Embed Size (px)
Citation preview

Transcription and Recombination: When RNAMeets DNA
Andres Aguilera and Helene Gaillard
Centro Andaluz de Biologıa Molecular y Medicina Regenerativa (CABIMER), Universidad de Sevilla,41092 Seville, Spain
Correspondence: [email protected]
A particularly relevant phenomenon in cell physiology and proliferation is the fact thatspontaneous mitotic recombination is strongly enhanced by transcription. The most accept-ed view is that transcription increases the occurrence of double-strand breaks and/or single-stranded DNA gaps that are repaired by recombination. Most breaks would arise as aconsequence of the impact that transcription has on replication fork progression, provokingits stalling and/or breakage. Here, we discuss the mechanisms responsible for the cross talkbetween transcription and recombination, with emphasis on (1) the transcription–replica-tion conflicts as the main source of recombinogenic DNA breaks, and (2) the formation ofcotranscriptional R-loops as a major cause of such breaks. The new emerging questions andperspectives are discussed on the basis of the interference between transcription and repli-cation, as well as the way RNA influences genome dynamics.
Homologous recombination (HR) is a con-served pathway responsible for the repair
of double-strand breaks (DSBs). In mitoticcells, DSBs may be induced by genotoxic agents,such as g irradiation or may occur spontane-ously, in most of the cases in association withreplication. Although HR represents one of thetwo main mechanisms of DSB repair, the otherbeing nonhomologous end joining (NHEJ),eukaryotic cells favor HR as the preferentialDSB repair pathway during the S/G2 phases ofthe cell cycle, when the sister chromatid is avail-able as template for error-free repair. Thus, HRevents are regulated at different steps along thecell cycle, among others by the cyclin-depen-dent kinase 1 during 50-end resection to guar-antee its occurrence at S/G2 (Heyer et al. 2010;
Huertas 2010). Consequently, spontaneous mi-totic recombination events are generally inter-preted as the result of DSB repair during S/G2,although it cannot be disregarded that recom-bination could also be initiated by single-stranded DNA (ssDNA) gaps generated duringreplication.
Spontaneous mitotic recombination might,in principle, take place anywhere in the genomewith similar probability. However, as for mu-tations, both recombination and chromosomebreakages occur more frequently in particularregions, referred to as hot spots. Those hot spotsmight arise from different local features, includ-ing the formation of non-B secondary struc-tures, chromatin compaction, DNA–proteinbarriers to replication, or low-replication initi-
Editors: Stephen Kowalczykowski, Neil Hunter, and Wolf-Dietrich Heyer
Additional Perspectives on DNA Recombination available at www.cshperspectives.org
Copyright # 2014 Cold Spring Harbor Laboratory Press; all rights reserved; doi: 10.1101/cshperspect.a016543
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
1
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

ation density (Aguilera and Garcia-Muse 2013).Nevertheless, the most extended and physio-logical relevant feature enhancing the probabil-ity of recombination is likely to be transcrip-tion (Aguilera 2002; Kim and Jinks-Robertson2012; Gaillard et al. 2013). From bacteria tohumans, a large body of evidence has accumu-lated showing that transcription stimulatesspontaneous recombination, a phenomenon re-ferred to as transcription-associated recombi-nation (TAR). As replication failures seem tobe the main source of recombinogenic DSBs,our actual view is that the major mechanismby which transcription stimulates recombina-tion is via DSBs or ssDNA gaps potentially gen-erated by the difficulties of the DNA replicationfork to progress through transcribed DNA se-quences (Aguilera and Gomez-Gonzalez 2008;Bermejo et al. 2012). Here, we review our actualknowledge of the cross talk between transcrip-tion and recombination with the aim of provid-ing mechanistic insights into the relevance forTAR as a natural source of genome instabilityalong with discussing emerging questions andperspectives.
TRANSCRIPTION STIMULATESRECOMBINATION FROM BACTERIATO HUMAN CELLS
The first evidence of TAR comes from l-phagestudies showing that recombination occurs intranscribed DNA regions and depends on Es-cherichia coli RNA polymerase (RNAP) activitybut not on RecA (Ikeda and Kobayashi 1977;Ikeda and Matsumoto 1979). Probably, themost influential reports on TAR were thoseshowing the identification of the recombinantDNA (rDNA) sequence HOT1 as a hot spot ofrecombination in Saccharomyces cerevisiae (Keiland Roeder 1984), and the following demon-stration that the ability of HOT1 to stimulateRad52-dependent ectopic recombination ofnon-rDNA sequences depends on the RNAPIactivity (Voelkel-Meiman et al. 1987; Stewartand Roeder 1989). Subsequent work on phagetransduction (Dul and Drexler 1988), ille-gitimate recombination in bacterial plasmids(Vilette et al. 1992), RNAPII-mediated recom-
bination between DNA repeats in S. cerevisiae(Thomas and Rothstein 1989), at the ADE6 lo-cus of Schizosaccharomyces pombe (Grimm et al.1991) and in rodent cells (Nickoloff andReynolds 1990) showed that TAR is a conservedfeature. A survey of these and later examples ofTAR from bacteria to mammals can be found ina recent review (Gaillard et al. 2013).
Although the possibility that the impactof transcription on features such as chromatinstructure or the channeling of DNA breaks intodifferent types of repair has been considered anexplanation of TAR, our current understandingfavors TAR being the consequence of an increasein recombinogenic DNA damage caused bytranscription. In yeast, DSBs generated by thehomothallic switching (HO) endonuclease in-ducerecombinationtosimilar levelsregardlessoftranscription being active or not (Weng et al.2000; Gonzalez-Barrera et al. 2002). In mamma-lian cells, transcription did not stimulate DSB-induced recombination (Taghian and Nickoloff1997). DSB-induced recombination and TARevents share similar features of reciprocal versusnonreciprocal events and are dependent on re-combinational DSB repair functions such asRad52 and Rad51 in yeast (Gonzalez-Barreraet al. 2002) or BRCA2 in mammals (Savolainenand Helleday 2009). Recently, human cell linesdefective in XPD, a DNA helicase subunit oftranscription factor II H (TFIIH) required fortranscription initiation and nucleotide excisionrepair, revealed that XPD is required for TAR, butnot for DSB-induced recombination (Savolai-nen et al. 2010). Although it is possible that afraction of TAR events does not initiate viaDSBs, we cannot exclude the possibility thatsome TAR events may also require XPD to beinitiated. All data, therefore, strongly supportthe idea that transcription stimulates recombi-nation by inducing recombinogenic damage atthe transcribed DNA sequence.
In support of this idea, it has been shownthat genotoxic agents such as 4-nitroquino-line 1-oxide or methyl methanesulfonate in-duce gene conversion synergistically with tran-scription (Garcia-Rubio et al. 2003). Althoughthe looser chromatin state of transcribed regionshas been proposed to provide a better substrate
A. Aguilera and H. Gaillard
2 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

for the action of genotoxic agents, there is nodefinitive evidence for this possibility. Also, itdoes not seem that transcription by itself in-creases DNA accessibility to endonucleases orother recombination enzymes, as high tran-scription reduced both HO cleavage efficiencyand strand invasion (Saxe et al. 2000; Gonzalez-Barrera et al. 2002). Instead, and according tothe “twin-supercoiled domain” model (Liu andWang 1987) in which transcription creates neg-ative and positive supercoils behind and aheadof the elongating RNAP, respectively, the prone-ness of the strands of negatively supercoileddouble-stranded DNA (dsDNA) to separatewould make DNA more accessible to genotoxicagents (Fig. 1A).
This model is supported by the reports ontranscription-associated mutagenesis. Mutage-nicity of a variety of external genotoxic agents isstrongly stimulated in actively transcribed DNAsequences (Aguilera 2002; Kim and Jinks-Rob-ertson 2012; Gaillard et al. 2013). Moreover, S.cerevisiae lacking topoisomerase 1 (Top1) activ-
ity undergoes a strong increase in recombina-tion leading to the loss of a marker inserted inthe rDNA (Christman et al. 1988), which hasbeen recently extended to inverted- and direct-repeat systems under the control of RNAPII(Garcia-Rubio and Aguilera 2012). These ob-servations, as well as the hypermutation engen-dered by the top1 mutation in yeast (Kim et al.2011; Lippert et al. 2011; Takahashi et al. 2011),would be consistent with better accessibility ofgenotoxic agents facilitated by negative DNAsupercoiling. Besides, nonrandom mutationclusters probably associated with long ssDNAwere found both in yeast grown under chronicaklylation damage and in human malignant tu-mors (Roberts et al. 2012, 2013; Alexandrov etal. 2013; Burns et al. 2013). These results suggestthat ssDNA patches are formed in proliferativecells, possibly during DSB repair, replication,or R-loop formation (see below). Yet, the factthat recombination is tightly regulated duringthe cell cycle, having its peak during the S/G2
phases, suggests that TAR, like any other form of
(–)
D
(+)
A C
B
(–) (+) (–) (+)
(–) (+)
Figure 1. Transcription intermediates can compromise genome integrity in prokaryotes and eukaryotes. (A) Ineukaryotes, mRNP biogenesis and export are coupled to transcription. Transcribed genes may also be anchoredto the nuclear pore. Negative supercoiling (2) accumulates behind the elongating RNAPII and facilitates theseparation of both strands, making the DNA more susceptible to genotoxic agents. Positive supercoiling (þ)accumulates ahead of the transcription machinery, in front of a head-on oncoming replication fork. (B) Thenascent messenger RNA (mRNA) might hybridize back to its DNA template behind the RNAPII, forming an R-loop. (C) In prokaryotes, cotranscriptional translation of the nascent mRNA may impede the formation of R-loop. (D) Cotranscriptional folding of nonprotein coding RNAs may reduce the probability of R-loop formationin both eukaryotes and prokaryotes.
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 3
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

mitotic recombination, is associated with repli-cation.
R-LOOPS AS MEDIATORS OF TAR
R-loops are structures in which an RNA seg-ment hybridizes with its DNA template, therebydisplacing the complementary strand, whichremains as a single-stranded loop (Fig. 1B). Itsexistence as a physiologically relevant interme-diate has been shown in many systems, includ-ing origin-independent replication in bacteria(Kogoma 1997), class-switching recombination(CSR) (Yu et al. 2003), or E. coli topA mutantsin which transcription-dependent formation ofR-loops behind the RNAP have been genetical-ly inferred (Drolet et al. 1995). More recently,R-loops were found at CpG island promotersand G-rich termination elements in humancells (Skourti-Stathaki et al. 2011; Ginno et al.2012, 2013) and at an Arabidopsis antisensepromoter, where it regulates the expression ofa long noncoding RNA (Sun et al. 2013).
The first demonstration that R-loops gener-ated during transcription induce genome insta-bility in the form of recombination was provid-ed in S. cerevisiae with mutants of the messengerRNA particle (mRNP) biogenesis and exportfactor THO (Huertas and Aguilera 2003). Thehyper-recombination phenotype of these mu-tants not only is transcription-dependent andcan be suppressed by mutations in the RNAPIImachinery that diminish transcription (Santos-Rosa and Aguilera 1995; Fan et al. 1996; Piruatand Aguilera 1996, 1998), but also relies on thenascent RNA molecule and its capacity to forman R-loop behind the elongating DNA polymer-ase (Huertas and Aguilera 2003). The relevanceof R-loops as an intermediate responsible fordifferent forms of instability is, nowadays, sup-ported by an increasing number of reports.These include the demonstration of the forma-tion of R-loops in DT40 chicken and HeLa cellsdepleted of the splicing factor ASF/SF2 (Li andManley 2005) or the transcription-dependentinstability manifested as gross-chromosomalrearrangements, g-H2AX foci, or hyper-recom-bination in yeast, Caenorhabditis elegans, andhuman cell lines mutated in or depleted of a
number of factors involved in RNA metabolism(Gomez-Gonzalez and Aguilera 2007; Gonza-lez-Aguilera et al. 2008; Paulsen et al. 2009; ElHage et al. 2010; Dominguez-Sanchez et al.2011; Wahba et al. 2011; Castellano-Pozo et al.2012; Stirling et al. 2012; Gavalda et al. 2013;Santos-Pereira et al. 2013). Interestingly, dif-ferent reports have provided evidence that ge-nome instability associated with DNA repeatswith the potential to form non-B DNA struc-tures, such as trinucleotide repeats (Grabczyket al. 2007; Lin et al. 2010), or common fragilesites (CFSs) in mammalian cells (Helmrich et al.2011) are also dependent on transcription andassociates with the formation of R-loops. Re-gardless of whether recombination may be trig-gered in such regions or not, the results confirmthat known hot spots of DNA fragility owe theirinstability to a process dependent on transcrip-tion that can be, either partially or completely,related to R-loop accumulation. The RNA, andspecifically the R-loop, has thus become a keyelement in all studies aimed at understandingthe cross talk between transcription and recom-bination (Aguilera and Garcia-Muse 2012).
Top1 restrains R-loop formation by coun-teracting the transient local accumulation ofnegative supercoils at transcribed regions thatfacilitate DNA strand opening (Drolet et al.1995; Tuduri et al. 2009; El Hage et al. 2010;Marinello et al. 2013; Teves and Henikoff 2014).Cotranscriptional mRNP biogenesis and pro-cessing in eukaryotes may also diminish thechances of the nascent RNA to hybridize backwith the template DNA behind the RNAP (Fig.1A,B). The suppression of the transcription-dependent hyper-recombination phenotypes ofTHO and THSC/TREX-2 mutants by overex-pression of heterogeneous nuclear ribonucleo-proteins able to bind RNA, such as Tho1, Nab2,or the RNA dependent on ATPase Sub2/UAP56,whose deletion also causes a strong transcrip-tion-dependent hyper-recombination (Fan et al.2001; Jimeno et al. 2002), would support thisview. We still know little about the structureof the eukaryotic mRNP, the posttranslationalmodifications that RNA-binding factors un-dergo during the RNA cycle, and how they canaffect the flexibility or hybridizing properties of
A. Aguilera and H. Gaillard
4 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

the nascent RNA. A speculative view based onthe known coupling of transcription with RNAprocessing could be that improper mRNP bio-genesis triggers a transcription checkpoint sig-nal that might affect transcription elongationtherefore causing a more stably bound RNAPto the DNA, interfering with other DNA pro-cesses such as transcription-coupled repair orreplication. In bacteria, it has been postulatedthat the nascent RNA is prevented from hybrid-izing with the DNA template by the cotran-scriptional nature of translation that disposesribosomes on the nascent mRNA in protein-en-coding genes (Fig. 1C) (Gowrishankar and Har-inarayanan 2004). The folding of the nascentRNA into secondary structures in nonprotein-encoding genes may fulfill the same function(Fig. 1D).
It is possible that transcription elongationimpairment linked to the formation of R-loopsmight cause TAR. However, the fact that notranscription defect was observed in ASF/SF2-depleted chicken DT40 cells (Li and Manley2005), the existence of THO mutants with atranscription defect that does not lead to in-creased TAR (Huertas et al. 2006), or the lackof correlation between transcription elongationdefects and TAR in a number of mRNA process-ing and export factors (Luna et al. 2005) wouldargue against such a possibility as a generalmechanism. Recently, it has also been conclud-ed that Rad51 is required for DNA–RNA hy-brids formed in trans (Wahba et al. 2013), but itis unclear at this point whether this is a mech-anism related to TAR.
The increasing evidence accumulated on therelevance of R-loops in different forms of ge-nome instability and existence of proteins withthe potential to remove the RNA moiety of R-loops suggest that cotranscriptional R-loopsmay be formed in wild-type cells proficient inmRNP biogenesis and surveillance. The obser-vation that the sen1-1 mutation of the Sen1/SETX putative RNA–DNA helicase, as reportedfor the S. pombe Sen1 (Kim et al. 1999), shows astrong TAR phenotype that correlates with tran-scription and R-loop accumulation argues infavor of this conclusion (Mischo et al. 2011).The questions remaining are whether all TAR
events, occurring in both wild-type and mutantcells, are mediated by R-loops, and whether anR-loop by itself can trigger the formation of aDSB and/or a ssDNA gap responsible for therecombination event.
Recently, a link between R-loops and theH3 S10 phosphorylation chromatin condensa-tion mark was uncovered in yeast, nematodes,and human cells (Castellano-Pozo et al. 2013),suggesting an additional possible mechanismby which R-loop can modulate genome dynam-ics. In the same study, the H3 K9me2 hetero-chromatin mark has also been associated withR-loops in C. elegans. In addition, DNA–RNAhybrids have been shown to mediate RNA in-terference–directed heterochromatin forma-tion at pericentromeric regions of S. pombe(Nakama et al. 2012). Hence, chromatin con-densation or heterochromatin taking place ator around R-loops could interfere with replica-tion and/or transcription, thus triggering geno-mic instability (Castellano-Pozo et al. 2013). Inagreement with this model, CFSs correlate withslow or incomplete replication at regions ofchromatin condensation and cotranscriptionalR-loops (Helmrich et al. 2011; Debatisse et al.2012). It is worth noting, however, that R-loopscoincide with increased chromatin deconden-sation in neurons (Powell et al. 2013). Thus, itseems likely that although the R-loop itselfmight destabilize nucleosomes, chromatin con-densation would occur around and extend fromthe R-loop. Further work will be required tounravel the mechanism by which R-loops alterchromatin structure and how this chromatinconnection modulates TAR.
THE SPECIFIC CASE OF CSR
Whereas TAR represents a global phenomenonof HR, CSR defines a specific type of develop-mentally controlled recombination occurringby NHEJ, not by HR, in vertebrate B cells, whichis worth mentioning here. CSR occurs betweenthe S regions located beside each C segment ofthe immunoglobulin (Ig) genes. Only when theS region is transcribed does it undergo a DSBevent that triggers the NHEJ process responsiblefor CSR (Fig. 2). The first indications that S
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 5
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

regions had the potential to form R-loops weregained using in vitro transcription assays (Rea-ban and Griffin 1990; Reaban et al. 1994; Dan-iels and Lieber 1995); definitive evidence of itsexistence in B cells was provided later by directmapping (Yu et al. 2003). Significantly, howev-er, CSR differs from TAR in important aspects.CSR is a process catalyzed by the activation-induced cytidine deaminase (AID) enzyme,which deaminates deoxycytidines (dCs) gener-ating high levels of deoxyuracils (dUs) that areprocessed by multiple base excision and mis-match repair events, resulting in the occurrenceof DSBs (Xu et al. 2012b). During transcrip-tion-dependent R-loop formation in the S re-gion, the high G content of the displaced DNAstrand would favor G-quadruplex (G4) forma-tion, thus stabilizing the R-loop (Duquette et al.2004). It has also been argued that the high Gcontent of the nascent RNA molecule may con-fer a structural difficulty on its assembly into aproper mRNP, enhancing their options to backhybridize with the DNA template and form anR-loop (Gomez-Gonzalez and Aguilera 2007).Finally, in contrast to TAR (see below), there is
no evidence so far that CSR requires replication.It is also worth noting that AID can induceDSB-mediated translocations like those respon-sible for Burkitt’s lymphoma, with breakpointsin the Ig S region and a G-rich region of c-MYC(Ramiro et al. 2004; Robbiani et al. 2008; Kleinet al. 2011), a phenomenon that has also beenreproduced in yeast THO mutants (Ruiz et al.2011). Therefore, although CSR is a paradig-matic case in which cotranscriptional R-loopsbecome a key substrate for the origin of insta-bility, it is not a model system to understandTAR as a major manifestation of the cross talkbetween transcription and HR.
REPLICATION–TRANSCRIPTIONCONFLICTS AS A SOURCE OF TAR
The way replication–transcription conflicts areresolved has been a main question in biologysince it became clear that most genes are dis-posed codirectionally with replication alongthe E. coli chromosome (Ellwood and Nomura1982). French (1992) reported that the move-ment of the replication fork was reduced in theribosomal operon of E. coli as a result of its hightranscriptional activity. Using an in vitro re-constituted T4 phage replication system, thelaboratory of B. Alberts showed that, whereasthe bacteriophage replication machinery couldovertake a codirectionally moving E. coli RNAPwithout major consequences, a stalled RNAPcaused the pausing of a head-on colliding DNApolymerase (Liu et al. 1993; Liu and Alberts1995). In theF29 bacteriophage, the replicationfork also stalled when the RNAP was arrestedin a head-on orientation (Elias-Arnanz andSalas 1999). In E. coli, the replication machinerywas able to displace a codirectionally elongat-ing RNAP, but not an oncoming RNAP (Pom-erantz and O’Donnell 2008). In the first case,the transient RNA–DNA hybrid formed dur-ing transcription could serve to reinitiate rep-lication (Pomerantz and O’Donnell 2010). Theability of transcription to pause or stall repli-cation in eukaryotes was first shown by 2D-gelelectrophoresis at RNAPIII-driven transcrip-tion in transfer RNA (tRNA) genes (Deshpandeand Newlon 1996). RNAPII transcription of a
Sμ Cμ
AID
AID
Sα
BER/MMR
DSB
NHEJ
Cα
CαSαSμ
Figure 2. CSR. Transcription of switch (S) regions ofthe Ig genes generates R-loops, providing ssDNA sub-strates for the activation-induced cytidine deaminase(AID). Subsequent sequential enzymatic activities ofdifferent DNA repair pathways, including base exci-sion and mismatch repair, would lead to DSBs that,by NHEJ, would complete the CSR event.
A. Aguilera and H. Gaillard
6 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

GAL fusion gene has a stronger effect on head-on replication than codirectional replicationencounters (Prado and Aguilera 2005). Ge-nome-wide evidence that transcription sup-poses an obstacle for the progression of forks,regardless of orientation, has been providedin yeast by determining the distribution of theRrm3 helicase, which works in association withthe replication fork (Azvolinsky et al. 2009).This study does not distinguish whether colli-sions might be stronger or more persistent un-der codirectional versus head-on orientation,but it certainly indicates that in vivo transcrip-tion in either direction may suppose an obstacleto replication fork progression. These and otherobservations, such as that E. coli replicationfork stop at a dG (deoxiguanosine)-dC tandemrepeat in which, normally, the RNAP stalls (Kra-silnikova et al. 1998), support the general con-clusion that transcription has a negative impacton replication, especially when both processesoccur in head-on orientation.
The identification of DNA helicases, suchas T4 dda and gp41 (Bedinger et al. 1983; Liuand Alberts 1995), E. coli DinG, Rep, and UvrD(Boubakri et al. 2010), yeast Rrm3 (Azvolinskyet al. 2006, 2009), and human RecQL5 (Li et al.2011), as well as the E. coli transcription-cou-pled repair factor Mfd (Pomerantz and O’Don-nell 2010), as factors that help bypass head-onstalled RNA polymerases confirms that cells usea battery of proteins to either prevent or resolveputatively harmful collisions. Removal of theseproteins has been shown to enhance recombi-nation in a transcription-dependent manner,supporting the idea that at least one majormechanism of TAR is mediated by transcrip-tion–replication collisions.
Clearly, in vivo evidence that replication islinked to TAR was provided by the observa-tion that transcription from an S-phase activepromoter causes hyper-recombination in yeast,whereas this is not the case of transcriptiondriven from G1-specific promoters (Prado andAguilera 2005). In such assays, the effect wasstronger in head-on collisions systems as com-pared with codirectional ones and stimulatedin the absence of the Rrm3 helicase. Accord-ingly, inversion of the E. coli rrn operon from
a codirectional to a head-on disposition withrespect to replication makes cell viability strictlydependent on the RecBC DSB repair protein(Boubakri et al. 2010). Evidence for the repli-cation dependency of TAR in mammals hasbeen provided in Chinese hamster cells, inwhich replication stress generated by thymidine,which slows down replication fork progression,enhances TAR at the HPRT gene (Gottipati etal. 2008). These and other data (Gaillard et al.2013) suggest that recombination stimulatedby transcription is a consequence of the difficul-ties of the replication fork to traverse throughtranscribed DNA regions. This phenomenonhas also been observed in tRNA genes flankedby direct repeats (de la Loza et al. 2009), as wellas in TAR mediated by R-loops. Thus, TAR inyeast THO complex mutants is only observedunder S-phase transcription, and replicationforks pause or stall in the DNA region in whicha hot spot of RNA–DNA hybrid formation atthe end of a lacZ gene has been identified (Hu-ertas et al. 2006; Wellinger et al. 2006). Indeed,the ability of R-loops to negatively affect pro-gression of replication forks has been confirmedin yeast, C. elegans, and bacteria (Gan et al. 2011;Castellano-Pozo et al. 2012; Santos-Pereira et al.2013). Therefore, it seems that regardless of thetype of TAR detected, whether or not stimulatedin mRNP biogenesis and export mutants or me-diated by R-loops, its origin is linked to a defec-tive replication progression.
THE rDNA REGION AS A PARADIGMOF TAR EVENTS
TAR takes place in the rDNA region, which isorganized in tandem repeats in most organismsto ensure the maintenance of its integrity. In-terestingly, many features underlying TAR arerecapitulated in the rDNA, such as the depend-ency on topoisomerase activities to restrain re-combination and R-loop formation (Kim andWang 1989; El Hage et al. 2010). In eukaryotes,DNA elements with replication fork barrierproperties (RFBs) are found in rDNA regions,as first shown in yeast (Brewer and Fangman1988). They associate with specialized proteinsto exert a programmed polar pausing of repli-
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 7
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

cation, thus avoiding frontal collisions withthe transcriptional machinery (Tsang and Carr2008). Structures with RFB capacity are alsofound at other genomic locations, includingreplication termination sites in E. coli (Khatriet al. 1989; Hill and Marians 1990), the matlocus in S. pombe (Dalgaard and Klar 2001),or transposons (Zaratiegui et al. 2011), indicat-ing that programmed fork block to avoid fron-tal encounters with transcriptional machineriesis a broadly used cellular strategy to avoid geno-mic instability.
In addition to the RFB, another importantmechanism contributing to rDNA stability in-volves the regulated recruitment of cohesin, amultifunctional protein complex that ensuressister chromatid cohesion during replicationand at sites of DSB, thus contributing to pre-vent genomic instability by favoring equal re-combination events (Nasmyth 2011; Dorsettand Strom 2012). In the yeast S. cerevisiae,only about half of the rDNA repeats are activelytranscribed, and repressed rDNA copies havebeen shown to be important for sister chromatidcohesion (Ide et al. 2010). rDNA silencing ismediated by the Sir2 protein and appears tocontrol transcription from E-pro, a bidirec-tional RNAPII promoter located adjacent tothe RFB (Santangelo et al. 1988; Ganley et al.2005). Interestingly, constitutive transcriptionfrom E-pro results in dissociation of cohesinfrom the rDNA spacer, and the proportion ofunequal recombination events was shown to in-crease in a conditional cohesin mutant, lead-ing to an increase in rDNA repeats expansion,marker loss, and extrachromosomal rDNA cir-cle formation (Kobayashi et al. 2004; Kobayashiand Ganley 2005). Hence, the activation of E-pro transcription has been shown to lead tocohesin dissociation at the rDNA. Similarly,transcription activation leads to cohesin delo-calization at other loci in yeast (Lengronneet al. 2004; Bausch et al. 2007). It is thus tempt-ing to speculate that transcription-dependentchanges of sister chromatid cohesion may alsocontribute to TAR. Indeed, the hyper-recombi-nation associated with cohesion mutations hasbeen observed at different loci (De Piccoli et al.2006; Alvaro et al. 2007).
TELOMERES AS A PUTATIVE REGION PRONETO TERRA-MEDIATED TAR
The ends of eukaryotic chromosomes not onlypose the “end-replication” problem, solved inyeast and proliferative tissues by the action ofthe telomerase, but they also need to be pro-tected by the association with specialized pro-teins and formation of a lariat structure, theT-loop, created by the invasion of the 30 ssDNAend into the duplex region to prevent the acti-vation of the DNA damage response (de Lange2009; O’Sullivan and Karlseder 2010). Replica-tion is challenged at telomeres, which, in thissense, resemble fragile sites (Sfeir et al. 2009).Although telomeres are heterochromatin struc-tures, they are transcribed by RNAPII intotelomeric repeat-containing RNA (TERRA)molecules in several eukaryotes from yeast tomammals (Feuerhahn et al. 2010; Bah and Az-zalin 2012). TERRA might interfere with repli-cation, either through collision between thetranscription and replication machineries orbecause TERRA might form R-loops with telo-meric DNA, which could be further stabilizedby G4-DNA structures in the displaced strandor G4-hybrid structures. Evidence in supportof this idea includes the observation that a frac-tion of TERRA remains associated with telo-meric chromatin (Azzalin et al. 2007; Schoeft-ner and Blasco 2008; Deng et al. 2009) anddysfunction of the Rat1 ribonuclease leadsto TERRA accumulation and telomere elonga-tion defects in S. cerevisiae, both phenotypesbeing suppressed by RNase H overexpression(Luke et al. 2008). Furthermore, both telomericDNA and TERRA adopt G4-DNA structures(Phan 2010; Xu et al. 2010; Lu et al. 2013),and RNA–DNA hybrid G4 structures may rep-resent a telomere component according to re-cent modeling studies (Xu et al. 2012a). Theexistence of telomeric R-loops has been recentlyshown in budding yeast in two independentstudies. Interestingly, those telomeric R-loopswere significantly increased in THO mutants,which suffer from short telomeres (Pfeifferet al. 2013), indicating that proper TERRA bio-genesis is important for telomere maintenance.Noticeably, THO subunits were identified at
A. Aguilera and H. Gaillard
8 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

human telomeres as well (Grolimund et al.2013). On the other hand, telomeric R-loopaccumulation was shown to promote recom-bination-mediated elongation events in theabsence of telomerase (Balk et al. 2013), thusbeing an important player in telomere dynam-ics. Further work will be required to dissect therole of telomeric R-loops in the regulation oftelomere function.
FROM TRANSCRIPTION–REPLICATIONCONFLICTS TO RECOMBINATION
The concept that the stalling of the replicationfork by transcription, whether or not mediatedby the formation of R-loops, is a major causeof TAR seems to be generally accepted. How-ever, the mechanism by which a stalled replica-tion fork can lead to the DSBs or ssDNA gapsresponsible for recombination remains unclear.In principle, different mutually nonexclusiveoptions could explain this phenomenon, ofwhich we review three.
First, as discussed above, the accumulationof negative supercoiled DNA during transcrip-tion may increase its susceptibility to endoge-nous genotoxic agents. The resulting DNA le-sions, unless properly repaired, will constitutephysical obstacles for the progression of thereplication fork. As a consequence, an ssDNAstretch would be created in front of the laggingor leading DNA polymerases, and replicationof the gapped region completed by recombina-tional template switching (Fig. 3A). In this case,although the recombination event would be rep-lication-dependent, the transcription machin-ery does not need to be present in the DNAduring replication, and a DSB may not neces-sarily be the initiation intermediate.
Second, during codirectional collisions, thereplication fork hitting the back of the RNAPwould probably pause or strongly reduce itsspeed, presumably without arrest in most cases.Once the elongating RNAP finishes its transcrip-tion cycle, replication could, in principle, resu-me normally. Alternatively, it is also possible thatthe replication fork coming from the other sidefinishes replication. However, the presence of aDNA lesion with the ability to block RNAPelon-
gation or requiring RNAP backtracking to resu-me elongation (Dutta et al. 2011), or mutantsdefective in 30-end RNA processing or transcrip-tion termination (Mischo et al. 2011), may makethis codirectional fork pausing permanent,leading to a replication arrest. A similar situa-tion could occur if an R-loop would haveformed behind the RNAP, in which case the rep-lication fork would hit the RNA:DNA hybriddirectly, presumably causing fork stalling (Fig.3B). Resumption of replication may then re-quire fork breakage, whether or not mediatedby a nuclease, replication restart thus occurringby a recombination-mediated mechanism. Al-ternatively, the hybrid might be used as primerto reinitiate DNA synthesis.
Third, during head-on collisions, replica-tion arrest likely becomes inevitable. It remainsunclear, however, whether or not a physical col-lision between the two polymerases takes place.The positive supercoiling generated between thetwo machineries, and its possible impact on nu-cleosome organization, may produce some sig-nals in response to which cells could block rep-lication fork progression avoiding physicalcollision (Fig. 3B). The observations that thehuman RECQL5 helicase interacts with RNAPIIand functions in transcription repression (Ay-gun et al. 2008, 2009; Kassube et al. 2013)suggest that direct inhibition of transcriptionelongation might provide a means to preventcollisions. RecQL5-depleted mammalian cellssuffer from strong increase in sister chromatidexchanges (Hu et al. 2005). Whether this phe-notype is caused by the RNAPII-regulating ac-tivity of RECQL5, a consequence of a more di-rect role of RecQL5 in DNA metabolism (Huet al. 2007), or a combination of both (Islamet al. 2010) remains yet an open question. It isthus possible that transcription may not behavejust as a protein–DNA barrier to replicationlike natural or even artificial recombinogenicRFBs (Horiuchi and Fujimura 1995; Lambertet al. 2005; Jacome and Fernandez-Capetillo2011). In this sense, genome-wide chromatinimmuno-precipitation (ChIP)-chip analyseshave revealed that Top2 accumulates in regionsin which replication forks encounter highlytranscribed genes (Bermejo et al. 2009). Also,
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 9
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
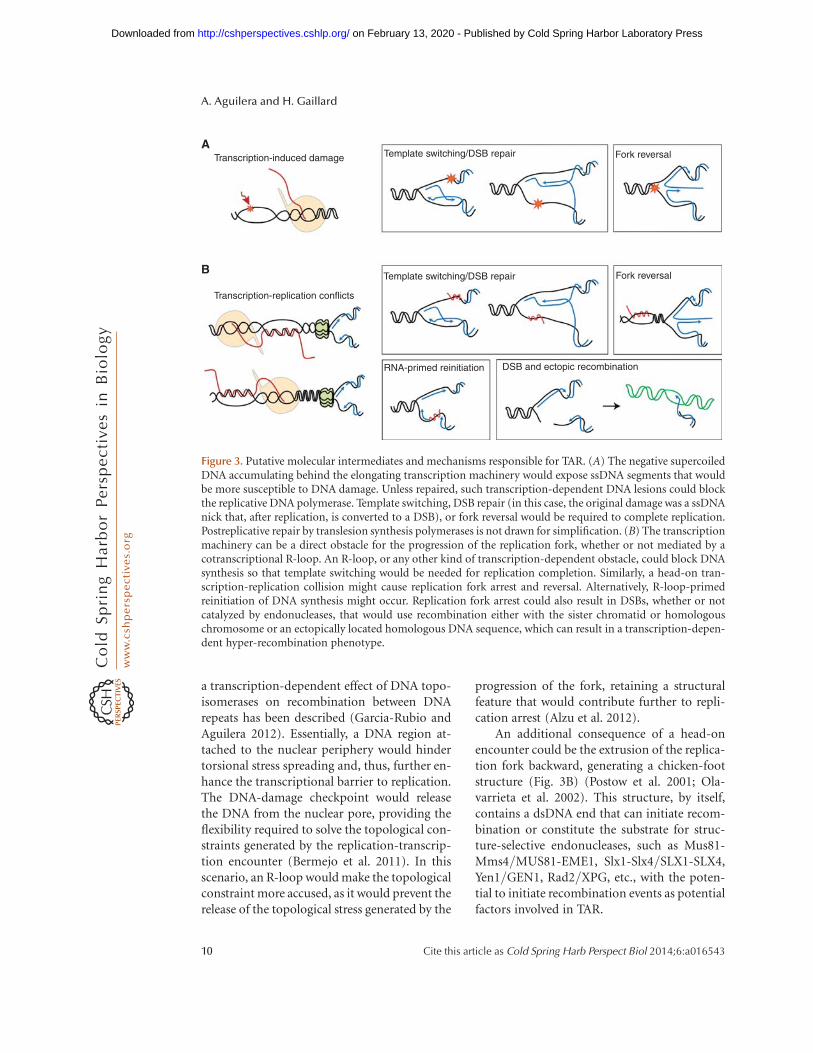
a transcription-dependent effect of DNA topo-isomerases on recombination between DNArepeats has been described (Garcia-Rubio andAguilera 2012). Essentially, a DNA region at-tached to the nuclear periphery would hindertorsional stress spreading and, thus, further en-hance the transcriptional barrier to replication.The DNA-damage checkpoint would releasethe DNA from the nuclear pore, providing theflexibility required to solve the topological con-straints generated by the replication-transcrip-tion encounter (Bermejo et al. 2011). In thisscenario, an R-loop would make the topologicalconstraint more accused, as it would prevent therelease of the topological stress generated by the
progression of the fork, retaining a structuralfeature that would contribute further to repli-cation arrest (Alzu et al. 2012).
An additional consequence of a head-onencounter could be the extrusion of the replica-tion fork backward, generating a chicken-footstructure (Fig. 3B) (Postow et al. 2001; Ola-varrieta et al. 2002). This structure, by itself,contains a dsDNA end that can initiate recom-bination or constitute the substrate for struc-ture-selective endonucleases, such as Mus81-Mms4/MUS81-EME1, Slx1-Slx4/SLX1-SLX4,Yen1/GEN1, Rad2/XPG, etc., with the poten-tial to initiate recombination events as potentialfactors involved in TAR.
Transcription-induced damage
Transcription-replication conflicts
B
A
RNA-primed reinitiation
Template switching/DSB repair
Template switching/DSB repair
DSB and ectopic recombination
Fork reversal
Fork reversal
Figure 3. Putative molecular intermediates and mechanisms responsible for TAR. (A) The negative supercoiledDNA accumulating behind the elongating transcription machinery would expose ssDNA segments that wouldbe more susceptible to DNA damage. Unless repaired, such transcription-dependent DNA lesions could blockthe replicative DNA polymerase. Template switching, DSB repair (in this case, the original damage was a ssDNAnick that, after replication, is converted to a DSB), or fork reversal would be required to complete replication.Postreplicative repair by translesion synthesis polymerases is not drawn for simplification. (B) The transcriptionmachinery can be a direct obstacle for the progression of the replication fork, whether or not mediated by acotranscriptional R-loop. An R-loop, or any other kind of transcription-dependent obstacle, could block DNAsynthesis so that template switching would be needed for replication completion. Similarly, a head-on tran-scription-replication collision might cause replication fork arrest and reversal. Alternatively, R-loop-primedreinitiation of DNA synthesis might occur. Replication fork arrest could also result in DSBs, whether or notcatalyzed by endonucleases, that would use recombination either with the sister chromatid or homologouschromosome or an ectopically located homologous DNA sequence, which can result in a transcription-depen-dent hyper-recombination phenotype.
A. Aguilera and H. Gaillard
10 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

CONCLUDING REMARKS
Despite many years of intensive research on TARusing a large battery of different systems, furthergenetic and molecular analyses are needed toidentify the intermediates involved. For exam-ple, a detailed and comparative analysis of TARusing different substrates leading to potentiallydifferent recombination events would be impor-tant to evaluate the prevalence of the kind ofinitiation event (DSB or ssDNA gap), recombi-nation event (reciprocal or nonreciprocal), andinvolved pathways (break-induced replication,single-strand annealing, synthesis-dependentstrand annealing, or DSB repair) in each case.It is unclear whether spontaneous TAR observedin wild-type cells is similar to that observed inmRNP processing mutants or mediated by R-loops. Although a genome-wide analysis in yeasthas shown that the sites of major accumulationof the Rrm3 helicase are the open reading framesof actively transcribed genes, implying that rep-lication has difficulties in traversing such DNAregions (Azvolinsky et al. 2009), a similar studyon the distribution of g-H2AX signals, as amarker of DSBs, has shown that they accumulatein other regions including tRNAs and promot-ers of protein-encoding genes (Szilard et al.2010). An explanation for this apparent dis-crepancy could be that transcription does notnecessarily induce DSBs, which would be con-sistent with recent work performed in mamma-lian cells (Gottipati et al. 2008) or the proposalthat most TAR events induced in yeast THOmutants, although being replication-dependent,would not occur via DSBs (Gomez-Gonzalezet al. 2009). Indeed, hpr1 mutants show a strongtranscription-dependent hyper-recombinationleading to deletions between direct repeats buta mild increase in other types of events. It hasbeen proposed that a DNA polymerase stalledin front of an R-loop may switch to anotherhomologous region and use it as a template toresume replication, possibly explaining why di-rect-repeat recombination is the most favoredtype of event in R-loop accumulating mutants(Fig. 3B) (Gomez-Gonzalez et al. 2009).
It seems, therefore, important to identifythe molecular event initiating recombination
at transcribed DNA that could explain the dif-ferent TAR events studied so far—by analogywith CFSs, specific chromosomal sites showingbreaks or constrictions in metaphase chromo-somes (Durkin and Glover 2007), which haverecently been shown to occur in very long geneslocated in low-replication initiation regions(Helmrich et al. 2011; Le Tallec et al. 2011; Le-tessier et al. 2011). An interesting and openquestion is the mechanism by which TAR couldbe induced in terminally differentiated cells.It could be that cotranscriptionally formedR-loops interfere with DNA-repair activitiesor transcriptional RNA-binding proteins mighthave a role in DNA repair itself. However,the emerging field involving small noncodingRNAs in the DNA damage response to DSBs(Wei et al. 2012; d’Adda di Fagagna 2013) pointsto novel connections between RNA and themaintenance of genome integrity that wouldneed to be further investigated in relation toTAR. Up to which point the nature of the tran-scriptional intermediates determine the out-come of recombination remains to be seen,but, no doubt, the more we know about thetype of event involved, the more we will learnabout the molecular nature of the cross talkbetween transcription and recombination andtheir connection to replication. This is of ma-jor interest in modern genetics and molecularbiology not only for the increasing relevanceof transcription in genome dynamics and, asrecently proposed, accelerated evolution (Paulet al. 2013), but also because of the importanceof genome integrity maintenance as a mecha-nism to prevent cancer and other genetic dis-eases.
ACKNOWLEDGMENTS
We thank R. Wellinger for helpful discussionsand D. Haun for style supervision. Researchin A.A.’s laboratory is funded by the SpanishMinistry of Economy and Competitiveness,the Junta de Andalucıa, and the EuropeanUnion (FEDER). We apologize to those whosework could not be cited because of space con-straints.
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 11
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

REFERENCES
Aguilera A. 2002. The connection between transcriptionand genomic instability. EMBO J 21: 195–201.
Aguilera A, Garcia-Muse T. 2012. R loops: From transcrip-tion byproducts to threats to genome stability. Mol Cell46: 115–124.
Aguilera A, Garcia-Muse T. 2013. Causes of genome insta-bility. Annu Rev Genet 47: 1–32.
Aguilera A, Gomez-Gonzalez B. 2008. Genome instability: Amechanistic view of its causes and consequences. Nat RevGenet 9: 204–217.
Alexandrov LB, Nik-Zainal S, Wedge DC, Aparicio SA, Beh-jati S, Biankin AV, Bignell GR, Bolli N, Borg A, Borresen-Dale AL, et al. 2013. Signatures of mutational processes inhuman cancer. Nature 500: 415–421.
Alvaro D, Lisby M, Rothstein R. 2007. Genome-wide anal-ysis of Rad52 foci reveals diverse mechanisms impactingrecombination. PLoS Genet 3: e228.
Alzu A, Bermejo R, Begnis M, Lucca C, Piccini D, Carote-nuto W, Saponaro M, Brambati A, Cocito A, Foiani M, etal. 2012. Senataxin associates with replication forks toprotect fork integrity across RNA-polymerase-II-tran-scribed genes. Cell 151: 835–846.
Aygun O, Svejstrup J, Liu Y. 2008. A RECQ5-RNA polymer-ase II association identified by targeted proteomic anal-ysis of human chromatin. Proc Natl Acad Sci 105: 8580–8584.
Aygun O, Xu X, Liu Y, Takahashi H, Kong SE, Conaway RC,Conaway JW, Svejstrup JQ. 2009. Direct inhibition ofRNA polymerase II transcription by RECQL5. J BiolChem 284: 23197–23203.
Azvolinsky A, Dunaway S, Torres JZ, Bessler JB, Zakian VA.2006. The S. cerevisiae Rrm3p DNA helicase moves withthe replication fork and affects replication of all yeastchromosomes. Genes Dev 20: 3104–3116.
Azvolinsky A, Giresi PG, Lieb JD, Zakian VA. 2009. Highlytranscribed RNA polymerase II genes are impediments toreplication fork progression in Saccharomyces cerevisiae.Mol Cell 34: 722–734.
Azzalin CM, Reichenbach P, Khoriauli L, Giulotto E,Lingner J. 2007. Telomeric repeat containing RNA andRNA surveillance factors at mammalian chromosomeends. Science 318: 798–801.
Bah A, Azzalin CM. 2012. The telomeric transcriptome:From fission yeast to mammals. Int J Biochem Cell Biol44: 1055–1059.
Balk B, Maicher A, Dees M, Klermund J, Luke-Glaser S,Bender K, Luke B. 2013. Telomeric RNA-DNA hybridsaffect telomere-length dynamics and senescence. NatStruct Mol Biol 20: 1199–1205.
Bausch C, Noone S, Henry JM, Gaudenz K, Sanderson B,Seidel C, Gerton JL. 2007. Transcription alters chromo-somal locations of cohesin in Saccharomyces cerevisiae.Mol Cell Biol 27: 8522–8532.
Bedinger P, Hochstrasser M, Jongeneel CV, Alberts BM.1983. Properties of the T4 bacteriophage DNA replica-tion apparatus: The T4 dda DNA helicase is required topass a bound RNA polymerase molecule. Cell 34: 115–123.
Bermejo R, Capra T, Gonzalez-Huici V, Fachinetti D, CocitoA, Natoli G, Katou Y, Mori H, Kurokawa K, Shirahige K,et al. 2009. Genome-organizing factors Top2 and Hmo1prevent chromosome fragility at sites of S phase tran-scription. Cell 138: 870–884.
Bermejo R, Capra T, Jossen R, Colosio A, Frattini C, Caro-tenuto W, Cocito A, Doksani Y, Klein H, Gomez-Gonza-lez B, et al. 2011. The replication checkpoint protects forkstability by releasing transcribed genes from nuclearpores. Cell 146: 233–246.
Bermejo R, Lai MS, Foiani M. 2012. Preventing replicationstress to maintain genome stability: Resolving conflictsbetween replication and transcription. Mol Cell 45: 710–718.
Boubakri H, de Septenville AL, Viguera E, Michel B. 2010.The helicases DinG, Rep and UvrD cooperate to promotereplication across transcription units in vivo. EMBO J 29:145–157.
Brewer BJ, Fangman WL. 1988. A replication fork barrier atthe 30 end of yeast ribosomal RNA genes. Cell 55: 637–643.
Burns MB, Temiz NA, Harris RS. 2013. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat Genet45: 977–983.
Castellano-Pozo M, Garcia-Muse T, Aguilera A. 2012. R-loops cause replication impairment and genome insta-bility during meiosis. EMBO Rep 13: 923–929.
Castellano-Pozo M, Santos-Pereira JM, Rondon AG, Bar-roso S, Andujar E, Perez-Alegre M, Garcia-Muse T, Agui-lera A. 2013. R loops are linked to histone H3 S10 phos-phorylation and chromatin condensation. Mol Cell 52:583–590.
Christman MF, Dietrich FS, Fink GR. 1988. Mitotic recom-bination in the rDNA of S. cerevisiae is suppressed by thecombined action of DNA topoisomerases I and II. Cell55: 413–425.
d’Adda di Fagagna F. 2013. A direct role for small non-cod-ing RNAs in DNA damage response. Trends Cell Biol 24:171–178.
Dalgaard JZ, Klar AJ. 2001. A DNA replication-arrest siteRTS1 regulates imprinting by determining the directionof replication at mat1 in S. pombe. Genes Dev 15: 2060–2068.
Daniels GA, Lieber MR. 1995. RNA:DNA complex forma-tion upon transcription of immunoglobulin switch re-gions: Implications for the mechanism and regulation ofclass switch recombination. Nucleic Acids Res 23: 5006–5011.
Debatisse M, Le Tallec B, Letessier A, Dutrillaux B, Brison O.2012. Common fragile sites: Mechanisms of instabilityrevisited. Trends Genet 28: 22–32.
de la Loza MC, Wellinger RE, Aguilera A. 2009. Stimulationof direct-repeat recombination by RNA polymerase IIItranscription. DNA Repair 8: 620–626.
de Lange T. 2009. How telomeres solve the end-protectionproblem. Science 326: 948–952.
Deng Z, Norseen J, Wiedmer A, Riethman H, LiebermanPM. 2009. TERRA RNA binding to TRF2 facilitates het-erochromatin formation and ORC recruitment at telo-meres. Mol Cell 35: 403–413.
A. Aguilera and H. Gaillard
12 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

De Piccoli G, Cortes-Ledesma F, Ira G, Torres-Rosell J, UhleS, Farmer S, Hwang JY, Machin F, Ceschia A, McAleenanA, et al. 2006. Smc5-Smc6 mediate DNA double-strand-break repair by promoting sister-chromatid recombina-tion. Nat Cell Biol 8: 1032–1034.
Deshpande AM, Newlon CS. 1996. DNA replication forkpause sites dependent on transcription. Science 272:1030–1033.
Dominguez-Sanchez MS, Barroso S, Gomez-Gonzalez B,Luna R, Aguilera A. 2011. Genome instability and tran-scription elongation impairment in human cells depletedof THO/TREX. PLoS Genet 7: e1002386.
Dorsett D, Strom L. 2012. The ancient and evolving roles ofcohesin in gene expression and DNA repair. Curr Biol 22:R240–R250.
Drolet M, Phoenix P, Menzel R, Masse E, Liu LF, Crouch RJ.1995. Overexpression of RNase H partially complementsthe growth defect of an Escherichia coli DtopA mutant:R-loop formation is a major problem in the absence ofDNA topoisomerase I. Proc Natl Acad Sci 92: 3526–3530.
Dul JL, Drexler H. 1988. Transcription stimulates recombi-nation: II. Generalized transduction of Escherichia coli byphages T1 and T4. Virology 162: 471–477.
Duquette ML, Handa P, Vincent JA, Taylor AF, Maizels N.2004. Intracellular transcription of G-rich DNAs inducesformation of G-loops, novel structures containing G4DNA. Genes Dev 18: 1618–1629.
Durkin SG, Glover TW. 2007. Chromosome fragile sites.Annu Rev Genet 41: 169–192.
Dutta D, Shatalin K, Epshtein V, Gottesman ME, Nudler E.2011. Linking RNA polymerase backtracking to genomeinstability in E. coli. Cell 146: 533–543.
El Hage A, French SL, Beyer AL, Tollervey D. 2010. Loss oftopoisomerase I leads to R-loop-mediated transcription-al blocks during ribosomal RNA synthesis. Genes Dev 24:1546–1558.
Elias-Arnanz M, Salas M. 1999. Resolution of head-on col-lisions between the transcription machinery and bacter-iophage F29 DNA polymerase is dependent on RNApolymerase translocation. EMBO J 18: 5675–5682.
Ellwood M, Nomura M. 1982. Chromosomal locations ofthe genes for rRNA in Escherichia coli K-12. J Bacteriol149: 458–468.
Fan HY, Cheng KK, Klein HL. 1996. Mutations in the RNApolymerase II transcription machinery suppress the hy-perrecombination mutant hpr1D of Saccharomyces cere-visiae. Genetics 142: 749–759.
Fan HY, Merker RJ, Klein HL. 2001. High-copy-numberexpression of Sub2p, a member of the RNA helicasesuperfamily, suppresses hpr1-mediated genomic insta-bility. Mol Cell Biol 21: 5459–5470.
Feuerhahn S, Iglesias N, Panza A, Porro A, Lingner J. 2010.TERRA biogenesis, turnover and implications for func-tion. FEBS Lett 584: 3812–3818.
French S. 1992. Consequences of replication fork movementthrough transcription units in vivo. Science 258: 1362–1365.
Gaillard H, Herrera-Moyano E, Aguilera A. 2013. Transcrip-tion-associated genome instability. Chem Rev 113: 8638–8661.
Gan W, Guan Z, Liu J, Gui T, Shen K, Manley JL, Li X. 2011.R-loop-mediated genomic instability is caused by im-pairment of replication fork progression. Genes Dev 25:2041–2056.
Ganley AR, Hayashi K, Horiuchi T, Kobayashi T. 2005. Iden-tifying gene-independent noncoding functional ele-ments in the yeast ribosomal DNA by phylogenetic foot-printing. Proc Natl Acad Sci 102: 11787–11792.
Garcia-Rubio ML, Aguilera A. 2012. Topological constraintsimpair RNA polymerase II transcription and causes in-stability of plasmid-borne convergent genes. Nucleic Ac-ids Res 40: 1050–1064.
Garcia-Rubio M, Huertas P, Gonzalez-Barrera S, Aguilera A.2003. Recombinogenic effects of DNA-damaging agentsare synergistically increased by transcription in Saccha-romyces cerevisiae. New insights into transcription-asso-ciated recombination. Genetics 165: 457–466.
Gavalda S, Gallardo M, Luna R, Aguilera A. 2013. R-loopmediated transcription-associated recombination intrf4D mutants reveals new links between RNA surveil-lance and genome integrity. PLoS One 8: e65541.
Ginno PA, Lott PL, Christensen HC, Korf I, Chedin F. 2012.R-loop formation is a distinctive characteristic of un-methylated human CpG island promoters. Mol Cell 45:814–825.
Ginno PA, Lim YW, Lott PL, Korf I, Chedin F. 2013. GC skewat the 50 and 30 ends of human genes links R-loop forma-tion to epigenetic regulation and transcription termina-tion. Genome Res 23: 1590–1600.
Gomez-Gonzalez B, Aguilera A. 2007. Activation-inducedcytidine deaminase action is strongly stimulated by mu-tations of the THO complex. Proc Natl Acad Sci 104:8409–8414.
Gomez-Gonzalez B, Felipe-Abrio I, Aguilera A. 2009. TheS-phase checkpoint is required to respond to R-loopsaccumulated in THO mutants. Mol Cell Biol 29: 5203–5213.
Gonzalez-Aguilera C, Tous C, Gomez-Gonzalez B, HuertasP, Luna R, Aguilera A. 2008. The THP1-SAC3-SUS1-CDC31 complex works in transcription elongation-mRNA export preventing RNA-mediated genome in-stability. Mol Biol Cell 19: 4310–4318.
Gonzalez-Barrera S, Garcia-Rubio M, Aguilera A. 2002.Transcription and double-strand breaks induce similarmitotic recombination events in Saccharomyces cerevi-siae. Genetics 162: 603–614.
Gottipati P, Cassel TN, Savolainen L, Helleday T. 2008. Tran-scription-associated recombination is dependent on rep-lication in mammalian cells. Mol Cell Biol 28: 154–164.
Gowrishankar J, Harinarayanan R. 2004. Why is transcrip-tion coupled to translation in bacteria? Mol Microbiol 54:598–603.
Grabczyk E, Mancuso M, Sammarco MC. 2007. A persistentRNA.DNA hybrid formed by transcription of the Frie-dreich ataxia triplet repeat in live bacteria, and by T7RNAP in vitro. Nucleic Acids Res 35: 5351–5359.
Grimm C, Schaer P, Munz P, Kohli J. 1991. The strong ADH1promoter stimulates mitotic and meiotic recombinationat the ADE6 gene of Schizosaccharomyces pombe. Mol CellBiol 11: 289–298.
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 13
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Grolimund L, Aeby E, Hamelin R, Armand F, Chiappe D,Moniatte M, Lingner J. 2013. A quantitative telomericchromatin isolation protocol identifies different telo-meric states. Nat Commun 4: 2848.
Helmrich A, Ballarino M, Tora L. 2011. Collisions betweenreplication and transcription complexes cause commonfragile site instability at the longest human genes. Mol Cell44: 966–977.
Heyer WD, Ehmsen KT, Liu J. 2010. Regulation of homol-ogous recombination in eukaryotes. Annu Rev Genet 44:113–139.
Hill TM, Marians KJ. 1990. Escherichia coli Tus protein actsto arrest the progression of DNA replication forks invitro. Proc Natl Acad Sci 87: 2481–2485.
Horiuchi T, Fujimura Y. 1995. Recombinational rescue of thestalled DNA replication fork: A model based on analysisof an Escherichia coli strain with a chromosome regiondifficult to replicate. J Bacteriol 177: 783–791.
Hu Y, Lu X, Barnes E, Yan M, Lou H, Luo G. 2005. Recql5and Blm RecQ DNA helicases have nonredundant roles insuppressing crossovers. Mol Cell Biol 25: 3431–3442.
Hu Y, Raynard S, Sehorn MG, Lu X, Bussen W, Zheng L,Stark JM, Barnes EL, Chi P, Janscak P, et al. 2007.RECQL5/Recql5 helicase regulates homologous recom-bination and suppresses tumor formation via disruptionof Rad51 presynaptic filaments. Genes Dev 21: 3073–3084.
Huertas P. 2010. DNA resection in eukaryotes: Decidinghow to fix the break. Nat Struct Mol Biol 17: 11–16.
Huertas P, Aguilera A. 2003. Cotranscriptionally formedDNA:RNA hybrids mediate transcription elongation im-pairment and transcription-associated recombination.Mol Cell 12: 711–721.
Huertas P, Garcia-Rubio ML, Wellinger RE, Luna R, Agui-lera A. 2006. An hpr1 point mutation that impairs tran-scription and mRNP biogenesis without increasing re-combination. Mol Cell Biol 26: 7451–7465.
Ide S, Miyazaki T, Maki H, Kobayashi T. 2010. Abundanceof ribosomal RNA gene copies maintains genome integ-rity. Science 327: 693–696.
Ikeda H, Kobayashi I. 1977. Involvement of DNA-depen-dent RNA polymerase in a recA-independent pathwayof genetic recombination in Escherichia coli. Proc NatlAcad Sci 74: 3932–3936.
Ikeda H, Matsumoto T. 1979. Transcription promotes recA-independent recombination mediated by DNA-depen-dent RNA polymerase in Escherichia coli. Proc NatlAcad Sci 76: 4571–4575.
Islam MN, Fox D III, Guo R, Enomoto T, Wang W. 2010.RecQL5 promotes genome stabilization through two par-allel mechanisms—Interacting with RNA polymerase IIand acting as a helicase. Mol Cell Biol 30: 2460–2472.
Jacome A, Fernandez-Capetillo O. 2011. Lac operator re-peats generate a traceable fragile site in mammalian cells.EMBO Rep 12: 1032–1038.
Jimeno S, Rondon AG, Luna R, Aguilera A. 2002. The yeastTHO complex and mRNA export factors link RNA me-tabolism with transcription and genome instability.EMBO J 21: 3526–3535.
Kassube SA, Jinek M, Fang J, Tsutakawa S, Nogales E. 2013.Structural mimicry in transcription regulation of human
RNA polymerase II by the DNA helicase RECQL5. NatStruct Mol Biol 20: 892–899.
Keil RL, Roeder GS. 1984. Cis-acting, recombination-stim-ulating activity in a fragment of the ribosomal DNA ofS. cerevisiae. Cell 39: 377–386.
Khatri GS, MacAllister T, Sista PR, Bastia D. 1989. The rep-lication terminator protein of E. coli is a DNA sequence-specific contra-helicase. Cell 59: 667–674.
Kim N, Jinks-Robertson S. 2012. Transcription as a source ofgenome instability. Nat Rev Genet 13: 204–214.
Kim RA, Wang JC. 1989. A subthreshold level of DNA topo-isomerases leads to the excision of yeast rDNA as extra-chromosomal rings. Cell 57: 975–985.
Kim HD, Choe J, Seo YS. 1999. The sen1þ gene of Schizo-saccharomyces pombe, a homologue of budding yeastSEN1, encodes an RNA and DNA helicase. Biochemistry38: 14697–14710.
Kim N, Huang SN, Williams JS, Li YC, Clark AB, Cho JE,Kunkel TA, Pommier Y, Jinks-Robertson S. 2011. Muta-genic processing of ribonucleotides in DNA by yeasttopoisomerase I. Science 332: 1561–1564.
Klein IA, Resch W, Jankovic M, Oliveira T, Yamane A, Na-kahashi H, Di Virgilio M, Bothmer A, Nussenzweig A,Robbiani DF, et al. 2011. Translocation-capture sequenc-ing reveals the extent and nature of chromosomal rear-rangements in B lymphocytes. Cell 147: 95–106.
Kobayashi T, Ganley AR. 2005. Recombination regulation bytranscription-induced cohesin dissociation in rDNA re-peats. Science 309: 1581–1584.
Kobayashi T, Horiuchi T, Tongaonkar P, Vu L, Nomura M.2004. SIR2 regulates recombination between differentrDNA repeats, but not recombination within individualrRNA genes in yeast. Cell 117: 441–453.
Kogoma T. 1997. Stable DNA replication: Interplay betweenDNA replication, homologous recombination, and tran-scription. Microbiol Mol Biol Rev 61: 212–238.
Krasilnikova MM, Samadashwily GM, Krasilnikov AS, Mir-kin SM. 1998. Transcription through a simple DNA re-peat blocks replication elongation. EMBO J 17: 5095–5102.
Lambert S, Watson A, Sheedy DM, Martin B, Carr AM.2005. Gross chromosomal rearrangements and elevatedrecombination at an inducible site-specific replicationfork barrier. Cell 121: 689–702.
Lengronne A, Katou Y, Mori S, Yokobayashi S, Kelly GP, ItohT, Watanabe Y, Shirahige K, Uhlmann F. 2004. Cohesinrelocation from sites of chromosomal loading to places ofconvergent transcription. Nature 430: 573–578.
Le Tallec B, Dutrillaux B, Lachages AM, Millot GA, BrisonO, Debatisse M. 2011. Molecular profiling of commonfragile sites in human fibroblasts. Nat Struct Mol Biol 18:1421–1423.
Letessier A, Millot GA, Koundrioukoff S, Lachages AM,Vogt N, Hansen RS, Malfoy B, Brison O, Debatisse M.2011. Cell-type-specific replication initiation programsset fragility of the FRA3B fragile site. Nature 470: 120–123.
Li X, Manley JL. 2005. Inactivation of the SR protein splicingfactor ASF/SF2 results in genomic instability. Cell 122:365–378.
A. Aguilera and H. Gaillard
14 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Li M, Xu X, Liu Y. 2011. The SET2-RPB1 interaction domainof human RECQ5 is important for transcription-associ-ated genome stability. Mol Cell Biol 31: 2090–2099.
Lin Y, Dent SY, Wilson JH, Wells RD, Napierala M. 2010. Rloops stimulate genetic instability of CTG.CAG repeats.Proc Natl Acad Sci 107: 692–697.
Lippert MJ, Kim N, Cho JE, Larson RP, Schoenly NE, O’SheaSH, Jinks-Robertson S. 2011. Role for topoisomerase 1 intranscription-associated mutagenesis in yeast. Proc NatlAcad Sci 108: 698–703.
Liu B, Alberts BM. 1995. Head-on collision between a DNAreplication apparatus and RNA polymerase transcriptioncomplex. Science 267: 1131–1137.
Liu LF, Wang JC. 1987. Supercoiling of the DNA templateduring transcription. Proc Natl Acad Sci 84: 7024–7027.
Liu B, Wong ML, Tinker RL, Geiduschek EP, Alberts BM.1993. The DNA replication fork can pass RNA polymer-ase without displacing the nascent transcript. Nature 366:33–39.
Lu W, Zhang Y, Liu D, Songyang Z, Wan M. 2013. Telomeres-structure, function, and regulation. Exp Cell Res 319:133–141.
Luke B, Panza A, Redon S, Iglesias N, Li Z, Lingner J. 2008.The Rat1p 50 to 30 exonuclease degrades telomeric repeat-containing RNA and promotes telomere elongation inSaccharomyces cerevisiae. Mol Cell 32: 465–477.
Luna R, Jimeno S, Marin M, Huertas P, Garcia-Rubio M,Aguilera A. 2005. Interdependence between transcriptionand mRNP processing and export, and its impact ongenetic stability. Mol Cell 18: 711–722.
Marinello J, Chillemi G, Bueno S, Manzo SG, Capranico G.2013. Antisense transcripts enhanced by camptothecin atdivergent CpG-island promoters associated with burstsof topoisomerase I-DNA cleavage complex and R-loopformation. Nucleic Acids Res 41: 10110–10123.
Mischo HE, Gomez-Gonzalez B, Grzechnik P, Rondon AG,Wei W, Steinmetz L, Aguilera A, Proudfoot NJ. 2011.Yeast Sen1 helicase protects the genome from transcrip-tion-associated instability. Mol Cell 41: 21–32.
Nakama M, Kawakami K, Kajitani T, Urano T, Murakami Y.2012. DNA–RNA hybrid formation mediates RNAi-di-rected heterochromatin formation. Genes Cells 17: 218–233.
Nasmyth K. 2011. Cohesin: A catenase with separate entryand exit gates? Nat Cell Biol 13: 1170–1177.
Nickoloff JA, Reynolds RJ. 1990. Transcription stimulateshomologous recombination in mammalian cells. MolCell Biol 10: 4837–4845.
Olavarrieta L, Hernandez P, Krimer DB, Schvartzman JB.2002. DNA knotting caused by head-on collision of tran-scription and replication. J Mol Biol 322: 1–6.
O’Sullivan RJ, Karlseder J. 2010. Telomeres: Protecting chro-mosomes against genome instability. Nat Rev Mol CellBiol 11: 171–181.
Paul S, Million-Weaver S, Chattopadhyay S, Sokurenko E,Merrikh H. 2013. Accelerated gene evolution throughreplication-transcription conflicts. Nature 495: 512–515.
Paulsen RD, Soni DV, Wollman R, Hahn AT, Yee MC, GuanA, Hesley JA, Miller SC, Cromwell EF, Solow-CorderoDE, et al. 2009. A genome-wide siRNA screen reveals
diverse cellular processes and pathways that mediate ge-nome stability. Mol Cell 35: 228–239.
Pfeiffer V, Crittin J, Grolimund L, Lingner J. 2013. The THOcomplex component Thp2 counteracts telomeric R-loopsand telomere shortening. EMBO J 32: 2861–2871.
Phan AT. 2010. Human telomeric G-quadruplex: Structuresof DNA and RNA sequences. FEBS J 277: 1107–1117.
Piruat JI, Aguilera A. 1996. Mutations in the yeast SRB2general transcription factor suppress hpr1-induced re-combination and show defects in DNA repair. Genetics143: 1533–1542.
Piruat JI, Aguilera A. 1998. A novel yeast gene, THO2, isinvolved in RNA pol II transcription and provides newevidence for transcriptional elongation-associated re-combination. EMBO J 17: 4859–4872.
Pomerantz RT, O’Donnell M. 2008. The replisome usesmRNA as a primer after colliding with RNA polymerase.Nature 456: 762–766.
Pomerantz RT, O’Donnell M. 2010. Direct restart of a rep-lication fork stalled by a head-on RNA polymerase. Sci-ence 327: 590–592.
Postow L, Ullsperger C, Keller RW, Bustamante C, Vologod-skii AV, Cozzarelli NR. 2001. Positive torsional straincauses the formation of a four-way junction at replicationforks. J Biol Chem 276: 2790–2796.
Powell WT, Coulson RL, Gonzales ML, Crary FK, Wong SS,Adams S, Ach RA, Tsang P, Yamada NA, Yasui DH, et al.2013. R-loop formation at Snord116 mediates topotecaninhibition of Ube3a-antisense and allele-specific chro-matin decondensation. Proc Natl Acad Sci 110: 13938–13943.
Prado F, Aguilera A. 2005. Impairment of replication forkprogression mediates RNA polII transcription-associatedrecombination. EMBO J 24: 1267–1276.
Ramiro AR, Jankovic M, Eisenreich T, Difilippantonio S,Chen-Kiang S, Muramatsu M, Honjo T, NussenzweigA, Nussenzweig MC. 2004. AID is required for c-myc/IgH chromosome translocations in vivo. Cell 118: 431–438.
Reaban ME, Griffin JA. 1990. Induction of RNA-stabilizedDNA conformers by transcription of an immunoglobu-lin switch region. Nature 348: 342–344.
Reaban ME, Lebowitz J, Griffin JA. 1994. Transcription in-duces the formation of a stable RNA.DNA hybrid in theimmunoglobulin a switch region. J Biol Chem 269:21850–21857.
Robbiani DF, Bothmer A, Callen E, Reina-San-Martin B,Dorsett Y, Difilippantonio S, Bolland DJ, Chen HT, Cor-coran AE, Nussenzweig A, et al. 2008. AID is required forthe chromosomal breaks in c-myc that lead to c-myc/IgHtranslocations. Cell 135: 1028–1038.
Roberts SA, Sterling J, Thompson C, Harris S, Mav D, ShahR, Klimczak LJ, Kryukov GV, Malc E, Mieczkowski PA, etal. 2012. Clustered mutations in yeast and in human can-cers can arise from damaged long single-strand DNAregions. Mol Cell 46: 424–435.
Roberts SA, Lawrence MS, Klimczak LJ, Grimm SA, FargoD, Stojanov P, Kiezun A, Kryukov GV, Carter SL, SaksenaG, et al. 2013. An APOBEC cytidine deaminase mutagen-esis pattern is widespread in human cancers. Nat Genet45: 970–976.
Transcription-Associated Recombination
Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543 15
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Ruiz JF, Gomez-Gonzalez B, Aguilera A. 2011. AID inducesdouble-strand breaks at immunoglobulin switch regionsand c-MYC causing chromosomal translocations in yeastTHO mutants. PLoS Genet 7: e1002009.
Santangelo GM, Tornow J, McLaughlin CS, Moldave K.1988. Properties of promoters cloned randomly fromthe Saccharomyces cerevisiae genome. Mol Cell Biol 8:4217–4224.
Santos-Pereira JM, Herrero AB, Garcia-Rubio ML, Marin A,Moreno S, Aguilera A. 2013. The Npl3 hnRNP preventsR-loop-mediated transcription-replication conflicts andgenome instability. Genes Dev 27: 2445–2458.
Santos-Rosa H, Aguilera A. 1995. Isolation and genetic anal-ysis of extragenic suppressors of the hyper-deletion phe-notype of the Saccharomyces cerevisiae hpr1 d mutation.Genetics 139: 57–66.
Savolainen L, Helleday T. 2009. Transcription-associated re-combination is independent of XRCC2 and mechanisti-cally separate from homology-directed DNA double-strand break repair. Nucleic Acids Res 37: 405–412.
Savolainen L, Cassel T, Helleday T. 2010. The XPD subunit ofTFIIH is required for transcription-associated but notDNA double-strand break-induced recombination inmammalian cells. Mutagenesis 25: 623–629.
Saxe D, Datta A, Jinks-Robertson S. 2000. Stimulation ofmitotic recombination events by high levels of RNApolymerase II transcription in yeast. Mol Cell Biol 20:5404–5414.
Schoeftner S, Blasco MA. 2008. Developmentally regulatedtranscription of mammalian telomeres by DNA-depen-dent RNA polymerase II. Nat Cell Biol 10: 228–236.
Sfeir A, Kosiyatrakul ST, Hockemeyer D, MacRae SL, Karl-seder J, Schildkraut CL, de Lange T. 2009. Mammaliantelomeres resemble fragile sites and require TRF1 for ef-ficient replication. Cell 138: 90–103.
Skourti-Stathaki K, Proudfoot NJ, Gromak N. 2011. Humansenataxin resolves RNA/DNA hybrids formed at tran-scriptional pause sites to promote Xrn2-dependent ter-mination. Mol Cell 42: 794–805.
Stewart SE, Roeder GS. 1989. Transcription by RNA poly-merase I stimulates mitotic recombination in Saccharo-myces cerevisiae. Mol Cell Biol 9: 3464–3472.
Stirling PC, Chan YA, Minaker SW, Aristizabal MJ, Barrett I,Sipahimalani P, Kobor MS, Hieter P. 2012. R-loop-medi-ated genome instability in mRNA cleavage and poly-adenylation mutants. Genes Dev 26: 163–175.
Sun Q, Csorba T, Skourti-Stathaki K, Proudfoot NJ, Dean C.2013. R-loop stabilization represses antisense transcrip-tion at the Arabidopsis FLC locus. Science 340: 619–621.
Szilard RK, Jacques PE, Laramee L, Cheng B, Galicia S,Bataille AR, Yeung M, Mendez M, Bergeron M, RobertF, et al. 2010. Systematic identification of fragile sites viagenome-wide location analysis of g-H2AX. Nat StructMol Biol 17: 299–305.
Taghian DG, Nickoloff JA. 1997. Chromosomal double-strand breaks induce gene conversion at high frequencyin mammalian cells. Mol Cell Biol 17: 6386–6393.
Takahashi T, Burguiere-Slezak G, Van der Kemp PA, BoiteuxS. 2011. Topoisomerase 1 provokes the formation of shortdeletions in repeated sequences upon high transcription
in Saccharomyces cerevisiae. Proc Natl Acad Sci 108: 692–697.
Teves SS, Henikoff S. 2014. Transcription-generated torsion-al stress destabilizes nucleosomes. Nat Struct Mol Biol 21:88–94.
Thomas BJ, Rothstein R. 1989. Elevated recombination ratesin transcriptionally active DNA. Cell 56: 619–630.
Tsang E, Carr AM. 2008. Replication fork arrest, recombi-nation and the maintenance of ribosomal DNA stability.DNA Repair 7: 1613–1623.
Tuduri S, Crabbe L, Conti C, Tourriere H, Holtgreve-Grez H,Jauch A, Pantesco V, De Vos J, Thomas A, Theillet C, et al.2009. Topoisomerase I suppresses genomic instability bypreventing interference between replication and tran-scription. Nat Cell Biol 11: 1315–1324.
Vilette D, Uzest M, Ehrlich SD, Michel B. 1992. DNA tran-scription and repressor binding affect deletion formationin Escherichia coli plasmids. EMBO J 11: 3629–3634.
Voelkel-Meiman K, Keil RL, Roeder GS. 1987. Recombina-tion-stimulating sequences in yeast ribosomal DNA cor-respond to sequences regulating transcription by RNApolymerase I. Cell 48: 1071–1079.
Wahba L, Amon JD, Koshland D, Vuica-Ross M. 2011. RN-ase H and multiple RNA biogenesis factors cooperate toprevent RNA:DNA hybrids from generating genome in-stability. Mol Cell 44: 978–988.
Wahba L, Gore SK, Koshland D. 2013. The homologousrecombination machinery modulates the formation ofRNA-DNA hybrids and associated chromosome instabil-ity. eLife 2: e00505.
Wei W, Ba Z, Gao M, Wu Y, Ma Y, Amiard S, White CI,Rendtlew Danielsen JM, Yang YG, Qi Y. 2012. A role forsmall RNAs in DNA double-strand break repair. Cell 149:101–112.
Wellinger RE, Prado F, Aguilera A. 2006. Replication forkprogression is impaired by transcription in hyper-recombinant yeast cells lacking a functional THO com-plex. Mol Cell Biol 26: 3327–3334.
Weng YS, Xing D, Clikeman JA, Nickoloff JA. 2000. Tran-scriptional effects on double-strand break-induced geneconversion tracts. Mutat Res 461: 119–132.
Xu Y, Suzuki Y, Ito K, Komiyama M. 2010. Telomeric repeat-containing RNA structure in living cells. Proc Natl AcadSci 107: 14579–14584.
Xu Y, Ishizuka T, Yang J, Ito K, Katada H, Komiyama M,Hayashi T. 2012a. Oligonucleotide models of telomericDNA and RNA form a hybrid G-quadruplex structure asa potential component of telomeres. J Biol Chem 287:41787–41796.
Xu Z, Zan H, Pone EJ, Mai T, Casali P. 2012b. Immunoglob-ulin class-switch DNA recombination: Induction, target-ing and beyond. Nat Rev Immunol 12: 517–531.
Yu K, Chedin F, Hsieh CL, Wilson TE, Lieber MR. 2003. R-loops at immunoglobulin class switch regions in thechromosomes of stimulated B cells. Nat Immunol 4:442–451.
Zaratiegui M, Vaughn MW, Irvine DV, Goto D, Watt S,Bahler J, Arcangioli B, Martienssen RA. 2011. CENP-Bpreserves genome integrity at replication forks paused byretrotransposon LTR. Nature 469: 112–115.
A. Aguilera and H. Gaillard
16 Cite this article as Cold Spring Harb Perspect Biol 2014;6:a016543
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

2014; doi: 10.1101/cshperspect.a016543Cold Spring Harb Perspect Biol Andrés Aguilera and Hélène Gaillard Transcription and Recombination: When RNA Meets DNA
Subject Collection DNA Recombination
Meiotic Recombination: The Essence of HeredityNeil Hunter Recombinational DNA Repair
An Overview of the Molecular Mechanisms of
Stephen C. Kowalczykowski
MaintenanceRegulation of Recombination and Genomic
Wolf-Dietrich HeyerHomologs during MeiosisRecombination, Pairing, and Synapsis of
Denise Zickler and Nancy Kleckner
Chromatin RemodelingFlexibility, Impact of Histone Modifications, and Initiation of Meiotic Homologous Recombination:
Lóránt Székvölgyi, Kunihiro Ohta and Alain Nicolas
MeiosisDNA Strand Exchange and RecA Homologs in
M. Scott Brown and Douglas K. Bishop
Recombination InitiationMechanism and Regulation of Meiotic
Isabel Lam and Scott KeeneyAneuploid Oocytes and Trisomy BirthsMeiosis and Maternal Aging: Insights from
al.Mary Herbert, Dimitrios Kalleas, Daniel Cooney, et
ProteinsThe Roles of BRCA1, BRCA2, and Associated Homologous Recombination and Human Health:
Rohit Prakash, Yu Zhang, Weiran Feng, et al.
Homeologous RecombinationMismatch Repair during Homologous and
Maria Spies and Richard Fishel
Cell Biology of Mitotic RecombinationMichael Lisby and Rodney Rothstein Amplification
Mechanisms of Gene Duplication and
Andrew B. Reams and John R. Roth
Homology-Directed RepairHomologous Recombination and DNA-Pairing and Annealing Processes in
Scott W. Morrical
at Functional and Dysfunctional TelomeresThe Role of Double-Strand Break Repair Pathways
Ylli Doksani and Titia de Lange
Mediators of Homologous DNA PairingAlex Zelensky, Roland Kanaar and Claire Wyman Recombination
Regulation of DNA Pairing in Homologous
Kwon, et al.James M. Daley, William A. Gaines, YoungHo
http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2014 Cold Spring Harbor Laboratory Press; all rights reserved
on February 13, 2020 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from



![Saizen [somatropin (rDNA origin) for injection] … · Saizen® [somatropin (rDNA origin) for injection] cool.click](https://img.pdfslide.net/doc/110x75/5b8977fc7f8b9abe1e8db089/saizen-somatropin-rdna-origin-for-injection-saizen-somatropin-rdna-origin.jpg)

























