Embed Size (px)
Citation preview

S.S. VirmaniZ.X. SunT.M. Mou
A. Jauhar AliC.X. Mao
Two-Line HybridRice Breeding
Manual

i
Two-Line HybridRice Breeding
Manual
S.S. VirmaniZ.X. SunT.M. Mou
A. Jauhar AliC.X. Mao
����������������������������� ��

ii
The International Rice Research Institute (IRRI) was established in 1960 bythe Ford and Rockefeller Foundations with the help and approval of the Gov-ernment of the Philippines. Today IRRI is one of 16 nonprofit internationalresearch centers supported by the Consultative Group on International Agri-cultural Research (CGIAR – www.cgiar.org).
IRRI receives support from several CGIAR members, including the WorldBank, European Union, Asian Development Bank, International Fund forAgricultural Development, Rockefeller Foundation, and agencies of thefollowing governments: Australia, Belgium, Canada, People’s Republic ofChina, Denmark, France, Germany, India, Islamic Republic of Iran, Japan,Republic of Korea, The Netherlands, Norway, Philippines, Portugal, Sweden,Switzerland, Thailand, United Kingdom, United States, and Vietnam.
The responsibility for this publication rests with the International RiceResearch Institute.
Copyright International Rice Research Institute 2003
Mailing address: DAPO Box 7777, Metro Manila, PhilippinesPhone: +63 (2) 580-5600, 845-0563, 844-3351 to 53Fax: +63 (2) 580-5699, 891-1292, 845-0606Email: [email protected] page: www.irri.orgRiceweb: www.riceweb.orgRiceworld: www.riceworld.orgCourier address: Suite 1009, Pacific Bank Building
6776 Ayala Avenue, Makati City, PhilippinesTel. (63-2) 891-1236, 891-1174, 891-1258, 891-1303
Suggested citation:Virmani SS, Sun ZX, Mou TM, Jauhar Ali A, Mao CX. 2003. Two-linehybrid rice breeding manual. Los Baños (Philippines): International RiceResearch Institute. 88 p.
Editing: Bill HardyCover design, print production coordinator, and page makeup and composition: George R. ReyesFigures and illustrations: Emmanuel Panisales and George R. ReyesCover photos: IRRI photos
ISBN 971-22-0185-6

iii
FOREWORD v
CHAPTER 1Hybrid rice and heterosis breeding 1Types of heterosis 1How is heterosis measured? 1Genetic basis of heterosis 2Molecular basis of heterosis 2Approaches for using heterosis 2Methods of using heterosis 3Hybrid rice 3
CHAPTER 2Male sterility systems in rice 5Cytoplasmic genetic male sterility 5Hybrid seed production using the 5
CMS systemEnvironment-sensitive genic 7
male sterilityChemically induced male sterility 12
CHAPTER 3Comparative organization of two- 15and three-line hybrid breedingprogramsSimilarities in three-line and two-line 15
breeding nurseriesDifferences in three-line and two-line 16
breeding nurseries
CHAPTER 4Inheritance of EGMS 19Procedure for carrying out inheritance 19
studies on the EGMS traitInheritance of TGMS 19Inheritance of PGMS and PTGMS 19
CHAPTER 5Breeding procedures for developing 23EGMS linesScreening of existing varieties for EGMS 23Induced mutagenesis 23Hybridization method 24
CHAPTER 6Characterizing EGMS lines under field 31and controlled conditionsCharacterization of EGMS lines under 31
field conditionsCharacterization of EGMS lines under 32
controlled conditionsEvaluation of EGMS lines 34
CHAPTER 7Developing pollen parents for two-line 37hybridsCharacteristic features of an elite pollen 37
parentBreeding methods for identifying pollen 37
parents for two-line hybridsNew strategies for developing pollen 38
parent lines
CHAPTER 8Combining ability nursery 41Definitions 41Type of lines to be evaluated 41Procedure using the line × tester design 41Composition of the combining 41
ability nurseryField layout 41Statistical analysis 41
Contents

iv
Analysis of variance 42Interpretation of results 44Using the results 44
CHAPTER 9Evaluating two-line hybrids 45Observation yield trial (OYT) 45Preliminary yield trials (PYT) 47Advanced yield trials (AYT) 50Multilocation yield trials (MLT) 50
CHAPTER 10Two-line hybrid rice seed production 53Multiplication of EGMS lines 53Differences between EGMS 54
and CMS line multiplicationSimilarity of CMS and EGMS 55
line multiplicationHigh-yielding techniques for PGMS line 55
multiplication (Chinese experience)High-yielding techniques for TGMS line 55
multiplication (Chinese experience)Three-line hybrid rice seed production 56Two-line hybrid rice seed 62
production on a large scale
CHAPTER 11Two-line rice hybrids: maintenance 65of genetic seed purity standards1. Using nucleus seeds of EGMS 65
for seed production2. Using anther culture for 66
purifying EGMS lines3. Transferring a recessive 66
marker gene into EGMS lines4. Insertion of a dominant marker 66
gene into the pollen parent
CHAPTER 12Future outlook for two-line 69rice hybrids
BIBLIOGRAPHY 71AUTHORS 76GLOSSARY 77APPENDIX I: IDENTIFYING MALE STERILITY 85APPENDIX II: PROTOCOL FOR ANTHER 87
CULTUREAPPENDIX III: DATA TO BE RECORDED 88
FOR HYBRID RICE EXPERIMENTS

v
Foreword
Hybrid rice technology has contributed signifi-cantly toward food security, environmental pro-tection, and employment opportunities in Chinafor the past 25 years. Since the mid-1990s, thistechnology has also been developed and intro-duced to farmers in India, Vietnam, the Philip-pines, Bangladesh, and the United States, eitherindependently or in close collaboration with IRRI.Several other countries, such as Egypt, Indonesia,Myanmar, Sri Lanka, Thailand, and the Republicof Korea, are now developing this technology incollaboration with IRRI.
The availability of adequately trained humanresources is an essential prerequisite for develop-ing and using hybrid rice technology. Hybrid ricebreeding uses several concepts, skills, and proce-dures that are strikingly different from those usedfor inbred rice breeding. Two male sterility sys-tems (the cytoplasmic genic male sterility and en-vironment-sensitive genic male sterility system)have been used extensively to develop commer-cial rice hybrids in China and elsewhere. Duringthe past 25 years, IRRI and China have offeredseveral short-term training courses jointly and in-dependently to develop the human resources incountries interested in developing this technol-ogy. In 1997, IRRI also published a “Hybrid RiceBreeding Manual” to serve the needs of thosetraining courses. This manual described conceptsand procedures to breed rice hybrids primarilyusing cytoplasmic genic male sterility and the fer-tility restoration system. Since then, considerableprogress has been made in breeding rice hybrids
using environment-sensitive genic male sterility.In August 2000, China and IRRI held a col-
laborative international training course on hybridrice breeding at the China National Rice ResearchInstitute in Hangzhou, which was funded by theIRRI-ADB project on “Development and Use ofHybrid Rice in Asia.” Several Chinese and IRRIscientists participated in this course as resourcepersons and provided the training materials fo-cusing on two-line hybrid breeding using the en-vironment-sensitive genic male sterility system.Based on these materials, and the experience ofhybrid rice scientists from China and IRRI, thistraining manual on the two-line hybrid breedingmethod has been prepared, which expands uponthe hybrid rice breeding manual published earlierby IRRI. The authors have described the conceptsand procedures stepwise and in a systematic man-ner so that trainees can learn them easily. Thisshould make an excellent manual for future hy-brid rice breeding training courses organized atIRRI, in China, and in other countries.
I compliment the authors for preparing thistraining manual and thank Bill Hardy for editingit. The assistance of the Asian Development Bank,which provided financial support under RETA6005 on Sustaining Food Security in Asia Throughthe Development of Hybrid Rice Technology, isgratefully acknowledged.
RONALD P. CANTRELL
Director General

1
CHAPTER 1
Hybrid rice and heterosisbreeding
Heterosis is a phenomenon in which F1 hybrids
derived from diverse parents show superiority overtheir parents in vigor, yield, panicle size, numberof spikelets per panicle, number of productivetillers, etc.
• Heterosis is expressed in the first genera-tion only.
• Heterosis varies according to the level ofparental diversity and or presence of het-erotic gene blocks in parental lines; indica× japonica crosses show maximum hetero-sis vis-à-vis any other combination betweenother subspecies. The crosses showing het-erosis in descending order are indica ×japonica > indica × javanica > japonica ×javanica > indica × indica > japonica ×japonica > javanica × javanica.
• Heterosis can be positive or negative. Bothpositive and negative heterosis can be use-ful depending on the trait, for example, posi-tive heterosis for yield and negative hetero-sis for growth duration.
• Farmers tend to use a lower seed rate forhybrids than for conventional varieties be-cause of their better seed quality and higherseed cost. However, it is necessary to pur-chase fresh seeds every season to raise a com-mercial crop.
Types of heterosisHeterosis is expressed in three ways, dependingon the reference used to compare the performanceof a hybrid:
• Mid-parent heterosis is the increase or de-crease in the performance of a hybrid in com-parison with the mid-parental value.
• Heterobeltiosis is the increase or decreasein the performance of a hybrid in compari-son with the better parent of the cross com-bination.
• Standard heterosis is the increase or decreasein the performance of a hybrid in compari-son with the standard check variety of theregion.
From the practical viewpoint, standard het-erosis is the most important because we aim todevelop hybrids that are better than the existinghigh-yielding varieties grown commercially byfarmers.
How is heterosis measured?Measurement of heterosis is quite simple. It is gen-erally expressed as the percent increase or decreasein the performance of a hybrid in comparison withthe reference variety or a parameter.
F1 – mid-parentMid-parent = × 100
heterosis (%) Mid-parent
F1 – better parent
Heterobeltiosis (%) = × 100 Better parent
F1 – check varietyStandard = × 100
heterosis (%) Check variety

2
Genetic basis of heterosis
Two major hypotheses have been proposed toexplain the genetic basis of heterosis: the domi-nance hypothesis (Davenport 1908) and over-dominance hypothesis (East 1908, 1936).
• Dominance hypothesis– This states that heterosis is due to the accu-
mulation of favorable dominant genes in ahybrid derived from the two parents (Fig.1).
This was demonstrated in a pea hybridwhose parents had different dominant genesfor node number and internodal length. Thehybrid was much taller than either parent.The increased height was due to the accu-mulation of four dominant genes in the hy-brid.
• Overdominance hypothesis– This states that the heterozygote (Aa) is more
vigorous and productive than either ho-mozygote (AA or aa). This has been provenin traits controlled by a single or a few genes.The heterozygote performs a given func-tion, over a range of environments, moreefficiently than either homozygote (East1936).
Studies on the genetic basis of heterosis forpolygenic traits in various crops have shown thatheterosis is the result of partial to complete domi-nance, overdominance, and epistasis, and it maybe a combination of all these (Comstock andRobinson 1952). Evidence of real overdominancefor quantitative traits is hard to find. However,apparent overdominance caused by nonallelicinteraction and linkage disequilibrium is a com-mon contributor to heterosis (Jinks 1983).
Heterosis may also be due to the specific posi-tive effects of the cytoplasm of the maternal par-ent on the nuclear component of the paternal par-ent. Differential heterosis observed between thesame pollen parent and cytoplasmic male sterile(CMS) lines from different cytosterility sources isan example of this kind of heterosis.
Molecular basis of heterosisSeveral molecular studies support the overdomi-nance hypothesis (Stuber et al 1992, Yu et al 1997,
Li et al 2000) except for a few that support thedominance hypothesis (Xiao et al 1995). Yu et al(1997) reported overdominance at several main-effect quantitative trait loci (QTLs) and a strongeradditive epistasis affecting grain yield and its com-ponents in F
3 progenies from the most widely grown
hybrid in China, Shan You 63. Zhang et al (2001)demonstrated the involvement of large numbersof two-loci interactions or epistasis as the geneticbasis of quantitative traits and heterosis. Further-more, Li et al (2000) concluded that most QTLsassociated with inbreeding depression and het-erosis in rice appeared to be involved in epistasis.And almost 90% of the QTLs contributing to het-erosis appeared to be overdominant. Zhang et al(2001) assessed the relationship between geneexpression and heterosis by assaying the patternsof differential gene expression in hybrids relativeto their parents in a diallel cross. The analysis re-vealed that differentially expressed fragments oc-curring in only one parent of the cross were posi-tively correlated with heterosis and fragments de-tected in F
1s but not in the respective parents were
negatively correlated with heterosis. A total of 384fragments recovered from gels were hybridizedwith mRNAs from seedling and flag-leaf tissuesand thereby Zhang et al (2000) detected an over-all elevated level of gene expression in the hybridcompared with the parents. Several fragmentsshowed a higher expression in the highly heterotichybrid than in the other hybrids.
Nonetheless, a lack of a clear understandingof the genetic or molecular basis of heterosis hasnot prevented plant breeders from exploiting thisphenomenon to raise crop yields.
Approaches for using heterosisCurrently, hybrid rice technology mainly usesintrasubspecific heterosis, that is, indica × indicaand japonica × japonica. The high-yielding
P1 P2AAbbCCdd × aaBBccDD
(2 dominant genes) (2 dominant genes)
F1AaBbCcDd
(4 dominant genes)
Fig. 1. Illustration of dominance hypothesis to explaingenetic basis for heterosis.

3
intrasubspecific hybrids yield nearly 15% to 20%more than the best inbred varieties grown undersimilar conditions. It has been quite difficult tocreate or widen the genetic difference among par-ents belonging to the intrasubspecific hybrids andthey have almost reached their yield ceiling.
Indica and japonica (both tropical and tem-perate) are the two main subspecies of Oryza sa-tiva in Asia. Among them, the indica and temper-ate japonica subspecies are the most apparentlydifferent in their morphological and agronomictraits apart from the genetic distance between them.Therefore, it is now well understood why the in-dica × japonica hybrids show maximum hetero-sis. But intersubspecific heterosis is limited be-cause of high spikelet sterility and long growthduration. With the discovery of wide compatibil-ity (WC) genes, it has been possible to exploitintersubspecific hybrids with normal seed settingand suitable growth duration. Efforts are underway in IRRI, China, and India to developintersubspecific hybrids. Recently, Chinese sci-entists have developed super high-yielding ricehybrids from crosses involving indica/indica-japonica derivative parents.
Most of the interspecific crosses in cultivatedspecies pertain to only O. sativa and O. glaberrima,which are heterotic but not so useful in terms ofyield and plant stature. Most interspecific hybrids,resulting from wide hybridization, result in ge-netic variability and bring in desirable genes forresistance to several biotic and abiotic stresses,for example, O. sativa × O. longistaminata, O. sa-tiva × O. rufipogon, and O. sativa × O. perennis.In rice, the interspecific F
1 hybrids cannot be used
commercially.
Methods of using heterosisThe three-line method is based on cytoplasmicgenic male sterility and the fertility restorationsystem and involves three lines—the CMS line(A), maintainer line (B), and restorer line (R)—forthe commercial production of rice hybrids. Theseed of the male sterile line is multiplied by cross-ing A and B lines in an isolation plot. Hybrid seedis produced by crossing the A line with an R linein isolation in another plot. Seed production tech-niques are now developed to produce up to 3 tha–1 (mean 1.2 t ha–1) of hybrid seed in the tropicsand up to 6 t ha–1 (mean 2.7 t ha–1) in subtropicaland temperate regions of China.
In the two-line method, the two lines are in-volved in a cross for hybrid rice seed production.One is a male sterile line in which male sterility isgenetically controlled by recessive genes, the ex-pression of which is influenced by environment(temperature, photoperiod, or both) and the otheris any inbred variety with a dominant gene forthat locus. The male sterile lines in which sterilityexpression is controlled by temperature are knownas thermosensitive genic male sterile (TGMS) linesand those in which expression is controlled bydaylength are called photoperiod-sensitive genicmale sterile (PGMS) lines.
Another two-line approach for hybrid riceseed production is by spraying chemical hybrid-izing agents (CHAs)—ethrel, ethyl 4′ fluorooxanilate, or sodium methyl arsenate—that selec-tively sterilize the male reproductive organs ofany one parent and planting the other line (notsprayed) close to the pollinator rows. China is theonly country that used CHAs such as sodium me-thyl arsenate and zinc methyl arsenate on a com-mercial scale. Because of the inefficient seed pro-duction related to nonsynchronous tillering andflowering as well as health hazards associated withthe use of arsenic compounds, CHA use in Chinawas discontinued.
To use three-line and two-line rice hybrids,farmers have to buy fresh seed every season. Thisseed is produced by a proficient seed productionagency in the public or private sector.
The one-line method involves the use of apo-mixis to develop F
1 hybrids. This represents true
breeding so that farmers can use the harvest fromthe hybrids as seed for the next crop as with anyinbred rice variety. Attempts to discover apomixishave not succeeded so far; however, research is stillunder way at IRRI, in China, and in some othercountries using genetic engineering techniques.
Hybrid riceWhat is hybrid rice?Hybrid rice is the commercial rice crop grown fromF
1 seeds of a cross between two genetically dis-
similar parents.• Good rice hybrids have the potential of
yielding 15–20% more than the best inbredvariety grown under similar conditions.
• To exploit the benefits of hybrid rice, farm-ers have to buy fresh seeds every croppingseason.

4
Why hybrid rice?The need for hybrid rice has been felt because
• Yield levels of semidwarf varieties of theGreen Revolution era have plateaued.
• The demand for rice is increasing rapidlywith the increase in population, especiallyin less developed countries.
• More and more rice has to be produced onless land and with less inputs.
• Hybrid rice varieties have already shown a15–20% higher yield potential than inbredrice varieties under farmers’ field conditionsin several countries.
• Hybrids have also shown an ability to per-form better under adverse conditions ofdrought and salinity.
How is hybrid rice developed?Hybrid rice is developed by exploiting the phe-nomenon of heterosis. Rice, being a strictly self-pollinated crop, requires the use of a male sterilitysystem to develop commercial rice hybrids. Malesterility (genetic or nongenetic) makes the pollenunviable so that rice spikelets are incapable ofsetting seeds through selfing. A male sterile lineis used as a female parent and grown side by sidewith a pollen parent in an isolated plot to producea bulk quantity of hybrid seed because of crosspollination with the adjoining fertile pollen par-ent. The seed set on male sterile plants is the hy-brid seed that is used to grow the commercial hy-brid crop.

5
CHAPTER 2
Male sterility systemsin rice
Male sterility can be defined as a condition inwhich the pollen grain is unviable or cannot ger-minate and fertilize normally to set seeds.
The following genetic and nongenetic malesterility systems are known for developing ricehybrids (Fig. 2):
• Cytoplasmic genetic male sterility• Environment-sensitive genic male sterility• Chemically induced male sterility
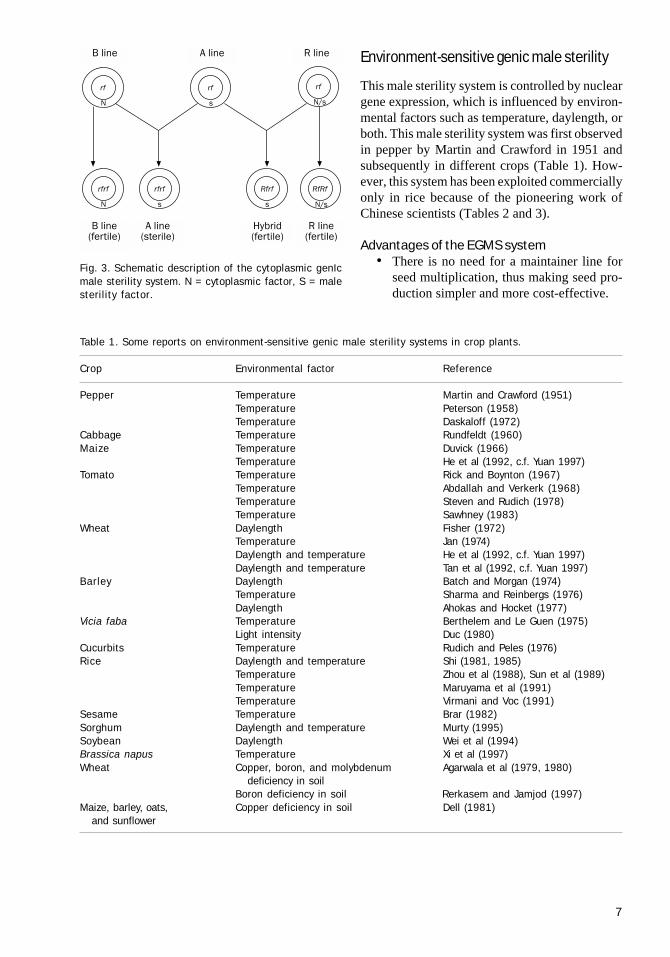
Cytoplasmic genetic male sterilityMale sterility is controlled by the interaction of agenetic factor S present in the cytoplasm andnuclear gene(s). It is now known that the malesterility factor S is located in the mitochondrialDNA. A line is male sterile when the male steril-ity-controlling factor S in the cytoplasm and re-cessive alleles (rf ) of fertility-restoring genes arepresent in the nucleus. The maintainer line (B line)is iso-cytoplasmic to the CMS line since it is simi-lar to it for nuclear genes but differs in cytoplas-mic factor (N), which makes it self-fertile, but ithas the capacity to maintain the sterility of the Aline when crossed with it. A restorer or R line pos-sesses dominant fertility-restoring genes (Rf) andit is dissimilar to or diverse from the A line. Cross-ing a restorer line as a pollen parent with a CMS(A) line as a female parent restores the fertility inthe derived F
1 hybrid.
• The restorer gene in the dominant homozy-gous (RfRf) or heterozygous (Rfrf) state canrestore the fertility in the F
1 hybrid despite
the presence of sterility factors in the cyto-plasm derived from the A line. The CMSsystem is illustrated in Figure 3.
Hybrid seed production usingthe CMS system
Hybrid seed production involves two steps: mul-tiplication of the CMS line and production ofhybrid seeds.
Multiplication of the CMS lineThis requires crossing of the CMS line with itsmaintainer line by outcrossing by hand (for a smallquantity of seed) or in the field under isolation byspace or time (to produce a bulk quantity of seed).For successful production of the CMS line, it isgrown in six or eight rows interspersed by tworows of a maintainer line in an alternating man-ner.
Because there is a small difference betweenthe growth duration of A and B lines, their sowingdates are adjusted to achieve good synchroniza-tion of their flowering. Several other techniques(such as flag-leaf clipping, GA
3 application, and
supplementary pollination by rope pulling or thebamboo pole method) are used to improve theoutcrossing rate and seed yield of the CMS line.
Production of hybrid seedsThis involves the use of CMS lines with a selectedrestorer line (R line) by growing them in a specificfemale:male ratio in the field under isolation byspace or time. The CMS line is usually grown ineight or ten rows interspersed with two rows ofrestorer lines in an alternating manner. The sow-ing dates of A and R lines are staggered to achievesynchronization of their flowering. To increasethe outcrossing rate and hybrid seed yield, thetechniques described in the previous section areused.
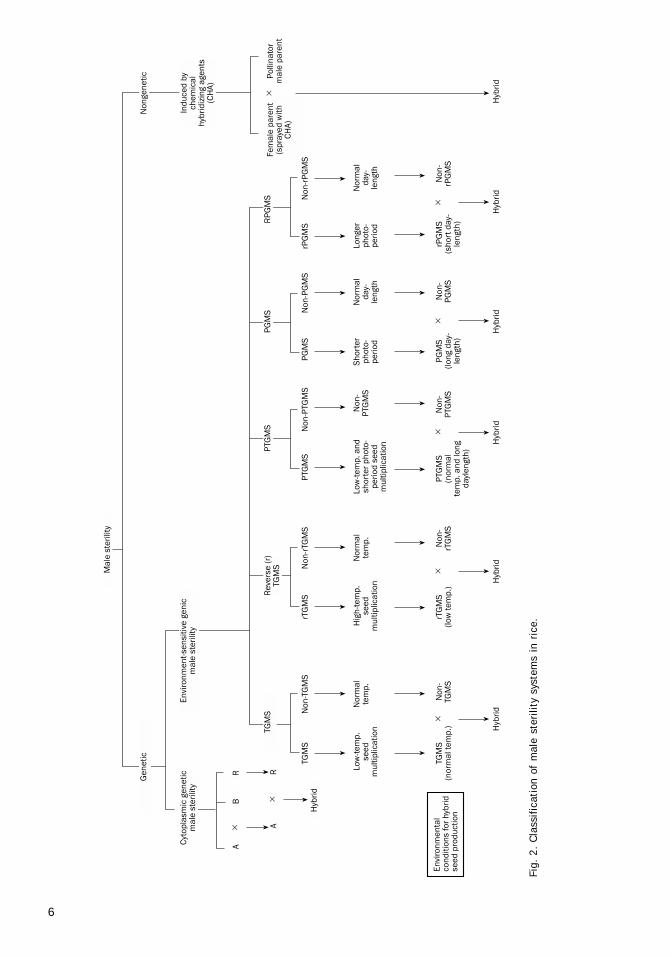
6
Fig.
2.
Clas
sific
atio
n of
mal
e st
erili
ty s
yste
ms
in r
ice.
���������
��
����� ��
����
����
� ����
� ����
� ����
� ����
� ����
����
��������
����
��
���
��
�����
������
����
��
������
�����
������
�������
���
�� ��
��
��
���
���
����
���
��� �
��
��
��
���
��
�����
����
���� �
��
��
��
������
!����
���
����
�"���
��#���
������
����
�� ������
����
�"���
��#���
������
����
!����
������
��������
����
�����
����
���"���
��#���
��
���
���
������
�����
�����
�
������
�� �
��
�
!�
����
���
�����
�
������
�� �
��
�
��
������
�����
��
�����
����
��
����
��
�����
��
������
�����
��
�����
����
����
$�%��������
����
�����
����
$��
��
&%����
���
#��
���
��'����
����
����
�����"
#��
��
($
�$
�
� ����
� �����
��#� �
��#
�����������
&%����
���
����%�� �
�#
�����������
��
��#
)�"
#����
#���
�#��
� �����*� ��
��
�����
������
��
�������
��
�
� ����
�����������
��
��
�
��
��
�#
7
Environment-sensitive genic male sterility
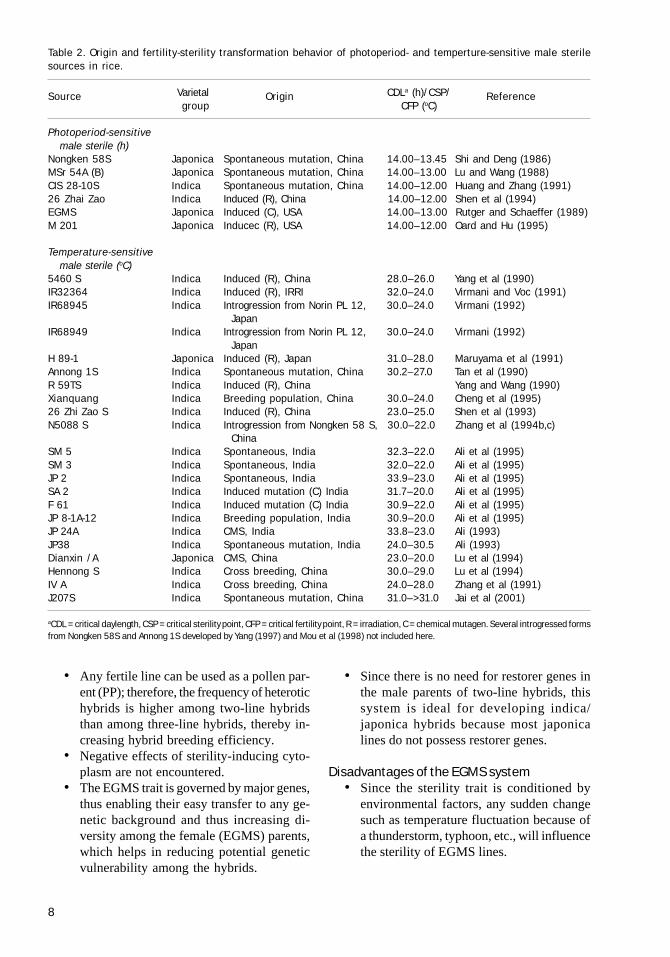
This male sterility system is controlled by nucleargene expression, which is influenced by environ-mental factors such as temperature, daylength, orboth. This male sterility system was first observedin pepper by Martin and Crawford in 1951 andsubsequently in different crops (Table 1). How-ever, this system has been exploited commerciallyonly in rice because of the pioneering work ofChinese scientists (Tables 2 and 3).
Advantages of the EGMS system• There is no need for a maintainer line for
seed multiplication, thus making seed pro-duction simpler and more cost-effective.
Fig. 3. Schematic description of the cytoplasmic genIcmale sterility system. N = cytoplasmic factor, S = malesterility factor.
(����
�
����� $����
����
�
����
�
����
�+�
��
�+�
��
�
��
����
�
(�����'������
�������������
$�����'������
� �����'������
Table 1. Some reports on environment-sensitive genic male sterility systems in crop plants.
Crop Environmental factor Reference
Pepper Temperature Martin and Crawford (1951)Temperature Peterson (1958)Temperature Daskaloff (1972)
Cabbage Temperature Rundfeldt (1960)Maize Temperature Duvick (1966)
Temperature He et al (1992, c.f. Yuan 1997)Tomato Temperature Rick and Boynton (1967)
Temperature Abdallah and Verkerk (1968)Temperature Steven and Rudich (1978)Temperature Sawhney (1983)
Wheat Daylength Fisher (1972)Temperature Jan (1974)Daylength and temperature He et al (1992, c.f. Yuan 1997)Daylength and temperature Tan et al (1992, c.f. Yuan 1997)
Barley Daylength Batch and Morgan (1974)Temperature Sharma and Reinbergs (1976)Daylength Ahokas and Hocket (1977)
Vicia faba Temperature Berthelem and Le Guen (1975)Light intensity Duc (1980)
Cucurbits Temperature Rudich and Peles (1976)Rice Daylength and temperature Shi (1981, 1985)
Temperature Zhou et al (1988), Sun et al (1989)Temperature Maruyama et al (1991)Temperature Virmani and Voc (1991)
Sesame Temperature Brar (1982)Sorghum Daylength and temperature Murty (1995)Soybean Daylength Wei et al (1994)Brassica napus Temperature Xi et al (1997)Wheat Copper, boron, and molybdenum Agarwala et al (1979, 1980)
deficiency in soilBoron deficiency in soil Rerkasem and Jamjod (1997)
Maize, barley, oats, Copper deficiency in soil Dell (1981) and sunflower
8
• Any fertile line can be used as a pollen par-ent (PP); therefore, the frequency of heterotichybrids is higher among two-line hybridsthan among three-line hybrids, thereby in-creasing hybrid breeding efficiency.
• Negative effects of sterility-inducing cyto-plasm are not encountered.
• The EGMS trait is governed by major genes,thus enabling their easy transfer to any ge-netic background and thus increasing di-versity among the female (EGMS) parents,which helps in reducing potential geneticvulnerability among the hybrids.
• Since there is no need for restorer genes inthe male parents of two-line hybrids, thissystem is ideal for developing indica/japonica hybrids because most japonicalines do not possess restorer genes.
Disadvantages of the EGMS system• Since the sterility trait is conditioned by
environmental factors, any sudden changesuch as temperature fluctuation because ofa thunderstorm, typhoon, etc., will influencethe sterility of EGMS lines.
Table 2. Origin and fertility-sterility transformation behavior of photoperiod- and temperture-sensitive male sterilesources in rice.
Source Varietal Origin CDLa (h)/CSP/ Referencegroup CFP (oC)
Photoperiod-sensitive male sterile (h)Nongken 58S Japonica Spontaneous mutation, China 14.00–13.45 Shi and Deng (1986)MSr 54A (B) Japonica Spontaneous mutation, China 14.00–13.00 Lu and Wang (1988)CIS 28-10S Indica Spontaneous mutation, China 14.00–12.00 Huang and Zhang (1991)26 Zhai Zao Indica Induced (R), China 14.00–12.00 Shen et al (1994)EGMS Japonica Induced (C), USA 14.00–13.00 Rutger and Schaeffer (1989)M 201 Japonica Inducec (R), USA 14.00–12.00 Oard and Hu (1995)
Temperature-sensitive male sterile (oC)5460 S Indica Induced (R), China 28.0–26.0 Yang et al (1990)IR32364 Indica Induced (R), IRRI 32.0–24.0 Virmani and Voc (1991)IR68945 Indica Introgression from Norin PL 12, 30.0–24.0 Virmani (1992)
JapanIR68949 Indica Introgression from Norin PL 12, 30.0–24.0 Virmani (1992)
JapanH 89-1 Japonica Induced (R), Japan 31.0–28.0 Maruyama et al (1991)Annong 1S Indica Spontaneous mutation, China 30.2–27.0 Tan et al (1990)R 59TS Indica Induced (R), China Yang and Wang (1990)Xianquang Indica Breeding population, China 30.0–24.0 Cheng et al (1995)26 Zhi Zao S Indica Induced (R), China 23.0–25.0 Shen et al (1993)N5088 S Indica Introgression from Nongken 58 S, 30.0–22.0 Zhang et al (1994b,c)
ChinaSM 5 Indica Spontaneous, India 32.3–22.0 Ali et al (1995)SM 3 Indica Spontaneous, India 32.0–22.0 Ali et al (1995)JP 2 Indica Spontaneous, India 33.9–23.0 Ali et al (1995)SA 2 Indica Induced mutation (C) India 31.7–20.0 Ali et al (1995)F 61 Indica Induced mutation (C) India 30.9–22.0 Ali et al (1995)JP 8-1A-12 Indica Breeding population, India 30.9–20.0 Ali et al (1995)JP 24A Indica CMS, India 33.8–23.0 Ali (1993)JP38 Indica Spontaneous mutation, India 24.0–30.5 Ali (1993)Dianxin /A Japonica CMS, China 23.0–20.0 Lu et al (1994)Hennong S Indica Cross breeding, China 30.0–29.0 Lu et al (1994)IV A Indica Cross breeding, China 24.0–28.0 Zhang et al (1991)J207S Indica Spontaneous mutation, China 31.0–>31.0 Jai et al (2001)
aCDL = critical daylength, CSP = critical sterility point, CFP = critical fertility point, R = irradiation, C = chemical mutagen. Several introgressed formsfrom Nongken 58S and Annong 1S developed by Yang (1997) and Mou et al (1998) not included here.

9
• The multiplication of EGMS lines and hy-brid seed production are restricted by spaceand season. This means that an EGMS lineis used in a given region and season.
Characteristic features of ideal EGMS lines• The proportion of male sterile plants in a
population of more than 1,000 plants dur-ing the critical sterility period should be100%.
• The pollen sterility of each male sterile plantshould be more than 99.5%.
• EGMS lines should have clearly defined ste-rility-fertility alteration regimes.
• The male sterile phase should last for morethan 4 consecutive weeks.
• Seed setting during the fertile phase shouldbe more than 30%.
• The critical temperature or photoperiod forinducing sterility should be as low as pos-sible for more stability of the EGMS lines,
for example, <27 ºC (maximum) or <13 hphotoperiod.
In addition, these lines should have a goodplant type possessing high yield, acceptable grainquality, pest and disease resistance, and adapt-ability to the target environment.
Classification of the EGMS systemDepending on the environmental factor(s) influ-encing expression of the sterility-inducing gene(s),EGMS is classified in the following categories(Figs. 4–7):
1. TGMS: temperature-sensitive genic malesterility
2. rTGMS: reverse temperature-sensitivegenic male sterility
3. PGMS: photoperiod-sensitive genic malesterility
4. rPGMS: reverse photoperiod-sensitivegenic male sterility
5. PTGMS: photothermosensitive genic malesterility
Table 3. Two-line rice hybrids released up to 2001 in China.
Hybrid Pedigree Type Year of release Cultivation region/crop
Ejingza No.1 N5088S/R187 Japonica 1995 Yangtze Valley/secondHuajingza No. 1 7001S/1514 Japonica 1995 Yangtze Valley/secondHuajingza No. 2 N5088S/65396 Japonica 2001 Yangtze Valley/second70 you 9 7001S/Wanhui 9 Japonica 1994 Yangtze Valley/second70 you 99 7001S/99 Japonica 1997 Yangtze Valley/second70 you 04 7001S/Xiushui 04 Japonica 1994 Yangtze Valley/secondPeiliangyou Teqing Pei-ai 64S/Teqing Indica 1994 Yangtze/single or secondPeiliangyou 288 Pei-ai 64S/R822 Indica 1996 Yangtze/single or secondPeiliangyou Yuhong Pei-ai 64S/Yuhong No. 1 Indica 1997 Yangtze/single or secondLiangyou Peiju Pei-ai 64S/9311 Indica 1999 Yangtze/single or secondLiangyou 923 W9593S/Shengyou No. 2 Indica 2001 Yangtze/single or secondLiangyou 681 Shuguang 612S/881 Indica 1999 Yangtze/single or secondXiangliangyou 68 Xiang 125S/D 68 Indica 1998 Yangtze/first8 Liangyou 100 Annong 810S/D100 Indica 1998 Yangtze/firstTainliangyou 402 TianfengS/R 402 Indica 1998 Yangtze/firstAn Liangyou 25 1356 S/Zao 25 Indica 1998 Yangtze/firstPeiza Shangqing Pei-ai 64S/Shanqing Indica 1997 South China/first and secondJinliangyou 36 HS-3/946 Indica 2000 South China/first and secondPeiza Shuangqi Pei-ai 64S/Shuangqizhan Indica 1998 South China/first and secondLiangyou 2163 SE21S/Minghui63 Indica 2000 South China/first and secondLiangyou 2186 SE21S/Minghui86 Indica 2000 South China/first and secondFu Liangyou 63 FJS-1/Minghui63 Indica 2000 South China/first and secondPei Liangyou 275 Pei-ai 64S/275 Indica 1999 South China/first and secondPei Liangyou 99 Pei-ai 64S/Gui99 Indica 1998 South China/first and secondPeiza Maosan Pei-ai 64S/Maosan Indica 2000 South China/first and secondPeiza Maoxuan Pei-ai 64S/Maoxuan Indica 2000 South China/first and secondSouth China/first and Pei-ai 64S/G67 Indica 2000 South China/first and second second Peiza 67Yunguang No. 8 N5088S/Yunhui 11 Japonica 2000 West China/single
10
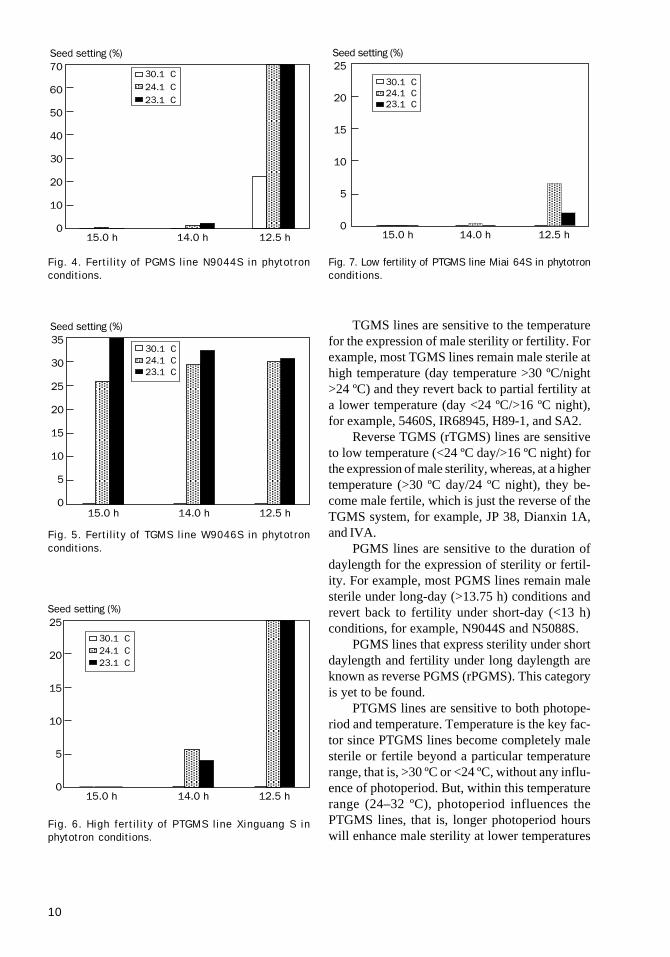
TGMS lines are sensitive to the temperaturefor the expression of male sterility or fertility. Forexample, most TGMS lines remain male sterile athigh temperature (day temperature >30 ºC/night>24 ºC) and they revert back to partial fertility ata lower temperature (day <24 ºC/>16 ºC night),for example, 5460S, IR68945, H89-1, and SA2.
Reverse TGMS (rTGMS) lines are sensitiveto low temperature (<24 ºC day/>16 ºC night) forthe expression of male sterility, whereas, at a highertemperature (>30 ºC day/24 ºC night), they be-come male fertile, which is just the reverse of theTGMS system, for example, JP 38, Dianxin 1A,and IVA.
PGMS lines are sensitive to the duration ofdaylength for the expression of sterility or fertil-ity. For example, most PGMS lines remain malesterile under long-day (>13.75 h) conditions andrevert back to fertility under short-day (<13 h)conditions, for example, N9044S and N5088S.
PGMS lines that express sterility under shortdaylength and fertility under long daylength areknown as reverse PGMS (rPGMS). This categoryis yet to be found.
PTGMS lines are sensitive to both photope-riod and temperature. Temperature is the key fac-tor since PTGMS lines become completely malesterile or fertile beyond a particular temperaturerange, that is, >30 ºC or <24 ºC, without any influ-ence of photoperiod. But, within this temperaturerange (24–32 ºC), photoperiod influences thePTGMS lines, that is, longer photoperiod hourswill enhance male sterility at lower temperatures
��
��
��
��
��
��
��
�
�� ���������
������ ������ ������
���������������������
Fig. 7. Low fertility of PTGMS line Miai 64S in phytotronconditions.
Fig. 4. Fertility of PGMS line N9044S in phytotronconditions.
Fig. 5. Fertility of TGMS line W9046S in phytotronconditions.
Fig. 6. High fertility of PTGMS line Xinguang S inphytotron conditions.
��
��
��
��
��
��
�
�
�� ���������
������ ������ ������
���������������������
��
��
��
��
�
�
�� ���������
������ ������ ������
���������������������
��
��
��
��
�
�
�� ���������
������ ������ ������
���������������������
11
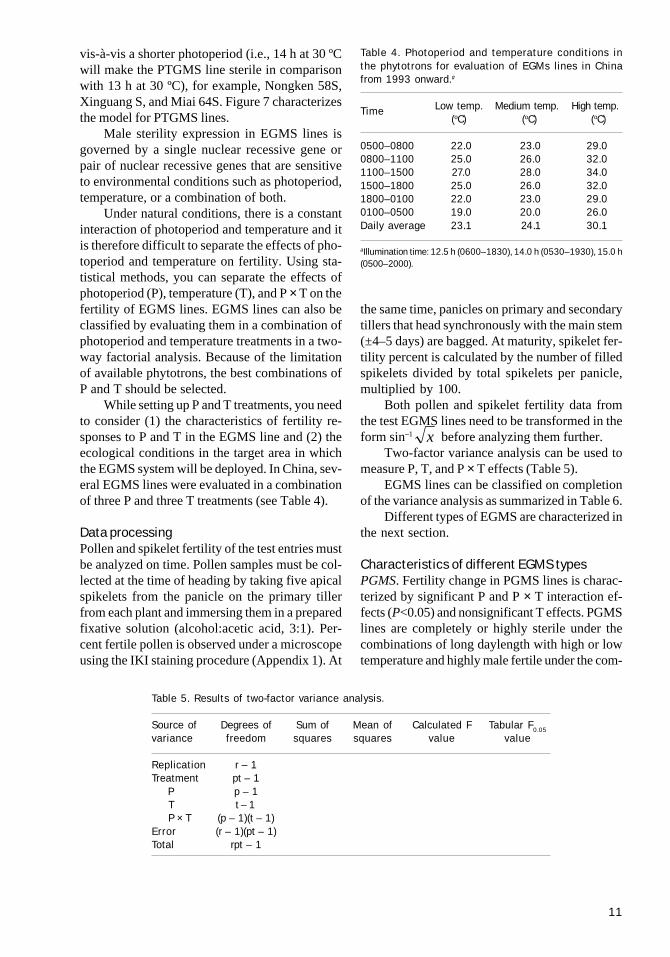
vis-à-vis a shorter photoperiod (i.e., 14 h at 30 ºCwill make the PTGMS line sterile in comparisonwith 13 h at 30 ºC), for example, Nongken 58S,Xinguang S, and Miai 64S. Figure 7 characterizesthe model for PTGMS lines.
Male sterility expression in EGMS lines isgoverned by a single nuclear recessive gene orpair of nuclear recessive genes that are sensitiveto environmental conditions such as photoperiod,temperature, or a combination of both.
Under natural conditions, there is a constantinteraction of photoperiod and temperature and itis therefore difficult to separate the effects of pho-toperiod and temperature on fertility. Using sta-tistical methods, you can separate the effects ofphotoperiod (P), temperature (T), and P × T on thefertility of EGMS lines. EGMS lines can also beclassified by evaluating them in a combination ofphotoperiod and temperature treatments in a two-way factorial analysis. Because of the limitationof available phytotrons, the best combinations ofP and T should be selected.
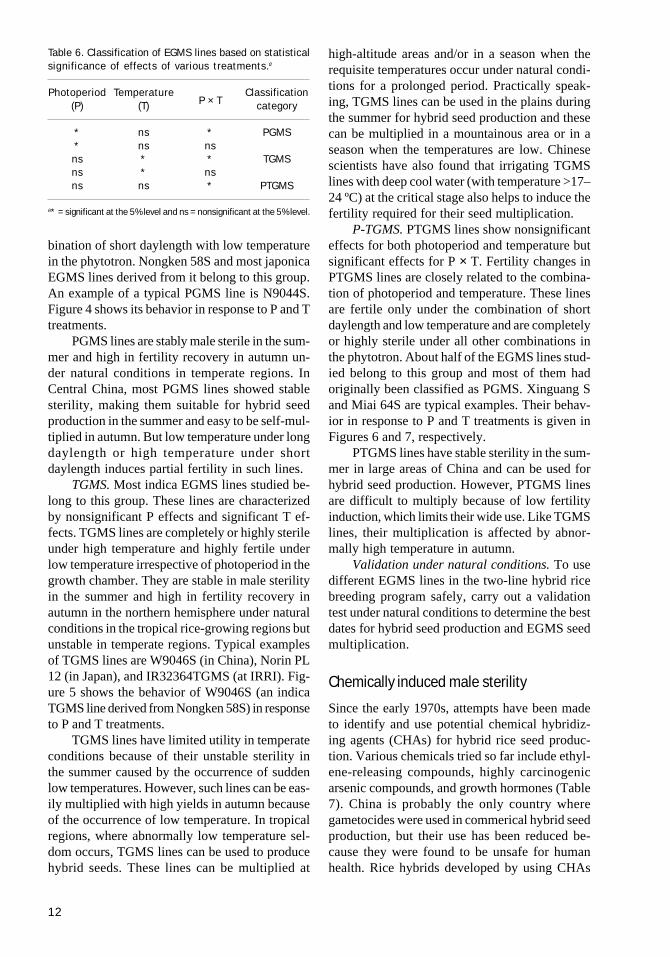
While setting up P and T treatments, you needto consider (1) the characteristics of fertility re-sponses to P and T in the EGMS line and (2) theecological conditions in the target area in whichthe EGMS system will be deployed. In China, sev-eral EGMS lines were evaluated in a combinationof three P and three T treatments (see Table 4).
Data processingPollen and spikelet fertility of the test entries mustbe analyzed on time. Pollen samples must be col-lected at the time of heading by taking five apicalspikelets from the panicle on the primary tillerfrom each plant and immersing them in a preparedfixative solution (alcohol:acetic acid, 3:1). Per-cent fertile pollen is observed under a microscopeusing the IKI staining procedure (Appendix 1). At
the same time, panicles on primary and secondarytillers that head synchronously with the main stem(±4–5 days) are bagged. At maturity, spikelet fer-tility percent is calculated by the number of filledspikelets divided by total spikelets per panicle,multiplied by 100.
Both pollen and spikelet fertility data fromthe test EGMS lines need to be transformed in theform sin–1 x before analyzing them further.
Two-factor variance analysis can be used tomeasure P, T, and P × T effects (Table 5).
EGMS lines can be classified on completionof the variance analysis as summarized in Table 6.
Different types of EGMS are characterized inthe next section.
Characteristics of different EGMS typesPGMS. Fertility change in PGMS lines is charac-terized by significant P and P × T interaction ef-fects (P<0.05) and nonsignificant T effects. PGMSlines are completely or highly sterile under thecombinations of long daylength with high or lowtemperature and highly male fertile under the com-
Table 4. Photoperiod and temperature conditions inthe phytotrons for evaluation of EGMs lines in Chinafrom 1993 onward.a
Time Low temp. Medium temp. High temp. (oC) (oC) (oC)
0500–0800 22.0 23.0 29.00800–1100 25.0 26.0 32.01100–1500 27.0 28.0 34.01500–1800 25.0 26.0 32.01800–0100 22.0 23.0 29.00100–0500 19.0 20.0 26.0Daily average 23.1 24.1 30.1
aIllumination time: 12.5 h (0600–1830), 14.0 h (0530–1930), 15.0 h(0500–2000).
Table 5. Results of two-factor variance analysis.
Source of Degrees of Sum of Mean of Calculated F Tabular F0.05variance freedom squares squares value value
Replication r – 1Treatment pt – 1
P p – 1T t – 1
P × T (p – 1)(t – 1)Error (r – 1)(pt – 1)Total rpt – 1
12
bination of short daylength with low temperaturein the phytotron. Nongken 58S and most japonicaEGMS lines derived from it belong to this group.An example of a typical PGMS line is N9044S.Figure 4 shows its behavior in response to P and Ttreatments.
PGMS lines are stably male sterile in the sum-mer and high in fertility recovery in autumn un-der natural conditions in temperate regions. InCentral China, most PGMS lines showed stablesterility, making them suitable for hybrid seedproduction in the summer and easy to be self-mul-tiplied in autumn. But low temperature under longdaylength or high temperature under shortdaylength induces partial fertility in such lines.
TGMS. Most indica EGMS lines studied be-long to this group. These lines are characterizedby nonsignificant P effects and significant T ef-fects. TGMS lines are completely or highly sterileunder high temperature and highly fertile underlow temperature irrespective of photoperiod in thegrowth chamber. They are stable in male sterilityin the summer and high in fertility recovery inautumn in the northern hemisphere under naturalconditions in the tropical rice-growing regions butunstable in temperate regions. Typical examplesof TGMS lines are W9046S (in China), Norin PL12 (in Japan), and IR32364TGMS (at IRRI). Fig-ure 5 shows the behavior of W9046S (an indicaTGMS line derived from Nongken 58S) in responseto P and T treatments.
TGMS lines have limited utility in temperateconditions because of their unstable sterility inthe summer caused by the occurrence of suddenlow temperatures. However, such lines can be eas-ily multiplied with high yields in autumn becauseof the occurrence of low temperature. In tropicalregions, where abnormally low temperature sel-dom occurs, TGMS lines can be used to producehybrid seeds. These lines can be multiplied at
high-altitude areas and/or in a season when therequisite temperatures occur under natural condi-tions for a prolonged period. Practically speak-ing, TGMS lines can be used in the plains duringthe summer for hybrid seed production and thesecan be multiplied in a mountainous area or in aseason when the temperatures are low. Chinesescientists have also found that irrigating TGMSlines with deep cool water (with temperature >17–24 ºC) at the critical stage also helps to induce thefertility required for their seed multiplication.
P-TGMS. PTGMS lines show nonsignificanteffects for both photoperiod and temperature butsignificant effects for P × T. Fertility changes inPTGMS lines are closely related to the combina-tion of photoperiod and temperature. These linesare fertile only under the combination of shortdaylength and low temperature and are completelyor highly sterile under all other combinations inthe phytotron. About half of the EGMS lines stud-ied belong to this group and most of them hadoriginally been classified as PGMS. Xinguang Sand Miai 64S are typical examples. Their behav-ior in response to P and T treatments is given inFigures 6 and 7, respectively.
PTGMS lines have stable sterility in the sum-mer in large areas of China and can be used forhybrid seed production. However, PTGMS linesare difficult to multiply because of low fertilityinduction, which limits their wide use. Like TGMSlines, their multiplication is affected by abnor-mally high temperature in autumn.
Validation under natural conditions. To usedifferent EGMS lines in the two-line hybrid ricebreeding program safely, carry out a validationtest under natural conditions to determine the bestdates for hybrid seed production and EGMS seedmultiplication.
Chemically induced male sterilitySince the early 1970s, attempts have been madeto identify and use potential chemical hybridiz-ing agents (CHAs) for hybrid rice seed produc-tion. Various chemicals tried so far include ethyl-ene-releasing compounds, highly carcinogenicarsenic compounds, and growth hormones (Table7). China is probably the only country wheregametocides were used in commerical hybrid seedproduction, but their use has been reduced be-cause they were found to be unsafe for humanhealth. Rice hybrids developed by using CHAs
Table 6. Classification of EGMS lines based on statisticalsignificance of effects of various treatments.a
Photoperiod TemperatureP × T
Classification(P) (T) category
* ns * PGMS* ns nsns * * TGMSns * nsns ns * PTGMS
a* = significant at the 5% level and ns = nonsignificant at the 5% level.

13
have been tested along with 3-line bred hybridsand were reported to give consistently comparableand often higher yields. Over the years, seed yieldshave increased from 0.4 t ha–1 with 40–60% seedpurity to 1.5 t ha–1 with 80–90% seed purity. CHAsmust be able to selectively induce total male ste-rility. The effectiveness of CHAs is highly stage-specific (i.e., these should be applied at the sta-men and pistil primordia formation stage or stageIV) and genotype-specific (i.e., the gametocidaleffect varies from variety to variety). In India,oxanilates, when sprayed at stage IV (meioticstage) of rice development, were found to be ef-fective and variety Pusa 150 was sterilized moreeffectively by the gametocidal spray than othervarieties, thus indicating genotype specificity (Ali1993). Figure 8 illustrates an example of two-linehybrid rice seed production using a CHA.
Properties of an ideal CHAAn ideal CHA should have the following proper-ties:
1. Wide-spectrum action to induce sterility insuccessively emerging panicles.
2. Selective and total sterilization of stamenswithout affecting ovular fertility.
3. Be less phytotoxic, noncarcinogenic, andwithout residual toxicity that could harmhuman beings and animals.
4. Be easy to apply and economical.
Advantages of the two-line approachvia CHA
1. A wide range of varieties can be used formaking superior hybrid combinations.
2. The method of seed production is simplerthan that of three-line breeding as it does
Table 7. List of chemical hybridizing agents and their efficacy on the rice plant.
Chemical Male sterility induction Reference
Ethrel Partial to high Perez et al (1973)Partial to high Cheng and Huang (1978, 1980)Partial to high Parmar et al (1979)Partial to high Chan and Cheah (1981)High Wang and Que (1981)Partial to high Kaul (1988)Partial to high Song et al (1990)
Ethrel + isourea High Kitaoka et al (1991)
ArsenatesSodium methyl arsenate (MG2) Complete Chen et al (1986), Ali (1990, 1993)Zinc methyl arsenate (MG1) Complete Anonymous (1978)Monosodium methane arsenate Complete Wang and Que (1981)
OxanilatesEthyl 4′ flouro oxanilate Complete Ali (1990, 1993)Ethyl 4′ metho oxyoxanilate Complete Ali (1990, 1993)Ethyl 4′ bromo oxanilate High Ali (1990, 1993)Ethyl 4′ chloro oxanilate High Ali (1990, 1993)
Other chemicals (with and without codes)RH 531 Complete Perez et al (1973)DPX 3778 Prevents anther dehiscence Long et al (1973)3-(-p-chlorophenyl) 6-methoxy-s-triazine High to complete Zhangzing and Chunnong (1980) –2-4 (1H,3H) dione tri-ethanolamineSodium sulfate Complete Wang et al (1981)HRG 626 Complete Takeoka et al (1990)Ammonia sulfonic acid High Chen (1985)HAC 123 + N 312 Complete Luo et al (1988)MHC Complete Song et al (1990)CRMS Complete Wang et al (1991a,b,c)Kasugamycin Partial to high Atsumi et al (1992)AOA High Astumi et al (1992)

14
not require development of three lines (A,B, and R).
3. If the CHA is not effective because ofnonsynchronization of flowering or con-tinuous rain during the critical stage, heavycrop losses can still be averted because theyield of the sprayed unaffected female wouldstill be good enough.
4. Partial CMS lines and EGMS lines can bemade completely sterile with a spray of anideal CHA.
5. The narrow genetic base for cytoplasmicgenic male sterility, inherent in three-linerice hybrids, ceases to be a problem in CHA-derived hybrids.
Disadvantages of CHAs1. Production of impure hybrid seeds if the
CHA is not effective because of unfavor-able weather conditions or nonsynchronizedtillering and growth.
2. Health hazards of some CHAs (such as zincmethyl arsenate or sodium methyl arsenate).
3. High cost of the chemicals.Fig. 8. Schematic description of the use of a CHA todevelop two-line hybrids.
,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,�-�-�-�-�-�-�-�-�,�,./ ,/ ./ ,/ ./ ,/ ./
�����������
����������
���� ���������
���� ���������
���� ���������
����#�������#�#������������ �0�,/���./�
�1�1 �1 �1
,1 � .10 � � �,/ ./0 ,2 .20��� , .
���%���/������ �����'�������������

15
CHAPTER 3
Comparative organizationof two- and three-line hybridbreeding programs
Organization of a hybrid breeding program is strik-ingly different from the conventional inbred breed-ing program in rice. Three-line and two-line hy-brid rice breeding have some similarities as wellas differences in their organization. These are de-scribed in this chapter.
Similarities in three-line and two-linebreeding nurseriesThe breeding nurseries that are identical in bothbreeding programs are as follows.
Source nurseryThis nursery contains elite lines that have the po-tential to become parents of commercial hybrids.Include the best available CMS, TGMS, and PGMSlines in this nursery. To raise the source nursery,grow 20 plants (with a single seedling per hill) perline in rows. Use these as single plants for test-crosses with the best available CMS or EGMSlines. To synchronize flowering and to make asmany testcrosses between elite lines as possibleand to make sterile lines, plant the sterile lines onthree dates with a 10-day interval.
Testcross nurseryThis nursery contains the testcrosses made in thesource nursery along with the single-plant prog-enies of the male parents used to make the test-crosses. After growing every ten pairs of testcrossand the corresponding male parent progenies,grow single rows of inbred and hybrid checks. In athree-line hybrid breeding program, screen thetestcross progenies for pollen sterility/fertility,spikelet fertility, and other agronomic traits toidentify potential maintainers and restorers and
heterotic hybrids. In the two-line hybrid breedingprogram, screen the testcross progenies for pollenfertility, spikelet fertility, and other agronomictraits to identify the potential elite male parentsand heterotic hybrids. Plant the F
1 and their male
parents side by side in three rows each with a singleplant per hill.
Combining ability nurseryThis breeding nursery contains a set of crossesderived from promising CMS and restorer lines(in the three-line breeding program) and promis-ing EGMS and elite pollen parent lines (in thetwo-line breeding program) made in a line × testerdesign to evaluate the parental lines for their com-bining ability, that is, their ability to produce su-perior progenies when crossed with several maleparents.
Yield evaluation nurseriesThe procedure adopted for heterosis evaluationfor two- and three-line breeding is similar. It con-tains the following five different trials:
1. Observational yield trial (OYT). This con-tains hybrids derived from commerciallyusable CMS lines (showing stable pollensterility, high general combining ability,and good outcrossing potential) and prom-ising restorer lines identified in the testcrossnursery. The trial also contains inbred checkvarieties of different growth duration.Groups of test hybrids are compared with aset of inbred checks and hybrids. Thus, thetest hybrids are unreplicated, whereas thecheck varieties and hybrids are replicatedacross different groups. The OYT uses anaugmented design in which test hybrids are

16
arranged in groups. Each plot is about 6 m2,in which 125–175 single plants per hill areincluded. Data on agronomic traits, yield,and disease and insect pest resistance arerecorded.
2. Preliminary yield trial (PYT). The prom-ising hybrids that yield 15–20% higher thanthe check varieties in the testcross nursery,combining ability nursery, and OYT are for-warded for further evaluation in the PYT.Hybrids are grouped according to their ma-turity duration. This trial uses a randomizedcomplete block design with 3–4 replica-tions, with an individual plot size of 7 m2.Data on agronomic traits, yield, disease andinsect resistance, and grain quality (of het-erotic hybrids only) are recorded. The plotdesign and data recorded are similar to thoseof the OYT.
3. Advanced yield trial (AYT). The promis-ing hybrid entries from the PYT are includedin the AYT. The plot design and data re-corded are similar to those of the PYT, butthe plot size is 10 m2.
4. Multilocation trials (MLT)/national hy-brid trial (NHT). The outstanding hybridentries in the AYT are nominated into theMLT/NHT. The experimental design anddata collected remain the same as in the AYT,except that the plot size is increased to 15–20 m2.
5. On-farm testing (OFT). One to two hybridsperforming consistently better than the re-gional check and local checks over 2–3years are recommended to the OFT. The plotarea is about 0.1 ha, with or without replica-tion. The best local check variety (hybrid)is used as the control.
Hybrids that pass all the above trials and haveresistance to major insect pests and diseases andacceptable grain quality are registered and/or re-leased.
Differences in three-line and two-linebreeding nurseries
1. Male sterile maintenance and evaluationnurseries
In three-line breeding, CMS lines are maintainedand evaluated in a CMS line maintenance andevaluation nursery by growing A and B lines sideby side. Their pollen sterility/fertility is monitoredand lines are evaluated on a single-plant basis forphenotypic acceptability (on a 1–9 scale, where 1= excellent and 9 = poor), days to 50% flowering,outcrossing ability, and other desired traits. TheCMS lines are maintained by hand-crossing of Aand B lines (on a single-plant basis) to maintaintheir purity. Hand-crossed seeds of the commer-cially usable CMS lines are produced in largerquantities (500–1,000 seeds) so that they can beused for nucleus seed production.
In the two-line breeding program, EGMS linesare maintained by selfing by growing them in ap-propriate daylength and temperature conditionsthat induce fertility. These EGMS lines are evalu-ated for sterility separately by growing them un-der suitable daylength and/or temperature condi-tions in the phytotron and/or field. Simulta-neously, they are also evaluated for phenotypicacceptability, flowering, outcrossing rate, andother desired traits in comparison with suitablechecks (such as popular commercial varieties,popular CMS lines, and the best available EGMSlines), which should be grown side by side.
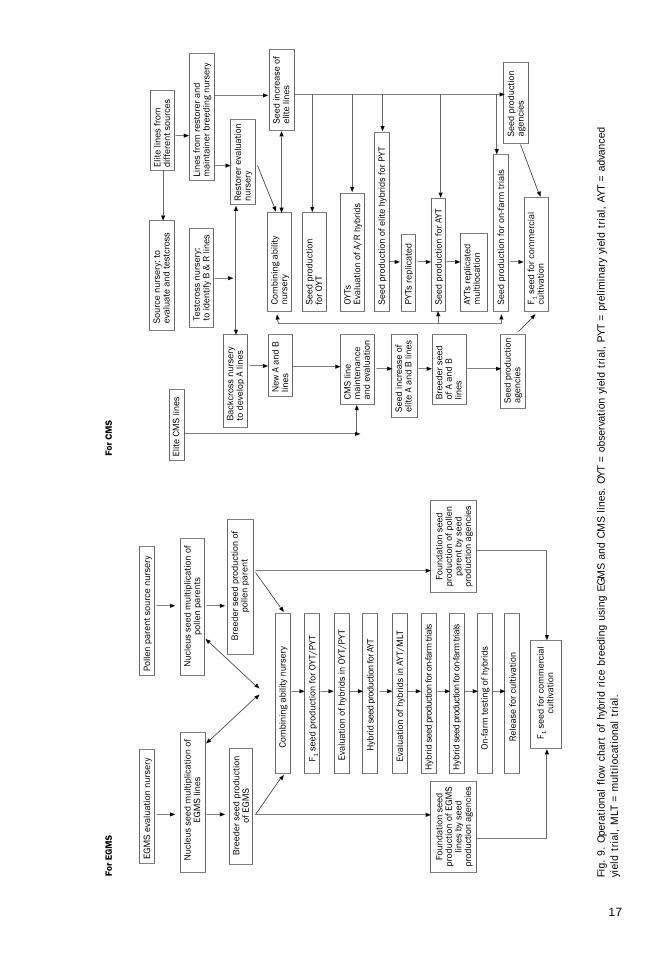
2. EGMS breeding nurseriesThese are described in the next chapter. A com-parative flow chart of two-line and three-line hy-brid rice breeding nurseries appears in Figure 9.
17
Fig.
9.
Ope
ratio
nal
flow
cha
rt o
f hy
brid
ric
e br
eedi
ng u
sing
EG
MS
and
CMS
lines
. OY
T =
obse
rvat
ion
yiel
d tr
ial,
PYT
= pr
elim
inar
y yi
eld
tria
l, AY
T =
adva
nced
yiel
d tr
ial,
MLT
= m
ulti
loca
tion
al t
rial.
������
�����
��� ��� ��
������
����� ��������� �
�
��� �
����������� ������������
�� ���
��� �
����������� ������������
�� ���
������
� ��������� �
�� �����
�� �
��� �
����������� ���������
������
� ��������� �
�� �����
�!��
" #����
�����
�� �
������
���!
��
$��� � ��
��� � �����
���
%���
�����
�����
�� �
����
�����
����
���
%���
�����
�����
�� �
�����&�'
(�
����
����
������ �
� �� �����
�&�'�� �
��(�
����
����
������ �
� �� �����
�����
����
����
�&�'�������
� �����
���
!����
����
�������
����
���
"���
��� �
����
�����
�� �
������&�'
� ���
������
�����
�� �
����
�� ��
"���
��� �
����
�����
�� �
�����������
�����������
�����
�� �
����
�� ��
�� ��
�$�'�� �
��
%�
)��������
�����������
���� ��
�
(�*
������
�%� �
��
$�'�� �
��� ����
���
����������
� ��
'���
� ���
�����
�� ��
������
�%�� ��
�
%���
�����
���������
�%� �
��
'���
�����
� ��
����
��
'���
�����
�+���
������
������
�������
�
������
����
���+
��� �
��� ���%�,���� ��
�
$��� � ��
��� � ��
�����
'���
�����
� ��
����
��
���
���
����
� ���������
���� �
�
'���
�����
� �������� ������ ������!
��
!�����
�� ����
'���
�����
� �������
��
������
�� ����
���� ��
�� ��
'���
�����
� �������
�����
�� ���
" #����
�����
���� ��
��� ��� �
�
'���
�����
� ��
����
��
'���
� ���
�����
�� ��
�� ��
�
�������������� ��
����� ��
�����
����������
�� ��� ��
�����
�����
���
�� ��
�� ��
�����
� �����
�����
��
�����
��
���
���
�
" #����
�����
���� ��
��� ��� �
�

19
CHAPTER 4
Inheritance of EGMS
Procedure for carrying out inheritancestudies on the EGMS trait
The following stepwise procedure explains howinheritance studies on the EGMS trait are con-ducted:
• Purify the EGMS lines and fertile parentsthrough bagging.
• Prepare all generations for genetic analysisand obtain sufficient seeds. The generationsinvolve P
1, P
2, F
1, BC
1F
1, BC
2F
1, and F
2.
• Plant different generations with 10–20 P1,
P2, and F
1 plants, 60–120 BC
1 plants, and
more than 200 F2 plants.
• Observe the pollen and spikelet sterility ofeach plant by putting the plants under ap-propriate sterile environmental conditionsduring the sensitive stage.
• Classify the sterile:fertile plants accord-ingly.
• Use the χ2 test to analyze the segregationpattern and determine the genes that con-trol EGMS.
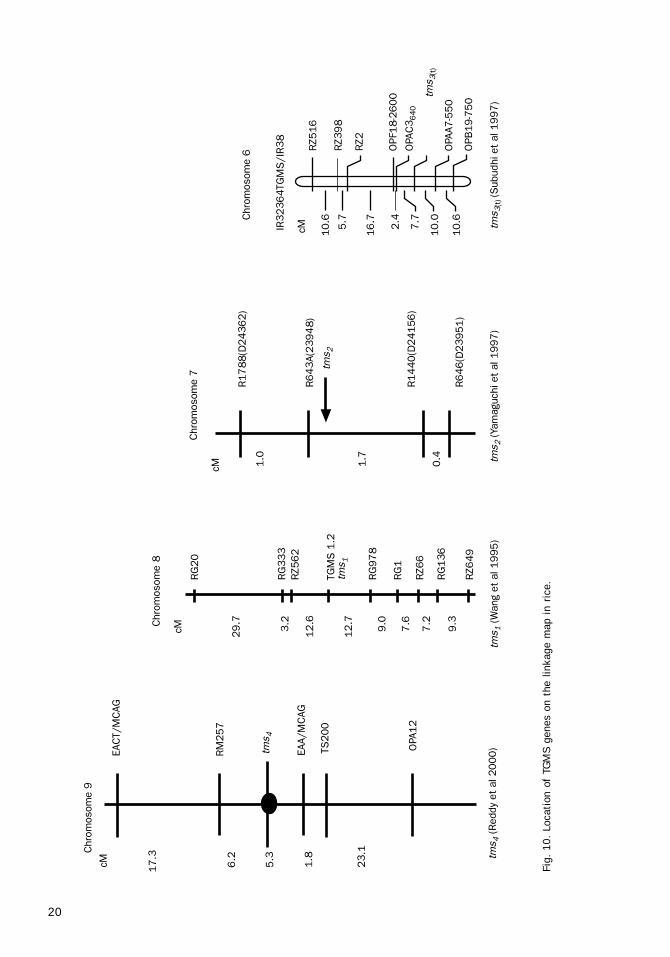
Inheritance of TGMSGenetic studies in Japan, at IRRI, and in India(Maruyama et al 1991, Borkakati and Virmani1993, Ali 1996) indicated that the TGMS trait inNorin PL 12, IR32364S, and SA 2 was controlledby a single recessive gene. Allelic relationshipstudies indicated that the TGMS genes in NorinPL 12 and IR32364S mutants were different.TGMS line 5460s developed in China carries thetms
1 allele on chromosome 8, whereas IR32364S
carries the tms3 allele on chromosome 6. The
TGMS allele in Norin PL 12 is designated as tms2
(Fig. 10). On the basis of this information, otherTGMS sources from India have also been studiedfor allelic relationships and Reddy et al (2000)found a new nonallelic TGMS trait designated astms
4 in SA 2, a sodium azide-induced TGMS mu-
tant. Other details are given in Table 8.The transfer of the Norin PL 12 gene (tms
2)
into IRRI cultivars such as IR68945S, IR68949S,and IR68294S revealed varied sterility-fertility–altering conditions, indicating a change in theexpression of the TGMS trait in a different ge-netic background.
Inheritance of PGMS and PTGMSSingle-locus genetic modelIn an initial study, many conventional japonicaand indica varieties were reciprocally crossed toNongken 58S (NK58S) and all the F
1s were fertile.
In F2 populations of reciprocal crosses between
NK58S and three conventional japonica rice lines,a 3:1 ratio of fertile:sterile plants was observed ineach population grown under long-day conditions.The sterility was therefore considered to be con-trolled by a single recessive nuclear gene. Yang etal (1992a) designated the PGMS gene as Ps. It wasalso noted that minor genes might also be respon-sible for the sterility, as few sterile F
2 plants were
100% sterile. Many other studies showed that fer-tility segregation in crosses between NK58S andNK58 was controlled by a single nuclear gene.
Two-loci genetic modelMany studies showed that PTGMS in NK58S-de-rived P(T)GMS lines was conditioned by reces-sive alleles at two loci.
The two-loci genetic control of PTGMS was
20
Fig.
10.
Loc
atio
n of
TG
MS
gene
s on
the
lin
kage
map
in
rice.
����
���
����
����
���
���
���
���
�
��
�
�
���
�
��
���
��������
��� �
���
��
��
���
���
���
���
��
��
������
����
��
�����
����
��
����
����
��
������
� �
��� �
���� ���
���� ���
���
����
����
�
��
�
�
�
����� �
��
����
����
�
�����
����
���
������
�����
����
������
� !"#
"$"#
%��
� !"#
"$"#
%��
��
� !"#
"$"#
%�
��� �
�&�
��� �
��'()
*�%&�(+��
����
��� �
��,(#
(*-� .
�%&�(+
����
����
� ��&����-/
-0 .�%&�(+����
��
��� !"#
"$"#
%��
����
���
���
��
���� ��
� ���
%001�%&�(+����
��2��������
���
�
2���
����
���
� �
����
�
�����

21
well demonstrated in the work of Mei et al (1990).The F
1, F
2, and BC
1F
1 populations were derived by
crossing NK58, NK58S(R), 1149, and NK58S witheach other. NK58S(R) is a fertility revertant mu-tant of NK58S, whereas l149 is a fertile line devel-oped from the cross of NK58S and japonica vari-ety Nonghu 26. A χ2 test indicated that sterility inNK58S was conditioned by recessive alleles attwo loci. The fertility-restoring alleles inNK58S(R) and l149 are allelic to each other, butthey are nonallelic to that in NK58.
Designating one gene as msph and the other asrfph, the genotypes of NK58S, NK58, and NK58S(R8), a fertility revertant mutant of NK58S, werethe following:
Nongken 58S: msph msph rfph rfph
Nongken 58: Msph Msph rfph rfph
Nongken 58S(R): msph msph Rfph Rfph
Complete dominance is displayed at the Msph
locus and plants possessing Msph would be fertile.When partial dominance is displayed at the Rfph
locus, plants possessing Rfph alone would be par-tially fertile.
Diverse genetic modelResults obtained from crosses between NK58S andjaponica varieties other than NK58 were quitecomplex. Although the fertility segregation in themajority of crosses was attributed to a single domi-nant gene or two dominant genes, it was also evi-dent that multiple genes were involved in the ge-netic control of PTGMS.
The two genes segregated independently andthe interaction between them varied greatly de-pending on the genetic background. Different seg-regation patterns such as 9 fertile:6 partially fer-tile:1 sterile, 9 fertile:3 partially fertile:3 partiallysterile:l sterile, and 9 fertile:3 partially fertile:4sterile were observed (Sheng 1992). A bimodaldistribution was generally observed in the fertil-
ity segregation but, on close scrutiny, the segre-gation was found to be continuously distributedin each population. Mei et al (1990) showed thatthe fertility segregation ratio varied greatly de-pending on the criteria for the classification ofsterile and fertile plants. Although Mei et al (1990)suggested that PTGMS was quantitatively inher-ited, other reports suggested that the complexityof PTGMS was conditioned by a major gene andmultiple minor genes, partial dominance of ster-ile alleles, and dosage effects. In addition, it wassuggested that both major and minor genes mightbe responsible for the genotypic difference in thelow-sterility-inducing temperature. The conceptthat the genetic control of PTGMS is now gov-erned by a set of multiple genes having major andminor effects is well accepted.
Allelic relationship studiesThe PTGMS genes in NK58S-derived japonicalines were allelic to the NK58S gene. In some ofthe NK58S-derived indica lines, genes that werenonallelic to NK58S were also found. AmongPTGMS lines having independent origins, the ste-rility in both Hengnong S-l and 5460S was con-trolled by two recessive nuclear genes, and that inAnnong S-l was controlled by a recessive nucleargene. The allelism test indicated that the genes inNK58S, Annong S-l, 5460S, and Hengnong S-lwere nonallelic to each other. It is now understoodthat five genes were involved in the genetic con-trol of NK58S, NK58S-derived P(T)GMS linesW6154S and W7415S, Annong S-1, and 5460S(Table 9).
Genetic mapping of PTGMSBy employing marker-gene stocks, Zhang et al(1990) found that the major gene for PTGMS msph
in NK58S was linked to d-1 on chromosome 5with a recombination fraction of 28.4% and it seg-regated independently from the other 33 markergenes located on the remaining 11 chromosomesof rice. This result was confirmed by the detection
Table 8. Different alleles for the TGMS trait amongavailable TGMS sources.
Gene symbol TGMS source
tms1 JP 1 and 5460Stms2 IR68945 (Norin PL 12)tms3 JP 8-8-1-S, 1C 10,
IR32364tms4 SA2tms5 Annong S-1rtms1 (reverse TGMS) J207 S
Table 9. Genotypes proposed for PTGMS in five lines.
Line Genotype
Nongken 58S S1S1 S5S5W6154S S2S2 S5S5W7415S S1S1 S4S4 Annong S-1 S2S25460S S3S3 S5S5
22
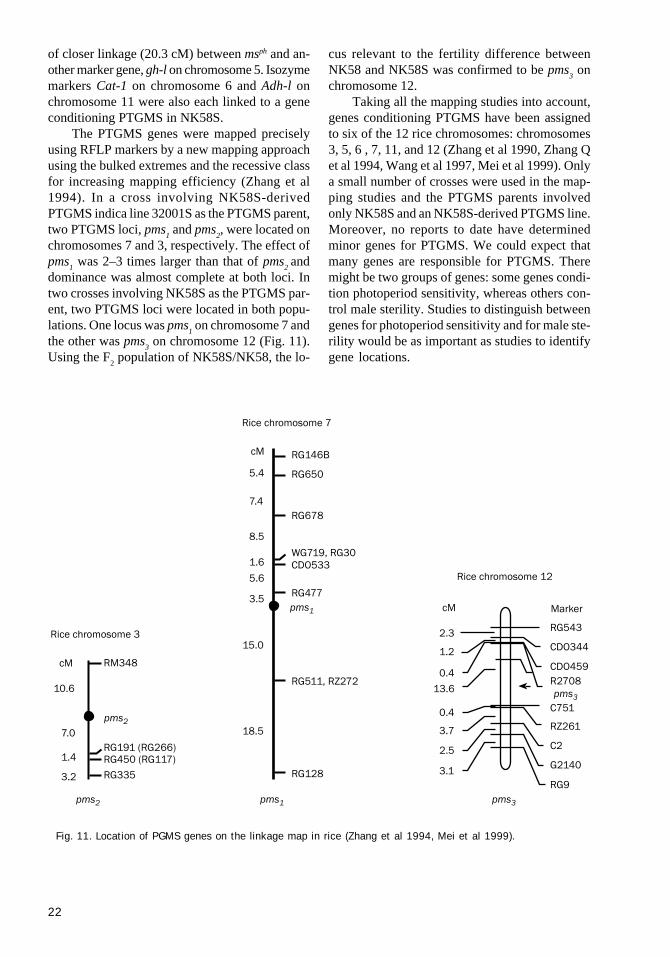
of closer linkage (20.3 cM) between msph and an-other marker gene, gh-l on chromosome 5. Isozymemarkers Cat-1 on chromosome 6 and Adh-l onchromosome 11 were also each linked to a geneconditioning PTGMS in NK58S.
The PTGMS genes were mapped preciselyusing RFLP markers by a new mapping approachusing the bulked extremes and the recessive classfor increasing mapping efficiency (Zhang et al1994). In a cross involving NK58S-derivedPTGMS indica line 32001S as the PTGMS parent,two PTGMS loci, pms
1 and pms
2, were located on
chromosomes 7 and 3, respectively. The effect ofpms
1 was 2–3 times larger than that of pms
2 and
dominance was almost complete at both loci. Intwo crosses involving NK58S as the PTGMS par-ent, two PTGMS loci were located in both popu-lations. One locus was pms
1 on chromosome 7 and
the other was pms3 on chromosome 12 (Fig. 11).
Using the F2 population of NK58S/NK58, the lo-
cus relevant to the fertility difference betweenNK58 and NK58S was confirmed to be pms
3 on
chromosome 12.Taking all the mapping studies into account,
genes conditioning PTGMS have been assignedto six of the 12 rice chromosomes: chromosomes3, 5, 6 , 7, 11, and 12 (Zhang et al 1990, Zhang Qet al 1994, Wang et al 1997, Mei et al 1999). Onlya small number of crosses were used in the map-ping studies and the PTGMS parents involvedonly NK58S and an NK58S-derived PTGMS line.Moreover, no reports to date have determinedminor genes for PTGMS. We could expect thatmany genes are responsible for PTGMS. Theremight be two groups of genes: some genes condi-tion photoperiod sensitivity, whereas others con-trol male sterility. Studies to distinguish betweengenes for photoperiod sensitivity and for male ste-rility would be as important as studies to identifygene locations.
Fig. 11. Location of PGMS genes on the linkage map in rice (Zhang et al 1994, Mei et al 1999).
�� �����
����
�������� ������������������
���
���
���
���
����
�����
�����������
����
����
����
���
��
��
���
���
���
�� ���� �
�� ��
�� ��
���������������
���������
���������� ����
���������� ����
�!�"��
���
��
���
������
���
���
���
���
�����
������
����������
����
��� �
��
�����
��
����
���������� �����
����

23
CHAPTER 5
Breeding proceduresfor developing EGMS lines
EGMS lines can be developed by any of the fol-lowing methods:
• Screening of existing varieties• Induced mutagenesis• Hybridization followed by pedigree selec-
tion, anther culture, backcrossing, andmarker-aided selection (MAS)
Screening of existing varieties for EGMSExisting varieties can be screened to detect spon-taneous EGMS mutants. A large number of ricegermplasm materials should be screened over sea-sons involving various high-temperature ordaylength regimes to identify any spontaneousmutants or lines showing differential pollen andspikelet sterility during different temperature ordaylength regimes occurring during the panicleinitiation stage onward. Rice germplasm consist-ing of photoperiod-sensitive varieties or varietiesadapted to high altitudes provides a higher prob-ability of success when using this method.
During this screening,1. Select plants in which earlier panicles are
partly fertile and later ones are almost ster-ile or vice versa according to the environ-mental conditions. These plants are easilyidentified by the combination of partly filledbending panicles and sterile erect paniclesin the same plant.
2. Study pollen sterility of younger paniclesand determine whether sterility is higherthan 99% (see Appendix 1 for the detailedprocedure).
3. Multiply the suspected plants by separat-ing the tillers and ratooning them.
4. Evaluate plants for their fertility behaviorunder different temperature (TGMS) andphotoperiod (PGMS) using the growth cham-ber or phytotron or under field conditions(as mentioned in Chapter 2).
Induced mutagenesisEGMS lines, particularly TGMS lines, can also bedeveloped by the mutation breeding method de-scribed below.
1. Select the best available high-yielding ricecultivars that are photoperiod-sensitive,cold-tolerant, and adapted to high altitudesfor inducing EGMS mutants. These linesstand a better chance for inducing suchmutations.
2. Select any of the physical (gamma rays, fastneutrons) or chemical (sodium azide, ethylmethane sulfonate, EMS, methyl methanesulfonate, MMS, N-methyl-N-nitrosourea,MNU, etc.) mutagens.
3. Treat the seed material with an appropriatedose of mutagen and grow it as the M
1 gen-
eration, for example, gamma rays—25 Kr;sodium azide—0.002 M, pH 3 for 6 h;EMS—0.1%, pH 7.0 for 6 h; MNU—1.5 mMfor 1 h.
4. Select 1 or 2 panicles from single plants inthe M
1 generation.
5. Grow the M2 progenies under appropriate
temperature (such as >30/24 ºC) anddaylength (such as >13.45 h) conditions andselect plants showing complete sterility ordifferential fertility of panicles within thesame plant. Multiply selected plants byseparating their stubbles and evaluate them

24
under appropriate fertility induction in thegrowth chamber (as shown in Tables 10 and11) or under natural field conditions. Thosethat revert to fertility are suspected EGMSplants. These may be TGMS, reverse TGMS,PTGMS, PGMS, or reverse PGMS depend-ing on their behavior.
Hybridization methodThis is an important and simple method to de-velop new EGMS lines in diverse genetic back-grounds by using already identified and charac-terized EGMS lines. Under this method, the known
EGMS source is crossed as a female parent to anyof the popular high-yielding rice varieties (HYV)possessing good combining ability, disease andpest resistance, and acceptable grain quality. Thismethod is generally used in a situation in whichthe available EGMS line is unadapted or unac-ceptable for one reason or another. The crossesmade above can be handled in any of the follow-ing procedures:
Pedigree selectiona) Pedigree selection procedure using method I
1. Grow F1 plants from the crosses made to
obtain F2 seeds.
Table 10. Simultaneous screening of suspected EGMS lines for identification of type of EGMS.a
Category of growth chamber settings Expected reaction of suspected EGMS on pollensterility/fertility
High temperature (32 oC) and long photoperiod (14 h) Completely sterile (PGMS/TGMS/PTGMS)Fertile (reject/reverse PGMS/reverse TGMS)
High temperature (32 oC) and short photoperiod (12 h) Completely sterile (TGMS/PTGMS/reverse PGMS)Fertile (PGMS/reverse TGMS)
Low temperature (24 oC) and long photoperiod (14 h) Completely sterile (PGMS/reverse TGMS)Fertile (TGMS/PTGMS/reverse PGMS)
Low temperature (24 oC) and short photoperiod (12 h) Completely sterile (reverse PGMS/reverse TGMS accordingly)Fertile (TGMS/PGMS/PTGMS)
aThe same set of lines must be placed in all four chambers simultaneously.
Table 11. Design of experiment to identify and differentiate PTGMS from TGMS lines.a
Photoperiod (daylength duration)Temperature (oC)
12 h 13 h 14 h
>32 Sterile (TGMS and PTGMS) Sterile (TGMS and PTGMS) Sterile (TGMS and PTGMS)(1) (2) (3)
30 • Fertile (partial) TGMS • Fertile (partial) (TGMS) • Fertile (partial) (TGMS)·• Less fertile than the value • Less fertile than the value
in box 4 (PTGMS) in boxes 4 and 5 (PTGMS)(4) (5) (6)
28 • Fertile (partial) (TGMS) • Fertile (partial) (TGMS) • Fertile (partial) (TGMS)• More fertile than the value • Less fertile than the value • Less fertile than the value in
in box 4 (PTGMS) in box 7 (PTGMS) boxes 7 and 8 (PTGMS)(7) (8) (9)
26 • Fertile (partial) (TGMS) • Fertile (partial) (TGMS) • Fertile (partial) (TGMS)• More fertile than the value • Less fertile than the value • Less fertile than the value in
in box 7 (PTGMS) in box 10 (PTGMS) box 11 (PTGMS)(10) (11) (12)
24 Fertile (TGMS and PTGMS) Fertile (TGMS and PTGMS) Fertile (TGMS and PTGMS)(13) (14) (15)
aLines that show influence of temperature and photoperiod are PTGMS, whereas the lines that show no influence of photoperiod are TGMS. Pollenfertility % is a better measure than spikelet fertility provided anthers from the top five spikelets at panicle emergence are used for making a squashpreparation with 1% IKI stain and the pollen fertility/sterility of the emerged panicle is related to the given treatment in the phytotron.

25
2. Grow F2 populations of the crosses made
under appropriate temperature (for TGMS)or daylength (for PGMS) conditions to iden-tify sterile single plants that combine mostof the useful traits of both parents. Grow thepopular HYV and the donor EGMS line be-side the F
2 population to select comparable
plants. Raise stubbles of the selected sterilesingle plants under fertility induction con-ditions to produce F
3 seeds. Seed setting on
the ratooned plants identified earlier as malesterile will confirm the EGMS trait.
3. Grow F3 progenies of the selected EGMS
plants under appropriate fertility inductiontemperature or daylength conditions to se-lect desirable plants and harvest F
4 seeds.
Concurrently evaluate the F3 progenies in
sterility-inducing temperature or daylengthconditions to select completely sterile F
3
progenies to advance the generation.4. Handle F
4 and F
5 progenies in the same man-
ner as F3 progenies.
5. To confirm stability for sterility of an EGMSline in any given generation, trials can beconducted under varying temperature anddaylength regimes in various places.
6. Characterize identified TGMS or PGMSlines for specific defined environmentalconditions at the sensitive stage both in thegrowth chamber and under field conditions.
7. Test the specific TGMS and PGMS linesacross various locations and seasons to iden-tify suitable locations for hybrid seed pro-duction and EGMS self seed multiplication.
b) Pedigree selection procedure using method IIThis method is deployed when we do not have alow temperature/shorter photoperiod conduciveto fertility near where the research is conducted orthe specific environmental condition is not avail-able during the same season of selection.
1. Grow the F2 population under a sterility-in-
ducing temperature regime. Raise the stan-dard controls as mentioned earlier besidesthe EGMS and the other elite parent usedfor hybridization to develop superior EGMSlines. Select desired fertile plants in the seg-regating population.
2. Grow the F3 to F
5 generations under steril-
ity-inducing temperature and select 8–10desirable fertile plants from the progenyrows segregating for sterility. The reason for
selecting so many fertile plants in the segre-gating population is to ensure the probabil-ity of selecting at least one heterozygousfertile plant that would segregate for steril-ity in the next generation.
3. Grow F5 and F
6 populations under sterility-
inducing temperature or longer photoperiod.Select the most desirable male sterile plantsand ratoon them.
4. Transfer the ratooned male sterile plants toa phytotron or glasshouse with a day/nighttemperature of 25/19 ºC or shorter daylength(12 h) to induce fertility.
5. Select those plants that revert to fertilityunder low-temperature or shorter photope-riod conditions and collect their seeds. Theseare suspected TGMS/PGMS plants.
6. Grow progenies of the suspected TGMS/PGMS plants under sterility-inducing tem-perature/photoperiod conditions in the fieldand select those plants as TGMS/PGMS thatgive completely male sterile progenies.
Backcrossing after hybridizationBackcrossing is the most suitable method whenwe need to transfer oligogenes in a recessive con-dition to an already established variety as the re-current parent.
The procedure involves the following steps:Select a stable and suitable TGMS/PGMS donorwith well-defined critical sterility/fertility pointsand cross it with an established variety to whichthe TGMS/PGMS trait has to be transferred.
1. Grow the F1 generation.
2. Grow the F2 generation to select suspected
EGMS plants; ratoon these plants to con-firm their TGMS/PGMS nature.
3. Backcross the EGMS plants with the recur-rent parent.
4. Repeat steps 2 and 3.5. After every two backcrosses, one generation
of selfing is required to verify that the reces-sive EGMS trait is being carried forward asdepicted in Figure 12. After six generationsof backcross, self the BC
6F
1 to select sterile
plants under high temperature (>30 ºC day/24 ºC night)/long photoperiod (14 hdaylength). Ratoon plants may be sent tothe growth chamber or suitable natural con-ditions conducive to fertility reversion.
6. The TGMS development procedure is shownin Figure 13 but the same is applicable to
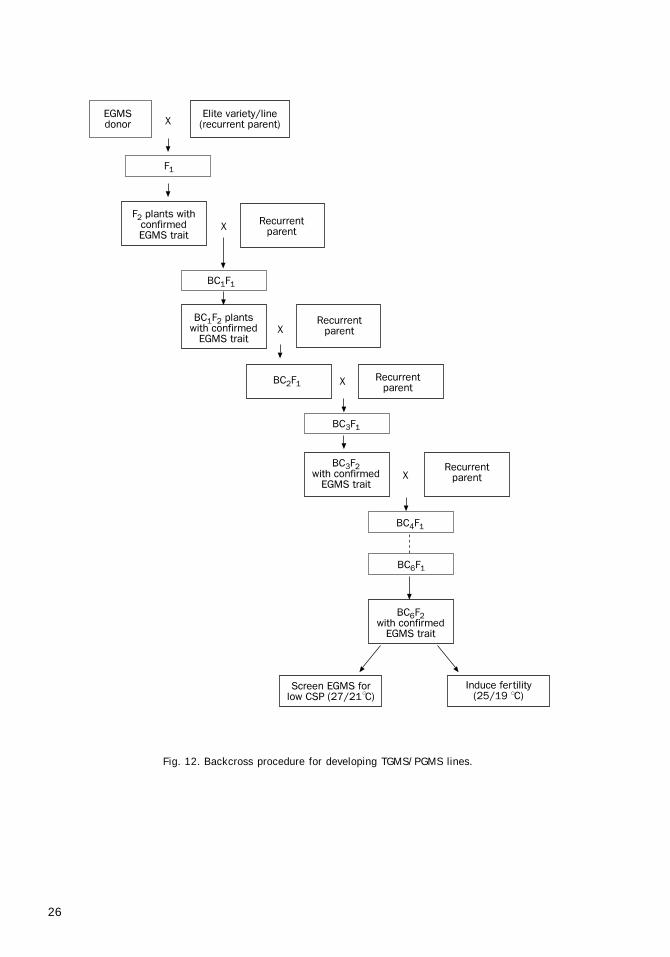
26
�����
����� �����������������������
������������������ !���
�����"#���������������$#���
� ������
��
���$�#�%����&�����'�� �����#��
(�����$#��
�����
������$�#�%���&������'�� �����#��
(�����$#��
(�����$#��
�����
��)��
��)�����&������'�� �����#��
(�����$#��
��*��
��������&������'�� �����#��
+
+
+
+
+
Fig. 12. Backcross procedure for developing TGMS/PGMS lines.

27
PGMS since the screening procedure variesonly under the BC
nF
2 (n = 1, 3, 5) genera-
tion. To develop PGMS through the back-cross procedure, long-day conditions (>14h) will be required for screening in theBC
1F
2, BC
3F
2, and BC
5F
2 generations to
identify the recessive sterile genotypes.
Anther culture or pollen culturefollowing hybridizationAnther culture or pollen culture accelerates thebreeding process and increases selection efficiency
in the breeding of EGMS lines over conventionaltechniques. To use this method, rice breedersshould work closely with a tissue culture expert.
1. Make specific crosses to combine stableTGMS or PGMS lines with promising localcultivars. Grow F
1 plants of these crosses.
2. Produce dihaploid lines through anther cul-ture using anthers or pollen grains from theF
1 plants, adopting standard available pro-
tocols for rice (Fig. 13). (See also AppendixII.)
Fig. 13. Procedure for producing and evaluating dihaploid EGMS lines.
,�������������-�#�&��������-�#�$�#������#%$�#��.����'����.�#����/����%%���������%$��#��%��
���$�#��#�&�%�$�#��
(.�#���
��#���%
�0��$���$�#�%
�������������"#��#����/���%%���������%$��#��%�#��$�#��/���%
0��"#��#������#���/��$�#��/���%
�#���%

28
3. Seeds obtained from the dihaploid plantsshould be sown and raised under sterility-conducive conditions for the selection ofmale sterile lines. Monitor the pollen steril-ity of anther/pollen culture–derived plants.
4. Ratoon sterile dihaploid plants and placethem under low temperature and shorter pho-toperiod to induce fertility and get selfedseeds.
5. Screen for TGMS and PGMS lines based onthe criteria for ideal EGMS lines.
6. TGMS lines such as 6442S, 1286S, HS-1,and HS-5 and one PTGMS line, Lu Guang2S, have been developed through antherculture and extensively tested in Jiangxi,Sichuan, and Fujian provinces of China (Fig.14).
���������������� �������������
�#���#�%��������%%�.
1�&��������
2�������#�
��������#���#�"#��#���
(.��#���%�
�3#������.�%��#���
�3#�������#%
0��.�#������$��$#.#��� 0��.�#���
,�/�$�#���%
�&#�#����4#���
��,� ����%
,��5��
/���.
���%%�.
0�/����%�$��������
EGMS gene transfer and pyramidingthrough marker-aided selection (MAS)after hybridizationSeveral closely linked markers have been identi-fied with TGMS and PGMS genes to use the MASapproach. These markers are listed in Table 12and can be used for selection of the TGMS/PGMStrait in segregating F
2 and backcross breeding
populations without actually screening them un-der specific field or phytotron conditions. TheMAS approach can enhance the speed and effi-ciency for selecting EGMS plants without expos-ing them to conditions for expression of the EGMSgene.
MAS also allows pyramiding of differentsources of TGMS or PGMS/PTGMS alleles into acommon genetic background. It would be inter-
Fig. 14. Procedure for indica two-line hybrid rice breeding through anther culture (modifiedfrom Zhu et al 1999).

29
esting to study the manifestation of pyramidedEGMS genes under different environmental con-ditions. Pyramiding different sources of allelesmay improve the EGMS lines for their critical ste-rility points (CSP) and their critical fertility points(CFP). A low CSP–low CFP line will be useful intropical conditions. Four different TGMS genes—tms
1, tms
2, tms
3, and tms
4—exist, while there are
three reported PGMS genes—pms1, pms
2, and pms
3.
Efforts are under way to pyramid different TGMSgenes in China, at IRRI, and in India (Fig. 15).
tms1 × tms2 tms3 × tms4
tms1/2 × tms3/4
tms1/2/3/4
Select with the help ofclose-flanking molecularmarkers
Test these pyramided linesunder different ecologicalsituations
Fig. 15. Procedure for TGMS gene pyramiding.
Table 12. Closely linked flanking molecular markers for EGMS genes.
Trait Gene Chromosome Closest flankingmarkers
TGMS tms1 8 RZ562–RG978tms2 7 R643A–R1440 (D24156)tms3 6 OPAC3640–OPAA7550tms4 9 RM257–TS200rtms1 10 RM239–RG257
PGMS/PTGMS pms1 7 RG477–RG511, RZ272pms2 3 RG348–RG191 (RG266)pms3 12 R2708–RZ261/C751

31
CHAPTER 6
Characterizing EGMS lines underfield and controlled conditions
Photoperiod varies in a gradual and periodicalmanner year-round based on the latitude and so-lar terms. For a given place, the photoperiod onone day over different years is the same and, on agiven date, the photoperiod in different locationsat the same latitude is also the same. Likewise, thedaylength at different latitudes in the northernhemisphere is the same on the spring (21 March)and autumn equinox (23 September) because thesun radiates vertically on the equator on thosedays (Table 13). The longest photoperiod is ob-served on the summer solstice, when the sun radi-ates vertically on the Tropic of Cancer, and theshortest photoperiod is on the winter solstice, whenthe sun radiates on the Tropic of Capricorn. Datain Table 13 also indicate that the higher the lati-tude, the longer the photoperiod on 22 June (sum-mer solstice) and the shorter the photoperiod on22 December (winter solstice). Likewise, in thesouthern hemisphere, similar conditions exist ex-cept that the winter and summer solstice changes.The winter solstice occurs in the southern hemi-sphere on 22 June when the daylength is shortest,whereas the summer solstice occurs on 22 Decem-ber when the daylength is longest. The higher thelatitude, the longer the observed photoperiod.Under the winter solstice on 22 June in the south-ern hemisphere, the higher the latitude, the lowerthe photoperiod or daylength (Table 13).
The temperature at any given location is in-fluenced by factors such as solar radiation, lati-tude, altitude, local topography, and atmosphericand marine phenomena such as El Niño and LaNiña. Many locations in the tropics still have lessinfluence from the abovementioned environmen-tal factors on temperature and therefore those lo-cations are quite stable in temperature and can be
easily identified based on 25 years of meteoro-logical data.
Characterization of EGMS linesunder field conditionsDetailed meteorological data (such as minimumand maximum temperature, daylength, sunshinehours, humidity, etc.) are essential to characterizeEGMS lines at a given location. It is better if dataof the past 10–15 years are available. To charac-terize given EGMS lines, the following procedurecan be used:
1. Identify 3–4 distinct periods of high andlow temperatures during the year. Likewise,determine the longer and shorter daylengthdurations during the year and over loca-tions.
2. Seed/plant EGMS lines at 15–25-day inter-vals in such a way that their heading coin-cides with the high temperature or longerphotoperiod.
3. Study pollen fertility of the EGMS linesfrom the top five spikelets of primarypanicles under the microscope.
4. Relate the pollen sterility data to tempera-tures/daylengths prevailing during the pe-riod of 15–25 days before heading (“track-ing technique”). The temperature ordaylength that is just sufficient to make theplant completely pollen sterile must benoted among the several temperature ordaylength regimes to which plants were ex-posed during the period. Such a tempera-ture point or daylength at which completepollen sterility is obtained is termed thecritical sterility point and the number of days
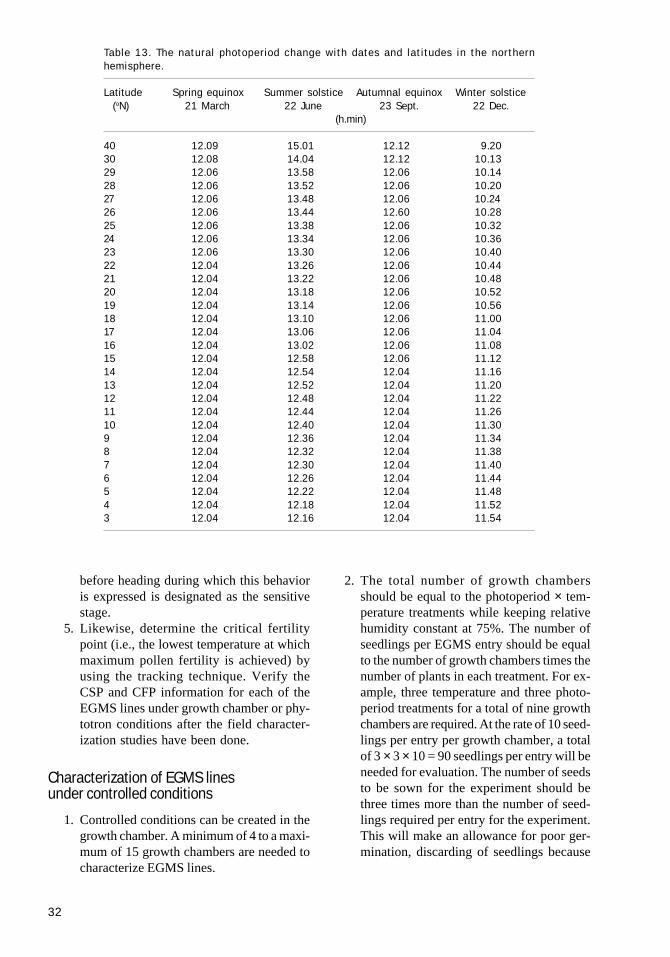
32
before heading during which this behavioris expressed is designated as the sensitivestage.
5. Likewise, determine the critical fertilitypoint (i.e., the lowest temperature at whichmaximum pollen fertility is achieved) byusing the tracking technique. Verify theCSP and CFP information for each of theEGMS lines under growth chamber or phy-totron conditions after the field character-ization studies have been done.
Characterization of EGMS linesunder controlled conditions
1. Controlled conditions can be created in thegrowth chamber. A minimum of 4 to a maxi-mum of 15 growth chambers are needed tocharacterize EGMS lines.
Table 13. The natural photoperiod change with dates and latitudes in the northernhemisphere.
Latitude Spring equinox Summer solstice Autumnal equinox Winter solstice (oN) 21 March 22 June 23 Sept. 22 Dec.
(h.min)
40 12.09 15.01 12.12 9.2030 12.08 14.04 12.12 10.1329 12.06 13.58 12.06 10.1428 12.06 13.52 12.06 10.2027 12.06 13.48 12.06 10.2426 12.06 13.44 12.60 10.2825 12.06 13.38 12.06 10.3224 12.06 13.34 12.06 10.3623 12.06 13.30 12.06 10.4022 12.04 13.26 12.06 10.4421 12.04 13.22 12.06 10.4820 12.04 13.18 12.06 10.5219 12.04 13.14 12.06 10.5618 12.04 13.10 12.06 11.0017 12.04 13.06 12.06 11.0416 12.04 13.02 12.06 11.0815 12.04 12.58 12.06 11.1214 12.04 12.54 12.04 11.1613 12.04 12.52 12.04 11.2012 12.04 12.48 12.04 11.2211 12.04 12.44 12.04 11.2610 12.04 12.40 12.04 11.309 12.04 12.36 12.04 11.348 12.04 12.32 12.04 11.387 12.04 12.30 12.04 11.406 12.04 12.26 12.04 11.445 12.04 12.22 12.04 11.484 12.04 12.18 12.04 11.523 12.04 12.16 12.04 11.54
2. The total number of growth chambersshould be equal to the photoperiod × tem-perature treatments while keeping relativehumidity constant at 75%. The number ofseedlings per EGMS entry should be equalto the number of growth chambers times thenumber of plants in each treatment. For ex-ample, three temperature and three photo-period treatments for a total of nine growthchambers are required. At the rate of 10 seed-lings per entry per growth chamber, a totalof 3 × 3 × 10 = 90 seedlings per entry will beneeded for evaluation. The number of seedsto be sown for the experiment should bethree times more than the number of seed-lings required per entry for the experiment.This will make an allowance for poor ger-mination, discarding of seedlings because

33
of nonuniform growth, and varied growthduration of EGMS lines.
3. Sowing and nursery management. Use thefollowing procedures for sowing and nurs-ery management.• Sow the seeds of EGMS lines with simi-
lar duration and a standard set ofcontrols in the nursery at the same time.
• If there is a significant difference ingrowth duration among the EGMS lines,sow these in two stages (the first stage forlate-maturing material and the secondstage for early maturing material).
• Manage the nursery using routine man-agement to eliminate obvious mixed-riceseedlings and weeds.
• Select uniform and healthy rice seedlingsat the 5-leaf stage for transplanting twoplants per plastic pot with a single plantper hill.
• Use any convenient method to label thepots. As an example, 5-digit labeling canbe used for each plant, that is, 30817means number 3 photoperiod/tempera-ture treatment, 08 is the number of theEGMS line, and 17 is the number of theindividual plant.
• Treat high-photosensitive japonica andindica EGMS lines with a short-day pre-treatment (10 h/28 ºC of photodurationand 14 h/25 ºC of dark duration for 10days) to speed up their development.
• Adjust growth chambers prior to their usefor the experiment. These should be al-lowed to function for one or two daysusing the specified temperature and rela-tive humidity level. A placement planfor each of the pots within a growthchamber should be prepared and thechambers should be sprayed with appro-priate chemicals a few days before mov-ing the plants to control pests and dis-eases.
• Keep the progenies of suspected EGMSplants in separate growth chambers withvarying day and night temperature re-gimes and separate daylength durations.For easy adaptation of the EGMS lines,temperature or daylength should be keptas available in the natural situations ex-emplified as follows:
The lines identified to adapt to specific situationscan be deployed accordingly.
• Place the plants in growth chambers at thecritical stage. The critical stage for photo-period or temperature sensitivity is during5–15 days after panicle initiation (PI). Thesuspected EGMS plants grown in pots areobserved for PI. The plants that are used fordetermining PI by physical opening mustnot be used for the experiment.
• Plants must be placed inside the growthchamber at the PI stage.
• Plants must be placed for 2 weeks or up toheading in the growth chamber for treatment.
• After the plants are treated in the growthchamber, they need to be examined for pol-len sterility from the primary tiller (see Ap-pendix1).
• Observations on pollen sterility must be re-corded in relation to temperature and pho-toperiod.
• Collect a pollen sample from the primarytiller and bag the same panicle for spikeletfertility percent (bagged). Likewise, the pol-len and spikelet sterility of the first threepanicles from each plant need to be recordedseparately as panicles 1, 2, and 3.
• The CSP and CFP vary from genotype togenotype. It is therefore essential to knowthe CSP and CFP of each EGMS line beforeit is used for seed production (Table 2).
• Sterile plants that remain completely sterilein different temperature or photoperiod re-gimes are not considered as EGMS typesand are discarded.
• The EGMS lines that have been strictlyevaluated in phytotrons for their fertility al-teration are advanced for evaluation underdifferent ecological conditions to furtherascertain their suitability for hybrid seedproduction and self seed multiplication.
Situation 1 (maximum-minimum difference of 6 ºC)
Day temperature (ºC) 24 25 26 27 28 30 32
Night temperature (ºC) 18 19 20 21 22 24 26
Situation 2 (maximum-minimum difference of 8 ºC)
Day temperature (ºC) 24 25 26 27 28 29 30 31 32
Night temperature (ºC) 16 17 18 19 20 21 22 23 24

34
Evaluation of EGMS lines
EGMS lines are also evaluated for their pheno-typic acceptability, outcrossing rate, and combin-ing ability as per the methods described for evalu-ating CMS lines. The operational flow chart de-scribing the procedure for using the EGMS sys-tem is presented in Figure 9.
After a thorough examination of the EGMSlines in the growth chamber for their fertility al-teration behavior, they need to be evaluated acrossseveral selected environments. The purpose of thisevaluation is primarily to identify suitable regionsor locations for EGMS self seed multiplicationand hybrid seed production. This evaluation alsoprovides information on their range of adaptabil-ity and their resistance to diseases and insect pests.The key to successful multilocation evaluationlies in the timely layout of the trials and consis-tent observations. It is therefore highly recom-mended that one researcher be in charge of themultilocation evaluation.
Selection of locations based on environmentThe location must represent the rice produc-tion region and must be in the vicinity of theresearch institute/station, with specialistsequipped with minimum scientific instru-ments (e.g., microscope, refrigerator, hot dryoven, daily weather-recording gadgets, seedgerminators, etc.) required for evaluatingEGMS lines.Criteria used for identifying locationsa. Locations that have a different latitude
within the same longitude.b. Locations with distinct differences in to-
pography and temperature within the samelatitude.
c. Locations with different elevations withinthe same region.
There should be proper representation of eco-logical conditions for better understanding of thebehavior of EGMS lines under differentagroclimatic conditions.
For the unified evaluation of EGMS lines, thesowing and transplanting dates must be in accor-dance with the respective locations identified be-cause of the different ecological conditions (Fig.16). For all other purposes, a standard evaluation
procedure must be adopted for drawing meaning-ful conclusions, that is, planting design, pollenand spikelet fertility, and their method of data col-lection should be consistent.
The identification of appropriate areas andseasons for large-scale commercial production ofTGMS and hybrid seed needs special attention,unlike for the PGMS lines, for which the daylengthis quite stable across a given region. On the basisof analysis of about 10–15 years of meteorologi-cal data, areas and ideal periods for seed produc-tion in different parts of the world can be identi-fied.
• Places located 500 and 700 m above sealevel are highly suitable during May to Sep-tember for hybrid seed production and dur-ing November to March for TGMS selfseed production.
• In the choice of place, in hills or coastalplains or interior plains and plateau regions,extra care needs to be taken to ensure thatthe temperature range is <40 to > 16 ºC, be-yond which physiological sterility occurs.
• Four weeks of stable high temperature (>30/>24 ºC day/night) are required for hybridrice seed production interspersing the sen-sitive stage of stage II (secondary branchprimordial) to stage IV (stamen and pistilprimordial stage) of the panicle developmentstage of rice (Fig. 16).
• Similarly, four weeks of low temperature(<26/>16 ºC night) are required for TGMSself seed multiplication, interspersing thesensitive stage of stage II (secondary branchprimordial stage) to stage IV (stamen andpistil primordial stage) of the panicle de-velopment stage of rice (Fig. 16).
• The identified place must be suitable for ricecultivation with space or time isolation.
• Places with naturally chilled irrigation wa-ter of about >17–24 ºC can be useful forTGMS self seed multiplication under high-temperature fluctuations. The chilled irri-gation water is effective from stage IV (sta-men and pistil primordial stage) to stage V(meiotic division).
All the data collected should be stored in acomputer and sent to the researcher in charge foranalysis and interpretation.

35
B. EGMS self seed multiplication
Minimum of four continuous weeks with stable low temperature (<24/>16 oC day/night) or shortphotoperiod (<12 h).
Fig. 16. Graphic illustration for adjustment of sowing time for EGMS for hybrid seed and self seed productionin ideal locations.
Ideal locations with four consecutive weeks with stable temperature or photoperiod
A. Hybrid seed production
Minimum of four continuous weeks with stable high temperature (>30/24 oC day/night) or longphotoperiod (>14 h)
�������������� ��
�� �� �� �� �� �� �� �� �� �� ��
��� ��� ���� ���� ������������ �������� ������� ��������
• Adjust sowing date accordingly so that 15–25 days before flowering falls between the second and thirdweek of high temperature.
• Adjust sowing date accordingly so that 15–25 days before flowering falls between second and thirdweek of low temperature.
��� ��� ���� ���� ������������ �������� ������� ��������
�� �� �� �� �� �� �� �� �� �� ��
�������������� ��

37
CHAPTER 7
Developing pollen parentsfor two-line hybrids
A pollen parent line is defined as a male parentalline that has the ability to restore the fertility inthe F
1 of the EGMS line under a sterile phase re-
gime that ensures complete male sterility. Mostconventional inbred lines are the source of pollenparent lines or pollen parents. For hybrid rice seedproduction using EGMS, any pollen parent linecan restore fertility in the F
1, unlike CMS lines,
for which a set of restorer lines alone can restorefertility in the F
1. It was estimated that about 97%
of the japonica and indica inbred lines restoredthe fertility of japonica PGMS (such as Nongken58S) and indica TGMS (such as W6154S) lines.
Characteristic features of anelite pollen parentThe following are the characteristic features of anideal elite pollen parent:
• Strong fertility-restoring ability. When across is made with the EGMS line, the hy-brids have a normal seed setting percentage(>80%) and are less affected by changes inenvironmental conditions.
• Good general combining ability. Whencrossed with different EGMS lines, the F
1
hybrids from many crosses perform well.• Good agronomic characters. The pollen
parent should be a high-yielding inbred linewith favorable traits for outcrossing, for ex-ample, good anther dehiscence, good an-ther protrusion, large anther size, high pol-len load, etc.
• Genetic distance. Considerable genetic dis-tance from the EGMS lines will be the keyto enhanced heterosis.
Breeding methods for identifying pollenparents for two-line hybrids
The following breeding methods can be used toidentify pollen parents for two-line hybrids:screening available elite inbred lines by testcross-ing with available EGMS lines, hybridization,induced mutagenesis, and anther culture. Amongthese methods, the testcross and hybridizationmethods are the most common.
Screening available elite inbred lines bytestcrossing with available EGMS linesThis is the most effective method to screen forsuitable pollen parents from existing ricegermplasm. In China, most pollen parents for two-line hybrid rice breeding were screened from ex-isting inbred lines. For example, the pollen parentin hybrid Liangyou Peijiu (Peiai64S/9311) wasYangdao b (9311), a medium-season indica vari-ety developed by the Lixiahe Agricultural Insti-tute, Jiangsu Province. The high-yielding indicavariety Teqing developed by the Rice Institute,Guangdong Academy of Agricultural Sciences,was used as the pollen parent for the TGMS linePeiai64S and developed a hybrid, Liangyou Peite,in Hunan Province.
Three fundamental steps are required in mak-ing testcrosses, as discussed below.
1. Preliminary testcross. Use existing elitelines or varieties to cross to EGMS lines,then conduct a preliminary evaluation ofthe F
1 hybrids based on their seed setting,
yield components, grain quality, and resis-tance to diseases and insect pests, etc. At

38
least 10 single plants are needed for a pre-liminary testcross.
2. Re-testcross. From the preliminary testcrossdata, the varieties or lines that performedwell in terms of strong restoration and thatshowed no segregation for agronomic char-acters can be advanced to the re-testcross toconfirm the results of the preliminary test-cross. About 50–100 single plants areneeded for the re-testcross. In the re-testcross,yield will be considered as the key evaluat-ing factor.
3. Identification. Only excellent pollen par-ents (varieties or lines) are selected for fur-ther use. After identification of suitable pol-len parents, hybrid seeds will be producedfor field evaluation and multilocation tri-als.
HybridizationThis is an important and very popular method forbreeding pollen parents for two-line hybrids whenthere is a lack of such parents among existing va-rieties or lines.
1. Single cross. Under this method, two variet-ies or lines with desired traits are crossedand, in the F
2, selection is exercised to iden-
tify single plants that possess the desiredtraits. The standard selection pedigree pro-cedure is followed to fix the lines. At theF
5–F
6 generation, a testcross with EGMS
lines is made. The best-performing pollenparent showing high fertility restoration andheterosis in testcross F
1s is selected for the
production of hybrid seeds and subsequentevaluation in the field.
2. Multiple crosses. This procedure uses twoor more varieties or lines to develop idealpollen parent lines. The procedure is almostthe same as the single-cross method exceptthat in the F
2 generation the selections must
be made to combine the favorable traits fromall the parents used in the multiple cross.
3. Backcross. For a pollen parent line with sev-eral good traits except for one or two unde-sirable traits, the backcross method of breed-ing is the most suitable for removing andreplacing the undesirable traits with favor-able traits from the donor parent. Select astable and suitable variety or line as the traitdonor and cross it with the pollen parentline (recurrent parent) that needs to be im-
proved. Raise the F1 generation and back-
cross it with the recurrent parent and BC1F
1
seeds obtained. The selection strategy mayvary according to the dominant or recessivetrait. For a dominant trait, it is simple to se-lect the plants with the target trait in thebackcross generations. In the BC
5F
1 genera-
tion, self them once. After selfing, the se-lected plants are used to test-cross to screenthe strong F
1 hybrids for evaluation. For the
recessive trait, care must be taken to ensurethat the trait is selected and confirmed byselfing after every two generations of back-crossing.
New strategies for developingpollen parent linesThe main difference in pollen parent and restorerlines is that the former may not have special re-storer genes, that is, Rf, like the latter, therebymaking them more convenient for using the het-erosis between indica and japonica lines in two-line hybrid breeding.
To overcome hybrid sterility, longer growthduration, taller plant types, and incomplete grainfilling, wide-compatibility genes can be used inthe development of pollen parent lines.
Exploiting intersubspecific indica ××××× japonicacrosses through the wide-compatibilitysystemWhat is wide compatibility? It is well known thatindica and japonica crosses result in hybrid steril-ity despite their high heterosis for various agro-nomic traits. But, it was found that certain indicaand japonica hybrids showed normal spikelet fer-tility. One or both parents of these crosses mustpossess a dominant wide-compatability gene (S5n)and such lines are designated as wide-compatibil-ity varieties (WCVs). When such WCVs arecrossed with indica or japonica, the hybrids shownormal spikelet fertility.
The concept of wide compatibility in rice wasfirst introduced by Ikehashi and Araki (1986) toexplain hybrid sterility at the subspecies level.To study the interactions between the sporophyteand gametophyte, Ikehashi and Araki (1986) con-ducted a survey for WCVs via a triple testcross.They found that varieties such as Ketan Nangka,Calotoc, and CPSLO-17 showed wide compatibil-ity. The fertility of their hybrids was linked to the

39
cytochrome (C) and waxy (wx) gene. The WC genewas located on chromosome 6 and three alleles ofthe locus were identified: S-5i from indica rice, S-5j from japonica rice, and S-5n from WC rice. Thesterility resulting from crosses between indica andjaponica rice is due to the interaction between thealleles S-5i and S-5j. The S-5i/S-5j genotype pro-duces semisterile panicles because of the partialabortion of female gametes carrying the allele S-5j. Such abortion does not occur in S-5n/S-5i andS-5n/S-5j genotypes. The donor of S-5n is referredto as a WCV (Table 14).
Developing pollen parents with wide com-patibility. Pollen parents with wide compatibilitywere developed by introducing the WC gene intoconventional indica and japonica rice.
Pollen parents are generally crossed with WClines and then selection is made in the segregat-ing generation using marker genes linked withthe WC trait. Sometimes, the backcross method isused when the traits of WCVs are not ideal.
In China, at the National Rice Research Insti-tute (CNRRI), pollen parent R2070 with WC wassuccessfully developed by using the crossMinghui 63 (Lunhui422 and javanica lineWL1312). R2070 can be used in both three-lineand two-line hybrid rice breeding. The three-linehybrid II You 2070 (II-32A/R2070) and two-linehybrid Guangya 2 (M2S/R2070) performed wellin the yield trial and in farmers’ fields and bothwere registered in Zhejiang Province.
To evaluate indica/tropical japonica hybrids,• Make several crosses between TGMS lines
(WC) and tropical japonica varieties. If sometropical japonicas have the WC gene, theycan be crossed to any indica TGMS line.
• Evaluate the hybrids in the OYT and a se-ries of trials to identify the most promisinghybrids with enhanced heterosis. The besthybrids that are grown in the locality shouldbe used as standard checks. Emphasis shouldbe given to monitoring spikelet fertilityduring the evaluation.
Developing tropical japonica pollen parents,the same method described for transferring thedesirable tropical japonica pollen parent can befollowed. However, the recipient tropical japonicashould possess a WC gene.
When developing desirable indica pollenparents possessing wide compatibility, the fre-
quency of elite indica lines is quite high. Hence,these can be good male parents developing in-dica/tropical japonica hybrids, if only WC genesare transferred into them. The procedure for this isas follows:
• Select indica lines with very good restoringability and WC donors with a marker gene(a purple apiculus or Amp32).
• Make crosses between the indica line andWC donors.
• Grow F1 and evaluate all crosses for spikelet
fertility. Choose the highly fertile cross forfurther selection.
• Grow F2 and select good recombinants look-
ing like the restorer but with the apiculuspigmentation or Amp32 allele.
• Grow F3 and F
4 and select the best plants in
the best families. Keep track of the originalplant type of the pollen parent and the Amp32
allele.• In the F
5, test-cross the selected lines on a
single-plant basis with a tropical japonicaEGMS line having no WC gene.
• Evaluate the testcross progenies for fertil-ity. Select lines that show high fertility. Theyare the pollen parents possessing WC genes.
Once the requisite pollen parents are devel-oped, they can be test-crossed with the best avail-able female parents and the experimental hybridscan be evaluated as discussed in Chapter 9 on“Evaluating two-line hybrids.”
Table 14. Wide-compatibility varieties in differentvarietal groups.
Indica Japonica Tropicaljaponica
BPI 76 NK 4 BantenDular (aus) Norin PL9 CalotocN22 (aus) 02428 CP-SLO
Ketan NangkaMoroberekanPalawanPadi BujangPendecIR64446-7-3-2-2IR65598-112-2

41
CHAPTER 8
Combining ability nursery
Assessing the combining ability of parental linesis extremely useful in a hybrid breeding program,especially when many prospective parental linesare available and the most promising ones are tobe identified on the basis of their ability to givesuperior hybrids. The line × tester method(Kempthorne 1957) is commonly used for thispurpose.
Definitions• Combining ability refers to the ability of a
genotype to transfer its desirable traits to itsprogenies.
• General combining ability (GCA) is the av-erage performance of a parent in a series ofcrosses.
• Specific combining ability (SCA) is the de-viation in the performance of a hybrid fromthe performance predicted on the basis ofthe general combining ability of its parents.
Type of lines to be evaluated• The most stable CMS and EGMS lines pos-
sessing high phenotypic acceptability anda fair to excellent outcrossing rate.
• Effective restorers/pollen parents adapted tothe target area.
Procedure using theline × tester design
• Let us suppose we have “l” lines (elite pol-len parents) and “t” testers (elite CMS and/or EGMS lines).
• All the l lines should be crossed to each ofthe t testers to produce l × t experimentalhybrids.
Composition of the combining abilitynursery
• All the l × t hybrids along with the parents(lines + testers).
• Suitable check varieties may also be in-cluded for working out standard heterosis.
Field layout• Choose a fairly homogeneous plot for grow-
ing the combining ability nursery in a repli-cated trial using a randomized completeblock design (RCBD).
• Use several replications to ensure a mini-mum of 12 degrees of freedom for error tohave statistically valid comparisons.
• Plant a single seedling per hill with a spac-ing of 20 ×15 or 20 × 20 cm.
• Plot size may depend on the amount of F1
seed available. However, a minimum of 50plants per plot is essential. The larger theplot size, the better it is for evaluation.
• Avoid collecting data from border plants.Each three-row plot of hybrids can be flankedby a border row of a check variety.
Statistical analysis• If we have five lines (pollen parent lines)
and four testers (EGMS lines), the total num-ber of crosses will be 1 × t = 5 × 4 = 20.

42
• Let these 20 crosses along with five linesand four testers (29 entries) be tested in anRCBD with three replications with onecheck variety (Fig. 17).
Analysis of variance (Grand total)2
Correction factor = (CF) Total no. of observations
Total sum of squares (TSS) = Σ Yij2 – CF
Σ Y.J2
Replication SS (RSS) = – CF t Σ Yi2
Replication SS (RSS) = – CF r
Error SS (ErSS) = TSS – TrSS – RSS
Analysis of variance tablea
Source df SS MSS F
Replications (r – l) [2] RSS/2Treatments (t – l) [28] TrMSS/28Error (r – l)(t – l) [56] ErMSS/56Total (rt – l) [86]
adf = degrees of freedom, SS = sum of squares,MSS = mean sum of squares, F = F value.
To test the significance of the genotypic dif-ference, compare the calculated F (TrMSS/ErMSS)
with the table value of F for 28 and 56 degrees offreedom at the 5% or 1% level of significance.
Treatment SS can be further partitioned intoSS from parents, SS from crosses, and SS from theinteraction of parents vs crosses.
Σ C2ij + Σ P2iiTreatment SS = – CF
r
Cij = observation for ijth crossPii = observation for ith parent r = number of replications
SS from Σ C2ijcrosses = – CF (crosses with 19 df)
r
Σ P2iiSS from parents = – CF (parents
r with 8 df)
SS from interaction = TrSS – SS (crosses) –of parents vs crosses SS (parents) (with 1
df)
ANOVA with parents and crosses
Source df SS MSS F
Replications 2Treatments 28Crosses 19Parents 8Parents vs crosses 1Error 56
Test all sources of variation against error vari-ance.
Fig. 17. Layout of combining ability trial.
Rep 1 Rep 2 Rep 3
5 25 15 18 13 24 C 4 21 6 25 17 27 14 716 9 11 2 28 19 14 23 13 26 19 10 1 20 1212 21 1 17 8 5 28 16 9 17 21 29 3 16 2
3 7 23 4 27 29 7 3 11 10 26 6 23 8 C10 20 C 26 29 12 1 18 22 20 29 13 5 24 2214 22 6 24 19 27 25 15 8 2 9 18 15 11 4
1–20 hybrids, 21–25 lines, and 26–29 testers. C = check variety.

43
Line ××××× tester analysis
Construct a two-way table.
Testers Lines
1 2 3 4 Total
1 Yij Yi..2345Total Y.J. Y..
Σ Yi2..SS from lines = – CF (crosses)
r × t
where r = replications and t = testers.
Σ Y.J.2
SS from testers = – CF (crosses) 1 × r
SS from lines × testers = SS (crosses) – SS (lines)– SS (testers)
ANOVA for line ××××× tester analysis
Source df SS MSS F
Lines 4Testers 3Lines × testers 12Error 56
ANOVA for line ××××× tester analysis includingparents
Source df SS MSS F
Replications 2Treatments 28Parents 8Parents vs crosses 1Crosses 19Lines 4Testers 3Lines × testers 12Error 56Total 86
Note: MSS from lines and MSS from testers are tobe tested against MSS from lines × testers. MSSfrom lines × testers is to be tested against MSSfrom error.
Sometimes line × tester analysis is done byusing cross means (means of crosses over replica-tions). In that case, MS from error that is used fortesting the significance of MS (lines × tester) shouldbe divided by the number of replications beforetesting.
Estimation of GCA effects:
i) GCA effects of lines
Yi.. Y..gi = –
tr ltr
where Yi.. = total of ith line over testers, Y.. =grand total, and ltr = number of lines, testers, andreplications.
Work out GCA effects for g1 to g
5. See whether
Σ gi = 0.
ii) GCA effects of testers
Σ Y.j. Y..gt = –
lr ltr
where Y.j. = total of jth tester over lines, Y.. =grand total, and ltr = number of lines, testers, andreplications.
Work out GCA effects for g5 to g
9. See whether
Σ gt = 0.
iii) Estimation of SCA effects
Yij. Y.. Y.j. Y..Sij = – – +
r rt rl ltr
where Yij. = value of jth line with ith tester, Yi.. =total of ith line over all testers, Y.j. = total of jthtester over all lines, Y.. = grand total, and ltr =number of lines, testers, and replications.
Work out SCA effects for all hybrids. Seewhether Σ
i Σ
j S
ij = 0.

44
Testing the significance of combining abilityeffects
SE (standard error) (GCA for lines) =
SE (standard error) (GCA for testers) =
SE (standard error) (SCA effects) =
SE (standard error) (gi – gj) line =
SE (standard error) (gi – gj) testers =
SE (standard error) (Sij – Skl) =
Me is the error mean sum of squares.
Interpretation of results• The statistical significance of treatments
indicates that the entries have genotypic dif-ferences between them. If the treatment dif-ferences are significant, we can use furtherpartitioning.
2Me r
Me rt
Me rl
Me r
2Me rt
2Me rl
• Partitioning of treatment SS into SS fromcrosses and parents helps to test the signifi-cance of these two components individu-ally.
• The parents with higher positive significantGCA effects are considered to be good gen-eral combiners, whereas those with negativeGCA effects are considered to be poor gen-eral combiners.
• The hybrids with significant SCA effects ina positive direction are considered to be themost promising ones.
Using the results• The EGMS lines with good GCA are chosen
for developing experimental hybrids for test-ing in observation yield trials.
• The pollen parents with good GCA will beused for crossing with EGMS lines to pro-duce experimental hybrids for testing inobservation yield trials.
• Hybrids with higher positive significantSCA effects are chosen for evaluation in pre-liminary yield trials.

45
CHAPTER 9
Evaluating two-linehybrids
Many hybrids developed through the two-lineapproach using EGMS lines are heterotic. There-fore, an elaborate evaluation of the hybrids is re-quired to identify the most promising ones in termsof yield heterosis per se and their suitability forhigh seed yields and self seed yields of the femaleparent during its multiplication. Experimentalhybrids should be evaluated in a series of trials,using an appropriate statistical design to ensureunbiased comparisons. The choice of design de-pends on the number of entries and the quantityof hybrid seed available. During the initial stages,the number of entries to be tested is large and thequantity of hybrid seed available is limited; there-fore, the hybrids are tested in unreplicated trials.However, in subsequent evaluation, hybrids needto be tested in replicated trials with a larger plotsize. The performance of hybrids may be location-specific. Therefore, it is necessary to conductmultilocation trials to identify hybrids havingwide adaptability and those that are specificallyadapted to certain locations. Testing the perfor-mance of hybrids in farmers’ fields along with lo-cal check varieties of the region is necessary be-fore these hybrids are released for commercial cul-tivation.
Observation yield trial (OYT)CompositionRe-testcrosses are made between the commerciallyusable EGMS lines and effective restorers identi-fied in the testcross nursery. Use three to four checkvarieties representing different growth durations(such as very early, early, medium, and late).
Experimental design and field layout• Since the number of experimental rice hy-
brids is large (100–500) and the amount ofhybrid seed is limited, it is convenient toconduct the OYT by using the augmenteddesign.
• In this design, the whole experimental areais divided into several blocks.
• The check varieties are replicated in eachblock, whereas the test entries are not repli-cated but are assigned to the remaining plotsrandomly.
• The yields of test entries are adjusted forblock differences based on the yield of checkvarieties in each block.
• The block size is determined as follows:
If c = number of check varieties, v = numberof test hybrids, and b = number of blocks, the num-ber of test entries in a block (n) = v/b, the numberof plots/block (P) = c + n, and the total number ofplots (N) = b (c + n).
• The total number of blocks should ensure atleast 12 df for error in ANOVA.
12b > + 1 c – 1
• Let us take 40 hybrids and four check vari-eties.
12The number of blocks = + 1
4 – 1
12 = + 1 = 5
3
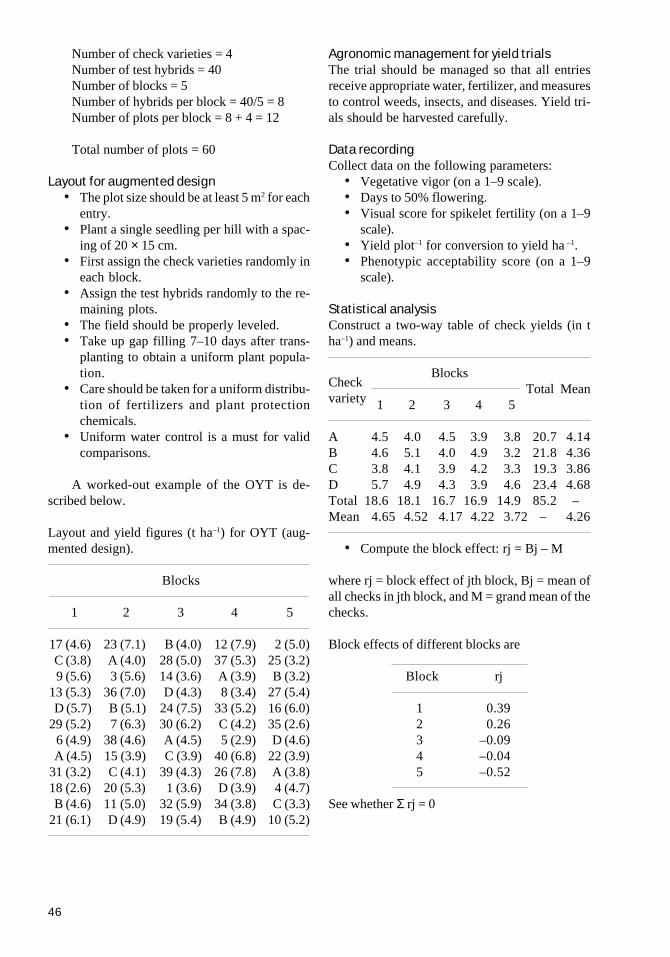
46
Number of check varieties = 4Number of test hybrids = 40Number of blocks = 5Number of hybrids per block = 40/5 = 8Number of plots per block = 8 + 4 = 12
Total number of plots = 60
Layout for augmented design• The plot size should be at least 5 m2 for each
entry.• Plant a single seedling per hill with a spac-
ing of 20 × 15 cm.• First assign the check varieties randomly in
each block.• Assign the test hybrids randomly to the re-
maining plots.• The field should be properly leveled.• Take up gap filling 7–10 days after trans-
planting to obtain a uniform plant popula-tion.
• Care should be taken for a uniform distribu-tion of fertilizers and plant protectionchemicals.
• Uniform water control is a must for validcomparisons.
A worked-out example of the OYT is de-scribed below.
Layout and yield figures (t ha–1) for OYT (aug-mented design).
Blocks
1 2 3 4 5
17 (4.6) 23 (7.1) B (4.0) 12 (7.9) 2 (5.0)C (3.8) A (4.0) 28 (5.0) 37 (5.3) 25 (3.2)9 (5.6) 3 (5.6) 14 (3.6) A (3.9) B (3.2)
13 (5.3) 36 (7.0) D (4.3) 8 (3.4) 27 (5.4)D (5.7) B (5.1) 24 (7.5) 33 (5.2) 16 (6.0)
29 (5.2) 7 (6.3) 30 (6.2) C (4.2) 35 (2.6)6 (4.9) 38 (4.6) A (4.5) 5 (2.9) D (4.6)A (4.5) 15 (3.9) C (3.9) 40 (6.8) 22 (3.9)
31 (3.2) C (4.1) 39 (4.3) 26 (7.8) A (3.8)18 (2.6) 20 (5.3) 1 (3.6) D (3.9) 4 (4.7)B (4.6) 11 (5.0) 32 (5.9) 34 (3.8) C (3.3)
21 (6.1) D (4.9) 19 (5.4) B (4.9) 10 (5.2)
Agronomic management for yield trialsThe trial should be managed so that all entriesreceive appropriate water, fertilizer, and measuresto control weeds, insects, and diseases. Yield tri-als should be harvested carefully.
Data recordingCollect data on the following parameters:
• Vegetative vigor (on a 1–9 scale).• Days to 50% flowering.• Visual score for spikelet fertility (on a 1–9
scale).• Yield plot–1 for conversion to yield ha –1.• Phenotypic acceptability score (on a 1–9
scale).
Statistical analysisConstruct a two-way table of check yields (in tha–1) and means.
BlocksCheck Total Meanvariety 1 2 3 4 5
A 4.5 4.0 4.5 3.9 3.8 20.7 4.14B 4.6 5.1 4.0 4.9 3.2 21.8 4.36C 3.8 4.1 3.9 4.2 3.3 19.3 3.86D 5.7 4.9 4.3 3.9 4.6 23.4 4.68Total 18.6 18.1 16.7 16.9 14.9 85.2 –Mean 4.65 4.52 4.17 4.22 3.72 – 4.26
• Compute the block effect: rj = Bj – M
where rj = block effect of jth block, Bj = mean ofall checks in jth block, and M = grand mean of thechecks.
Block effects of different blocks are
Block rj
1 0.392 0.263 –0.094 –0.045 –0.52
See whether Σ rj = 0
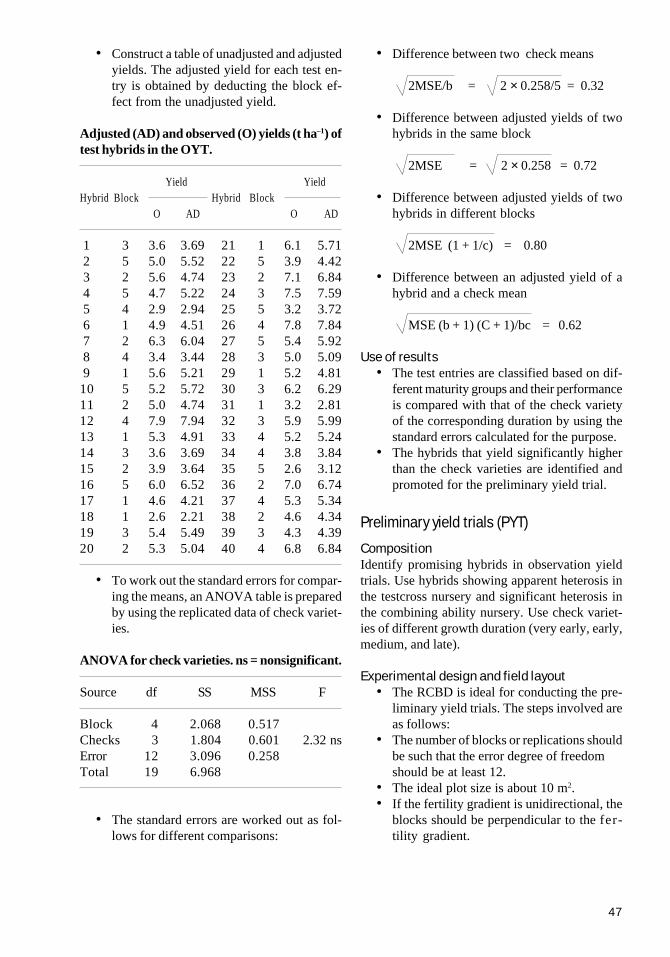
47
• Construct a table of unadjusted and adjustedyields. The adjusted yield for each test en-try is obtained by deducting the block ef-fect from the unadjusted yield.
Adjusted (AD) and observed (O) yields (t ha–1) oftest hybrids in the OYT.
Yield Yield Hybrid Block
O ADHybrid Block
O AD
1 3 3.6 3.69 21 1 6.1 5.71 2 5 5.0 5.52 22 5 3.9 4.42 3 2 5.6 4.74 23 2 7.1 6.84 4 5 4.7 5.22 24 3 7.5 7.59 5 4 2.9 2.94 25 5 3.2 3.72 6 1 4.9 4.51 26 4 7.8 7.84 7 2 6.3 6.04 27 5 5.4 5.92 8 4 3.4 3.44 28 3 5.0 5.09 9 1 5.6 5.21 29 1 5.2 4.8110 5 5.2 5.72 30 3 6.2 6.2911 2 5.0 4.74 31 1 3.2 2.8112 4 7.9 7.94 32 3 5.9 5.9913 1 5.3 4.91 33 4 5.2 5.2414 3 3.6 3.69 34 4 3.8 3.8415 2 3.9 3.64 35 5 2.6 3.1216 5 6.0 6.52 36 2 7.0 6.7417 1 4.6 4.21 37 4 5.3 5.3418 1 2.6 2.21 38 2 4.6 4.3419 3 5.4 5.49 39 3 4.3 4.3920 2 5.3 5.04 40 4 6.8 6.84
• To work out the standard errors for compar-ing the means, an ANOVA table is preparedby using the replicated data of check variet-ies.
ANOVA for check varieties. ns = nonsignificant.
Source df SS MSS F
Block 4 2.068 0.517Checks 3 1.804 0.601 2.32 nsError 12 3.096 0.258Total 19 6.968
• The standard errors are worked out as fol-lows for different comparisons:
• Difference between two check means
2MSE/b = 2 × 0.258/5 = 0.32
• Difference between adjusted yields of twohybrids in the same block
2MSE = 2 × 0.258 = 0.72
• Difference between adjusted yields of twohybrids in different blocks
2MSE (1 + 1/c) = 0.80
• Difference between an adjusted yield of ahybrid and a check mean
MSE (b + 1) (C + 1)/bc = 0.62
Use of results• The test entries are classified based on dif-
ferent maturity groups and their performanceis compared with that of the check varietyof the corresponding duration by using thestandard errors calculated for the purpose.
• The hybrids that yield significantly higherthan the check varieties are identified andpromoted for the preliminary yield trial.
Preliminary yield trials (PYT)CompositionIdentify promising hybrids in observation yieldtrials. Use hybrids showing apparent heterosis inthe testcross nursery and significant heterosis inthe combining ability nursery. Use check variet-ies of different growth duration (very early, early,medium, and late).
Experimental design and field layout• The RCBD is ideal for conducting the pre-
liminary yield trials. The steps involved areas follows:
• The number of blocks or replications shouldbe such that the error degree of freedomshould be at least 12.
• The ideal plot size is about 10 m2.• If the fertility gradient is unidirectional, the
blocks should be perpendicular to the fer-tility gradient.
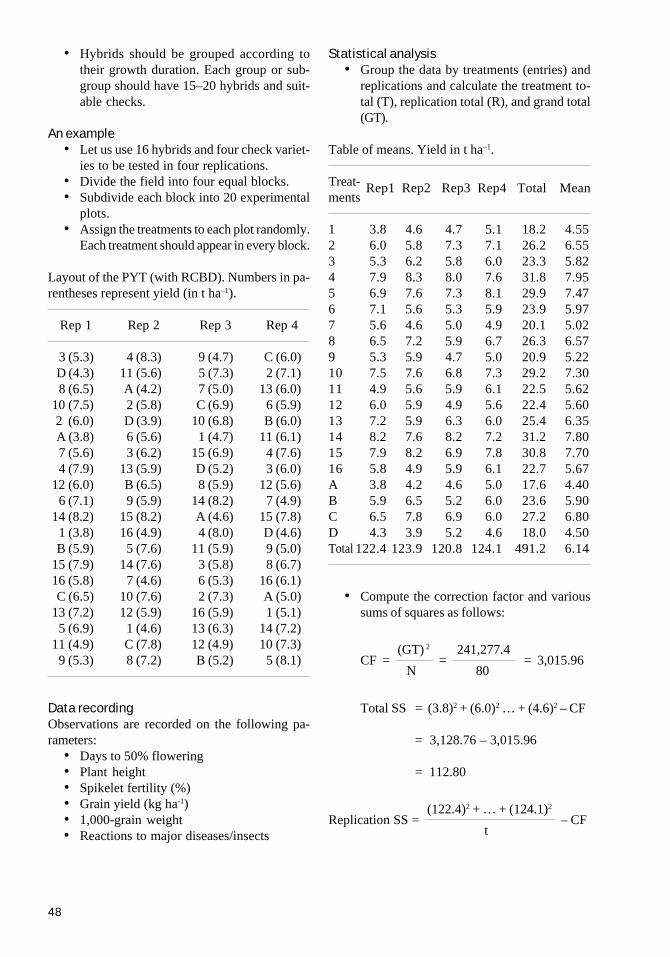
48
• Hybrids should be grouped according totheir growth duration. Each group or sub-group should have 15–20 hybrids and suit-able checks.
An example• Let us use 16 hybrids and four check variet-
ies to be tested in four replications.• Divide the field into four equal blocks.• Subdivide each block into 20 experimental
plots.• Assign the treatments to each plot randomly.
Each treatment should appear in every block.
Layout of the PYT (with RCBD). Numbers in pa-rentheses represent yield (in t ha–1).
Rep 1 Rep 2 Rep 3 Rep 4
3 (5.3) 4 (8.3) 9 (4.7) C (6.0) D (4.3) 11 (5.6) 5 (7.3) 2 (7.1) 8 (6.5) A (4.2) 7 (5.0) 13 (6.0)
10 (7.5) 2 (5.8) C (6.9) 6 (5.9) 2 (6.0) D (3.9) 10 (6.8) B (6.0) A (3.8) 6 (5.6) 1 (4.7) 11 (6.1) 7 (5.6) 3 (6.2) 15 (6.9) 4 (7.6) 4 (7.9) 13 (5.9) D (5.2) 3 (6.0)
12 (6.0) B (6.5) 8 (5.9) 12 (5.6) 6 (7.1) 9 (5.9) 14 (8.2) 7 (4.9)
14 (8.2) 15 (8.2) A (4.6) 15 (7.8) 1 (3.8) 16 (4.9) 4 (8.0) D (4.6) B (5.9) 5 (7.6) 11 (5.9) 9 (5.0)15 (7.9) 14 (7.6) 3 (5.8) 8 (6.7)16 (5.8) 7 (4.6) 6 (5.3) 16 (6.1) C (6.5) 10 (7.6) 2 (7.3) A (5.0)13 (7.2) 12 (5.9) 16 (5.9) 1 (5.1) 5 (6.9) 1 (4.6) 13 (6.3) 14 (7.2)11 (4.9) C (7.8) 12 (4.9) 10 (7.3) 9 (5.3) 8 (7.2) B (5.2) 5 (8.1)
Data recordingObservations are recorded on the following pa-rameters:
• Days to 50% flowering• Plant height• Spikelet fertility (%)• Grain yield (kg ha-1)• 1,000-grain weight• Reactions to major diseases/insects
Statistical analysis• Group the data by treatments (entries) and
replications and calculate the treatment to-tal (T), replication total (R), and grand total(GT).
Table of means. Yield in t ha–1.
Treat- Rep1 Rep2 Rep3 Rep4 Total Meanments
1 3.8 4.6 4.7 5.1 18.2 4.552 6.0 5.8 7.3 7.1 26.2 6.553 5.3 6.2 5.8 6.0 23.3 5.824 7.9 8.3 8.0 7.6 31.8 7.955 6.9 7.6 7.3 8.1 29.9 7.476 7.1 5.6 5.3 5.9 23.9 5.977 5.6 4.6 5.0 4.9 20.1 5.028 6.5 7.2 5.9 6.7 26.3 6.579 5.3 5.9 4.7 5.0 20.9 5.2210 7.5 7.6 6.8 7.3 29.2 7.3011 4.9 5.6 5.9 6.1 22.5 5.6212 6.0 5.9 4.9 5.6 22.4 5.6013 7.2 5.9 6.3 6.0 25.4 6.3514 8.2 7.6 8.2 7.2 31.2 7.8015 7.9 8.2 6.9 7.8 30.8 7.7016 5.8 4.9 5.9 6.1 22.7 5.67A 3.8 4.2 4.6 5.0 17.6 4.40B 5.9 6.5 5.2 6.0 23.6 5.90C 6.5 7.8 6.9 6.0 27.2 6.80D 4.3 3.9 5.2 4.6 18.0 4.50Total 122.4 123.9 120.8 124.1 491.2 6.14
• Compute the correction factor and varioussums of squares as follows:
(GT) 2 241,277.4CF = = = 3,015.96
N 80
Total SS = (3.8)2 + (6.0)2 … + (4.6)2 – CF
= 3,128.76 – 3,015.96
= 112.80
(122.4)2 + … + (124.1)2
Replication SS = – CF t

49
= 3,016.32 - 3,015.96
= 0.36
(18.2)2 + (26.2)2 + … (18.0)2
Treatment = – CFSS r
= 3,110.82 – CF
= 94.86
Error SS = Total SS – RSS – TrSS
= 112.80 – 0.38 – 94.86
= 17.58
• Compute the mean sum of squares by divid-ing each sum of squares by its correspond-ing degree of freedom.
RSS 0.36Replication MSS = = = 0.12 r – 1 3
Tr.SS 94.86Treatment MS = = = 4.99
t – 1 19
Er.SS 17.58Error MS = = = 0.30
(r – 1)(t – 1) 57
• Compute the F value for testing the treat-ment differences.
Treatment MS 4.99F value = =
Error MS 0.30
= 16.6
• Compare the calculated F value with thetable F value.
• Prepare the analysis of variance table byincluding all the computed values.
Analysis of variance (ANOVA) table.
Computed Table FSource DF SS MSS Fa
5% 1%
Replication 3 0.36 0.12Treatment 19 94.86 4.99 16.6**Error 57 17.58 0.30Total 79 112.80
a**A highly significant F value indicates that the test en-tries differ significantly among themselves.
• Compute the coefficient of variation (CV)
Error MSCV = × 100
GM1
0.30= × 100 6.14
= 8.92
• Compute the critical difference (CD)
2 × EMSCD = t 0.05 ×
r
= 3.44 × 0.387
= 1.33
• The hybrids with a difference of more thanthe CD value from the check variety are con-sidered significantly superior to the checkvariety.
Use of results• The performance of the hybrids is compared
with that of the check variety of correspond-ing duration or the highest-yielding checkvariety.
1GM = grand mean.

50
• The hybrids that have a significantly higheryield than the check variety are identifiedand promoted to the advanced yield trial. Asignificant yield advantage of more than 1 tha–1 would also be the ideal criterion for se-lecting the best hybrids after testing the sig-nificance.
Advanced yield trials (AYT)CompositionIdentifying promising hybrids in the preliminaryyield trials. Use three or four check varieties ofdifferent duration (very early, early, medium, andlate).
Experimental design and field layout• The RCBD is ideal for conducting the AYT.• The number of entries in the AYT is much
lower than in the PYT. It is helpful to in-crease the plot size to 15 m2.
• Entries should be divided into at least twomaturity groups (1—very early and early;2—medium and late).
• The field layout and agronomic manage-ment are similar to those for the PYT.
• Data recording is essential.
The following observations are recorded forthe AYT:
• Plant height• Days to 50% flowering• Panicles m–2
• Number of filled grains panicle–1
• Spikelet fertility (%)• Yield ha–1
• 1,000-grain weight• Reactions to major pests and diseases• Remarks on special features• Statistical analysis
The method of statistical analysis is the same asthe one explained for the preliminary yield trials.
Use of results• The performance of the hybrids is compared
with that of the check variety of correspond-ing duration or the highest-yielding checkvariety.
• The hybrids that have significantly higheryield (>1 t ha–1) than the check variety arepromoted for multilocation trials.
• Mere statistical significance is not sufficientto consider a hybrid as promising. There-fore, an advantage of about 1 t ha–1 over thecheck variety is specified, which would re-sult in a real benefit to farmers.
Multilocation yield trials (MLT)The major objective of multilocation yield trialsis to identify the hybrids that have a wider adapt-ability or those that are specifically adapted to aparticular location. This exercise is essential ashybrids perform differently in different environ-ments. This also provides an opportunity for breed-ers to see the performance of hybrids bred by themin other locations, even though these hybrids mayfail to perform well in the location where they aredeveloped. The concept of multilocation yieldtrials has really improved the efficiency of ricebreeders and this is more so in hybrid rice breed-ing.
CompositionIdentify the promising hybrids in the AYT fromdifferent centers, including those introduced fromabroad. Use three or four check varieties of differ-ent duration (very early, early, medium, and late).If the trials are constituted based on duration, itwould suffice to include a check variety of corre-sponding duration in the trial.
Experimental design and field layout• The locations for MLT should be selected
carefully so that each location serves as adistinct environment. The location selectedshould be in the proposed target area for thecultivation of hybrid rice.
• An RCBD is most commonly used for con-ducting MLT.
• It is necessary to have common guidelinesfor agronomic management and data col-lection from different centers.
General guidelines for conducting MLT• Specify the design to be adapted—an RCBD
with four replications.• Specify the entries and the check varieties.
Besides the common check, each center canchoose a local check for comparison.
• The trial should be conducted during thesame season at all locations.

51
• Specify the seedling age at transplanting—21–25 days old.
• The spacing adopted should be uniform—20 × 20 or 20 × 15 cm.
• Specify planting a single seedling per hill.• The fertilizer dose may depend on the na-
tive soil fertility and recommendations inthe local area.
• Plant protection should be need-based.• The plot size should be uniform in all the
locations to the extent possible (at least 15m2).
Agronomic managementAgronomic management should be uniform in alllocations so as to have valid comparisons, exceptfor some specific recommendations made for aparticular location.
Data recordingData sheets are circulated to all the cooperatorsfor collecting data on important parameters, suchas
• Plant height• Days to 50% flowering• Panicles m–2
• Number of filled grains panicle–1
• Spikelet fertility (%)• Yield plot–1
• Yield ha–1
• 1,000-grain weight• Reactions to pests and diseases• Weather data of each location
The statistical analysis for G × E interactionsand interpretation of results are covered in thechapter on G × E analysis.
Use of results• The hybrids with higher yield potential and
wider adaptability are identified based onstability analysis. These are promoted foron-farm testing in different areas, before theirrelease for commercial cultivation.
• Those hybrids that are found to be suitablefor a particular location are promoted foron-farm testing in that particular regiononly.

53
CHAPTER 10
Two-line hybrid riceseed production
The production of two-line rice hybrids involvestwo major steps: (1) multiplication of EGMS linesand (2) hybrid rice seed production.
Each of these steps requires specific environ-mental conditions for seed production. Therefore,the locations/seasons for producing the two typesof seed have to be different. In Chapter 5, we sug-gested that suitable specific locations must beidentified for elite EGMS lines for their self seedmultiplication. Locations must also be identifiedthat are suitable for hybrid seed production usingthe same EGMS lines.
Multiplication of EGMS linesEGMS lines, if multiplied continuously for sev-eral generations without any selection, may seg-regate for CSP, thereby causing major problemsin maintaining purity of the hybrid seeds. There-fore, nucleus and breeder seed production mustbe taken up on a continual basis.
Method I• Nucleus seed production of an EGMS
(TGMS or PGMS) line begins in the fertil-ity-inducing environment. Seeding ofTGMS or PGMS lines is arranged in such away that the sensitive stage occurs when thetemperature or photoperiod is favorable fora higher seed set.
• At the time of flowering, about 100 typicalplants are selected from the population ofan EGMS (TGMS or PGMS) line and theirpanicles are bagged. The selection processshould be completed within 1 week.
• After the harvest, the selected plants arescored for spikelet fertility (based on the
main panicle) and 50 plants with higherspikelet fertility (above 30%) are selected.
• Progenies of the selected plants are grownin the sterility-inducing environment. About30 seeds are taken from each of the selectedplants to grow single-row progenies and theremaining seeds are stored carefully. Thebalance of the seeds of the progenies thatare uniform and completely male sterilemust be marked and bulked to form thenucleus seed (Fig. 18).
• Nucleus seed of the EGMS line is used forproducing breeder seed under strict isola-tion. Breeder seed for the EGMS line is pro-duced in the fertility-inducing environment.
• The breeder seed produced under the directsupervision of the plant breeder has highgenetic purity and is used for producingfoundation seed of parental lines, which inturn will be used for producing hybrid seed.
Method II• Select a completely male sterile plant with
typical characteristics of the original EGMSline under a sterility-inducing environment.
• Ratoon the selected plant and clone it for asmany plants as you need. Multiply the ra-tooned stubbles under a fertility-inducingenvironment. The nucleus seed will be har-vested from the ratooned stubbles.
• The nucleus seed is used for producingbreeder seed and the latter for producingfoundation seed.
• Preserve the selected stubbles under favor-able temperature conditions with good man-agement as long as you need them. The newnucleus seed will be produced continuously.

54
• With this system of multiplication, theEGMS line continues in the same genera-tion and it will not segregate for its CSP. Itsgeneral combining ability will also notchange.
In the tropics, since ratooning may not al-ways be successful because of the high incidenceof diseases and insects, method II can be risky.
• Foundation seed production. Fresh breederseed should be used by seed product ionagencies to produce foundation seed ofTGMS/PGMS lines in a strict isolation areawith suitable temperature and/or daylengthconditions.
Differences between EGMSand CMS line multiplication
• The fertility of the CMS line is controlledby the CMS gene and a pair of recessivenuclear fertility restorer genes. Expressionof male sterility of CMS lines is stable overenvironments. The fertility of EGMS linesis controlled by a recessive nuclear genealone, the expression of which is influencedby environmental conditions.
• The maintenance of the CMS line is throughits cross pollination with the maintainer line.The yield of CMS seeds through this pro-cess largely depends on outcrossing and
�������������
��� �������������������������
�������������������
��� ������������
������������
������������������������������������������������������������ ������������������������� ����������������� ��������������� �������!���� ���������������������� ��������������������� �������"�����������������������������������
���"�������������������"��������������������������������� ���������������������#�����������������������
�������� ���������� ��������!�������������������������������$���� ������������������"���������� ������������������������������������
���������������������������������#�����������������������������������������
�����������������������������������������������������������������
%�����������������&�������������������������������������������������������������� ��������������������
�����
�����
�����
�����
�����
�����
Fig. 18. Procedure for nucleus and breeder seed production of TGMS lines(Virmani et al 1997a).

55
other related traits of the CMS line. Themaintenance of the EGMS lines, in contrast,is quite simple since it is through selfingunder fertility-inducing environments, es-pecially during the sensitive panicle devel-opment phase (stage II to stage VI). The seedyield of EGMS lines largely depends uponthe CFP and the favorable environment dur-ing fertility alteration.
• The CMS line is purified through pair cross-ing (A × B) in which the A and B lines areselected based on their morphological char-acteristics. The purity of the EGMS line ismaintained by evaluating the morphologi-cal characters and the CFP under specificlocations.
• The yield of CMS line multiplication isabout 2 t ha–1 under suitable conditions andit is unstable because of the changingweather conditions during the floweringperiod. Under favorable conditions, theyield of EGMS line multiplication couldreach 4–6 t ha–1. This yield is higher be-cause it occurs due to self-pollination com-pared to seed yield in the CMS line, whichoccurs due to cross-pollination.
Similarity of CMS and EGMSline multiplication
• CMS line multiplication (A × B) requiresstrict isolation to prevent contamination bystray pollen. Under favorable environmen-tal conditions, the EGMS line becomes fer-tile and seed setting can be as high as 75%;however, some male sterile spikelets can stillreceive pollen from another source. There-fore, strict isolation must also be providedto multiply pure EGMS lines.
• Strict roguing is necessary for CMS andEGMS line multiplication.
High-yielding techniques for PGMS linemultiplication (Chinese experience)Autumn-season multiplicationShort daylength can induce PGMS lines to be-come fertile under proper temperature conditions.In autumn, the daylength is getting shorter and ahigh percentage of seed setting can be achieved ifthe sensitive stage occurs under this condition.
However, seed quality becomes inferior, especiallywhen the temperature falls sharply in late autumn.
Winter-season multiplicationin lower-latitude areasFor PGMS self seed multiplication, shortdaylength and low temperature in lower-latitudeareas in the winter season are the most suitableenvironmental conditions. Seed quality is betterthan in late autumn because the temperature gradu-ally rises at the time of seed maturity.
High-yielding techniques for TGMS linemultiplication (Chinese experience)Spring-season multiplicationThe fertility of TGMS lines is mainly influencedby temperature. TGMS lines should be sown inearly spring so that the sensitive stage matchesthe temperature for fertility induction based onlocal meteorological data. The yield may not bestable because of the occurrence of abnormallyhigh temperature, and the long growing periodmay result in nonmatching of the sensitive stagewith the proper temperature regime.
Autumn-season multiplicationThe temperature is low enough for fertility induc-tion in the autumn season. The TGMS lines shouldbe sown at the proper time to make the sensitivestage coincide with the low-temperature period inorder to obtain higher yields. Seed quality may beinfluenced by low temperature in the late matu-rity period.
High-altitude multiplicationIn high-altitude areas (with altitudes from 800 to1,000 m), moderate temperature and longerdaylength conditions are suitable for the shortgrowing period for PGMS and TGMS line self seedmultiplication.
Chilled-water irrigationResults showed that chilled-water (>17 ºC) irriga-tion can induce TGMS lines to produce fertilepollen similar to ambient low temperatures. Theadvantage of chilled-water irrigation is that it pro-vides flexibility for adjusting the sensitive stage.The TGMS self seed production is high and morestable since the water temperature is relativelystable and controllable.

56
Seed plot selection. A sufficient water supplywith suitable temperature is essential to the estab-lishment of a seed plot for TGMS self seed multi-plication. The water temperature should be higherthan 17–18 ºC but lower than the CFP of the TGMSline to be multiplied.
Season selection. The joint effect of lowertemperature and chilled-water irrigation is goodfor increasing EGMS self seed yield.
Irrigation time. Chilled-water irrigation startsfrom the young panicle differentiation of the sta-men and pistil primordial (stage IV) and goes tothe meiotic division of the pollen mother cell(stage VI).
Three-line hybrid rice seed productionThe three-line hybrid rice seed production systeminvolving CMS lines is a relatively stable methodacross normal rice-growing conditions, whereasthe two-line hybrid rice seed production systeminvolving EGMS lines has environmental limita-tions requiring strict adherence to the proper tim-ing according to season and location. For success-ful hybrid rice seed production, the male sterilitytrait of the female parent and effective pollen loadof the male parent with proper flowering synchro-nization are the key factors. For three-line seedproduction, only the flowering synchronizationbetween the A and R lines is important for higherhybrid seed yields, whereas, for two-line seed pro-duction, both the male sterility expression underthe sterile phase and the flowering synchrony be-tween the EGMS and pollen parent influence seedpurity and bring about higher hybrid seed yields.
Two safe-period determinationsfor hybrid seed production
• In three-line hybrid rice seed production,favorable climatic conditions for pollina-tion are called the safe flowering period,which is important for increasing the out-crossing rate. In two-line seed production,the two safe-periods refers to the conduciveenvironmental conditions that support firstthe induction of complete male sterility andsecond the facilitation of proper pollenmovement from the pollen parent and fer-tilization of the EGMS parent.
• Therefore, the first safe-period determinesseed purity, while the second safe-period de-termines hybrid seed yield.
• The first safe-period, the sensitive stage offertility alteration, must be given priority ifthere is a need for an adjustment accordingto the prevailing environment at a givenlocation.
The desirable climatic conditions for polli-nation are as follows: (1) temperature of 23–35 ºC(minimum-maximum) in a day, (2) relative humid-ity around 70–90%, and (3) no continuous rains(that last more than 3 days) during the pollinationperiod.
Isolation of the hybridseed production plot
• Terrain isolation. The selected seed plot iseither isolated by mountains or hills or byother natural barriers.
• Time isolation. The hybrid seed plot shouldflower 20–30 days earlier or later than anyother plot that could be a source of straypollen for the seed production plot.
• Distance isolation. Keep a distance of 200m between the seed plot and any other ricepollen source.
Determining the seedinginterval for synchronization
• Time method. The seeding interval is deter-mined by the difference in growth durationbetween the two parental lines. The one withlonger duration must be sown early accord-ing to the number of days of difference be-tween the two parents in terms of days to50% flowering.
• Leaf number method. The total leaf numberof a variety is relatively stable at the samesite and in the same season in different years.The rate of growth in terms of leaf number isinfluenced by environmental temperature.By observing the leaf number of the earlyseeded parental line, you could determinethe seeding date of the later one because theleaf number difference is rather stable be-tween the two parents.
• Heading date prediction• Remaining leaf number method. The
young panicle differentiation starts fromthe reciprocal third-leaf emergence(keeping the flag leaf as the first leaf andthe leaf before the flag leaf as the 2ndleaf and the leaf before the 2nd leaf asthe 3rd leaf) and the time taken for com-

57
plete panicle differentiation is about 30days from the start to heading. Thismeans that the parental line would headafter 30 days, when the reciprocal thirdleaf emerges.
• Stripping the young panicle. The youngpanicle differentiation could be dividedmorphologically into eight stages andeach stage takes about 3–4 days for de-veloping (Table 15, Fig. 19).
Production of hybrid seedsfor preliminary yield trialsFor preliminary yield trials, the hybrid seed re-quirement is usually small, but numerous hybridsare produced simultaneously.
Planting techniques. To produce a small quan-tity of hybrid seeds for the OYT or PYT, the fol-lowing are the planting designs: (1) chimney iso-lation, (2) modified chimney isolation, and (3) theisolation-free method.
i. Chimney isolation procedure• The desired parental lines are sown on dif-
ferent dates to obtain synchronous flower-ing.
• Twenty-five-day-old seedlings of EGMS(female parent) and pollen parent lines areplanted in alternate rows of five plants eachat a spacing of 15 ×15 cm (Fig. 20).
• Frames of 1 × 1 × 1 m are prepared witheither iron or aluminum angles.
• Cubicles of 1 × 1 × 1 m are stitched withmuslin cloth, with a flap at the top.
• The metal frame is placed around a 1-m2 area
where the EGMS (female parent) and pollenparent lines are planted just before flower-ing.
• The frame is covered with a muslin clothbag to prevent cross pollination.
• During the flowering period, the pollenplants are shaken to increase seed settingon the EGMS line. This can be facilitatedby opening the flap.
• Pollen parent plants are harvested first andthreshed separately. The EGMS line is har-vested and threshed later to avoid possibleseed admixture.
ii. Modified chimney isolation procedureThe chimney isolation method has been modifiedto overcome the problem of synchronization andto simplify the supplementary pollination. Thebasic layout is the same as that of the chimneymethod, except for the following differences:
• The EGMS and pollen parents are sown ondifferent days to achieve maximum flower-ing synchronization.
• At the boot-leaf stage of the parental lines,2-m-high barriers are erected to cover thethree sides of a 1-m2 plot, leaving a gap of20 cm from the ground. The open side iscovered by the barrier of the opposite plot.The space between the opposite plots is con-venient for cultural operations, includingsupplementary pollination.
• Supplementary pollination is done by us-ing sticks 3–4 times per day at peak anthe-sis during the flowering period of 7–10 days.
Table 15. Morphological stages in rice panicle development.
Morphological character Morpho-physiological stages of ricepanicle development (equivalent)
Invisible First bract primordium differentiation(stage I) and primary branchprimordium (stage II)
Little white hairs Secondary branch primordiumdifferentiation (stage III)
More large hairs Stamen and pistil primordiumdifferentiation (stage IV)
Can see the spikelet individually Mid meiotic stage (stage V)The lemma and palea are visible Late meiotic stage (stage VI)Spikelet has full size Pollen tetrad stage (stage IV)Panicle has green color Pollen maturation (stage VII)Panicle is enclosed in leaf sheath Heading stage (stage VIII, stage IX)
and is about to emerge

58
Fig. 19. Development stages of panicle formation to flowering.
Stage no. Development Approx. days Approx. paniclestage before heading length (mm)
I Panicle primordia 30 0.2II Primary branch primordium 27 0.4III Secondary branch primordium 24 1.5IV Stamen and pistil primordia 20 2V Pollen mother cells 17 10–25VI Meiotic division 12 80VII Mature pollen 6 190–250VIII Ripe stage of pollen 4 260XI Completed spikelets 1–2 270X Flowering –

59
Fig. 20. Position of EGMS line and pollen parent line in the chimney isolation procedure.
�!��
�!�
�#�
��
��� �
��
��� ���
#���
'�������� �������������
iii. Isolation-free methodAn isolation-free method developed at the Inter-national Rice Research Institute has been foundto be more practical and popular in tropical coun-tries. This method is ideal for producing smallquantities of hybrid seed required for the OYTand PYT.
• Selected pollen parent lines are grown sideby side in 5 × 3-m plots. In each pollen par-ent line plot, four rows of pollen parentplants are planted as border rows at 20 × 20-cm spacing to provide isolation from ad-joining plots. Four vacant spaces 40 cm inwidth are left in the middle of the plot, whichare interspersed by single rows of pollenparent line plants. About 68 EGMS plantscan be planted in these spaces at the time offlowering (Fig. 21).
• EGMS lines of experimental hybrids are stag-gered five times at 8–10-day intervals to en-sure a continuous supply of EGMS plants atthe flowering stage to synchronize the flow-
ering of pollen parents in different seed pro-duction plots.
• When primary tillers of EGMS and pollenparent lines are in the boot-leaf stage, theirflag leaves are clipped except for the twooutermost border rows of pollen parent lines,which act as a barrier for pollen from ad-joining plots.
• Three to five days after leaf clipping, theEGMS lines are uprooted (preferably in themorning, that is, 0600—0800) and areplanted in the vacant spaces of the plots.
• To enhance outcrossing, supplementarypollination is advocated at the peak anthe-sis period. Care should be taken to shakeonly those pollen parent lines that are flank-ing the EGMS lines.
• The pollen parent line is harvested first andthreshed separately, followed by the EGMSline bearing the hybrid seeds.
• By adopting this method, 3–5 g of hybridseed can be obtained from each EGMS/CMS

60
Fig. 21. Layout for isolation-free system for producing seeds of experimental two-line hybrids.
���������� �� ���� ���������� �� ����
���������� �� ���� ���������� �� ����
���������������
���������������
��������������#���
#��
�
plant. A plot with 15–40 EGMS plants canyield 50–200 g of hybrid seed, which willbe enough to conduct the OYT for two sea-sons (20 g per season) and replicated PYTsalso for two seasons (100 g per season).
Seed production for AYT and MLT• Strict isolation method. The hybrid seed
required for conducting the AYT should behighly pure. About 1–2 kg of seed is re-quired for this purpose. Therefore, the seedhas to be produced in a larger area (100–200-m2 plots) under strict isolation to en-
sure purity. The method is described below.• Isolation. A space isolation of 50 m is ideal
for hybrid seed production, which meansthat within this range no other rice varietiesshould be flowering except the pollen par-ent. If it is difficult to get space isolation, atime isolation of more than 21 days wouldserve the purpose. Distance isolation can bereduced to 30–40 m if the hybrid seed pro-duction plot is surrounded by an additional15–20 rows of pollen parents.
• Seeding sequence. Parental lines of hybridcombinations differ in their growth duration.

61
Therefore, they have to be seeded on differ-ent dates so that their flowering will be syn-chronous. A late parent is sown first and anearly parent is sown later, the difference be-ing equal to the difference in their growthduration. The EGMS line is seeded onlyonce so as to match the environmental con-ditions that favor complete male sterility.The pollen parent is seeded three times with3-day intervals, such that the difference be-tween the second sowing of the pollen par-ent and that of the EGMS line is equal to theseeding interval between the parental lines.
• Row ratio and layout. The optimum rowratio for hybrid seed production is 2–3males:8–10 females. Pollen parent seedlingsare evenly mixed and planted in three rows,at a spacing of 15 × 15 cm, leaving a spacefor an EGMS line in between. The EGMSseedlings are planted with a spacing of 30 ×15 cm. The spacing between the EGMS lineand the adjacent pollen parent line shouldbe 20 cm. Row direction should be perpen-dicular to the wind direction (Fig. 22).
• Roguing. Roguing is an important opera-tion in a hybrid seed production plot to en-sure purity of hybrid seeds. Rogues can beidentified as those that are out of their rowand early in booting, and based on othermorphological characters. The off-typesobserved during different growth stages areto be removed. Before flowering, roguing isessential, especially in experimental hybridseed production plots. Roguing at flower-ing is also extremely important as pollenfrom off-type plants can cause irreparabledamage through cross pollination with malesterile plants.
• GA3 spray. Spraying of GA
3 is recommended
to obtain good panicle exsertion. A dose of40–60 g ha–1 by a knapsack sprayer or 15–20 g ha–1 by a ULV sprayer is recommendedfor desired results. The spray liquid requiredis 500 L and 20 L for the knapsack and ULVsprayer, respectively. GA
3 should be sprayed
two times, the first when 15–20% of thetillers have started heading and the second2 days after the first spraying or when 35–40% of the panicles of the seed parent haveemerged.
• Supplementary pollination. At the time offlowering, supplementary pollination isdone by shaking the pollen parents with ei-ther a rope or bamboo sticks. This operationhas to be done 3–4 times daily at peak an-thesis for 6–10 days. The supplementarypollination technique using bamboo poleswill substantially increase the experimen-tal hybrid seed yield.
• Harvesting and threshing. Extreme careshould be taken while harvesting andthreshing the hybrid rice plots. Harvest andthresh the pollen parent first. Thoroughlycheck and remove any panicles of the pol-len parent separately. The seed should bedried, processed, bagged, and properly la-beled. The rogues must be removed beforethe flowering of the parental lines and be-fore harvest. The male rows or the pollenparent lines must be harvested carefully andthreshed separately. Before threshing, thethreshing machine must be properly cleanedto avoid seed admixture. The labels must becarefully placed on each bag of hybrid seedalong with the parentage, date of harvest,and field location.
Two-line hybrid rice seedproduction on a large scaleIsolation from a pollen source is equally impor-tant for large-scale seed production, especially inthe initial stages, for a newly released hybrid. To-tal space isolation is the best way for large-plotseed production. But, a minimum of 200 m dis-tance from any other rice pollen source is essen-tial for ensuring seed purity.
Sowing and transplantingFor proper flowering synchronization, the maleparental line should be raised on three differentsowing dates to match the estimated heading dateof the female line. Such a sowing plan will pro-vide a continuous supply of pollen during the flow-ering of the female (EGMS) parent (Fig. 23).
At the time of transplanting, the top one-thirdpart of the leaves may be chopped off from eachuprooted nursery bundle of the pollen parent forits easy identification. The pollen parent can betransplanted first in two rows with a space of 45
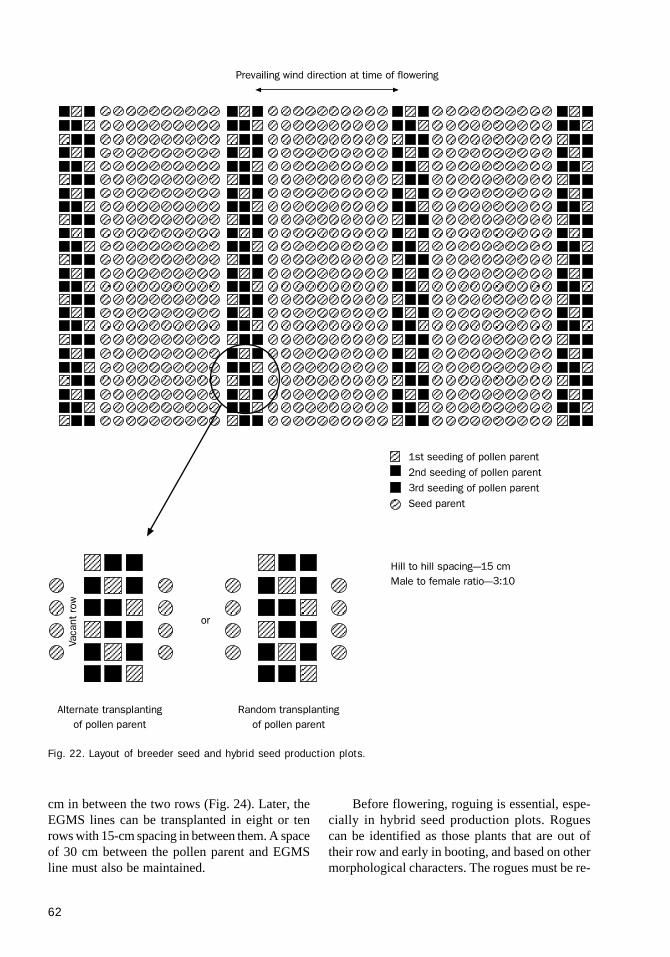
62
Fig. 22. Layout of breeder seed and hybrid seed production plots.
��������� ��������������������������������
��������� �����������������#������� �����������������&������� ���������������������������
(����������������� )��������������������)
%����������������� ����������������
*��������������������� ����������������
+��
����
��
��
�!��&,��
cm in between the two rows (Fig. 24). Later, theEGMS lines can be transplanted in eight or tenrows with 15-cm spacing in between them. A spaceof 30 cm between the pollen parent and EGMSline must also be maintained.
Before flowering, roguing is essential, espe-cially in hybrid seed production plots. Roguescan be identified as those plants that are out oftheir row and early in booting, and based on othermorphological characters. The rogues must be re-

63
moved before the flowering of the parental linesand before harvest.
Supplementary pollinationtechniquesSupplementary pollination techniques (i.e., push-ing the plant over to facilitate pollen movement)using rope or bamboo poles will substantially in-crease seed yield.
HarvestingThe male rows or the pollen parent lines must beharvested carefully and threshed separately. Be-fore threshing, the threshing machine must be prop-erly cleaned to avoid seed admixture. The labelsmust be carefully placed on each bag of hybridseed along with the parentage, date of harvest,and field location for further seed processing andpackaging for distribution to farmers.
Problems and their solutionsin hybrid seed productionMale sterility of EGMS lines, particularly TGMSand PTGMS lines, is highly influenced by tem-
perature rather than by photoperiod. Among theproblems, the most important is the adverse effectof temperature fluctuations caused by sudden/un-foreseen local weather changes.
During hybrid seed production at high-tem-perature locations, a sudden drop in temperature(below the CSP) can be disastrous because of re-version to the fertile phase resulting in selfing inthe female parent or the TGMS line. Hence, caremust be taken to use TGMS lines of low CSP andlow CFP.
Equally important is the identification of lo-cations with stable temperature based on severalyears of meteorological data.
Further, the high-temperature regime can beprolonged for a minimum of 4 weeks during thesensitive phase.
Likewise, higher temperature (above the CSP)during TGMS seed multiplication at low-tempera-ture locations can result in reduced percentageseed set, thus seriously affecting seed yield.
Aside from chemical remedies, the real solu-tion to these problems lies in developing stableTGMS lines adapted to reasonable fluctuations intemperature during hybrid seed production andTGMS seed multiplication.
Rather than using a strictly temperature-sen-sitive sterility system, it is desirable to use TGMSlines slightly influenced by photoperiod becausethey are flexible regarding temperature fluctua-tions and are more stable for fertility-sterility ex-pression.
15 cm
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P
E E E E E E E E E E P P E E E E E E E E E E P P15 cm 45 cm30 cm
Fig. 24. Field layout for two-line hybrid seed production. E = EGMS line and P =pollen parent line.
Pollen parent (3rd) ------------------------------------------
Pollen parent (2nd) -------------------------------------------
Pollen parent (1st) ---------------------------------------------
EGMS female parent flowering
Fig. 23. Proper flowering synchronization between pollenand seed parent in a hybrid seed production plot.

65
CHAPTER 11
Two-line rice hybrids:maintenance of geneticseed purity standards
Compared to three-line hybrids, the developmentof two-line hybrids was mainly limited by theirpurity. The purity of two-line rice is determinedby the uniformity of the fertility transformation ofEGMS lines, which is probably controlled by bothmajor and minor genes. These minor genes are notalways homozygous during the selection andevaluation of EGMS lines, which thus segregateduring seed reproduction, which leads to the dif-ference in the critical temperature of fertility al-teration among individual plants within lines.When the environmental temperature is above thecritical sterility point (CSP), the whole popula-tion of a TGMS line shows complete male steril-ity and looks uniform. However, once the tem-perature decreases, a difference among individualplants appears. Plants with a lower CSP are sterile,whereas plants with a higher CSP show partial fer-tility or even complete fertility. During hybridseed production, plants with a low CSP are sterileand produce hybrid seeds, but plants with a highCSP become fertile and produce self seed at lowtemperature. This causes a mixture of hybrid seedswith self seeds, which results in a nonuniform hy-brid rice production field. Therefore, purificationof EGMS lines is critical to the production of two-line hybrid seed. The following methods are rec-ommended for purification of EGMS lines:
1. Use nucleus seeds directly from the researchcenter for seed reproduction of EGMS lines.
2. Apply anther culture for the purification ofEGMS lines.
3. Introduce a recessive morphological markerinto the EGMS line or a dominant morpho-logical marker gene into the pollen parentline by gene transformation or conventional
backcross breeding methods. The pseudo-hybrids can be identified and eliminated inthe nursery, thereby increasing the purity ofhybrid rice in the field.
1. Using nucleus seeds of EGMSfor seed production
The nucleus seeds of EGMS lines produced mustbe used for only five generations since there canbe a gradual change in CSP levels.
The procedure for purifying EGMS lines is asfollows:
• Select about 100 plants with typical mor-phological characters of the original linefrom a population with less purity and plantthem in pots.
• Transfer the pots into a glasshouse or phy-totron with the controlled temperature ofCSP until heading, when the selected plantsdevelop into the secondary rachis–branchprimordial differentiation stage (stage III).
• Select the plants with 100% sterility. Whenthe plants begin heading, investigate pol-len and spikelet fertility under the micro-scope.
• Ratoon the selected plants and adjust thecontrolled-temperature to CFP till heading.The self-pollinated seeds harvested arecalled nucleus seeds.
• Plant nucleus seeds from each selected plantagain in rows. Compare their agronomictraits and fertility with those of the originalline and harvest the seeds from the rows thatare identical to the original line in bulk.

66
2. Using anther culture forpurifying EGMS lines
Applying anther culture to purify EGMS lines byproducing instant dihaploids that are completelyhomozygous for the EGMS trait can be highlyuseful for the proper maintenance of any givenEGMS line. Such an approach will be highly use-ful for hybrid seed production.
EGMS lines that are genetically homozygousand uniform in morphology have a better photo-and thermoperiod response to fertility alteration.Those showing a significant difference in fertilityamong individual plants are difficult to purify ef-fectively by conventional methods. Such linesshould undergo anther culture for purification.
Choose a suitable sowing date to makeEGMS plants head at the fertile phase. Alterna-tively, use the ratooned plants that passed theevaluation test in the growth chamber for purifi-cation. Make the ratooned EGMS plants tiller asmuch as possible to increase their panicle numberfor anther culture. Appendix II contains the de-tailed procedure for anther culture.
3. Transferring a recessivemarker gene into EGMS lines
A recessive morpho-logical marker is usedto distinguish thep s e u d o - h y b r i dplants. The markermust have (1) single-gene control, (2) aclearly visible pheno-type, (3) no obviousnegative effect onother useful traits, and(4) no influence onEGMS trait expres-sion.
The phenol reac-tion gene (ph gene) isone such marker thatcould be incorpo-rated into TGMSlines. Paddy grains ofvarieties possessingthis gene, whentreated with solutionsof phenolic com-
pounds (such as phenol, catechol, hydroquinone,pyrogallol, and tyrosine), become uniformlyblack. A monogenic recessive gene controls theexpression of this trait. If the pollen parent hasthis gene in the homozygous dominant form, theselfed seed of the TGMS line mixed with the hy-brid seed can be identified easily by the appear-ance of black grains after staining treatment. Thetrue hybrid F
1 seed material will appear brownish
white after staining. These two classes of grainscan then be separated easily by a color-sortingmachine (Virmani and Maruyama 1995). Thismethod can also be used by seed certifying agentsto determine seed purity percentage.
The backcross method is also used to transferthe recessive marker gene into EGMS lines. Theprocedure is illustrated in Figure 25 using the palegreen leaf marker gene (pgl) as an example.
4. Insertion of a dominant markergene into the pollen parent
A dominant morphological marker is used to dis-tinguish the pseudo-hybrid plants and it must havethe following characteristics: (1) be controlled bya single gene, (2) have a visible phenotype, (3)have no negative effect on any other useful trait,
March 1991 IGM 19 × 8902S (with pgl)
July 1991 F 1
Oct. 1991 8902S × F2 (plants with pale green leaves)
March 1992 B1F1 (out of 22 lines, 15 showed fertilitytransformation and M2S was the bestamong them)
Aug. 1992 B1F2 (36 individual sterile plants were selectedfrom 661 plants with pale green leaves)
March 1993 B1F3 (22 lines with uniform characters and betterfertility were selected from 36 lines)
Aug. 1993 B1F4 (22 lines were sown in intervals. After 1.5months, the best one—M2S—was selected)
M2S indica P(T)GMS line with pale greenleaves
Fig. 25. Procedure for transfer of recessive marker gene pgl into EGMS lines.

67
and (4) have no influence on the restoration abil-ity of the pollen parent line.
The protocol for transferring the herbicide(Basta) resistance gene (Bar) into a pollen parentusing genetic transformation and/or backcrossingis described below.
a) Screen the pollen parents used widely inthe commercial hybrids for their regenera-tion ability and those possessing good re-generation ability are selected.
b) For gene transformation, use mature seedsand immature embryos as explants. The Bargene is inserted by a particle gun orAgrobacterium-mediated transformation.
c) Transplant about 50 transgenic plants (T0)
into the soil using appropriate agronomicmanagement procedures.
d) Spray herbicide (Basta) on the regeneratedplants to kill the sensitive plants. Conductmolecular analysis (such as Southern blot-ting) of the resistant plants.
e) Harvest separately seeds (T1) from T
0 plants
that were identified as single-copy insertedtransgenic plants.
f) Sow T1 seeds on a line basis. The seedling
number of each line should be more than50. Meanwhile, sow the original pollen par-ent line as a control. In addition, EGMSlines should be sown timely using routineagronomic management practices.
g) Spray herbicide Basta on the T1 plants in
the nursery and select the surviving seed-lings from the lines that show a 3:1 segre-gation ratio for resistance to sensitivity.Then transplant them in the field as a singleplant per hill. The ideal transgenic plantswill be the ones that show no significantdifference from their original pollen parentin phenotype.
h) At the heading stage, cross the transgenicplants (pollen parent + Bar gene) with theEGMS line and their control (pollen parentline) with the EGMS line at the same timeand harvest their F
1 hybrid seeds.
i) Noncrossed panicles from the transgenicplants should also be bagged, then harvestself and crossed seeds (T
2) plant by plant.
j) Raise 24 F1 plants per replication, replicated
three times. Measure agronomic traits suchas plant height, tiller number, resistance todiseases and pests, and yield. The perfecttransgenic lines should have nonsignifi-cant differences with their control in F
1 char-
acters.k) Sow T
2 seeds derived from T
1 plants (nearly
50 plants are required for each T1-derived
plot). Spray Basta in the nursery, select oneplot in which all seedlings are resistant toBasta from each line, and transplant them.
l) Harvest the selfed seeds from the Basta-re-sistant plants; these will give a transgenicpollen parent possessing the Bar gene.
m) In the next year, use these transgenic Barlines to test the production potential of thetwo-line transgenic hybrid. If the produc-tion potential of the hybrid is acceptable,produce the bulk F
1 seed. Carry out the nor-
mal hybrid trial to ascertain transgenic re-storer lines with practical production po-tential.
n) Seed the bulk seeds of transgenic two-linehybrids in the seedbed and spray Basta onthe seedbed to kill the selfed EGMS seed-lings and other nonhybrid plants. Only truehybrid seedlings carrying the Bar gene willsurvive in the seedbed, which can be trans-planted to the field.
Special note: Any field experiments withtransgenic plants need proper approval and per-mission from the government and must be done inan isolated area for testing. Transgenic plants andtheir derived seeds are forbidden to serve as par-ents of a cross, animal feed, and human food with-out government approval. This approach, there-fore, cannot be used freely by plant breeders. As aresult, it has limited practical application.
The purity standards of two-line hybrids arebased on grade, varietal purity, seed purity, ger-mination percentage, and percent moisture con-tent. The standards for China appear in Table 16.
The criteria of breeder seeds and foundationseeds of three-line hybrids appear in Tables 17and 18.
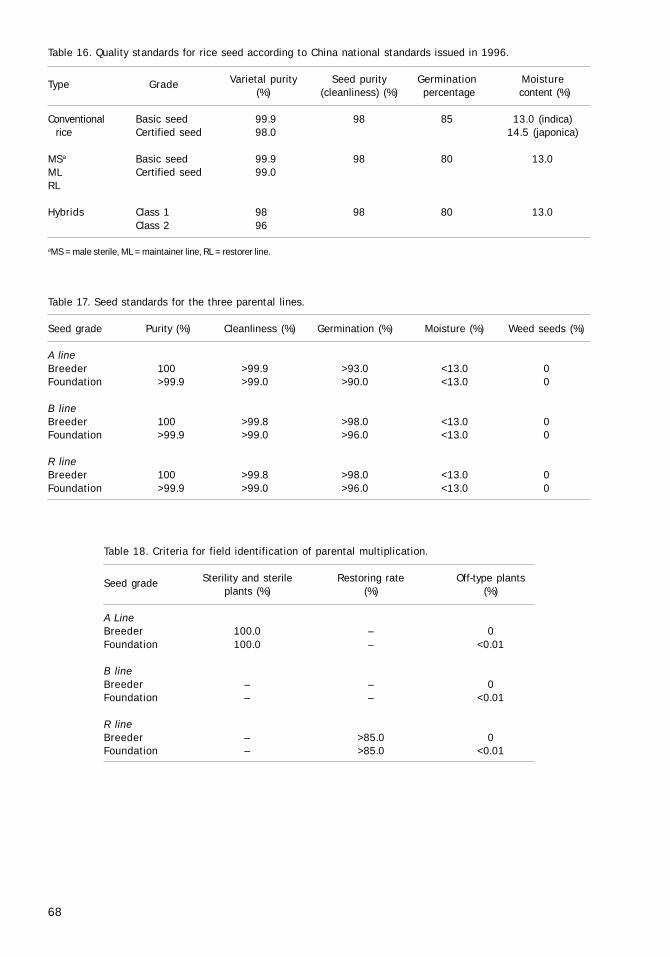
68
Table 16. Quality standards for rice seed according to China national standards issued in 1996.
Type Grade Varietal purity Seed purity Germination Moisture(%) (cleanliness) (%) percentage content (%)
Conventional Basic seed 99.9 98 85 13.0 (indica) rice Certified seed 98.0 14.5 (japonica)
MSa Basic seed 99.9 98 80 13.0ML Certified seed 99.0RL
Hybrids Class 1 98 98 80 13.0Class 2 96
aMS = male sterile, ML = maintainer line, RL = restorer line.
Table 17. Seed standards for the three parental lines.
Seed grade Purity (%) Cleanliness (%) Germination (%) Moisture (%) Weed seeds (%)
A lineBreeder 100 >99.9 >93.0 <13.0 0Foundation >99.9 >99.0 >90.0 <13.0 0
B lineBreeder 100 >99.8 >98.0 <13.0 0Foundation >99.9 >99.0 >96.0 <13.0 0
R lineBreeder 100 >99.8 >98.0 <13.0 0Foundation >99.9 >99.0 >96.0 <13.0 0
Table 18. Criteria for field identification of parental multiplication.
Seed grade Sterility and sterile Restoring rate Off-type plantsplants (%) (%) (%)
A LineBreeder 100.0 – 0Foundation 100.0 – <0.01
B lineBreeder – – 0Foundation – – <0.01
R lineBreeder – >85.0 0Foundation – >85.0 <0.01

69
CHAPTER 12
Future outlook fortwo-line rice hybrids
The exploitation of hybrid vigor in rice has shownthe way to increasing rice production after yieldstagnation with the use of semidwarf inbred ricevarieties in irrigated ecosystems. The deploymentof three-line rice hybrids in China and elsewherein Asia has substantially increased the hopes ofsustaining Asian food security. Hybrid rice tech-nology has caught the attention of rice farmersoutside China and, during the next ten years, sev-eral countries should have a large area coveredwith rice hybrids. There is a continuous need toreduce the cost and increase the efficiency of hy-brid rice seed production. The discovery of theEGMS system in China and later in Japan, at IRRI,and in India has improved the chances of substan-tially reducing the cost of seed production by us-ing two-line rice hybrids. These hybrids also helpto increase heterosis beyond the level of three-line rice hybrids. The two-line hybrids have al-ready created an impact in China, with their areareaching 2.6 million ha. Among the several two-line rice hybrids, Liangyou PeiJiu (Peiai64S/9311)gave the highest average yields of 11 tha–1 on 10-ha farms in two successive years. Thehighest yield recorded was about 12.1 t ha–1, anew record in these areas, clearly revealing theenhanced potential of two-line rice hybrids. Out-side China, two-line rice hybrids are also beingdeveloped at IRRI and in Vietnam, India, Korea,the Philippines, Thailand, and Egypt.
Future research priorities include the follow-ing:
1. Development of stable EGMS lines. Stableelite EGMS lines with a precise fertility al-teration mechanism hold the key to successin developing two-line commercial hybrid
rice. The underlying genetic mechanism offertility alteration needs to be understoodclearly to properly enhance the efficacy ofEGMS seed multiplication and hybrid riceseed production. Breeding of TGMS lineswith a low CSP is important for developingtwo-line commercial rice hybrids in the trop-ics. The genetic characterization of the lociof the EGMS genes from different sourcesin relation to closely tagged molecular mark-ers is useful for marker-assisted selection.
2. Use of anther culture to develop and/orpurify elite EGMS lines. Anther culture tech-niques involving dihaploidization can beused to expedite the development and/orpurification of EGMS lines possessing ma-jor genes and QTLs in influencing thePGMS/TGMS trait.
3. Breeding for super high-yielding two-linehybrids. Two-line hybrid rice technologyinvolving EGMS lines allows the choice ofa wider range of parental combinations andavoids the negative effects of male-steril-ity-inducing cytoplasm. Rice scientists atIRRI and in China can now use new planttype (NPT) lines developed in the tropicaljaponica and indica/tropical japonica back-ground as male and/or female parents to de-velop hybrids with enhanced heterosis.Two-line breeding technology can over-come the major problems of wide incom-patibility and the narrow range of restorersin exploiting indica-japonica heterosis.
4. Incorporation of hybrids with resistance tobiotic stress. Two-line rice hybrids possess-ing multiple resistance to diseases and in-

70
sects can be developed more expeditiouslythan three-line hybrids since the desired re-sistance genes need to be incorporated intwo rather than three parental lines.
5. Abiotic stress tolerance. Hybrid rice tech-nology so far has been used under the irri-gated rice ecosystem. Environmental deg-radation resulting from salinity and watershortages has posed a major threat to sus-taining food production. Researchers at IRRIand in Egypt, India, and China have foundthat hybrid rice technology can be extendedto saline-prone irrigated conditions becausehybrids have performed exceedingly wellunder moderate to high saline soil condi-tions. Since the high cost of seed is a majorconstraint to the adoption of this technol-ogy by resource-poor farmers and two-linehybrid breeding and the seed production
approach make it more cost-effective, thistechnology can be adopted by resource-poorfarmers.
6. Quality. The negative influence of WA cy-toplasm on certain quality parameters (suchas grain chalkiness) allows the alternativeuse of EGMS-based two-line hybrid ricetechnology to overcome such drawbacks.
A multidisciplinary approach in developingsuperior EGMS lines and pollen parents can helpto develop two-line rice hybrids suitable for thedifferent ecological situations in which rice isgrown. Despite the promise that two-line hybridrice technology holds, it would be wise to have aharmonious balance in using two-line and three-line hybrids and conventional rice varieties in anappropriate manner in national rice productionprograms.

71
Abdallah AA, Verkerk K. 1968. Growth, floweringand fruit set of the tomato at high temperature.Neerl. J. Agric. Sci. 16:71-76.
Agarwala SC, Chatterjee C, Sharma PN, Sharma CP,Nautiyal N. 1979. Pollen development in maizeplants subjected to molybdenum deficiency. Can.J. Bot. 57:1946-1950.
Agarwala SC, Sharma PN, Chatterjee C, Sharma CP.1980. Copper deficiency induced changes inwheat anthers. Proc. Indian Natl. Sci. Acad. Sect.B 2:172-176.
Ahokas H, Hockett EA 1977. Male sterile mutant ofbarley. IV. Different fertility levels of msgqci (cv.Vantage), an ecoclinal response. Barley Genet.Newsl. 7:10-11.
Ali J. 1990. Studies on gametocides in rice (Oryzasativa L.). M.Sc. thesis submitted to the IndianAgricultural Research Institute, New Delhi, In-dia. 124 p.
Ali J. 1993. Studies on temperature sensitive genicmale sterility and chemical induced male sterilitytowards development of two-line hybrids in rice(Oryza sativa L.). Ph.D. thesis submitted to theIndian Agricultural Research Institute, New Delhi,India. 138 p.
Ali J, Siddiq EA, Zaman FU, Abraham MJ, Ahmed I.1995. Identification and characterization of tem-perature sensitive genic male sterile sources inrice (Oryza sativa L.). Indian J. Genet. 55(3):243-259.
Ali J. 1996. Characterisation and genetics of tempera-ture sensitive male sterile line SA 2 in rice. Ab-stract: PI-102. 2nd International Crop ScienceCongress, New Delhi, India.
Anonymous. 1978. Studies on male sterility of riceinduced by male gametocide no. 1. Acta Bot. Sin.230:303-313.
Atsumi S, Takeo N, Yasuo K. 1992. Sterility in riceinduced by chemical treatment. Biosci. Biotech.Biochem. 56(1):1619-1622.
Batch JJ, Morgan, DG. 1974. Male sterility induced inbarley by photoperiod. Nature 250:165-167.
Berthelem P, Le Guen J. 1975. Rapport d’activite-sta-tion d’amelioration des plantes. INRA Rennes(France): 1971-74. (As reported in Kaul, 1988.)
Borkakati RP, Virmani SS. 1993. Inheritance of athermosensitive genic male sterile mutant of in-dica rice. Rice Genet. Newsl. 10:92-94.
Brar DS. 1982. Male sterility in sesame. Indian J.Genet. 42:23-27.
Chan YW, Cheah CH. 1981. Evaluation of ethrel as aselective gametocide in the breeding of rice(Oryza sativa L.). In: Proceedings of the 4th In-ternational SABRAO Congress, Malaysia. p 24.
Chen P. 1985. Studies on the effect of amidosulfonicacid (a male sterility agent) to induce male steril-ity of rice. (Hua-nan-nung-yeh da hsueh-huseh-pao.) J. South China Agric. Univ. 6(2):62-68.
Chen PL, Chen B, Chen JL, Ho LK. 1986. Arseniccontent and castration effect in reaction to malegametocide no. 2 in rice plants. (Hua-nan-nung-yeh da hsueh-huseh-pao.) J. South China Agric.Univ. 7(4):22-27.
Cheng YK, Huang, CS. 1978. Some properties of cy-toplasmic male sterile rice. J. Agric. Res. Taiwan27:267-290.
Cheng YK, Huang CS. 1980. Studies on the cytoplas-mic–genic male sterility in cultivated rice (Oryzasativa L.). II. Morphological-histological investi-gations on functional male sterility. J. Agric. Res.China 29:67-80.
Cheng S, Sun Z, Si H, Zhuo L. 1995. Response tophotoperiod and temperature in photo-thermo pe-riod sensitive genic male sterile line Xinguang S(Oryza sativa L. subsp. indica). Chinese J. RiceSci. 9(2): 87-91.
Cheng SH, Si HM, Zhou LS, Sun ZX. 1996. Classifi-cation of environmentally induced genetic malesterile lines of rice based on their fertility responsesto photoperiod and temperature. J. Agric. Sci.(Camb.) 127:161-167.
Bibliography

72
Chu CC, Wang CC, Sun CS, Hsu C, Yin KC, Chu Y.1975. Establishment of an efficient medium foranther culture of rice through comparative ex-periments on the nitrogen source. Sci. Sin. 18:659-668.
Comstock RE, Robinson HF. 1952. Estimation of av-erage dominance of genes. In: Heterosis. Ames,Iowa (USA): Iowa State College Press. p 494-516.
Daskaloff S. 1972. Male sterile pepper (C. annuum)mutants and their utilization in heterosis breeding.Proceedings of Eucarpia meeting. Capsicum7:205-210.
Davenport CB. 1908. Degeneration, albinism and in-breeding. Science 28:454-455.
Dell B. 1981. Male sterility and anther structure in cop-per deficient plants. Ann. Bot. 48:599-608.
Duc G. 1980. Effect of environment on instability oftwo sources of cytoplasmic male sterility in fababeans. FEBS Lett. 2:29-30.
Duvick DN. 1966. Influence of morphology and steril-ity on breeding methodology. In: Frey KJ, editor.Plant breeding. Vol. I. Ames, Iowa (USA): IowaState University Publications. p 85-138.
East EM. 1908. Inbreeding in corn. Rep. ConnecticutAgric. Exp. Stn. for 1907. p 419-428.
East EM. 1936. Heterosis. Genetics 21:375-397.Fisher JE 1972. The transformation of stamens to ova-
ries and of ovaries to inflorescences in I. aestivumL. under short day treatment. Bot. Gaz. 133:78-85.
Ikehashi H, Araki H. 1984. Varietal screening for com-patibility types revealed in F
1 sterility of crosses
in rice. Jpn. J. Breed. 34(3):304-312.Ikehashi H, Araki H. 1986. Genetics of F
1 sterility in
remote crosses in rice. In: Rice genetics. Manila(Philippines): International Rice Research Insti-tute. p 119-130.
Jan CC. 1974. Genetic male sterility in wheat (Triti-cum aestivum L.): expression, stability, inherit-ance and practical use. Ph.D. thesis. Universityof California, Davis, Calif., USA.
Jia JH, Zhang DS, Li CY, Qu XP, Wang SW,Chamarerk V, Nguyen HT, Wang B. 2001. Mo-lecular mapping of the reverse thermo-sensitivegenic male sterile gene (rtms 1) in rice. Theor.Appl. Genet. 103(4):607-612.
Jinks JL. 1983. Biometrical genetics of heterosis. In:Frankel R, editor. Heterosis: reappraisal of theoryand practice. Berlin (Germany): Springer-Verlag.
Kaul MLH. 1988. Male sterility in higher plants. Mono-graph on Theoretical and Applied Genetics 10.Berlin (Germany): Springer-Verlag. 1,005 p.
Kempthorne O. 1957. An introduction to genetic statis-tics. London (UK): John Wiley and Sons, Inc.
Kitaoka M, Ota K, Ogawa M. 1991. The synergisticeffects of ethephon and isourea in male sterility inrice. Jpn. J. Crop Sci. 60(3):349-354.
Li X, Pan XB, Chen ZX, Yu HX, Cheng ZK, Giu MH1995. Anther culture of photosensitive male ster-ile rice and the breeding of Miai 64S. J. JiangsuAgric. Coll. 16(4):7-12.
Li XL, Xie R, Deng XH, He GH, Yang ZL, Zuo YS,Wu LJ. 1994a. Comparison of the effect of antherculture in a photoperiod (temperature) sensitivegenic male sterile line and a wide compatibilityrestoring line in rice. Southwest China J. Agric.Sci. 8 (Special Issue):88-93.
Li RH, Cai HW, Wang XK. 1994b. A specific es-terase band found in Annong 1-S. Int. Rice Res.Newsl. 19(3):6-7.
Li ZK, Luo LJ, Mei HW, Shu QY, Tabien R, ZhongDB, Ying CS, Stansel JW, Khush GS, PatersonAH. 2000. Genetic basis of inbreeding depres-sion and heterosis in rice (Oryza sativa L.). In:Paper presented at the International Rice Re-search Conference, 31 March-3 April 2000. LosBaños (Philippines): International Rice ResearchInstitute.
Long JD, Weed MB, Fitzgerald DJ, Barrier GE. 1973.Chemical control of pollination. In: Proceedingsof the American Society of Agronomy, Madison,Wis. (USA). p 69.
Lu X, Wang J. 1988. Fertility transformation and ge-netic behavior of Hubei photoperiod-sensitivegenic male sterile rice. In: Hybrid rice. Los Baños(Philippines): International Rice Research Insti-tute. p 129-138.
Lu XG, Virmani SS, Yang RC. 1998. Advances intwo-line hybrid rice breeding. In: Virmani SS,Siddiq EA, Muralidharan K, editors. Advances inhybrid rice technology. Proceedings of the ThirdInternational Symposium on Hybrid Rice, 14-16November 1996, Hyderabad, India. Los Baños(Philippines): International Rice Research Insti-tute. p 89-98.
Lu XG, Zhang ZG, Maruyama, K, Virmani SS. 1994.Current status of two-line method of hybrid ricebreeding In: Virmani SS, editor. Hybrid rice tech-nology: new developments and future prospects.Manila (Philippines) International Rice ResearchInstitute. p 37-39.
Luo Z, Zhong A, Zhou Q. 1988. Effects of a new nontoxic chemical gametocide on rice. In: Hybridrice. Manila (Philippines): International Rice Re-search Institute. p 147-156.
Martin JA, Crawford JH. 1951. Several types of steril-ity in Capricum frutescens. Proc. Am. Soc. Hort.Sci. 57:335-338.

73
Maruyama K, Araki H, Kato H. 1991. Thermosensitivegenetic male sterility induced by irradiation. In:Rice genetics II. Manila (Philippines): Interna-tional Rice Research Institute. p 227-235.
Mei G, Wang X, Wang, M. 1990. Genetical analysisof the photoperiod sensitive male sterility ofNongken 58s and its derivatives. J. HuazhongAgric. Univ. 9(4):400-406.
Mei CS, Zhang JY, Wu GN. 1988. A preliminary re-port on increasing of green pollen plants in indicarice. Jiangsu J. Agric. Sci. 4(2):45-48.
Mei MH, Dai KK, Xu CG, Zhang Q. 1999. Mappingand genetic analysis of the genes for photoperiod-sensitive genic male sterility in rice using the origi-nal mutant Nongken58S. Crop Sci. 39:1711-1715.
Mou TM, Li CH, Yang GC, Lu XG. 1998. Breedingand characterizing indica PGMS and TGMS linesin China. In: Virmani, SS, Siddiq EA,Muralidharan K, editors. Advances in hybrid ricetechnology. Proceedings of the Third InternationalSymposium on Hybrid Rice, 14-16 November1996, Hyderabad, India. Manila (Philippines): In-ternational Rice Research Institute. p 79-88.
Murty UR. 1995. Breeding two line hybrids in Sor-ghum bicolor (L.) Moench. Cereal Res. Comm.23:397-402.
Oard JH, Hu J. 1995. Inheritance and characterizationof pollen fertility in photoperiodically sensitive ricemutants. Euphytica 82:17-23.
Pan XG, Zhu DY, Yin JH, Ding XH 1993. A study onthe technological procedures for breeding dual-purpose genic male-sterile rice. Acta Agric.Jiangxi, China 5(Supplement):1-5.
Parmar KS, Siddiq EA, Swaminathan MS. 1979.Chemical induction of male sterility in rice. In-dian J. Genet. Plant Breed. 39:529-541.
Perez AT, Chang TT, Beachell HM, Vergara BS,Marciano AP. 1973. Induction of male sterility inrice with ethrel and RH-531. SABRAO Newsl.5:133-139.
Peterson PA. 1958. Cytoplasmically inherited malesterility in Capsicum. Am. Nat. 92:111-119.
Reddy OUK, Siddiq EA, Sarma NP, Ali J, HussainAJ, Nimmakayala P, Ramasamy P, Pammi S,Reddy AS. 2000. Genetic analysis of tempera-ture-sensitive male sterility in rice. Theor. Appl.Genet. 100:794-801.
Rerkasem B, Jamjod S. 1997. Boron deficiency in-duced male sterility in wheat (Triticum aestivumL.) and its implications for plant breeding.Euphytica 96:257-262.
Rick CM, Boynton JE. 1967. A temperature sensitivemale sterile mutant of the tomato. Am. J. Bot.54:601-611.
Rudich, J, Peles A. 1976. Sex expression in water-melon as affected by photoperiod and tempera-ture. Sci. Hort. 5:339-344.
Rundfeldt H. 1960. Untersuchungen zur zuchtung desKopfkohls (B. olerecea L. var. Capitata). Z.Pflanzenzuchtung 44:30-62. (As reported in Kaul,1988.)
Rutger JN, Schaeffer GW. 1989. An environmentallysensitive genetic male sterile mutant in rice. In:Annual Meeting of the American Society ofAgronomy, Las Vegas, Nev., USA. Agron. Abstr.p 98.
Sawhney VK. 1983. Temperature control of male ste-rility in a tomato mutant. J. Hered. 74:51-54.
Sharma RK, Reinbergs E. 1976. Male sterility genesin barley and their sensitivity to light and tempera-ture intensity. Ind. J. Genet. Plant Breed. 3 2:408-410.
Shen YW, Kai QH, Gao M. 1993. A new thermo sen-sitive radiation induced male sterile rice line. RiceGenet. Newsl. 10:97-98.
Shen Y, Gao, M, Cai Q. 1994. A novel environment-induced genic male sterile (EGMS) mutant in in-dica rice. Euphytica 76:89-96.
Sheng XB. 1992. Genetics of photoperiod sensitivegenic male sterility of Nongken 58S (Oryza sa-tiva L.). Chinese J. Rice Sci. 6(1):5-14.
Shi MS. 1981. Preliminary report of breeding and utili-zation of late japonica natural double purpose line.J. Hubei Agric. Sci. 7:1-3.
Shi MS. 1985. The discovery and the study of the pho-tosensitive recessive male sterile rice (Oryza sa-tiva L. subsp. japonica). Sci. Agric. Sin. 2:44-48.
Shi MS, Deng JY. 1986. The discovery, determinationand utilization of Hubei photosensitive genic malesterile rice (Oryza sativa L. subsp. japonica). ActaGenet. Sin. 13(2):107-112.
Song MT, Kim, JK, Choe ZR. 1990. Effects of chemi-cals on inducing grain sterility of rice. Korean J.Crop Sci. 35(4):309-314.
Stevens MA, Rudich J. 1978. Genetic potential forovercoming physiological limitations on adaptabil-ity, yield and quality in tomato. Hort. Sci. 13:673-678.
Stuber CW, Lincoln SE, Wolff DW, Helentjaris T,Lander ES. 1992. Identification of genetic factorscontributing to heterosis in a hybrid from two elitemaize inbred lines using molecular markers. Ge-netics 132:823-839.
Subudhi PK, Borkakati RP, Virmani SS. Huang N.1997. Molecular mapping of a thermosensitivegenetic male sterility gene in rice using bulkedsegregant analysis. Genome 40:188-194.
Sun ZX, Min SK, Xiong ZM. 1989. A temperaturesensitive male sterile line found in rice. RiceGenet. Newsl. 6:116-117.
Takeoka YI, Ito M, Yamamura S, Saka H. 1990. Mor-phogenetic alterations of rice floral organs as af-fected by spray of chemical hybridizing agentHGR-626. Jpn. J. Crop Sci. 59(3):528-534.

74
Tan ZC, Li YY, Chen LB, Zhou GQ. 1990. Studies onecology and adaptability of dual-purpose line AnNong S-1. Hybrid Rice 3:35-38.
Virmani SS, Maruyama K. 1995. Some genetic toolsfor hybrid breeding and seed production in selfpollinated crops. In: Sharma et al, 1995 editors.Genetic research and education: current trendsand next fifty years. New Delhi (India): IndianSociety of Genetics and Plant Breeding. p 708-728.
Virmani SS, Voc PC. 1991. Induction of photo- andthermo-sensitive male sterility in indica rice.Agron. Abstr. 119.
Virmani SS. 1992. Transfer and induction ofthermosensitive genic male sterile mutant in in-dica rice. In: Proceedings of the Second Interna-tional Symposium on Hybrid Rice, 21-25 Apr.1992. Manila (Philippines): International RiceResearch Institute.
Virmani SS, Viraktamath BC, Lopez MT. 1997b.Nucleus and breeder seed production ofthermosensitive genic male sterile lines. Int. RiceRes. Newsl. 22(3):26-27.
Virmani SS, Viraktamath BC, Casal CL, Toledo RS,Lopez MT, Manalo JO. 1997a. Hybrid rice breed-ing manual. Manila (Philippines): InternationalRice Research Institute. 151 p.
Wang B, Wang JZ, Wu W, Zheng HG, Yang ZY, XuWW, Ray JP, Nguyen HT. 1995. Tagging andmapping the thermosensitive genic male sterilegene in rice (Oryza sativa L.) with molecularmarkers. Theor. Appl. Genet. 91(6-7):1111-1114.
Wang X, Que RF. 1981. Induction of pollen sterility byethylene and gametocides in rice. ActaPhytophysiol. Sin. 7:381-383.
Wang X, Que, RF, Xu MF. 1981. Preliminary study onthe effect of AWN on the induction of male steril-ity in rice. Zheijiang Nongye Kexue (ZheijiangAgric. Sci.) 3:135-139.
Wang X, Yu M, Tao LX, Yao F. 1991a. The malesterility in rice plant induced by CRMS. I. Effectof CRMS on male sterility in rice plants. ChineseJ. Rice Sci. 5(2)49-55.
Wang, X, Yu M, Tao LX, Yao F. 1991b. The malesterility in rice plant induced by CRMS. II. Physi-ological effects of CRMS on inducing male steril-ity in rice. Chinese J. Rice Sci. 5(3)104-108.
Wang, X, Yu M, Tao LX, Yao F. 1991c. The malesterility in rice plant induced by CRMS. III. Effectof CRMS on free proline content in anthers of riceand its possible relation to abortion of anther. Chi-nese J. Rice Sci. 5(4):169-174.
Wang XM, Wang MQ, Mei GZ, Wu HG, Duan WJ,Wang WJ. 1991. Photoperiod conditioned malesterility and its inheritance in rice. In: Rice genet-ics II. Proceedings of the Second InternationalRice Genetics Symposium, 14-18 May 1990.Manila (Philippines): International Rice ResearchInstitute. p 217-226.
Wang FP, Mei MH, Xu CG, Zhang Q. 1997. pms1 isnot the locus revelant to fertility difference be-tween the photoperiod-sensitive male sterile riceNongken 58S and normal rice Nongken58. ActaBot. Sin. 39:922-925.
Wei BG, Sun GC, Chang JW, Jiang CX. 1994. Thedetermination of photoperiod-temperature sensi-tive male sterile soybean. Proceedings of ThirdNational Youth Symposium on Crop Genetics andBreeding. Edited by China Agricultural ScientechPress. p 185-189.
Xi DW, Cheng WJ, Yi DL, Yu DR, Ning ZL, DengXX, Li M. 1997. Genetic analysis on the TGMSline in rape of B. napus Xiangyou 91S. In: Lu RL,Cao XB, Liao FM, Xin YY, editors. Proceedingsof the International Symposium on Two-LineSystem of Heterosis Breeding in Crops, 6-8 Sep-tember 1997, Changsha, China. p 215-220.
Xiang YW, Zhang ZX, Zhang AZ, Zhou XM, Wie R.1993. Selection and breeding of photosensitivegenic male sterile rice by anther and somatic cellculture. J. Southwest Agric. Univ. 15(5):462-465.
Xiao J, Li J, Yuan LP, Tanksley SD. 1995. Domi-nance is the major genetic basis of heterosis inrice as revealed by QTL analysis using molecu-lar markers. Genetics 140:745-754.
Xiao GY, Yuan LP, Fu, XQ, Deng XP. 1997. Studieson effect of water temperature on male fertility ofthe thermo-sensitive genic male sterile (TGMS)lines in rice under simulated low air temperatureconditions in high summer. In: Lu RL, Cao XB,Liao FM, Xin YY, editors. Proceedings of theInternational Symposium on Two-Line Systemof Heterosis Breeding in Crops, 6-8 September1997, Changsha, China. p 64-69.
Xie R, Deng XH, Li XL, Luo GC, Wu LJ, He XY.1993. Studies on fertility transition in the antherculture-derived progeny of photothermo-sensitivegenic male sterile rice. J. Southwest Agric. Univ.15(5):450-453.
Yamaguchi Y, Ikeda R, Hirasawa H, Minami M,Ujikara A. 1997. Linkage analysis ofthermosensitive genic male sterility gene, tms
2, in
rice (Oryza sativa L.). Breed. Sci. 47:371-373.Yang RC, Wang NY, Mang K, Chau Q, Yang RR,
Chen S. 1990. Preliminary studies on applicationof indica photo-thermo genic male sterile line5460S in hybrid rice breeding. Hybrid Rice 1:32-34.

75
Yang YQ, Wang L. 1990. The breeding ofthermosensitive male sterile rice R59TS. Sci.Agric. Sin. 23(2):90.
Yang RC, Liang K, Wang N, Chen S. 1992a. A reces-sive gene in indica rice 5460S for thermosensitivegenic male sterility. Rice Genet. Newsl. 9:56-57.
Yang RC, Wang NY, Mang, K, Chau Q, Yang, RR,Chen S. 1992b. Preliminary studies on applica-tion of indica photo-thermo genic male sterile line5460S in hybrid rice breeding. Hybrid Rice 1:32-34.
Yang ZP. 1997. Inheritance of photoperiod genic malesterility and breeding of photoperiod sensitivegenic male sterile lines in rice (Oryza sativa L.)through anther culture. Euphytica 94:93-99.
Yin HQ. 1999. Scientific progress made in the two linesystem of hybrid rice research and production inChina. Hunan Agric. Res. Newsl. 6(3):4-8.
Yu SB, Zi JX, Xu CG, Tan YF, Gao YJ. 1997. Impor-tance of epistasis as the genetic basis of heterosisin an elite rice hybrid. Proc. Natl. Acad. Sci. USA94:9226-9231.
Yuan LP. 1997. Exploiting crop heterosis by two-linesystem hybrids: current status and future pros-pects. In: Proceedings of the International Sym-posium on Two-Line System of Heterosis Breed-ing in Crops, 6-8 September 1997, Changsha,China.
Yuan SC, Zhang ZG, He HH, Zen HL, Lu KY, LianJH, Wang BX. 1993. Two photoperiodic reac-tions in photoperiod-sensitive genic male sterilerice. Crop Sci. 33:651-660.
Zhang M, Liang C, Huang Y, Chen B. 1994. Fertilityexpression of photoperiod temperature sensitivegenic male sterile rice in Guangzhan and the re-sponse to photoperiod and temperature treatments.J. Trop. Subtrop. Bot. 2(4):100-107.
Zhang Z, Zeng H, Yang J, Yuan SC, Zhang D. 1994.Conditions inducing fertility alteration and eco-logical adoption of photoperiod-sensitive genicmale sterile rice. Field Crops Res. 38(2):111-120
Zhang ZG, Yuan SC, Zen HL, Li YZ, Li ZC, WeiCL. 1990. Preliminary analysis on fertilitychanges and adaptability of HPGMR at differentaltitudes in Yuanjiang. J. Huazhong Agric. Univ.9(4):348-354.
Zhang KT, Fu HY. 1982. Effect of high temperaturetreatment on male sterility in sorghum. Acta Genet.(China) 9:71-77.
Zhang NY, Xue QZ. 1996. Development of photope-riod sensitive genic male sterile lines using antherculture in rice (Oryza sativa L.). J. Zhejiang Agric.Univ. 22(5):474-480.
Zhang Q, Shen BZ, Dai XK, Mei MH, Saghai-MaroofMA, Li ZB. 1994. Using bulked extremes andrecessive class to map genes for photoperiod sen-sitive genic male sterility in rice. Proc. Natl. Acad.Sci. USA 91:8675-8679.
Zhang Q, Hua J, Yu S, Xiong L, Xu C. 2001. Geneticand molecular basis of heterosis in rice. In: KhushGS, Brar DS, Hardy B, editors. Rice genetics IV.Proceedings of the Fourth International Rice Ge-netics Symposium, 22-27 October 2000, LosBaños, Philippines. New Delhi (India): SciencePublishers, Inc., and Los Baños (Philippines): In-ternational Rice Research Institute. p 173-188.
Zhang ZG, Yuan, SC. 1989. Effect of twilight on twophotoperiod reactions in Hubei photoperiod sen-sitive male sterile rice. Chinese J. Rice Sci.3(3):107-112.
Zhang ZG, Lu XG, Yuan LP. 1992a. Reflections onidentification of light and temperature propertiesof fertility transformation of photoperiod sensitivemale sterile rice. Hybrid Rice 6:29-32.
Zhang ZG, Yuan, SC, Xu CZ. 1987. The influence ofphotoperiod on fertility changes of Hubei photo-period sensitive genetic male sterile rice. ChineseJ. Rice Sci. 1(3):137-143.
Zhang ZG, Yuan SC, Zen HL, Li YZ, Zhang DP.1992b. Studies on model of photo-thermo reac-tion of fertility alteration in photosensitive geneticmale sterile rice. J. Huazhong Agric. Univ. 11:1-6.
Zhang ZG, Yuan SC, Zen HL, Li YZ, Li ZC, Wei CL.1990. Preliminary analysis on fertility changes andclimatic adaptation of HPGMR at different alti-tudes in Yuan Jiang. J. Huazhong Agric. Univ.9:348-354.
Zhang ZG, Yuan SC, Zen HL, Li YZ, Li ZG, WeiCL. 1991. Preliminary observations of fertilitychanges in a new type temperature sensitive malesterile rice IVA. Hybrid Rice 1:31-34.
Zhangxing Z, Chunnong H. 1980. A study of malesterility induced in rice (Oryza sativa L.) by 3-(p-chlorophenyl) 6-methoxy-S-triazine-2,4-(1H, 3H)Dione. J. Huazhong Univ. 4:81-87.
Zhu DY, Sun ZX, Pan XG, Ding XH, Shen XH, WanY, Pan H, Yin JH, Alejar MS, Torrizo LB, DattaSK. 1988. Use of anther culture in hybrid ricebreeding. In: Virmani SS, Siddiq EA,Muralidharan K, editors. Advances in hybrid ricetechnology. Proceedings of the Third InternationalSymposium on Hybrid Rice, 14-16 November1996, Hyderabad, India. Manila (Philippines): In-ternational Rice Research Institute. p 265-281.
Zhou TB, Xiao HC, Lei DY, Duan QZ. 1988. Thebreeding of indica photoperiod male sterile lines.J. Hunan Agric. Sci. 6:16-18.

76
Authors
S.S. Virmani is plant breeder and deputy head,Plant Breeding, Genetics, and Biochemistry Divi-sion, IRRI; Sun Zongxiu is a research professor atthe Key Laboratory for Rice Biology of the Min-istry of Agriculture and at the China National RiceResearch Institute, Hangzhou; Mou Tongmin is aprofessor in the Agronomy Department ofHuazhong Agricultural University in Wuhan,Hubei; Jauhar Ali is national coordinator of theHybrid and Molecular Rice Breeding Program ofIran at the Rice Research Institute of Iran in Rasht;Chang Xiang Mao was a plant breeder at IRRI andis now senior rice breeder cum chief of the Inter-national Project Office at the Rice Research Insti-tute of the Guangxi Academy of Agricultural Sci-ences in Nanning City, Guangxi, China.

77
Glossary
AA line – the male sterile parent involving cyto-
plasmic or cytoplasmic genetic male steril-ity. It is also known as a CMS line.
adaptability – the ability of a genotype to adjustto a given environment and give a reason-ably good yield.
allo-plasmic lines – CMS or restorer lines thathave different cytoplasms.
anther – terminal part of the stamen that containsthe pollen grains (male gametes).
anthesis – the action of opening of a flower orspikelet of rice.
apiculus – a small acute point or tip of a rice spike-let; extension of the lemma or palea.
apomixis – a kind of asexual reproductionthrough seed in which the embryo developsfrom a maternal cell without fertilization. Theresulting seed has the same genetic constitu-tion as that of the seed parent.
apparent heterosis – subjective superiority of ahybrid over its parents or a check varietybased on visual observation.
augmented design – a statistical design used forevaluation of genotypes in which the checkvarieties are replicated and the test entries arenot replicated but are allotted randomly tothe blocks.
auricles – the small paired ear-like appendageson either side of the base of the rice leaf bladethat may not be present in older leaves.
awn – a bristle-like extension of varying lengthoriginating from the lemma of the spikelet.
BB line – the fertile counterpart parent of the male
sterile A line of a cytoplasmatic or cytoplas-mic genetic male sterility system that is usedas a male parent to maintain the latter. It isalso known as a maintainer line.
backcross method – a breeding method in whichthe F
1 hybrid is again crossed with either of
its parents. The resulting progeny is called abackcross progeny.
backcross nursery – breeding nursery in whichmale sterile plants identified among the test-crosses (CMS ´ elite lines) are crossed withthe respective male parents to transfer cyto-plasmic male sterility into the nuclear geno-type of the elite line.
boot – a rapidly growing panicle enveloped bythe flag-leaf sheath. In tissue culture, this re-fers to the panicle collected when the distancebetween the collar of the flag leaf and sub-tending leaf is about 7 to 8 cm.
booting – bulging of the flag-leaf sheath becauseof the growing panicle inside.
border rows – the recommended number of rowsof the male parental line grown on all sides ofthe hybrid seed production plot to minimizecontamination by outcrossing with stray pol-len.
bract – a leaf from the axis from which a flowerarises.
breeder seed – breeder seed is the seed of the high-est genetic purity and is produced by theagency sponsoring a variety; it is used to pro-duce foundation seed.

78
Ccaryopsis – a small one-seeded dry indehiscent
fruit with a thin membranous pericarp adher-ing so closely to the seed that fruit and seedare incorporated in one body, forming asingle grain, as in wheat and barley. In rice,brown rice is the caryopsis.
certified seed – seeds used for commercial cropproduction produced from foundation, regis-tered, or certified seeds under the regulationof a legally constituted agency. In hybrid rice,it is F
1 seed produced directly from CMS ×
restorer lines grown as per certification stan-dards.
CHA (chemical hybridizing agent) – any chemi-cal that is used to induce male sterility inplants.
check variety – any popular or high-yielding va-riety widely grown in a region.
chemical mutagen – any chemical used to inducemutations artificially.
chemical hybridizing agent (CHA) – this is anychemical formulation, that is, auxins, antiaux-ins, growth regulators, arsenicals, oxanilates,ethylene-releasing compounds, halogenatedaliphatic acids, etc., that has the ability toselectively sterilize the male gametes with-out affecting ovular fertility and is not phy-totoxic.
CMS – the CMS line is governed by genetic fac-tors present in the mitochondria of the cyto-plasm responsible for inducing selective malesterility. But its pistil is normal and it canproduce seeds when pollinated by any nor-mal plant.
combining ability – the ability of a genotype (in-bred, pure line, or synthetic) to transfer itsdesirable traits to its progeny: general—av-erage performance of a strain in a series ofcrosses; specific—deviation from perfor-mance predicted on the basis of general com-bining ability of parental lines.
correlation coefficient – a measure of the degreeof association between two variables that iscomputed as the ratio of the covariance of thetwo variables to the products of their stan-dard errors. Its values vary between –1 and+1.
covariance – the mean of the product of the de-viation of two varieties from their individualmeans.
critical difference – a statistical parameter com-puted to test whether the observed differencesbetween the means of entries are significantor not.
critical fertility point (CFP) – the critical tem-perature or photoperiod experienced by theEGMS line during the sensitive stage resultsin maximum pollen and spikelet fertility.
critical sterility point (CSP) – the critical tem-perature or photoperiod experienced by theEGMS line during the sensitive stage resultsin complete pollen and spikelet sterility.
cross fertilization – the fertilization of an eggnuclei (ovule) of one parent by the pollen ofanother parent.
cytoplasm – all the protoplasm of the cell exceptthe nucleus.
cytoplasmatic heredity/inheritance – the trans-mission of characters from parent to offspringthrough the cytoplasm of the germ cell.
Ddaylength - number of light hours in a day.diallel mating – a mating design in plant breed-
ing in which a set of parents is crossed in allpossible combinations.
dihybrid – a hybrid of two different genes; het-erozygous for two pairs of alleles.
diploid (2n) – an organism having two chromo-somes of each kind.
disomic – a plant having one or more chromo-somes duplicated, but not the entire genome.
diverse – having or capable of having variousforms or qualities.
dominance – intra-allelic/intragenic interactionwith complete suppression of the effects ofone allele by another.
Eeffective accumulated temperature (EAT) – the
total effective temperature in centigrade re-ceived by the plant from seeding to flower-ing. It is useful for predicting flowering.
EAT = Mean daily temperature (°C) – tem-perature higher than 30 °C – temperature oflower limit (18 °C)
emasculation – the process of removal of anthersfrom the florets so as to make the plant malesterile.

79
elite line – an improved breeding line or a variety.endosperm – the nutritive tissue of the ripened
ovary. It consists of the aleurone layer andthe starchy tissue, and serves as the source offood for the germinating embryo.
environmental genic male sterility (EGMS) –male sterility–fertility transformation con-trolled by environmental factors such as tem-perature and photoperiod.
epistasis – the interaction of different genes in theexpression of a trait.
FF
1 – abbreviation for the first filial generation,
usually the hybrid between two homozygousparents.
fertility restoration – an ability of a genotype torestore fertility to its progeny when crossedto a CMS line.
fertilization – fusion of the nuclei of male andfemale gametes.
flag leaf – the uppermost leaf (of rice plant) origi-nating just below the panicle base.
flag-leaf clipping – a method of cutting 1/2 to2/3 of the flag leaf from its tip in CMS andrestorer lines to facilitate easy pollen dispersal.
floret – a unit of the spikelet, which includes thelemma, palea, and the flower.
flower, rice – the reproductive organ consistingof lemma, palea, two lodicules, six stamens,and the pistil.
foundation seed – seed stock produced frombreeder seed by or under the direct control ofa breeder or a research station. Foundationseed is the source of certified seed, either di-rectly or through registered seed.
GGA
3 – a form of gibberellic acid that is sprayed onCMS lines to obtain good panicle exsertion.
gamete – a mature reproductive male or femalegerm cell, sperm, or egg specialized for fer-tilization.
gametic (tissue or generation) – having “n” num-ber of chromosomes (haploid), in contrast tozygotic tissue with 2n (diploid).
gametocide – organic or inorganic chemicals usedfor killing the functional sexual parts (pol-len, ovule) of the plant. These may be selec-tive for male or female parts.
gametophytic – in this system, the sterility/fertil-ity reaction is imparted to the pollen by thegenetic constitution of the pollen itself andis controlled by a single gene, which may havea large number of allelic forms.
genetic purity – trueness to type; seeds/plantsconfirming to the characteristics of the line/variety/hybrid as described by the breeder.
genetic shift – change in the genetic makeup ofthe line/variety/hybrid if grown over a longperiod, particularly in areas outside its adap-tation.
genic male sterility – the type of male sterilitygoverned entirely by the nuclear genes. It maybe transmitted by either the male or femaleparent.
germination – the resumption of growth by theembryo and development of the young plantfrom the seed. Germination, precisely, is theemergence and development from the seedembryo of those essential structures that, forthe kind of seed being tested, indicate theability to develop into a normal plant underfavorable conditions in the soil.
grain – the ripened ovary and its associated struc-tures.
Hheading (flowering), rice – growth stage of the
rice plant marked by the emergence of thepanicle from the boot followed by anthesis.
heritability – broadly, the proportion of observedvariance that is inherited, the remainder be-ing due to environmental effects. Strictly, theproportion of variance caused by the addi-tive effect of genes.
heterobeltiosis – refers to the phenomenon inwhich an F
1 hybrid obtained by the crossing
of two genetically dissimilar parents showssuperiority over the better parent in one or acombination of characters.
heterosis – refers to the phenomenon in which anF
1 hybrid obtained by the crossing of two ge-
netically dissimilar parents shows superior-ity over mid-parental values in one or a com-bination of characters.
heterosis (standard) – refers to the phenomenonin which the F
1 hybrid obtained by the cross-
ing of two genetically dissimilar parentsshows superiority over the best standard checkprevailing at that time in one or a combina-tion of characters.

80
heterosis breeding – a method of breeding to de-velop an F
1 hybrid obtained by the crossing
of two genetically dissimilar parents.heterozygote – an individual having different al-
leles for any gene pair and producing twokinds of gametes.
heterozygous – hybrid for any gene pair, with dif-ferent alleles for the gene being considered.
hill – a group of rice plants directly adjacent toeach other because the seeds or seedlings wereplanted together. A hill may also consist ofonly one plant.
hybrid – the product of a cross between geneti-cally dissimilar parents.
hybrid rice – the F1 seed of rice bred for commer-
cial use.hybrid vigor – increased vigor of the hybrid over
its parents in one or more characteristics.hybridization – a breeding method in which two
varieties are crossed to produce new variabil-ity and desired recombinants. The hybrids areallowed to self-pollinate and the segregatingpopulations are handled by an appropriatemethod.
Iinbred – an individual resulting from the mating
of closely related parents or by selfing.inbred line – a nearly homozygous line produced
by continued self-fertilization.inbreeding – the interbreeding of closely related
individuals occurring naturally (as in a closedpopulation) or as a deliberately chosen sys-tem of breeding and serving especially to pre-serve and fix desirable characters or to elimi-nate unfavorable characters from a suitablyselected stock but tending to bring about anunwanted decline (as in size, vigor, or fertil-ity) through the fixation of undesirable andoften recessive characters when the initialstock is in any way defective.
indoor growth cabinets – small indoor chamberswherein temperature, humidity, and light areartificially controlled.
intersubspecific hybrid – a cross between differ-ent subspecies of a crop. For example, in rice,hybrids between indica and japonica lines areconsidered as intersubspecific hybrids.
isolation – the separation of one group from an-other so that mating between or among groupsis prevented.
isolation (barrier) – the separation between twogroups can be provided by topography sur-face features or artificial/natural obstacles toa height of at least 2.5 m for rice.
isolation-free method – a method of producinghybrid seed for experimental purposes with-out isolation but by providing crop barriersof 2–4 rows of the restorer lines.
isolation (space) – separation is provided by keep-ing a certain distance between two groups. Aspace isolation of 50–100 meters is ideal forhybrid rice seed production.
isolation (time) – separation is provided by grow-ing two groups at different times of the cropseason so that one group is already mature(stopped providing pollen) when the othergroup reaches flowering. Generally, a periodof 21 days’ difference in flowering is suffi-cient for rice.
isoplasmic – these are the CMS or restorer linesdiffering in nuclear genetic constitution buthaving common cytoplasm.
Lleaf number – total number of leaves developed
on the main culm of a plant, which is a char-acteristic feature of each variety.
lodicules – the two scale-like structures adjoin-ing the base of the palea that control the open-ing of the lemma and palea during anthesis.
Mmaintainer line – a pollinator variety is used to
pollinate a CMS line and produce progeniesthat remain male sterile. If there is no main-tainer line, the male sterile line cannot bemaintained and multiplied generation aftergeneration.
male sterility – absence or nonfunction of pollenin plants.
mature grain stage (rice) – stage occurring dur-ing the ripening phase when the inside of thegrain is at first watery but later turns milky inconsistency.
milling yield – the estimate of the quantity ofhead rice and of total milled rice that can beproduced from a unit of rough rice. It is gen-erally expressed in percentage.
multilocation trial – yield trials conducted indifferent locations to study the adaptabilityof varieties/hybrids over environments.

81
Nnuclear genes – genes located on the chromo-
somes.nucleus – a small quantity of genetically pure seed
produced under the strict supervision of theplant breeder.
Ooff-type – the plants/seeds of the same crop devi-
ating significantly from the characteristics ofthe variety/hybrid as described by the breeder.
one-line breeding – this uses apomixis as a meansto fix the heterosis of F
1 hybrids into true-
breeding hybrids and is also known as theone-line breeding method.
outcrossing rate – the extent of cross pollinationmeasured on the basis of seed set to the totalnumber of spikelets.
outdoor growth cabinets – the small cabinets lo-cated outside where temperature and humid-ity are artificially controlled while light pro-vided is natural.
ovary – the bulbous basal portion of the pistilscontaining one ovule.
overdominance – superiority of the heterozygoteAa over either homozygote AA or aa.
Ppanicle – the terminal component of a rice plant
that bears the rice spikelets.panicle development – the growth stage of the
rice plant in which the spikelets become dis-tinguishable and the panicle extends upwardinside the flag-leaf sheath.
panicle exsertion – growth stage of the rice plantmarked by the emergence of the panicle fromthe boot.
panicle exsertion rate – the extent to which thepanicle is exserted out of the flag leaf.
panicle initiation (rice) – growth stage that startswhen the primordium of the panicle has dif-ferentiated and becomes visible.
partial restorer – a pollinator variety used topollinate a male sterile line to produce F
1 male
fertile progenies, which produce partial seedset upon selfing.
pedigree – the record of the ancestry of an indi-vidual or a cultivar.
pedigree nursery – a nursery consisting of segre-gating families in different generations de-rived from different crosses.
PGMS – photoperiod sensitive genic male sterileline. The genic male sterile plants that re-spond to the photoperiod or duration ofdaylength in terms of pollen fertility and ste-rility behavior.
phenotypic acceptability – breeders’ shorthandto record their observations on overall accept-ability of breeding lines or populations. Thiscan be done using an acceptability score of1–9. For example, 1 = excellent plant typeand absence of diseases. Promote to the nextlevel of testing and spread to other breedingprograms. 3 = very good appearance. Promoteto next level of testing. 5 = fair appearance,but has a few essential shortcomings (tooearly maturity, etc.). Use as parent in hybrid-ization block. 7 = poor appearance, but has afew important traits that make it suitable as adonor. Make a few crosses. 9 = poor. Discard.
photoperiod – duration of daylength.pistils – the female reproductive organ consisting
of the ovary, style, and stigma.plant growth substances – natural and synthetic
compounds that elicit growth and develop-mental or metabolic responses. These sub-stances are usually not metabolites in thesense that they are not intermediates or prod-ucts of the pathways they control, and theyare active at very low concentrations.
planting ratio – the ratio in which the male andfemale parental lines are planted to make acrossing block in hybrid seed production ormaintenance of the CMS line.
plumule – the leaves of the young plant in anyembryo It is enclosed by the coleoptile.
pollen – a mature reproductive male germ cell(microsporocyte) specialized for fertilization.
pollen fertility/sterility – the ratio of fertile/strerilepollen grains to the total pollen grainscounted in 3–4 fields under a microscope andexpressed in percentage. Fertility/sterility ofpollen grains is determined by their stainabil-ity with 1% IKI stain.
Pollen fertility/sterility gradiation
% sterile pollen Category % fertile pollen
0–20 Fully fertile 81–10021–40 Fertile 61–8041–70 Partially fertile 31–6071–90 Partially sterile 11–3091–99 Sterile 1–20
100 Completely sterile 0

82
pollen load – the amount of air-borne pollen perliter per hour at peak anthesis on a specifiedday.
pollen parent – male parent of a cross combina-tion.
pollination – transfer of pollen from the anther tothe stigma of a flower.
progeny – offspring; individuals resulting from amating.
pure line – a line that has been rendered almosthomozygous by repeated self-pollination overgenerations.
purity – the composition by weight of the samplebeing tested and, by inference, the composi-tion of the seed lot; the identity of variouskinds of seeds and inert matter constitutingthe sample.
Rrandom mating – a system in which every indi-
vidual plant in a population has an equalchance of becoming pollinated by any otherindividual.
randomization – allotting treatments to differentplots without any bias.
recurrent selection – a method of breeding de-signed to concentrate favorable genes scat-tered among several individuals by selectingin each generation among the progenies pro-duced by random mating of the selected indi-viduals (or their selfed progenies) of the pre-vious generation.
replication – repeating the experiment under iden-tical conditions with the objective of reduc-ing the experimental error.
restorer line – a pollinator variety is used to pol-linate the male sterile line to produce F
1 prog-
enies that are male fertile and thus produceseeds on selfing.
retestcross – a cross made beween a cytoplasmaticmale sterile line and a test variety (identifiedto be a restorer in the testcross) to recheck thepotentialities of the F
1 to give normal seed
set upon selfing.retestcross nursery – breeding nursery to evalu-
ate the retestcross F1s and corresponding male
parents.ripening phase (syn. maturity phase, grain-fill-
ing phase) – the period from pollination toharvest.
rogue – a variation from the standard of a varietyor strain. Roguing is the removal of undesir-able individuals to purify the stock.
row ratio – the proportion of seed parents andpollen parents planted to maintain cytoplas-mic male sterile lines or to produce F
1 hybrid
seed in a seed production plot.
Ssecondary tillers – tillers arising from primary
tillers.second leaf – the first differentiated leaf with blade
and sheath.seed – the fertilized and ripened ovule of a seed
plant comprising an embryonic plant accom-panied by a store of food (as endosperm orperisperm), enclosed in a protective seed coat,and capable under suitable conditions of in-dependent development into a plant.
seed dormancy – the ability of mature seeds todelay their germination after reaching physi-ological maturity.
seed parent – a female parent of a cross combina-tion.
seed viability – in general, the state of being alive;ability of the seed to germinate and producenormal seedlings.
seedbed – the bed on which rice seeds are sown,consisting of soil (wetbed method) or bananaleaves, plastic sheets, or concrete floor(“dapog method”).
seeding sequence – the order of seeding the pa-rental lines based on their growth duration sothat they reach flowering at the same time.
seedling (rice) – from seed germination to earlytillering; a juvenile plant.
self-fertilization – fusion of male and female ga-metes from the same individual.
source nursery – breeding nursery where all thegenetic material, including sources impart-ing cytoplasmic male sterility, genotypeswith specific traits useful for a hybrid breed-ing program, and elite rice lines showing highgeneral and specific combining ability, ismaintained for use in a hybrid breeding pro-gram.
spikelet – the basic unit of the rice inflorescenceconsisting of the two sterile lemmas, therachilla, and the floret.
spikelet fertility – the number of filled spikeletsto the total number of spikelets on a panicle.

83
sporophytic – in this system, sterility/fertility isimparted to the pollen by the mother plantupon which the pollen is borne and the geno-type of the pollen has no bearing per se. Itmay be controlled by more than one gene withmultiple alleles.
staggered planting – planting the restorer line ondifferent dates to maintain a uniform and regu-lar supply of the pollen to the spikelets of acytoplasmic male sterile line that continuesto bloom for a longer period.
stamen – the male reproductive organ consistingof the anther and the filament.
sterile – failing to produce or incapable of pro-ducing offspring.
stigma – the apex of the pistil of a flower, uponwhich pollen is deposited at pollination.
stigma exsertion rate – the proportion of spike-lets with exserted stigma (either on one or onboth sides) to the total number of spikelets ina panicle.
supplementary pollination – a method of shak-ing the male parent at the time of peak anthe-sis so as to disperse pollen grains to increasethe seed set on a CMS line. This is particu-larly necessary when the wind velocity is lessthan optimum (2–3 m sec–1).
synchronization (anthesis) – refers to the simul-taneous opening of the spikelets of the seedand pollen parents.
synchronization (flowering) – refers to the simul-taneous flowering of seed and pollen parentsdespite having different growth durations.
TTGMS (thermosensitive genic male sterile) line
– the genic male sterile plants that respond tothe temperature in terms of their fertility/ste-rility behavior.
testcross – a cross made between a cytoplasmicmale sterile line and a test variety to identifymaintainers and restorers.
testcross nursery – breeding nursery where F1 prog-
enies of cytoplasmic male sterile lines andtest varieties are screened for pollen sterility/fertility and spikelet fertility to identifymaintainers and restorers.
thermosensitivity – sensitivity of a genotype tovarying temperature regimes in terms of pol-len or spikelet sterility/fertility.
three-line breeding – a breeding strategy to de-velop hybrids uses three important lines—CMS lines (A lines), maintainer lines (B lines),and restorer lines (R lines)—in a two-step seedproduction system as follows: (1) CMS linemultiplication from an A × B cross in the fieldthrough natural outcrossing and (2) hybrid(A × R) seed production.
tiller – a vegetative branch of the rice plant com-posed of roots, culm, and leaves, which mayor may not develop a panicle.
tillering – growth stage of the rice plant that ex-tends from the appearance of the first tilleruntil the maximum number is reached.
topcross – a cross between a selection, line, clone,etc., and common pollen parent is called thetopcross of a tester parent.
two-line breeding – breeding methodology inwhich only two lines, a male sterile line (pho-tosensitive, thermosensitive, or chemicallyinduced) and a pollen parent, are used to pro-duce F
1 hybrids.
Uuniformity – the extent of similarity between the
individuals of a population.
Vvariance – the mean squared deviation of variet-
ies from their mean.vegetative phase – the period from germination
to panicle initiation.viability – the ability to grow and develop.vigor – the capacity for natural growth and sur-
vival, as of seed, plants, or animals.volunteer plants – unwanted plants growing from
the seed (may or may not be the same crop)that remains in the field from a previous crop.
Wwide compatibility – the ability of a genotype to
produce normally fertile progeny whencrossed with both indica and japonica testers.
wide hybridization – a process of crossing be-tween distantly related species.

85
Identifying male sterility
When rice plants start heading, male sterility isidentified with the following methods:
1. Visual inspectionAt the complete flowering stage, observe thecolor and plumpness of the anthers in themale sterile plants directly with the nakedeye. Shake the panicles slightly to examinethe dehiscence of anthers. Pay attention todetecting any pore dehiscence that occursat the basal part of the anthers. Male steril-ity in rice plants is expressed by anthers thatare whitish/pale yellow, shriveled, andnondehiscent.
2. Seed set on baggingWhen the plants just start heading but theirflorets have yet to flower, cover the panicleswith glassine paper bags to check whetherself seed setting has occurred. In practice,two panicles are bagged for each plant. Af-ter 25 days, observe the seed setting in thebagged panicles. If no seed is set, the plantis considered to be completely male sterile.When a few seeds (i.e., 5–20%) are produced,the plant is considered to be partially malesterile. The spikelet fertility percent is cal-culated as the number of filled spikelets di-vided by the total number of spikelets (i.e.,filled and unfilled spikelets) per panicle ona per plant basis multiplied by 100.
3. Pollen viability study under themicroscopeCollect five spikelets at the time of flower-ing and squash the anthers in 1–2 drops of1% acetocarmine or 1% IKI (iodine potas-sium iodide) stain. When the plant just starts
heading but its flowers have yet to flower,sample the five apical spikelets and immersethem in a tube with a screw cap containingfixative solution (alcohol:acetic acid, 3:1)and store them at room temperature. Formicroscopic observation under the light mi-croscope, rinse the spikelets with distilledwater, place the anthers from three or fivespikelets on a slide, and crush them on theslide with a drop of 1% IKI stain. Observa-tions for each microscopic field must havemore than 60 pollen grains and be averagedfor five different fields. The fertile pollengrains will be spherical and darkly stained,whereas the sterile pollen grains will be ei-ther unstained and spherical or unstainedand irregular in shape. Pollen fertility per-cent is calculated as the number of stainedspherical pollen grains with normal shapeand size to the total number of pollen grainsexpressed in percentage. Many of the EGMSlines in the completely sterile phase, that is,under very high-temperature or long-pho-toperiod conditions, become pollenless andonly the anther bag remains.
Classifying male sterility accordingto the morphology of sterile pollen
1. Typical abortion typeThe pollens are irregular in shape; some aretriangular, some are shuttle shaped, etc.They are unstained with IKI solution. Pol-len abortion occurs mainly at the one-nucleus stage. So, this type is also calledthe uninucleate abortion type. The CMSlines of the WA type correspond to this type.
Appendix I

86
2. Spherical abortion typePollens are spherical and unstainable withIKI solution. Pollen abortion occurs ap-proximately at the two-nuclei stage. So, thistype is also called the binucleate abortiontype. The Hong-Lian type CMS lines arerepresentative of this type.
3. Stained abortion typePollens are spherical, but partially or lightlystained with IKI solution. Pollen abortionoccurs mainly at the three-nuclei stage, sothe trinucleate abortion type is its othername. The boro-type CMS line are includedin this type.
4. No-pollen typesThere are no pollen grains, indicating thatabortion occurs even before the pollenmother cell formation stage. An example isSA2.
5. Antherless typeUnder extreme cases, this has been observedin certain EGMS lines with no anthers at all,leaving the female organs functional andintact.
For most EGMS lines, fertility to sterility al-teration behavior has been well documented, es-pecially for different pollen types with the gradualchange in environmental conditions. When anEGMS line is placed during its sensitive phaseunder extremely high-temperature (e.g., >35 ºC)or long-photoperiod (15 h) conditions, no-pollentypes and typical abortion types are observed.While under relatively less higher temperature nearits CSP level (e.g., >32 ºC) or photoperiod >14 h,the unstained spherical and abortive types areobserved. And with a further lowering of tempera-ture or photoperiod, the fertile stained sphericalpollen types and unstained spherical pollen typesare observed. As the EGMS lines experience theenvironment that approaches the CFP level, thatis, <24 ºC or <13.5 h, the fertile stained sphericalpollen grains are observed in a higher percentage.This gradual change can be observed with chang-ing environmental conditions within an EGMSline by collecting regularly the pollen grains uponflowering from subsequent flowering panicles.

87
Protocol for anther culture
This section is based on Xiang et al (1993), Li etal (1994a, 1995), and Pan et al (1993).
1. Young panicles at the meiosis stage asplant material for in vitro irradiation (0.5to 3.0 KR). The panicles were pretreatedfor 7 d under 6–10 h of irradiation, fol-lowed by incubating anthers on M8 me-dium with 2 mg L–1 2,4-D, 2 mg L–1 NAA,1 mg L–1 KT, and 5% sucrose.
2. Select and plate the anthers in N6 mediafrom the F
1 hybrids of TGMS × non-TGMS
or PGMS × non-PGMS (Chu et al 1975) orin M8 media (Mei et al 1988).
3. To obtain a higher success rate of usefuldihaploids, a large number of anthers mustbe plated in N6 or M8 media for more than5,000 petriplates, with 100 anthers on eachpetriplate (6 cm). Callus initiation is <7%and the regeneration rate is <20%.
4. The callus obtained must be tried for theregeneration of plants.
5. Regenerated plants should be hardened in1/2 MS liquid media followed by activatedcharcoal-sterilized soil medium, adoptingstandard procedures.
6. Regenerants should be taken to the fieldand raised under special fertility-condu-cive conditions.
7. Vigorous-looking plants that may bedihaploids should be selected and theirseeds kept separate. Weak plants shouldbe treated with colchicine for chromosomedoubling since they may be haploids.
8. Seeds obtained should be sown and raisedunder sterility-conducive conditions forthe selection of male sterile lines.
9. Sterile plants should be ratooned andbrought to low temperature or shorter pho-toperiod for inducing fertility to get selfseeds.
10. TGMS and PGMS lines should be screenedbased on the criteria for ideal EGMS lines.
11. Four TGMS lines, 6442S, 1286S, HS-1,and HS-5, and one PTGMS line, Lu Guang2S, have been developed through antherculture and extensively tested in Jiangxi,Sichuan, and Fujian provinces of China.
Appendix II
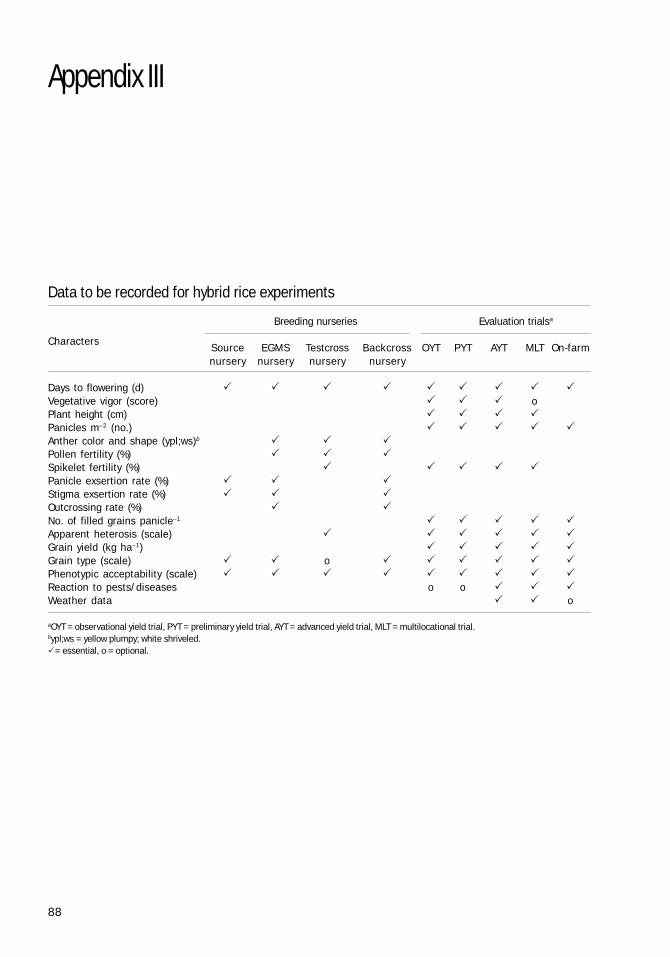
88
Appendix III
Data to be recorded for hybrid rice experiments
Breeding nurseries Evaluation trialsa
Characters Source EGMS Testcross Backcross OYT PYT AYT MLT On-farmnursery nursery nursery nursery
Days to flowering (d) ! ! ! ! ! ! ! ! !Vegetative vigor (score) ! ! ! oPlant height (cm) ! ! ! !Panicles m–2 (no.) ! ! ! ! !Anther color and shape (ypl;ws)b ! ! !Pollen fertility (%) ! ! !Spikelet fertility (%) ! ! ! ! !Panicle exsertion rate (%) ! ! !Stigma exsertion rate (%) ! ! !Outcrossing rate (%) ! !No. of filled grains panicle–1 ! ! ! ! !Apparent heterosis (scale) ! ! ! ! ! !Grain yield (kg ha–1) ! ! ! ! !Grain type (scale) ! ! o ! ! ! ! ! !Phenotypic acceptability (scale) ! ! ! ! ! ! ! ! !Reaction to pests/diseases o o ! ! !Weather data ! ! o
aOYT = observational yield trial, PYT = preliminary yield trial, AYT = advanced yield trial, MLT = multilocational trial.bypl;ws = yellow plumpy; white shriveled.!= essential, o = optional.





























