology (tomatoes). In Puerto Rico Univ. Agric. Expt. Sta. Ann. Rpt. (1933/1934): 131-132.
19. Cook M.T., 1935. Annual report of the division of botany and plant pathology (wilt disease of tomato). In Puerto Rico Univ. Agric. Expt. Sta. Ann. Rpt. (1934/1935): 25-26.
20. Cordeil J., Digat B., 1967. Etude de la résistance variétale de la tomate au flétrissement bactérien en Guadeloupe et en Guyane française. Proc. of the 5th Annual Meeting of the Caribbean Food Crop Society, Paramarimbo, Surinam, 5: 91-98.
21. Daly, P. 1976. "IRAT L3" une nouvelle variété de tomate combinant plusieurs résistance aux maladies. Agronomie Tropicale 31 (4):398-402.
22. Danesh D., Aarons S., Mcgill G.E., Young N.D., 1994. Genetic dissection of oligogenic resistance to bacterial wilt in tomato. Molecular Plant Microbe Interaction 7: 464-471.
23. Daunay M.C., 1977. Une résistance de la tomate à trois maladies vasculaires. Mémoire de fin d‟Etudes, Ecole Nationale des Ingénieurs des Techniques Horticoles, Angers, FRA. Ed. INRA. 72 pp + annexes.
24. Deanon, J. R. Jr. 1988. Biochemical bases of screening tomato for bacterial wilt resistance. p.83-99 In: Current problems on fruits and vegetables. Philippines Council for Agriculture Forestry and Natural Resources Research and Development, Los Banos, Laguna, Philippines.
25. Deanon, Pers. com to Mew & Ho, 1977. 26. Deberdt P., Olivier J., Thoquet P., Grimsley N., Prior P., 1999a. Quantitative
resistance loci to bacterial wilt located on tomato chromosome 6 in near isogenic lines for the Mi gene. Proc. of the First Australasian Soilborne Disease Symp., R.C. Magarey (Ed), Bureau of Sugar Experiment Stations, Brisbane, Australia: 93-94.
27. Deberdt, P., Quénéhervé, P., Darrasse, A., and Prior, P. 1999b. Increased susceptibility to bacterial wilt in tomatoes by nematode galling and the role of the Mi gene in resistance to nematodes and bacterial wilt. Plant Pathology 48:408-414.
28. Digat, B., and M., Derieux. 1968. A study of the varietal resistance to bacterial wilt. p.95-101 in Proceedings of the Caribbean Food Crops Society, 6th Annual meeting, St Augustine, Trinidad.
29. Empig L.T., Calub A.G., Katigbak M.M., Deanon J.R., 1962. Screening tomato, eggplant, and pepper varieties and strains for bacterial wilt (P. solanacearum E.F.S.) resistance. Philippine Agriculturist (46): 303-314.
30. Fegan M., Prior P., 2005. How complex is the “Ralstonia solanacearum species complex”. In: Bacterial Wilt: the Disease and the Ralstonia solanacearum species

25
complex. C. Allen, P. Prior and C. Hayward (Eds), APS Press, St. Paul, Minnesota, USA , 449-462.
31. Gilbert J.C., Chinn J.T., 1969. Bacterial wilt resistant combinations. Rept. Tomato Genet. Coop. 19: 11.
32. Gilbert J.C., Tanaka J.S., 1965. Horticultural refinement of multiple disease resistant tomatoes in Hawaii. Hawaii Farm Science 14(1): 4-6.
33. Gilbert J.C., Tanaka J.S., Takeda K.Y., 1974. „Kewalo‟ tomato. Hortscience 9: 481-482.
34. Grimault V., Schmit J., Prior P., 1993. Some characteristics involved in bacterial wilt (Pseudomonas solanacearum) resistance in tomato. p.112-119 In: Bacterial wilt. ACIAR proceedings No. 45, edited by G. L. Hartman and A. C. Hayward. ACIAR, Canberra.
35. Hai T.T.H., Esch E, Wang J.F., 2008. Resistance to Taiwanese race 1 strains of Ralstonia solanacearum in wild tomato germplasm. European Journal of Plant Pathology 122 (4): 471-479.
36. Hanson P., 2010. pers. com. 37. Hanson, P. M., Licardo, O., Hanudin, Wang, J. F., and Chen, J. T. 1998. Diallel
analysis of bacterial wilt resistance in tomato derived from different sources. Plant Disease 82:74-78.
38. Hanson, P. M., Wang, J. F., Licardo, O., Hanudin, Mah, S. Y., Hartman, G. L., Lin, Y. C., and Chen, J. T. 1996. Variable reactions of tomato lines to bacterial wilt evaluated at several locations in Southeast Asia. HortScience 31:143-146.
39. Henderson W.R., Jenkins S.F., 1972a. „Venus‟ and „Saturn‟, tomato varieties resistant to Southern bacterial wilt. Hortscience 7: 346.
40. Henderson W.R., Jenkins S.F., 1972b. „Venus‟ and „Saturn‟, two new tomato varieties combining desirable horticultural features with southern bacterial wilt resistance. Bulletin 444 of Agricultural Experiment Station, North Carolina State University (Raleigh), July 1972.
41. Hutton, Samuel F., Jay W. Scott, Yang, Wencai, Sim, Sung-Chur, Francis, David M., and Jones, Jeffrey B. 2010. Identification of QTL Associated with Resistance to Bacterial Spot Race T4 in Tomato. Theor. Appl. Genet. 121 (7):1275-1287.
42. IRAT, 1964. Annual Report, Guadeloupe Martinique. Ed. IRAT, p. 433. 43. IRAT, 1965. Annual Report, Guadeloupe. Ed. IRAT, p. 563. 44. IRAT, 1974. Annual Report, Martinique. Ed. IRAT, p.134. 45. IRAT, 1975. Annual Report, Martinique. Ed. IRAT, p.141. 46. Jaunet, T. X., and Wang, J. F. 1999. Variation in genotype and aggressiveness of
Ralstonia solanacearum race 1 isolated from tomato in Taiwan. Phytopathology 89 (4):320-327.
47. Jaworski C.A., Phatak S.C., Ghate S.R., Gitaitis R.D., Widrlechner M.P., 1987. GA 1565-2-4 BWT, GA 219-1-2 BWT, GA 1095-1-4 BWT, and GA 1405-1-2 BWT Bacterial Wilt Tolerant Tomato. HortScience 22(2): 324-325.
48. Kaan F., 1976. Mémoire sur les travaux réalises. CRAAG, Station d‟Amélioration des Plantes, Guadeloupe. Doc. Interne.
49. Kaan F., Beramis M., Messiaen, C.M., 1969. Recherche de variétés de tomates aux Antilles. Proceed. VIIth Ann. Meeting Caribb. Food Crops Society, 7 : 173-181.

26
50. Kaan F., Laterrot H., 1977. Mise en évidence de la relation entre des résistances de la tomate à deux maladies vasculaires: le flétrissement bactérien (Pseudomonas solanacearum E.F. Sm.) et la Fusariose pathotype 2 (Fusarium oxysporum f.sp. lycopersici (Sacc.) Snyd. & Hans). Ann. Amélior. Plantes 27(I) : 25-34.
51. Kaan F., Laterrot H., Anaïs G., 1975. Etude de 100 variétés de tomate en fonction de l‟adaptation climatique et de la résistance à sept maladies sévissant aux Antilles. Nouv. Agron. Antilles-Guyane, (1,2) : 123-138.
52. Kelman, A., 1953. The bacterial wilt caused by Pseudomonas solanacearum. A literature review and bibliography. North Carolina Agricultural Experiment Station, Tech. Bul. N°99: 194 pp.
53. Krausz J.P., Thurston H.D., 1975. Breakdown of resistance to Pseudomonas solanacearum in tomato. Phytopathology 65: 1272-1274.
54. Kuriyama T., & Kuniyasu K, 1974. Studies on the breeding of disease resistant tomato by interspecific hybridization. III. On the breeding of a new tomato resistant to bacterial canker caused by Corynebacterium michiganense. Bull. of the Veg. and Ornament. Crops Research Station A, 1: 93-107.
55. Laterrot, H., Kaan, J.F. 1978. Resistance to Corynebacterium michiganense of lines bred for resistance to Pseudomonas solanacearum. Report of the Tomato Genetics Cooperative 28.
56. Laterrot, H., Brand, R., Daunay, M. C. 1978. La résistance à Corynebacterium michiganense chez la tomate. Annales Amélioration des Plantes 28 (5):579-591.
57. Lebeau A., Daunay M.C., Frary A., Palloix A., Wang J.F., Dintinger J., Chiroleu F., Wicker E., Prior P., 2011. Bacterial wilt resistance in tomato, eggplant and pepper: genetic resources challenged with the multifaceted Ralstonia solanacearum species complex. Phytopathology 101(1): 154-165.
58. Lin, W.-C., Liu, C.-F., Wu, J.-W., Cheng, M.-L., Lin, Y.-M., Yang, N.-S., Black, L., Green, S.K., Wang, J.-F., and Cheng, C.-P. 2004. Transgenic Tomato Plants Expressing the Arabidopsis NPR1 Gene Confer Enhanced Resistance to A Spectrum of Fungal and Bacterial Diseases. Transgenic Research 13:567-581.
59. Mangin, B., Thoquet, P., Olivier, J., and Grimsley, N. H. 1999. Temporal and multiple quantitative trait loci analyses of resistance to bacterial wilt in tomato permit the resolution of linked loci. Genetics 151:1165-1172.
60. Mejía, L., Garcia, B. E., Fulladolsa, A. C., Ewert, E. R., Wang, J.-F., Scott, J. W., Allen, C. and Maxwell, D. P. 2009. Evaluation of recombinant inbred lines for resistance to Ralstonia solanacearum in Guatemala and preliminary data on PCR-based tagging of introgressions associated with bacterial wilt-resistant Line, Hawaii 7996. Rept. Tomato Genet. Coop. 59:32-4.
61. Messiaen C.M., 1989. Environmental influences on the severity of tomato bacterial wilt in the French West Indies: Interactions with varietal resistance. Pp 235-238 In: Tomato and Pepper Production in the Tropics. Proceed. Internat. Symp. on Integrated Management Practices; S.K. Green (Sc. Ed.), T.D. Griggs & B.T. Mc Lean (Publ. Eds), AVRDC, Shanhua, Taiwan.
62. Messiaen, C. M., Laterrot, H., and Kaan, F. 1978. Cumulate resistances to Pseudomonas solanacearum and to Meloidogyne icognita with determinate growth in tomato. p.48-51 in Vegetables for the hot humid tropics. Mayaguez Institute of Tropical Agriculture, Puerto Rico

27
63. Mew T.W., Ho W.C., 1977. Effect of soil temperature on resistance of tomato cultivars to bacterial wilt. Phytopathology 67: 909-911.
64. Mohamed M.El.S., Umaharan P., Phelps R.H., 1997. Genetic nature of bacterial wilt resistance in tomato (Lycopersicon esculentum Mill.) accession LA 1421. Euphytica 96: 323-326.
65. Mohanakumaran N., Gilbert J.C., Young R.L., 1967. Bacterial wilt resistant tomato lines with unusually high content of the alkaloid tomatin. Rept. Tomato Genet. Coop. 17: 41.
66. Nakaho K., Inoue H., Takayama T., Miyagawa H., 2004. Distribution and multiplication of Ralstonia solanacearum in tomato plants with resistance derived from different origins. J. Gen. Plant Pathol. 70: 115-119.
67. Opena R.T., Hartman G.L., Chen J.T., Yang C.H. 1992. Breeding for bacterial wilt resistance in tropical tomato. p.44-50 in: Proceedings of the 3rd international conference on plant protection in the tropics. Malayasian Plant Protection Society, Kuala Lumpur.
68. Opena R.T., Green S.K., Talekar N.S., Chen J.T., 1989. Genetic improvement of tomato adaptability to the tropics: progress and future prospects. Pp 71-85 In: Tomato and Pepper Production in the Tropics. Proceedings of the International Symposium on Integrated Management Practices, S.K. Green (Sc. Ed.), T.D. Griggs & B.T. Mc Lean (Publ. Eds), AVRDC, Shanhua, Taiwan.
69. Panthee, D and Gardner R.G. 2010. Identification of useful sources of bacterial wilt resistance in tomato: A challenge. Proc. 25th Annual Tomato Disease Workshop. P. 18[Abstr.]
70. Peter, K. V., Gopalakrishnan, T. R., Rajan, S., and Sadhan Kumar, P. G. 1993. Breeding for resistance to bacterial wilt in tomato, eggplant and pepper. p.183-190 in Bacterial wilt. ACIAR proceedings No. 45, edited by G. L. Hartman and A. C. Hayward. ACIAR, Canberra.
71. Prior P., Grimault V., Schmit J., 1994. Resistance to bacterial wilt (Pseudomonas solanacearum) in tomato: present status and prospects. In: Bacterial Wilt, the disease and its causative agent Pseudomonas solanacearum, A.C. Hayward & G.L. Hartman (eds.), Wallingford (UK), CAB International : 209-223.
72. Roque A., 1935. Annual Report of the Assistant Pathologist (wilt disease of tomatoes). Puerto Rico Univ. Agr. Expt. Sta., Ann. Rpt. (1934-1935): 31-33.
73. Rouamba A., Laterrot H., Moretti A., 1988. A case of relation between resistances to Pseudomonas solanacearum and Verticillum pathotype 2. Rept. Tomato Genet. Coop. 38: p.43.
74. Schmidt R., 1936. Tomato breeding. N.C. Agri. Expt. Sta. Ann. Rep. 59: 68. 75. Schmidt R., 1937. Tomato breeding. N.C. Agri. Expt. Sta. Ann. Rep. 60: 55-56. 76. Scott, J.W. 1997. Tomato improvement for bacterial disease resistance for the
tropics: A contemporary basis and future prospects. In: Proc. First International Conference on Processing Tomato/First International Symposium on Tropical Tomato Diseases, Recife, Brazil, ASHS Press, Alexandria, VA USA. p. 117-123.
77. Scott, J.W., Jones J.B., 1986. Sources of resistance to bacterial spot (Xanthomonas campestris pv. vesicatoria (Doidge) Dye) in tomato. HortScience 21(2):304-306.

28
78. Scott, J.W., Jones J.B., Somodi G.C., 2003. Development of a large fruited tomato with a high level of resistance to bacterial wilt (Ralstonia solanacearum). Rept. Tomato Genet. Coop 53:36-37.
79. Scott, J.W., Jones J.B., Somodi G.C., Chellemi D.O., Olson S.M., 1995a. „Neptune‟, a heat-tolerant, bacterial wilt-tolerant tomato. HortScience 30(3):641-642.
80. Scott, J.W., Jones J.B., Somodi G.C., Stall R.E., 1995b. Screening tomato accessions for resistance to Xanthomonas campestris pv. vesicatoria, race T3. HortScience 30(3):579-581.
81. Scott, J.W., Jones J. B., Vallad G. E., 2010. Breeding for Bacterial Wilt Resistance in Tomato: The Struggle Continues. Proc. 25th Annual Tomato Disease Workshop. P.19[Abstr.]
82. Scott J.W., Somodi G.C., Jones J.B., 1988. Bacterial spot resistance is not associated with bacterial wilt resistance in tomato. Proc. Fla. State Hort. Soc. 101: 390-392.
83 Scott, J.W., G.E. Vallad, and J.B. Jones. 2009. High level of resistance to bacterial wilt (Ralstonia solanacearum) obtained in large-fruited tomato breeding lines derived from Hawaii 7997. Acta Horticulturae 808:269-274.
84. Scott, J.W., J.F. Wang, and P.M. Hanson. 2005. Breeding tomatoes for resistance to bacterial wilt, a global view. In: Proceeding of the First International Symposium on Tomato Diseases, Orlando, Florida, USA. Acta Hort. (ISHS) 695:161-172.
85. Sonoda R.M., Augustine J.J., Volin R.B., 1979. Bacterial wilt of tomato in Florida: history, status, and sources of resistance. Proc. Fla. State Hort. Sci. 92:100-102.
86. Suzuki, I., Sugahara, Y., Kotani, A., Todaka, S., and Shimada, H. 1964. Studies on breeding eggplants and tomatoes for resistance to bacterial wilt. I. Investigations on method of evaluating the resistance and on the source of resistance in eggplants and tomatoes. Bulletin-Horticultural Research Station Ministry of Agriculture and Forestry Series A 3:77-106.
87. Theis T., 1950. Tomato diseases. Puerto Rico Univ. Agr. Expt. Sta., Ann. Rpt. (1950): 9.
88. Thoquet, P., Olivier, J., Sperisen, C., Rogowsky, P., Laterrot, H., and Grimsley, N. 1996a. Quantitative trait Loci determining Resistance to Bacterial Wilt in Tomato Cultivar Hawaii7996. Molecular Plant-Microbe Interactions 9:826-836.
89. Thoquet, P., Olivier, J., Sperisen, C., Rogowsky, P., Prior, P., Anaïs, G., Mangin, B., Bazin, B., Nazer, R., and Grimsley, N. 1996b. Polygenic resistance of tomato plants to bacterial wilt in the French West Indies. Molecular Plant-Microbe Interactions 9:837-842.
90. Thurston H.D., 1976. Resistance to bacterial wilt (Pseudomonas solanacearum). Proc. 1rst Int. Conference and Worskshop on the ecology and control of bacterial wilt caused by Pseudomonas solanacearum. Raleigh, July 1976: 58-62.
91. Vaneechoutte M, Kämpfer P, De Baere T, Falsen E, Verschraegen G. 2004. Wautersia gen. nov., a novel genus accommodating the phylogenetic lineage including Ralstonia eutropha and related species, and proposal of Ralstonia [Pseudomonas] syzygii (Roberts et al. 1990) comb. nov. Int J Syst Evol Microbiol. 54(Pt 2):317-27.
92. Walter J.M., 1967. Hereditary resistance to disease in tomato. Annual Review of Phytopathology, vol.5: 131-162.

29
93. Wang, J.-F., P. Hanson, and J.A. Barnes. 1998. Worldwide evaluation of an international set of resistance sources to bacterial wilt in tomato. In: P. Prior, C. Allen and J. Elphinstone (eds.), Bacterial Wilt Disease: Molecular and Ecological Aspects, Springer-Verlag, Berlin 269-275.
94. Wang, J. F., Olivier, J., Thoquet, P., Mangin, B., Sauviac, L., and Grimsley, N. H. 2000. Resistance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan is controlled mainly by a major strain-specific locus. Molecular Plant-Microbe Interactions 13 (1):6-13.
95. Wang HanRong; Ru ShuiJiang; Wang LianPing; Fang Li; Ren HaiYing; Feng ZhongMen, 2009. Control of tomato bacterial wilt with grafting. Acta Agriculturae Zhejiangensis 21(3): 283-287.
96. Warmke H.E., Cruzado H.J., 1949. Tomato breeding. Puerto Rico Fed. Expt. Sta. Ann. Rpt. For 1949: 9.
97. Wicker E., Grassart L., Coranson-Beaudu R., Mian D., Guilbaud C., Fegan M., Prior P., 2007. Ralstonia solanacearum strains from Martinique (French West Indies) exhibiting a new pathogenic potential. Appl. & Environ. Microbiology 73(21): 6790-6801.
98. Winstead N.N., Kelman A., 1952. Inoculation techniques for evaluating resistance to Pseudomonas solanacearum. Phytopathology 42: 628-634.
99. Yabuuchi E, Kosako Y, Oyaizu H, Yano I, Hotta H, Hashimoto Y, Ezaki T, Arakawa M. 1992. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol Immunol.36(12):1251-75.
100.Yabuuchi E, Kosako Y, Yano I, Hotta H, Nishiuchi Y. 1995. Transfer of two Burkholderia and an Alcaligenes species to Ralstonia gen. Nov.: Proposal of Ralstonia pickettii (Ralston, Palleroni and Doudoroff 1973) comb. Nov., Ralstonia solanacearum (Smith 1896) comb. Nov. and Ralstonia eutropha (Davis 1969) comb. Nov.Microbiol Immunol. 39(11): 897-904.
101.Yarnell S.H., 1948. The southern tomato exchange program. Proc. Am. Soc. Hort. Sci. 52: 375-382.

30

31
Figure 3: Pedigree of ‘MR4’ & ‘NC 72 TR 4-4’ (and of ‘Venus’ and ‘Saturn’) (North Carolina State University material) (Taken from Daunay, 1977, based on W.R. Henderson & E. Echandi, pers. com. to H. Laterrot in the 1970s, and on Hendersons & Jenkins, 1972)

32

33

34

35

36

37
Figure 9 : Pedigree of ‘CRA74’, ‘Carette’ & ‘Caraibo’, (INRA, Guadeloupe material) (Taken from Anais, 1986)

38

39

40

41
Research Papers TGC REPORT VOLUME 60, 2010 Preliminary Observations on the Effectiveness of five Introgressions for Resistance to Begomoviruses in Tomatoes Luis Mejía, Rudy E. Teni, Brenda E. García, Ana Cristina Fulladolsa, and Luis Méndez, Facultad de Agronomía, Universidad de San Carlos de Guatemala; Sergio Melgar, Escuela de Biología, Universidad de San Carlos de Guatemala and Douglas P. Maxwell, Department of Plant Pathology, University of Wisconsin-Madison Introduction: Tomato-infecting begomoviruses have remained a major constraint to tomato production in many parts of the tropical and sub-tropical regions. Management strategies have involved i) extensive use of insecticides to control the whitefly vector, Bemisia tabaci, ii) production of virus-free transplants, iii) host-free periods, iv) use of whitefly-proof fabric cover in micro- and macro-tunnels for 30 or more days, and v) use of moderately resistant hybrids. Over the past decade the availability of moderately resistant hybrids has increased and most private seed companies devote resources to the incorporation of begomovirus-resistance genes into their new generation of hybrids.
Several genes for resistance to begomoviruses have been identified in the last two decades (Ji et al., 2007). Zamir et al. (1994) described the first resistance gene, Ty1, originating from the wild species Solanum chilense accession LA1969, to a region located between 4 cM and 10 cM in the short arm of chromosome 6. Later, gene, Ty2, was incorporated into the genome of the tomato from the wild species Solanum habrochaites by Hanson et al. (2000) and the introgression was located between 84 and 91 cM in chromosome 11. More recently, Ji et al. (2007b) reported gene, Ty3, in an introgression derived from S. chilense accession LA2779 and located at 19 to 25 cM in chromosome 6. An introgression in this same region from S. chilense accession LA1932 was designed Ty3a (Ji et al., 2007a). The resistant lines from the accession LA1932 had yet another gene, Ty4, which was recently located in the upper half of chromosome 3 (Ji et al., 2008) near 82 cM (Maxwell, personal communication). A major QTL, Ty5, was recently mapped in the TY172 breeding line with introgressions from S. peruvianum from approximately 16 to 46 cM on chromosome 4 (Anbinder et al., 2009). The potential for pyramiding begomovirus-resistance introgressions from different accessions of S. chilense, from different wild species such as S. peruvianum or S. habrochaites was discussed by Vidavski et al. (2008). Using several sources of resistance in one hybrid may overcome the selection of virus variants with novel genome combinations that may be more aggressive on tomatoes with one begomovirus-resistance introgression (García-Andrés et al., 2009). Because of the need to understand how different resistance genes respond to begomoviruses, this project was initiated to evaluate the effectiveness of begomovirus-resistance introgressions (Ty1, Ty2, Ty3, Ty3a, and Ty4) for conferring resistance to begomoviruses in a field trial in Guatemala. It is expected that this information will

42
provide a rationale for pyramiding begomovirus-resistance introgressions into hybrids that will have more durable resistance. Materials and Methods:
Germplasm: The breeding lines used in these experiments were: Gh13, homozygous for Ty3; Gc143-2, homozygous for Ty1 and Ty3; Gc171, homozygous for Ty3a and Ty4 (provided by J. Scott, University of Florida); CLN2116 homozygous for Ty2 (provided by P. Hanson, The World Vegetable Center) and Gh188-2, also homozygous for Ty2, and Gc21-a, homozygous for Ty3a and lacking Ty4. Gh13 was selected from the hybrid, Favi9 (Vidavsky and Czosnek, 1988). Gc143-2 was selected from a cross of Gc9 (provided by J. Scott, University of Florida) by a susceptible commercial hybrid. Gc21-a was obtained by several cycles of selfing from a cross between Gc171 with a susceptible commercial hybrid and an individual F2 plant. Gh188-2 was selected from a commercial hybrid with Ty2 introgression. The susceptible germplasm, HUJ-VF, without any of the introgressions for resistance, was used in several crosses. HUV-VF was provided by F. Vidavsky, The Hebrew University of Jerusalem. The F2 population used in the segregation of genes Ty3a and Ty4 was obtained from a hybrid produced by a cross between the resistant line Gc171 and a slightly resistant inbred (Gh44).
PCR Methods for detection of the introgressions associated with resistance to begomoviruses: DNA extraction was as reported by García et al. (2008). PCR-based molecular markers have been developed for the detection of Ty2, Ty3, Ty3a, and Ty4. Garcia et al. (2007) developed a co-dominant SCAR (Sequence Characterized Amplified Region) marker from the RFLP T0302 marker at 89 cM for the detection of the Ty2 introgression. The Ty3 and Ty3a introgressions were monitored with the co-dominant SCAR marker P6-25 (Ji et al., 2007a). The Ty4 introgression was detected with PCR primers developed by Y. Ji and J. Scott (personal communication, Ji et al., 2008). The presence of the Ty1 introgression in Gc143-2 was determined by sequencing the PCR fragments associated with the RFLP TG97 marker (García and Maxwell, unpublished). Field Evaluation of disease severity for the different populations: Each entry was replicated three times with 5 plants per replication. Four-week-old seedlings were transplanted into a field near Sanarate, Guatemala, where high levels of viruliferous whiteflies were present. In this area, at least 7 bipartite tomato-infecting begomoviruses have been identified (Nakhla et al., 2004) as well as the monopartite begomovirus, Tomato yellow leaf curl virus (Mejía and Maxwell, unpublished results). Each plant was scored at about 30-days after transplanting in either January 2009 or October 2009 using a disease severity index (DSI) from zero to six: 0, no symptoms; 1, extremely slight symptoms; 2, slight symptoms; 3, moderate symptoms; 4, severe symptoms with deformed leaves; 5, severe symptoms with stunted plant; and 6, very severe symptoms, no marketable fruit and very stunted plant.

43
Development of populations with different introgressions: Combinations of introgressions Ty3a and Ty4: An F2 population was obtained
from a F1 hybrid produced from the cross between line Gc171 (Ty3a/Ty3a, Ty4/Ty4, Scott and Schuster, 2007) by a slightly resistant line (Gh44, ty3/ty3, ty4/ty4). Leaf samples were collected from 77 individual plants for molecular analysis. The genotype of these plants was determined with the PCR primers P6-25F2 and P6-25R5 (SCAR marker P6-25) for introgression Ty3a and a PCR-based marker for Ty4 introgression (Ji et al., 2008). Individual F2 plants were selected that were dominant for one or both genes (RR, Ty3a/Ty3a, Ty4/Ty4; RS, Ty3a/Ty3a, ty4/ty4; SR, ty3/ty3, Ty4/Ty4 and SS, ty3/ty3, ty4/ty4). These plants were selfed to produce F3 seeds. Twelve F3 families, Gc171, Gh44 and a susceptible commercial control C-SS (ty3/ty3, ty4/ty4) were planted in a field to determine their phenotype, i.e., the level of resistance to begomoviruses was determined in January 2009. Each entry was replicated three times with 5 plants per replication.
Combination of introgressions for Ty2 and Ty3: F2 seed was obtained from the cross between resistant line Gh13 (Ty3/Ty3, ty2/ty2, RS-C, Martin et al., 2007), and line CLN2116 (ty3/ty3,Ty2/Ty2, SR-C). The genotype of the F2 plants was determined using molecular markers and individuals with one, both or none of the introgressions for resistance (genotypes RR, Ty3/Ty3,Ty2/Ty2; SR, ty3/ty3, Ty2/Ty2; RS, Ty3/Ty3, ty2/ty2 and SS, ty3/ty3, ty2/ty2) were allowed to self. The genotype of each F3 family was verified and seventeen F3 families were transplanted into a field along with the two parental lines and a susceptible control. Each entry was replicated three times with 5 plants per replication to determine their phenotype and DSIs were taken in October 2009.
Combination of introgressions Ty1, Ty3 and Ty2: F2 seed was obtained from the cross between resistant line Gc143-2 (Ty1/Ty1-Ty3/Ty3, ty2/ty2) and line Gh188-2, (ty1/ty1-ty3/ty3, Ty2/Ty2). The genotype of the F2 plants was determined using molecular markers for Ty3 and Ty2 and individuals with one, both or none of the introgressions were identified (RR, Ty3/Ty3, Ty2/Ty2; RS, Ty3/Ty3, ty2/ty2; SR, ty3/ty3, Ty2/Ty2 and SS, ty3/ty3, ty2/ty2). The Ty1 introgression was not determined as it is linked to Ty3. F3 seed was collected from individual F2 plants of different genotypes and the F3 families were transplanted in the field. Sixteen F3 families and both parental lines were planted and each entry was replicated three times and 5 plants per replication. Plants were scored for their DSIs in October 2009.
Combination of introgressions Ty3a and Ty2: Gc21-a (RS, Ty3a/Ty3a, ty2/ty2), was crossed with line Gh188-2 (SR, ty3/ty3, Ty2/Ty2). Heterozygous individuals were evaluated for resistance (HH, Ty3a/ty3, Ty2/ty2), along with the parental genotypes. Gc21-a (RS) and Gh188-2 (SR) were also crossed to the susceptible line HUJ-VF (ty2/ty2, ty3/ty3). The genotypes of the F1 populations were verified and their resistance phenotype determined in the field in October 2009. Three hybrids [Gc21-a X Gh188-2 (HH, Ty3a/ty3, Ty2/ty2), Gc21-a X HUJ-VF (HS, Ty3a/ty3, ty2/ty2) and Gh188-2 X HUJ-VF (SH, ty3/ty3, Ty2/ty2)], two parental lines and a susceptible commercial control (SS) were planted with three replications of 5 plants each. Plants were scored for their DSIs in October 2009.
Combination of introgressions Ty3a and Ty3: Line Gc21-a (Ty3a/Ty3a) was crossed with line Gh13 (Ty3/Ty3). The level of resistance in the hybrid (Ty3a/Ty3)

44
was evaluated in relation to the parental genotypes. The genotype of the F1 families was determined and subsequently transplanted in the field for the evaluation of their phenotype of resistance. The hybrid (Gc21-a X Gh13), both parental lines, and a susceptible control were planted in with three replications of five plants each. Plants were scored for their DSIs in October 2009. Results and Discussion: Natural inoculation with begomoviruses in a field in Guatemala with high populations of the whitefly vector was used to evaluate various combinations of begomovirus-resistance introgressions for their effectiveness to provide resistance to multiple begomoviruses. Evaluations of the introgressions Ty3a and Ty4: The highly resistant inbred line, Gc171 (Ty3a/Ty3a, Ty4/Ty4), was crossed with a slightly resistant genotype (Gh44, ty3/ty3, ty4/ty4) (Garcia, et al., 2008a). Among the 77 F2 plants analyzed, 17 were found to be homozygous Ty3a/Ty3a, 12 homozygous ty3a/ty3a, and 48 heterozygous Ty3a/ty3a. With relation to Ty4 introgression, 10 plants were found to be homozygous Ty4/Ty4, 42 homozygous ty4/ty4, and 25 heterozygous Ty4/ty4. Seven F3 families were evaluated: three with genotype ty3/ty3, Ty4/Ty4 (SR), and four with genotype Ty3a/Ty3a, ty4/ty4 (RS) (Fig. 1).
Fig. 1. Disease severity index (DSI) and standard errors for F3 families of genotypes RR (4 families Ty3a/Ty3a, Ty4/Ty4), RS (4 families, Ty3a/Ty3a, ty4/ty4), SR (4 families, ty3/ty3, Ty4/Ty4), C-SS (commercial susceptible control), R-RR (parent, Gc171), S-SS (Gh44).
0
1
2
3
4
5
6
7
RR RS SR R-RR S-SS C-SS
DSI
F3 Families and Parents

45
The average DSI for the RS F3 families with the Ty3a introgression was 2.1, while the SR F3 families with the Ty4 introgression was 3.6, which is similar to the slightly resistant parent, Gh44 (S-SS). These results indicate that there is a larger effect on resistance by Ty3a than by Ty4 to the multiple bipartite begomoviruses present in this area. This is consistent with the report by Ji et al. (2008) where the Ty4 introgression had a lesser effect on resistance to Tomato yellow leaf curl virus than the Ty3a introgression.
Combinations of introgression Ty2 and Ty3: Seventeen F3 families were obtained from individual F2 plants arising from the cross of Gh13 X CLN2116 (F1, Ty3/ty3, Ty2/ty2). The F3 families were evaluated for symptom development 36 days after transplanting (Fig. 2).
Fig. 2. Disease severity index (DSI) with standard errors for the F3 families of genotypes RR (6 families, Ty3/Ty3, Ty2/Ty2), SR (5 families, ty3/ty3, Ty2/Ty2), RS (3 families, Ty3/Ty3, ty2/ty2), SS (3 families, ty3/ty3, ty2/ty2), SR-C (CLN2116, ty3/ty3, Ty2/Ty2), RS-C (Gh13, Ty3/Ty3, ty2/ty2) and SS-C (susceptible commercial hybrid control, ty3/ty3, ty2/ty2).
0
1
2
3
4
5
6
RR SR RS SS SR-C RS-C SS-C
DS
I
F3 Families and Parents

46
The parents had average DSIs of 0.6 and 3.8 for Gh13 and CLN2116, respectively. A commercial hybrid with neither introgression had a DSI of 5.2. The average DSI of the 6 RR families was 1.6, which was greater than the most resistant parent, Gh13 (DSI 0.6), and was similar to the DSI for the 3 RS families, which had an average DSI of 1.4. The average DSI of the 5 SR families (DSI 2.9) was similar to that for the 3 SS families (DSI 2.6).
These results indicate that Ty3 introgression from Gh13 was mainly responsible for the observed level of virus resistance in the F3 families and adding the Ty2 introgression resulted in no increase in virus resistance. The susceptible control had a DSI of 5.1, while the 3 F3 families without the introgressions (ty3/ty3, ty2/ty2) had a DSI of 2.6, indicating the presence of other unknown genes for resistance. Of interest was the range of average DSIs within families of one genotype (Fig. 3). For the 6 RR families, the average DSI ranged from 0.8 to 2.3. For the 3 SS families the range was also great, 1.8 to 3.6, for the 3 RS the range was 1.1 to 1.9. For the 5 SR families the range was 2.1 to 3.2. One explanation for these differences among F3 families with the same genotype for the two introgressions could be that there was segregation of other modifying resistance genes in the F2 plants used to generate the F3 families.
Fig. 3. Disease severity index (DSI) and standard errors for the individual F3 families with the genotypes as listed in Fig. 2 along with the parents and susceptible commercial control.
0
1
2
3
4
5
6
DS
I
F3 Families and Parents

47
Combination of introgressions Ty1-Ty3 and Ty2: Individual F2 plants of different genotype obtained from the F1 (Ty1-Ty3/ty1-ty3, Ty2/ty2) of the cross of Gc143-2 X Gh188-2 were selfed to produce 16 F3 families. The DSI for these families was determined in the field (Fig. 4).
Fig. 4. Average disease severity index (DSI) and standard errors for 16 F3 families of genotypes RR (5 families, Ty1-Ty3/Ty1-Ty3, Ty2/Ty2), RS (3 families, Ty1-Ty3/Ty1-Ty3, ty2/ty2), SR (3 families, ty1-ty3/ty1-ty3, Ty2/Ty2), SS (5 families, ty1-ty3/ty1-ty3, ty2/ty2), and the parents: RS-C (Gc143-2, Ty1-Ty3/Ty1-Ty3, ty2/ty2) and SR-C (Gh188-2, ty1-ty3/ty1-ty3, Ty2/Ty2).
0
1
2
3
4
5
6
RR RS SR RS-C SR-C SS
DS
I
F3 Families and Parents

48
The average DSI for RS families with the T1-Ty3 introgression was 1.7 while DSI for those SR families with the Ty2 introgression was 4.8. RR families with both introgressions for resistance had an average DSI of 1.4 and those with neither introgression had DSI of 3.6. There was no difference in the average DSI for the RR and SR families. Surprisingly, the DSI for the SR families was higher than the DSI for the SS families. These observations indicated that Ty2 introgression provides no effective resistance to these begomoviruses.
An important observation was that there was a considerable range in the average DSI for families with the same genotype for these two introgressions (Fig. 5). For example, one RR family had a DSI of 3.2, which was greater than the other four RR families (range of 0.3 to 1.4). Within the RS families, one family had a DSI of 0.2 and another RS family had a DSI of 3.8. The genotype for the markers in these families, i.e., the presence or absence of the PCR fragments corresponding to the S. lycopersicum size or the introgression size, was reconfirmed by additional testing in the laboratory. Explanations to consider are that there could have been a recombination event between the PCR marker and the resistance gene for these introgressions or that other genes controlling resistance were segregating within the F2 plants.
Fig. 5. Average disease severity index (DSI) and standard errors for individual F3 families for each genotype and the two parents. See Fig. 3 for codes.
-1
0
1
2
3
4
5
6
DS
I
F3 Families and Parents

49
Combinations of genes Ty3a and Ty2: Line Gc21-a (DSI 0.7), homozygous for Ty3a, was crossed to line Gh188-2 (DSI 5.2), homozygous for Ty2; and the DSI for the F1 (HH, Ty3a/ty3, Ty2/ty2) was 0.4. Lines Gc21-a and Gh188-2 were also crossed with susceptible line HUJ-VF (ty3/ty3, ty2/ty2). The resulting heterozygous F1‟s, Ty3a/ty3, ty2/ty2 (HS) and ty3/ty3, Ty2/ty2 (SH) had DSIs of 2.9 and 4.2, respectively (Fig. 6). Thus, the Ty2 gene along either in a homozygous or heterozygous condition did not confer an adequate level of resistance. The Ty3a introgression in the heterozygous condition conferred a moderate level of resistance, but when both resistance introgressions were present in the heterozygous condition the F1 was highly resistant.
Fig. 6. Average disease severity index (DSI) and standard errors for the heterozygous genotypes HH (Ty2/ty2, Ty3a/ty3), SH (ty3/ty3, Ty2/ty2), HS (Ty3a/ty3, ty2/ty2) and homozygous parental lines RS (Ty3a/Ty3a, ty2/ty2), SR (ty3/ty3, Ty2/Ty2), and susceptible commercial control SS (ty2/ty2, ty3/ty3).
0
1
2
3
4
5
6
HH SH HS SR RS SS
DSI
F1 families and Parents
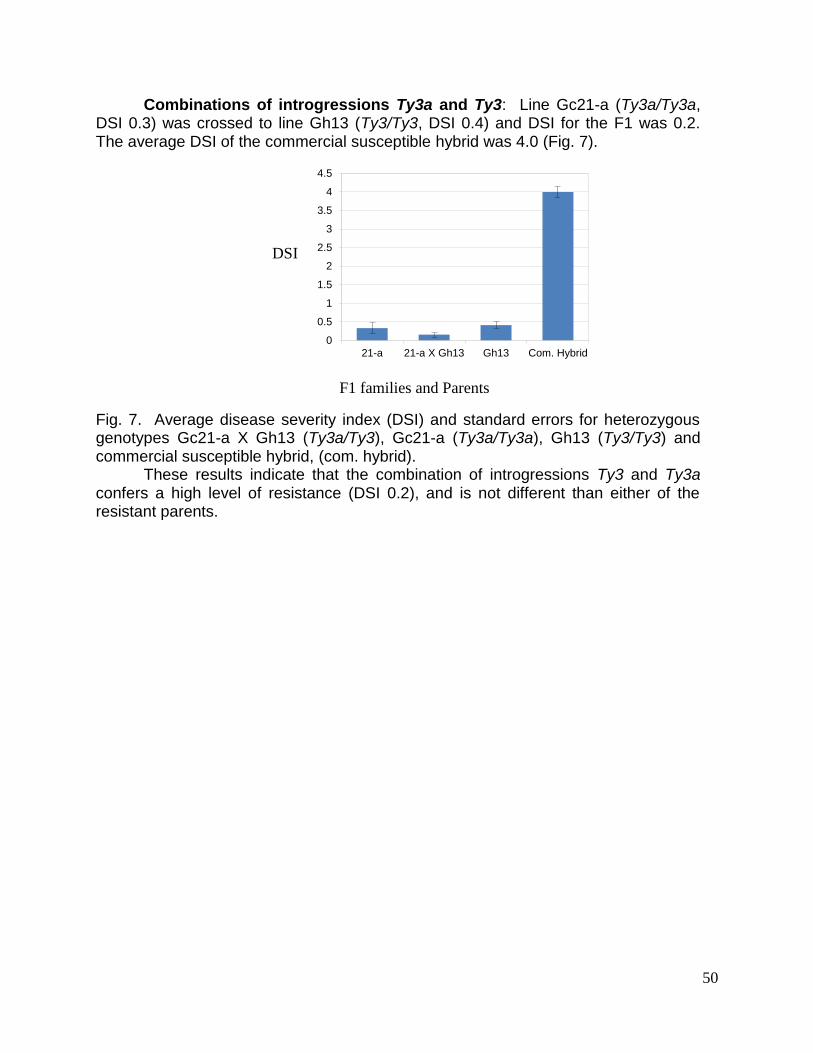
50
Combinations of introgressions Ty3a and Ty3: Line Gc21-a (Ty3a/Ty3a, DSI 0.3) was crossed to line Gh13 (Ty3/Ty3, DSI 0.4) and DSI for the F1 was 0.2. The average DSI of the commercial susceptible hybrid was 4.0 (Fig. 7).
Fig. 7. Average disease severity index (DSI) and standard errors for heterozygous genotypes Gc21-a X Gh13 (Ty3a/Ty3), Gc21-a (Ty3a/Ty3a), Gh13 (Ty3/Ty3) and commercial susceptible hybrid, (com. hybrid).
These results indicate that the combination of introgressions Ty3 and Ty3a confers a high level of resistance (DSI 0.2), and is not different than either of the resistant parents.
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
21-a 21-a X Gh13 Gh13 Com. Hybrid
DSI
F1 families and Parents

51
Conclusions: It should be recognized that not all of the comparisons contained all of the genotypes that would have been useful for making conclusions, that viruliferous whiteflies populations would have existed at different times, that several bipartite begomoviruses as well as Tomato yellow leaf curl virus would have been present, and that weather conditions may have impacted symptom expression. In general, the following observations were considered important concerning the various introgressions: 1) Ty3a was a more effective source of resistance than Ty4. 2) Ty2 either in the homozygous or heterozygous condition was not an effective source of resistance unless it was associated with another resistance gene, such as Ty3a. 3) The heterozygous families with Ty3a were not as resistant as families that were homozygous for this introgression (Fig. 6). 4) F1 hybrids with two different introgressions were highly resistant, such as Ty3a and Ty2 (Fig. 6) or Ty3a and Ty3 (Fig. 7). 5) There was considerable variation for the DSIs among the F3 families with same genotype for an introgression, which indicated that other genes were important in conditioning resistance. This was most notable in the F3 families from the cross of Gc143-2 by Gh188-2 (Fig. 5). Others have reported that several genes are involved in highest level of begomovirus resistance expression (Anbinder et al., 2009; Vidavsky and Czosnek, 1998; Zamir et al., 1994) and Anbinder et al. (2009) found that there were minor QTLs for resistance associated with both the resistant and susceptible parent used in the cross for their molecular analysis of resistance. 6) For the near future it is not enough to only use molecular markers for breeding for begomovirus resistance, but field evaluations with high levels of viruliferous whiteflies will continue to be an important part of any tomato breeding program for begomovirus resistance. Acknowledgements: This project was funded in part by grant FODECYT 54-07 to L. Mejía, by Facultad de Agronomía, Universidad de San Carlos, and by the College of Agricultural and Life Sciences, University of Wisconsin-Madison. Appreciation is expressed to Semillas Tropicales, S.A. for assistance with production of the seedlings and the field facilities.

52
Literature Cited: Abinder, I., Reuveni, M., Azari, R., Paran,I., Nahon, S., Shlomo, H., Chen, L., Lapidot,
M., and Levin, I. 2009. Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 119:519-530.
Garcia, B.E., Graham, E., Jensen, K.S., Hanson, P., Mejía, L., and Maxwell, D.P. 2007. Co-dominant SCAR for detection of the begomovirus-resistance Ty2 locus derived from Solanum habrochaites in tomato germplasm. Tom. Gen. Coop. Rept. 57:21-24.
Garcia, B.E., Barillas, A.C., Maxwell, D.P., and Mejia, L. 2008a. Genetic analysis of an F2 population for the segregation of two introgressions associated with the begomovirus-resistant parent, Gc171. Tomato Genetic Cooperative Report 58:18-21.
García, B.E., Mejía, L, Melgar, S., Teni, R., Sánchez-Pérez, A., Barillas, A.C., Montes, L., Keuler, N.S., Salus, M.S., Havey, M.J., and Maxwell, D.P. 2008b. Effectiveness of the Ty-3 introgression for conferring resistance in F3 families of tomato to bipartite begomoviruses in Guatemala. Tom. Genetic Coop. 58:22-28.
García-Andrés, S., Tomás, D.M., Navas-Castillo, J., and Moriones, E. 2009. Resistance-driven selection of begomoviruses associated with the tomato yellow leaf curl disease. Virus Research 146:66-72.
Hanson, P.M., Bernacchi, D., Green, S., Tanksley, S.D., Muniyappa, V., Padmaja, A.S., Chen, H.M., Kuo, G., Fang, D., and Chen, J.T. 2000. Mapping of a wild tomato introgression associated with tomato yellow leaf curl virus resistance in a cultivated tomato line. J. Amer. Soc. Hort. Sci. 125:15-20.
Ji, Y., Salus, M.S., van Betteray, B., Smeets, J., Jensen, K., Martin, C.T., Mejía, L., Scott, J.W., Havey, M.J., and Maxwell, D.P. 2007a. Co-dominant SCAR markers for detection of the Ty-3 and Ty-3a loci from Solanum chilense at 25 cM of chromosome 6 of tomato. Rept. Tomato Genetic Coop. 57:25-28.
Ji, Y., Schuster, D.J., and Scott, J.W. 2007b. Ty3, a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato. Mol. Breeding 20:271-284.
Ji, Y., Scott, J.W., Hanson, P., Graham, E., and Maxwell, D.P. 2007c. Sources of resistance, inheritance and location of genetic loci conferring resistance to members of the tomato-infecting begomoviruses. In: Czosnek, H. (ed), Tomato yellow leaf curl virus Disease. Springer, The Netherlands, pp. 343-362.
Ji, Y., Scott, J.W., Maxwell, D.P., and Schuster, D.J. 2008. Ty-4, a Tomato yellow leaf curl virus resistance gene on chromosome 3 of tomato. Tomato Genet. Coop. Rep. 58:29-31.
Martin, C.T., Salus, M.S., Garcia, B.E., Jensen, K.S., Montes, L., Zea, C., Melgar, S., El Mehrach, K., Ortiz, J., Sanchez, A., Havey, M.J., Mejía, L., and Maxwell, D.P. 2007. Evaluation of PCR-based markers for scanning tomato chromosomes for introgressions from wild species. Rept. Tomato Genetic Coop. 57:31-34.
Mejía, L., Teni, R.E., Vidavski, F., Czosnek, H., Lapidot, M., Nakhla, M.K., and Maxwell, D.P. 2005. Evaluation of tomato germplasm and selection of breeding lines for resistance to begomoviruses in Guatemala. Acta Hort. 695:251-255.

53
Nakhla, M.K., Sorenson, A., Mejía, L., Ramírez, P., Karkashian, J.P., and Maxwell, D.P. 2005. Molecular Characterization of Tomato-Infecting Begomoviruses in Central America and Development of DNA-Based Detection Methods. Acta Hort. 695:277-288.
Scott, J.W., and Schuster, D.J. 2007. Gc9, Gc171, and Gc173 begomovirus resistant inbreds. Tomato Cooperative Genetics Report 57:45-46.
Vidavski, F., Czosnek, H., Gazit, S., Levy, D., and Lapidot, M. 2008. Pyramiding of genes conferring resistance to Tomato yellow leaf curl virus from different wild tomato species. Plant Breeding 127:625-631.
Vidavsky, F., and Czosnek, H. 1998. Tomato breeding lines immune and tolerant to tomato yellow leaf curl virus (TYLCV) issued from Lycopersicon hirsutum. Phytopathology 88:910-914.
Zamir, D., Michelson, I., Zakay, Y., Navot, N., Zeidan, N., Sarfatti, M., Eshed, Y., Harel, E., Pleban, T., van-Oss, H., Kedar, N., Rabinowitch, H.D., and Czosnek, H. 1994. Mapping and introgression of a tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Genet. 88:141-146.

54
Research Papers TGC REPORT VOLUME 60, 2010 Preliminary report on association of ‘Candidatus Liberibacter solanacearum’ with field grown tomatoes in Guatemala
Luis Mejía, Amilcar Sánchez, and Luis Méndez, Facultad de Agronomía, Universidad de San Carlos de Guatemala; D. P. Maxwell, Department of Plant Pathology, University of Wisconsin-Madison; R. L. Gilberston, Department of Plant Pathology, University of California-Davis; V.V. Rivera and G.A. Secor, Department of Plant Pathology, North Dakota State University, Fargo. Introduction A new disease of tomatoes has received considerable attention in the local newspapers in Guatemala. Locally, it is referred to “Paratrioza disease”, which refers to the insect associated with symptomatic plants. The symptoms on tomatoes are flower abortion, purple margins of youngest leaves, upward cupping of leaves, thickened stems and retarded internode growth, and stunting of the plants (Fig. 1). In Mexico, a disease of tomato with similar symptoms is called permanent damage disease or permanent yellowing disease (daňo permanente del tomate; Páramo Menchaca, 2007) and Munyaneza et al. (2009) reported that Candidatus Liberibacter solanacearum was associated with these plants. This unculturable bacterium is transmitted by the tomato/potato psyllid (Bactericerca (Paratrioza) cockerelli).
Fig. 1. Typical symptoms associated with “Paratrioza disease” of tomato in Guatemala. Image taken December 2009 and shows flower abortion, purple leaf margins, cupping of leaves and thickened petioles and stems.
In a tomato field (about 0.7 ha, at 1,500 m, Department of Sacatepéquez) where symptom incidence was over 90%, samples of young leaves with typical symptoms were collected in December 2009. DNA was extracted at San Carlos University, Guatemala City (Garcia et al., 2007). PCR was performed at North Dakota State

55
University with 16s rRNA primers,CLi.po.F/O12c and PCR fragments sequenced using protocols reported by Secor et al. (2009). A PCR fragment of about 1,100 bp was obtained and sequenced in both directions. After correction by visual proofreading, a 966-nt region (contact D.P. Maxwell for sequence) was submitted to a BLAST analysis at the National Center for Biotechnology Information data base; and a 100% nucleotide identity was obtained with the 16s-23s rRNA intergenic spacer region for „Candidatus Liberibacter solanacearum’ from potato from Guatemala (FJ395205, Secor et al., 2009) and tomato from Sinaloa, Mexico (FJ957897, Munyaneza et al., 2009b). These sequences also had a 100% nucleotide identity with „Ca. L. solanacearum‟ from tomato in New Zealand (EU834130, Liefting et al., 2009a, 2009b), from bell pepper collected in Sinaloa, Mexico (FJ957896, Munyaneza et al., 2009a) and „Candidatus Liberibacter psyllaurous‟ (synonym „Ca. L. solanacearum‟, see discussion in Secor et al., 2009) from Zebra chip symptomatic potatoes in California (FJ498802, Crosslin and Bester, 2009). In Central America, Rehman et al. (2010) report the widespread occurrence of „Ca. L. solanacerum‟ (GQ926922) and its potato psyllid vector in potato fields in Honduras. Pair wise comparison of this sequence from „Ca. L. solanacearum‟ from potato with the Guatemalan „Ca. L. solanacearum’ sequence from tomato showed that there was a difference of two SNP between the sequences (678 nt), and this might indicate a different geographic origin of these two pathogens. Subsequently to samples collected in December 2009, samples were collected from tomatoes with either typical begomovirus symptoms and/or purple/yellowing symptoms on younger leaves in the Department of El Progresso in March 2010. The samples were prepared for transport to the University of California-Davis using AgDia absorption strips. This is a method to capture total nucleic acids from plant tissue in which sap is prepared from the target tissue, applied to an absorbent matrix on the end of a plastic 'stick', and allowed to dry prior to transport. DNA extracts were prepared from the 12 samples and tested for the presence of Liberobacter and begomovirus infection by PCR. 'Ca. Liberibacter sp.' was detected in 4 of 12 samples, and these samples showed symptoms of stunted and distorted growth; older leaves were yellow and brittle and younger leaves were upcurled with yellowing and vein purpling. Begomovirus infection was detected in all 12 samples. Additionally, 49 tomato samples were collected from March to June, 2010 and assayed for Liberibacter sp. using PCR primers CLi.po.F/O12c (Secor et al., 2009). The expected size fragment for Liberibacter sp. was obtained from 17 samples and no fragments were obtained with the other 32 samples. Positive PCR samples were collected from the following Departments: Sacatepéquez, Guatemala and Baja Verapaz,. These PCR fragments will be sequenced for more definitive identification. Universal PCR primers for phytoplasma (Smart et al., 1996) were used with the DNA samples from tomato collected in Guatemala and all were negative. Because of the purple top symptoms, phytoplasma were originally suspected as being present.
In the last two years, there has been considerable effort devoted to understanding the etiology of Zebra Chip (see Secor et al., 2009) and the new Candidatus Liberibacter sp. associated with solanaceous plants (see Liefting et al., 2009a, 2009b). These efforts plus the report here indicate that this unculturable bacterium transmitted by the tomato/potato psyllid will result in a serious disease for

56
tomatoes and peppers grown in the field or greenhouse (Brown et al., 2010) in Central America.
No universally accepted name exists for this disease on tomatoes, as illustrated by the use of different names: tomato vein-greening in Arizona (Brown et al., 2010), permanent yellowing in Mexico (Munyaneza et al., 2009b) and Paratrioza disease in Guatemala. At least from our observations in Guatemala several symptoms are notable: flower abortion, purpling of the leaf margins, stem thickening, yellowing of younger leaves, and stunting. An internet search reveals that there is a substantial number of sources using „permanente del tomate‟ or permanent yellowing disease of tomato. Thus, it is proposed that this diseased be named Liberibacter permanente del tomate or Liberibacter yellowing disease of tomato. This would distinguish this disease from psyllid yellows (Brown et al., 2010).

57
Literature Cited: Brown, J.K., Rehman, M., Rogan, D., Martin, R.R., and Idris, A.M. 2010. First report of
„Candidatus Liberibacter psyllaurous‟ (synonym „Ca. L. solanacearum‟ associated with „tomato vein-greening‟ and „tomato psyllid yellows‟ dieseses in commercial greenhouses in Arizona. Plant Dis. 94:376.
Crosslin, J.M., and Bester, G. 2009. First report of „Candidatus Liberibacter psyllasurous‟ in Zebra chip symptomatic potatoes from California. Plant Dis. 93:551.
Garcia, B.E., Graham, E., Jensen, K.S., Hanson, P., Mejía, L., and Maxwell, D.P. 2007. Co-dominant SCAR for detection of the begomovirus-resistance Ty2 locus derived from Solanum habrochaites in tomato germplasm. Tom. Gen. Coop. Rept. 57:21-24.
Liefting, L.W., Weir, B.S., Pennycook, S.R., and Clover, G.R.G. 2009a. „Candidatus Liberibacter solanacearum‟ associated with plants in the family Solanaceae. Internat. J. System. and Evolut. Microbiol. 59:2274-2276.
Liefting, L.W., Sutherland, P.W., Ward, L.I., Paice, K.L., Weir, B.S., and Clover, G.R.G. 2009b. A new „Candidatus Liberibacter‟ species associated with diseases of Solanaceous crops. Plant Dis. 93:208-214.
Munyaneza, J.E., Sengoda, V.G., Crosslin, J.M., Garzón-Tiznado, J.A., and Cardenas-Valenzuela, O.G. 2009a. First report of „Candidatus Liberibacter solanacearum‟ in pepper plants in México. Plant Dis. 93:1079.
Munyaneza, J.E., Sengoda, V.G., Crosslin, J.M., Garzón-Tiznado, J.A., and Cardenas-Valenzuela, O.G. 2009b. First report of „Candidatus Liberibacter solanacearum‟ in tomato plants in México. Plant Dis. 93:1079.
Páramo Menchaca, V. Estrategia integrada: control de Paratrioza, pulgón saltador o psílido de la papa y el tomate. Productores de Hortalizas. April, 2007.
Rehman, M., Melgar, J.C., Rivera, J.M., Idris, A.M., and Brown, J.K. 2010. First report of „Candidatus Liberibacter psyllaurous‟ or „Ca. Liberibacter solanacearum‟ associated with severe foliar chlorosis, curling, and necrosis and tuber discoloration of potato plants in Honduras. Plant Dis. 94:376.
Secor, G.A., Rivera, V.V., Abad, J.A., Clover, G.R.G, Liefting, L.W., Li. X., and De Boer, S.H. 2009. Association of „Candidatus Liberibacter solanacearum‟ with Zebra Chip disease of potato established by graft and psyllid transmission, electron microscopy, and PCR. Plant Dis. 93:574-583.
Smart, C.D., Schneider, B., Blomquist, C.L., Guerra, L.J., Harrison, N.A., Ahrens, U., Lorenz, K.-H., Seemuller, E., and Kirkpatrick, B.C. 1996. Phytoplasm-specific PCR primers based on sequences of the 16s-23s rRNA spacer region. Applied and Environ. Microbiol. 62:2988-2993.

58
Research Papers TGC REPORT VOLUME 60, 2010 Study of epidermal cell size of petals and stamens in tomato species and hybrids using confocal laser-scanning microscopy Christopher Lofty, Julian Smith, Pravda Stoeva-Popova Department of Biology, Winthrop University, Rock Hill SC 29733 E-mail: [email protected] Introduction
The phenomenon of cytoplasmic male sterility (CMS) has been described and the genetics underlying the phenomemon studied in many species. Whether arising spontaneously, as the result of mutations, or through alloplasmic incompatibilities in interspecific crosses, the main effect of CMS is on the development of stamens and pollen, leading to aberrant stamens with no pollen or aborted pollen (Kaul 1988). Other changes correlated with the CMS phenotype are changes in the second whorl affecting petal size and color (Andersen 1963, 1964; Petrova et al. 1999; Farbos et al. 2001; Leino et al. 2003)
In the tomato, CMS does not occur naturally. CMS has been reported in interspecies hybrids. Andersen (1963, 1964) reported the emergence and increase of pollen abortion in F1 and backcrosses of the crosses between Solanum lycopersicum, S. cheesmaniiae (formerly L. chesmanii f. typicum and f. minor) or S. habrochaites (formerly L. hirsutum f. glabratum) used as pistillate parents and S. pennellii as the recurrent pollinating parent. Pleiotropic effects of the CMS phenotype included the reduction of anther length and size, and the lengthening of the filaments. The anther size was negatively correlated to the percent of aborted pollen.
Similar results were observed by Valkova-Atchkova (1980) in crosses involving S. peruvianum as pistillate parent and S. pennellii and S. habrochaites (formerly L. hirsutum f. typicum) as pollinating parents. Further introgression of the nuclear genome of the recurrent parents confirmed the stability of the CMS phenotype over many generations (Petrova et al. 1999, Stoeva et al. 2007).
As a preliminary step to dissecting morphological (and underlying genetic) changes occurring in the CMS phenotype, this study has focused on the comparative analysis of the size of epidermal cells from abaxial and adaxial sides of petals and stamens of mature flowers from CMS-pennellii line (0% stainable pollen), the isonuclear S. pennellii (100% pollen fertility), and the cultivated tomato S. lycopersicum. Materials and methods Plant material
In the study the following genotypes were used: Solanum lycopersicum (cv. Merkurii), Solanum pennellii (LA716), and CMS-pennellii (CMS line) previously described in Petrova et al. (1998) and Radkova (2002). To determine the size of the epidermal cells, fully expanded flowers in stage 20 according to the classification of Burkhin et al. (2003) were collected from plants grown in the same environmental

59
chamber with 24C/ 20C day/night temperature and 18/6 hours day/night photoperiod (Fig. 1). Tomato flower preparation for confocal microscopy: A 0.1% aqueous solution of calcofluor white (SIGMA) was made (Pringle 1991). Excess solution was kept in the freezer and defrosted as needed. Excised floral structures (petals, anthers or filaments) were immersed in approximately 5 ml of 0.1% calcofluor solution and placed under vacuum until rapid boil was achieved. The sunken specimens were allowed to soak for 20 minutes in the dark and then were rinsed in dH2O for 10 minutes. Specimens were dissected and placed on slides. For the anthers, spacers of approximately 0.5 mm – 1 mm thick were used to ensure the specimens were not smashed by the coverslip. Coverslips were placed on top and fixed with nail polish at each corner. Calcofluor staining of epidermal cell walls was observed using the DAPI channel of the confocal laser-scanning microscope (CLSM) (FV1000). Images were taken at the planes in which cell wall margins contacted each other. Cells were
counted in image frames measuring from 150 m X 150 m to 300 m X 270 m, with the frame size adjusted as needed to include from five to twenty cells in each frame.
Ten to fifteen cells from both sides (abaxial and adaxial) of each floral structure
were measured using ImageJ (Rasband, 2009) to obtain cell areas (in m2). The ANOVA and Tukey's LSD statistical tests (Minitab, Inc.) were performed separately for each epidermal floral surface across genotypes to determine if any significant difference existed between the mean surface areas of studied epidermal cells. Results Petal abaxial cell surface area was significantly different among the three genotypes (p < 0.001) with CMS line having the largest mean cell size and S. pennellii having the smallest (Figs. 2, 3). Petal adaxial cell sizes differed significantly with S. lycopersicum and S. pennelli both being larger than the CMS line (Fig 3). Although no statistical test was done, abaxial cells were smaller than the adaxial in the two species S. pennellii and S. lycopersicum. On the contrary, the abaxial cells in the CMS line were larger in comparison to the adaxial cells.
Epidermal cell area on both the adaxial and the abaxial surfaces of the anthers was significantly different (p< 0.001) among the three genotypes (Figs.4, 5), with S. lycopersicum having the largest mean cell size for both, and the CMS line, the smallest.
Among the filament epidermal cells, the abaxial cells in the CMS line and cultivated tomato were not significantly different, but both were significantly bigger than those in S. pennellii (p< 0.001) (Fig.6, 7). There were no significant differences in the mean size of the epidermal cells on the adaxial surface of the filaments of the three strains (p=0.252) (Fig.5, 6). There was a large variation (Fig. 7) in the size of the adaxial epidermal cells in S. pennellii and the CMS line, while the size of the adaxial cells of filament in the cultivated tomato showed less variation. Discussion
According to published data, cytoplasmic male sterility has a considerable effect on the size and color of petals and stamens (Kaul 1988, Farbos et al. 2001, Leino et al. 2003). Our previous studies had also shown that in comparison to S. pennellii, the CMS

60
flowers have smaller and lighter green petals and anthers and longer filaments (Fig 1; Petrova et al 1998). In addition to tomato nuclear male sterility mutants (reviewed in Gorman and McCormick 1997) ), similar changes of flower structures have been reported for tomato plants grown at low temperatures (Lozano et al. 1998) and in loss-of-function transgenic plants for the B-class genes required for petal and stamen identity (De Martino et al. 2006).
The size of plant organs is determined by cell division and cell expansion and elongation. Our investigation of the size of epidermal cells shows that the smaller size of the anthers in the CMS line (Fig. 1; Stoeva-Popova et al., unpublished) can be explained largely by the reduction in the size of the cells. The epidermal cells on both surfaces of the CMS anther were significantly smaller in comparison to S. pennellii: respectively 51% smaller on the adaxial side, and 42.3% on the abaxial surface. On the other hand, no corresponding effect of cytoplasmic male sterility on cell size in the petals was observed On the abaxial surface of the petals of CMS line, the cells were 81.5% larger than in S. pennellii, while on the adaxial side, they were 32.2 % smaller. According to our measurements (Stoeva et al., unpublished data) the CMS-pennellii filaments are several magnitudes longer than the filaments of S. pennellii (Fig. 1). This is not merely a consequence of increased cells size, as the filament epidermal cells of CMS are statistically larger than those of S. pennellii only on the abaxial side. CMS-pennellii and S. pennellii share the same nuclear genome and cytoplasmic male sterility affects expression of nuclear genes ( reviewed in Linke, Börner 2005, Chase 2006). Our results above show that cytoplasmic male sterility does not equally affect the development of petals, stamens and filaments.
Our study has shown that there are significant differences in the size of epidermal cells between the two species: the red-fruited cultivated tomato and the green-fruited S. pennellii. Significant differences were determined for the epidermal cells of the anthers and petals. The epidermal cells of the anthers of the cultivated tomato were larger, which is an indication that the anthers of S. pennellii have greater number of epidermal cells. The same conclusion could be drawn from the study of the abaxial epidermal cells of the petals and the filaments. Since the two species are not closely related it will be interesting to investigate other representatives of the tomato clade and to determine if the size of epidermal cells of flower structures can be indicative of species relatedness.
Although no statistical analysis was carried out, our data show one consistent feature across all genotypes: the largest epidermal cells were observed on anthers, while the ones with the smallest surface area were epidermal cells of petals.

61
References
Andersen W.R. (1964). Evidence for plasmon differentiation in Lycopersicon. Report Tomato Genet. Coop. 14:4-6
Andersen, W.R. (1963). Cytoplasmic sterility in hybrids of Lycopersicon esculentum and Solanum pennellii. Report Tomato Genet. Coop. 13:7-8
Burkhin V., Hernould M., Gonzalez N., Chevalier C., Mouras A. (2003). Flower development schedule in tomato Lycopersicon esculentum cv. sweet cherry. Sex. Plant. Reprod. 15:311-320
Chase Ch. (2006). Cytoplasmic male sterility: a window to the world of plant mitochondrial-nuclear interactions. Trends in Genetics 23 (3):81-90
de Martino G., Pan I., Emmanuel E., Levy A., Irish V.F. (2006). Functional analysis of two tomato APETALA3 genes demonstrate diversification in their roles in regulating floral development. Plant Cell 18:1833-1845
Farbos I., Mouras A., Bereterbide A., Glimelius K. (2001). Defective cell proliferation in the floral meristems of alloplasmic plants of Nicotiana tabacum leads to abnormal floral organ development and male sterility. Plant Journal 26:131-142.
Gorman S.W., McCormick S. (1997). Male sterility in tomato. Critical Reviews in Plant Sciences 16(1):31-53
Kaul M.L.H (1988). Male sterility in higher plants. In: Monographs on Theor. Appl. Genet. 10. Springer Verlag Berlin
Leino M., Teixeira R., Landgren M., Glimelius K. (2003). Brassica napus lines with rearranged Arabidopsis mitochondria display CMS and a range of developmental aberrations. Theor. Appl. Genet. 106:1156-1163
Linke B., Börner T. (2005). Mitochondrial effects on flower and pollen development. Mitochondrion 5:389-402
Lozano R., Angosto T., Gomez P., payan C., Huijer P., Salinas J., Martinez-Zapater J.M. (1998). Tomato flower abnormalities induced by low temperatures are associated with changes of expression of MADS-box genes. Plant Physiology 117:91-100
Petrova M., Vulkova Z., Gorinova N., Izhar S., Firon N., Jacquemin J.-M., Atanassov A., Stoeva P. (1999). Characterization of cytoplasmic male sterile hybrid line between Lycopersicon peruvianum Mill. x Lycopersicon pennellii Corr. and its crosses with the cultivated tomato. Theor. Appl. Genet. 98:825-830
Pringle JR. (1991) Staining of bud scars and other cell wall chitin with calcofluor. Methods in Enzymology 4:732-5
Radkova M. (2002). Morphological, cytogenetic and molecular genetic studies of cytoplasmic male sterility in genus Lycopersicon. PhD Thesis, AgroBioInstitute, Sofia, Bulgaria
Rasband W.S., (2009) ImageJ. U. S. National Institutes of Health, Bethesda, Maryland, USA, http://rsb.info.nih.gov/ij.

62
Stoeva P., Dimaculangan2D., Radkova M., Vulkova Z. (2007). Towards cytoplasmic male sterility in cultivated tomato. Journal of Agricultural, Food and Environmental Sciences 1(1): http://www.scientificjournals.org/journals2007/articles/1058.htm
Valkova-Achkova Z. (1980). L. peruvianum a source of CMS. Rep. Tomato Genet. Coop. 30:36
Authors’ contributions: P.S-P. designed the experiment and provided the plant material. C.L. prepared all of the specimens and measured the specimens using techniques designed in collaboration with J.SIII. C.L. drafted the Methods and Results sections, and performed the statistical analysis under the supervision of JSIII. P.S-P. drafted the Introduction and Discussion. All three authors contributed to the final editing and read and approved the manuscript before submission. Fig.1: Flowers of Solanum lycopersicum (A), S. pennellii (B) and CMS-pennellii (C)
A B
C

63
Fig 2: CLSM photographs of epidermal cells from abaxial and adaxial surfaces of petals from flowers (genotypes and magnification as indicated on pictures)
Fig 3: Mean epidermal surface area of petals. Bars with the same letter are statistically equal

64
Fig. 4: CLSM photographs of epidermal cells from abaxial and adaxial surfaces of mature anthers (genotypes and magnification as indicated on pictures
Fig. 5: Mean epidermal surface area of anthers. Bars with the same letter are statistically equal

65
Fig. 6: CLSM photographs of epidermal cells from abaxial and adaxial surfaces of filaments from mature stamens (genotypes and magnification as indicated on pictures)
Fig 7: Mean epidermal surface area of filaments. Bars with the same letter are statistically equal

66
TGC REPORT VOLUME 60, 2010 Revised List of Wild Species Stocks Chetelat, R. T. C.M. Rick Tomato Genetics Resource Center, Dept. of Plant Sciences, University of California, One Shields Ave., Davis, CA 95616 The following list of 1,196 accessions of wild tomatoes and allied Solanum species is a revision of the list published in TGC vol. 57, 2007. Other types of TGRC stocks are catalogued in TGC 58 (monogenic mutants) and TGC 59 (miscellaneous stocks). Inactive accessions have been dropped and new collections added to the present list. The new material includes populations of wild or feral cherry tomato (S. lycopersicum ‘cerasiforme‟) from Mexico (LA4352, LA4353), a stock of S. pimpinellifolium containing the sun gene introgressed from S. lycopersicum, and populations of S. peruvianum from the Azapa valley of northern Chile (LA4445-LA4448). Seed samples will be provided, upon request, for research, breeding or educational purposes. Some accessions may be temporarily unavailable for distribution during seed multiplication. In general, only small quantities of seed will be provided: 25 seed per accession for the self-pollinated accessions, 50 for the outcrossers or facultative accessions, and 5-10 for the allied Solanum species. These seed samples should be sufficient for researchers to produce larger quantities of seed, if needed. Accessions are grown for seed increase in the UC-Davis greenhouses, except for cherry tomatoes and certain populations of S. pimpinellifolium, which are grown in the field. The population sizes used for seed multiplication depend on the mating system, and are designed to maintain genetic diversity within accessions (see guidelines at http://tgrc.ucdavis.edu). The following tables are ordered by species name, using the classification system of Peralta et al. (2008)11, but with the equivalent Lycopersicon names listed as well. Although only brief collection site data can be presented here, more detailed records are available from our website, including geographic coordinates, images, and donor information. An appendix table lists the accessions belonging to the core subsets for each species.
S. arcanum (L. peruvianum or L. peruvianum var. humifusum)
LA0378 Cascas Cajamarca Peru
LA0385 San Juan (Rio Jequetepeque) Cajamarca Peru
LA0389 Abra Gavilan Cajamarca Peru
LA0392 Llallan Cajamarca Peru
LA0441 Cerro Campana La Libertad Peru
LA1027 Cajamarca Peru
11
Peralta, I. E., D. M. Spooner, S. Knapp (2008) Taxonomy of wild tomatoes and their relatives
(Solanum sect. Lycopersicoides, sect. Juglandifolia, and sect. Lycopersicon; Solanaceae).
Systematic Botany Monographs 84: 1-186.

67
S. arcanum (L. peruvianum or L. peruvianum var. humifusum)
LA1031 Balsas Amazonas Peru
LA1032 Aricapampa La Libertad Peru
LA1346 Casmiche La Libertad Peru
LA1350 Chauna Cajamarca Peru
LA1351 Rupe Cajamarca Peru
LA1360 Pariacoto Ancash Peru
LA1394 Balsas - Rio Utcubamba Amazonas Peru
LA1395 Chachapoyas Amazonas Peru
LA1396 Balsas (Chachapoyas) Amazonas Peru
LA1626 Mouth of Rio Rupac Ancash Peru
LA1708 Chamaya to Jaen Cajamarca Peru
LA1984 Otuzco La Libertad Peru
LA1985 Casmiche La Libertad Peru
LA2150 Puente Muyuno (Rio Jequetepeque) Cajamarca Peru
LA2151 Morochupa (Rio Jequetepeque) Cajamarca Peru
LA2152 San Juan #1 (Rio Jequetepeque) Cajamarca Peru
LA2153 San Juan #2 (Rio Jequetepeque) Cajamarca Peru
LA2157 Tunel Chotano Cajamarca Peru
LA2163 Cochabamba to Yamaluc Cajamarca Peru
LA2164 Yamaluc Cajamarca Peru
LA2172 Cuyca Cajamarca Peru
LA2185 Pongo de Rentema Amazonas Peru
LA2326 Above Balsas Amazonas Peru
LA2327 Aguas Calientes Cajamarca Peru
LA2328 Aricapampa La Libertad Peru
LA2330 Chagual La Libertad Peru
LA2331 Agallapampa La Libertad Peru
LA2333 Casmiche La Libertad Peru
LA2334 San Juan Cajamarca Peru
LA2388 Cochabamba to Huambos (Chota) Cajamarca Peru
LA2548 La Moyuna (Magadalena) Cajamarca Peru
LA2550 El Tingo, Chorpampa (Rio Jequetepeque) Cajamarca Peru
LA2553 Balconcillo de San Marcos Cajamarca Peru
LA2555 Marical - Castilla La Libertad Peru
LA2565 Potrero de Panacocha a Llamellin Ancash Peru
LA2566 Cullachaca Ancash Peru
LA2582 San Juan (4x) Cajamarca Peru
LA2583 (4x)
LA2917 Chullchaca Ancash Peru
LA4316 Kuntur Wasi Cajamarca Peru
S. cheesmaniae (L. cheesmanii)
LA0166 Santa Cruz: Barranco, N of Punta Ayora Galapagos Islands Ecuador

68
S. cheesmaniae (L. cheesmanii)
LA0421 Cristobal: cliff East of Wreck Bay Galapagos Islands Ecuador
LA0422 San Cristobal: Wreck Bay, Puerto Baquerizo Galapagos Islands Ecuador
LA0428 Santa Cruz: Trail Bellavista to Miconia Zone Galapagos Islands Ecuador
LA0429 Santa Cruz: Crater in highlands Galapagos Islands Ecuador
LA0434 Santa Cruz: Rambech Trail Galapagos Islands Ecuador
LA0437 Isabela: Ponds North of Villamil Galapagos Islands Ecuador
LA0521 Fernandina: Inside Crater Galapagos Islands Ecuador
LA0522 Fernandina: Outer slopes Galapagos Islands Ecuador
LA0524 Isabela: Punta Essex Galapagos Islands Ecuador
LA0528B Santa Cruz: Academy Bay Galapagos Islands Ecuador
LA0529 Fernandina: Crater Galapagos Islands Ecuador
LA0531 Baltra: Barranco slope, N side Galapagos Islands Ecuador
LA0746 Isabela: Punta Essex Galapagos Islands Ecuador
LA0749 Fernandina: North side Galapagos Islands Ecuador
LA0927 Santa Cruz: Academy Bay Galapagos Islands Ecuador
LA0932 Isabela: Tagus Cove Galapagos Islands Ecuador
LA1035 Fernandina: Low elevation Galapagos Islands Ecuador
LA1036 Isabela: far north end Galapagos Islands Ecuador
LA1037 Isabela: Alcedo East slope Galapagos Islands Ecuador
LA1039 Isabela: Cape Berkeley Galapagos Islands Ecuador
LA1040 San Cristobal: Caleta Tortuga Galapagos Islands Ecuador
LA1041 Santa Cruz: El Cascajo Galapagos Islands Ecuador
LA1042 Isabela: Cerro Santo Tomas Galapagos Islands Ecuador
LA1043 Isabela: Cerro Santo Tomas Galapagos Islands Ecuador
LA1138 Isabela: E of Cerro Azul Galapagos Islands Ecuador
LA1139 Isabela: W of Cerro Azul Galapagos Islands Ecuador
LA1402 Fernandina: W of Punta Espinoza Galapagos Islands Ecuador
LA1404 Fernandina: W flank caldera Galapagos Islands Ecuador
LA1406 Fernandina: SW rim caldera Galapagos Islands Ecuador
LA1407 Fernandina: caldera, NW bench Galapagos Islands Ecuador
LA1409 Isabela: Punta Albermarle Galapagos Islands Ecuador
LA1412 San Cristobal: opposite Isla Lobos Galapagos Islands Ecuador
LA1414 Isabela: Cerro Azul Galapagos Islands Ecuador
LA1427 Fernandina: WSW rim of caldera Galapagos Islands Ecuador
LA1447 Santa Cruz: Darwin Station-Punta Nunez Galapagos Islands Ecuador
LA1448 Santa Cruz: Puerto Ayora, Pelican Bay Galapagos Islands Ecuador
LA1449 Santa Cruz: Darwin Station, Seismo Station Galapagos Islands Ecuador
LA1450 Isabela: Bahia San Pedro Galapagos Islands Ecuador
LA3124 Santa Fe: near E landing Galapagos Islands Ecuador
S. chilense (L. chilense)
LA0130 Moquegua Moquegua Peru
LA0294 Tacna Tacna Peru

69
S. chilense (L. chilense)
LA0456 Clemesi Moquegua Peru
LA0458 Tacna Tacna Peru
LA0460 Palca Tacna Peru
LA0470 Taltal Antofagasta Chile
LA1029 Moquegua Moquegua Peru
LA1030 Tarata Rd. Tacna Peru
LA1782 Quebrada de Acari Arequipa Peru
LA1917 Llauta (4x) Ayacucho Peru
LA1930 Quebrada Calapampa Arequipa Peru
LA1932 Minas de Acari Arequipa Peru
LA1938 Quebrada Salsipuedes Arequipa Peru
LA1958 Pampa de la Clemesi Moquegua Peru
LA1959 Huaico Moquegua Moquegua Peru
LA1960 Rio Osmore Moquegua Peru
LA1961 Toquepala Tacna Peru
LA1963 Rio Caplina Tacna Peru
LA1965 Causuri Tacna Peru
LA1967 Pachia, Rio Caplina Tacna Peru
LA1968 Cause Seco Tacna Peru
LA1969 Estique Pampa Tacna Peru
LA1970 Tarata Tacna Peru
LA1971 Palquilla Tacna Peru
LA1972 Rio Sama Tacna Peru
LA2404 Arica to Tignamar Tarapaca Chile
LA2405 Tignamar Tarapaca Chile
LA2406 Arica to Putre Tarapaca Chile
LA2731 Moquella Tarapaca Chile
LA2737 Yala-yala Tarapaca Chile
LA2739 Nama to Camina Tarapaca Chile
LA2746 Asentamiento-18 Tarapaca Chile
LA2747 Alta Azapa Tarapaca Chile
LA2748 Soledad Tarapaca Chile
LA2749 Punta Blanca Antofagasta Chile
LA2750 Mina La Despreciada Antofagasta Chile
LA2751 Pachica (Rio Tarapaca) Tarapaca Chile
LA2753 Laonzana Tarapaca Chile
LA2754 W of Chusmisa Tarapaca Chile
LA2755 Banos de Chusmisa Tarapaca Chile
LA2757 W of Chusmisa Tarapaca Chile
LA2759 Mamina Tarapaca Chile
LA2762 Quebradas de Mamina a Parca Tarapaca Chile
LA2764 Codpa Tarapaca Chile
LA2765 Timar Tarapaca Chile

70
S. chilense (L. chilense)
LA2767 Chitita Tarapaca Chile
LA2768 Empalme Codpa Tarapaca Chile
LA2771 Above Poconchile Tarapaca Chile
LA2773 Zapahuira Tarapaca Chile
LA2774 Socorama Tarapaca Chile
LA2778 Chapiquina Tarapaca Chile
LA2779 Cimentario Belen Tarapaca Chile
LA2780 Belen to Lupica Tarapaca Chile
LA2879 Peine Antofagasta Chile
LA2880 Quebrada Tilopozo Antofagasta Chile
LA2882 Camar Antofagasta Chile
LA2884 Ayaviri Antofagasta Chile
LA2887 Quebrada Bandurria Antofagasta Chile
LA2888 Loma Paposo Antofagasta Chile
LA2891 Taltal Antofagasta Chile
LA2930 Quebrada Taltal Antofagasta Chile
LA2931 Guatacondo Tarapaca Chile
LA2932 Quebrada Gatico, Mina Escalera Antofagasta Chile
LA2946 Guatacondo Tarapaca Chile
LA2949 Chusmisa Tarapaca Chile
LA2952 Camiña Tarapaca Chile
LA2955 Quistagama Tarapaca Chile
LA2980 Yacango Moquegua Peru
LA2981A Torata to Chilligua en route to Puno Moquegua Peru
LA3111 Tarata Tacna Peru
LA3112 Estique Pampa Tacna Peru
LA3113 Apacheta Tacna Peru
LA3114 Quilla Tacna Peru
LA3115 W of Quilla Tacna Peru
LA3153 Desvio Omate (Rio de Osmore) Moquegua Peru
LA3155 Quinistaquillas Moquegua Peru
LA3355 Cacique de Ara Tacna Peru
LA3356 W of Tacna Tacna Peru
LA3357 Irrigacion Magollo Tacna Peru
LA3358 Rio Arunta-Cono Sur Tacna Peru
LA3784 Rio Chaparra Arequipa Peru
LA3785 Terras Blancas Arequipa Peru
LA3786 Alta Chaparra Arequipa Peru
LA4106 Taltal Antofagasta Chile
LA4107 Catarata Taltal Antofagasta Chile
LA4108 Caleta Punta Grande Antofagasta Chile
LA4109 Quebrada Canas Antofagasta Chile
LA4117A San Pedro - Paso Jama Antofagasta Chile

71
S. chilense (L. chilense)
LA4117B San Pedro - Paso Jama Antofagasta Chile
LA4118 Toconao Antofagasta Chile
LA4119 Socaire Antofagasta Chile
LA4120 Cahuisa Tarapaca Chile
LA4121 Pachica - Poroma Tarapaca Chile
LA4122 Chiapa Tarapaca Chile
LA4127 Alto Umayani Tarapaca Chile
LA4129 Pachica (Rio Camarones) Tarapaca Chile
LA4132 Esquina Tarapaca Chile
LA4319 Alto Rio Lluta Tarapaca Chile
LA4321 Quebrada Cardones Tarapaca Chile
LA4324 Estacion Puquio Tarapaca Chile
LA4327 Pachica, Rio Camarones Tarapaca Chile
LA4329 Puente del Diablo, Rio Salado Antofagasta Chile
LA4330 Caspana Antofagasta Chile
LA4332 Rio Grande Antofagasta Chile
LA4334 Quebrada Sicipo Antofagasta Chile
LA4335 Quebrada Tucuraro Antofagasta Chile
LA4336 Quebrada Cascabeles Antofagasta Chile
LA4337 Quebrada Paposo Antofagasta Chile
LA4338 Quebrada Taltal, Estacion Breas Antofagasta Chile
LA4339 Quebrada Los Zanjones Antofagasta Chile
S. chmielewskii (L. chmielewskii)
LA1028 Casinchihua Apurimac Peru
LA1306 Tambo Ayacucho Peru
LA1316 Ocros Ayacucho Peru
LA1317 Hacienda Pajonal Ayacucho Peru
LA1318 Auquibamba Apurimac Peru
LA1325 Puente Cunyac Apurimac Peru
LA1327 Sorocata Apurimac Peru
LA1330 Hacienda Francisco Apurimac Peru
LA2639B Puente Cunyac Apurimac Peru
LA2663 Tujtohaiya Cusco Peru
LA2677 Huayapacha #1 Cusco Peru
LA2678 Huayapacha #2 Cusco Peru
LA2679 Huayapacha #3 Cusco Peru
LA2680 Puente Apurimac #1 Cusco Peru
LA2681 Puente Apurimac #2 Cusco Peru
LA2695 Chihuanpampa Cusco Peru
LA3642 Ankukunka Cusco Peru
LA3643 Colcha Cusco Peru
LA3644 Puente Tincoj Cusco Peru

72
S. chmielewskii (L. chmielewskii)
LA3645 Boca del Rio Velille Cusco Peru
LA3648 Huallapachaca Apurimac Peru
LA3653 Matara Apurimac Peru
LA3654 Casinchigua to Chacoche Apurimac Peru
LA3656 Chalhuani Apurimac Peru
LA3658 Occobamba Apurimac Peru
LA3661 Pampotampa Apurimac Peru
LA3662 Huancarpuquio Apurimac Peru
S. corneliomulleri (L. peruvianum or L. peruv. f. glandulosum)
LA0103 Cajamarquilla, Rio Rimac Lima Peru
LA0107 Hacienda San Isidro, Rio Canete Lima Peru
LA0364 9 Km W of Canta Lima Peru
LA0366 12 Km W of Canta Lima Peru
LA0444 Chincha #1 Ica Peru
LA0451 Arequipa Arequipa Peru
LA1133 Huachipa Lima Peru
LA1271 Horcon Lima Peru
LA1274 Pacaibamba Lima Peru
LA1281 Sisacaya Lima Peru
LA1283 Santa Cruz de Laya Lima Peru
LA1284 Espiritu Santo Lima Peru
LA1292 San Mateo Lima Peru
LA1293 Matucana Lima Peru
LA1294 Surco Lima Peru
LA1296 Tornamesa Lima Peru
LA1304 Pampano Huancavelica Peru
LA1305 Ticrapo Huancavelica Peru
LA1331 Nazca Ica Peru
LA1339 Capillucas Lima Peru
LA1373 Asia Lima Peru
LA1377 Navan Lima Peru
LA1379 Caujul Lima Peru
LA1473 Callahuanca, Santa Eulalia valley Lima Peru
LA1551 Rimac Valley, Km 71 Lima Peru
LA1552 Rimac Valley, Km 93 Lima Peru
LA1554 Huaral to Cerro de Pasco, Rio Chancay Lima Peru
LA1609 Asia - El Pinon Lima Peru
LA1646 Yaso Lima Peru
LA1647 Huadquina, Topara Ica Peru
LA1653 Uchumayo, Arequipa Arequipa Peru
LA1677 Fundo Huadquina, Topara Ica Peru
LA1694 Cacachuhuasiin, Cacra Lima Peru

73
S. corneliomulleri (L. peruvianum or L. peruv. f. glandulosum)
LA1722 Ticrapo Viejo Huancavelica Peru
LA1723 La Quinga Ica Peru
LA1744 Putinza Lima Peru
LA1910 Tambillo Huancavelica Peru
LA1937 Quebrada Torrecillas Arequipa Peru
LA1944 Rio Atico Arequipa Peru
LA1945 Caraveli Arequipa Peru
LA1973 Yura Arequipa Peru
LA2717 Chilca Lima Peru
LA2721 Putinza Lima Peru
LA2724 Huaynilla Lima Peru
LA2962 Echancay Arequipa Peru
LA2981B Torata to Chilligua en route to Puno Moquegua Peru
LA3154 Otora-Puente Jahuay Moquegua Peru
LA3156 Omate Valley Moquegua Peru
LA3219 Catarindo (Islay) Arequipa Peru
LA3637 Coayllo Lima Peru
LA3639 Ccatac Lima Peru
LA3664 Nazca grade Ica Peru
LA3666 La Yapa Ica Peru
S. galapagense (L. cheesmanii f. minor)
LA0317 Bartolome Galapagos Islands Ecuador
LA0426 Bartolome: E of landing Galapagos Islands Ecuador
LA0436 Isabela: Villamil Galapagos Islands Ecuador
LA0438 Isabela: coast at Villamil Galapagos Islands Ecuador
LA0480A Isabela: Cowley Bay Galapagos Islands Ecuador
LA0483 Fernandina: inside crater Galapagos Islands Ecuador
LA0526 Pinta: W Side Galapagos Islands Ecuador
LA0527 Bartolome: W side, Tower Bay Galapagos Islands Ecuador
LA0528 Santa Cruz: Academy Bay Galapagos Islands Ecuador
LA0530 Fernandina: crater Galapagos Islands Ecuador
LA0532 Pinzon: NW side Galapagos Islands Ecuador
LA0747 Santiago: Cape Trenton Galapagos Islands Ecuador
LA0748 Santiago: E Trenton Island Galapagos Islands Ecuador
LA0929 Isabela: Punta Flores Galapagos Islands Ecuador
LA0930 Isabela: Cabo Tortuga Galapagos Islands Ecuador
LA1044 Bartolome Galapagos Islands Ecuador
LA1136 Gardner-near-Floreana Islet Galapagos Islands Ecuador
LA1137 Rabida: N side Galapagos Islands Ecuador
LA1141 Santiago: N crater Galapagos Islands Ecuador
LA1400 Isabela: N of Punta Tortuga Galapagos Islands Ecuador
LA1401 Isabela: N of Punta Tortuga Galapagos Islands Ecuador

74
S. galapagense (L. cheesmanii f. minor)
LA1403 Fernandina: W of Punta Espinoza Galapagos Islands Ecuador
LA1408 Isabela: SW volcano, Cape Berkeley Galapagos Islands Ecuador
LA1410 Isabela: Punta Ecuador Galapagos Islands Ecuador
LA1411 Santiago: N James Bay Galapagos Islands Ecuador
LA1452 Isabela: E slope, Volcan Alcedo Galapagos Islands Ecuador
LA1508 Corona del Diablo Islet (near Floreana) Galapagos Islands Ecuador
LA1627 Isabela: Tagus Cove Galapagos Islands Ecuador
LA3909 Bartolome: tourist landing Galapagos Islands Ecuador
S. habrochaites (L. hirsutum, L. hirsutum f. glabratum)
LA0094 Canta-Yangas Lima Peru
LA0361 Canta Lima Peru
LA0386 Cajamarca Cajamarca Peru
LA0387 Santa Apolonia Cajamarca Peru
LA0407 Mirador, Guayaquil Guayas Ecuador
LA1033 Hacienda Taulis Lambayeque Peru
LA1223 Alausi Chimborazo Ecuador
LA1252 Loja Loja Ecuador
LA1253 Pueblo Nuevo - Landangue Loja Ecuador
LA1255 Loja (Pedestal district) Loja Ecuador
LA1264 Bucay Chimborazo Ecuador
LA1265 Rio Chimbo Chimborazo Ecuador
LA1266 Pallatanga Chimborazo Ecuador
LA1295 Surco Lima Peru
LA1298 Yaso Lima Peru
LA1347 Empalme Otusco La Libertad Peru
LA1352 Rupe Cajamarca Peru
LA1353 Contumaza Cajamarca Peru
LA1354 Contumaza to Cascas Cajamarca Peru
LA1361 Pariacoto Ancash Peru
LA1362 Chacchan Ancash Peru
LA1363 Alta Fortaleza Ancash Peru
LA1366 Cajacay Ancash Peru
LA1378 Navan Lima Peru
LA1391 Bagua to Olmos Cajamarca Peru
LA1392 Huaraz - Casma Road Ancash Peru
LA1393 Huaraz - Caraz Ancash Peru
LA1557 Huaral to Cerro de Pasco, Rio Chancay Lima Peru
LA1559 Desvio Huamantanga-Canta Lima Peru
LA1560 Matucana Lima Peru
LA1624 Jipijapa Manabi Ecuador
LA1625 S of Jipijapa Manabi Ecuador
LA1648 Above Yaso Lima Peru

75
S. habrochaites (L. hirsutum, L. hirsutum f. glabratum)
LA1681 Mushka Lima Peru
LA1691 Yauyos Lima Peru
LA1695 Cacachuhuasiin, Cacra Lima Peru
LA1696 Huanchuy to Cacra Lima Peru
LA1717 Sopalache Piura Peru
LA1718 Huancabamba Piura Peru
LA1721 Ticrapo Viejo Huancavelica Peru
LA1731 Rio San Juan Huancavelica Peru
LA1736 Pucutay Piura Peru
LA1737 Cashacoto Piura Peru
LA1738 Desfiladero Piura Peru
LA1739 Canchaque to Cerran Piura Peru
LA1740 Huancabamba Piura Peru
LA1741 Sondorilla Piura Peru
LA1753 Surco Lima Peru
LA1764 West of Canta Lima Peru
LA1772 West of Canta Lima Peru
LA1775 Rio Casma Ancash Peru
LA1777 Rio Casma Ancash Peru
LA1778 Rio Casma Ancash Peru
LA1779 Rio Casma Ancash Peru
LA1918 Llauta Ayacucho Peru
LA1927 Ocobamba Ayacucho Peru
LA1928 Ocana Ayacucho Peru
LA1978 Colca Ancash Peru
LA2092 Chinuko Chimborazo Ecuador
LA2098 Sabianga Loja Ecuador
LA2099 Sabiango to Zozoranga Loja Ecuador
LA2100 Sozorango Loja Ecuador
LA2101 Cariamanga Loja Ecuador
LA2103 Lansaca Loja Ecuador
LA2104 Pena Negra Loja Ecuador
LA2105 Jardin Botanico, Loja Loja Ecuador
LA2106 Yambra Loja Ecuador
LA2107 Los Lirios Loja Ecuador
LA2108 Anganumo Loja Ecuador
LA2109 Yangana #1 Loja Ecuador
LA2110 Yangana #2 Loja Ecuador
LA2114 San Juan Loja Ecuador
LA2115 Pucala Loja Ecuador
LA2116 Las Juntas Loja Ecuador
LA2119 Saraguro Loja Ecuador
LA2124 Cumbaratza Zamora-Chinchipe Ecuador

76
S. habrochaites (L. hirsutum, L. hirsutum f. glabratum)
LA2128 Zumbi Zamora-Chinchipe Ecuador
LA2144 Chanchan Chimborazo Ecuador
LA2155 Maydasbamba Cajamarca Peru
LA2156 Ingenio Montan Cajamarca Peru
LA2158 Rio Chotano Cajamarca Peru
LA2159 Atonpampa Cajamarca Peru
LA2167 Cimentario Cajamarca Cajamarca Peru
LA2171 El Molino Piura Peru
LA2174 Rio Chinchipe, San Augustin Cajamarca Peru
LA2175 Timbaruca Cajamarca Peru
LA2196 Caclic Amazonas Peru
LA2204 Balsapata Amazonas Peru
LA2314 San Francisco Amazonas Peru
LA2321 Chirico Amazonas Peru
LA2324 Leimebamba Amazonas Peru
LA2329 Aricapampa La Libertad Peru
LA2409 Miraflores Lima Peru
LA2552 Las Flores Cajamarca Peru
LA2556 Puente Moche La Libertad Peru
LA2567 Quita Ancash Peru
LA2574 Cullaspungro Ancash Peru
LA2648 Santo Domingo Piura Peru
LA2650 Ayabaca Piura Peru
LA2651 Puente Tordopa Piura Peru
LA2722 Puente Auco Lima Peru
LA2812 Lambayeque Lambayeque Peru
LA2855 Mollinomuna, Celica Loja Ecuador
LA2860 Cariamanga Loja Ecuador
LA2861 Las Juntas Loja Ecuador
LA2863 Macara Loja Ecuador
LA2864 Sozorango Loja Ecuador
LA2869 Matola-La Toma Loja Ecuador
LA2975 Coltao Ancash Peru
LA2976 Huangra Ancash Peru
LA3794 Alta Fortaleza Ancash Peru
LA3796 Anca, Marca Ancash Peru
LA3854 Llaguén La Libertad Peru
LA3862 Purunuma Loja Ecuador
LA3863 Sozoranga Loja Ecuador
LA3864 Yangana Loja Ecuador
LA4137 Barrio Delta, Cajamarca Cajamarca Peru

77
S. huaylasense (L. peruvianum)
LA0110 Cajacay Ancash Peru
LA1358 Yautan Ancash Peru
LA1364 Alta Fortaleza Ancash Peru
LA1365 Caranquilloc Ancash Peru
LA1981 Vocatoma Ancash Peru
LA1982 Huallanca Ancash Peru
LA1983 Rio Manta Ancash Peru
LA2068 Chasquitambo Ancash Peru
LA2561 Huallanca Ancash Peru
LA2562 Canon del Pato Ancash Peru
LA2563 Canon del Pato Ancash Peru
LA2575 Valle de Casma Ancash Peru
LA2808 Huaylas Ancash Peru
LA2809 Huaylas Ancash Peru
S. juglandifolium
LA2120 Sabanilla Zamora-Chinchipe Ecuador
LA2134 Tinajillas Zamora-Chinchipe Ecuador
LA2788 Quebrada La Buena Antioquia Colombia
LA3322 Quito Pinchincha Ecuador
LA3323 Manuel Cornejo Astorga Pichincha Ecuador
LA3324 Sabanillas Zamora-Chinchipe Ecuador
LA3325 Cosanga Napo Ecuador
LA3326 Sicalpa Chimborazo Ecuador
S. lycopersicoides
LA1964 Chupapalca Tacna Peru
LA1966 Palca Tacna Peru
LA1990 Palca Tacna Peru
LA2385 Chupapalca to Ingenio Tacna Peru
LA2386 Chupapalca Tacna Peru
LA2387 Lago Aricota (Tarata) Tacna Peru
LA2407 Arica to Putre Tarapaca Chile
LA2408 Above Putre Tarapaca Chile
LA2730 Moquella Tarapaca Chile
LA2772 Zapahuira Tarapaca Chile
LA2776 Catarata Perquejeque Tarapaca Chile
LA2777 Putre Tarapaca Chile
LA2781 Desvio a Putre Tarapaca Chile
LA2951 Quistagama Tarapaca Chile
LA4018 Lago Aricota Tacna Peru
LA4123 Camina Tarapaca Chile
LA4126 Camina - Nama Tarapaca Chile

78
S. lycopersicoides
LA4130 Pachica (Rio Camarones) Tarapaca Chile
LA4131 Esquina Tarapaca Chile
LA4320 Rio Lluta Tarapaca Chile
LA4322 Quebrada Cardones Tarapaca Chile
LA4323 Putre Tarapaca Chile
LA4326 Cochiza, Rio Camarones Tarapaca Chile
S. lycopersicum (L. esculentum var. cerasiforme)
LA0168 New Caledonia Fr. Oceania
LA0292 Santa Cruz Galapagos Islands Ecuador
LA0349 Unknown
LA0384 Chilete (Rio Jequetepeque) Cajamarca Peru
LA0475 Sucua Morona-Santiago Ecuador
LA0476 Sucua Morona-Santiago Ecuador
LA1025 Oahu: Wahiawa Hawaii USA
LA1203 Ciudad Vieja Guatemala
LA1204 Quetzaltenango Guatemala
LA1205 Copan Honduras
LA1206 Copan Ruins Honduras
LA1207 Mexico
LA1208 Sierra Nevada Colombia
LA1209 Colombia
LA1226 Sucua Morona-Santiago Ecuador
LA1227 Sucua Morona-Santiago Ecuador
LA1228 Macas, San Jacinto de los Monos Morona-Santiago Ecuador
LA1229 Macas Plaza Morona-Santiago Ecuador
LA1230 Macas Morona-Santiago Ecuador
LA1231 Tena Napo Ecuador
LA1247 La Toma Loja Ecuador
LA1268 Chaclacayo Lima Peru
LA1286 San Martin de Pangoa Junin Peru
LA1287 Fundo Ileana #1 Junin Peru
LA1289 Fundo Ileana #3 Junin Peru
LA1290 Mazamari Junin Peru
LA1291 Satipo Granja Junin Peru
LA1307 Hotel Oasis, San Francisco Ayacucho Peru
LA1308 San Francisco Ayacucho Peru
LA1310 Hacienda Santa Rosa Ayacucho Peru
LA1311-1 Santa Rosa Puebla Ayacucho Peru
LA1311-10 Santa Rosa Puebla Ayacucho Peru
LA1311-11 Santa Rosa Puebla Ayacucho Peru

79
S. lycopersicum (L. esculentum var. cerasiforme)
LA1311-12 Santa Rosa Puebla Ayacucho Peru
LA1311-13 Santa Rosa Puebla Ayacucho Peru
LA1311-14 Santa Rosa Puebla Ayacucho Peru
LA1311-15 Santa Rosa Puebla Ayacucho Peru
LA1311-16 Santa Rosa Puebla Ayacucho Peru
LA1311-17 Santa Rosa Puebla Ayacucho Peru
LA1311-18 Santa Rosa Puebla Ayacucho Peru
LA1311-19 Santa Rosa Puebla Ayacucho Peru
LA1311-2 Santa Rosa Puebla Ayacucho Peru
LA1311-3 Santa Rosa Puebla Ayacucho Peru
LA1311-4 Santa Rosa Puebla Ayacucho Peru
LA1311-5 Santa Rosa Puebla Ayacucho Peru
LA1311-6 Santa Rosa Puebla Ayacucho Peru
LA1311-7 Santa Rosa Puebla Ayacucho Peru
LA1311-8 Santa Rosa Puebla Ayacucho Peru
LA1311-9 Santa Rosa Puebla Ayacucho Peru
LA1312-2 Paisanato Cusco Peru
LA1312-3 Paisanto Cusco Peru
LA1312-4 Paisanato Cusco Peru
LA1314 Granja Pichari Cusco Peru
LA1320 Hacienda Carmen Apurimac Peru
LA1323 Pfacchayoc Cusco Peru
LA1324 Hacienda Potrero, Quillabamba Cusco Peru
LA1328 Rio Pachachaca Apurimac Peru
LA1334 Pescaderos Arequipa Peru
LA1338 Puyo Napo Ecuador
LA1372 Santa Eulalia Lima Peru
LA1385 Quincemil Cusco Peru
LA1386 Balsas Amazonas Peru
LA1387 Quincemil Cusco Peru
LA1388 San Ramon Junin Peru
LA1420 Lago Agrio Napo Ecuador
LA1421 Santa Cecilia Napo Ecuador
LA1423 Near Santo Domingo Pichincha Ecuador
LA1425 Villa Hermosa Cauca Colombia
LA1426 Cali Cauca Colombia
LA1429 La Estancilla Manabi Ecuador
LA1453 Kauai: Poipu Hawaii USA
LA1454 Mexico
LA1455 Gral Teran Nuevo Leon Mexico
LA1456 Papantla Vera Cruz Mexico
LA1457 Tehuacan Puebla Mexico
LA1458 Huachinango Puebla Mexico

80
S. lycopersicum (L. esculentum var. cerasiforme)
LA1461 University Philippines, Los Banos Philippines
LA1464 El Progreso, Yoro Honduras
LA1465 Taladro, Comayagua Honduras
LA1467 Cali Cauca Colombia
LA1468 Fte. Casa, Cali Cauca Colombia
LA1479 Sucua Morona-Santiago Ecuador
LA1480 Sucua Morona-Santiago Ecuador
LA1481 Sucua Morona-Santiago Ecuador
LA1482 Segamat Malaysia
LA1483 Trujillo Saipan
LA1509 Tawan Sabah Borneo
LA1510 Mexico
LA1511 Siete Lagoas Minas Gerais Brazil
LA1512 Lago de Llopango El Salvador
LA1519 Vitarte Lima Peru
LA1540 Cali to Popayan Cauca Colombia
LA1542 Turrialba Costa Rica
LA1543 Upper Parana Brazil
LA1545 Becan Ruins Campeche Mexico
LA1546 Papantla Vera Cruz Mexico
LA1548 Fundo Liliana Junin Peru
LA1549 Chontabamba Pasco Peru
LA1569 Jalapa Vera Cruz Mexico
LA1574 Nana Lima Peru
LA1619 Pichanaki Junin Peru
LA1620 Castro Alves Bahia Brazil
LA1621 Rio Venados Hidalgo Mexico
LA1622 Lusaka Zambia
LA1623 Muna Yucatan Mexico
LA1632 Puerto Maldonado Madre de Dios Peru
LA1654 Tarapoto San Martin Peru
LA1655 Tarapoto San Martin Peru
LA1662 El Ejido Merida Venezuela
LA1667 Cali Cauca Colombia
LA1668 Acapulco Guerrero Mexico
LA1673 Nana Lima Peru
LA1701 Trujillo La Libertad Peru
LA1705 Sinaloa Mexico
LA1709 Desvio Yojoa Honduras
LA1710 Cariare Limon Costa Rica
LA1711 Zamorano Honduras

81
S. lycopersicum (L. esculentum var. cerasiforme)
LA1712 Pejibaye Costa Rica
LA1713 CATIE, Turrialba Costa Rica
LA1909 Quillabamba Cusco Peru
LA1953 La Curva Arequipa Peru
LA2076 Naranjitos Bolivia
LA2077 Paco, Coroica La Paz Bolivia
LA2078 Mosardas Rio Grande de Sol Brazil
LA2079 Maui: Kihei Hawaii USA
LA2080 Maui: Kihei Hawaii USA
LA2081 Maui: Kihei Hawaii USA
LA2082 Arenal Valley Honduras
LA2085 Kempton Park S. Africa
LA2095 La Cidra Loja Ecuador
LA2121 Yacuambi-Guadalupe Zamora-Chinchipe Ecuador
LA2122A Yacuambi-Guadalupe Zamora-Chinchipe Ecuador
LA2122B Yacuambi-Guadalupe Zamora-Chinchipe Ecuador
LA2122C Yacuambi-Guadalupe Zamora-Chinchipe Ecuador
LA2122D Yacuambi-Guadalupe Zamora-Chinchipe Ecuador
LA2123A La Saquea Zamora-Chinchipe Ecuador
LA2123B La Saquea Zamora-Chinchipe Ecuador
LA2126A El Dorado Zamora-Chinchipe Ecuador
LA2126B El Dorado Zamora-Chinchipe Ecuador
LA2126C El Dorado Zamora-Chinchipe Ecuador
LA2126D El Dorado Zamora-Chinchipe Ecuador
LA2127 Zumbi Zamora-Chinchipe Ecuador
LA2129 San Roque Zamora-Chinchipe Ecuador
LA2130 Gualaquiza Zamora-Chinchipe Ecuador
LA2131 Bomboiza Zamora-Chinchipe Ecuador
LA2135 Limon Santiago-Morona Ecuador
LA2136 Bella Union Santiago-Morona Ecuador
LA2137 Tayusa Santiago-Morona Ecuador
LA2138A Chinimpini Santiago-Morona Ecuador
LA2138B Chinimpini Santiago-Morona Ecuador
LA2139A Logrono Santiago-Morona Ecuador
LA2139B Logrono Santiago-Morona Ecuador
LA2140A Huambi Santiago-Morona Ecuador
LA2140B Huambi Santiago-Morona Ecuador
LA2140C Huambi Santiago-Morona Ecuador
LA2141 Rio Blanco Santiago-Morona Ecuador
LA2142 Cambanaca Santiago-Morona Ecuador
LA2143 Nuevo Rosario Santiago-Morona Ecuador
LA2177A San Ignacio Cajamarca Peru

82
S. lycopersicum (L. esculentum var. cerasiforme)
LA2177B San Ignacio Cajamarca Peru
LA2177C San Ignacio Cajamarca Peru
LA2177E San Ignacio Cajamarca Peru
LA2177F San Ignacio Cajamarca Peru
LA2205A Santa Rosa de Mirador San Martin Peru
LA2205B Santa Rosa de Mirador San Martin Peru
LA2308 San Francisco San Martin Peru
LA2312 Jumbilla #1 Amazonas Peru
LA2313 Jumbilla #2 Amazonas Peru
LA2392 Jakarta Indonesia
LA2393 Mercedes Canton Hoja Ancha Guanacaste Costa Rica
LA2394 San Rafael de Hoja Ancha Guanacaste Costa Rica
LA2402 Florianopolis Santa Catarina Brazil
LA2411 Yanamayo Puno Peru
LA2587 (4x, origin unknown)
LA2616 Naranjillo Huanuco Peru
LA2617 El Oropel Huanuco Peru
LA2618 Santa Lucia, Tulumayo Huanuco Peru
LA2619 Caseria San Augustin Loreto Peru
LA2620 La Divisoria Loreto Peru
LA2621 3 de Octubre Loreto Peru
LA2624 Umashbamba Cusco Peru
LA2625 Chilcachaca Cusco Peru
LA2626 Santa Ana Cusco Peru
LA2627 Pacchac, Chico Cusco Peru
LA2629 Echarate Cusco Peru
LA2630 Calzada Cusco Peru
LA2631 Chontachayoc Cusco Peru
LA2632 Maranura Cusco Peru
LA2633 Huayopata Cusco Peru
LA2635 Huayopata Cusco Peru
LA2636 Sicre Cusco Peru
LA2637 Sicre Cusco Peru
LA2640 Molinopata Apurimac Peru
LA2642 Molinopata Apurimac Peru
LA2643 Bella Vista Apurimac Peru
LA2660 San Ignacio de Moxos Beni Bolivia
LA2664 Yanahuana Puno Peru
LA2665 San Juan del Oro Puno Peru
LA2666 San Juan del Oro Puno Peru
LA2667 Pajchani Puno Peru
LA2668 Cruz Playa Puno Peru
LA2669 Huayvaruni #1 Puno Peru

83
S. lycopersicum (L. esculentum var. cerasiforme)
LA2670 Huayvaruni #2 Puno Peru
LA2671 San Juan del Oro, Escuela Puno Peru
LA2673 Chuntopata Puno Peru
LA2674 Huairurune Puno Peru
LA2675 Casahuiri Puno Peru
LA2683 Consuelo Cusco Peru
LA2684 Patria Cusco Peru
LA2685 Gavitana Madre de Dios Peru
LA2686 Yunguyo Madre de Dios Peru
LA2687 Mansilla Madre de Dios Peru
LA2688 Santa Cruz near Shintuyo #1 Madre de Dios Peru
LA2689 Santa Cruz near Shintuyo #2 Madre de Dios Peru
LA2690 Atalaya Cusco Peru
LA2691 Rio Pilcopata Cusco Peru
LA2692 Pilcopata #1 Cusco Peru
LA2693 Pilcopata #2 Cusco Peru
LA2694 Aguasantas Cusco Peru
LA2696 El Paramillo, La Union Valle Colombia
LA2697 Mata de Cana, El Dovio Valle Colombia
LA2698 La Esperanza de Belgica Valle Colombia
LA2700 Aoti, Satipo Junin Peru
LA2702 Kandy #1 Sri Lanka
LA2709 Bidadi, Bangalore Karnataka India
LA2710 Porto Firme Brazil
LA2782 El Volcan #1 - Pajarito Antioquia Colombia
LA2783 El Volcan #2 - Titiribi Antioquia Colombia
LA2784 La Queronte Antioquia Colombia
LA2785 El Bosque Antioquia Colombia
LA2786 Andes #1 Antioquia Colombia
LA2787 Andes #2 Antioquia Colombia
LA2789 Canaveral Antioquia Colombia
LA2790 Buenos Aires Antioquia Colombia
LA2791 Rio Frio Antioquia Colombia
LA2792 Tamesis Antioquia Colombia
LA2793 La Mesa Antioquia Colombia
LA2794 El Libano Antioquia Colombia
LA2795 Camilo Antioquia Colombia
LA2807 Taypiplaya Yungas Bolivia
LA2811 Cerro Huayrapampa Apurimac Peru
LA2814 Ccascani, Sandia Puno Peru
LA2841 Chinuna Amazonas Peru
LA2842 Santa Rita San Martin Peru
LA2843 Moyobamba mercado San Martin Peru

84
S. lycopersicum (L. esculentum var. cerasiforme)
LA2844 Shanhao San Martin Peru
LA2845 Mercado Moyobamba San Martin Peru
LA2871 Chamaca Sud Yungas Bolivia
LA2873 Lote Pablo Luna #2 Sud Yungas Bolivia
LA2874 Playa Ancha Sud Yungas Bolivia
LA2933 Jipijapa Manabi Ecuador
LA2977 Belen Beni Bolivia
LA2978 Belen Beni Bolivia
LA3135 Pinal del Jigue Holguin Cuba
LA3136 Arroyo Rico Holguin Cuba
LA3137 Pinares de Mayari Holguin Cuba
LA3138 El Quemada Holguin Cuba
LA3139 San Pedro de Cananova Holguin Cuba
LA3140 Los Platanos Holguin Cuba
LA3141 Guira de Melena La Habana Cuba
LA3162 N of Copan Honduras
LA3452 CATIE, Turrialba Turrialba Costa Rica
LA3623 Tablones Manabi Ecuador
LA3633 Botanical garden Ghana
LA3652 Matara Apurimac Peru
LA3842 El Limon, Maracay Araguay Venezuela
LA3843 El Limon, Maracay Aragua Venezuela
LA3844 Algarrobito Guarico Venezuela
LA4133 Makapuu Beach, Oahu Hawaii USA
LA4352 Bamoa Sinaloa Mexico
LA4353 Guasave Sinaloa Mexico
S. neorickii (L. parviflorum)
LA0247 Chavinillo Huanuco Peru
LA0735 Huariaca Huanuco Peru
LA1319 Abancay Apurimac Peru
LA1321 Curahuasi Apurimac Peru
LA1322 Limatambo Cusco Peru
LA1326 Rio Pachachaca Apurimac Peru
LA1329 Yaca Apurimac Peru
LA1626A Mouth of Rio Rupac Ancash Peru
LA1716 Huancabamba Piura Peru
LA2072 Huanuco Huanuco Peru
LA2073 Huanuco, N of San Rafael Huanuco Peru
LA2074 Huanuco Huanuco Peru
LA2075 Huanuco Huanuco Peru
LA2113 La Toma Loja Ecuador
LA2133 Ona Azuay Ecuador

85
S. neorickii (L. parviflorum)
LA2190 Tialango Amazonas Peru
LA2191 Campamento Ingenio Amazonas Peru
LA2192 Pedro Ruiz Amazonas Peru
LA2193 Churuja Amazonas Peru
LA2194 Chachapoyas West Amazonas Peru
LA2195 Caclic Amazonas Peru
LA2197 Luya Amazonas Peru
LA2198 Chachapoyas East Amazonas Peru
LA2200 Choipiaco Amazonas Peru
LA2201 Pipus Amazonas Peru
LA2202 Tingobamba Amazonas Peru
LA2315 Sargento Amazonas Peru
LA2317 Zuta Amazonas Peru
LA2318 Lima Tambo Amazonas Peru
LA2319 Chirico Amazonas Peru
LA2325 Above Balsas Amazonas Peru
LA2403 Wandobamba Huanuco Peru
LA2613 Matichico-San Rafael Huanuco Peru
LA2614 San Rafael Huanuco Peru
LA2615 Ayancocho Huanuco Peru
LA2639A Puente Cunyac Apurimac Peru
LA2641 Nacchera Apurimac Peru
LA2727 Ona Azuay Ecuador
LA2847 Suyubamba Amazonas Peru
LA2848 Pedro Ruiz Amazonas Peru
LA2862 Saraguro-Cuenca Azuay Ecuador
LA2865 Rio Leon Azuay Ecuador
LA2913 Uchucyaco - Hujainillo Huanuco Peru
LA3651 Matara Apurimac Peru
LA3655 Casinchigua to Chacoche Apurimac Peru
LA3657 Casinchigua to Pichirhua Apurimac Peru
LA3660 Murashaya Apurimac Peru
LA3793 Huariaca to San Rafael Huanuco Peru
LA4020 Gonozabal Loja Ecuador
LA4021 Guancarcucho Azuay Ecuador
LA4022 Pueblo Nuevo Azuay Ecuador
LA4023 Paute Azuay Ecuador
S. ochranthum
LA2118 San Lucas Loja Ecuador
LA2160 Acunac Cajamarca Peru
LA2161 Cruz Roja Cajamarca Peru
LA2162 Yatun Cajamarca Peru

86
S. ochranthum
LA2166 Pacopampa Cajamarca Peru
LA2203 Pomacochas San Martin Peru
LA2682 Chinchaypujio Cusco Peru
LA3649 Curpahuasi-Pacaipampa Apurimac Peru
LA3650 Choquemaray Apurimac Peru
S. pennellii (L. pennellii, L. pennellii var. puberulum)
LA0716 Atico Arequipa Peru
LA0750 Ica to Nazca Ica Peru
LA0751 Sisacaya Lima Peru
LA1272 Pisaquera Lima Peru
LA1273 Cayan Lima Peru
LA1275 Quilca road junction Lima Peru
LA1277 Trapiche Lima Peru
LA1282 Sisacaya Lima Peru
LA1297 Pucara Lima Peru
LA1299 Santa Rosa de Quives Lima Peru
LA1302 Quita Sol Ica Peru
LA1303 Pampano Huancavelica Peru
LA1340 Capillucas Lima Peru
LA1356 Moro Ancash Peru
LA1367 Santa Eulalia Lima Peru
LA1376 Sayan Lima Peru
LA1515 Sayan to Churin Lima Peru
LA1522 Quintay Lima Peru
LA1649 Molina Ica Peru
LA1656 Marca to Chincha Ica Peru
LA1657 Buena Vista to Yautan Ancash Peru
LA1674 Toparilla Canyon Lima Peru
LA1693 Quebrada Machurango Lima Peru
LA1724 La Quinga Ica Peru
LA1732 Rio San Juan Huancavelica Peru
LA1733 Rio Canete Lima Peru
LA1734 Rio Canete Lima Peru
LA1735 Rio Canete Lima Peru
LA1809 El Horador (playa) Piura Peru
LA1911 Locari Ica Peru
LA1912 Cerro Locari Ica Peru
LA1920 Cachiruma (Rio Grande) Ayacucho Peru
LA1926 Agua Perdida (Rio Ingenio) Ica Peru
LA1940 Rio Atico, Km 26 Arequipa Peru
LA1941 Rio Atico, Km 41 Arequipa Peru
LA1942 Rio Atico, Km 54 Arequipa Peru

87
S. pennellii (L. pennellii, L. pennellii var. puberulum)
LA1943 Rio Atico, Km 61 Arequipa Peru
LA1946 Caraveli Arequipa Peru
LA2560 Santa to Huaraz Ancash Peru
LA2580 Valle de Casma Ancash Peru
LA2657 Bayovar Piura Peru
LA2963 Acoy Arequipa Peru
LA3635 Omas Lima Peru
LA3665 Ica to Nazca (Rio Santa Cruz) Ica Peru
LA3778 Palpa to Nazca Ica Peru
LA3788 Rio Atico, Km 10 Arequipa Peru
LA3789 Rio Atico, Km 26 Arequipa Peru
LA3791 Caraveli Arequipa Peru
S. peruvianum (L. peruvianum)
LA0098 Chilca Lima Peru
LA0111 Supe Lima Peru
LA0153 Culebras Ancash Peru
LA0370 Hacienda Huampani Lima Peru
LA0371 Supe Lima Peru
LA0372 Culebras #1 Ancash Peru
LA0374 Culebras #2 Ancash Peru
LA0445 Chincha #2 Ica Peru
LA0446 Atiquipa Arequipa Peru
LA0448 Chala Arequipa Peru
LA0453 Yura Arequipa Peru
LA0454 Tambo Arequipa Peru
LA0455 Tambo Arequipa Peru
LA0462 Sobraya Tarapaca Chile
LA0464 Hacienda Rosario Tarapaca Chile
LA0752 Sisacaya Lima Peru
LA1161 Huachipa Lima Peru
LA1270 Pisiquillo Lima Peru
LA1278 Trapiche Lima Peru
LA1300 Santa Rosa de Quives Lima Peru
LA1333 Loma Camana Arequipa Peru
LA1336 Atico Arequipa Peru
LA1337 Atiquipa Arequipa Peru
LA1368 San Jose de Palla Lima Peru
LA1369 San Geronimo Lima Peru
LA1474 Lomas de Camana Arequipa Peru
LA1475 Fundo 'Los Anitos', Barranca Lima Peru
LA1513 Atiquipa Arequipa Peru
LA1517 Irrigacion Santa Rosa Lima Peru

88
S. peruvianum (L. peruvianum)
LA1537 Azapa Valley Tarapaca Chile
LA1556 Hacienda Higuereta Lima Peru
LA1616 La Rinconada Lima Peru
LA1675 Toparilla Canyon Lima Peru
LA1692 Putinza Lima Peru
LA1913 Tinguiayog Ica Peru
LA1929 La Yapana (Rio Ingenio) Ica Peru
LA1935 Lomas de Atiquipa Arequipa Peru
LA1947 Puerto Atico Arequipa Peru
LA1949 Las Calaveritas Arequipa Peru
LA1951 Ocona Arequipa Peru
LA1954 Mollendo Arequipa Peru
LA1955 Matarani Arequipa Peru
LA1975 Desvio Santo Domingo Lima Peru
LA1977 Orcocoto Lima Peru
LA1989 (self-fertile selection, origin unknown)
LA2573 Valle de Casma Ancash Peru
LA2581 Chacarilla (4x) Tarapaca Chile
LA2732 Moquella Tarapaca Chile
LA2742 Camarones-Guancarane Tarapaca Chile
LA2744 Sobraya Tarapaca Chile
LA2745 Pan de Azucar Tarapaca Chile
LA2770 Lluta Tarapaca Chile
LA2834 Hacienda Asiento Ica Peru
LA2955B Quistagama Tarapaca Chile
LA2959 Chaca to Caleta Vitor Tarapaca Chile
LA2964 Quebrada de Burros Tacna Peru
LA3218 Quebrada Guerrero (Islay) Arequipa Peru
LA3220 Cocachacra Arequipa Peru
LA3636 Coayllo Lima Peru
LA3640 Mexico City Mexico
LA3781 Quebrada Oscollo (Atico) Arequipa Peru
LA3783 Rio Chaparra Arequipa Peru
LA3787 Alta Chaparra Arequipa Peru
LA3790 Caraveli Arequipa Peru
LA3795 Alta Fortaleza Ancash Peru
LA3797 Anca, Marca (Rio Fortaleza) Ancash Peru
LA3799 Río Pativilca Ancash Peru
LA3853 Mollepampa La Libertad Peru
LA3858 Canta Lima Peru
LA3900 (CMV tolerant selection)
LA4125 Camina Tarapaca Chile
LA4128 Pachica (Rio Camarones) Tarapaca Chile

89
S. peruvianum (L. peruvianum)
LA4317 Rio Lluta, desembocadura Tarapaca Chile
LA4318 Sora - Molinos, Rio Lluta Tarapaca Chile
LA4325 Caleta Vitor Tarapaca Chile
LA4328 Pachica, Rio Camarones Tarapaca Chile
LA4445 Azapa Valley, 27 km from Arica Tarapaca Chile
LA4446 Azapa Valley, Km 37 from Arica Tarapaca Chile
LA4447 Azapa Valley, Km 27 and Km 37 from Arica Tarapaca Chile
S. pimpinellifolium (L. pimpinellifolium)
LA0100 La Cantuta (Rimac Valley) Lima Peru
LA0114 Pacasmayo La Libertad Peru
LA0121 Trujillo La Libertad Peru
LA0122 Poroto La Libertad Peru
LA0369 La Cantuta (Rimac Valley) Lima Peru
LA0373 Culebras #1 Ancash Peru
LA0375 Culebras #2 Ancash Peru
LA0376 Hacienda Chiclin La Libertad Peru
LA0381 Pongo La Libertad Peru
LA0391 Magdalena (Rio Jequetepeque) Cajamarca Peru
LA0397 Hacienda Tuman Lambayeque Peru
LA0398 Hacienda Carrizal Cajamarca Peru
LA0400 Hacienda Buenos Aires Piura Peru
LA0411 Pichilingue Los Rios Ecuador
LA0412 Pichilingue Los Rios Ecuador
LA0413 Cerecita Guayas Ecuador
LA0417 Puna Guayas Ecuador
LA0418 Daule Guayas Ecuador
LA0420 El Empalme Guayas Ecuador
LA0442 Sechin Ancash Peru
LA0443 Pichilingue Los Rios Ecuador
LA0480 Hacienda Santa Inez Ica Peru
LA0722 Trujillo La Libertad Peru
LA0753 Lurin Lima Peru
LA1236 Tinelandia, Santo Domingo Pichincha Ecuador
LA1237 Atacames Esmeraldas Ecuador
LA1242 Los Sapos Guayas Ecuador
LA1243 Co-op Carmela Guayas Ecuador
LA1245 Santa Rosa El Oro Ecuador
LA1246 La Toma Loja Ecuador
LA1248 Hacienda Monterrey Loja Ecuador
LA1256 Naranjal Guayas Ecuador
LA1257 Las Mercedes Guayas Ecuador
LA1258 Voluntario de Dios Guayas Ecuador

90
S. pimpinellifolium (L. pimpinellifolium)
LA1259 Catarama Los Rios Ecuador
LA1260 Pueblo Viejo Los Rios Ecuador
LA1261 Babahoyo Los Rios Ecuador
LA1262 Milagro Empalme Guayas Ecuador
LA1263 Barranco Chico Guayas Ecuador
LA1269 Pisiquillo Lima Peru
LA1279 Cieneguilla Lima Peru
LA1280 Chontay Lima Peru
LA1301 Hacienda San Ignacio Ica Peru
LA1332 Nazca Ica Peru
LA1335 Pescaderos Arequipa Peru
LA1341 Huampani Lima Peru
LA1342 Casma Ancash Peru
LA1343 Puente Chao La Libertad Peru
LA1344 Laredo La Libertad Peru
LA1345 Samne La Libertad Peru
LA1348 Pacasmayo La Libertad Peru
LA1349 Cuculi Lambayeque Peru
LA1355 Nepena Ancash Peru
LA1357 Jimbe Ancash Peru
LA1359 La Crau Ancash Peru
LA1370 San Jose de Palla Lima Peru
LA1371 Santa Eulalia Lima Peru
LA1374 Ingenio Ica Peru
LA1375 San Vicente de Canete Lima Peru
LA1380 Chanchape Piura Peru
LA1381 Naupe Lambayeque Peru
LA1382 Chachapoyas to Balsas Amazonas Peru
LA1383 Chachapoyas to Bagua Amazonas Peru
LA1384 Quebrada Parca Lima Peru
LA1416 Las Delicias Pichincha Ecuador
LA1428 La Estancilla Manabi Ecuador
LA1466 Chongoyape Lambayeque Peru
LA1469 El Pilar, Olmos Lambayeque Peru
LA1470 Motupe to Desvio Olmos-Bagua Lambayeque Peru
LA1471 Motupe to Jayanca Lambayeque Peru
LA1472 Quebrada Topara Lima Peru
LA1478 Santo Tome (Pabur) Piura Peru
LA1514 Sayan to Churin Lima Peru
LA1520 Sayan to Churin Lima Peru
LA1521 El Pinon, Asia Lima Peru
LA1547 Chota to El Angel Carchi Ecuador
LA1561 San Eusebio Lima Peru

91
S. pimpinellifolium (L. pimpinellifolium)
LA1562 Cieneguilla Lima Peru
LA1571 San Jose de Palle Lima Peru
LA1572 Hacienda Huampani Lima Peru
LA1573 Nana Lima Peru
LA1575 Huaycan Lima Peru
LA1576 Manchay Alta Lima Peru
LA1577 Cartavio La Libertad Peru
LA1578 Santa Marta La Libertad Peru
LA1579 Colegio Punto Cuatro #1 Lambayeque Peru
LA1580 Colegio Punto Cuatro #2 Lambayeque Peru
LA1581 Punto Cuatro Lambayeque Peru
LA1582 Motupe Lambayeque Peru
LA1583 Tierra de la Vieja Lambayeque Peru
LA1584 Jayanca to La Vina Lambayeque Peru
LA1585 Cuculi Lambayeque Peru
LA1586 Zana, San Nicolas La Libertad Peru
LA1587 San Pedro de Lloc La Libertad Peru
LA1588 Laredo to Barraza La Libertad Peru
LA1589 Viru to Galunga La Libertad Peru
LA1590 Viru to Tomaval La Libertad Peru
LA1591 Ascope La Libertad Peru
LA1592 Moche La Libertad Peru
LA1593 Puente Chao La Libertad Peru
LA1594 Cerro Sechin Ancash Peru
LA1595 Nepena to Samanco Ancash Peru
LA1596 Santa to La Rinconada Ancash Peru
LA1597 Rio Casma Ancash Peru
LA1598 Culebras to La Victoria Ancash Peru
LA1599 Huarmey Ancash Peru
LA1600 Las Zorras, Huarmey Ancash Peru
LA1601 La Providencia Lima Peru
LA1602 Rio Chillon to Punchauca Lima Peru
LA1603 Quilca Lima Peru
LA1604 Horcon Lima Peru
LA1605 Canete - San Antonio Lima Peru
LA1606 Tambo de Mora Ica Peru
LA1607 Canete - La Victoria Lima Peru
LA1608 Canete - San Luis Lima Peru
LA1610 Asia - El Pinon Lima Peru
LA1611 Rio Mala Lima Peru
LA1612 Rio Chilca Lima Peru
LA1613 Santa Eusebia Lima Peru
LA1614 Pampa Chumbes Lima Peru

92
S. pimpinellifolium (L. pimpinellifolium)
LA1615 Piura to Simbala Piura Peru
LA1617 Tumbes South Tumbes Peru
LA1618 Tumbes North Tumbes Peru
LA1628 Huanchaco La Libertad Peru
LA1629 Barrancos de Miraflores Lima Peru
LA1630 Fundo La Palma Ica Peru
LA1631 Planta Envasadora San Fernando (Moche) La Libertad Peru
LA1633 Co-op Huayna Capac Ica Peru
LA1634 Fundo Bogotalla #1 Ica Peru
LA1635 Fundo Bogotalla #2 Ica Peru
LA1636 Laran Ica Peru
LA1637 La Calera Ica Peru
LA1638 Fundo El Portillo Lima Peru
LA1645 Banos de Miraflores Lima Peru
LA1651 Vivero, La Molina Lima Peru
LA1652 Cieneguilla Lima Peru
LA1659 Pariacoto Ancash Peru
LA1660 Yautan to Pariacoto Ancash Peru
LA1661 Esquina de Asia Lima Peru
LA1670 Rio Sama Tacna Peru
LA1676 Fundo Huadquina, Topara Ica Peru
LA1678 San Juan Lucumo de Topara Ica Peru
LA1679 Tambo de Mora Ica Peru
LA1680 La Encanada Lima Peru
LA1682 Montalban - San Vicente Lima Peru
LA1683 Miramar Piura Peru
LA1684 Chulucanas Piura Peru
LA1685 Marcavelica Piura Peru
LA1686 Valle Hermosa #1 Piura Peru
LA1687 Valle Hermoso #2 Piura Peru
LA1688 Pedregal Piura Peru
LA1689 Castilla #1 Piura Peru
LA1690 Castilla #2 Piura Peru
LA1697 Hacienda Quiroz, Santa Anita Lima Peru
LA1719 E of Arenillas El Oro Ecuador
LA1720 Yautan Ancash Peru
LA1728 Rio San Juan Ica Peru
LA1729 Rio San Juan Ica Peru
LA1742 Olmos-Marquina Lambayeque Peru
LA1781 Bahia de Caraquez Manabi Ecuador
LA1921 Majarena Ica Peru
LA1923 Cabildo Ica Peru
LA1924 Piedras Gordas Ica Peru

93
S. pimpinellifolium (L. pimpinellifolium)
LA1925 Pangaravi Ica Peru
LA1933 Jaqui Arequipa Peru
LA1936 Huancalpa Arequipa Peru
LA1950 Pescadores Arequipa Peru
LA1987 Viru-Fundo Luis Enrique La Libertad Peru
LA1992 Pichicato Lima Peru
LA1993 Chicama Valley? Lima Peru
LA2093 La Union El Oro Ecuador
LA2096 Playa Loja Ecuador
LA2097 Macara Loja Ecuador
LA2102 El Lucero Loja Ecuador
LA2112 Hacienda Monterrey Loja Ecuador
LA2145 Juan Montalvo Los Rios Ecuador
LA2146 Hacienda Limoncarro La Libertad Peru
LA2147 Yube Cajamarca Peru
LA2149 Puente Muyuno Cajamarca Peru
LA2170 Pai Pai Cajamarca Peru
LA2173 Cruz de Huayquillo Cajamarca Peru
LA2176 Timbaruca Cajamarca Peru
LA2178 Tororume Cajamarca Peru
LA2179 Tamboripa - La Manga Cajamarca Peru
LA2180 La Coipa Cajamarca Peru
LA2181 Balsa Huaico Cajamarca Peru
LA2182 Cumba Amazonas Peru
LA2183 Corral Quemado Amazonas Peru
LA2184 Bagua Amazonas Peru
LA2186 El Salao Amazonas Peru
LA2187 La Caldera Amazonas Peru
LA2188 Machugal #1 Amazonas Peru
LA2189 Machugal #2 Amazonas Peru
LA2335 (4x)
LA2340 (4x)
LA2345 (doubled haploid, origin unknown)
LA2346 (doubled haploid, origin unknown)
LA2347 (doubled haploid, origin unknown)
LA2348 Trujillo La Libertad Peru
LA2389 Tembladera Cajamarca Peru
LA2390 Chungal Cajamarca Peru
LA2391 Chungal to Monte Grande Cajamarca Peru
LA2401 Moxeque Ancash Peru
LA2412 Fundo Don Javier, Chilca Lima Peru
LA2533 Lomas de Latillo Lima Peru
LA2576 Valle de Casma Ancash Peru

94
S. pimpinellifolium (L. pimpinellifolium)
LA2578 Tuturo Ancash Peru
LA2585 (4x, origin unknown)
LA2628 Echarate Cusco Peru
LA2645 Desvio Chulucanas-Morropon Piura Peru
LA2646 Chalaco Piura Peru
LA2647 Morropon-Chalaco Piura Peru
LA2652 Sullana Piura Peru
LA2653 San Francisco de Chocon Querecotillo Piura Peru
LA2655 La Huaca to Sullana Piura Peru
LA2656 Suarez Tumbes Peru
LA2659 Castilla, Univ. Nac. de Piura Piura Peru
LA2718 Chilca Lima Peru
LA2725 Tambo Colorado Ica Peru
LA2805 cv. Indehiscent Currant
LA2831 Rio Nazca Ica Peru
LA2832 Chicchi Tara Ica Peru
LA2833 Hacienda Asiento Ica Peru
LA2836 Fundo Pongo Ica Peru
LA2839 Tialango Amazonas Peru
LA2840 San Hilarion de Tomaque Amazonas Peru
LA2850 Santa Rosa, Manta Manabi Ecuador
LA2851 La Carcel de Montecristo Manabi Ecuador
LA2852 Cirsto Rey de Charapoto Manabi Ecuador
LA2853 Experiment Station, Portoviejo-INIAP Manabi Ecuador
LA2854 Jipijapa Manabi Ecuador
LA2857 Isabela: Puerto Villamil Galapagos Islands Ecuador
LA2866 Via a Amaluza Loja Ecuador
LA2914A Urb. La Castellana, Surco Lima Peru
LA2914B La Castellana, Surco Lima Peru
LA2915 El Remanso de Olmos Lambayeque Peru
LA2934 Carabayllo Lima Peru
LA2966 La Molina Lima Peru
LA2974 Huaca del Sol La Libertad Peru
LA2982 Chilca #1 Lima Peru
LA2983 Chilca #2 Lima Peru
LA3123 Santa Cruz: summit Galapagos Islands Ecuador
LA3158 Los Mochis Sinaloa Mexico
LA3159 Los Mochis Sinaloa Mexico
LA3160 Los Mochis Sinaloa Mexico
LA3161 Los Mochis Sinaloa Mexico
LA3468 La Molina Vieja Lima Peru
LA3634 Santa Rosa de Asia Lima Peru
LA3638 Ccatac Lima Peru

95
S. pimpinellifolium (L. pimpinellifolium)
LA3798 Río Pativilca Ancash Peru
LA3803 Pacanguilla La Libertad Peru
LA3852 Atinchik, Pachacamac Lima Peru
LA3859 TYLCV resistant selection „hirsute‟
LA3910 Near tortoise preserve, Santa Cruz Galapagos Islands Ecuador
LA4027 Olmos-Jaen Road Lambayeque Peru
LA4138 El Corregidor, La Molina Lima Peru
S. sitiens (S. rickii)
LA1974 Chuquicamata Antofagasta Chile
LA2876 Chuquicamata Antofagasta Chile
LA2877 El Crucero Antofagasta Chile
LA2878 Mina La Exotica Antofagasta Chile
LA2885 Caracoles Antofagasta Chile
LA4105 Mina La Escondida Antofagasta Chile
LA4110 Mina San Juan Antofagasta Chile
LA4112 Aguada Limon Verde Antofagasta Chile
LA4113 Estacion Cere Antofagasta Chile
LA4114 Pampa Carbonatera Antofagasta Chile
LA4115 Quebrada desde Cerro Oeste de Paqui Antofagasta Chile
LA4116 Quebrada de Paqui Antofagasta Chile
LA4331 Cerro Quimal Antofagasta Chile

96
Appendix. Wild species core collections. The accession numbers included in the core subsets for each species are listed below. In addition, this table lists accessions in the SolCAP core which are derived from the TGRC (wild species only). The „species sampler‟ subset includes 2-3 accessions from each species group.
S. arcanum
LA0441
LA1346
LA1360
LA1626
LA1708
LA1984
LA2152
LA2163
LA2172
LA2185
LA2326
LA2328
LA2553
S. cheesmaniae
LA0428
LA0429
LA0531
LA1039
LA1041
LA1406
LA1407
LA1409
LA1412
LA1450
S. chilense
LA1930
LA1932
LA1958
LA1960
LA1963
LA1967
LA1969
LA1971
LA2748
LA2750
LA2753
LA2759
LA2765
S. chilense
LA2771
LA2778
LA2880
LA2884
LA2930
LA2946
LA3114
S. chmielewskii
LA1028
LA1306
LA1316
LA1317
LA1325
LA1330
LA2663
LA2677
LA2680
LA2695
LA0103
S. corneliomulleri
LA0107
LA0444
LA1292
LA1305
LA1331
LA1339
LA1647
LA1677
LA1910
LA1937
LA1945
LA1973
S. galapagense
LA0317
LA0438
LA0483
LA0526
LA1136

97
S. galapagense
LA1137
LA1141
LA1401
LA1410
S. habrochaites
LA0407
LA1223
LA1266
LA1347
LA1353
LA1361
LA1363
LA1559
LA1624
LA1718
LA1721
LA1731
LA1753
LA1777
LA1918
LA1928
LA2098
LA2103
LA2109
LA2119
LA2128
LA2155
LA2158
LA2167
LA2174
LA2204
LA2329
LA2409
LA2650
LA2864
S. huaylasense
LA1364
LA1365
LA1982
LA2808
S. lyc. cerasiforme
LA0292
LA1204
S. lyc. cerasiforme
LA1206
LA1228
LA1231
LA1268
LA1286
LA1307
LA1314
LA1320
LA1323
LA1338
LA1385
LA1388
LA1420
LA1425
LA1429
LA1453
LA1456
LA1461
LA1464
LA1482
LA1483
LA1509
LA1511
LA1542
LA1543
LA1620
LA1622
LA2078
LA2095
LA2131
LA2138A
LA2308
LA2392
LA2402
LA2621
LA2670
LA2675
LA2688
LA2709
LA2710
LA2783
LA2845
LA2871

98
S. lyc. cerasiforme
LA4133
LA0247
S. neorickii
LA1319
LA1322
LA1626A
LA1716
LA2113
LA2133
LA2190
LA2198
LA2319
LA2325
S. pennellii
LA0716
LA0751
LA1272
LA1277
LA1356
LA1367
LA1376
LA1656
LA1674
LA1724
LA1732
LA1733
LA1926
LA1946
LA2580
LA2963
S. peruvianum
LA0153
LA0446
LA0752
LA1274
LA1336
LA1474
LA1954
LA2732
LA2744
S. pimpinellifolium
LA0373
LA0400
S. pimpinellifolium
LA0411
LA0417
LA0442
LA1237
LA1245
LA1246
LA1261
LA1279
LA1301
LA1335
LA1371
LA1375
LA1478
LA1521
LA1547
LA1576
LA1578
LA1582
LA1584
LA1586
LA1590
LA1593
LA1599
LA1602
LA1606
LA1617
LA1659
LA1683
LA1689
LA1924
LA1936
LA2102
LA2173
LA2181
LA2183
LA2401
LA2533
LA2852
SolCAP
LA0166
LA0317
LA0373
LA0407

99
SolCAP
LA0422
LA0438
LA0446
LA0716
LA0722
LA1028
LA1037
LA1141
LA1208
LA1237
LA1246
LA1269
LA1272
LA1274
LA1282
LA1283
LA1290
LA1301
LA1314
LA1322
LA1331
LA1338
LA1340
LA1346
LA1406
LA1455
LA1457
LA1464
LA1478
LA1512
LA1542
LA1545
LA1547
LA1549
LA1569
LA1578
LA1582
LA1589
LA1617
LA1620
LA1621
LA1623
LA1632
SolCAP
LA1654
LA1656
LA1668
LA1674
LA1701
LA1712
LA1732
LA1777
LA1809
LA1912
LA1926
LA1930
LA1941
LA1946
LA1953
LA1963
LA1973
LA2076
LA2077
LA2078
LA2093
LA2099
LA2126A
LA2131
LA2135
LA2137
LA2163
LA2181
LA2184
LA2185
LA2190
LA2308
LA2312
LA2411
LA2533
LA2560
LA2561
LA2626
LA2632
LA2633
LA2660
LA2663
LA2664

100
SolCAP
LA2675
LA2744
LA2779
LA2788
LA2792
LA2852
LA2880
LA2930
LA2932
LA2951
LA3136
LA3137
LA3650
LA3795
LA4331
Species Sampler
LA0528
LA0716
LA0722
LA1037
LA1223
LA1226
LA1274
LA1293
LA1326
LA1589
LA1777
LA1926
LA1932
LA1982
LA2150
LA2663
LA2884
LA2930
LA3661

101
Membership List Aarden, Harriette Monsanto Holland BV, Dept. Tomato Breeding.
Leeuwenhoekweg 52, CZ Bergschenhoek, 2661 [email protected]
Alger, Hillary Johnny's Selected Seeds, USA [email protected] ARC Veg and Orn Plant Inst. Atanassiva, Bistra Institute of Genetics, Prof. D. Kostov, BAS, Plovdivsko
Choss 13km, Sofia, BULGARIA, 1113 [email protected] Augustine, Jim BHN Research/ BHN Seed, PO Box 3267, Immokalee, Fl, USA,
34143 [email protected] Beck Bunn, Teresa Monsanto/Seminis, 37437 State Hwy 16, Woodland, CA,
USA, 95695 [email protected] Beckles, Diane M. University of Cal- Davis, Plant Sciences- MS3, One Shields Ave,
Davis, CA, USA, 95616 [email protected] Buonfiglioli, Carlo Della Rimembranze nr. 6A, San Lazzaro di Savena, Bologna, ITALY, 40068 [email protected] Burdick, Allan 3000 Woodkirk Dr., Columbia, MO, USA, 65203 [email protected] California Tomato Research Institute, Inc. Library 18650 E. Lone Tree Rd., Escalon,
CA, USA, 95320-9759 Carli, Stefano Nunhems Italy, via Ghiarone 2, S.Agata, Bolongnese, ITALY,
40019 [email protected] Carrijo Iedo, Valentim Rua Joao Angelo do Pinho 77, Apto 102, Betim, MG, BRAZIL, 32.510-040 [email protected] Chen, Dei Wei Bucolic Seeds Co. Ltd., P.O. 2-39, Tantzu, Taichung Co., TAIWAN, 427 [email protected] Chetelat, Roger University of California, Dept of Veg Crops, One Shields Ave,
Davis, CA, USA, 95616-8746 [email protected] . Cornell University, Albert R Mann Library, Serials Unit/Acquisition Div, Ithaca, NY, USA,
14853

102
Coulibaly, Sylvaine Nunhems USA, 7087 E. Peltier Rd, Acampo, CA, USA, 95220 [email protected] Cuartero, Jesus E.E. LaMayora- CSIC, Plant Breeding Dept., Algarrobo-Costa, Malaga, SPAIN, 29760 [email protected] deHoop, Simon Jan East West Seed Co. Ltd, PO Box 3, Bang Bua Thong, Nonthaburi, THAILAND, 11110 [email protected] Dick, Jim Tomato Solutions, 23264 Mull Rd, Chatham, Ontario, CANADA, N7M 5J4 [email protected] Fernandez-Munoz, Rafael E.E. LaMayora- CSIC, Plant Breeding Dept,
Algarrobo-Costa, Malaga, SPAIN, 29750 [email protected] Fisher, Dr. Dave K. Fisher Farms, 48244 Wesley Chapel Rd, Richfield, NC,
USA, 28137 [email protected] Foolad, Majid Penn State University, Dept. of Horticulture, 102 Tyson Blvd.,
University Park, PA, USA, 16802 [email protected] Fowler, C. Wayne, 2840 70th St SW, Naples, FL, USA, 34105 [email protected] Frank A. Lee Library, NYS Agriculture Experimental Station, 630 W. North St, Geneva,
NY, USA, 14456-1371 Georgiev, Hristo Atanasov Urbanizacion, COSTA JARDIN,
Residencial LA GRACIOSA, C/. LA NUBE 20, Telde, SPAIN, 35215 [email protected]
Gorin, Anthony Technisem, 7 au du Gargliano, Zac des Gatines, FRANCE, 91600 [email protected] Grazzini, Rick GardenGenetics LLC, 131 Mendels Way, Bellefonte, PA, USA, 16823 [email protected] Hanson, Peter AVRDC, PO Box 42, Shanhua, Tainan, TAIWAN,
REPUBLIC of CHINA, 741 [email protected] Hayashi, Masako Yaguchi Asahi Industries, Biol.Engineering Lab, 222 Watarase,
Kamikawa, Kodama-gun, Saitama-ken, JAPAN, 367-0394 [email protected]
Hernandez, Rogelio Harris Moran, 2092 Mission Drive, Naples, FL, USA, 34109 [email protected]

103
Hoogstraten, Jaap Seminis Veg Seeds, Postbus 97, 6700 AB Wageningen, THE NETHERLANDS [email protected]
Hotzev, Amit AB-SEEDS, ltd., P.O. Box 1, Teradion Ind. Zone, D.N. MISGAV, ISRAEL, 20179 [email protected] Hutton, Sam University of Florida, Gulf Coast Research and Education Center, 14625 County Rd 672, Wimuama, FL, USA, 33598 [email protected] Ignatova, Svetlana Box 15, Moscow E-215, RUSSIA, 105215 [email protected] Inai, Shuji Nippon Del Monte Corp., Research and Development, 3748 Shimizu-Cho Numata-shi, Gunma-ken, JAPAN, 378-0016 [email protected] Indian Institute of Horticultural Research, Bangalore, INDIA Jahrmann, Torben Semillas Fito, Centre de biotecnologia, Riera d/Agell, 11, Cabrera de Mar, Barcelona, SPAIN, 8349 [email protected] Johnston, Rob Johnny's Selected Seeds, 955 Benton Ave, Winslow, ME, USA, 4901 [email protected] Kuehn , Michael Harris-Moran Seed Co, 25757 County Rd 21A, Esparto, CA, USA, 95627 [email protected] Lewis, Mark Sakata Seed America, 105 Boronda Rd, Salinas, CA, USA, 93907 [email protected] Liao, Charle Farmer Seed and Ag Co., Ltd., P.O. Box 45, Siu Swei, TAIWAN, 504 [email protected] Liedl, Barbara WVSU, 201 ACEOP Admin Bldg, PO Box 1000, Institute, WV, USA 25112-1000 [email protected] Majde, Mansour Gautier Semences, Route d' Avignon, Eyragues, FRANCE, 13630 [email protected] Maris, Paul DeRuiter Seeds, R&D NL BV, Leeuwenhoekweg 52, Bergschenhoek, THE NETHERLANDS, 2661CZ [email protected] Massoudi, Mark Ag Biotech Inc., P.O. Box 1325, San Juan Bautista, CA, USA,
95045 [email protected] Maxwell, Douglas P. University of WI, Madison, 7711 Midtown Rd, Verona, WI,
USA, 53593 [email protected]

104
McCaslin, Mark FLF Tomatoes, 18591 Mushtown Rd, Prior Lake, MN, USA, 55372 [email protected] McGlasson, Barry University of Western Sydney, Centre for Plant and Food Science, Locked Bag 1797, Penrith South DC, NSW, AUSTRALIA, 1797 [email protected] McGuire, Cate Arcadia Biosciences, Inc. , 220 Cousteau Pl Ste #105, Davis, CA, USA, 95618 Merk, Heather Penn State University., Dept of Horticulture, 103 Tyson Building, University Park, PA, USA, 16802 [email protected] Min, Chai Beijing Vegetable Research Center (BVRC), PO Box 2443, Beijing, PEOPLES REPUBLIC of CHINA, 100089 [email protected] Myers, Jim Oregon State University, Dept. of Horticulture, rm 4017, Ag & Life Sci
Bldg., Corvallis, OR, USA, 97331 [email protected] Nadal, Michael Danson Seed Co, 10851 Woodbine St, Los Angeles, CA, USA, 90034-7675 Nakamura, Kosuke Kagome Co. Ltd., 17 Nishitomiyama, Nasushiobarashi,
Tochigi, JAPAN, 329-2762 [email protected] North Carolina State University, NCSU Library, Campus Box 7111, Raleigh, NC, USA,
27695-0001 Ouyang, Wei Magnum Seeds, Inc., 5825 Sievers Road, Dixon, CA, USA, 95620 [email protected] Ozminkowski , Richard Heinz N.A., PO Box 57, Stockton, CA, USA, 95201 [email protected] Panthee, Dilip R. N.C. State U., Mountain Hort Crops Res & Ext Center,
455 Research Dr, Mills River, NC, USA, 28759 [email protected] Peters , Susan Nunhems USA, 7087 E. Peltier Rd., Acampo, CA, USA, 95220 [email protected] Picard, Madame Florence Vilmorin, Service documentation, Route du Manoir,
La Menitre, FRANCE, 49250 Purdue University Library TSS, Unit Serials, 504 W. State St, West Lafayette, IN, USA,
47907-2058

105
Randhawa, Parm California Seed and Plant Lab, 7877 Pleasant Grove Rd, Elverta, CA, USA, 95626 [email protected] Rascle, Christine Clause Tezier, Domaine de Maninet, Route de Beaumont, Valence, FRANCE, 26000 [email protected] Saito, Atsushi National Institute of Vegetable and Tea Science, 360 Kusawa, Ano, Tsu, JAPAN, 514-2392 [email protected] Sasaki, Seiko Plant Breeding Station of Kaneko Seeds, 50-12,
Furuichi-machi 1-chome, Maebashi City,Gunma, JAPAN, 371-0844 Scott, Jay University of Florida, Gulf Coast Research and Education Center, 14625
County Rd 672, Wimuama, FL, USA, 33598 [email protected] Semences, Gautier Gautier Semences, BP1, 13630, EYRAGUES, FRANCE [email protected] Semillas Fito c/ Selva de Mar 111, Barcelona, SPAIN, 8019 [email protected] Seno, Akiyoshi American Takii Inc., 11492 S. Ave D, Yuma, AZ, USA, 85365 [email protected] Sharma, R.P. University of Hyderabad, Dept. of Plant Sciences, School of Life
Sciences, Hyderabad, INDIA, 500 046 [email protected] Shintaku, Yurie 2-10-2, Shimizu, Suginami-ku, Tokyo, JAPAN, 167-0033 Shupert, David Syngenta Seeds, 10290 Greenway Rd, Naples, FL, USA, 34114 [email protected] Stack, Stephen Colorado State University., Biology, 1878 Campus Delivery, Fort
Collins, CO, USA, 80523-1878 [email protected] Stamova, Liliana 1632 Santa Rosa St., Davis, CA, USA, 95616 [email protected] Stamova, Boryana 2825 Bidwell St, Apt 4, Davis, CA, USA, 95618 Stevens, Mikel Brigham Young University, 275 Widtsoe Bldg, PO. Box 25183,
Provo, UT, USA, 84602 [email protected] Stoeva-Popova, Pravda Winthrop University, Department of Biology, 202 Life
Sciences Building, Rock Hill, SC, USA, 29732 [email protected]

106
Stommel, John USDA-ARS, Genetic Improvement Fruits & Vegetables Laboratory, Bldg. 010A, BARC-West, 10300 Baltimore Ave., Beltsville, MD, USA, 20705 [email protected] Takizawa, Kimiko Japan Horticultural Production and Research Inst., 2-5-1 Kamishiki Matsudo-shi, Chiba, JAPAN, 270-2221 [email protected] Thomas, Paul 4 Juniper Court, Woodland, CA, USA, 95695 Thome, Catherine United Genetics Seeds Co., 764 Carr Ave., Aromas, CA, USA, 65004 [email protected] Tong, Nankui Campbell's Soup Co, Veg. Research & Dev Center, 28605 County
Road 104, Davis, CA, USA, 95691 [email protected] University of California Riverside, Serv/Serials Technical, PO Box 5900, Riverside, CA,
USA, 92517-5900 University of New Hampshire Library, Serials Unit, 18 Library Way, Durham, NH, USA, 03824-3520 University of Wisconsin, Steenbock Library, 550 Babcock Dr, Madison, WI, USA, 53706 [email protected] van Schriek, Marco Keygene N.V., P.O. Box 216, Wageningen,
THE NETHERLANDS, 6700AE [email protected]
Vecchio, Franco Nunhems Italy SRL, Via Ghiarone 2, Sant' Agata, Bolognese (BO), ITALY, 40019 Verbakel, Henk Nunhems Netherlands BV, R& D Library, PO Box 4005, Haelen, THE NETHERLANDS, 6080 AA [email protected] Vinals, Fernando Nuez COMAV, Ciudad Politecnica de la Innovacion, Edificio 8-E.,
Excalera J. 3a Planta, Camino de Vera S/N, Valencia, SPAIN, 46022 [email protected]
Volin, Ray Western Seed Americas, Inc., 15165 Dulzura Ct, Rancho Murieta, CA, USA, 95683-9120 [email protected] Wang, Wendy Xi'an Jinpeng seeds co. ltd., A803 of YuDao Hua Cheng,
No. 8 Feng Cheng 1 road, Xi'an City, Shaan'xi, PR CHINA, 710018 [email protected] WA State University Libraries, SEA Serial record, 100 Diary Rd, Pullman, WA, USA 99164-0001

107
AUTHOR INDEX Chetelat, R. T. 66
Daunay M.C. 6
Fulladolsa, Ana Cristina 41
García, Brenda E. 41
Gilberston, R. L. 54
Hanson P 54
Laterrot H. 54
Lofty, Christopher 58
Maxwell, Douglas P. 41, 54
Mejía, Luis 41, 54
Melgar, Sergio 41
Méndez, Luis 41,54
Rivera, V.V. 54
Sánchez, Amilcar 54
Scott, J.W. 6
Secor, G.A. 54
Smith, Julian 58
Stoeva-Popova, Pravda 58
Teni, Rudy E. 41
Wang J.-F 6