Embed Size (px)
Citation preview

J. Plant Res. 108 : 235-240, 1995 Journal of Plant Research ~) by The Botanical Society of Japan 1995
Transient Expression of #-Glucuronidase in Plastids of Various Plant Cells and Tissues Delivered by a Pneumatic Particle Gun
Motoaki Seki'*, Naoki Shigemoto', Mamoru Sugita 2, Masahiro Sugiura ~, Hans-Ulrich Koop 3, Kohei Irifune' and Hiromichi Morikawa'
i Graduate Department of Gene Science, Faculty of Science, Hiroshima University, Higashi-Hiroshima, 724 Japan 2 Center for Gene Research, Nagoya University, Nagoya, 464 Japan 3 Laboratory for Cell Biology and Cell Culture, University of Munich, Institute of Botany, Menzinger Str.67, W-8000 Munich 19,
Federal Republic of Germany
Chloroplast expression plasmids pTRBCL-GUS (tobacco rbcL promoter-gusA-tobacco rbcL terminator) and pHHU3004 (spinach 'x gene' promoter-gusA-spin- ach rbcL terminator) and a control nuclear expression plasmid pBI221 (CaMV 35S promoter-gusA-NOS ter- minator) were introduced separately into cultured cells and tissues of tobacco and Arabidopsis thaliana, as well as into cultured cells of the lower land plants liverwort and hornwort by a pneumatic particle gun. The pTRBCL-GUS and pHHU3004 plasmids produced many blue spots in the BY-2 cells and the roots of Arabidopsis thaliana, but not in any of the green cells or tissues. The results suggest that the pTRBCL-GUS and pHHU3004 plasmids are expressed more in proplasUds and amylo- plasts than in chloroplasts. GUS activities of the BY-2 cells bombarded with pTRBCL-GUS and pHHU3004 were insensitive to o-amanitin treatment (10 and 50 /Lg/ ml), while that of the cells with pBI221 greatly decreased by the same treatment. Hence, it is likely that the pTRBCL-GUS and pHHU3004 plasmids were substantial- ly expressed in the proplasUds.
Key words : a-amaniUn m #-glucuronidase-- Parti- cle gun m Plastid transformation ~ Transient expression
The particle gun method is convenient for transforming plastids (Svab et al. 1990). Only a few reports of plastid
Abbreviations: CaMV, cauliflower mosaic virus; CIM, callus- inducing medium; CRM, callus regeneration medium; 2, 4-D, 2, 4-dichlorophenoxyacetic acid ; GUS, ,8-glucuronidase ; LS, Linsmaier and Skoog; MS, Murashige and Skoog; MU, 4- methylumbelliferone ; MUG, 4-methyl umbellifery-p-D-glucur- onide ; NAA, 1-naphthaleneacetic acid ; NOS, nopaline synthase ; SIM, shoot-inducing medium ; X-Gluc, 5-bromo-4- chloro-3-indolyl-.8-D-glucuronide. * Present address: Laboratory of Plant Molecular Biology,
The Institute of Physical and Chemical Research (RIKEN), Tsukuba Life Science Center, 3-1-1 Koyadai, Tsukuba, Ibara- ki 305, Japan.
transformation by this method in higher plants, however, have been published (Daniell et al. 1990, Ye et al. 1990, Daniell et al. 1991, Seki et al. 1992, Staub and Maliga 1993).
Elsewhere (Seki et al. 1992), we reported preliminary results of the transient expression of the GUS (Jefferson 1987) gene in plastids of cultured tobacco (Nicotiana tabacum L. cv. Bright Yellow-2) cells on bombardment of chloroplast expression plasmids in which the GUS gene is driven by the promoter of the tobacco rbcL and spinach 'x gene' (Westhoff 1985). We here report the transient plastid expression of the GUS gene by bombardment of expression plasmids to cultured cells and tissues of tobacco and Arabidopsis thaliana, as well as to cultured cells of the lower land plants, liverwort and hornwort. We also discuss the effects of (~-amanitin, an inhibitor of RNA polymerase II (Jendrisak 1980) on GUS expression in bombarded BY-2 cells.
Materials and Methods
Plasmid DNA Chloroplast expression plasmid pTRBCL-GUS (Seki et
al. 1992 ; Fig. 1) was constructed. The plasmid pTRBCL- GUS has the GUS gene under the control of the promoter region (55,922 to 57,431) and the 3'-untranslated region (59,029 to 59,289) of the tobacco rbcL gene (Shinozaki and Sugiura 1982). Numbering conforms to the sequence reported by Shinozaki et al. (1986). The chloroplast expression plasmid pHHU3004 (SpOrlein et al. 1991, Fig. 1) has the GUS gene under the control of the spinach 'x gene' (Westhoff 1985) promoter and the 3"-untranslated region of the spinach rbcL gene. The nuclear expression plasmid pBI221 (Clontech, Palo Alto, CA, Fig. 1) has the GUS gene under the control of the CaMV 35S promoter and the NOS polyadenylation signal.
Plant materials Suspension-cultured white cells of tobacco (N. tabacum

236 M. Seki et aL
L. cv. Bright Yellow-2) were subcultured weekly as report- ed elsewhere (lida et al. 1990). A suspension culture of cells (ca. 100 mg fresh weight), made 4days after sub- culture, was spread on a filter paper (ADVANTEC TOYO No. 2, 5.5 cm diameter) then bombarded as reported else- where (lida et al. 1990).
Suspension-cultured green cells of tobacco (N. tabacum cv. Samsun NN) were subcultured weekly, as reported previously (Takeda et al. 1989), on modified LS (Linsmaier and Skoog 1965) basal medium containing 10 /zM NAA, 1/zM kinetin, and 3% sucrose. A suspension culture of cells (ca. 250 mg fresh weight), made 6 days after subculture, was spread on the filter paper then bombarded.
Suspension-cultured green cells of liverwort (Mar- chantia paleacea var. diptera) were subcultured on NA-MS medium at two-week intervals as reported elsewhere (Takio et al. 1988). A suspension culture of cells (ca. 90 mg fresh weight), made 4days after subculture, was spread on the filter paper then bombarded.
Suspension-cultured green cells of hornwort (Anthoce- ros punctatus) were subcultured essentially as reported previously (Ono et al. 1988) at 3-week intervals on the medium consisting of MS (Murashige and Skoog 1962) inorganic salts, MS vitamins, 1/zM 2, 4-D, 4% glucose, and 0.1% calcium carbonate. A suspension culture of cells (ca. 125 mg fresh weight), made 10 days after sub- culture, was spread on the filter paper then bombarded.
Leaf sections (1 cm X 0.5 cm) of tobacco (N. tabacum cv. Petit Havana line SR1) were spread in a plastic Petri dish (5.2 cm internal diameter) containing RMOP medium (Svab et al. 1990). After being cultured for 3 days, the sections (ca. 350 mg fresh weight) were bombarded.
Leaf sections (1 cm• of Arabidopsis thaliana, collection number C24, were spread in the Petri dish containing CRM (Seki et al. 1991a). After being cultured for 3 days, the sections (ca. 360 mg fresh weight) were bombarded.
Root sections (0.5-1.0 cm long) of Arabidopsis thaliana, collection number C24, were spread on the filter paper on CIM that consisted of B5 (Gamborg et al. 1968) inorganic salts, B5 vitamins, 3% sucrose, 0.5 mg/I 2, 4-D, and 0.05 mg/I kinetin. After being cultured for 3 days (Seki et al. 1991b), the sections (ca. 150 mg fresh weight) were bom- barded.
Green calli from Arabidopsis thaliana, collection number C24, were induced by culturing root sections in CIM for 6 days then in SIM (Seki et al. 1991b) that consisted of B5 inorganic salts, B5 vitamins, 3% sucrose, and 5 mg/I N 8 (z/2-isopentenyl)adenine for 14 days. The calli (ca. 650 mg fresh weight) were spread in the Petri dish containing SIM then bombarded.
Gene delivery to the cells The pneumatic particle acceleration device and the
method of gene delivery to the cells and tissues were essentially as reported previously (lida et al. 1990, Seki et al. 1991a), except that the gold particles (1.1/zm in diame-
ter, Tokuriki Honten Co., Tokyo, Japan) were used and the accelerating pressure was 115 kg/cm 2 for BY-2 cells and leaves of tobacco and Arabidopsis thaliana, 150 kg/cm 2 for suspension-cultured cells of liverwort and hornwort, and 200 kg/cm 2 for the other samples. All the samples were spread in a circle (35 mm diameter) on the filter paper or the Petri ,dish and subjected to particle bombard- ment.
Histochemical assay of GUS expression After being bombarded, the samples were assayed
histochemically for GUS expression according to the procedures of lida et al. (1990) and Seki et al. (1991a) as modified by Kosugi et al. (1990). Samples were cultured on 0.8% agar medium for 3 days at 26C under a 16 hr light/8 hr dark cycle then in X-Gluc solution for 5 days at 37C, after which the number of blue spots of the GUS- expressing cells were counted under a microscope (x40, Nikon, SMZ-10).
Investigation of the intracellular location of the blue stain (indigotin)
After bombardment, BY-2 cells were cultured at 26C for 1 day under a 16 hr light/8 hr dark cycle then incubated with X-gluc solution for 2 hr. After their collection by centrifugation, the cells were fixed for 24 hr below 3C, with ethanol : acetic acid (3 : 1, v/v), after which they were stained with aceto-orcein (1% [w/v] orcein in 45% Iv/v] acetic acid) for 5 min then observed under a light micro- scope.
Fluorometric GUS assay Bombarded BY-2 cells were suspended in 4 ml of
culture medium and incubated for 24 hr at 26C in the dark with shaking at 130 rpm. The cells then were assayed fluorometrically for GUS activity, essentially as reported elsewhere (Jefferson 1987), with MUG as the substrate.
Examination of the effects of inhibitor of RNA polymerase II on GUS activity
After bombardment, BY-2 cells were suspended in 4 ml of culture medium with or without various concentrations of oL-amanitin. ~-amanitin was added at a final concen- tration of 10, 50, or 100/zg/ml. These cells were cultured for 24 hr in the dark with shaking at 130 rpm then assayed fluorometrically for GUS activity as described previously.
Results and Discussion
Table 1 gives the number of blue spots of GUS-express- ing cells per circle (35 mm diameter) for various plant cells and tissues bombarded with chloroplast (pTRBCL-GUS and pHHU3004) and nuclear (pBI221) expression plasmids (Fig. l). Plasmids pTRB.CL-GUS and pBI221 were expressed in all the cells and tissues, except the suspension-cultured cells of hornwort. Plasmid pHHU3004 was expressed only in cultured white (BY-2) cells of tobacco, and the roots and green calli of

Plastid Transformation by Particle Bombardment 237
S pro /
- - tobacco rbeL ter NOS t~r
spinach tobacco psbB pro spinach 'x gene' pro
NPTII 4 ~ rbcL pro ~ \ \ N ~ ' ~
pet[} ter~ ~ \ spinach
6.3 Kb 10 Kb
GUS barley p replieon
( S p 6 r l e i n ct a/.,1991)
Fig. 1. Structure of the chloroplast expression plasmids pTRBCL-GUS and pHHU3004, and the nuclear expression plasmid pBI221.
GUS
spinach i r-bcL ter
Table1. Number of GUS-expressing cells counted in various cells and tissues bombarded with chloroplast and nuclear expression plasmids
Plasmids
Arabidopsis thaliana Cultured white Cultured green Cultured green Cultured green Tobacco cells of tobacco cells of tobacco cells of cells of (N. tabacum
(Nicotiana (N. tabacum Marchant ia Anthoceros cv. SR1) tabacum L. Green cv. BY-2) cv. Samsun NN) pa leacea punctatus Leaves Leaves Roots calli
pTRBCL-GUS 15t3_+1077 ~) 28+14 24• 0 18_+6 15_+10 190• 43• pHHU3004 938• 1 _+1 0 0 0 0 13_+10 3_+0 pBI221 3892+328 427_+1 4878_+73 0 150• 694+_10 1128• 147•
1) Average of two experiments_deviation.
Arabidopsis thaliana. No blue spots were detected in cells bombarded with noncoated gold particles (data not shown). In all the bombarded cells, the pTRBCL-GUS plasmid gave more blue spots than pHHU3004, except for the case of suspension-cultured cells of hornwort (Table 1).
Interestingly, when the chloroplast expression plasmids pTRBCL-GUS and pHHU3004 were introduced, many blue spots showed up in the BY-2 cells and roots of Arabidopsis thaliana, but not in the leaves of tobacco and Arabidopsis thaliana or in cultured green cells of tobacco and liverwort (Table 1). These results suggest that the pTRBCL-GUS and pHHU3004 plasmids are expressed more in proplastids and amyloplasts than in chloroplasts.
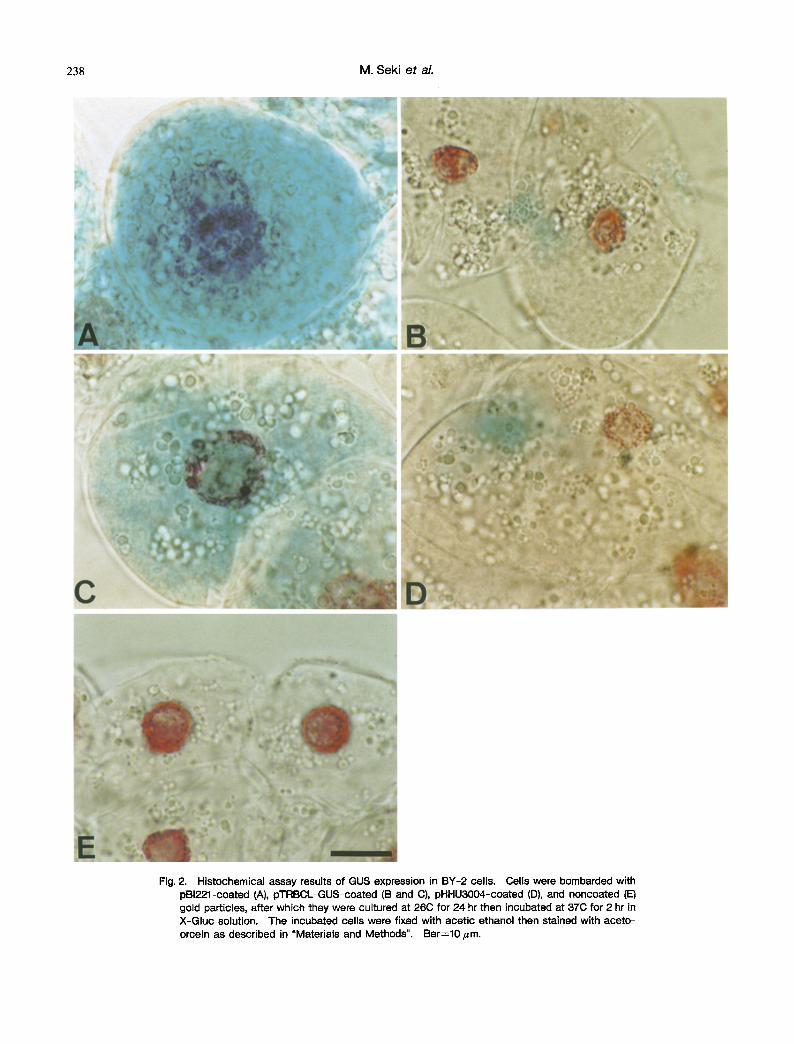
As the tobacco psbA promoter has been shown to function in the nuclear compartment (Cornelissen and Vandewiele, 1989), we examined whether the introduced GUS gene was expressed in the plastids or not. To determine the intracellular location of the blue stain (in- digotin) in the BY-2 cells, we made a histochemical assay of GUS expression in the bombarded cells then stained them with aceto-orcein. Cells bombarded with pBI221
showed an intense blue color that was distributed evenly throughout the cytosol (Fig. 2A). In cells bombarded with pHHU3004, the color was less intense and was localized in the proplastids (Fig. 2D). In some cells bombarded with pTRBCL-GUS, the blue color was not intense and was localized in the proplastids (Fig. 2B), whereas in others it was distributed throughout the cytosol (Fig. 2C). Cells bombarded with noncoated gold particles showed no blue color (Fig. 2E). We concluded that pTRBCL-GUS and pHHU3004 were expressed mostly in the plastids, as also has been reported for tobacco (Ye et al. 1990) and wheat (Dan• et al. 1991). Whether the plasmid DNA can be delivered into proplastids with or without delivery of gold particles into proplastids must await further analysis. It is noteworthy that BY-2 cells bombarded with the chloroplast expression plasmids pTRBCL-GUS and pHHU3004 showed blue spots as early as 2 hr after staining. This differs from the result reported by Ye et al. (1990) that cultured tobacco NT1 cells bombarded with chloroplast expression plasmids showed no blue spots until 4 to 5 days after staining.
Bombarded BY-2 cells were also subjected to a fluor-
238 M. Seki et aL
Fig. 2. Histochemical assay results of GUS expression in BY-2 cells. Cells were bombarded with pBI221-coated (A), pTRBCL-GUS-coated (B and C), pHHU3OO4-coated (D), and noncoated (E) gold particles, after which they were cultured at 26C for 24 hr then incubated at 37(3 for 2 hr in X-Gluc solution. The incubated cells were fixed with acetic ethanol then stained with aceto- orcein as described in "Materials and Methods". Bar=lO/zm.
Plastid Transformation by Particle Bombardment 239
i t -
[]11
"•1.5
i l . 0 -
0 . 5 -
~ 0 . C 0 10 50 100
Concentration of a-amanit in {tLtg/ml}
Q
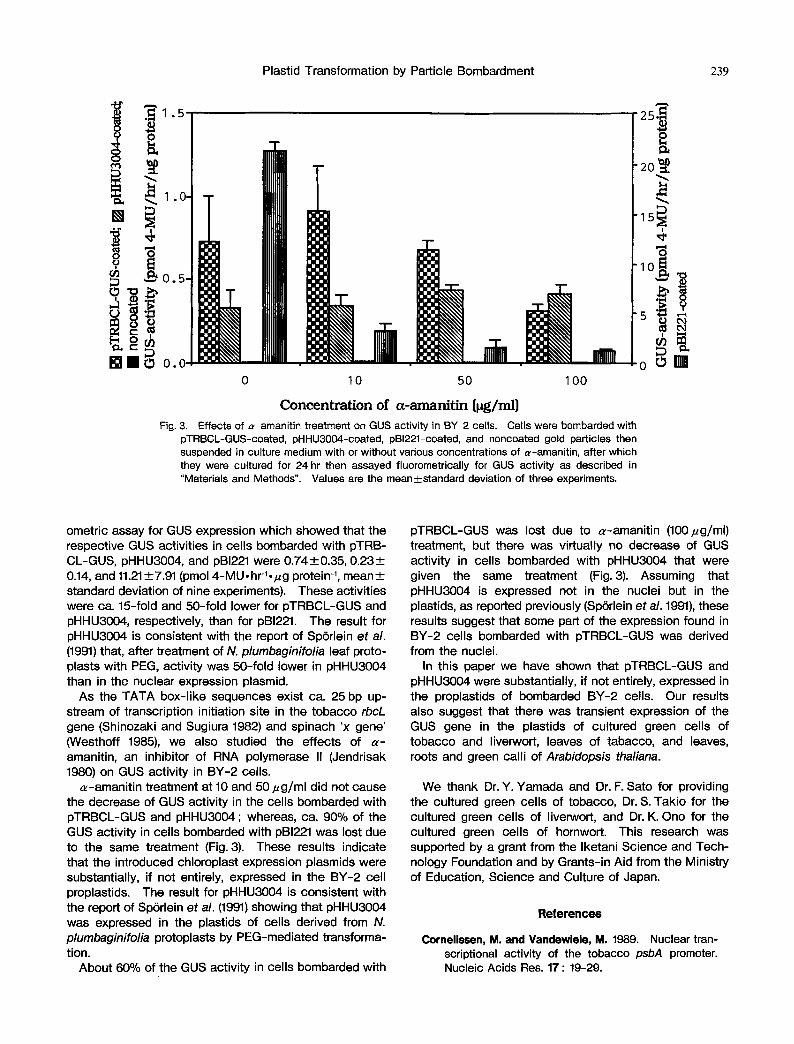
Fig. 3. Effects of ~z-amanitin treatment on GUS activity in BY 2 cells. Cells were bombarded with pTRBCL-GUS-coated, pHHU3004-coated, pBI221-coated, and noncoated gold particles then suspended in culture medium with or without various concentrations of ~-amanit in, after which they were cultured for 24 hr then assayed fluorometrically for GUS activity as described in "Materials and Methods". Values are the mean+standard deviation of three experiments.
25.
" 2 0 ~
4
,_~-o
0g,
ometric assay for GUS expression which showed that the respective GUS activities in cells bombarded with pTRB- CL-GUS, pHHU3004, and pBI221 were 0.74+__0.35, 0.23___ 0.14, and 11.21 _+7.91 (pmol 4-MU.hr-1-/zg protein -1, mean+_ standard deviation of nine experiments). These activities were ca. 15-fold and 50-fold lower for pTRBCL-GUS and pHHU3004, respectively, than for pBI221. The result for pHHU3004 is consistent with the report of Sp6rlein et al. (1991) that, after treatment of N. plumbaginifolia leaf proto- plasts with PEG, activity was 50-fold lower in pHHU3004 than in the nuclear expression plasmid.
As the TATA box-like sequences exist ca. 25 bp up- stream of transcription initiation site in the tobacco rbcL gene (Shinozaki and Sugiura 1982) and spinach 'x gene' (Westhoff 1985), we also studied the effects of ~- amanitin, an inhibitor of RNA polymerase II (Jendrisak 1980) on GUS activity in BY-2 cells.
o~-amanitin treatment at 10 and 50/zg/ml did not cause the decrease of GUS activity in the cells bombarded with pTRBCL-GUS and pHHU3004 ; whereas, ca. 90% of the GUS activity in cells bombarded with pBI221 was lost due to the same treatment (Fig. 3). These results indicate that the introduced chloroplast expression plasmids were substantially, if not entirely, expressed in the BY-2 cell proplastids. The result for pHHU3004 is consistent with the report of Sp6rlein et al. (1991) showing that pHHU3004 was expressed in the plastids of cells derived from N. plumbaginifolia protoplasts by PEG-mediated transforma- tion.
About 60% of the GUS activity in cells bombarded with
pTRBCL-GUS was lost due to ~-amanitin (100~g/ml) treatment, but there was virtually no decrease of GUS activity in cells bombarded with pHHU3004 that were given the same treatment (Fig. 3). Assuming that pHHU3004 is expressed not in the nuclei but in the plastids, as reported previously (Sp6rlein et al. 1991), these results suggest that some part of the expression found in BY-2 cells bombarded with pTRBCL-GUS was derived from the nuclei.
In this paper we have shown that pTRBCL-GUS and pHHUS004 were substantially, if not entirely, expressed in the proplastids of bombarded BY-2 cells. Our results also suggest that there was transient expression of the GUS gene in the plastids of cultured green cells of tobacco and liverwort, leaves of tabacco, and leaves, roots and green calli of Arabidopsis thaliana.
We thank Dr. Y. Yamada and Dr. F. Sato for providing the cultured green cells of tobacco, Dr. S. Takio for the cultured green cells of liverwort, and Dr. K. Ono for the cultured green cells of hornwort. This research was supported by a grant from the Iketani Science and Tech- nology Foundation and by Grants-in Aid from the Ministry of Education, Science and Culture of Japan.
References
Cornelissen, M. and Vandewiele, M. 1989. Nuclear tran- scriptional activity of the tobacco psbA promoter, Nucleic Acids Res, 17 : 19-29,

240 M. Seki et aL
Daniell, H., Krishnan, M. and McFadden, B.F. 1991. Tran- sient expression of ~'-glucuronidase in different cellu- lar compartments following biolistic delivery of foreign DNA into wheat leaves and calli. Plant Cell Rep. 9 : 615-619.
Daniell, H., Vivekananda, J., Nielsen, B.L., Ye, G.N., Tewari, K.K. and Sanford, J.C. 1990. Transient for- eign gene expression in chloroplasts of cultured tobacco cells after biolistic delivery of chloroplast vectors. Proc. Natl. Acad. Sci. USA 87 : 88-92.
Gamborg, O.L, Miller, R.A. and Ojima, K. 1968. Nutrient requirements of suspension cultures of soybean root cells�9 Exp. Cell Res. 50: 151-158.
lida, A., Seki, M., Kamada, M., Yamada, Y. and Morikawa, H. 1990. Gene delivery into cultured plant cells by DNA-coated gold particles accelerated by a pneu- matic particle gun. Theor. Appl. Genet. 80: 813- 816.
Jefferson, R.A. 1987. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol. Biol. Rep. 5: 387-405.
Jendrisak, J. 1980. The use of ~z-amanitin to inhibit in vivo RNA synthesis and germination in wheat em- bryos. J. Biol. Chem. 255: 8529-8533.
Kosugi, S., Ohashi, Y., Nakajima, K. and Arai, Y. 1990. An improved assay for #-glucuronidase in transfor- med cells : methanol almost completely suppresses a putative endogenous ,6-glucuronidase activity. Plant Sci. 70 : 133-140.
Linsmaier, E.M. and Skoog, F. 1965. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant. 18 : 100-127.
Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15 : 473-497.
Ono, K., Murasaki, Y. and Takamiya, M. 1988. Induction and morphogenesis of cultured cells of bryophytes. J. Hattori Bot. Lab. 65: 391-401.
Sek|, M., Komeda, Y., lida, A., Yamada, Y. and Morikawa, H. 1991a. Transient expression of /~-glucuronidase in Arabidopsis thaliana leaves and roots, and Brassica napus stems using a pneumatic particle gun. Plant Mol. Biol. 17 : 259-263.
Seki, M., Shigemoto, N., Komeda, Y., Imamura, J., Yamada, Y. and Morikawa, H. 1991b. Transgenic Arabidopsis thaliana plants obtained by particle-bom- bardment-mediated transformation. Appl. Microbiol.
Biotechnol. 36 : 228-230. Seki, M., Shigemoto, N., Sugita, M., Sugiura, M., Keep,
H.U., Irifune, K. and Morikawa, H. 1992. Chloroplast transformation by particle bombardment. In N. Mu- rata, ed., Research in Photosynthesis, Vol. III, Kluwer Academic Publishers, Dordrecht. pp. 413-416.
Shinozaki, K., Ohme, M., Tanaka, M., Wakasugi, T., Haya- shida, N., Matsubayashi, T., Zaita, N., Chunwongse, J., Obokata, J., Yamaguchi-Shinozaki, K., Ohto, C., Tora- zawa, K., Meng, B. Y.,Sugita, M., Deno, H., Kamoya- shira, T., Yamada, K., Kusuda, J., Takaiwa, F., Kato, A., Tohdoh, N., Shimada, H. and Sugiura, M. 1986. The complete nucteotide sequence of the tobacco chloroplast genome : its gene organization and expression. EMBO J. 5: 2043-2049.
Shinozaki, K. and Sugiura, M. 1982. The nucleotide sequence of the tobacco chloroplast gene for the large subunit of ribulose-t, 5-bisphosphate carboxy- lase/oxygenase. Gene 20 : 91-102.
Sp6rlein, B., Streubel, M., Dahlfeld, G, Westhoff, P. and Koop, H.U. 1991. PEG-mediated plastid transforma- tion: a new system for transient gene expression assays in chloroplasts. Theor. Appl. Genet. 82: 717-722.
Staub, J.M. and Maliga, P. 1993. Accumulation of D1 polypeptide in tobacco plastids is regulated via the untranslated region of the psbA mRNA. EMBO J. 12 : 601-606.
Svab, D., Hajdukiewitz, P. and Maliga, P. 1990. Stable transformation of plastids in higher plants. Proc. Natl. Acad. Sci. USA 87 : 8526-8530.
Takeda, S., Sato, F. and Yamada, Y. 1989. Photosynth- etic characteristics of photoautotrophically cultured cells of tobacco. Plant Cell Physiol. 30 : 885-891.
Takio, S., Takami, S. and Hino, S. 1988. Photosynthetic ability of dark-grown Marchantia paleacea cells in suspension culture. J. Plant Physiol. 132 : 195-198.
Westhoff, P. 1985. Transcription of the gene encoding the 51 kd chlorophyll a-apoprotein of the photosystem II reaction center from spinach. Mol. Gen. Genet. 201 : 115-123.
Ye, G.N., Daniell, H. and Sanford, J.C. 1990. Optimization of delivery of foreign DNA into higher-plant chloro- plasts. Plant Mol. Biol. 15 : 809-819.
(Received January 7, 1995 : Accepted April 20, 1995)