Embed Size (px)
Citation preview

― ―1
日大生活科研報(Rep. Res. Inst. Sci for Liv., Nihon Univ.)36;1~13, 2013 業績 第779号
イ セ エ ビ の 死 後 変 化
三橋 富子※1
・小嶋絵梨花※2
・川久 有紀※3
The Postmortem Changes of a Japanese Spiny Lobster
Tomiko M※1
ITSUHASHI, Erika K※2
OJIMA and Yuki K※3
AWAKU
ABSTRACTThe postmortem changes of a Japanese spiny lobster (Panulirus japonicus) stored at 4℃ were inves-
tigated to find the best time for eating “sashimi”. K value of a Japanese spiny lobster had been kept in very low and even at 96h postmortem it was around 20%, but at that time it already had reached early stage of decomposition accessed by sensory evaluation. The Japanese spiny lobster showed a temporal increase in firmness up to 3h postmortem which subsequently decreased up to 10h postmortem. The amount of IMP, umami nucleotide, was very small during storage time, but another umami nucleotide AMP, increased dramatically up to 24h or 48h postmortem and kept the largest amount in ATP relation compounds during storage. The amounts of glycine, alanine, serine or glutamic acid, which show sweet or umami taste were the largest at 0h postmortem, while histidine, lysine or arginine, which are bitter taste amino acids, increased with the storage time, so the total amount of free amino acid increased up to 96h postmortem. Glycinebe-taine, shows sweet taste, increased up to 10h postmortem and then decreased along storage time.
Considering the conditions of meat texture and taste components, the best time for eating a Japanese spiny lobster as “sashimi” was from 3h to 10h postmortem.
論 文
※1 日本大学短期大学部(三島校舎)教授 Junior College (Mishima Campus), Nihon University, Professor※2 日本大学短期大学部(三島校舎)助手 Junior College (Mishima Campus), Nihon University, Assistant※3 日本大学短期大学部(三島校舎)非常勤助手 Junior College (Mishima Campus), Nihon University, Provisional
Assistant
1.目 的日本人は魚介類を好み、中でもエビはよく消費される食材の1つである。新鮮なエビを刺身や寿司など生で食べることが多いが、生食するに当たり鮮度が重要になってくる。魚介類の死後の鮮度変化は一般に畜肉に比べて進行が速いことが知られているが、中でもエビ・カニ類(甲殻類)の鮮度変化は硬骨魚に比べて早いと言われている。死後の比較的早期における鮮度低下の指標として
K値が用いられているが、K値はアデノシン三リン酸(ATP)関連物質の組成比から算出されている。ATPとその関連化合物の死後に起こる分解過程においてエビは魚類と異なり、アデノシン一リン酸(AMP)
以降の分解経路が2つある。1つは、魚類と同じくイノシン酸(IMP)を経由する経路と、もうひとつはアデノシン(AdR)を経由する分解が起こるといわれている1)。また、このATP関連物質の中のIMPとAMPはうま味の成分としても知られている。IMP、AMPと同様に遊離アミノ酸であるグルタミン酸もうま味の成分として知られているが、エビの遊離アミノ酸は、グリシン、アルギニン、プロリンを多量に含み、これらのアミノ酸の組み合わせが味を作り出している。グリシンが多いほど美味であり、グリシン、アラニンが甘味、アルギニンとグルタミン酸がカニらしさを与える成分であると言う説もある。また、グリシンベタインもカニらしい味を形成すると

― ―2
言われており、エビの場合でも味に対し何らかの役割を果たしていると考えられている2)。生魚の美味しさはうま味成分とともに、テクス
チャーも関係が深い。魚介類は死後数分から十数時間で硬直し、硬直継続時間は5~₂₂時間であると言われている3)。刺身や寿司など魚介類を生食する場合は、まず第一に衛生学的な鮮度が重要であるが、同時に生の魚介類のテクスチャーと、味に関与する化学物質の量が最も良好な状態で食すことが望ましいと考えられる。そこで、静岡県の下田市で蓄養されているイセエビを試料とし、死後硬直の進行と核酸関連物質の変化との関連、刺身としての美味しさという観点から呈味物質としてのアミノ酸、ベタイン類、の定量、鮮度判定の指標としてpHとK値、エビ肉のテクスチャー・硬さの測定、また、物性の変化に関与すると思われる高分子たんぱく質の変化を電気泳動で検討し、生食に最適な時間の検討を試みた。
2.方 法2.1 試 料試料は下田市で蓄養されているイセエビ(Panulirus
japonicus)とし、重量6₀₀g前後のものを使用した。活イセエビを購入し即殺後、採取した腹部筋肉は、0時間(以下0hと記す)、3時間(以下3h)、6時間(以下6h)、₁₀時間(以下₁₀h)、₂₄時間(以下₂₄h)、₄₈時間(以下₄₈h)、₉6時間(以下₉6h)4℃で保存し測定した。供試エビは各時間とも3個体を用いた。
2.2 p H 測 定エビの腹部筋肉にpHメーター(TOHO Model IQ₁₅₀)の電極を差し込み測定した。3~5ヶ所測定し、平均値を求めた。
2.3 レオメーターによる硬さおよび凝集性の測定エビの腹部筋肉を5mm角に切断し、レオメーター
(FUDOHNRM-₂₀₁₀J-CW)の咀嚼試験を用いて測定する。測定条件は、クリアランス₁.₅mm、プランジャー径3₀mmの平丸型、レンジ₂₀₀₀g、テスト速度6cm/minで、圧縮は筋繊維と直角に背側から行った。サンプルは、1尾につき5個ずつ測定し、平均値を求めた。
2.4 各種成分定量用試料の調製エビ肉2gに冷₁₀%過塩素酸(PCA)を4ml加え、氷冷下で2分間ホモジナイズ(KINEMATIKA社製 POLYTORON PT3₁₀₀)した後、冷却遠心分離して上清を氷中のビーカーに入れる。沈殿物に冷₁₀%PCAを4ml加え、ホモジナイズと遠心分離を3回繰り返す。3回の上清を合わせKOHにて中和し、氷冷下に静置して、十分に過塩素酸カリウムの沈殿を生成させた後、沈殿を冷却遠心分離して取り除き、蒸留水で₂₅mlに定容した。₀.₄₅μmフィルターでろ過し、サンプルチューブに入れて-₈₅℃で冷凍保存し、使用時に室温解凍した。
2.5 HPLCによるATPおよびATP関連物質の定量前述の2.4で作成した試料を用いた。
〈分析条件〉装置:二液グラジェントコントローラー(SSC-66₀₀ センシュー科学)カラム:STR ODS-Ⅱ(₄.6mmφ×₁₅₀mm)移動相:₁₀₀mMりん酸(トリエチルアンモニウム)緩衝液(pH6.₈)/アセトニトリル=₁₀₀/₁(v/v)温度:₄₀℃、流量:₁.₀ml/min、検出器波長₂6₀nm、試料注入量:₂₀μl標準物質は、ATP、ADP、AMP、IMP、HxR、Hx、AdR(和光純薬)を用いた。₀.₁mg/mlの溶液を等量ずつ混合し、各標準物質が₁₀μg/mlになるように希釈した。各ATP関連物質の定量とK値は下記の式で求めた。各ATP関連物質(μmol/g)=サンプル当該ATP関連物質の面積÷標準物質のATP関連物質の面積×標準物質の濃度(₁₀μg/ml)×標準物質の純度÷分子量×₂₅ml÷2gK値=(HxR+Hx)/(ATP+ADP+AMP+IMP+AdR+HxR+Hx)×₁₀₀%
2.6 アミノ酸分析システムによる遊離アミノ酸定量試料は2.4で作成した試料に同量のアミノ酸希釈
液を加えたものを用いた。〈分析条件〉装置:島津高速液体クロマトグラフ -R₇A/LC-₁₀Aアミノ酸分析システムカラム:Shim-pack Amino-Na移動相:アミノ酸分析用移動相キットNa型(P/N₂₂₈-₂₁₁₉₅-₉₄)反応液:アミノ酸分析キットOPA試薬(P/N₂₂₈-₂₁₁₉₅-₉3)

― ―3
流量:₀.3ml/min、励起波長:3₅₀nm、蛍光波長:₄₅₀nm、温度:6₀℃、試料注入量:₁₀μl試料希釈液:(₀.₂NNa+(くえん酸三ナトリウム)pH₂.₂₀)標準物質:アミノ酸混合液(和光純薬製H型)と和光純薬製タウリンを混合し、各アミノ酸₀.₁μmol/mlのものを使用。遊離アミノ酸(μmol/ml)=サンプル当該アミノ酸の面積÷標準物質のアミノ酸の面積×標準物質濃度(₀.₁μmol/ml)×2(希釈倍率)×₂₅ml÷2gで算出した。
2.7 HPLCによるベタイン類の定量2.7.1 グリシンベタインの定量試料は2.4で作成した試料液をイオン交換カラ
ム、アンバーライトCG-₄₀₀(酢酸型)とアンバーライト IRC-₅₀(H+型)の1:1の混合物に通して前処理し、以下の分析条件にて分析した。装置:RI-₂₀₀₀検出器カラム:Inertsil NH₂(ジーエル サイエンス)移動相:アセトニトリル:水=6₅:3₅(v/v)溶液温度:₄₀℃、流量:₁.₀ml/min、試料注入量:₂₀μlグリシンベタイン標準液:和光純薬(株)のグリシンベタイン₀.₀₁gを₁₀mlに定容して原液を調整。グリシンベタイン(mg/g)=サンプルの面積÷標準物質のグリシンベタインの面積×標準物質の濃度(1mg/ml)×標準物質の純度×₂₅ml÷2gで算出した。2.7.2 ホマリンおよびトリゴネリンの定量試料は2.4で作成した試料を用い、以下の分析条
件で分析した。装置:二液グラジェントコントローラー(SSC-66₀₀ センシュー科学)カラム:SCX-₁₂₅3-P (センシュー科学)移動相:₀.₀3Mりん酸二水素カリウム(pH₂.₁)温度:₄₀℃、流量:₁.₀ml/min、検出器波長:₂6₂nm、試料注入量:₂₀μl標準液:トリゴネリン(和光純薬)₀.₀₁gを₁₀mlに定容したものを標準原液とし、₁₀倍希釈、₁₀₀倍希釈のものを使用。ホマリンはトリゴネリンと分子量が同一で、溶出位置は違うが濃度と面積は同一と考えて下記の同一式より算出した。ホマリンまたはトリゴネリン(mg/g)=サンプル面積÷標準物質のトリゴネリンの面積×標準物質の濃度(₀.₁mg/mlまたは₀.₀₁mg/ml)×標準物質の純度×₂₅ml÷2gで算出した。
2.8 SDS‐ポリアクリルアミドゲル電気泳動2.8.1 電気泳動サンプルの調製エビ肉₀.₂gをホモジナイザーカップに入れ、可溶
化液(2%SDS、₁₀mMりん酸buffer-pH₇.₀)3.₈₈mlおよび、酵素阻害剤PMSF、ロイペプチン、E-6₄(各₁₀₀μg/ml)を各₀.₀₄mlずつ加えた。ホモジナイザー(井内盛栄堂)でホモジナイズ(₂₀₀₀rpm、₁₀ストローク)して可溶化した後、遠心分離して不溶物を除いた。上澄み液1mlにトラッキングダイ(3₀mM Tris-HCl pH₈.₀ ₀.3mM、EDTA、3%SDS、3₀%グリセロール、₀.₀₀₁%ピロニンY)₀.₅ml、2-メルカプトエタノール₀.₁mlを加え、混合し、₅₀℃で₂₀分間加熱した。この試料液は凍結保存(-₈₅℃)し、使用時に₅₀℃で₂₀分間加熱解凍して用いた。2.8.2 ゲルの調整:
上記の組成で作った3~5%グラディエントアクリルアミドゲルを3₇℃で重合させた。2.8.3 電気泳動条件装置:Mini-PROTEAN Ⅱ cell(日本バイオ・ラッド・ラボラトリー(株))泳動buffer:₅₀mMトリス,₀.3₈₄Mグリシン,₀.₁%SDS,₁₀mM2-メルカプトエタノール溶液泳動サンプル量:₂₀μl泳動:3.₅mA/ミニスラブゲル1枚₁₇~₂₀時間染色・脱色:₀.₁% コマジーブリリアントブルー,₄₀%エタノール,7%酢酸溶液で一昼夜染色し、脱色液(₄₀%エタノール,7%酢酸溶液)で脱色した。
3.実験結果および考察3.1 pHの変化
pHの経時変化を図1に示した。pHは₂₄hまでに急激に低下して極限pHに達していた。その後₄₈h以降に徐々にpHは上昇し始めたが、初期のpHまでには達しなかった。活魚の筋肉はpH₇.₂~₇.₄とほぼ中性であるが、死後は嫌気的な条件下で筋肉中のグリコーゲンが乳酸に分解され、生成された乳酸が蓄積することによっ
ゲル組成〈4枚分〉 3% 5%
3₀%アクリルアミド ₂.₄ml ₄.₀ml
₀.₂M EDTA ₀.₂₄ml ₀.₂₄ml
₂₀% SDS ₀.₁₂ml ₀.₁₂ml
₁.₅M Tris-HCl(pH₈.₀) ₄.₈ml ₄.₈mlTEMED ₀.₀₁6ml ₀.₀₁6ml
₁₀%過硫酸アンモニウム ₀.₂₄ml ₀.₂₄ml
蒸留水 ₁6.₂ml ₁₁.₀ml
グリセロール ― 3.6ml

― ―4
てpHが低下する。最も下がった時のpHが極限pHである。一般にグリコーゲンの含有量によってpHの低下程度は異なり、白身魚では6.₀~6.₄程度以下には下がらないが、赤身魚は₅.₄~₅.6まで低下する。乳酸の蓄積により低下したpHは、細菌の繁殖によってアンモニアやトリメチルアミンなどが生成するため再び上昇する4)。しかし、ATPが消費されるときに生成するプロトンの増加によってpHが低下するという説の方が現在では有力である5)。
3.2 硬さおよび凝集性の変化硬さの経時変化を図2に示した。硬さは、0hか
ら3hにかけて上昇し、3hで最も硬く、その後₁₀hにかけて低下し、以後₉6hまで大きな変化はなかった。凝集性の経時変化を図3に示した。凝集性は6hまで上昇した後、₁₀hにかけて低下し、₂₄hまでは変化なく、以後上昇した。イセエビの死後の物性の変化は各個体の値の相違が大きく、標準偏差も大きいため有意差は認められなかったが、硬さと凝集性の全体的な変化から見て、死後3hでは死後硬直を反映して肉が硬くなっていたと考えられる。
3.3 ATPおよびATP関連物質の変化ATP関連物質は主として筋肉の運動エネルギーを
供給するATPと、それから生じるADP、AMP、IMP、HxR、Hxである。ATP、ADP、AMPまでが筋肉のエネルギー代謝と関係するため、死後、魚種に関わりなく急速に低下するのは当然である。また、ATP関連物質のIMPおよびAMPは遊離アミノ酸のグルタミン酸と同様にうま味成分であると考えられており1)、ATP関連物質の消長は肉のうま味や鮮度と関連があると報告されている6)。
IMP以降のもう一つの分解経路の産物であるアデノシンも同時に定量したが、3検体のうち、1検体しかアデノシンが検出されなかった。また、上述の物性値の変化から、死後変化の個体差が大きいことが推測されたので、3検体それぞれのATP関連物質の経時変化を図4、5、6に示した。試料中のATP関連物質の溶出時間は、標準物質の溶出時間とよく一致していた。松本はクルマエビの即殺直後のATPは6.₄μmol/gで2日目に3.3μmol/gになり₁₁日目に消失、AMPおよびIMPは即殺直後にはほとんど検出されなかったが、速やかに上昇し2日目にそれぞれ2μmol/gであり、AMPは4日以降、IMPは5日以降に減少し、HxRおよびHxは即殺直後存在しなかったが、4日目までに徐々に増加し、Hxのみ4日以降に急激
な増加がみられたと報告している7)。本研究では、3検体において、ATP関連物質の消長はかなり異なっていた。即殺直後のATPは試料Aで₀.₀3μmol/g、試料Bで₄.3₈μmol/g、試料Cで6.₁3μmol/gであり、試料Aを除いては、前述の結果と近い値であった。即殺が致死条件であったが、試料Aはうまく即殺できず、苦悶死して暴れたため、ATPが消耗していたのではないかと考えられる。ATP関連物質の総量に対するATPの割合は試料Bで₅6%、試料Cで₇3%を占めていた。このことから、本研究で用いたイセエビは輸送で疲労したものではなく、きわめて生きのよいものであると判断できた。試料Aは、0hからAMPの蓄積が多く、₂₄hには減少していた。また、試料Aでのみアデノシンが検出されたが、経時的な変化は見られなかった。試料Bは、ATPの消失が₉6hと遅く、試料Cは₂₄hにかけて急速に減少し、₄₈hにはほぼ消失していた。試料B、CともにATPの分解に伴いAMPが増加し、₉6hまで高濃度で残存していた。即殺後のIMPは₀.₀₉~₀.3₀μmol/gと低値であり、3検体とも₉6hに上昇のきざしは見えるが、多量の蓄積は見られなかった。一般に魚類ではIMPが、軟体動物ではAMPが蓄積するが、エビ類については、クルマエビ7)ではAMPとIMPが同程度蓄積し、バナメイエビ1)
では低温ではIMPよりAMPの方が多く蓄積すると報告されている。しかしながら、本研究のイセエビではIMPは保存初期にはほとんど存在せず、AMPの蓄積が目立っていた。このことから、エビの種類によって蓄積するATP分解物が違う可能性が考えられた。また、3検体においてHxはHxRより生成量が少ないため、イセエビにおいてはHxRが蓄積すると考えられた。さらにATP関連物質の終期の産物である、HxRおよびHxの生成は極めて緩慢で、生成量も少なかった。
3.4 K値の変化魚の鮮度を判定する方法としては、細菌学的方法、化学的方法あるいは誘電率の変化などを利用した物理的な方法などがあるが、初期の鮮度の指標としてよく用いられるものにK値がある。これは死後の時間経過とともに筋肉中のATP(アデノシン三リン酸)が順次ADP→AMP→IMP→HxR→Hxへと変化していくことに基づくもので、ATP関連物質の全体に占めるHxRとHx量の合計の比によって鮮度を判定する。K値は死後の時間経過に伴って徐々に上昇する8)。エビの場合AMP以降の分解が、IMPを経由する経路とは別にアデノシンを経由する経路もあるといわれ

― ―5
ている。そこで、本研究で測定したATPおよびATP関連物質の分析値からK値を算出した。また、アデノシンが検出された検体については、アデノシンを加えたATP関連物質の全体に占めるHxRとHx量の合計の比で算出した。K値の経時変化を図7に示した。3検体とも₄₈hまではほぼ一定であり、₉6hで急激に上昇していた。K値の上昇過程は魚種や保存温度、致死条件、季節、部位によって異なるが、即殺魚は₁₀%以下であり、K値が₂₀%であれば刺身として極めて良好な鮮度8)といえる。イセエビ3検体は、₄₈hまでは₁₀%以下(試料Aの0hを除き)であり、新鮮な値を示しているが、₉6h後には急速に上昇していた。K値は₉6hでも2検体が₂₀%以下で生食可能であると考えられるが、官能的には、3検体とも₉6hには鼻につく強いエビ臭と微かな腐敗臭が感じられ、ぬめりも認められたため、生食可能な状態ではなく、初期腐敗の状態であったと思われる。エビ類においては、官能的な初期腐敗に達した時点から急激なK値の増加が起こるとされており1)、₉6hにK値が急上昇した結果とよく一致している。そのため、今回のイセエビでは、₄₈hまではK値からみても新鮮な状態であったと言える。
3.5 遊離アミノ酸の変化味を決める成分の1つとして遊離アミノ酸がかかわっている。エビ類の筋肉エキスに含まれる遊離アミノ酸について、グリシンがもっとも多く、ついでアルギニン、プロリン、セリン、アラニンが多いと報告されている9)。また、即殺直後のクルマエビの遊離アミノ酸の総量は₄₀₁₉mg/₁₀₀gであり、主要な遊離アミノ酸はグリシンが最も多く(総量の₄3%)、次いでアルギニン、プロリン、グルタミン酸、アラニン、タウリンで、これらで総量の₉₇%を占めている。これらの遊離アミノ酸は2日目までに減少し、アミノ酸総量も大きく減少したが、4日目にプロリン、グリシン、アラニン、アルギニンが増加し、総量も大きく増加したと報告されている7)。今回測定したイセエビの遊離アミノ酸(図8)においても、グリシンが₁₀μmol/g前後と多く、次いでアルギニン、セリン、アラニンの順になり(プロリンは今回の分析条件では検出できない。)一致していた。ただヒスチジンが₁₀μmol/g前後と最も多く、また即殺以降のリジンの急激な増加の二点は一致していなかった。遊離アミノ酸の総量は経時的に増加傾向にあるが、₄₈hに減少し、₉6hにグリシン、アラニン、アルギニンが増加し、総量も増加していた。これは前述のク
ルマエビの経時変化と一致していた。しかしながら、本実験の遊離アミノ酸総量は約₅₀μmol/g前後で、同じイセエビの遊離アミノ酸組成から算出した₂₁₀mol/g9)に比べて少なく、充分に抽出されていなかった可能性が考えられた。
3.6 ベタインの変化魚介類の組織中で量的に多い有機塩基は、第4級アンモニウム塩基化合物および尿素である。前者にはベタイン類、トリメチルアミン、トリメチルアミオキシドなどが含まれる。ベタイン類には多くの種類(鎖状ベタインと環状ベタイン)があるが、よく知られているグリシンベタインは鎖状ベタインに、ホマリン、トリゴネリンは環状ベタインに属している6)。グリシンベタインは一般魚には少なく、海産軟骨魚に比較的多く含まれるが、軟体動物、甲殻類でははるかに上回っている。ホマリンは海産無脊椎動物にのみに比較的多く分布し、軟体動物、節足動物の筋肉に多い₁₀)。一方トリゴネリンはエンドウ、コーヒー豆など植物体に多いが、動物体にも存在が認められている₁₁)。また、味質に関係あるのはグリシンベタインで、甘味を呈し、魚介類の甘味の強さやフレーバーエンハンサー(風味増強剤)としての役割を担っていると報告されている₁₂)。そこで、本研究ではグリシンベタインとホマリン、トリゴネリンを定量した。それぞれの経時変化を図9、₁₀に示した。グリシンベタインは、3hから徐々に増加し₁₀hで最も多かった。その後は徐々に減少していた。イセエビの筋肉中のグリシンベタインの含量は、₉6₀mg/₁₀₀gと報告₁3)されており、本研究では₂₀₀~3₀₀mg/₁₀₀g前後と少ない結果となった。ホマリンは即殺後3hまで減少し、₁₀hまで一定に保ちその後は減少した。トリゴネリンは₁₀hまで減少し、その後は徐々に増加していた。またトリゴネリンはホマリンよりも少ないと報告されている₁3)が、本研究でもホマリンの₁/₁₀以下と少なく、一致していた。ベタイン定量の筋肉抽出液はアミノ酸定量と同様のものを使用しているので、抽出不十分であることがベタインでも確認された。
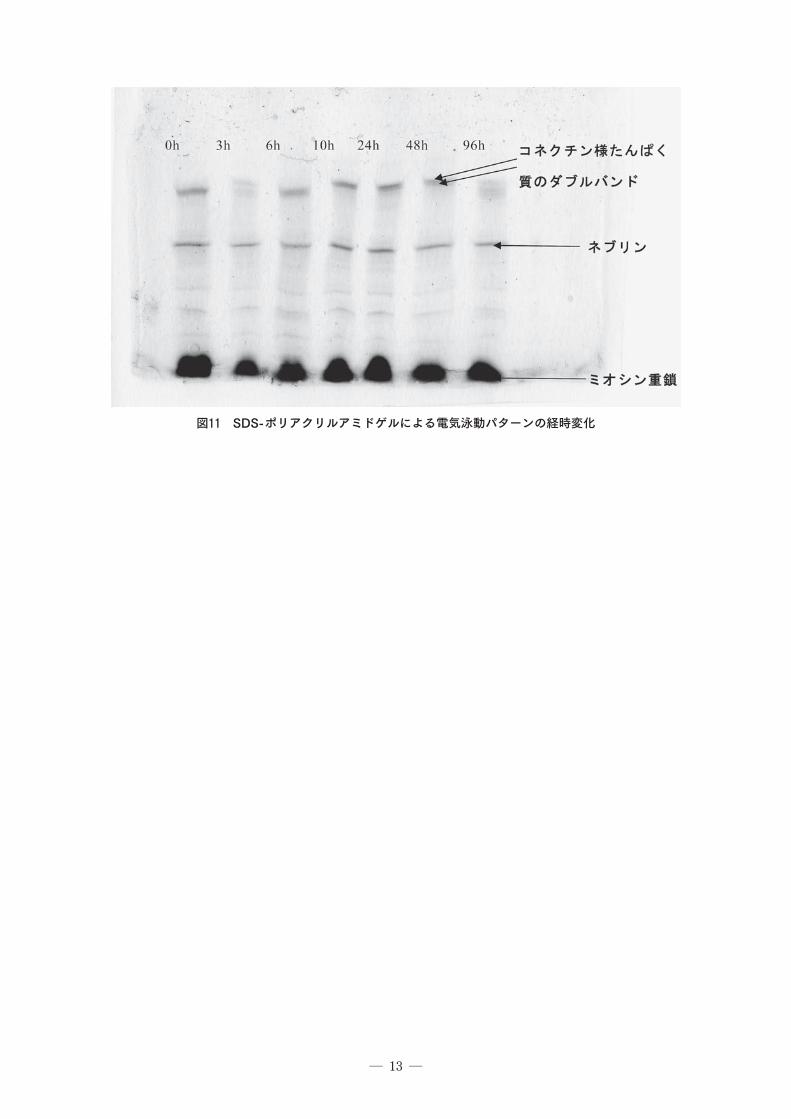
3.7 電気泳動法パターンについてコネクチンは筋原線維のZ線とミオシンフィラメントを連結する長い線維で、肉の解硬・軟化に伴う弾力性の喪失はコネクチンの死後の性状変化と関連していると推定される。コネクチンにはαとβがあり、β-コネクチンはα-コネクチンの分解物である。

― ―6
コネクチンと同様に巨大な分子量を有するネブリンは、細いフィラメントの配列を調整するたんぱく質である。また、ネブリンの消失は筋原線維の構造の脆弱化と関連があると考えられている₁₄)。これらの高分子タンパク質の分解は魚肉の軟化と関係があると考えられる。そこでコネクチンの分解を電気泳動パターンで検討した。各保存時間毎の電気泳動図を₁₁に示した。すべての時間でミオシン重鎖とネブリンが見られた。₄₈hまではコネクチン様たんぱく質のダブルバンドが見られたが、₉6hでは高分子のバンドは見られず、低分子化していた。ネブリンは魚の報告₁₅)の移動度とほぼ一致していたので、同一のものと考えられるが、コネクチン様たんぱく質のダブルバンドは魚と比べると低分子であった。エビ肉の軟化は₁₀hに完了していることからこの巨大たんぱく質の低分子化とは時間的に一致していないため、他の因子が肉の軟化に関係しているのではないかと考えられる。
4.ま と め日本人は新鮮なエビを刺身や寿司など生で食べることが多く、鮮度が重要になってくる。魚介類の死後の鮮度変化は畜肉に比べて進行が速く、生食するに当たり、鮮度には強い関心がもたれている。そこで、イセエビを試料とし、即殺後の経時的な死後変化について検討した。検討項目は、エビの死後硬直の進行と核酸関連物質の変化との関連、刺身としてのおいしさという観点から、肉の硬さと呈味物質としてアミノ酸、ベタイン類量の変化、鮮度の指標としてpHとK値、物性の変化に関与すると思われる高分子たんぱく質の変化についてである。
pHは₂₄hまでに急激に低下して極限pHに達していた。その後₄₈h以降に徐々にpHは上昇し始めたが、初期のpHには達しなかった。硬さは、0hから3hにかけて上昇し、最も硬く、
その後₁₀hまで低下した。物性値は個体差が大きかったので硬さや凝集性に有意差は見られず、また保存期間中の変化も大きくなかった。
ATP関連物質はATPの消失に伴い、うま味物質とされるAMPが増加していた。ATPの消失も個体差が大きく、一個体は即殺直後にATPが消失しており、苦悶死が疑われた。しかしながら、核酸関連物質の消長から算出したK値は3個体とも大きな差は見られず、₄₈hまでは₁₀%以下と低値であった。₄₈hから急激に上昇したが、₉6hにおいても₂₀%前後とK値と
しては可食であったが、官能的には初期腐敗に達していた。
ATP関連物質と同じく呈味に関する遊離アミノ酸は、グリシンやセリン、グルタミン酸などの甘味やうま味を呈するアミノ酸の割合が0hで多かった。₉6hでアミノ酸の総量が増加しているが、その中でも苦味に属するアルギニンやヒスチジン、リジンの増加が多かった。グリシンベタインは₁₀hが最大値であるが、含有
量は₂₀₀~3₀₀mg/₁₀₀g前後と少なかった。ホマリン、トリゴネリンは0hが最も多かった。電気泳動パターンの経時変化では₄₈hまではコネ
クチン様たんぱく質のダブルバンドが見られたが、₉6hでは低分子化していた。エビ肉の軟化は₁₀hに完了していることからこの巨大たんぱく質の低分子化とは時間的に一致していないため、他の因子が肉の軟化に関係しているのではないかと考えられた。これらの結果から、味覚およびテクスチャーを合わせて考えると3h~₁₀hが刺身をおいしく食べることが出来るのではないかと考えられる。テクスチャーでは、3hで一時的な硬さの上昇がみられたが、貯蔵期間中大きな変動は見られなかった。そのため、うま味成分の多い₁₀hが最もおいしく食べられると考えられる。
5.参考文献1) 小山法希、松川雅仁、島田昌彦、佐藤良一:バナメイエビの筋肉中のATP関連化合物の変化と味覚への影響 日水誌 74 pp.₁₀6₈-₁₀₇₄ (₂₀₀₈)
2) 東京水産大学第9回公開講座編集委員会:改訂増補 日本のエビ・世界のエビ (成山堂書店、東京) pp.₁6₂-₁₈3 (₁₉₈6)
3) 小関聡美、北上誠一、加藤登、新井健一:魚介類の死後硬直と鮮度(K値)の変化 東海大学紀要海洋学部 第4巻 第2号 pp.3₁-₄6 (₂₀₀6)
4) 露木英男:食品学各論 (共立出版、東京) p.₂₀₀ (₁₉₉₈)
5) 阿部宏喜:魚の科学 (朝倉書店、東京) p.₄₇ (₁₉₉₇)
6) 坂口守彦:魚介類の含窒素低分子成分とおいしさ 日水誌 67 pp.₇₈₇-₇₉3 (₂₀₀₁)
7) 松本美鈴:甲殻類筋肉の生化学的死後変化とあらい調理に関する研究 東京水産大学博士学位論文 食品生産学 課程博士 甲第33号

― ―7
pp.₁₁-₁₄ (₁₉₉₁)8) 阿部宏喜:魚の科学 (朝倉書店、東京) pp.₄₅-₄6 (₁₉₉₇)
9) 藤田真夫、遠藤金次、清水亘:水産動物肉に関する研究ⅩⅩⅩⅩⅦ クルマエビの筋肉中のエキス成分の季的変化 近畿大学農学部紀要 第5号 pp.6₇-₇3 (₁₉₇₂)
₁₀) 坂口守彦:非タンパク態窒素化合物、魚介類の微量成分(池田静徳編)、恒星社厚生閣、東京 pp.₂-3₁ (₁₉₈₁)
₁₁) 田口寛、阪口宗、嶋林幸英:各種食品のキノリン酸、トリゴネリンおよびN´-メチルニコチンアミドの含量ならびに加熱によるそれらのニコチン酸ニコチンアミドへの変換 ビタミン 60 pp.₅3₇-₅₄6 (₁₉₈6)
₁₂) 笠松千夏:水産軟体動物の食味に関する研究 博士学位論文 お茶の水女子大学大学院 人間文化研究科人間環境学専攻 博甲第3₄₁号 pp.₄3-₅₁ (₂₀₀3)
₁3) 坂口守彦:魚介類のエキス成分 水産学シリーズ 72 pp.₇₉-₈₉ (株)恒星社厚生閣 東京 (₁₉₈₈)
₁₄) 関伸夫:魚類の死後硬直 水産学シリーズ 86 解硬に伴う筋肉の性状変化 p.36 (株)恒星社厚生閣 東京 (₁₉₉₁)
₁₅) 三橋富子:魚肉物性の死後変化におよぼすコネクチン、ネブリンの役割 博士学位論文 お茶の水女子大学大学院 人間文化研究科人間環境学専攻 博甲第₂₈₇号 pp.63-₇₀ (₂₀₀₂)
図1 pHの経時変化
図2 硬さの経時変化

― ―8
図3 凝集性の経時変化
図4 ATP関連物質の経時変化:イセエビA
0 3 6 10 24 48 96Hx 0.19 0.15 0.12 0.22 0.08 0.14 0.23IMP 0.30 0.24 0.30 0.24 0.08 0.13 0.89HxR 0.74 0.55 0.29 0.60 0.18 0.37 1.43AMP 5.96 6.29 4.93 9.53 3.75 5.45 3.27ADP 0.74 0.50 0.55 1.28 0.37 0.54 0.45ATP 0.03 0.02 0.08 0.08 0.06 0.01 0.02AdR 0.03 0.02 0.01 0.27 0.01 0.01 0.01合計 7.98 7.75 6.29 12.23 4.53 6.66 6.30

― ―9
図5 ATP関連物質の経時変化:イセエビB
0 3 6 10 24 48 96Hx 0.11 0.13 0.13 0.08 0.09 0.12 0.26IMP 0.18 0.09 0.17 0.09 0.09 0.12 1.26HxR 0.06 0.09 0.13 0.19 0.18 0.22 0.76AMP 0.78 1.31 1.15 2.01 2.64 3.42 2.21ADP 2.26 2.01 2.12 2.22 1.97 2.14 0.49ATP 4.38 3.29 3.12 2.90 2.45 2.35 0.22合計 7.77 6.93 6.81 7.50 7.42 8.38 5.20
図6 ATP関連物質の経時変化:イセエビC
0 3 6 10 24 48 96Hx 0.35 0.07 0.20 0.08 0.24 0.08 0.19IMP 0.04 0.05 0.13 0.06 0.19 0.05 0.47HxR 0.08 0.04 0.20 0.02 0.46 0.12 0.38AMP 0.20 0.19 4.35 0.31 7.00 2.14 3.45ADP 1.51 1.02 5.02 1.19 2.55 1.74 0.95ATP 6.13 5.16 4.41 1.44 0.70 0.25 0.04合計 8.30 6.52 14.31 3.09 11.14 4.38 5.48

― ―10
図7 K値の経時変化
0 3 6 10 24 48 96K値:A 11.6 9.0 6.6 6.7 5.7 7.6 26.4K値:B 2.2 3.3 3.7 3.6 3.7 4.1 19.6K値:C 5.1 1.7 2.8 3.0 6.3 4.5 10.5

― ―11
図8 遊離アミノ酸の経時変化
0 3 6 10 24 48 96Tau 0.2 0.6 0.4 0.2 0.4 0.2 0.5Thr 0.1 0.1 0.1 0.1 0.1 0.1 0.1Ser 4.3 2.1 1.7 2.2 2.1 2.7 4.0Glu 0.5 0.4 0.3 0.2 0.3 0.0 0.3Gly 10.6 9.4 8.3 8.6 8.5 7.7 9.0Ala 2.7 1.2 0.9 0.9 1.0 0.9 1.3Cys 0.6 0.5 0.5 0.5 0.4 0.4 0.5Val 0.9 0.7 0.6 0.6 0.6 0.5 0.5Met 0.2 0.4 0.4 0.4 0.4 0.5 0.7Ile 0.4 0.4 0.3 0.4 0.3 0.4 0.6Leu 0.3 0.2 0.4 0.3 0.3 0.4 0.4Tyr 2.6 2.3 4.9 3.7 3.0 4.2 3.8Phe 2.9 3.7 5.8 4.2 4.0 4.0 4.4His 10.5 11.2 9.7 10.9 11.3 10.7 16.6Lys 2.1 6.1 5.8 12.6 13.4 12.1 10.6Arg 4.1 6.3 5.9 6.1 7.0 5.1 8.1合計 42.91 45.35 45.96 51.93 53.04 49.66 61.57

― ―12
図9 グリシンベタインの経時変化
0 3 6 10 24 48 96グリシンベタイン 2.20 2.07 2.26 3.12 2.60 2.66 1.93
図10 環状ベタインの経時変化
0 3 6 10 24 48 96ホマリン 1.71 1.20 1.19 1.18 1.43 1.36 1.78トリゴネリン 0.13 0.09 0.08 0.07 0.09 0.09 0.10
― ―13
図11 SDS-ポリアクリルアミドゲルによる電気泳動パターンの経時変化

― ―20
Fig.2 Mitotic metaphases chromosomes of wild and breed Freshwater angelfish (Pterophyllum). ① P. scalare (wild, Peru) ② P. scalare (wild, Surinum) ③ P. scalare (wild, Tefe) ④ P. scalare (breed, common) ⑤ P. scalare (breed, marble) ⑥ P. scalare (breed, diamond) ⑦ P. altum (wild, Negro) ⑧ P. dumerilii (wild, Guiana). A: the metaphase of Giemsa staining; B: Ag-NORʼs. Arrows indicate the position the Ag-NORʼs. Scale indicate in each metaphases 5μm.
⑤-A ⑤-B
⑥-A ⑥-B
⑦-A ⑦-B
⑧-A ⑧-B





























