Embed Size (px)
Citation preview

ESTUDO DE CARACTERIZAÇÃO MOLECULAR DO VÍRUS VARICELA ZOSTER
EM MANAUS, AMAZONAS
DANIELE DE ARAÚJO SAMPAIO
MANAUS
2015
UNIVERSIDADE DO ESTADO DO AMAZONAS FUNDAÇÃO DE MEDICINA TROPICAL DOUTOR. HEITOR VIEIRA DOURADO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL MESTRADO EM DOENÇAS TROPICAIS E INFECCIOSAS

i
DANIELE DE ARAÚJO SAMPAIO
ESTUDO DE CARACTERIZAÇÃO MOLECULAR DO VÍRUS VARICELA ZOSTER
EM MANAUS, AMAZONAS
Dissertação apresentado ao Programa de Pós-Graduação em Medicina Tropicalda Universidade do Estado do Amazonas em Convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, para obtenção do título de Mestre em Doenças Tropicais e Infecciosas.
Orientador (a): Profª Dra. Maria Paula Gomes Mourão
Co-orientador (a): Dra. Michele de Souza Bastos
MANAUS 2015

ii
Ficha Catalográfica
S192 Sampaio, Daniele de Araújo.
Estudo de caracterização molecular do vírus varicela Zoster em Manaus,
Amazonas ./Daniele de Araújo Sampaio. -- Manaus : Universidade do Estado do
Amazonas, Fundação de Medicina Tropical, 2015.
xiii, 53f. : il.
Dissertação (Mestrado) apresentada ao Programa de Pós- Graduação
em Medicina Tropical da Universidade do Estado do Amazonas –
UEA/FMT, 2015.
Orientadora: Profa. Dra. Maria Paula Gomes Mourão
Título em inglês: Molecular characterization study of varicella zoster virus
in Manaus Amazon.
1.VZV 2 Varicela. 3.Genótipo 4. Manaus Título.
CDU: 616.9(811.3)(043)
Ficha Catalográfica elaborada pela Bibliotecária Maria Eliana N Silva,lotada na Escola Superior de
Ciências da Saúde - UEA

iii
ESTUDO DE CARACTERIZAÇÃO MOLECULAR DO VÍRUS VARICELA ZOSTER
EM MANAUS, AMAZONAS
DANIELE DE ARAÚJO SAMPAIO
“Esta Dissertação foi julgada adequada para obtenção do Título de Mestre em Doenças Tropicais e Infecciosas, aprovada em sua forma final pelo Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas em convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado”.
Banca Julgadora:
Maria Paula Gomes Mourão
Presidente
RajendranathRamasawmy
Membro
Felipe Gomes Naveca
Membro

iv
AGRADECIMENTOS
Agradeço a Deus por sua proteção e direcionamento para alcançar esta
conquista.
À minha família, pelo apoio e confiança para que eu concretizasse este
projeto. Minha mãe, meu pai e meus irmãos, por priorizarem minha educação acima
de tudo e por acreditarem em meus sonhos, entendendo-me nos momentos de
ausência, dando-me apoio e carinho.
Ao professor Dra. Maria Paula Gomes , minha orientadoar, por todos os
ensinamentos e paciência ao longo desses anos, contribuindo de maneira intensa no
meu aprendizado durante esta jornada.
À professora Dra. Michele Bastos, minha co-orientadora que me possibilitou
aprendizagens únicas, sempre me incentivando. Muito obrigada pela confiança e por
acreditar em meu potencial.
Ao professor Dr. RajendranathRamasawmy pela contribuição na construção deste
trabalho, incentivo e ensinamentos incansáveis.
Ao professor Dr. Felipe Naveca pela contribuição na construção deste
trabalho, incentivo e ensinamentos incansáveis.
A todos os professores Universidade do Estado do Amazonas/Programa de
Pós-graduação em Medicina Tropical, por todo aprendizado e atenção dispensado
no repasse dos conhecimentos durante as aulas e extra sala de aula.
À Fundação de Amparo à pesquisa di Amazonas(FAPEAM)pelo apoio
financeiro por meio de bolsa de estudo.

v
Aos membros da secretaria acadêmica da Pós-graduação em Medicina
Tropical e a todos os funcionários da Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado, pelo apoio e incentivo.
A todos os funcionários do da gerencia de virologia a Gerente Marcia
Castilhoe toda sua equipe, pela atenção e tempo disponibilizados.
Aos colegas da turma de Mestrado ano 2014, por todo aprendizado e
amizade, durante esta jornada de estudo.
Muito obrigada !

vi
RESUMO
O Virus varicela zoster (VZV) pertence a família Herpesviridae, sua espécie é conhecida como humano - 3 (HHV-3) e causa duas doenças distintas conhecidas: catapora (varicela) e herpes zoster. Na maioria dos países, entre 2 e 4% das crianças são hospitalizadas por complicações da varicela.O VZV pode causar algumas complicações no Sistema Nervoso Central (SNC), já em pacientes com AIDS existe um total de 71% dos casos, e na sua maioria causa encefalite.O VZV foi inicialmente classificado como neurotropico, mas experimentos ultilizando células T em animais mostra o tropismo para células T importantes para replicação e liberação de viriões.Epidemias são mais prevalentes em climas temperados, que ocorre na maioria das vezes durante o inverno e primavera. Isolante de varicela e HZ pode ser estudado em cinco genótipos distintos de VZV de áreas geográficas específicas, o domínio regional de genótipos específicos pode ter sido estabelecido pelo clima e outros fatores.O objetivo desse projeto é realizar estudo de caracterização molecular do vírus Varicela Zoster detectado em amostras clinica de pacientes atendidos em uma unidade terciaria de saúde no Amazonas. Foram incluidas 12 amostrasidentificadas no período de 2010 a 2014, na Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT-HVD), para a triagem foram aplificadas usando a região ORF8 e para o sequenciamento foi amplificado as regiões ORF22, 38 e 54. Todas as amostras foram sequenciadas e feito a análise filogenética . Seis amostras foram identificadas as cepas selvagem do VZV. Na região da ORF54 em umas das amostras foi observado mudanças na posição 95241 o que não descarta a possibilidade de mutações. E o clado 5 foi sugerido para duas amostras da região ORF22.
PalavrasChaves:VZV, varicela, genótipos, Manaus.

vii
ABSTRACT
VZV belongs to the Herpesviridae family, its kind is known as human - 3 (HHV-3) and causes two distinct diseases known: chicken pox (varicella) and shingles. In most countries, between 2 and 4% of children are hospitalized for complications of varicela.O VZV can cause some complications in the central nervous system (CNS), in AIDS patients there is a total of 71% of cases, and its Most cause encephalitis. The VZV was initially classified as a neurotropic but ultilizando T cell experiments on animals show tropism for T cells important for replication and release of virions. Epidemics are more prevalent in temperate climates, which occurs most often during the winter and spring. sample isolatedof chickenpox and HZ can be studied in five different genotypes of VZV specific geographic areas, the regional domain specific genotypes may have been set by the climate and other factors. The objective of this project is to conduct study of molecular characterization of the Varicella Zoster virus detected in clinical samples from patients treated in a tertiary health unit in the Amazon. Specifics are to identify the current VZV genotype in Manaus, make comparison study with viral strains from other regions and determine whether there is a relationship with severe disease according to genotype identify and differentiate the strains of wild-type and vaccine .Included were 12 samples, Included 12 samples were identified in the period 2010 to 2014 , at the Fundação de Medicina Tropical Dr. Heitor Vieira Dourado( FMT - HVD ) for screening DNA samples were amplified using the ORF8 region and the sequencing was amplified the regions Orf22 38 and 54. All samples were sequenced and made phylogenetic analysis. Six samples of the wild strains of VZV have been identified. In the region of ORF54 in one of the samples was observed changes in position 95241 which does not rule out the possibility of mutations. Clade 5 and has been suggested for the two samples ORF22 region.
Keywords:VZV , varicella , genotypes , Manaus.

viii
RESUMO LEIGO
O Virus varicela zoster (VZV) pertence a família Herpesviridae, que pode ser de origem selvgem (vírus VZV) ou vacinal (Oka vacina) humano - 3 (HHV-3) e causa duas doenças conhecidas como catapora e herpes zoster. Na maioria dos países, entre 2 e 4% das crianças são hospitalizadas por complicações da varicela. Esse vírus pode causar algumas complicações no Sistema Nervoso Central (SNC), sendo as mais comuns a inflamação e a infecção no cérebro. É um virus que apresenta propensão para infectar células que são importantes para processo de duplicação do DNA e liberação de outros vírus. Epidemias são mais prevalentes em lugares de clima temperados. O VZV pode ser estudado em cinco genótipos distintos, conforme as áreas geográficas específicas, o domínio regional de genótipo, que são os genes específicos, pode ter sido estabelecido pelo clima e outros fatores. O objetivo dessa dissertação foi realizar estudo de caracterização molecular do vírus Varicela Zoster detectado em amostras de vesíscula de pacientes atendidos em uma unidade terciaria de saúde no Amazonas. Foram incluidas 12 amostras identificadas no período de 2010 a 2014, na Fundação de Medicina Tropical Dr. Heitor Vieira Dourado(FMT-HVD); para a triagem foi extraído o DNA e aplicadas técnica biomoleculares, e para o sequenciamento e feita análise filogenética. Em 6/12 (50%) das amostras identificou-se as cepas do vírus VZV. Na região da ORF54 em umas das amostras foi observado mudanças na posição 95241 o que evidencia a possibilidade de mutações. E duas amostras 5 e 7 da região ORF22 teve um agrupamento que inclui um ancestral comum classificado nos genótipos do VZV como clado 5.
PalavrasChaves:VZV, varicela, genótipos, Manaus.

ix
LISTA DE FIGURAS
Figura 1 Estrutura do vírus Varicela............................................................................................ 17
Figura 2 Organização do genoma do vírus da varicela-zoster.................................................... 20 Figura 3
Sistema de nomenclatura do VZV, adotado em 25 de julho de 2008, Whitechapel, Londres, Reino Unido................................................................................................... 21
Figura 4 Árvore filogenética mostrando os cinco grandes clades VZV...................................... 22
Figura 5 Mapa da distribuição geográfica dos clades do VZV................................................... 28
Figura 6 Representação da patogênese do VZV....................................................................... 24
Figura 7 Imagem do gel de agarose dos fragmentos de 275 pb da região ORF8 do VZV........ 34
Figura 8
Imagem do gel de agarose dos fragmentos de 447, 350 e 222pb das egiões ORF22, 38 e 54 respectivamente do VZV................................................................... 34
Figura 9 Imagem do gel de agarose do resultado da purificação com Polietilenoglicol (PEG).......................................................................................................................... 35
Figura 10 Imagem do gel de agarose do resultado da purificação com KIT.............................. 35
Figura 11 Representação do resultados das sequencias das regiões ORF 22, 38 e 54 alinhadas com a cepaDumas...................................................................................................... 37
Figura 12 Representação do ORF38 analizadas pela enzima de restrição PstI........................ 38
Figura 13 Representação do ORF54 analizadas pela enzima de restrição BglII. O (Y) significa a presença de (C) ou (T )............................................................................................. 39
Figura 14 Arvore filogenética do VZV baseada na incluindo as amostras 05 e 07....................... 41

x
LISTA DE TABELAS E QUADROS
Tabela 1 Tabela de descrição taxonômica do VZV..................... 15
Tabela 2 Tabela de primers usados para PCR........................... 31
Tabela 3 Tabela de primers usados para PCR para genotigem. 31
Tabela 4 Tabela de analise de variações do genótipo do VZV.... 40

xi
LISTA DE ABREVIATURAS, SÍMBOLOS E UNIDADES DE MEDIDA
˚C Graus Celsius A Adenina AIDS Síndrome da Imunodeficiência Adquirida C Citosina CEP Comitê de Ética e Pesquisa CMV Citomegalovírus DNA Ácido desoxirribonucleico EBV Epstein-Barr EUA Única Região Curta FAPEAM Fundação de Amparo a Pesquisa do Estado do Amazonas FMT-
HVD Fundação de Medicina Tropical Doutor Heitor Vieira Dourado G Guanina HHV Herpes Vírus Humano HIV Vírus da Imunodeficiência Humana HSV Herpes Simples Vírus HZ Herpes Zoster ICTV International Committee on Taxonomy of Viruses IRL Repetições Internas Longas IRS Repetições Internas Curtas KSHV Herpes Vírus Humano associado ao sarcoma de Kaposi Nm Nanômetro Nº Número ORF Open Reading Frame(Fase de leitura aberta) PCR Polymerase chain reaction (Reação em cadeia da polimerase) Pb Base-pair R Região RFLP Restriction Fragment Length Polymorphism SNC Sistema Nervoso Central SNP Single Nucleotide Polymorphism T Timina (T) Triangulação TRL Terminais Longos TRS Terminais Curtos UL Região Longa VZV Vírus de Varicela Zoster Α Alfa Β Beta Γ Gama

xii
SUMÁRIO
1 INTRODUÇÃO.......................................................................................................... 14
1.1A família Herpesviridae.......................................................................................... 14
1.2 Varicela Zoster..................................................................................................... 15
1.2.1 Estrutura do virion VZV..................................................................................... 16
1.2.1.1 Proteínas do virion........................................................................................... 17
1.2.1.2 Tegumento....................................................................................................... 18
1.2.1.3 Capsídeo.......................................................................................................... 18
1.2.1.4 Envelope.......................................................................................................... 18
1.2.1.5 Core.................................................................................................................. 19
1.2.2 Genoma do VZV.................................................................................................. 19
1.2.2.1 Genes do VZV.................................................................................................. 20
1.2.2.2 Filogenia e Genótipos VZV.............................................................................. 21
1.3 Complicações no Sistema Nervoso Central (SNC) do VZV................................... 22 1.4 Tropismo..................................................................................................................
23 1.5 Vacina Contra Varicela............................................................................................
25 1.6 Epidemiologia..........................................................................................................
26 1.7 Epidemiologia Molecular.......................................................................................
26
2 OBJETIVOS............................................................................................................ 29 2.1 Geral ......................................................................................................................
29 2.2 Específicos ............................................................................................................ 29
3 MATERIAIS E MÉTODOS......................................................................................... 30
3.1 Modelo de Estudo................................................................................................... 37
3.2 Seleção das amostras........................................................................................... 37
3.3 Método de diagnóstico molecular.......................................................................... 30
3.3.1 Detecção do genoma viral da família Herpesviridae........................................... 30
3.3.1.1 Extração de DNA............................................................................................. 30
3.3.1.2 Amplificação do DNA PCR.............................................................................. 30
3.3.2 Sequenciamento de nucleotídeo....................................................................... 31
3.3.2.1 Amplificação do DNA PCR............................................................................... 31
3.3.2.1.1 Purificação do produto da PCR..................................................................... 32

xiii
3.3.3 Genotipagem do vírus VZV ................................................................................ 32
3.3.4 Sequeciamento e analise filogenética................................................................. 32
3.4 Aspectos éticos....................................................................................................... 33
3.5 Financiamento........................................................................................................ 33
4 RESULTADOS.......................................................................................................... 34
4.1 Seleção de amostra e identificação do VZV.......................................................... 34
4.2 Amplificação do DNA para sequenciamento ........................................................ 34
4.3 Cronograma de Execução.................................................................................... 35
5DISCUSSÃO............................................................................................................. 42
6CONCLUSÃO............................................................................................................ 44
7 REFERÊNCIAS......................................................................................................... 45
8 ANEXOS.................................................................................................................... 55

1
1 INTRODUÇÃO
1.1 A FAMÍLIA Herpesviridae
Os virus já são patógenos claramente conhecidos e que estão relacionados com o
homem e outras espécies há mais de 200 milhões de anos1.Mais de 300 diferentes
herpesvírus foram descobertos até agora, alguns pela sua associação com doenças e
outros usando a Reação em Cadeia da Polimerase (PCR)8.
Existe um agrande variação de vírus, devido à ampla ocorrência e diversidade, de
acordo com o International Committee on Taxonomy of Viruses-ICTV2 (2015) a família
Herpesviridaeé classificadaem 3 subfamílias: Alphaherpesvirinae, Bethaherpesvirinae e
Gammaherpesvirinae10.Esses membros têm como hospedeiros os répteis, aves e
mamíferos13 (Tabela 1).
Dentro da família Herpesviridae existem 8 espécies de vírus que podem infectar o
ser humano. A subfamília Alphaherpesvirinaeque inclui os Herpesvírus Simples tipo I e
II(HSV-1 e HSV-2), responsáveis, respectivamente, pela infecção damucosa labial e
genital e são vulgarmente conhecidos como Herpes. Essasubfamília inclui também o
gênero Varicellovirus contendo a espécie Herpesvirus humano - 3 (HHV-3), que causam
duas doenças conhecidas como catapora e herpes zoster9-10 (Tabela 1).
A segunda subfamília Bethaherpesvíruscompreendeao gêneroCytomegalovirus
ouCytomegalovirus Humanoque tem como espécie Herpesvirus humano-5 (HHV-5), que
na infecção primária está associada à síndrome da mononucleose. E nessa mesma
subfamília inclui o gênero Roseolovirus com a espécieHerpesvirus Humano-6A, 6B e 7
(HHV6, 6B e 7), responsáveis pela doença infantil infecciosa roséola9-10,3 (Tabela 1).

2
A subfamília Gammaherpesvirinae apresenta o gêneroLymphocryptoviruse sua
espécieé o Herpesvirus Humano-4 também conhecido comoEpstein-Barr (EBV), agente
etiológico da infecção popularmente conhecida como doença do beijo ou mononucleose
infecciosa, além de estar envolvido na patogênese de alguns tumores, como o linfoma de
Burkitt e o carcinoma nasofaringeal, eo gênero Rhadinovirustem, a espécie Herpesvirus
Humano-8 (HHV-8 ou KSHV), associado ao sarcoma de Kaposi 9-10(Tabela 1).
Tabela1 –Tabela de descrição taxonômica do VZV2
Fonte: www.ictvonline.org (2015).
1.2 Varicela Zoster
O Vírus Varicela Zoster (VZV) causa duas doenças distintas conhecidas:
varicela (catapora) e herpes zoster. A nomenclatura aprovada pelo ICTV2 é
Herpesvirus humano - 3 (HHV-3).
Não se sabe ao certo a origem da palavra catapora, mas uma possível
derivação do termo catapora é do antigo Inglês “gican” designando-os pox "coceira"
para diferenciá-lo de doenças como a varíola. As palavras zoster e herpes são
derivadas do grego e latin, que significa, respectivamente, um cinto ou cinta, e é,
obviamente, uma característica relacionada distribuição das erupções no corpo do
hospedeiro1.
Família Subfamília Gêneros Espécies
Herpesviridae
Alphaherpesvirinae
Simplexvirus Human herpesvirus 1 Human herpesvirus 2
Varicellovirus Human herpesvirus 3
Bethaherpesvirinae
Cytomegalovirus Human herpesvirus 5
Roseolovirus
Human herpesvirus 6A Human herpesvirus 6B Human herpesvirus 7
Gammaherpesvirinae Lymphocryptovirus Human herpesvirus 4

3
Historicamente, a relação entre as etiologias de varicelae herpes zoster foi
sugerida pela primeira vez por von Bo'kay em1892, a partir de observação em
crianças que após a exposição a um adulto com Herpes Zoster desenvolveram a
Varicela14.
A varicela (catapora) é a manifestação da infecção primária com VZV e é
mundialmente umas das doenças mais conhecidas1, frequente na infância associada
à febre euma erupção vesicular, e estabelece latência nas células dos gânglios da
raiz dorsal, após a infecção primária4. O VZV é o único entre os herpes vírus
humano queinfecta através da lesão cutânea de pessoas infectadas e atinge o trato
respiratório por inalação de partículas virais,causando diversas complicações e risco
a vida do hospedeiro5-7.Estudos em crianças com varicela e que desenvolveram
trombocitopenia mostram que é consequência da destruição das plaquetas através
da resposta imune. Trombocitopenia durante varicela aguda está associada com
sangramento em lesões de pele, petéquias, púrpura, epistaxe, hematúria e
hemorragia gastrointestinal4,81.
Herpes zoster é uma manifestação de reativação do vírus VZVque antes
estava latente no sistema nevos central e, embora seja uma doença que raramente
ameaça a vida, a maior preocupaçãoda população épor causa da dor que ela causa,
não só devido à lesão aguda, mas também aneuralgia pós-herpética, a qual pode
ser muito debilitante e é notoriamente difícil de tratar1.
O vírus quando reativado pode se espalhar com um padrão centrípeto (ou
seja, em direção à medula espinhal ou o cérebro), um padrão centrífugo (isto é, em
direção à pele), ou ambos. A detecção do vírus em neurônios, oligodendrócitos,
células meníngeas, células ependimais, ou na parede do vaso sanguíneo exige
muitas vezes uma combinação de características morfológicas, imuno-histoquímica,
hibridação in situ, e PCR. A análise de PCR do líquido cefalorraquidiano continua a
ser o esteio para o diagnóstico das complicações neurológicas de VZV durante a
vida59.

4
1.2.1 Estrutura do virion VZV
A morfologia do virion de VZV pode ser vista usando microscopia eletrônica
tradicional, contendo um diâmetro médio de 150-200 nanômetros1,11.O virion é
formado por quatro elementos essenciais: umenvelope lipídico esférico que
apresenta glicoproteínas embutido, com o número de triangulação simetria
icosaédrica (T) igual a 16 icosahedral proteína do capsídeo, um núcleo (core)que
contém o DNA genômico de fita dupla linear, umcápsideque consiste em 162
capsômeros e um tegumento amorfo que rodea o capsídeo. As glicoproteínas
complexas são incorporados no envelope lipídico11,17. (Figura 1).
Figura 1. Estrutura do vírus Varicela. Fonte: Adaptado de Quinlivan;Breuer, 200563.
1.2.1.1 Proteínas do vírion
O VZV induz as células a produzirem cinco ou mais glicoproteínas, que são
conhecidas como gB (gp II), gC (gp IV), gE (gp I), gH (gp III) e gL, elas envolve o
envelope lipídico e durante a replicação viral são expressas nas membranas da
célula, permitindo que o vírus possa interagir com o seu ambiente.1,20
O grupo das glicoproteínas gB (gp II), induzem a artuculação do sistema
complemento dependente e independente de anticorpos neutralizantes, além de ser
a segunda glicoproteína mais abundante e imunogênicas do envelope do VZV21-25.

5
As glicoproteínas gC (gp IV), são reativadas com anticorpos anti-peptideos.
Os genes para estas pequenas glicoproteínas estão localizado na região Us com 39
kDa ORF no centro da região do VZV31.
As glicoproteínas gE (gp I), são as mais freqüentes do envelope do VZV25-26.
Elas provocam a fativação dosistema complemento-dependente neutralizando
anticorpos21, 27-30.
O grupo das glicoproteínas gH (gp III), também induz na formação do sistema
complemento22,28-29, sendo é a terceira mais abundante das glicoproteínas do
envelope do VZV23.
As propriedades que já são conhecidas das glicoproteínas do VZV se
assemelham com as dos HSV e pode fornecer uma visão mais aprofundada dentro
das funções biológicas dos imunógenos20.
1.2.1.2 Tegumento
É uma estrutura amorfa localizada entre o capsídeo e o envelope do vírus.
Apresentaum complexo de massa proteica e contém enzimas que controlam a
replicação do vírus e regulação da função celular1.Os tegumentos de
alphaherpesviruses contem mais de 20 proteínas codificadas pelo vírus32-33.
Essas proteínas do tegumento têm amplas funçõesincluindo a regulação viral,
expressão do gene da célula hospedeira durante a replicação, transporte de viriões
para o núcleo pelo recrutamento de células motores durante sua entrada e saída, e
montagem da partícula viral infecciosa34.
1.2.1.3 Capsídeo

6
Existem três formas: A (nulo), B (intermediário) e C (maduro). Proteínas de
montagem presentes na forma B são perdidas durante a inserção de DNA para
produzir o C1.
Caracteristicamente, a cápside detodos vírus herpes têm um diâmetro de 100-
110 nm. Os 150 capsômeros hexaméricas são 9,5 x 12,5 nm na seção longitudinal,
um canal aberto de 4 nm de diâmetro corre via superfície ao longo do eixo longo36.
1.2.1.4 Envelope
O VZV é um vírus envelopado. Uma estrutura membranosa complexa
derivada de porções de membranas celulares alteradas1. O envelope doherpesvírus
tem uma aparência típica trilaminar37.
1.2.1.5 Core
O Core do VZV contém DNA genômico de fita dupla linear17, que tem uma
forma toróide38-39.A baixa frequência de virions que apresentam essa morfologia
pode-se considerar suaconfiguração espacial do core no interior docapsídeo15.O
core pode também explicar a variação na forma do núcleo eminúmeros estudos40-41.
1.2.2 Genoma do VZV
A sequência completa do DNA do VZV foi publicada pela primeira vez em
1986 por Davison e Scott e em uma revisão geral da biologia molecular do VZVque é
dada por Davison em 19911.
Como todos os herpesvírus, oVZV tem um DNA linear de fita dupla, o genoma
é de tamanho variável, mas a sequência que foi obtida compreende 124.884 pb e

7
contem pelo menos 72 estrutura de leituras abertas (open reading frame ORF) que
contém 70 genes distribuídos igualmente entre as duas cadeias de DNA12,.
As estimativas de densidade flutuante, bem como de dados de sequências,
mostra um conteúdo de guanina (G) +citosina (C) de 46%, o que é muito mais baixa
do que a maioria das herpesviroses como 67% em herpes simples 1 (HSV) e 58%
no DNA de citomegalovírus (CMV). Dentro do genoma existem, porém, regiões ricas
em (G+ C), notavelmente as regiões de repetição1,18.
O genoma consiste em umaregião única longa (UL) delimitada por terminais
longos (TRL), repetições internas longas(IRL),e uma região única curta (EUA)
delimitada por curto interno (IRS) e repetições de terminais curtos (TRS)12(Figura 2).
O mesmo tem cinco regiões de repetição. A região 1 (R1) está localizado na
fase de leitura aberta (ORF) 11, a R2 está localizado na ORF14 (glicoproteína C),a
R3 em ORF22, a R4 entre ORF62 e a origem de replicação viral, e R5 entre o
ORF60 e 61. O comprimento das regiões de repetição varia entre as diferentes
estirpes de VZV e tem sido utilizada para distinguir-las12.
Figura 2. Organização do genoma do vírus da varicela-zoste. Fonte: Quinlivan;Breuer, 200563.
Apesar da motivação para estudar este vírusde importância médica, o
conhecimento da biologia molecular do VZV é rudimentar em comparação com o dos
outros quatro vírus de herpes que infectam os seres humanos: vírus herpes simplex
tipo 1 (HSV-1) e tipo 2 (HSV-2 ), citomegalovírus e vírus de Epstein-Barr (EBV)12.

8
1.2.2.1 Genes doVZV
A partir da descrição do genoma referente ao VZV, sabe-se que codifica pelo
menos 70 genes, três (ORF62, 63, 64) estão presentes em ambas as regiões de
repetição curtas. As funções de muitas das proteínas que eles codificam já foram
caracterizadas e estão localizados no tegumento do virione regulam a transcrição
viral12.19.
Os genes que foram identificados, em sua maioria, por analogia com o HSV1,
encontram-se listados com todos os 70 genes35. Apêndice A.
Em comum com outras herpesviroses, a síntese de proteínas do VZV envolve
amplamente três fases designadas: início imediato (α) que estão localizados no
tegumento do virion e regulam a transcrição do vírus, inícial (β) e tardio (γ).
1.2.2.2 Filogenia e Genótipos VZV
Apesar da homogeneidade do genoma de VZV, estirpes deste virus foram
distinguidas por meio de análise de vários tipos de mutações que dão origem ou
eliminam os fragmentos de polimorfismode restrição estável (RFLPs), ou resulta em
alterações na composição das regiões de repetição variáveis. A identificação de
polimorfismos de nucleotídeo único (SNPs) também permitiu a classificação das
cepas VZV geograficamente relacionadas em distintos genótipos ou clados42.
Uma vez que a nomenclatura do VZV em clados-genótipos foi baseada em
diferentes métodos de tipagem molecular, uma nova nomenclatura universal foi
introduzida mais recentemente, separando os genótipos em cinco clades principais
(1-5) com dois clades provisórios (VI e VII)43(Figura 3).

9
Figura 3- Sistema de nomenclatura do VZV, adotado em 25 de julho de 2008, Whitechapel, Londres, Reino Unido.Fonte: Adaptado de JUDITH BREUER et al.,201043.
Isolados de varicela zoster e HZ podem ser estudado em cinco genótipo
distinto de VZV de áreas geográficas específicas: Clado 1, o genótipo C (E1 / A);
Clado 2, genótipo J (B); Clado 3, genótipo B (E2 A / D); Clado 4, J2 genótipo (M2 /
B); Clado 5, genótipo A1 (M1)45.
Usando esta nova nomenclatura, a rede filogenética de cepas VZV ilustra as
relações evolutivas entre diferentes clados. Para verificar a diversidade genética do
VZV, osdiferentes termosgenótipode clado têm sido utilizados na literatura. Embora
a definição de genótipo refere-se a determinados alelos em loci especificados, e
clado significa um único “ramo” na árvore filogenética,44 e essa rede de filogenia do
VZV é baseada nas sequencias completas já identificadas (Figura 4).
Até o presente momento, a maioria dos dados de genotipagem foram na
análise deapenas alguns loci no genoma de VZV, e apenas 23 sequências do
genoma completotêm sido relatadas44.Referências de Genotipagem do VZV
foramfornecidas por vários países,o que estabeleceu de uma nomenclatura comum
que facilitará o intercâmbio de informações43.

10
Figura 4– Árvore filogenética mostrando os cinco grandes clades VZV.Arvore filogenética do VZV
baseada em sequencias completas. Clado 1 Incluem 10 cepas: referencia Dumas (NC001348), Netherlands; BC(AY548171), British Columbia, Canada; 36 (DQ479958), New Brunswick, Canada;49 (DQ479959), New Brunswick, Canada; referencia cepa MSP (AY548170),Minnesota, EUA; 32p5 (DQ479961), Texas, EUA; Kel (DQ479954), Iowa, EUA; SD(DQ479953), South Dakota, EUA; NH293 (DQ674250), EUA; SVETA (EU154348),Russia; clado 3 incluem 4 cepas: referencia 03-500 (DQ479957), Alberta,Canada; 11 (DQ47995), New Brunswick, Canada; 22 (DQ479956), New Brunswick,Canada; reference strain HJO (AJ871403), Germany; clado 2 incluem um cepa: referencia strain pOka (AB097933), Japão; clado 5 incluem 5 cepas referência CA123strain (DQ457052), California, USA; clado4 incluem 2 cepas: referencia 8(DQ479960), New Brunswick,
Canada and referencia DR (DQ452050), EUA.Fonte: JONAS SCHMIDT-CHANASIT et al.,201144.
1.3 Complicações no Sistema Nervoso Central (SNC) causado pelo VZV
O vírus varicela zoster pode causar algumas complicações no Sistema
Nervoso Central (SNC) que incluem: ataxia cerebelar, meningoencefalite, mielite
transversa e meningite asséptica, em crianças são raras, ocorrem em menos de 1%,
devido aos programas de vacinação em todo o mundo14.Sintomas causados pelo
VZV mais comuns são manifestados em meningite asséptica, meningoencefalite e
encefalite49,61.Quadro 1.
Os seres humanos são os únicos reservatórios para esses vírus.
Epidemiologicamente, HHV pode causar infecçõesem lactentes, crianças ou
adultos55. Em indivíduos comSíndrome da Imunodeficiência Adquirida (AIDS)
quando o vírus da imunodeficiência humana (HIV)já danificou o sistema imune
celular o herpesvírus no período latente pode ser reativado e afetar posteriormente a
progressão da doença e sobrevidacausando uma doença oportunista58.

11
As manifestações neurológicas predominantesdo VZV em pacientes com
AIDS são concomitantecom a erupção por herpes zoster em 71% dos casos,
encefalite (37%), mielite(24%), radiculite (21%), meningite (17%), enecrose aguda de
retina (12%)60.
Os herpesvírus apresentam tropismo pelo sistema nervoso central, seja por
disseminação hematogênica ou transmissão neuronal56.A característica mais
importante do HHV é sua latência no tecido neuronal antes de serem reativado por
algum estímulo57. A maioria dos pacientes que têm varicela, mesmo aquelescom
complicações neurológicas, recuperam-se da doença16.
1.4 Tropismo
A resposta adquirida é importante durante a infecção do VZV, mas a resposta
inata que faz o controle da replicação viral, contribui significativamente na
persistencia do virus na população83.
A resposta celular inata faz parte da regulação patogênica da pele. Na
infecção do VZV é formada lesões na pele ao longo de 10 – 21 dias, considerado o
período de incubação da varicela82.
O VZV foi inicialmente classificado como neurotropico, mas experimentos
ultilizando células T em animais mostra o tropismo para células T importantes para
replicação e liberação de viriões84-66.
O VZV Infecta o hospedeiro atingindo a mucosa. A replicação local é seguida
de propagação nas amídalas onde o VZV ganha acesso as células T. As células T
infectadas seguem aos locais cutâneos de replicação. O VZV estabelece latência
nos glânglios sensoriais. A reativação da latência permite a segunda faze da
replicação ocorrendo na pele83.

12
Figura 6.Representação da patogênese do VZV.
Alguns estudos com xenoenxertos mostram a característica importante do
tropismo do VZV. As células T infectadas não se unem as células T não infectadas,
por isse motivo elas formam viriões completos e liberam os virus infecciosos87,88.
O VZV tem a habilidade em infectar células T de memória sem interromper
seu fluxo normal, por isso as lesões são desenolvidas, assim essas células são
consideradas células associadas a viremia89-91(Figura 6).
Em geral compreender o princípio da patogenese do VZV ao nível molecular
tem um potencial para produzir novas estratégias para tratar e previnir a infecçãodo
VZV83.
1.5 Vacina Contra Varicela
Takahashi et. al. em 1970 foi o primeiro a desenvolver e testar a vacina contra
varicela a partir de vírus atenuados de estirpes Oka de VZV. Essa reprodução foi
feita a partir do métodopadrão da época:o víru foi isolado e passado em fibroblastos

13
de embrião de porco-da-índia e expandido para produção de vacinas em células
WI384,68-69.
A vacina contra a varicela é a primeira contra hepesvirus que foi aprovada
para uso clínico em vários países70. Os primeiros a serem vacindados no Japão
foram pessoas saudáveis, em seguida foram crianças imunodeprimidas incluindo as
que tinham leucemia e tiveram bons resultados68, 70.
Em 1980 ensaios clínicos da vacina foram iniciados nos Estados Unidos
levando asua aprovação pelo Food and Drug Administration em 199571,72. Antes da
vacina contra varicela existiam 4 milhões de casos de varicela por ano no USA, após
ser licenciada consequentemente o índice de varicela reduziu para 76-87% no
período de 1995 a 200073.
Antes de desenvolver a vacina houve muitos questionamentos, um deles era
a possibilidade de que o vírus da vacina iria se tornar latente, possivelmente
resultando no desenvolvimento posterior de HZ, já que a vacina contem o vírus
infeccioso74.
Já existem estudos que mostram a reativação do vírus em crianças
leucêmicas imunizadas, porém a gravidade do herpes zoster nessas crianças
vacinadas foi significativamente menor do que nas crianças leucêmicas com
infecção natural do VZV75. Em um adulto saudável que recebeu a vacina, mesmo
após exposição ao vírus, não apresentou sintomas, porém mais tarde teve um
episódio de herpes zoster causado pelo tipo selvagem VZV77. Atualmente, para
melhor diferenciar o vírus atenuado (Oka) do vírus infeccioso da varicela, técnicas de
PCR utilizando RFLPs são desenvolvidas, e sãoconsideradas muito importantes em
estudos epdemiológicos66.
Pesquisas apontam que a eficácia da vacina não diminui substancialmente ao
longo do tempo, alcança 97% no primeiro ano e 84% oito anos após a vacinação78.
Em relação àproteção ela é modesta contra a infecção, moderada contra doença
clínica e muito bem contra a doença grava79. Indicações e contraindicações para a

14
administração de vacina têm sidoestabelecidas pela American Academy of
Pediatrics80.
1.6 Epidemiologia
A varicela é uma das doenças transmissíveis mais comuns, é
predominantemente uma doença da infância, acomete principalmente a faixa de
idade de 4 – 10 anos1.
O VZV tem uma ampla distribuição geográfica, pode ser detectada em
qualquer um dos cinco continentes. É o únicovírus dentre a família Herpesviridae
que causa doença em humanos, que podeser transmitido através da via aérea,
apresenta um padrão de sazonalidade, e suas epidemias anuais são mais
prevalentes em climas temperados, e ocorrem na maioria das vezes durante o
inverno e a primavera,45.
VZV pode ser transmitido diretamente após contato com as lesões na pele de
pessoas infectadas, o vírus propaga-se rapidamente a outros indivíduos
susceptíveis. O índice de HZ aumenta com a idade, apontando com uma idade por
volta de 50 anos o índice é de aproximadamente 3 casos / 1000 pessoa-ano. Com
80 anos de idade o índice alcança 10 casos / 1000 pessoa-ano4,45.
Pessoas de climas tropicais são grupos considerados de alto risco para
adquirir a varicela quando emigram para os países de alta endemicidade do VZV,
influenciando sua epidemiologia. A varicela não é uma doença de notificação
obrigatória no Brasil, o que torna sua prevalência difícil de ser estimada. A
epidemiologia do VZV pode ser medida a partir de estudos de soro prevalência ou
de casos notificados em geral50-52.
1.7 Epidemiologia Molecular

15
Muitos laboratórios desenvolveram metodologias de reação em cadeia da
polimerase (PCR)para melhor compreender a patogênese do VZV45.
Algunsdemonstram a detecção do VZV na saliva e outros no sangue46-47. Em
alguns estudos iniciais, o DNA do VZV foi caracterizado pelo RFLP, que analisou
variações de cepas entre o tipo selvagem e o tipo vacina. Os marcadores RFLP de
VZV são os ORF 38 (PstI), 54 (BglII) e 62 (Smal) são considerados para vacinas e
estudos epidemiológicos44.
A maioria dos estudos mostram que as cepas do tipo selvagem na América
do Norte eEuropa foram caracterizadoscomo PstI+ BglI-, Cepas africanas e asiáticas
foram BglI+, Japonês Oka-likecepas do tipo selvagem PstI+/ PstI-BglI+Smal-e a
cepavacinal OkaPstI-BglI+Smal+66,92.
Estudos de pesquisa genômica do VZV levaram à identificação de vários
genótipos, que circulam de forma estável estabelecendo distribuição global, e
analisando esse fluxo a observação é que esta distribuição está em fluxo em alguns
países devido a imigração44.
Esses estudos de genotipagem do VZV vêm sendo realizados para verificar
os seus genótipos e em que área geográfica estão localizados. Em um estudo
recente feito por Jonas Schmidt-Chanasit em 201144 mostra a distribuição dos
clados de acordo com a nomenclatura universal (Figura 5).
Nos países da Europa foi observado a presença do clade 1 em 50% das
amostras e 37% para o clade 3. Em conjunto,os clades 1 e 3 representaram 87%
das cepas circulantes de VZV na Europa44. Na África o mais abundante encontrado
foi o clade 5 com 100%, No entanto, deve considerar-se que apenas alguns vírus
foram avaliados a partir de apenas quatro países africanos44.
Por causa da longa história de colonização europeia os clades 1e 3, foram
detectadas com um total de 74%na Oceania. Esses mesmos clades são dominante
nas Américas com 87% incluindo estudos realizados no Canadá, México e Reino
Unido44.

16
Ao contráriode todos os outros continentes, o clade 2 do VZV
representadominância na Ásia. De um total de 134 amostras analisadas, 99 (74%)
pertencem ao clade já mencionado44.
Figura 5 - Mapa da distribuição geográfica dos clades do VZV. Fonte: JONAS SCHMIDT-CHANASIT et al.,201144.
Um genótipo recombinante proposto no Brasil,53é provavelmente uma cepa do
genótipo M. Dos 10 isolados obtidos no Brasil testados em laboratório, 8 foram
considerados da linhagens do genótipo M e tinham padrões semelhantes as dos
SNP´s descritos por Muir et ai54.
A análise genotípica poderia levar àidentificação de fatores de virulência e,
consequentemente, para a melhoria dacompreensão da biologia do VZV104. Outra
questão importanteque pode ser solucionado usando esta abordagem é a avaliação
do direcionamento e predisposição da filogenia do VZV na população vacinada e
não vacinada54.
Em princípio, o domínio regional de claodos específicos do VZV pode ser
dependente de fatores ambientais, condições evolutivas, interações e ou importação
do vírusatravés da imigração ou de viagens. As abordagens inovadoras para a
vigilância global de clados VZV são ferramentas importantes para resolver estas
perguntas.

17
2 OBJETIVOS
2.1 Geral
Realizar estudo de caracterização molecular do vírus Varicela Zoster
detectado em amostras clinicas de pacientes atendidos em uma unidade terciaria de
saúde no Amazonas.
2.2 Específicos
Identificar o genótipo do VZV circulante em Manaus.
Identificar e diferenciar as cepas dos tipode selvagem e vacinal.
Realizar estudo de comparação com cepas virais de outras regiões e com a
vacinal, com a finalidade de identificar mutações no genoma.

18
3 MATERIAIS E MÉTODOS
3.1 Modelo de Estudo
Estudo descritivo de caracterização molecular dos casos de VZV identificados
no período de 2010 a 2014, na Fundação de Medicina Tropical Dr. Heitor Vieira
Dourado(FMT-HVD).
3.2 Seleção das amostras
Toda amostra clínica de vesículas de pele sugestivas de catapora e/ou herpes
zoster, que foram confirmadas por PCR para o vírus VZV, selecionadas no período
de janeiro de 2010 a dezembro de 2014, foram incluídas no estudo de
caracterização molecular.
3.3 Método de diagnóstico molecular
3.3.1 Detecção do genoma viral da família Herpesviridae
3.3.1.1Extração de DNA.
O DNA viral foi extraído utilizando QIAamp Viral DNA Mini Kit (QIAamp Blood
DNA Mini Kit, Qiagen), seguindo-se as instruções do fabricante.
3.3.1.2 Amplificação do DNA PCR
A amplificação gênica foi realizada pela técnica de PCR, e permitiu identificar
o vírus a partir da região ORF8 do VZV, utilizando os primers VP22, VM20, descritos
por MARKOULATOS, et al. (2001)65. Tabela 2.

19
Tabela 2–Tabela de primers usados para PCR65
Primers Sequências (5'→ 3' ) Vírus Região do Posição Comprimento
Genoma Amplificado do Primer do Amplímero(bp)
VP22 CACACGATAATGCCTGATCGG VZV ORF 8
9796–9816 275
VM20 TGCTGATATTTCCACGGTACAGC 10049–10071
A detecção do produto da PCR foi realizada através de eletroforese em gel de
agarose 2% corado com brometo de etídio, sendo registrado em sistema de foto
documentação digital.
3.3.2Sequenciamento de nucleotídeo
3.3.2.1 Amplificação do DNA PCR
A PCR foi otimizada, ultilizando seis pares de primer´s para amplificar
osfraguimentos das regiões conservadas do VZV, como anteriormente já descrito
54(Tabela 3).Para os pares de prime´s Nla/Fok e PstA/B foi realizado dez ciclo de
amplificação a 95°C por 30 segundos e 63°C por 1 minuto, trinta ciclos a 95°C por
30 segundos, 60°C por 30 segundos e 72°C por 30 segundos, seguido por um ciclo
de 72°C durante 10 minutos, usando o termocicladoreppendorf (Marstercycler
gradiente).E para o primer p22R1f/p22R1r foi realizado vinte e cinco ciclo de
amplificação a 95°C por 1 minuto, 50°C por 1 minuto e 72°C por 3 minutos, seguido
por um ciclo de 72°C durante 10 minutos, usando o termociclador(Applied
Biosystens).
Tabela 3–Tabela de primers usados para PCR para genotigem54
PRIMER
Nome sequencia (5`→ 3`) fraguimentos ORF
Nla GGAACCCCTGCACCATTAAA 222 54
Fok TCCCTTCATGCCCGTTACAT
Pst A TTGAA CAATCACGAACCGTT 350 38
Pst B CGGGTGAACCGTATTCTG AG
p22R1f GGG TTT TGT ATG AGC GTT GG 447 22
p22R1r CCC CCG AGG TTC GTA ATA TC

20
Foi realizado eletroforese em gel de agarose 1,8% corado com brometo de
etídio, para a detecção dos fraguimentos, sendo registrado em sistema de foto
documentação digital.
3.3.2.1.1 Purificação do produto da PCR
O produto amplificado foi purificado de acordo com os fraguimentos das
regiões conservadas do VZV escolhodas. Para os fraguimentos de tamanho 447 pb
e 350 pb foi realizado o método de precipitação de produto da PCR com
Polietilenoglicol (PEG),e os fraguimnetos de tanho 222 pb foi realizado pelo
kit.Confirmado pela eletrofose em gel de aragose 1,8%. Após a purificação foram
quantificadas usando NanoDrop 200 (thermo scintifo)e diluídas para uma
concentração de 10ng/µl devido ao tamanho do produto obtido para fazer o
sequenciamento.
3.3.3 Genotipagem do vírus VZV
3.3.4Sequeciamento e analise filogenética
O sequenciamento do DNA foi realizado pelo sequnciador (3130xl Genetic
analyzer), a reação de sequenciamnto dos produtos da PCR purificados foi usado o
BigDye Terminator, version 3.1. De cada primer (5 pmol/µl) foi usado 0,5µl, 10ng de
produto do DNA purificado e 2µl de Tampão. E iniciando o sequenciamento
utilizando no termociclador(Applied Biosystens).com 25 ciclos de 95°C de 10 s, 50°C
de 5 s, 60°C de 4 s e 1 ciclo com 4 min em ∞. E antes de ir para o sequenciador foi
feita uma precipitação com HIDI e incubada por 95°C por 3 min no gelo.
Todas as sequencias foram alinhadas e comparadas com a cepa Dumas
(GenBank numero de acesso X04370), usando o programa (Geneious 7.1.9).A
arvore filogenética foi do tipo concatenada sequenciadas as 3 ORF escolhidas.
Metodo de referência BAYESIANA com 1 milhão de geração e variação com sítio
invariável e analizado a cada 1000 gerações, modelo de substituição HKY85

21
ultilizando o programa (Model test 2.1.7), metodo AIC para escolher o modelo.
3.4 Aspectos éticos
Este projeto foi submetido e aprovado pelo Comitê de Ética em Pesquisa em
Seres Humanos (CEP) da FMTAM (Processo Nº 2891, de 5 de dezembro de 2007) e
está de acordo com as normas previstas na resolução Nº196/96, do Conselho
Nacional de Saúde.
3.5 Financiamento
Projeto aprovado no Edital 021/2011 Universal/FAPEAM
22
4 RESULTADOS
4.1 Seleção de amostra e identificação do VZV
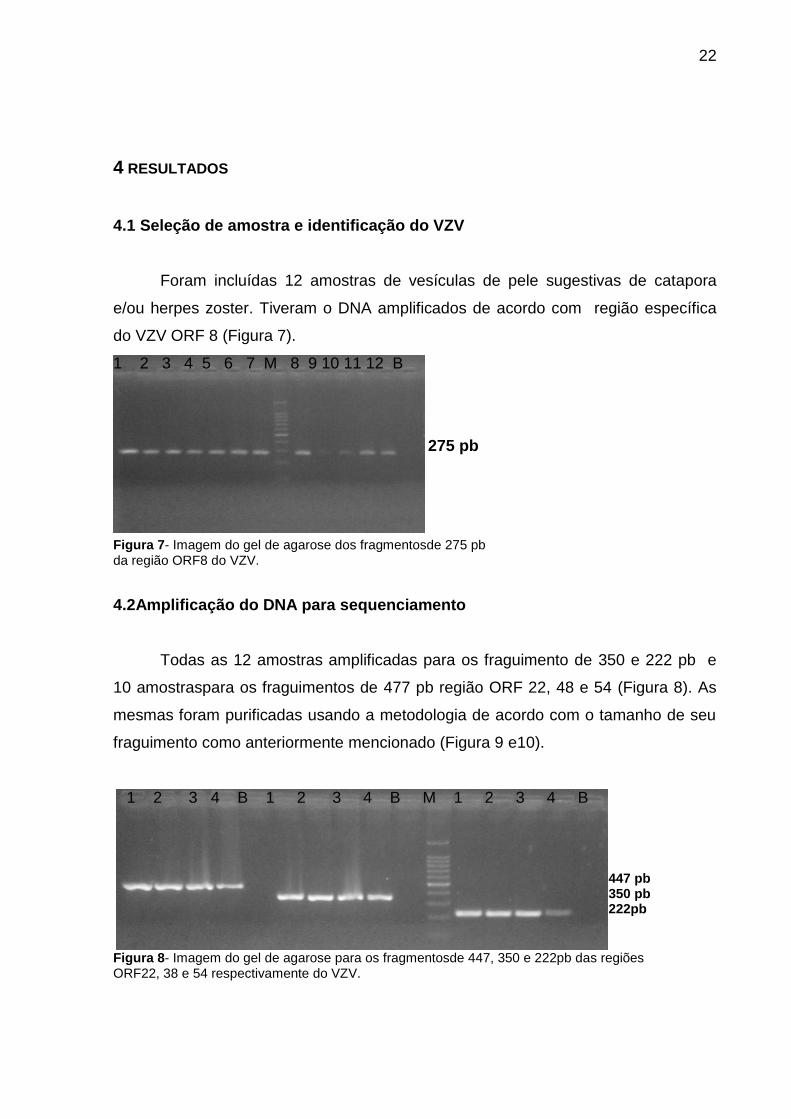
Foram incluídas 12 amostras de vesículas de pele sugestivas de catapora
e/ou herpes zoster. Tiveram o DNA amplificados de acordo com região específica
do VZV ORF 8 (Figura 7).
1 2 3 4 5 6 7 M 8 9 10 11 12 B
275 pb
Figura 7- Imagem do gel de agarose dos fragmentosde 275 pb da região ORF8 do VZV.
4.2Amplificação do DNA para sequenciamento
Todas as 12 amostras amplificadas para os fraguimento de 350 e 222 pb e
10 amostraspara os fraguimentos de 477 pb região ORF 22, 48 e 54 (Figura 8). As
mesmas foram purificadas usando a metodologia de acordo com o tamanho de seu
fraguimento como anteriormente mencionado (Figura 9 e10).
1 2 3 4 B 1 2 3 4 B M 1 2 3 4 B
447 pb 350 pb 222pb
Figura 8- Imagem do gel de agarose para os fragmentosde 447, 350 e 222pb das regiões ORF22, 38 e 54 respectivamente do VZV.
23
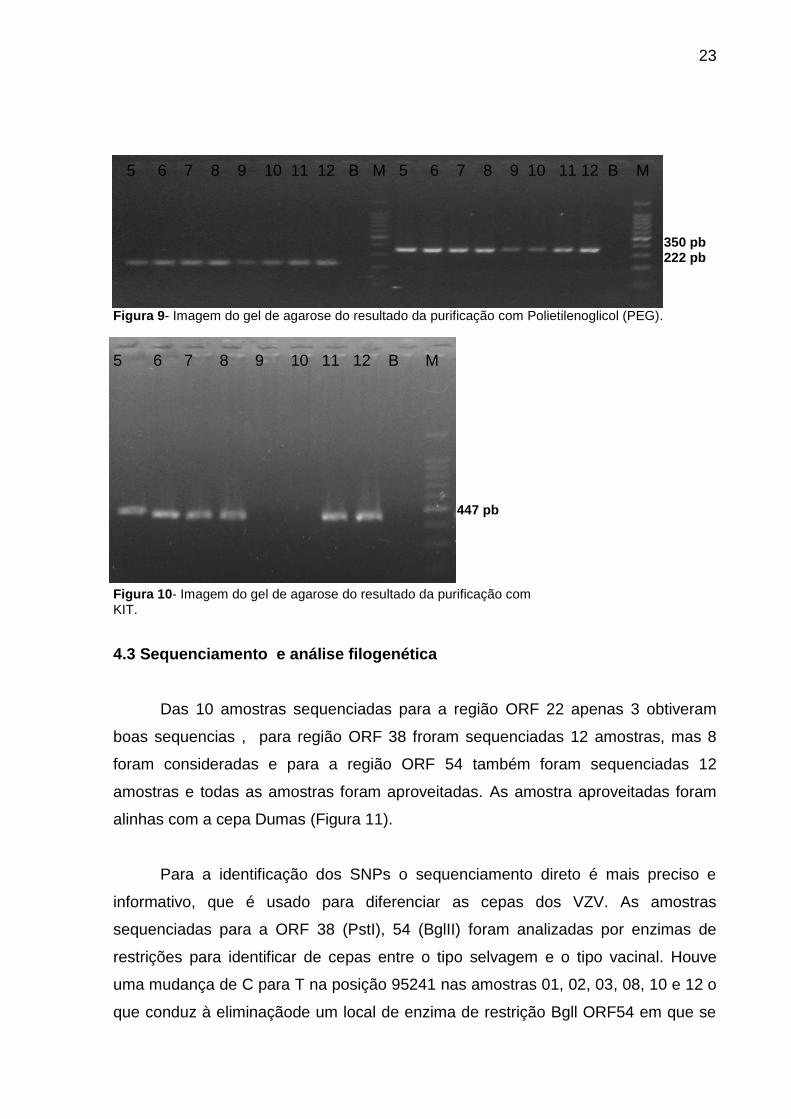
5 6 7 8 9 10 11 12 B M 5 6 7 8 9 10 11 12 B M
350 pb 222 pb
Figura 9- Imagem do gel de agarose do resultado da purificação com Polietilenoglicol (PEG).
5 6 7 8 9 10 11 12 B M
447 pb
Figura 10- Imagem do gel de agarose do resultado da purificação com KIT.
4.3 Sequenciamento e análise filogenética
Das 10 amostras sequenciadas para a região ORF 22 apenas 3 obtiveram
boas sequencias , para região ORF 38 froram sequenciadas 12 amostras, mas 8
foram consideradas e para a região ORF 54 também foram sequenciadas 12
amostras e todas as amostras foram aproveitadas. As amostra aproveitadas foram
alinhas com a cepa Dumas (Figura 11).
Para a identificação dos SNPs o sequenciamento direto é mais preciso e
informativo, que é usado para diferenciar as cepas dos VZV. As amostras
sequenciadas para a ORF 38 (PstI), 54 (BglII) foram analizadas por enzimas de
restrições para identificar de cepas entre o tipo selvagem e o tipo vacinal. Houve
uma mudança de C para T na posição 95241 nas amostras 01, 02, 03, 08, 10 e 12 o
que conduz à eliminaçãode um local de enzima de restrição Bgll ORF54 em que se

24
encontra em todas as cepas japonesas e que está ausente na maioria dos VZV
isolado no EUA (Figura 12 e 13).
Foram analizandas as amostras sequenciadas das ORF38 (PstI), 54 (BglII)
para a diferenciação das cepas selvagens das cepas vacinal, as amostras 01, 03,
04, 05, 07, 09 e 10 foram designadas tipo selvagem wt (wild-type), sendo que as
amostras 01, 03 e 10 são mais comuns na America Norte e Europa (Tabela 4).
A analise das variações do genótios usando dados da ORF54 mostra qua as
amostras 01, 02, 03, 08, 10 e 12 apresentão variações comuns para os clades 1, 3 e
VI (provisório). As amostras 05, 06, 07, 09 e 11 tem variações comuns para o clade
2, 4 e 5 (Tabela 4).

25
Figura 11- Representação do resultados das sequencias das regiões ORF 22, 38 e 54 alinhadas com a cepa Dumas12.
26
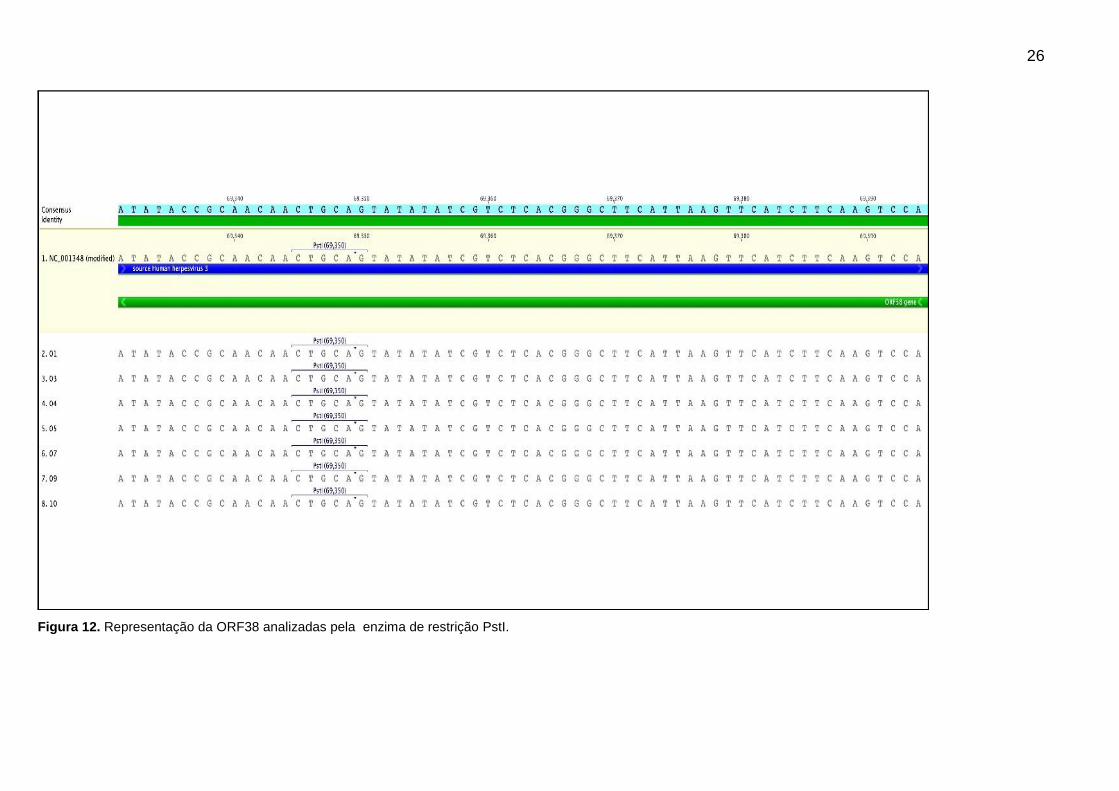
Figura 12. Representação da ORF38 analizadas pela enzima de restrição PstI.

27
Figura 13.Representação daORF54 analizadas pela enzima de restrição BglII. O (Y) significa a presença de (C) ou (T ).
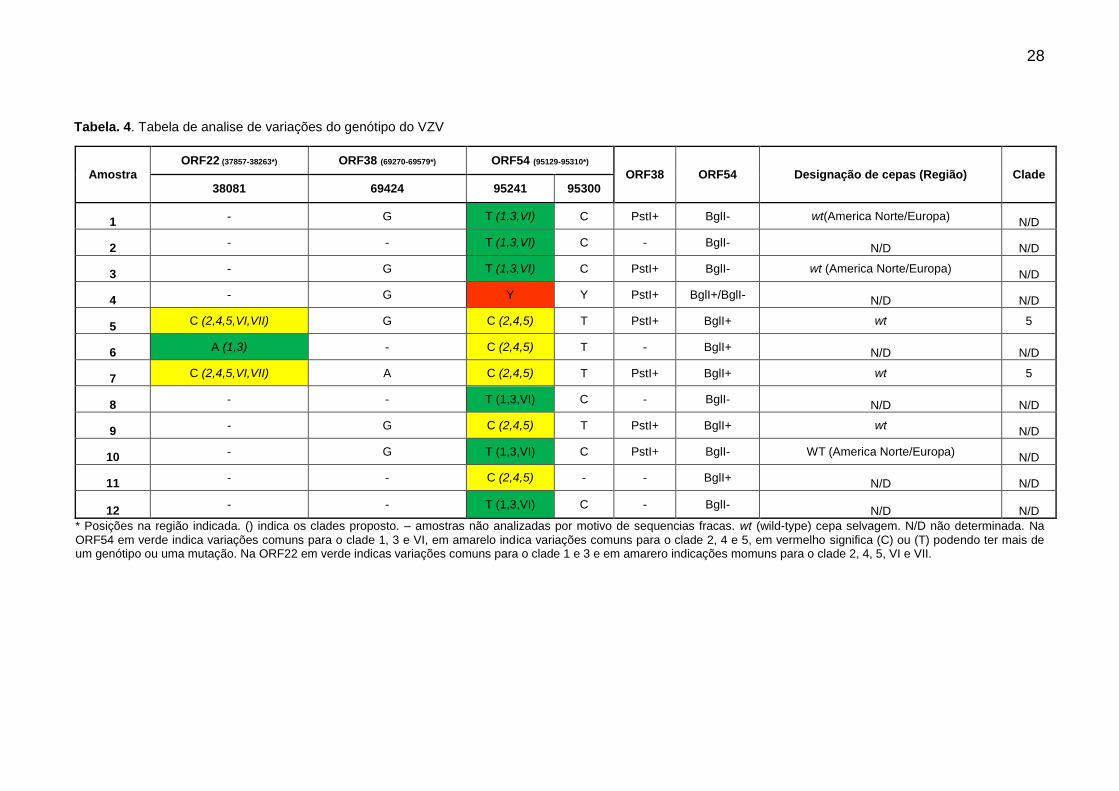
28
Amostra
ORF22 (37857-38263*) ORF38 (69270-69579*) ORF54 (95129-95310*)
ORF38 ORF54 Designação de cepas (Região) Clade
38081 69424 95241 95300
1 - G T (1,3,VI) C PstI+ BglI- wt(America Norte/Europa) N/D
2 - - T (1,3,VI) C - BglI- N/D N/D
3 - G T (1,3,VI) C PstI+ BglI- wt (America Norte/Europa) N/D
4 - G Y Y PstI+ BglI+/BglI- N/D N/D
5 C (2,4,5,VI,VII) G C (2,4,5) T PstI+ BglI+ wt 5
6 A (1,3) - C (2,4,5) T - BglI+ N/D N/D
7 C (2,4,5,VI,VII) A C (2,4,5) T PstI+ BglI+ wt 5
8 - - T (1,3,VI) C - BglI- N/D N/D
9 - G C (2,4,5) T PstI+ BglI+ wt N/D
10 - G T (1,3,VI) C PstI+ BglI- WT (America Norte/Europa) N/D
11 - - C (2,4,5) - - BglI+ N/D N/D
12 - - T (1,3,VI) C - BglI-
N/D N/D
* Posições na região indicada. () indica os clades proposto. – amostras não analizadas por motivo de sequencias fracas. wt (wild-type) cepa selvagem. N/D não determinada. Na
ORF54 em verde indica variações comuns para o clade 1, 3 e VI, em amarelo indica variações comuns para o clade 2, 4 e 5, em vermelho significa (C) ou (T) podendo ter mais de um genótipo ou uma mutação. Na ORF22 em verde indicas variações comuns para o clade 1 e 3 e em amarero indicações momuns para o clade 2, 4, 5, VI e VII.
Tabela. 4. Tabela de analise de variações do genótipo do VZV

29
Na ORF22 algumas variablilidades foram aparentes para apenas para um
clade. Amostra 06 apresenta variação comum para os clades 1 e 3, e as amostras
05 e 07 a proncípio apresentam variações comuns para os clades 2, 4, 5, VI e VII
esses dois ultimos considerados provisórios, porém, as mesmas amostras aprentam
todas as variações indicativa para o clade 5 (Tabela 4).
A arvore filogenética foi construída usando cepas referencia de sequências
(BLAST) limitando apenas na ORF22, indicando que a amostra 05 e 07
apresentaram a tendência para o clado 5 (Figura 14).
Figura 14.Arvore filogenética do VZV baseada na incluindo as amostras 05 e 07. Clado 1 Incluem 10 cepas:
referencia Dumas (NC001348), Netherlands; BC (AY548171), British Columbia, Canada; 36 (DQ479958), New Brunswick, Canada; 49 (DQ479959), New Brunswick, Canada; referencia cepa MSP (AY548170), Minnesota, EUA; 32p5 (DQ479961), Texas, EUA; Kel (DQ479954), Iowa, EUA; SD (DQ479953), South Dakota, EUA; NH293 (DQ674250), EUA; SVETA (EU154348), Russia; clado 3 incluem 4 cepas: referencia 03-500 (DQ479957), Alberta, Canada; 11 (DQ47995), New Brunswick, Canada; 22 (DQ479956), New Brunswick, Canada; reference strain HJO (AJ871403), Germany; clado 2 incluem 4 cepas: referencia strain pOka (AB097933), Japão, VariVax (DQ008355), VariVax (DQ008354), Oka, sustrain pOka (AB097932); clado 5 incluem 1 cepa referência CA123
strain (DQ457052), California, USA; clado 4 incluem 2 cepas: referencia 8 (DQ479960), New Brunswick, Canada and referencia DR (DQ452050), EUA.

30
5 DISCUSSÃO
A genotipagem é uma ferramenta epidemiológica útil para muitos humana
herpesvírus, embora não tenha geralmente levado a correlações entre variação de
sequência e patologia vírus95-99.
Protocolos de genotipagem destinados ao laboratório foram desenvolvido
para simples execução. Eles também devem ser capazes de produzir resultados a
partir da quantidade limitada de DNA tipicamente recuperado a partir de amostras
clínicas , por exemplo , vesicular. As amotras de vesicula mostrou que mantem rica
em DNA conservado, e a otmização do PCR anteriormente já descrito, amplificaram
as amostras de acordo com seus respectivos fragmentos, o que indica uma boa
reprodução do protocolo54.
O sistema de nomenclatura do VZV, adotado em 25 de julho de 2008,
também foi ultilizado neste trabalho para padronizar e melhor compreenção das
classificações dos clados. Nesse estudo com as regiões definidas ORF22, 54 e 38,
nas amostras em geral foi possível identificar variações importantes para todos os
clados
Eu executei alvo ORF de sequenciamento para uma variedade de isolados
distribuídos globalmente , começando com uma comparação entre o sítio PstI em
ORF38 e site da BglI em ORF54 . Análise com enzimas de restrição são importante
para a determinação das cepas do VZV, na diferenciação da cepa vacinal com a
cepa selvagem ultilizando as ORF´s 38 (PstI) e 54(BgtI). As cepas selvagem wt na
América do Norte e Europa é comum encontrar PstI+ BglI-, na África e Ásia BglI+, no
Japão PstI+/- BglI- Smal+ eavacinal PstI- BglI+Smal+62,92.
Em todas as amotras foram encontradas a presença do sítio PstI o que
evidencia a ausencia da cepa vacival entre as amostras, apresentando apenas a
cepa selvagem algumas encontradas nas Américas e Europa. Outros estudos
mostram que 70% de numeros de cepas analizadas foram observadas na Europa
para o clado 544. A presença dessas cepas pode ser explicada pelos principais

31
países de origem de imigrantes do Brasil, mostrando que entre 2015 a 2010, 25%
são dos Estados Unidos e 6% do Reino Unido105.
A região da ORF54(BgtI) pode ser ultilizado para diferenciar cepas da região
européia e América do Norte das japonesas, porem EstadosUnidos, Canadá, grande
parte da Europa , Austrália oriental e América do Sul, onde as cepas doVZV para os
clados 2, 4 e 5 não são tão encontrados.Por tanto a ORF54 apresentou apenas
algumas variações comuns encontradas em quase todos os clados e tambem houve
uma mudança de bases na região 95241 da amostra 04 podendo ser uma mutação
ou outros genótipos.
Este marcardor ORF22 derivado de 447 pb resolve genótipos distintos
que se correlacionam com a nossa compreensão da variabilidade VZV54. Para fazer
o mapeamento dos genótipos foi utilizado o protocolo de genotipagemdefinido por
outros estudos54,43. E duas amostras correspondente a região da ORF22 apresentou
uma linhagem que é definida para o clado 5. A presença desse clado provavelmente
é explicado pela questão das imigracões no Brasil nos ultimos anos.
Uma observação foi a descoberta de domínio geográfico de cepas do
genótipo M (clado 5) em áreas tropicais e subtropicais, como África (RDC, República
do Tchad,Marrocos) e na Ásia (Índia, Bangladesh, Nepal e China). E cepas mais
recentemente obtida a partir de Brasil, Argentina, CoteD'Ivoire, Etiópia, Zimbábue,
África do Sul, Tailândia, Vietnã,norte da China, Grã-Bretanha, Tajiquistão,
Cazaquistão, Uzbequistão,Quirguistão, Turcomenistão e Azerbaijão54.Outra
evidencia baseadas nessas mesmas amostras foi a árvore filigenética, que
apresentou a tendencia de linhagem também para o clado 5.
A origem de cepas do VZV poderia refletir a imunogenética do local da
população humana em que originalmente evoluiu.Referente a distribuição do
genótipo tambem reflete as qualidades demograficas.Além disso , foi sugerido que
os eventos de recombinação podeter influenciado a evolução dos diferentes clados
já existentes do VZV100-103.

32
6 CONCLUSÃO
De todas as amostras analizadas apenas nas 5 e 7 froram observadas
possível linhagem para o clado 5. A ávore filigenética mostra a mesma linhagem
para o clado 5.
Nesse estudo foi isolado o DNA do VZV para análises genéticas e
caracterização de cepas do VZV, e seis amostras encontradas foram consideradas
cepas selvagem o que evidencia a ausência de cepas vacinais Oka Vacina, entre as
amostras estudadas em Manaus.
Todas as amostras foram alinhadas e comparadas com a cepa Dumas, para
identificar os SNP´s que possa determinar os clados, mas apenas duas amostras 5 e
7 foram consideradas mais próximas do clado 5, e as mesmas foram comparadas
com outras cepas inclusive a vacinal, porém limitadas pela região ORF22 o que
possibilitou a formação da árvore filigenética.
Algumas amostras apresentaram alterações indeterminadas em algumas
posições, mas para defini-las é necessário refazer as sequências e analizar outras
vezes.

33
7 REFERÊNCIAS BIBLIOGRAFICAS
1. Arie jz, jangu eb, john rp, paul dg, barry ds. Principles and practice of clinical virology. 5ed. Englad:john wiley & sons ltd.:2004.
2. International committee on taxonomy of viruses[internet]. Ictv; ec 45, edinburgh, july 2013 [acesso em 04 abril 2014]. Disponível em: www.ictvonline.org.
3. Luzuriaga k, sullivan jl. Infectious mononucleosis. N engl j med [internet]. 2010 may 27;362(21):1993–2000. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20505178.
4. Arvin annm. Varicella-zoster virus. 1996;9(3):361–81.
5. Brunell pa. Transmission of chickenpox in a school setting prior to the bserved exanthem. Am j dis child 1989; 143: 1451–1452.
6. Tsolia m, gershon a a., steiberg sp, gelb l. Live attenuated varicella vaccine:
evidence that the virus is attenuated and the importance of skin lesions in transmission of varicella-zoster virus. J pediatr [internet]. 1990 feb;116(2):184–9. Available from: http://linkinghub.elsevier.com/retrieve/pii/s0022347605828720
7. Arvin am, moffat jf, redman r. Varicella-zoster virus: aspects of pathogenesis
and host response to natural infection and varicella vaccine. Adv virus res 1996; 46: 263–309.
8. Ehlers, b. 2008. Herpesviruses: discovery. In encyclopedia of virology. 3rd ed.
Vol. 2. B.w.j. Mahy & m.h.v. Van regenmortel, eds.: 420–429. Academic press. Oxford.
9. Davison aj. Evolution of the herpesviruses. Vet microbiol [internet]. 2002 apr
22;86(1-2):69–88. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11888691
10. Roberto p, stephens s. Capítulo 2 virologia. 1960;125–220.
11. Viralzone [internet]; acids res. 2011 jan;39 [acesso em 04 abril 2014]. Disponível em: viralzone.expasy.org/all_by_species/179.html
12. Davison a j, scott je. The complete dna sequence of varicella-zoster virus. J gen virol [internet]. 1986 sep;67 ( pt 9):1759–816. Available from:http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=421634&tool=pmcentrez&rendertype=abstract

34
13. King amq, adams mj, carstens eb, lefkowitz ej. Virus taxonomy.
14. Barnes dw, whitley rj. Cns diseases associated with varicella zoster virus and herpes simplex virus infection. Pathogenesis and current therapy. Neurol clin 1986;4(1):265–83.
15. Poultry r. Dna configuration in the core of marek ’ s disease virus mjr *. 1974;13(5):1148–50.
16. Document d. Title: varicella-zoster virus infections of the nervous system: clinical and pathologic correlates author(s): donald h. Gilden and b. K. Kleinschmidt-demasters source: 2001;6(june):1–12.
17. Cohen, j. I., and s. E. Straus. 1995. Varicella-zoster virus and its replication, p. 2525–2546. In b. Fields (ed.), virology, 3rd ed. Raven press, new york.
18. Tc s, io t. The characterization of varicella-zoster virus d n a (. 1966;0–3.
19. Ambagala apn, cohen ji. Varicella-zoster virus ie63, a major viral latency protein, is required to inhibit the alpha interferon-induced antiviral response. J virol [internet]. 2007 aug [cited 2014 apr 2];81(15):7844–51. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1951283&tool=pmcentrez&rendertype=abstract
20. Waters dj. Varicella-zoster characterization. 1985;71:62–71.
21. Edson, c. A., b. A. Hosler, c. A. Poodry, r. T. Schooley, d. J. Waters, and d. A. Thorley-lawson. 1985. Varicella-zoster virus envelope glycoproteins: biochemical characterization and identification in clinical material. Virology 145:62-71.
22. Edson cm, hosler ba, respess ra, waters dj. Cross-reactivity between herpes simplex virus glycoprotein b and a 63 , 000-dalton varicella-zoster virus envelope glycoprotein. 1985;

35
23. Keller pm, neff bj, ellis rw. Three major glycoprotein genes of varicella-zoster virus whose products have neutralization epitopes. 1984;52(1).
24. Vafai a, wroblewska z, wellish m, green m, gilden d. Analysis of three late varicella-zoster virus proteins, a 125,000-molecular-weight protein and gp1 and gp3. J virol [internet]. 1984 dec;52(3):953–9. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=254619&tool=pmcentrez&rendertype=abstract
25. Wroblewska z, gilden d, green m, devlin m, vafai a. Affinity-purified varicella-zoster virus glycoprotein gp1/gp3 stimulates the production of neutralizing antibody. J gen virol [internet]. 1985 aug;66 ( pt 8):1795–9. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2991442
26. Weigle, k. A., and c. Grose. 1983. Common expression of varicella-zoster viral glycoproteins in vitro and in chickenpox and zoster vesicles. J. Infect. Dis. 148:630-638.
27. Forghani b, dupuis kw, schmidt nj. With monoclonal antibodies . Varicella-zoster viral glycoproteins analyzed with monoclonal antibodies. 1984;52(1).
28. Forghani, b., n. J. Schmidt, c. K. Myoraku, and d. Gallo. 1982. Serological reactivity of some monoclonal antibodies to varicella-zoster virus. Arch. Virol. 73:311-317.
29. Grose c, edwards dp, friedrichs we, mcguire wl. Monoclonal antibodies against three major glycoproteins of monoclonal antibodies against three major glycoproteins of varicella-zoster virus. 1983;
30. Montalvo e a, parmley rt, grose c. Structural analysis of the varicella-zoster virus gp98-gp62 complex: posttranslational addition of n-linked and o-linked oligosaccharide moieties. J virol [internet]. 1985 mar;53(3):761–70. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=254704&tool=pmcentrez&rendertype=abstract

36
31. Davison a j, waters dj, edson cm. Identification of the products of a varicella-zoster virus glycoprotein gene. J gen virol [internet]. 1985 oct;66 ( pt 10:2237–42. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2995558
32. Mettenleiter tc, klupp bg, granzow h. Herpesvirus assembly: an update. Virus res [internet]. 2009 aug [cited 2014 mar 21];143(2):222–34. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19651457
33. Roizman b, knipe dm, whitley rj. 2007. Herpes simplex viruses, p 2501–2601.inknipe dm, howley pm, griffin de, lamb ra, martin ma, roizman b, straus se (ed), fields virology, 5th ed. Lippincott williams & wilkins, philadelphia, pa.
34. Kelly bj, fraefel c, cunningham al, diefenbach rj. Functional roles of the tegument proteins of herpes simplex virus type 1. Virus res [internet]. 2009 nov [cited 2014 apr 2];145(2):173–86. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19615419
35. Manuscript a, genome tvv. Nih public access. 2012;(table 1):1–14.
36. Wildy p. And watson d. H. Electron microscopic studies on the architecture of animal.
37. Epstein bma. Observations on the fine structure of mature h e r p e s s i m p l e x virus a n d o n t h e c o m p o s i t i o n o f its n u c l e o i d ( from the bland-sutton institute o / pathology , the middlesex hospital , london ) p y ~ s 1 to 5 ( received for pu. 1961;(13).
38. Furlong d, swift h, roizman b. Arrangement of herpesvirus deoxyribonucleic acid in the core. J virol [internet]. 1972 nov;10(5):1071–4. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=356578&tool=pmcentrez&rendertype=abstract
39. Nazerian k. Dna configuration in the core of marek ’ s disease virus dna configuration in the core of marek ' s disease virus. 1974;13(5).

37
40. Morgan byc, rose hm, am pd, jones ep. D e v e l o p m e n t of h e r p e s s i m p l e x virus * ( from the department of microbiology , college of physician ~ and surgeons , columbia university , new york ) materials and metkods hela cells were cultured in a medium consisting of earle ’ s ba. 1959;
41. Roizman, b., s. B. Spring, and j. Schwartz. 1969. The herpes virion and its precursors made in productively and in abortively infected cells. Proc. Nat. Acad. Sci. U.s.a. 28: 1890-1898.
42. Quinlivan m, breuer j. Molecular studies of varicella zoster virus. Rev med virol [internet]. 2006;16(4):225–50. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16791838
43. Breuer j, grose c, norberg p, tipples g, schmid ds. A proposal for a common nomenclature for viral clades that form the species varicella-zoster virus: summary of vzv nomenclature meeting 2008, barts and the london school of medicine and dentistry, 24-25 july 2008. J gen virol [internet]. 2010 apr [cited 2014 apr 2];91(pt 4):821–8. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2888159&tool=pmcentrez&rendertype=abstract
44. Schmidt-chanasit j, sauerbrei a. Evolution and world-wide distribution of varicella-zoster virus clades. Infect genet evol [internet]. Elsevier b.v.; 2011 jan [cited 2014 apr 2];11(1):1–10. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20817040
45. Gershon a a, gershon md, breuer j, levin mj, oaklander al, griffiths pd. Advances in the understanding of the pathogenesis and epidemiology of herpes zoster. J clin virol [internet]. Elsevier b.v.; 2010 may [cited 2014 apr 2];48 suppl 1:s2–7. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20510263
46. Mehta sk, tyring sk, gilden dh, cohrs rj, leal mj, castro v a, et al. Varicella-zoster virus in the saliva of patients with herpes zoster. J infect dis [internet]. 2008 mar 1 [cited 2014 apr 2];197(5):654–7. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2938732&tool=pmcentrez&rendertype=abstract

38
47. Gauthier a, breuer j, carrington d, martin m, r´emy v. Epidemiology and cost of herpes zoster and post-herpetic neuralgia in the united kingdom. Epidemiol infect 2009;137:38–47.
48. Miller e. Epidemiology, outcome and control of varicella-zoster infection. Rev med microbiol 1993;4:222–30.
49. Braun-falco m, hoffman m. Herpes zoster with progression to acute varicella
zoster virusmeningoencephalitis. Int j dermatol 2009;48:834-9.
50. Wharton m. The epidemiology of varicella-zoster. 1996;10(3):571–81.
51. Massad, e.; azevedo-neto, r.s.; burattini, m.n. Et al. - assessing the efficacy of
a mixed vaccination strategy against rubella in são paulo, brazil. Int. J. Epidem., 24: 842-850, 1995.
52. In v, paulo são, lúcia a, yu f, costa jm, amaku m, et al. Three year seroepidemiological study of varicella-zoster. 2000;42(3):125–8.
53. Muir wb, nichols r, breuer j. Phylogenetic analysis of varicella-zoster virus : evidence of intercontinental spread of genotypes and recombination phylogenetic analysis of varicella-zoster virus : evidence of intercontinental spread of genotypes and recombination. 2002;76(4).
54. Loparev vn, gonzalez a, tipples g, fickenscher h, torfason g, schmid ds, et al. Global identification of three major genotypes of varicella-zoster virus : longitudinal clustering and strategies for genotyping global identification of three major genotypes of varicella-zoster virus : longitudinal clustering and strategies for genotypi. 2004;
55. Bulakbasi n, kocaoglu m. Central nervous system infections of herpesvirus family. Neuroimaging clin n am [internet]. 2008 feb [cited 2014 apr 2];18(1):53–84; viii. Available from: http://www.ncbi.nlm.nih.gov/pubmed/18319155.
56. Roizman b, sears ae. Herpes simplex viruses and their replication. In: fields bn, knipe dm, editors. Virology. 2nd edition. New york: raven; 1990. P. 1795–841.

39
57. Rohman b, sears ae. Herpes simplex virus latency. 1987;
58. Palu g, benetti l, calistri a. Molecular basis of the interactions between herpes simplex viruses and hiv-1. Herpes 2001;8(2):50–5.
59. Kleinschmidt-demasters bk, gilden dh. Varicella-zoster virus infections of the nervous system: clinical and pathologic correlates. Arch pathol lab med [internet]. 2001 jun;125(6):770–80. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11371229
60. De la blanchardiere a, rozenberg f, caumes e, et al. Neurological complications of varicellazoster virus infection in adults with human immunodeficiency virus infection. Scand j infect dis 2000;32(3):263–9.
61. Persson a, bergström t, lindh m, namvar l, studahl m. Varicella-zoster virus cns
disease--viral load, clinical manifestations and sequels. J clin virol [internet]. 2009 nov [cited 2014 apr 2];46(3):249–53. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19709927
62. Oxman mn. Clinical manifestations of herpes zoster. In: arvin am, gershon aa,
editors. Varicella-zoster virus virology and clinical management. Cambridge, uk: cambridge university press; 2002. P. 246-75.
63. Quinlivan m, breuer j. Molecular and therapeutic aspects of varicella – zoster virus infection. 2005;7(15):1–24.
64. Davison aj, edson cm, ellis rw, forghani b. New common nomenclature for glycoprotein genes of varicella- zoster virus and their glycosylated products. 1986;57(3):1195–7.
65. Markoulatos p, georgopoulou a, siafakas n, plakokefalos e, tzanakaki g, kourea-kremastinou j. Laboratory diagnosis of common herpesvirus infections of the central nervous system by a multiplex pcr assay. 2001;39(12):4426–32.
66. Larussa p, lungu o, hardy i, gershon a, steinberg sp, silverstein s. Polymerase chain reaction products from restriction fragment length polymorphism of polymerase chain reaction products from vaccine and wild-type varicella-zoster virus isolates. 1992;
67. Larussa p, steinberg s, arvin a, dwyer d, burgess m, menegus m, et al. Polymerase chain reaction and restriction fragment length polymorphism analysis of varicella-zoster virus isolates from the united states and other parts of the world. 1998;178(suppl 1):64–6.

40
68. Takahashi m, otsuka t, okuno y, asano y, yazaki t, isomura s. Live attenuated
varicella vaccine used to prevent the spread of varicella in hospital. Lancet 1974;2:1288.
69. Gershon, a. A., s. P. Steinberg, p. Larussa, a. Ferrara, m. Hammerschlag, and l. Gelb. 1988. Immunization of healthy adults with live attenuated varicella vaccine. J. Infect. Dis. 158:132–137.
70. Ha, k., baba, k., ikeda, t., nishida, m. & yabuuchi, h. Application of live varicella vaccine to children with acute leukemia or other malignancies without suspension of anticancer therapy the world wide web at : application of live varicella vaccine to children with acute leukemia or other malignancies without suspension of anticancer therapy. (1980).
71. Krause, p. R. & klinman, d. M. Efficacy, immunogenicity, safety, and use of live attenuated chickenpox vaccine. J. Pediatr. 127, 518–25 (1995).
72. Annunziato pa, gershon aa. Primary immunization against varicella. In: arvin am, gershon aa, eds. Varicella-zoster virus: virology and clinical management. Cambridge, united kingdom: cambridge university press, 2000:460-76
73. Vazquez m. Varicella zoster virus infections in children after the introduction of live attenuated varicella vaccine. Curr opin pediatr 2004;161:80–4.
74. Takahashi, m. Et al. Development of varicella vaccine. J. Infect. Dis. 197 suppl 2, s41–4 (2008).
75. The new england journal of medicine downloaded from nejm.org at uea on april 10, 2014. For personal use only. No other uses without permission. Copyright © 1991 massachusetts medical society. All rights reserved. 1991;
76. Itor ed, ls ia. V v — t f s y. 2001;344(13):1007–9.
77. Hammerschlag, m. R., a. A. Gershon, s. P. Steinberg, l. Clarke, and l. D. Gelb. 1989. Herpes zoster in an adult recipient of live attenuated varicella vaccine. J. Infect. Dis. 160:535–537.
78. Vázquez m, shapiro ed. Varicella vaccine and infection with varicella-zoster virus. N engl j med [internet]. 2005 feb 3;352(5):439–40. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15689581.
79. Village eg. Varicella vaccine stanley a . Plotkin the online version of this article , along with updated information and services , is located on the world wide web at : illinois , 60007 . Copyright © 1996 by the american academy of pediatrics . All rights reserved . Print varicella vaccine. 1996;
80. Village eg. Recommendations for the use of live attenuated varicella vaccine committee on infectious diseases the online version of this article , along with updated information and services , is located on the world wide web at : illinois ,

41
60007 . Copyright © 1995 by the american academy of pediatrics . All rights reserved . Print recommendations for the use of live varicella vaccine. 1995;
81. Feusner, j. H., s. J. Slichter, and l. A. Harker. 1979. Mechanisms of thrombocytopenia in varicella. Am. J. Hematol. 7:255–262.
82. Arvin, am.; gilden, d. Fields virology. 6. Knipe, d.; howley, p., editors. Lippincott williams&wilkins; 2013. P. 2015-2057.
83. Zerboni, leigh, et al. "molecular mechanisms of varicella zoster virus pathogenesis." nature reviews microbiology 12.3 (2014): 197-210.
84. moffat jf, et al. The orf47 and orf66 putative protein kinases of varicella-zoster virus determine tropism for human t cells and skin in the scid-hu ouse. Proc natl acad sci usa. 1998; 95:11969–11974. [pubmed: 9751774].
85. moffat jf, et al. Attenuation of the vaccine oka strain of varicella-zoster virus
and role of glycoprotein c in alphaherpesvirus virulence demonstrated in the scid-hu mouse. J virol. 1998; 72:965–974. This study is the first report to model vzv pathogenesis using t cell and skinxenografts in scid mice. [pubmed: 9444989].
86. Ku cc, et al. Tropism of varicella-zoster virus for human tonsillar cd4+ t lymphocytes that express activation, memory and skin homing markers. J virol. 2002; 76:11425–11433. [pubmed:12388703]
87. Moffat jf, et al. The orf47 and orf66 putative protein kinases of varicella-zoster virus determine tropism for human t cells and skin in the scid-hu mouse. Proc natl acad sci usa.1998; 95:11969–11974. [pubmed: 9751774].
88. Schaap a, et al. T-cell tropism and the role of orf66 protein in pathogenesis of varicella-zostervirus infection. J virol. 2005; 79:12921–12933. [pubmed: 16188994].
89. koropchak cm, et al. Investigation of varicella-zoster virus infection of lymphocytes by in situhybridization. J virol. 1989; 63:2392–2395. [pubmed: 2539528].
90. Ozaki t, et al. Viremic phase in nonimmunocompromised children with varicella.
J pediatr. 1984;104:85–87. [pubmed: 6317835].
91. Arvin am, et al. Varicella-zoster virus t cell tropism and the pathogenesis of skin infection. Currtop microbiol immunol. 2010; 342:189–209. [pubmed: 20397071].
92. Loparev, v.n., argaw, t., krause, p.r., takayama, m., schmid, d.s., 2000. Improved identification and differentiation of varicella–zoster virus (vzv) wild-type strains and an attenuated varicella vaccine strain using a vzv open readingframe 62-based pcr. J. Clin. Microbiol. 38, 3156–3160.

42
93. Adams, s. G., d. E. Dohner, and l. D. Gelb. 1989. Restriction fragment differences between the genomes of the oka varicella vaccine virus andamerican wild-type varicella-zoster virus. J. Med. Virol. 29:38–45.
94. Larussa, p.s., gershon, a.a., 2001. Biologic and geographic differences betweenvaccine and clinical varicella–zoster virus isolates. Arch. Virol. Suppl. 41–48.
95. Chou, s., and k. M. Dennison. 1991. Analysis of interstrain variation in cytomegalovirus glycoprotein b sequences encoding neutralizing epitopes.j. Infect. Dis. 163:1229–1234.
96. Cone, r., m. L. Huang, r. C. Ackman, and l. Corey. 1996. Coinfection with
human herpesvirus 6 variants a and b in lung tissue. J. Clin. Microbiol.34:877–881.
97. Davis, c. L., d. Field, d. Metzgar, r. Saiz, p. A. Morin, i. L. Smith, s. A.spector, and c. Wills. 1999. Numerous length polymorphisms at short tandemrepeats in human cytomegalovirus. J. Virol. 73:6265–6270.
98. Franti, m., j.-t. Aubin, l. Poirel, a. Gautheret-dejean, d. Candotti, j.-m. Huraux, and h. Agut. 1998. Definition and distribution analysis of glycoproteinb gene alleles of human herpes virus 7. J. Virol. 72:8725–8730.
99. Meng, y. X., t. J. Spira, g. J. Bhat, c. J. Birch, j. D. Druce, b. R. Edlin, r. Edwards, c. Gunthel, r. Newton, f. R. Stamey, c. Wood, and p. E. Pellett. 1999. Individuals from north america, australasia, and africa are infectedwith four different genotypes of human herpesvirus 8. Virology 261:106–119.
100. Norberg, p., liljeqvist, j.a., bergstro¨m, t., sammons, s., schmid, d.s., loparev, v.n., 2006. Complete-genome phylogenetic approach to varicella–zoster virus evolution: genetic divergence and evidence for recombination. J. Virol. 80, 9569– 9576.
101. Peters, g.a., tyler, s.d., grose, c., severini, a., gray, m.j., upton, c., tipples, a., 2006. A full-genome phylogenetic analysis of varicella–zoster virus reveals a novel origin of replication-based genotyping scheme and evidence of recombinationbetween major circulating clades. J. Virol. 80, 9850–9860.
102. Schmidt-chanasit, j., olschla¨ger, s., gu¨ nther, s., jaeger, g., bleymehl, k., scha¨d, s.g., heckel, g., ulrich, r.g., doerr, h.w., 2008b. Molecular analysis of varicella– zoster virus strains circulating in tanzania demonstrating the presence ofgenotype m1. J. Clin. Microbiol. 46, 3530–3533.
103. Mcgeoch, d.j., 2009. Lineages of varicella–zoster virus. J. Gen. Virol. 90, 963–969.
104. Santos, r. A., c. C. Hatfield, n. L. Cole, j. A. Padilla, j. F. Moffat, a. M. Arvin, w. T. Ruyechan, j. Hay, and c. Grose. 2000. Varicella-zoster virus ge escape
43
mutant vzv-msp exhibits an accelerated cell-to-cell spread phenotypein both infected cell cultures and scid-hu mice. Virology 275:306–317.
105. Ibge – instituto brasileiro de geografia e estatistica. Disponível em: <http:/www.ibge.gov.br>. Acesso em: 20 out. 2015.
8 ANEXOS GENES VZV
VZV gene products
VZV gene
HSV-1 homolog
VZV protein function
1 None Membrane protein
2 None
3 UL 55
4 UL 54 Transactivator, tegument protein, expressed in latency
5 UL 53 gK
6 UL 52
7 UL 51
8 UL 50 Deoxyuridine triphosphatase
9 UL 49 Tegument protein
9ª UL 49 Syncytia formation, putative Gn
10 UL 48 Transactivator, tegument protein
11 UL 47
12 UL 46
13 None Thymidylate synthetase
14 UL 44 Gc
15 UL 43
16 UL 42 Putative small subunit of viral DNA polymerase
17 UL 41 Induces RNA cleavage
18 UL 40 Ribonucleotide reductase, small subunit
19 UL 39 Ribonucleotide reductase, large subunit
20 UL 38

44
21 UL 37 Nucleocapsid protein, expressed in latency
22 UL 36
23 UL 35 Capsid assembly
24 UL 34
25 UL 33
26 UL 32
27 UL 31
28 UL 30 DNA polymerase
29 UL 29 ssDNA binding protein, expressed in latency
30 UL 28
31 UL 27 Gb
32 None Probable substrate for ORF47 kinase
33 UL 26 Protease
33.5 UL 26.5 Assembly protein
34 UL 25
35 UL 24 Cell-to-cell fusion
36 UL 23 Thymidine kinase
37 UL 22 Gh
38 UL 21
39 UL 20
40 UL 19 Major nucleocapsid protein
41 UL 18
42/45 UL 15
43 UL 17
44 UL 16
46 UL 14
47 UL 13 Protein kinase, tegument protein
48 UL 12 Putative Dnase
49 UL 11 Virion protein
50 UL 10 Gm
51 UL 9 Origin binding protein
52 UL 8
53 UL 7
54 UL 6 Putative portal protein
55 UL 5
56 UL 4
57 None Cytoplasmic protein
58 UL 3
59 UL 2 Uracil-DNA glycosylase
60 UL 1 Gl, chaperone for Gh
61 ICP 0 Transactivator, transrepressor
62,71 ICP4 Transactivator, tegument protein, expressed in latency
63,7 US1 (ICP22) Tegument protein, transrepressor, inhibits interferon- alpha, expressed in latency
64,69 US10
65 US9 Virion protein
66 US3 Protein kinase, expressed in latency

45
67 US7 (Gi) Gi
68 US8 (Ge) Ge
S/L None Cytoplasmic protein (also referred to as ORF0)
Fonte: ANDREW J. DAVISON et al.,1986.




![[PPT]FAMILIA: HERPESVIRIDAE - CAO-Blog 4toAño ... · Web viewFAMILIA: HERPESVIRIDAE ESTRUCTURA: ADN CAPSIDE ENVOLTURA 162 CAPSOMER ESPIGAS DE GLICOPROTEINAS 150 – 300 NM LINEAL](https://img.pdfslide.net/doc/110x75/5ae5426c7f8b9ae1578c114a/pptfamilia-herpesviridae-cao-blog-4toao-viewfamilia-herpesviridae-estructura.jpg)