Embed Size (px)
Citation preview

اىجمسح اىجضائشح اىذمقشاطح اىشعثح
اىعيمصاسج اىرعيم اىعاى اىثذس
République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la recherche scientifique
Université d’ORAN
Faculté des Sciences
Département de Biotechnologie
Mémoire de MAGISTER
En Biotechnologie
Option: Intérêt des microorganismes
en Agriculture et en Agro-alimentaire
Présentée par
TLEMSANI MOKHTARIA
Intitulé
Contribution à l'étude du flétrissement vasculaire du pois
chiche (Cicer arietinum L.) causé par Fusarium oxysporum
Schelcht. Emend. Snyd. & Hans. f. sp. ciceri (Padwick):
caractérisation, lutte biologique et comportement variétal
Soutenu le /11/2010 devant la commission d’examination :
Président:
Examinateurs:
Rapporteur:
Mme
Fortas Z.
M. Djibaoui R.
M. Kacem M.
M. Bellahcene M.
Professeur
Maitre de conférences
Maitre de conférences
Professeur
Université d'Oran
Université de Mostaganem
Université d'Oran
Université de Mostaganem

Dédicace
A mes parents pour les efforts et les sacrifices faits pour que je puisse réaliser ce
travail.
A ma sœur et mes frères: Fatiha, Mohammed, Nabil, Hocine en témoignage de leur
soutien et leurs encouragements incessants.
Pour les moments extraordinaires passés ensemble je tiens à remercier mes amis et mes
collègues: Gabed Noujoud, Kébayli Hakima, Bouslama Nabila, Fatima Zitouni, Fatima
Taharia, Sidhoum Warda, Harir Mohamed pour leur fidèle soutien pendant les années
d’études et pour tous les excellents souvenirs.
A tous ce que j’ai oubliés, mais qui se reconnaîtront ici.
Tlemsani Mokhtaria

Remerciement
Je tiens d’abord à adresser toute ma gratitude au Professeur Bellahcene M. sans
lequel ce travail n’aurait pu aboutir, je le remercie pour sa gentillesse, son soutien et pour le
fait de m’avoir fait partager son expérience, je lui adresse toute ma reconnaissance pour sa
patience, sa disponibilité et sa participation active lors de la rédaction de mémoire.
Je remercie vivement Madame le Professeur Fortas Z., chef d'équipe au laboratoire de
Biologie des Microorganismes et Biotechnologie à l'université d'Oran Es- Sénia, pour le fait
de m’avoir accueilli dans son équipe de laboratoire et pour ses conseils toujours pertinents et
son soutien moral et professionnel pendant le mémoire mais aussi pendant toutes les années
d’études.
Mes remerciements vont également à Monsieur Djibaoui R., Maitre de conférences à
l'université de Mostaganem et Monsieur Kacem M., Maitre de conférences à l'université
d'Oran Es- Sénia, qui ont acceptés d’examiner ce travail avec bienveillance.
J’exprime également mes remerciements et ma reconnaissance à Monsieur Labdi A.,
directeur de l'INRA de Sidi Bél-Abbès et Monsieur Zénai R., directeur de l'ITGC de Saida
pour m'avoir offert le matériel végétal nécessaire pour ce travail.
Je remercie également Monsieur Bekkar A. A., enseignant à l'université de Mascara,
de m'avoir aidé dans la réalisation de l'étude statistique.
Je remercie M. Bensehla, Professeur à l'université d'Oran, de m'avoir accueilli dans
son laboratoire, pendant la réalisation de l'étude biométrique.
Un grand merci pour Monsieur Miloudi, enseignant au département de chimie,
université d'Oran, qui m’a appris les méthodes d’analyse chimique de laboratoire nécessaires
pour ce travail, je leur adresse tout mon respect pour sa disponibilité et sa gentillesse.
Je tien à remercie M. Abdi Y, sous directeur de la station météorologique d'Oran, pour
m'avoir facilité l'obtention des données climatiques.
Je remercie vivement M. Djibaoui R. Maitre de conferences à l'université de
Mostaganem m'avoir aidé dans l'identification des actinommycètes.
Je leur serai toujours reconnaissante pour leur aide et leurs conseils et je pense
spécialement à Mmes
Bouchentouf L, Dib S, Mejdoub H, Tahri A, Tahari F, Kébayli H.
Enfin, que tous ceux, de prés ou de loin, dont l'assistance ou les conseils m'ont permis
de mener à bien ce travail, trouvent ici l'expression de ma profonde gratitude.

Résumé
Au cours des prospections, nous avons constaté que les attaques du flétrissement du
pois chiche dû à l’agent causal Fusarium oxysporum f. sp. ciceri dans 03 régions (Tlemcen,
Ain Temouchent et Mostaganem) peuvent prendre une grande ampleur lorsque les conditions
climatiques sont favorables. Ainsi, l'incidence et la gravité déterminées dans ce présent travail
sont élevées surtout dans les localités d’Ain Témouchent et Tlemcen. Les symptômes
manifestés ont consistés en un jaunissement tardif qui s'exprime sous forme d'un jaunissement
vasculaire progressif de la plante du bas en haut.
Parallèlement, les diverses techniques d’isolement et d’identification employées ont
permis d’élucider certains aspects morphologiques: culturales et biométriques chez 17 isolats
de FOC (Fusarium oxysporum f. sp. ciceri). L’étude des caractéristiques morphologiques a
permis de distinguer 03 morphotypes sauvages différents: cotonneux, ras muqueux, duveteux
et une variabilité dans l'aspect et la pigmentation entre les cultures jeunes et âgées. Ainsi
l'étude biométrique a révélée des variabilités dans la longueur et la largeur des microconidies
(5.55 - 8.44 x 2.28 -2.78 µm) et des macroconidies (18.2-29.64 x 2.81- 4.03 µm). L'étude
physiologique de 17 isolats de FOC nous a permis de déterminer que le milieu le plus
favorable pour la croissance de FOC est le milieu pois chiche et que la température optimale
pour ce parasite est autour de 30°C et un pH de 5.5 est le plus favorable. Nous avons
démontré aussi que l'obscurité apparait adéquate pour le FOC que l'incubation sous lumière
continue et ceci induit la pigmentation. Concernant la détermination de la source de carbone
nous avons remarqué que le glucose est la meilleur source, en revanche la peptone s'est révèle
être la meilleur source d'azote.
Le test de confrontation directe nous a permis de selectionner 06 souches
d'actinomycètes antagonistes vis-à-vis 03 isolats de FOC. Ainsi les tests de filtrats de culture
nous ont permis de confirmer la production des antibiotiques par ces isolats antagonistes.
L'identification microscopique et culturale nous a permis de les classer dans le groupe des
Streptomyces. L'isolat de Streptomyces ACT2 qui a présenté des pourcentages d'inhibition
non négligeables contre les 03 isolats de FOC en boite de Petri a été retenu pour des tests in
vivo. Ce test a été conduit sous serre en vue de lutter contre la fusariose du pois chiche en
utilisant 03 variétés (ILC-482, Col-27, PPC-25).

Les résultats obtenus par ce dernier test ont montré la capacité de l'isolat Streptomyces
ACT2 de réduire le degré de la maladie de la fusariose du pois chiche chez ces 03 variétés en
comparaison avec les témoins inoculés utilisés.
Le test de l'agressivité nous a permis d'identifier des indices de la maladie sur ILC-482
(variété connue par sa sensibilité au FOC) variaient entre (32.33 et 92.12%). Suite à ce
résultat nous avons classé: 06 isolats comme peu agressifs, 02 moyennement agressifs, 07
agressifs et 02 uniquement ont été classés comme très agressifs.
L'étude du comportement variétal de 11 variétés du pois chiche vis à vis le FOC a
révélée un comportement variable et a permis de sélectionner 04 variétés tolérantes qui sont:
F7-43, F10-13, Fedjoj, 2G-03.
Mots clés : Cicer arietinum L., Fusarium oxysporum f. sp. ciceri, flétrissement vasculaire,
Antagonisme, Actinomycètes, agressivité.

Liste des Tableaux
Tableau 1: Principaux pays producteurs du pois chiche dans le monde, campagne 2007-
2008…………………………………………………………………………………………….8
Tableau 2: Répartition géographique de la culture du pois chiche dans le monde………........9
Tableau 3: Rendement et production mondiale du pois chiche en comparaison avec d'autres
cultures, campagne 2004-2005………………………………………………………………10
Tableau4: Composition chimique des grains du pois chiche en comparaison avec quelques
légumineuses alimentaires et le blé ………………......................................................................10
Tableau 5: Composition chimique de pois chiche…………………………………………...11
Tableau6:Principales maladies cryptogamiques chez le pois chiche dans le monde………..12
Tableau7: Légumineuses alimentaires cultivées en Algérie, superficies, production,
rendements 'moyenne 1993-2002)……………………………………………………………14
Tableau 8: Evolution de la superficie, de la production et du rendement et semences du pois
chiche en Algérie durant la compagne 1998-2008de la culture du pois chiche………………15
Tableau9: Quelques publications sur l'utilisation des actinomycètes en lutte biologique contre
les pathogènes telluriques…………………………………………………………………….33
Tableau 10: Résistance multiple chez Cicer sp……………………………………………...42
Tableau11: Principales espèces de Cicer utilisées dans les programmes d'amélioration de la
résistance du pois chiche contre le flétrissement vasculaire………………………………….43
Tableau 12: Températures en degrés °C et pluviométrie en mm mensuelles des chefs lieux
des wilayas d'étude en 2009…………………………………………………………………..45
Tableau13: Parcelles prospectées des 03 régions choisies…………………………………..46
Tableau14: Répartition des isolats de FOC isolés à partie de tiges du pois chiche………….49
Tableau15: Sites d'échantillonnage des sols pour l'isolement des actinomycètes…………...56

Tableau16: Incidence et gravité du flétrissement vasculaire dans les parcelles de la culture de
pois chiche prospectées……………………………………………….....................................67
Tableau 17. Nombres des isolats obtenus par région et par partie de plantes infectés
………………………………………………………………………………………………...69
Tableau 18: Caractéristiques culturales des morphotypes sauvages de 17 isolats de
FOC…………………………………………………………………………………………...78
Tableau 19: Mensurations en µm des conidies……………………………………................82
Tableau 20: Nombre des souches d'actinomycètes isolés sur milieu ISP2………................120
Tableau 21: Pourcentages d'isolement des actinomycètes à partir des deux dilutions
effectuées……………………………………………………………………………………121
Tableau 22: Pourcentages d'inhibition de 06 isolats d'actinomycètes sur les 03 FOC……..124
Tableau 23: Diamètres d'inhibition de FOC par les filtrats………………………………...126
Tableau 24: Caractères culturaux des souches d'actinomycètes après 14 jours
d'incubation………………………………………………………………………………….134
Tableau 25. Degrés de suppression de la maladie par la souche ACT2 en utilisant 03 variétés
du pois chiche………………………………………………………………………………..139
Tableau 26: Classification de 17 isolats de FOC selon leur agressivité…………................151
Tableau 27: Réponse des cultivars du pois chiche vis à vis de FOC………………………152
Tableau 28: Résultats du test de réisolement des 03 FOC…………………………………154
Tableau29: Répartition des isolats de FOC en fonction des morphotypes, la pigmentation du
thalle, la croissance mycélienne, les mensurations des conidies et le degré d'agressivité…..156
Tableau 30: Matrice de corrélation calculée sur huit paramètres étudiés……......................157

Liste des figures
Figure 1: Plante du pois chiche……………..............................................................................5
Figure 2: Champ du pois chiche……………………………………………............................5
Figure 3: Description de la plante du pois chiche……………………………………………..5
Figure 4: Cycle infectieux de Fusarium oxysporum................................................................23
Figure 5: Distribution typique de plante du pois chiche infestés par le FOC………………..26
Figure 6: Symptôme de la fusariose du pois chiche………………………............................27
Figure 7: Observation microscopique des actinomycètes…....................................................30
Figure 8: Localisation des sites de prélèvement des isolats de FOC………………………...47
Figure 9:Cultures du pois chiche présentant des foyers malades…………………................65
Figure 10: Plants du pois chiche présentant une attaque de la fusariose…….. ……………..66
Figure11: Réduction des ramifications secondaires sur le système racinaire des plantes
malades du pois chiche par apport aux racines des plantes saines…………............................66
Figure 12. Colonies de Fusarium oxysporum f. sp. ciceri…………………………...............68
Figure 13. Histogrammes présentant la répartition des isolats de FOC par région et par partie
de plante…………………………………………………………………................................72
Figure 14: Morphotype du thalle de FOC cotonneux………………………………………..78
Figure 15: Morphotype du thalle de FOC duveteux sauvage………………………………..79
Figure 16: Morphotype du thalle de FOC ras muqueux……………………………………..79
Figure 17: Morphotype du thalle de FOC ras sénescent……………………………………..81
Figure 18: Observation microscopique de FOC……………………………………………..81
Figure 19: Observation microscopique des chlamydospores colorée au bleu de cotton sous
microscope optique x 2000…………………………………………………………………...81
Figure 20: Estimation de la croissance mycélienne en cm sur milieu de culture PDA (FOC1,
FOC2, FOC3, FOC4, FOC5, FOC6)………………………………………............................95
Figure 21: Estimation de la croissance mycélienne en cm sur milieu de culture PDA (FOC7,
FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………..............................96
Figure 22: Estimation de la croissance mycélienne en cm sur milieu de culture PDA (FOC13,
FOC14, FOC15, FOC16, FOC17)……………………………………………………………97

Figure 23: Influence de la température sur la croissance mycélienne des isolats de FOC
(FOC1, FOC2, FOC3, FOC4, FOC5, FOC6)………………………………………………...98
Figure 24: Influence de la température sur la croissance mycélienne des isolats de FOC
(FOC7, FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………………….99
Figure 25: Influence de la température sur la croissance mycélienne des isolats de FOC
(FOC13, FOC14, FOC15, FOC16, FOC17)………………………………………………...100
Figure 26: Influence du pH sur la croissance mycélienne de FOC (FOC1, FOC2, FOC3,
FOC4, FOC5, FOC6)………………………………………………………………………..101
Figure 27: Influence du pH sur la croissance mycélienne de FOC (FOC7, FOC8, FOC9,
FOC10, FOC11, FOC12)…………………………………………………............................102
Figure 28: Influence du pH sur la croissance mycélienne de FOC (FOC13, FOC14, FOC15,
FOC16, FOC17)…………………………………………………………..............................103
Figure 29: Influence de la lumière et de l'obscurité sur la croissance mycélienne de FOC
(FOC1, FOC2, FOC3, FOC4, FOC5, FOC6)……………………………………………….104
Figure 30: Influence de la lumière et de l'obscurité sur la croissance mycélienne de FOC
(FOC7, FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………………...105
Figure 31: Influence de la lumière et de l'obscurité sur la croissance mycélienne de FOC
(FOC13, FOC14, FOC15, FOC16, FOC17)………………………………………………...106
Figure 32: Influence de la source d'azote sur la croissance mycélienne de FOC (FOC1,
FOC2, FOC3, FOC4, FOC5, FOC6)………………………………………………………..107
Figure 33: Influence de la source d'azote sur la croissance mycélienne de FOC (FOC7,
FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………............................108
Figure 34: Influence de la source d'azote sur la croissance mycélienne de FOC (FOC13,
FOC14, FOC15, FOC16, FOC17)…………………………………………………………..109
Figure 35: Influence de la source de carbone sur la croissance mycélienne de FOC (FOC1,
FOC2, FOC3, FOC4, FOC5, FOC6)………………………………………………………..110
Figure 36: Influence de la source de carbone sur la croissance mycélienne de FOC (FOC7,
FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………............................111
Figure 37: Influence de la source de carbone sur la croissance mycélienne de FOC (FOC13,
FOC14, FOC15, FOC16, FOC17)…………………………………………………………..112
Figure 38: Influence des milieux de culture sur la croissance mycélienne de FOC (FOC1,
FOC2, FOC3, FOC4, FOC5, FOC6)…………………………………..................................113
Figure 39: Influence des milieux de culture sur la croissance mycélienne de FOC (FOC7,
FOC8, FOC9, FOC10, FOC11, FOC12)…………………………………............................114

Figure 40: Influence des milieux de culture sur la croissance mycélienne de FOC (FOC13,
FOC14, FOC15, FOC16, FOC17)…………………………………………………………..115
Figure 41: Diversité des actinomycètes isolés à partir des sols arides du sud
algérien………………………………………………………………………………………122
Figure 42: Pourcentages d'inhibition par série de couleurs d'actinomycètes ……................125
Figure 43: Résultats du test de confrontation entre les isolats de FOC et les souches
d'actinomycètes…...................................................................................................................125
Figure 44: Résultats du test de filtrat entre les isolats de FOC et les actinomycètes………127
Figure 45: Résultats du test de puits entre les isolats de FOC et les actinomycètes………..127
Figure 46: Résultats du test des disques imbibés entre les isolats de FOC et la souche
ACT2………………………………………………………………………………………...128
Figure 47: Coloration Gram+ de la souche ACT2 observée au microscope optique………132
Figure 48: Observation microscopique des colonies d'actinomycètes à l'aide du microscope
optique……………………………………………………………………………………….135
Figure 49: Résultat de la séparation de la molécule antibiotique…………………………..138
Figure 50: Résultats de l'effet protectif de la souche ACT2 chez le cultivar ILC-482 …….141
Figure 51: Résultats de l'effet protectif de la souche ACT2 chez le cultivar Col27 ……….142
Figure 52: Résultats de l'effet protectif de la souche ACT2 chez le cultivar PPC-25 ……..143
Figure 53: Résultats du test d'agressivités des isolats testés de FOC………………………150
Figure 54: Résultats du test de la résistance variétale, montrant 02 variétés tolérantes …...152
Figure 55:Test de la résistance variétale montrant symptôme de la fusariose chez la variété
ILC-482…...............................................................................................................................153
Figure 56: Photo de réisolement de FOC à partir des fragments de plants du pois
chiche………………..............................................................................................................153

Liste des abréviations
Cm: Centimètre
f. sp. : Forme spécialisée.
FOC: Fusarium oxysporum f. sp. Ciceri.
ICARDA: International Center for Agricultural in the Dry Areas.
Ha: Hectare.
ILC: International Legume Cicer.
ITGC: Institue Technique des Grande Culture.
INRA: Institut National de Recherches Agronomique
Mm: Millimètre.
Mt: Millions de tonnes.
µm: micromètre.
pH: Potentiel d'hydrogène.
Qx: Quintaux.
°C: Degré Celsius.
T: Témoin.
JC: Jésus Christ.
FAO: Food Agriculture Organisation.
Mg: Milligramme.
Gr: Gramme.
Prod.: production.
ADN: Acide Désoxyribonucléique.
G: Guanine.
C: Cytosine
IAA: Indole-3-Acétique Acide
R: Résistante
S: Sensible

Table des Matières
Introduction…………………………………………………………………………………...1
Première partie: Synthèse bibliographique
1. Plante hôte: pois chiche (Cicer arietinum L.)
1.1. Origine et Historique de pois chiche……………………………………………................4
1.2. Description de la plante de pois chiche…………………………………………................4
1.3. Cycle et saison de culture de pois chiche………………………………………………….5
1.4. Type de cultivars chez le pois chiche……………………………………………………...6
1.4.1. Macrosperma……………………………………………………………................6
1.4.2. Microsperma……………………………………………………………………….6
1.5. Classification et Taxonomie de Cicer arietinum L………………………..........................6
1.6. Caractéristiques agronomiques……………………………………………………………7
1.6.1. Les exigences climatiques…………………………………………………………..7
1.6.1.1. La température……………………………………………………………..7
1.6.1.2. La pluviométrie…………………………………………………………….7
1.6.1.3. La lumière………………………………………………………………….7
1.6.2. Les exigences édaphiques………………………………………………………….8
1.6.2.1. Type du sol………………………………………………………................8
1.6.2.2. Humidité…………………………………………………………................8
1.6.2.3. Nutrition minérale………………………………………………………….8
1.6.2.3. pH du sol………………………………………………………...................8
1.7. Répartition géographique, aire de culture et production de pois chiche dans le monde…..8
1.8. Intérêts de pois chiche
1.8.1. Intérêt économique………………………………………………………………….9
1.8.2. Intérêt agronomique……………………………………………….........................10
1.8.3. Intérêt nutritionnelles……………………………………………………………...10
1.9. Le stress biotique chez le pois chiche…………………………………………………...11
1.10. Situation de pois chiche en Algérie……………………………………..........................12
1.10.1. Intérêt culturale et importance des légumineuses en Algérie…………................13

1.10.2. Aire de culture et production de pois chiche en Algérie………………...............14
1.10.3. Problème phytosanitaire de pois chiche en Algérie……………………………...15
1.10.3.1. L'anthracnose…………………………………………………..................15
1.10.3.2. Pourriture racinaires……………………………………………................16
1.10.3.3. Complexe de flétrissement………………………………………………..16
2. Agent pathogène: Fusarium oxysporum f. sp. ciceri
2.1. Généralité sur le genre Fusarium………………………………………………………...17
2.2. Généralité sur Fusarium oxysporum……………………………………..........................17
2.3. Fusarium oxysporum f. sp. ciceri (FOC)………………………………………………...18
2.3.1. Historique de FOC………………………………………………………………….18
2.3.2. Présentation de FOC………………………………………………………………..19
2.3.3. Description et clés d'identification de FOC………………………...........................19
2.3.4. Classification, biologie et cycle de reproduction du champignon………………….20
2.3.5. Mécanismes d'infection et de colonisation
2.3.5.1. Mode d'infection…………………………………………….......................21
2.3.5.2. Mode de colonisation……………………………………………................21
2.3.6. Inoculum primaire, mode de survie et de transmission de FOC…………………...22
2.3.7. Gamme d'hôte……………………………………………………………...............24
2.3.8. Races physiologiques et pathotypes de FOC……………………............................24
2.3.9. Distribution de la maladie et des races pathologiques dans le monde……………..25
2.3.10. Spécialisation et résistance chez le FOC…………………………........................25
2.3.11. Origine de FOC et source de variabilité………………………………………….25
2.4. La maladie de la fusariose de pois chiche…………………………………......................26
2.4.1. Dégâts causés par la fusariose de pois chiche……………………………………..26
2.4.2. Description des symptômes de la fusariose de pois chiche……….........................27
2.4.3. Localisation de la fusariose de pois chiche………………………..........................28
2.4.4. Répartition géographique et importance économique du flétrissement en
Algérie………………………………………………………………………………………...28
2.4.5. Facteurs influençant la maladie du flétrissement………………………................28
3. L'agent antagoniste (les actinomycètes)
3.1. Les actinomycètes
3.1.1. Présentation et caractéristiques des actinomycètes………………………………..30
3.1.2. Ecologie des actinomycètes……………………………………………………….30
3.1.3. Importance économiques des actinomycètes……………………………………...31
3.1.4. Taxonomie des actinomycètes…………………………………………………….32
3.1.5. Utilisation des actinomycètes en lutte biologique………………………................32
3.1.6. Mode d'action des actinomycètes…………………………………….....................34

3.1.7. Possibilité d'utilisation des actinomycètes contre le FOC………………………...34
3.2. Le genre Streptomyces
3.2.1. Caractéristiques du genre Streptomyces…………………………………………...35
3.2.2. Cycle de différentiation des Streptomyces………………………………………...35
3.2.4. Classification des Streptomyces…………………………………………………...36
3.2.5. Les avantages de l'utilisation des Streptomyces en lutte biologique……................36
4. Moyens de lutte
4.1. Pratiques culturales……………………………………………………………................38
4.2. Méthodes chimiques……………………………………………………………………..39
4.3. Méthodes biologiques…………………………………………………………................39
4.3.1. Utilisation des microorganismes…………………………………………………..40
4.3.2. Méthodes génétiques
4.3.2.1. La résistance variétale……………………………………………………..40
4.3.2.2. Méthodes de défense naturelle…………………………………………….41
4.3.2.3. Résistance induite…………………………………………………………42
4.3.2.4. Utilisation des espèces sauvages de Cicer sp……………………………...42
4.3.3. La lutte intégrée………………………………………………………...................44
4.4. Application du phénomène d'antagonisme et ses limites………………………………...44
Deuxième Partie: Matériels et méthode
1. Caractérisation des symptômes de la fusariose et identification de l'agent causal
(FOC)
1.1. Les
prospections…………………………………………………………………………………..45
1.2. Estimation de l'indice et de la gravité de la maladie…………………………..................46
1.3. Obtention du matériel fongique……………………………………………….................47
1.3.1. Isolement de l'agent causal………………………………………………................48
1.3.2. Purification et conservation de l'agent pathogène………………………................48
1.4. Détermination de la forme spéciale ciceri…………………………………….................49
1.4.1. Préparation du matériel fongique………………………………………................50
1.4.2. Préparation du matériel végétal…………………………………………………...50
1.4.3. Technique d'inoculation des plantules du pois chiche…………………................50
1.4.4. Notation des symptômes………………………………………………………….51

1.5. Caractérisation morphologique de l'agent pathogène……………………………………51
1.5.1. Caractérisation culturale et description des morphotypes………………………...51
1.5.2. Etude biométrique des conidies…………………………………………………..52
1.6. Etude physiologique des isolats de FOC………………………………………...............52
1.6.1. Recherches des conditions optimales de croissance mycélienne des isolats……..52
1.6.1.1. Estimation de la croissance de FOC………………………………………52
1.6.1.2. Effet de l'obscurité et la lumière sur le FOC…………………………….53
1.6.1.3. Effet de la température………………………………..................53
1.6.1.4. Effet du pH……………………………………………................53
1.6.2. Influence des facteurs nutritionnels sur la croissance de FOC…………………...53
1.6.2.1. Effet des milieux de cultures………………………………….................54
1.6.2.2. Influence de la source de carbone sur le FOC…………………………….54
1.6.2.3. Influence de la source d'azote sur le FOC………………………………...55
1.7. Répétition et traitement statistique…………………………………………….................55
2. Recherche de moyens de lutte
2.1. Obtention des bactéries actinomycètes………………………………………..................55
2.1.1. Prélèvement des échantillons du sol……………………………………………….56
2.1.2. Isolement, purification et conservation des actinomycètes………………………...56
2.1.2.1. Isolement……………………………………………………........................56
2.1.2.2. Purification des isolats d'actinomycètes……………………......................56
2.1.2.3. Conservation des isolats d'actinomycètes………………………................57
2.2. Etude du phénomène d'antagonisme FOC/ Actinomycètes……………………………...57
2.2.1. Méthode de confrontation directe………………………………………………….57
2.2.2. Mise en évidence de l'action des métabolites d'actinomycètes…………………….58
2.2.2.1. Test de filtrat de culture…………………………………………………….58
2.2.2.1.1. Préparation des filtrats de culture d'actinomycètes………………..58
2.2.2.1.2. Incorporation des filtrats de culture d'actinomycètes……………...59
2.2.2.2. Technique de puits………………………………………………………….59
2.2.2.3. Technique des disques du papier Wattman…………… …….......................59
2.2.2.3.1. Préparation des filtrats de culture d'actinomycètes….......................59
2.2.2.3.2. Extraction de la molécule antifongique……………………………60
2.2.2.3.3. Test de l'activité de l'extrait………………………….......................60
2.3. Identification des isolats d'actinomycètes antagonistes………………………………...60
2.3.1. Etude macroscopique……………………………………………………………….61
2.3.2. Etude microscopique………………………………………………………………..61
2.4. Essai d'extraction et de séparation de la molécule antifongique………………................61

2.4.1. Préparation des filtrats de culture des actinomycètes……………………………...61
2.4.2. Extraction liquide-liquide…………………………………………….....................61
2.4.3. Chromatographie sur couche mince……………………………………..................61
2.5. Test de lutte biologique in vivo………………………………………………................62
2.5.1. Préparation de l'inoculum…………………………………………………………..62
2.5.2. Obtention des plantules du pois chiche…………………………………………….62
2.5.3. L'estimation des symptômes………………………………………………………63
3. Détermination de l'agressivité et du comportement variétal
3.1. Détermination de l'agressivité…………………………………………………………..63
3.2. Etude du comportement variétal………………………………………………………..63
3.2.1. Estimation des symptômes………………………………………….......................64
3.2.2. Réisolument du champignon pathogène……………………………......................64
Troisième Partie: Résultats et discussions
1. Caractérisation des symptômes de la fusariose et identification de l'agent causal
(FOC)
1.1. Importance du flétrissement du pois chiche dans les 03 régions d'études
1.1.1. Résultats
1.1.1.1. Observation des symptômes au champ…………………………………….65
1.1.1.2. Etude de l'indice et de la gravité de la maladie…………………………….67
1.1.1.3. Détection et isolement de l'agent pathogène……………………………….68
1.1.1.4. Détermination de la forme spéciale ciceri…………………………………70
1.1.2. Discussion
1.1.2.1. Observation des symptômes………………………………………………..70
1.1.2.2. Indice et gravité de la maladie……………………………………………..72
1.2.2.3. Détection et isolement de l'agent pathogène……………………………….74
1.1.2.4. Détermination de la forme spéciale ciceri…………………………………75
1.2. Etude morphologique de FOC
1.2.1. Résultats
1.2.1.1. Caractérisation physique…………………………………………………...77
1.2.1.1.1. Caractéristiques culturales…………………………......................77
1.2.1.1.1.1. Description des morphotypes rencontrés……………..77
1.2.1.1.1.2. Pigmentation du thalle………………………………..80
1.2.1.1.2. Caractéristiques microscopiques et biométriques des spores…….80
1.2.1.2.1.1. Etude microscopique de FOC…………......................80
1.2.1.2.1.2. Etude biométrique……………....................................82

1.2.2. Discussion
1.2.2.1. Caractérisation physique……………………………………………….83
1.2.2.1.1. Caractéristiques culturales…………………………...............83
1.2.2.1.2. Etude microscopique et biométrique
1.2.2.1.2.1. Etude microscopique……………………………..87
1.2.2.1.2.2. Etude biométrique………………………………..89
1.2.1.2. Résultats de la caractérisation physiologique………………………….91
1.2.1.2.1. Recherche des conditions optimales………………………….91
1.2.1.2.1.1. Estimation de la croissance mycélienne de 17
FOC…………………………………………………………………………………...91
1.2.1.2.1.2. Effet de la température…………………………….92
1.2.1.2.1.3. Effet du pH…………………...................................92
1.2.1.2.1. 3.Effet de la lumière et de l'obscurité……………......93
1.2.1.2.2. Influence des facteurs nutritionnelle sur la croissance de
FOC…………………………………………………………………………………...93
1.2.1.2.2.1. Effet du milieu de culture………………………..93
1.2.1.2.2.2. Effet de la source de carbone……………………93
1.2.1.2.2.3. Effet de la source d'azote………………………...94
1.2.1.3. Discussion des résultats de la caractérisation physiologique
1.2.1.3.1. Recherche des conditions optimales…………………………116
1.2.1.3.1.1. Estimation de la croissance mycélienne…………..116
1.2.1.3.2. Effet de la température…………................116
1.2.1.3.3. Effet du pH………………………………..117
1.2.1.3.4. Effet de l'action de la lumière et de
l'obscurité……………………………………………………………………………………118
1.2.1.3.2. Influence des facteurs nutritionnels sur la croissance du
FOC………………………………………………………………………………………….118
1.2.1.3.2.1. Effet des milieux de cultures…………................118
1.2.1.3.2.2. Effet de la source de carbone……………………118
1.2.1.3.2.3. Effet de la source d'azote………………………...119
2. Recherche de moyens de lutte biologique
2.1. Isolement et étude de la diversité des actinomycètes
2.1.1. Résultats
2.1.1.1. Isolement des actinomycètes…………………………………………….125
2.1.1.2. Caractéristiques des actinomycètes isolés…………………………..126
2.1.2. Discussion
Etude d'antagonisme FOC/ Actinomycète

2.2.1. Résultats
2.2.1.1. Tests de confrontation directe……………………………................129
2.2.1.2. Test des filtrats……………………………………………………...131
2.2.2.3. Test des puits……………………………………………………….132
2.2.1.4. Test des disques imbibés…………………………………................132
2.2.2. Discussion
2.2.2.1. Tests de confrontation directe………………………………………..133
2.2.2.2. Test des filtrats……………………………………………………….136
2.2.2.3. Test des disques imbibés…………………………………..................137
2.3. Identification des actinomycètes présélectionnés
2.3.1. Résultats
2.3.1.1. Etude des caractères culturaux……………………………………….138
2.3.1.2. Observations microscopiques………………………………………..141
2.3.2. Discussion
2.3.2.1. Etude des caractères culturaux………………………………………..141
2.3.2.1. Observation microscopique…………………………………………...142
2.4. Essai d'extraction de la molécule antibiotique………………………………............144
2.5. Essai d'antagonisme sous serre……………………………………………………….146
2.5.1. Résultats
2.5.2. Discussion
3. Etude de l'agressivité des isolats de FOC et du comportement variétal……………..159
3.1. Résultats
3.1.1. Etude de l'agressivité…………………………………………………………………159
3.1.2. Etude du comportement variétal……………………………………………...............162
3.2. Discussion
3.2.1. Détermination de l'agressivité ………………………………………………………..165
3.2.2. Etude du comportement variétal……………………………………………...............165

4. Relation entre l'agressivité, les morphotypes, la pigmentation du thalle, le diamètre de
la croissance mycélienne, les mensurations des conidies………………………………...121
4.1. Résultats
4.1.1. Relation entre l'agressivité, la morphologie et la pigmentation du thalle…………….121
4.1.2. Relation entre l'agressivité et le diamètre de croissance du thalle……........................122
4.1.3. Relation entre l'agressivité et les mensurations des conidies…………........................122
4.2. Discussion
Conclusion et perspectives………………………………………………………................166
Références bibliographiques
Annexes

Introduction

Introduction
Introduction
Les légumineuses alimentaires représentent de part la superficie qu'elles occupent, une
place importante dans le système agraire et l'agroéconomie de nombreux pays du monde, en
raison des caractéristiques biologiques des racines dans la fixation de l'azote atmosphérique
par les nodosités bactériennes (Bacha et Ounane, 2003) et leur capacité d'adaptation à des
conditions pédoclimatiques difficiles, ainsi que leur faible exigence culturale. Du point de vue
nutritionnel, la richesse des légumineuses en protéines permet de corriger dans une certaine
mesure les carences en protéines animales, ainsi que le déséquilibre alimentaire des
populations qui ont tendance à ce nourrir exclusivement de céréales. Selon Obaton, (1980), un
hectare de légumineuses alimentaires produit un tonne de protéines, soit 10 fois plus qu'une
production d'un élevage à viande sur la même surface.
Les légumineuses alimentaires regroupent plusieurs espèces, parmi celles-ci, nous
avons le pois chiche (Cicer arietinum L.), dont la production et l’exploitation répondent à
divers fonctions, telles que : la consommation humaine, l’alimentation animale et la
fertilisation des sols notamment dans les pays en voie de développement. L'intérêt du pois
chiche réside dans leur teneur élevée en protéines, de sa haute valeur nutritive en complément
celle des céréales. Le pois chiche joue aussi un rôle important dans les systèmes de cultures
en contribuant à l'amélioration de la fertilité du sol par les reliquats d'azote qu'ils laissent et en
font ainsi un excellent précédent cultural (Bacha et Ounane, 2003).
L’Algérie, comme beaucoup de pays en voie de développement attribue une place de
choix à cette culture dotée d’une bonne valeur nutritive, les légumes secs telles que le pois
chiche, la lentille et le petit pois se placent après les céréales. Malgré les efforts déployés, la
production nationale reste encore très insuffisante. Cette insuffisance est liée à
l’accroissement démographique, à une stagnation des superficies de culture et aux problèmes
phytosanitaires (Toulaiti, 1988). De ce fait, chaque année l’Algérie à recours aux importations
pour satisfaire les besoins de la population.
Le pois chiche souffre de nombreuses difficultés, en dehors des conditions
d’environnement et le non maitrise des techniques culturales qui sont des causes non
négligeables de la faiblesse de la production, il semble que le problème majeur reste celui de
l’aspect phytosanitaire souvent attribué à des maladies fongiques telles que l’anthracnose, les
pourritures racinaires et le flétrissement.

Introduction
Ces différents problèmes associés à l'immense besoin en protéines dans l'alimentation
humaine ont amené les chercheurs à s'intéresser à cette légumineuse alimentaire.
Il est donc clair que l’amélioration des rendements est liée à une bonne connaissance
de ces maladies et leurs agents causals. Actuellement, les producteurs sont confrontés à une
recrudescence de problèmes pathologiques associés à la présence de Fusarium oxysporum.
Mais des doutes quant à la caractérisation des symptômes n’ont pas permis d’identifier la
forme spéciale impliquée dans le développement de cette épidémie affectant l’ensemble des
régions productrices en Algérie. Cette maladie constitue une énorme contrainte pour la
culture, cela dans de nombreuses régions où elle est pratiquée. Néanmoins, on connait peu
d’informations sur l’étiologie et l’importance de cette maladie en Algérie. Seuls des
observations préliminaires et fragmentaires ont suggéré qu’il peut s’agir d’un complexe
semblable à celui du flétrissement et de la pourriture racinaire, qui réduit sévèrement la
production du pois chiche dans la majorité des régions où cette culture est pratiquée.
Dans le cadre de notre étude, étant donné de nombreuses lacunes dans les
connaissances à la fois de l'agent pathogène, des processus pathologiques qu'il induit lors de
l'attaque du pois chiche ainsi que les moyens de lutte, nos nous somme intéressé au parasite,
sous un triple aspect: son caractérisation morphologique et physiologique, les moyens de lutte
et l'étude du comportement variétal.
Le premier chapitre de notre travail correspond à une mise au point bibliographique
sur le pois chiche, l’agent causal du flétrissement (Fusarium. oxysporum f. sp. ciceri), l'agent
antagoniste (Les actinomycètes) et les moyens de lutte contre cette maladie. Le deuxième
chapitre correspond à notre étude expérimentale, l'étape initiale a consisté à regrouper, une
collection d'isolats afin qu'elle puisse, raisonnablement, être considéré comme représentative
de la diversité. La collection de FOC, nous a servira de support à l'ensemble de nos
recherches que nous avons structurées de la manière suivante:
1. Caractérisation morphologique, biométrique et physiologique
Après avoir observé une attaque de ce champignon dans différentes régions où cette
culture est bien développée, nous nous sommes efforcés d’apporter une contribution à la
connaissance de la maladie.
Au cours des prospections effectuées, nous nous sommes intéressés à l’estimation de
l’importance de la maladie dans différentes régions où la culture du pois chiche est pratiquée

Introduction
et une identification de l’agent pathogène par un biotest sur plantule d'une variété sensible en
conditions contrôlées à fin de vérifier le Postulat de Koch.
Cette approche nous a conduits également à déterminer quelques caractéristiques
morphologiques (morphotype et pigmentation du thalle), biométriques (mensurations des
conidies produites en culture) de FOC et sa croissance mycélienne en boite de Petri en jouant
sur différents facteurs afin de rechercher les conditions optimales qui favorisent sa croissance
et son développement.
2. Etude du phénomène d'antagonisme entre FOC/Actinomycètes
Nous avons réalisé des essais de confrontations in vitro entre l’agent causal et des
espèces d’actinomycètes isolées des milieux extrêmes, dans le but de sélectionner quelques
germes antagonistes. Un autre test réalisé in vivo entre l'agent pathogène et une souche
d'actinomycète antagoniste dans le but d'évaluer le degré protectif des plantes du pois chiche
par cette souche antagoniste.
3. Etude du comportement variétal
Dans ce chapitre, nous nous sommes intéressés à l'étude du comportement de 17
isolats de FOC vis–à-vis une variété sensible afin de déterminer l'agressivité de nos isolats, en
suite nous avons procédé à l'étude du comportement variétal de 11 variétés du pois chiche vis-
à-vis le FOC dans le but de sélectionner quelques cultivars tolérants.

Chapitre 1: Synthèse bibliographique

Synthèse bibliographique
17
1. Plante hôte : Pois chiche (Cicer arietinum L.)
1.1. Origine et Historique du pois chiche (Cicer arietinum)
le pois chiche (Cicer arietinum L.) est originaire du Sud Est de la Turquie
(Ladisinsky, 1975) et le Nord Est de la Syrie (Staginnus et al., 1999). Il a été cultivé
pour la première fois à environ 7000 - 8000 avant JC (Sharma et Muehlbauer, 2007;
Staginnus et al., 1999). Le nom Cicer est originaire du latin, dérivé du mot grec
« Kikus » qui désigne fort ou solide (Singh et Diwakar, 1995). Duschak, (1871) a
signalé que l’origine du mot Cicer est Hébreu « Kirkes », où le mot « Kikar » désigne
rond. Le mot « arietinum » est aussi latin, traduit du mot grec « Krios », une allusion
de la forme des grains qui ressemblent à la tête de bélier (Van Der Maesen, 1987). Le
pois chiche est appelé garbenzo en espagnol, Kichar ou Chicher en allemand, chana
en Hindi et chickpea, gram ou bengal gram en anglais (Singh et Diwakar, 1995), il est
appelé « nakhut » ou « nohut» en Turquie, Romanie, Bulgarie et Afghanistan (Van
Der Maesen, 1987).
Le pois chiche a été utilisé en alimentation et en médicine par Homer dans
l’Iliade (1000-800 avant JC) à Rome, en Inde et dans la littérature européenne du
moyen âge (Van Der Maesen, 1972). La culture a été propagée avec le groupe
fondateur de l'agriculture de croissant fertile vers l’Europe et l’Asie de l'Ouest depuis
5500 avant JC (Harris, 1998; Harlan, 1992). Le pois chiche a été disséminé depuis ce
temps pour devenir une culture importante des environnements subtropicaux et la
région méditerranéenne (Muehlbauer et Rajesh, 2008; Kumar et Abbo, 2001; Zohary
et Hopf, 2000). Les pois chiches de types Kabuli ont été transféré vers l’Inde depuis la
région méditerranéenne dans le 18ème
siècle et les types Desi ont été importé au Kenya
par les émigrés indiens durant le 19eme
siècle (Van Der Maesen, 1972). Le pois chiche
a été transféré vers les deux Amériques par les espagnoles et les portugais
(Muehlbauer et Rajesh, 2008).

Synthèse bibliographique
18
1.2. Description de la plante du pois chiche (Cicer arietinum L.)
Le pois chiche est une plante annuelle, herbacée avec des branches diffusées et
propagées (Muehlbauer et Rajesh, 2008; Yadav et al., 2007; Staginnus et al., 1999;
Labdi et al., 1996).
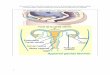
Figure 1. Plante de pois chiche (Patankar, 2000) Figure 2. Champ de pois chiche (Patankar, 2000)
Figure 3. Description de la plante du pois chiche (Singh et Diwakar, 1995)
Bourgeon de
fleur
Feuilles
Gousse
Pédoncule
Pédicelle
Branches I aires
Tige
Nodules
Racines

Synthèse bibliographique
19
1.3. Cycle et saison de la culture du pois chiche
Dans le bassin méditerranéen, le pois chiche est considéré comme une
culture du printemps. La plante se développe vigoureusement et complète son cycle
évolutif en 04 mois (El-Aoufir, 2001). La pollinisation chez le pois chiche est
complétée dans le stade de la formation des bourgeons des fleurs, avant que les
abeilles visitent les fleurs ouvertes aux champs (Van Der Maesen, 1972).
Certains cultivars du pois chiche à maturité précoce peuvent compléter leur
cycle de vie dans 65 jours. Cependant les cultivars à maturité tardée demandent
environ 120 jours.
Les variétés du pois chiches cultivés en hiver peuvent demander plus que 180
jours de la date de plantation jusqu'à la maturité (Muehlbauer et Rajesh, 2008).
1.4. Type de cultivars chez le pois chiche
L’espèce Cicer arientinum L., présente une variabilité phénotypique et
génotypique, ce dernier se divise en 02 types: Kabuli et Desi (Singh, 1985). Les pois
chiches de types Desi sont cultivés dans le sud de l'Asie où il représente la majorité de
la production. Cependant les types Kabuli dominent la production dans la plupart des
autres régions et spécialement dans l'hémisphère de l'Ouest (Muehlbauer et Rajesh,
2008). Les deux types sont destinés à la consommation humaine (Malhotra et al.,
1987; Muehlbauer et Singh, 1987).
1.4.1. Macrosperma (Kabuli type)
Ce type est cultivé dans la région méditerranéenne (Ohri et Pal, 1991) et
couvre 15% de la surface réservée au pois chiche (Babar et al., 2009; Muehlbauer et
Rajesh, 2008; Singh et Jauhar, 2005; Singh et Diwakar, 1995).
1.4.2. Microsperma (Desi type)
Il est cultivé principalement dans le subcontinent Indien (Ohri et Pal, 1991) et
compte environ 85% de la surface du pois chiche, il a souvent une petite forme (Erler
et al., 2009; Babar et al., 2009; Singh et Jauhar, 2005; Iruela et al., 2002; Singh et
Diwakar, 1995 ; Singh, 1985). Quelques auteurs ajoutent un troisième type qui est le

Synthèse bibliographique
20
type Culabi qui se caractérise par des graines lisses, de couleur claire, ressemblant au
pois, mais avec un bec caractéristique (Braun et al., 1988).
1.5. Classification et Taxonomie de Cicer arietinum L.
La famille des Fabacae comptant plus que 700 genres et 1800 espèces (Polhill
et Raven, 1981). Le genre Cicer L. comptant 44 espèces (Yadav et al., 2007), 9
espèces annuelles et 35 espèces pérennes. Ces espèces sont divisées en 04 sections:
Monocicer, Chamaecicer, polycicer et Acanthocicer (Van Der Maesen, 1987;
Valcilova et al., 2002). Le pois chiche (Cicer arietinum L.) appartient au genre Cicer
à la classe des Dicotylédones, à la sous-classe des Dialypétales, l’ordre des rosales,
famille de Fabaceae, la sous-famille des Papilionaceae, Régne : Plantae et à la
section Monocicer (Bock, 2009; Yadav et al., 2007; Staginnus et al., 1999; Singh et
Diwakar, 1995; Moreno et Cubero, 1978).
1.6. Caractéristiques agronomiques
1.6.1. Les exigences climatiques: le pois chiche se cultive entre 20°N et 40°N dans
l'hémisphère nord, et à petit échelle entre 10°N et 20°N, ces environnements diffèrent
dans la photopériode, la température et les précipitations (Singh et Diwakar, 1995).
1.6.1.1. La température: Les graines du pois chiche germent à une température
optimum entre 28 à 33°C (Singh et Diwakar, 1995; Covelle et al., 1986), mais elles
peuvent germer entre 10 et 45°C (Singh et Diwakar, 1995). Le pois chiche est une
plante à climat intermédiaire, la température optimale exigée varie entre 18°C et 29°C
le jour et 20°C la nuit (Girrard, 1985; Verret, 1982). Le pois chiche souffre dans les
environnements chauds (35°C-18°C jour/nuit) (Lopez-Bellido et al., 2004). Selon
Nielson, (2001), des températures plus de 32°C limitent le rendement en grains du
pois chiche en accélérant sa maturité. De même, les températures élevées de la
floraison à la maturité des variétés à semi retardées conduit à la réduction de la taille
des graines et du rendement (Lopez- Bellido et al., 2004).
1.6.1.2. La pluviométrie: Peu de besoins en eau, résistant assez bien au stress
hydrique, le pois chiche ne demande qu’une pluviométrie moyenne (Singh et
Diwakar, 1995; Singh et Bushan, 1979). Sa consommation en eau a été estimée entre
110 et 240 mm par an pour produire des rendements en grains allant de 9 à 30 qx / ha

Synthèse bibliographique
21
(Singh et Bushan, 1979). Le pois chiche est cultivé principalement comme culture de
précipitation (en hiver dans les climats subtropicaux et en printemps dans la région
méditerranéenne et les climats tempérés).
1.6.1.3. La lumière: Le pois chiche est une plante de jour long, mais fleuri dans
toutes les photopériodes (Smith-son et al., 1985; Summerfield et al., 1979). La plus
part des légumineuses à grains sont des plantes qui préfèrent le soleil et réagissent à
l’ensoleillement en fournissant un grand rendement (Vincent et Gregory, 1974). Il a
été aussi rapporté que l'intensité de la lumière et la durée d'éclairement sont des
facteurs importants pour la nodulation et la fixation de l'azote (Lie, 1971).
1.6.2. Exigences édaphiques
1.6.2.1. Type du sol: Le pois chiche se cultive dans différents types de sols (Khan et
al., 2009; Babar et al., 2009; Yusuf et al., 2002), mais il semble qu'il préfère les sols
meubles, profonds, plus ou moins argileux avec une bonne capacité de rétention
(Singh et Diwakar, 1995; Moolani et Chandra, 1970) ou des sols limoneux profond
qui lui fournit des sels solubles (Moolani et Chandra, 1970). Ces sols retiennent plus
de 200 mm d'humidité sur une longueur d’environ 1m (Saxena, 1987).
1.6.2.2. Humidité du sol: Les graines du pois chiche germent à un niveau d’humidité
de 15% pendant 5 à 6 jours (Singh et Diwakar, 1995).
1.6.2.3. Nutrition minérale: Le pois chiche exige plusieurs éléments minéraux tels
que le nitrogène (N), le phosphore (P), le soufre (S) et le zinc (Zn). Le pois chiche
montre une sensibilité vis-à-vis de la déficience en Zn (Khan et al., 1998; Ahlawat,
1990; Sakal et al., 1988).
1.6.2.4. pH du sol: Le pH optimum du sol pour que le maximum des nutriments soit
assimilable est entre 5.7 et 9 (Singh et Diwakar, 1995; Mahler et al., 1988; Braun et
al., 1988).

Synthèse bibliographique
22
1.7. Répartition géographique, aire de culture et de production du pois chiche
dans le monde
Le pois chiche est cultivé dans 49 pays (Chakraborti et al., 2006) et dans les
05 continents ce qui le rend la 2ème
légume sec dans le monde (17.1% du total) après
le petits pois (Pisum sativum L.) (Berger et al., 2003). Il se cultive dans les régions
semi aride et tropicales (Staginnus et al., 1999).
Tableau 1. Principaux pays producteurs du pois chiche dans le monde, campagne 2007-2008 (FAO,
2010)
Pays Production (tonnes) Pays Production (tonnes)
Inde 5, 970,000 Myanmar 225,000
Pakistan 842,000 Canada 215,000
Turquie 523,553 Ethiopie 190,000
Australie 313,000 Mexique 165,000
Iran 310,000 Iraq 85,000
Total : 9, 313,043
La superficie de la culture du pois chiche dans le monde est de 11.67 millions
d’hectares et sa production totale avoisine 9.31 millions tonnes avec un rendement
moyen de 800 kg/ ha (FAO, 2007). L’Asie est le continent le plus important dans la
production du pois chiche avec plus de 90% de la surface totale et de la production
mondiale (Babar et al., 2009). L'Inde représente le plus grand pays producteur avec
une production estimée à 6.0 millions de tonnes par an et recouvre plus de la moitié de
la production mondiale (Muehlbauer et Rajesh, 2008).
Tableau 2. Répartition géographique de la culture du pois chiche dans le monde
Continents Régions Pays Références bibliographiques
Asie Sud Est de l'Asie Inde – Pakistan -
Myanmar
(Rekha et Thiruvengadam, 2009; Muehlbauer
et Rajesh, 2008; Yadav et al., 2007; Ram et
Prem, 2005; Flandez - Galvez et al., 2003;
Valcilova et al., 2002; Staginnus et al.,
1999).
Centre et Ouest de
l'Asie
Turquie – Iran –
Iraq - Syrie
Afrique Nord de l'Afrique Maroc - Tunisie (Ben Mbarek et al., 2009; Rekha et
Thiruvengadam, 2009; Muehlbauer et
Rajesh, 2008; Yadav et al., 2007; Valcilova
et al., 2002; Staginnus et al., 1999;
Ladizinsky et Adler, 1976).
Est de l'Afrique Ethiopie - Malawi-
Tanzanie
Amériques Amérique du nord Canada - USA. (Rekha et Thiruvengadam, 2009; Muehlbauer
et Rajesh, 2008; Yadav et al., 2007; Flandez Amérique du sud Mexique

Synthèse bibliographique
23
- Galvez et al., 2003; Valcilova et al., 2002;
Staginnus et al., 1999).
Australie Australie Australie (Rekha et Thiruvengadam, 2009; Muehlbauer
et Rajesh, 2008; Valcilova et al., 2002;
Staginnus et al., 1999)
Europe Sud de l'Europe Espagne (Rekha et Thiruvengadam, 2009; Muehlbauer
et Rajesh, 2008; Yadav et al., 2007;
Valcilova et al., 2002; Staginnus et al., 1999;
Singh, 1985; Ladizinsky et Adler, 1976).
1.8. Intérêts du pois chiche
1.8.1. Intérêt économique: Les légumineuses alimentaires constituent un composant
important du régime alimentaire, spécialement dans les pays sous développés où elles
représentent environ 90% de la consommation globale (Hassan, 2006).
Tableau 3. Rendement et production mondiale du pois chiche en comparaison avec d’autres cultures
(campagne 2004-2005) (FAO, 2005)
Cultures Rendements
(Kg/ha)
Production (Mt)
Maïs 4.707 692.034.184
Blé 2.898 626.466.585
Riz 4.004 614.654.895
Soja 2.292 209.531.558
Arachides 1.447 36.492.147
Haricots 0.709 25.419.286
Petits pois 1.757 20.721.735
Pois chiche 0.818 9.172.530
Lentille 1.007 4.031.837
1.8.2. Intérêts agronomiques: La capacité symbiotique que possède le pois chiche
d’utiliser l’azote atmosphérique pour sa croissance, leur rend comme culture
préférable de l’agriculture durable en réduisant la dépendance au fertilisant azoté
(Babar et al., 2009; Khan et al., 2009; Hassan, 2006; Flandez-Galvez et al., 2003). Il a
été également rapporté que cette culture réduit l'inoculum potentiel des maladies
racinaires d’origine tellurique (Flandez-Galvez et al., 2003).
1.8.3. Intérêts nutritionnels: Le pois chiche a une importance économique
significative. Ces pailles ont une valeur de fourrage en comparaison avec les autres
pailles communément utilisées pour l’alimentation du bétail (Rekha et
Thiruvengadam, 2009; Malhotra et al., 2000). Le pois chiche constitue une source très
importante de protéines végétales qui peuvent corriger le déficit en protéines animales

Synthèse bibliographique
24
(Ben Mbarek et al., 2009; Rekha et Thiruvengadam, 2009; Chérif et al., 2007;
Hassan, 2006; Singh et Singh, 1992).
Tableau 4. Composition chimique des grains du pois chiche en comparaison avec quelques
légumineuses alimentaires et le blé (pour 100 g de MS) (Aykroyd et Doughty, 1982)
Blé Pois Pois chiche Lentille Fève Espèces
370 330 358 346 343 Calories (g)
13 22.2 20.1 24.2 23.4 Protéines (g)
2 1.4 4.5 1.8 2 Matière grasse (g)
68 60.1 61.5 60.8 60.2 Glucides (g)
2.5 2.7 2.5 3.1 7.8 Cellulose (g)
60 70 149 56 90 Calcium(g)
1 4.3 7 6.1 3.6 Fer (mg)
0.13 0.72 0.4 0.5 0.54 Thiamine (mg)
0.04 0.15 0.18 0.21 0.29 Riboflavine (mg)
------ 4 5 3 4 Vit C (mg)
Le pois chiche est une bonne source de carbohydrates et de protéines qui
constituent ensemble environs 80% du poids sec de la graine. L’amidon est le
principal carbohydrate chez le pois chiche, il contient aussi une quantité considérable
en acide gras. Les triglycérides et les phospholipides sont les composants prédominant
des lipides chez le pois chiche (Singh, 1985). Les acides gras majeurs chez le pois
chiche sont les acides: linoléique, oléique et palmitique (Ling et Robinson, 1976).
Tableau 5. Composition chimique du pois chiche (Singh et Jauhar, 2005; Williams et Singh, 1987)
Eléments gr ou mg/100g
Protéines digestibles (gr) 23
Carbohydrates (gr) 64
Amidon (gr) 47
lipide (Ac. Linoleique et oléique) (gr) 5
Fibres bruts (gr) 6
Sucres solubles (gr) 6
Cendre (mg) 3
Phosphore (mg) 343
Calcium (mg) 186
Magnésium (mg) 141
Fer (mg) 7
Zinc (mg) 3

Synthèse bibliographique
25
I. 9. Le stress biotique chez le pois chiche
Le facteur primordial dans la réduction du rendement du pois chiche reste
l’aspect phytosanitaire attribué souvent à des agents pathogènes, dont les plus
importants sont les agents telluriques (Jimenez-Diaz et al., 1989). Parmi les nombreux
organismes nuisibles qui réduisent la croissance et la production du pois chiche,
l’agent de l’anthracnose, de la pourriture racinaire et du flétrissement (Rekha et
Thiruvengadam, 2009; Flandez-Galvez et al., 2003).
Tableau 6. Principales maladies cryptogamiques chez le pois chiche dans le monde
Maladies Agents responsables Références
anthracnose Ascochyta rabiei (Pass) Labr (Merzoug et al., 2009; Chérif et al., 2007; Mazur et
al., 2004; Singh et al., 1998; Singh et al., 1994;
Susanne et Wolfgang, 1990; Singh, 1990; Haware et
al.,1986).
Pourriture
sèche
Rhizoctonia sp. (Merzoug et al., 2009; Mazur et al., 2004; Rouibah,
1989; Trapero-Casas et Jimenez-Diaz, 1985; Singh et
Mehrotra, 1980).
Pourriture noire Fusarium oxysporum f. sp. Pisi (Merzoug et al., 2009; Mazur et al., 2004; Trapero-
Casas et Jimenez-Diaz, 1985).
Flétrissement
vasculaire
Fusarium oxysporum f. sp.
ciceri
(Merzoug et al., 2009; Chérif et al., 2007; Mazur et
al., 2004; Singh et al., 1998; Singh et al., 1994;
Trapero-Casas et Jimenez-Diaz, 1985; Mani et Sethi,
1984).
Pourriture
racinaire noire
Fusarium solani (Mart.) Sacc. (Merzoug et al., 2009; Mazur et al., 2004; Trapero-
Casas et Jimenez-Diaz, 1985; Mani et Sethi, 1984).
Pourriture du
collet
Sclerotium rolfsii Sacc. Chérif et al., (2007).
Fonte de semi Pythium debaryanum Hesse,
Pythium irregulare Buisman,
Pythium ultimum Trow
(Kainer et Hannan, 1983; Trapero Casas et al., 1990).
Pouriture
racinaire
F.acuminatum,F.
arthrosporioides, F. avenaceu ,
F. solani .f. sp. Eumartii
Merzoug et al., (2009).
Complexe du
flétrissement
F. oxysporum f.sp. ciceri, F.
solani, Verticillium albo-atrum,
Rhizoctonia bataticola et R.
solani
(Fahim et al., 1987; Trapero et Jimez-Diaz, 1985;
Grewal, 1982).
1. 10. Situation du pois chiche en Algérie
En Algérie les espèces de légumineuses alimentaires les plus cultivées sont la
lentille (Lens culinaris L.), le pois chiche (Cicer arietinum L), le pois (Pisum sativum
L), la fève (Vicia faba L.) et l'haricot (Phasiolus L.). Les légumineuses alimentaires
ont reçu beaucoup d’attention de la part des services agricoles pour augmenter les

Synthèse bibliographique
26
superficies et améliorer les niveaux de rendements. Cependant les résultats obtenus
n’ont pas été à la hauteur des efforts consentis (Bouzerzour et al., 2003).
Le pois chiche (Cicer arietinum) en Algérie, vient en seconde place après
l'haricot avec une superficie de 14.6% et occupe la troisième place en production
environ 15.6%. Cependant, les productions n'ont pas évalué au contraire ils sont
régressé pour atteindre les niveaux les plus faibles dans le monde (4qx/ha) (Mahrez et
al., 2010; Abdelguerfi et al., 2001).
Compte tenu des problèmes que pose la culture du pois chiche en Algérie,
particulièrement du point de vue comportement variétal vis-à-vis des facteurs
biotiques (champignons, insectes, virus) et abiotiques (sécheresse, gelées, froids). La
collecte et l'évaluation adéquate des ressources génétiques locales devient
indispensable pour pouvoir créer des variétés nouvelles ayant un bon rendement,
adaptées aux variations climatiques et résistantes aux maladies (Abdelgherfi et al.,
2000).
L’Algérie porte peu de variabilité pour le pois chiche. Les variétés ou
populations cultivées méritent, cependant, des études sérieuses pour en déterminer les
caractéristiques qui font qu’elles soient encore appréciées par les agriculteurs
(Bouzerzour et al., 2003). Parmi les caractéristiques utilisables immédiatement, et qui
sont successibles d’exister dans le germoplasme local il y a la capacité de la fixation
symbiotique de l’azote de l’air, la hauteur de la plante, la résistance à l’égrenage, la
résistance aux maladies cryptogamiques et la tolérance aux stress abiotiques (Singh et
al., 1998).
Malheureusement, peut d'intérêt est porté à la variabilité qui caractérise le
pois chiche. Les instituts de recherche ont sélectionné 20 cultivars du pois chiche dont
9 sont en multiplication (Bouzerour et al., 2003). Compte tenu du fait que l’Algérie ne
fait pas partie de l’aire de distribution du genre Cicer, on définit le pois chiche local
comme tous cultivars ou variétés introduits par de nombreuses civilisations, au fil du
temps ces cultivars se sont adaptés à certaines conditions édapho-climatiques.
Contrairement au pois chiche local qui est très hétérogène, à caractères génétiques
inconnus et peu utilisé, celui introduit est généralement homogène, à caractères

Synthèse bibliographique
27
génétiques connus et commercialisés à grande échelle (Abdelgherfi et al., 2000).
Parmi les variétés les plus cultivées en Algérie on trouve les variétés ILC-3279
(Chetoui1) et ILC-482 (Cehtoui 2) introduites par l'ICARDA en 1988.
1.10.1. Intérêt cultural et importance des légumineuses en Algérie
L’espèce Cicer arietinum L. est connue en Algérie depuis des milliaires
(Labdi, 1990), elle occupe une grande place dans les habitudes alimentaires de la
population algérienne (soupes, sauces, plats, …) (Abdelgherfi et al., 2001). Avec
l'augmentation de la demande, les autorités nationales ont recours à l'importation
(Labdi, 1990). En Algérie, les espèces cultivées de légumineuses alimentaires ont
bénéficié de peu d’intérêt dans le domaine des ressources phylogénétiques, les
espèces du pois chiche, lentille, haricot sec et pois sec locaux en Algérie présentent
des intérêts non négligeables (Abdelgherfi et al., 2001).
En Algérie, les légumineuses alimentaires sont utilisées dans la rotation avec
les céréales car elles enrichissent le sol en azote, ils sont aussi cultivées parce qu'elles
constituent une importante source protéique susceptible de remplacer les protéines
animales difficilement accessibles pour une large couche de la population algérienne
(Melakhssou, 2007). La place des légumineuses alimentaires dans le système agraire
n'a pas toujours été importante, leur superficie totale entre 1993 -2002 avoisine 82 301
hectares. Les espèces les plus cultivées par ordre d'importance sont: la fève et les
féveroles, le pois chiche et le pois sec. Les rendements moyens enregistrés pour ces
trois espèces sont très faibles, de l'ordre de 3 à 5 qx /ha entre 1993-2002.
Tableau 7. Légumineuses alimentaires cultivées en Algérie, superficie, production et rendement
(Moyenne 1993-2002)
Cultures Superficies Production Rendement
(Qx/Ha) Ha % Qx %
Fève/Féverole 40299 48.96 207042 50.27 5.13
Pois chiche 30487 37.04 161799 39.28 5.30
Pois sec 8627 10.48 29793 7.23 3.45
Lentilles 1271 1.54 5021 1.22 3.95
Haricot sec 1240 1.50 6480 1.57 5.22
Gesse 377 0.46 1732 0.42 4.59
Total 82301 100 411867 100 5.00

Synthèse bibliographique
28
1.10.2. Aire de culture et production du pois chiche en Algérie
On connait quatre zones principales de culture de pois chiche en Algérie
(Benzohra, 2009):
Pleines littorales et sublittorales (pluviométrie, plus de 600 mm/an).
Plaines d’altitude 700 à 900 m (pluviométrie plus de 600 mm/an).
Hautes plaines telliennes (pluviométrie entre 400 et 600 mm/an).
Plaines basses telliennes (pluviométrie entre 400 et 500 mm/an).
Tableau 8. Evolution des superficies, productions, rendement et semences du pois chiche en Algérie
durant la campagne 1998-2008 (FAO, 2010).
Années Superficie
(Ha)
Production
(tonnes)
Rendement
(Hg/Ha)
Semence
(tonnes)
1998 29550 18143 6139 1386
1999 27720 13070 4715 974
2000 19480 6661 3419 964
2001 19290 12312 6382 966
2002 19330 14971 7744 1142
2003 22850
19102 8359 1153
2004 23079 16367 7091 1167
2005 23348 13727 5879 1062
2006 21252 12706 5978 1034
2007 20681 14294 6911 1000
2008 20000 15000 7500 1000
1.10.3. Problèmes phytosanitaires du pois chiche en Algérie
Le pois chiche peut être attaqué par de nombreux parasites. Cependant, tous ne
sévissent pas en Algérie et tous n’ont pas le même impact sur la culture. Nous allons
donner un rapide aperçu de ceux que l’on rencontre fréquemment dans les cultures en
Algérie. Parmi les principales maladies du pois chiche, on rencontre l’anthracnose, la
pourriture racinaire et le flétrissement (Ben Freha et al., 2010; Merzoug et al., 2009;
Zaim, 2007; Bekkar, 2007; Pande et al., 2007; Labdi, 1990).
1.10.3.1. L’anthracnose
Cette maladie est causée par Ascochyta rabiei, ce champignon est transmis par
les semences (Haware et al., 1986), mais peut se maintenir pendant deux ans dans les

Synthèse bibliographique
29
résidus de culture si les conditions climatiques lui sont favorables (température allant
de 9 à 29°C et fortes hygrométries) (Pande et al., 2005).
La maladie peut en quelques jours détruire entièrement une culture réalisée
avec une variété sensible (Chen et al., 2004; Haware et al., 1981). La pratique du
semis précoce, selon plusieurs auteurs impose l’utilisation d’une variété tolérante à
l’anthracnose, ainsi que le traitement de semences avec des produits adéquats (Pande
et al., 2005; Muehlbauer et Kaiser, 1994 ; Reddy et al., 1980). Selon Nene et Reddy,
(1987), les symptômes caractéristiques sont la présence de tâches brunâtres à
rougeâtres avec un halo clair au centre, présentant des points noirs de taille variables
sur feuilles, même les tiges et les gousses peuvent être atteintes en présentant les
mêmes symptômes (Markell et al., 2008; Haware et al., 1986).
1.10.3.2. Pourritures racinaires
On distingue deux types de pourritures: sèche et noire.
Pourriture sèche : elle fut citée la première fois en Inde, dont l’agent causal est le
genre Rhizoctonia spp. Selon Nene et al., (1981), d’autres espèces sont responsables
des pourritures. Cependant, ces dernières années, ces espèces sont beaucoup moins
fréquentes par rapport au genre Rhizoctonia. D’après Rouibah, (1989), cette maladie
débute par un dessèchement brusque de la plante, qui évolue du bas vers le haut, les
feuilles prennent une couleur jaune palle, alors que les racines se dessèchent et
deviennent facilement cassables.
Pourriture noire : Selon, Nene et Reddy, (1987) et Trapero et Jimez-Diaz, (1985), ce
genre de pourriture est causé par le Fusarium oxysporum f. sp. pisi. Cette maladie se
caractérise par des symptômes au niveau des organes aériens, similaires à ceux du
flétrissement, tandis qu’au niveau des organes souterrains on distingue une pourriture
noire sans décoloration du système vasculaire (Alvarez et Briner, 1987). D’après
Trapero et Jimez-Diaz, (1985), l’importance du jaunissement foliaire est directement
liée avec la sévérité des attaques de la partie souterraine. Selon toujours ces mêmes
auteurs, ce pathogène cause aussi la détérioration des semences et la mort des
plantules en post émergence.

Synthèse bibliographique
30
1.10.3.3. Complexe du flétrissement
Le flétrissement est un complexe de plusieurs agents pathogènes du pois
chiche, plusieurs auteurs signalent notamment le F. oxysporum f.sp. ciceri, F. solani,
Verticillium albo-atrum, Rhizoctonia bataticola et R. solani (Labdi, 1990; Fahim et
al., 1987; Trapero et Jimez-Diaz, 1985; Grewal, 1982).
Plusieurs auteurs tels que Alvarez et Moreno, (1984); Allen, (1983); Nene et
al., (1981) et Haware, (1990) ont signalés que le terme « complexe du flétrissement »
définit deux maladies bien distinctes, le flétrissement et les pourritures racinaires.
Selon Jimenez-Diaz et al., (1989), les pertes annuelles dues à ces deux types de
maladies sont estimées à environ 10 millions de dollars. En Tunisie, le flétrissement a
entrainé la destruction de la moitié de la campagne 1982 – 1983 (Rouibah, 1989).
2. L’agent pathogène (Fusarium oxysporum f. sp. ciceri)
2.1. Généralités sur le genre Fusarium
Le genre Fusarium comptant principalement des espèces phytopathogènes
(Agrios, 2005; Pieckova et Jesenska, 1999), nécrotrophiques, d'origine tellurique
causants des maladies sérieuses chez les plantes dans le monde (Agrios, 2005).
Cependant, ce genre regroupe des espèces appartenant à des agents causants des
mycoses chez l'être humain (Pieckova et Jesenska, 1999). La production de
mycotoxine est un phénomène très commun chez ce genre (trichothcenes, zearalenon,
fumonisins) (Pieckova et Jesenska, 1999). Les espèces de F. oxysporum causent
principalement le flétrissement vasculaire de plusieurs espèces de plantes en causant
des pertes de rendement importantes (Agrios, 2005; Schouten et al., 2004). Cependant
plusieurs espèces du genre Fusarium, spécialement F. solani cause la pourriture des
racines et des tiges et même des semences, cette pourriture est accompagnée par une
production de mycotoxines (Agrios, 2005).

Synthèse bibliographique
31
2.2. Généralités sur le Fusarium oxysporum
Fusarium oxysporum Schlecht est un champignon d'origine tellurique
cosmopolite, présente une très grande diversité génétique et écologique (Jimenez -
Gasco et al., 2004; Di-Pietro et al., 2003; Di-Pietro, 1998; Beckman, 1987;
Armstrong et Armstrong, 1981; Booth, 1971). Cette espèce inclut plus d’une centaine
de formes spéciales et races qui sont chacune spécifique d’une plante hôte, souvent
des espèces végétales d’intérêt horticole, maraîcher ou agronomique. F. oxysporum
inclut également des populations non pathogènes pour lesquelles aucune plante hôte
n’a été identifiée à ce jour (Barik et al., 2010; Arroyo et al., 2003).
Parmi cette population, on rencontre des parasites opportunistes et des agents
antagonistes (Summerbell et Schroers, 2002). Cette espèce est présente dans la
plupart des sols cultivés dans le monde (Odds et al., 1998; Boutati et Anaissie, 1997).
Les espèces phytopathogènes de F. oxysporum causent des maladies sérieuses sur les
cultures, en causant spécialement le flétrissement vasculaire (Di-Pietro, 1998; Tello et
Lacasa, 1990).
Cette maladie attaque plusieurs plantes pérennes et annuelles, aussi bien dans
les zones tempérées que dans les zones tropicales causant leur jaunissement et leur
flétrissement (Di-Pietro et al., 2003; Di-Pietro, 1998; Namiki et al., 1994; Beckman,
1987).
Le champignon produit trois types de spores: les chlamydospores, les
macroconidies et les microconidies (Booth, 1971). Le haut niveau de spécificité des
souches pathogéniques dans F. oxysporum a conduit au développement des formes
spéciales conçu pour permettre une bonne différentiation de ces souches similaires
morphologiquement (Cunnington et al., 2009). Les souches qui ont une gamme
d'hôtes identiques sont assignées dans un groupe intraspécifique, nommé formae
speciales (Armstrong et Armstrong, 1981; Snyder et Hansen, 1940), créer par Snyder
et Hansen (Agrios, 2005). Les formae speciales sont distinguables par leur capacité à
causer les symptômes du flétrissement sur une gamme d'hôte de plante limité
taxonomiquement (Armstrong et Armstrong, 1981; Snyder et Hansen, 1940).

Synthèse bibliographique
32
Actuellement, cette espèce se compose de plus de 120 formes spéciales correspondant
aux hôtes qu'elles infectent (Agrios, 2005), chacune de ces formes peut être divisé en
races physiologiques et chaque race physiologique montre un modèle caractéristique
de virulence sur des variétés différentielle dans la même espèce de plante hôtes
(Agrios, 2005; Armstrong et Armstrong, 1981).
2.3. Fusarium oxysporum f. sp. ciceri (FOC)
2.3.1. Historique
Fusarium oxysporum a été décrit pour la première fois par Matuo et Ishigami
en (1958) à partir d'une plante soufrant du flétrissement vasculaire S. melongena
(Solanaceae). Tel que cité par plusieurs auteurs, l’agent causal responsable du
flétrissement du pois chiche est Fusarium oxysporum (Schliecht) f. sp. ciceri (Hans.)
Snyd et Hansen (Cabrera et al., 1985; Halila et al., 1984; Sharma et Gupta, 1983;
Nene et al., 1978), signalé depuis 1910 (Erwin, 1958). Le diagnostique de cette
maladie n’a été complété qu’en 1940 par Padwick (Padwick, 1940). La maladie a été
signalée en premier lieu dans seulement 14 pays.
Les premières recherches sur cette maladie ont débuté en Inde et Myanmar
dans les années 1920 puis en Mexique, des confusions dans l'identification du
flétrissement du pois chiche ont été très répandue, jusqu'à que Nene et al., (1981) ont
clarifié l'identification de FOC (Singh, 1987). Actuellement, cette maladie a été
rapporté dans au moins 33 pays (Singh et al., 2002).
2.3.2. Présentation de FOC
Le filament mycélien de Fusarium oxysporum Schlecht f. sp. ciceri (Hanz)
Snyd. et Hansen est hyalin, septé et uninuclé (Dickinson, 1932), il provoque des
dégâts systémiques par pénétration dans les racines de la plante du pois chiche et
obstruction des vaisseaux. Il bloque ou réduit le passage de l'eau et des éléments
nutritifs vers les feuilles et cause le flétrissement vasculaire (Sharma et Muehlbauer,
2007), ainsi le F. oxysporum ciceri est inféodé au genre Cicer (Kraiser, 1994). Le
FOC se transmis par semence et par le sol (Kraft et al., 1994; Pande et al., 2007),

Synthèse bibliographique
33
survie à travers les chlamydospores dans les grains et les débris des plantes mortes
(Haware et al., 1978) et par conséquent il est difficile de le contrôler par l'utilisation
des produits chimiques et la rotation des cultures (Shah et al., 2009). Le pathogène
peut rester en survie dans le sol en absence de son hôte pendant au moins 6 ans
(Stevenson et al., 1995; Haware et al., 1996). L’instabilité morphologique de
Fusarium oxysporum f. sp. ciceri sur milieu de culture est un phénomène très
commun (Nelson et al., 1983; Haware et al., 1978).
2.3.3. Description et clés d'identification de FOC
Le genre Fusarium oxysporum peut être définit par les critères
morphologiques, incluant la forme de la microconidie et macroconidie, la structure du
conidiophore (fausse tète sur des phialides courts formés sur l'hyphe) et la formation
de chlamydospores. Sur le milieu CLA (Carnation Leaf-piece Agar), les
macroconidies de Fusarium oxysporum ciceri sont formées en sporodochie orange
pale qui naissent à partir de monophialides sur des conidiophores branchés et parfois
sur des monophialides sur l'hyphe. La longueur des macroconidies est courte à
moyenne, courbée à légèrement droite, à paroi épaisse et présentant souvent 03
septations.
Les microconidies sont formés abondamment en fausse tête sur des
monophialides courtes, ils peuvent être ovales, elliptiques ou réniformes et sont
souvent sans septation (Leslie et Summerell, 2006).
Chez la plupart des isolats, les chlamydospores sont formées abondamment et
rapidement (2 à 4 semaines), mais elle peut être lente (4 à 6 semaines) et parfois
n'existe pas chez certains isolats. Les chlamydospores sont souvent unique ou en
paires, mais peuvent être en groupe ou en petite chaine. Ils peuvent être terminaux ou
intercalaire. Sur milieu PDA, la morphologie des colonies varie largement. Fusarium
oxysporum produit un pigment sur agar-agar, qui varie de claire au violet foncé, mais
quelques isolats ne le produisent pas. Le mycélium peut être cotonneux, épars ou
abondant, et varie de la couleur blanchâtre au violet. Les macroconidies sont produites

Synthèse bibliographique
34
en abondance. Des sclérotes marron claire à bleu foncé ou violette peuvent être
produite en abondance chez quelques isolas (Leslie et Summerell, 2006).
Fusarium oxysporum peut être distingué de F. solani, par la formation des
microconidies en fausses tète sur des monophialides longues, et se distingue de F.
subglutinans par des microconidies sur des polyphialides et ne forme pas de
chlamydospores (Leslie et Summerell, 2006). Cette identification morphologique pose
un problème à cause de la diversité des isolats non pathogéniques et saprophytiques
dans le sol, et aussi ne diffère pas les races de FOC, c'est pourquoi des méthodes
moléculaires ont été mise au point pour surmonter ce problème (Jimenez-Gasco et
Jimenez-Diaz, 2003).
2.3.4. Classification, biologie et cycle de reproduction du champignon
Fusarium oxysporum (Schlecht.) emend. Snyder et Hansen, se distingue des
autres espèces de Fusarium par la production abondante de microconidies,
rassemblées en fausse tète à partir de monophialides courtes. Seule la reproduction
asexuée est connue chez cette espèce (Jimenez-Gasco et al., 2004), ce qui la place
dans le groupe des Deutéromycètes. En fait, F. oxysporum est un Deutéromycètes
telluriques appartenant à la sous classe des Hyphomycètes et à la famille des
Tuberculariacées, il fait partie de la section Elegans (Nelson et al., 1983; Messiaen et
Cassini, 1968; Booth, 1971).
2.3.5. Mécanismes d'infection et de colonisation
2.3.5.1. Mode d’infection
Le champignon peut survivre dans le sol sous forme de mycélium ou spore en
absence de son hôte (Agrios, 2005). La chlamydospore est la forme dormante dans les
tissus en décomposition. Le cycle de vie de Fusarium oxysporum (Figure 4.)
commence dans le sol en présence de l'hôte. Le stimulus de la germination serait les
racines de la plante hôte, en le contactant avec des débris non colonisés de la plante
fraîche (Delgado-Jarana, 2005). Le tube germinatif des spores ou du mycélium
pénètre dans les racines de plantes saines cultivées dans un sol contaminé. L'entrée

Synthèse bibliographique
35
est soit directe, à travers les parois ou opportuniste (Cunnington et al., 2009; Agrios,
2005; Aboul-Soud et al., 2004). Le mycélium prend un chemin intercellulaire à
travers le cortex, et entre dans les vaisseaux du xylème, dans lequel il se multiplie en
causant les symptômes du flétrissement (Agrios, 2005; Aboul-Soud et al., 2004).
2.3.5.2. Mode de colonisation
Après infection des racines de l'hôte, le champignon traverse le cortex et entre
dans les tissues du xylème (Cunnington et al., 2009). Généralement, le mycélium
migre à travers la partie inférieure de la plante vers la tige et la couronne (Aboul-Soud
et al.., 2004), ensuite le pathogène se branche aux vaisseaux du xylème où le
mycélium produit des microconidies (Cunnington et al., 2009; Halila et al., 2009;
Aboul-Soud et al., 2004), les microconidies se détachent et se transportent dans le
système vasculaire (sève) via le flux de transpiration (Aboul-Soud et al., 2004). A ce
stade les microconidies germent et le mycélium pénètre dans la paroi du vaisseau
adjacent et il devient systémique dans les tissues de l'hôte (Cunnington et al., 2009).
L'économie de l'eau des plants infectés est éventuellement compromis
sévèrement par le blocage des vaisseaux conductrices par la germination des
microconidies, le résultat est la clôture des stomates, flétrissement et mort des feuilles,
suivie par la mort de la plante entière (Halila et al., 2009; Cunnington et al., 2009;
Aboul-Soud et al., 2004).
2.3.6. Inoculum primaire, mode de survie et de transmission de FOC
Le FOC est un pathogène d'origine tellurique, cependant quelques rapport
indiquent que ce pathogène peut être transmit par les graines (Richerdson, 1990;
Haware et al., 1986). Cela suppose que l'augmentation de la densité de l'inoculum de
FOC dans le sol est aussi contribuée par le mélange des graines infectées avec les
graines saines utilisées pendant le semis (Pande et al., 2007). La dissémination du
champignon peut se produire selon différentes formes: les résidus de plants infectés
(racines, tiges, feuilles), le sol et les graines. Les spores peuvent être disséminées par
le vent et la pluie (Cunnington et al., 2009; Booth, 1971).

Synthèse bibliographique
36
Le FOC est fréquemment considéré comme agent de maladies monocycliques
gouvernées fortement par la densité et la distribution de l’inoculum primaire. C'est
pour cela, le but principal d'une stratégie de contrôle est de réduire la quantité de cet
inoculum (Rekha et al., 2000).
Les champignons du sol ont, quant à eux, des rayons de disséminations très
faibles. Ils évoluent par le développement de leur mycélium, attiré par les sécrétions
racinaires des plantes. Ils bénéficient néanmoins d’autres atouts: des formes de
conservation performantes qui peuvent rester viables plusieurs années dans le sol et
passer ainsi le cap de rotations longues. Ils peuvent en outre être disséminés d’une
parcelle à l’autre par le biais du matériel de travail du sol (Cunnington et al., 2009;
Gwata et al., 2006).
L’homme est en effet lui aussi un vecteur, notamment par le transport
de semences contaminées, ou du fait d’échanges commerciaux de matières premières
végétales. Par ces différents vecteurs, les maladies peuvent être transportées sur des
grandes distances, d’une parcelle à l’autre mais aussi d’un pays à l’autre (Cunnington
et al., 2009; Gwata et al., 2006).

Synthèse bibliographique
37
Figure 4. Cycle infectieux de Fusarium oxysporum, d'aprés Agrios, (2005)
Anneau des vaisseaux
décolorés d'un xylème
secondaire
Mycélium et
conidies
dans des
vaisseaux
Des vaisseaux de
xylème chez une tige
ou un pétiole sains
Des vaisseaux détruits chez
une tige ou pétiole infectés
Pénétration à travers les
blessures- le mycélium
entre dans les vaisseaux
Les conidies
forment des
sporodochies sur
les feuilles mortes
Mycélium ou tubes
germinatifs
attaquent les racines
Spore en germination
Microconidies
Macroconidies Chlamydospore
s
Mycélium
Spores formées par le mycélium dans le sol Plant entièrement flétris et mort
Les branches basses
commencent à flétrir Tous les stades sont présents dans les tissues et les sols infectés
Pénétration à travers les
cassures formées par
racines latérales
Mycélium dans les
vaisseaux
Le mycélium
bouche les
vaisseaux

Synthèse bibliographique
30
2.3.7. Gamme d'hôte
L'hôte principale de FOC est le genre Cicer (Jimenez-Gasco et al., 2004; Nene
et al., 1996; Haware et al., 1986), mais quelques auteurs ont signalé que ce
champignon peut attaquer d’autres espèces de plantes, telles que: Cajanus cajan (Pois
de pigeon), Lens culinaris ssp. (Lentille) et Pisum sativum (petits pois) (Haware et
Nene, 1982).
2.3.8. Races physiologiques et pathotypes de FOC
Actuellement , 08 races du pathogène (race 0, 1A,1B/C, 2, 3, 4, 5 et 6) ont été
identifié par la réaction sur 10 lignées du pois chiche (Cunnington et al., 2009; Sharma
et Muehalbauer, 2007; Ansari et al., 2001; Garcia- Pedrajas et al., 1999; Jimenez-Diaz
et al., 1993; Jimenez- Diaz, 1989; Cabrera De La Colina et al., 1985; Haware et Nene,
1982). Haware et Nene, (1982) sont les premiers qui ont identifiés les races 1, 2, 3 et
4 de F. oxysporum f. sp. ciceri en Inde. 03 races du pathogène ont été ensuite
identifiées dans le sud de l’Espagne: 0, 5, et 6 (Jiménez-Díaz et al., 1989). La race 1 a
été subdivisé en deux races, nommées race 1A en Inde et la race 1B/C en Espagne,
basée sur la variation dans la réaction aux lignées des plantes hôtes différentielles
(Jimenez-Diaz et al., 1993; Trapero-Casas et Jimenez-Diaz, 1985). Rahman et al.,
(1998) ont suggéré l'occurrence de deux races additionnelle en Inde et cette suggestion
a été appuyée par une autre étude faite par le même auteur.
Les 8 races ont une distribution géographique distincte (Honnaredy et Dubey,
2006; Jimenez-Gasco et al., 2004). La maladie est caractérisée par deux types de
symptômes, nommée: flétrissement vasculaire et jaunissement (Dubey et al., 2007;
Kelly et al., 1994; Jimenez-Diaz et al., 1989). Bien que la caractérisation des races est
importante, la distinction initiale la plus crucial est entre les pathotypes associés
flétrissement - jaunissement (Garcia-Pedrajas et al., 1999) en se basant sur les
symptômes de la maladie qu'il induit dans les tests pathologiques (Trapero-Casas et
Jimenez-Diaz, 1985). Les deux symptômes sont le résultat des infections vasculaires et
les travaux ont montré que le pathotype jaunissement (Yellowing) de FOC est moins
agressive que le pathotype flétrissement (Wilting) (Jimenez-Gasco et al., 2004).

Synthèse bibliographique
31
2.3.9. Distribution de la maladie et des races pathologiques dans le monde
La maladie est largement propagée dans les régions de culture du pois chiche
dans le monde, elle a été rapporté dans au moins 33 pays (Singh et al., 2002). Nene et
Reddy, (1987) ont rapporté que la maladie couvre le nord de l'Amérique, l’Europe, le
Moyen orient et le Sud Est de l'Asie. La distribution globale est corrélée avec la
présence des races de FOC. La fusariose a été rapporté dans la plupart des régions de
culture du pois chiche: le subcontinent Indien, L'Iran, le Pérou, la Syrie, l'Ethiopie, le
Mexique, l’Espagne, la Tunisie, la Turquie, USA (Halila et Strange, 1996; Jalali et
Chand, 1992; Haware, 1990; Nene et al., 1989; Nene et Reddy, 1987; Westerlund et
al., 1974) et l'Algérie (Mehrez et al., 2010; Ben Freha et al., 2010; Bekkar, 2007).
2.3.10. Spécialisation chez le FOC et résistance variétale
La résistance race spécifique chez le pois chiche est gouvernée par plusieurs
gènes qui contrôlent la reconnaissance race de FOC/variété du pois chiche. Plusieurs
sources de la résistance race spécifique ont été identifiées (Sharma et al., 2005) et
quelques unes ont été exploitées pour l'amélioration des cultivars du pois chiche
résistant au flétrissement (Singh et Jimenèz-Diaz, 1996). La résistance à la fusariose
du pois chiche peut être exprimé avant que le FOC gagne l'entrée dans le xylème des
plantes ou même avant (Farooq et al., 2005).
2.3.11. Origine de FOC et source de variabilité
Le génome de FOC entier est lié, et se transmettre en unité d'une génération à
la génération suivante, et leurs populations naturelles devraient être clonale dans la
nature. La reproduction forcée chez le Fusarium oxysporum interdit toute
recombinaison régulière, et la variation génétique doit résulter principalement de
l'accumulation des mutations (Jimenez-Gasco et al., 2004). Les isolats de la même
forme spéciale qui partagent le même hôte partagent des similarités génétiques plus
que les autres de différente spécialisation. L'interprétation évolutionnaire de ce
concept est que les formes spéciales sont monophylétique et que les isolats qui
partagent le même hôte sont tous dérivés d'un seul génotype pathologique. Les races
peuvent être dérivé de mutations ponctuelles dans ce génotype sélectionné par la

Synthèse bibliographique
32
pression sélective imposée par la culture de variétés à gènes résistants (Arroyo et al.,
2003).
Cette clonalité chez le Fusarium oxysporum a été associée avec la
compatibilité végétative (VCG): capacité des isolats fongiques d'établir des
hétérokayons stables par anastomose entre les hyphes homokaryotiques adjacents (Di
Primo et al., 2001; Klein et Correll, 2001; Gordon et Martyn, 1997).
2.4. La maladie de la fusariose du pois chiche
Le flétrissement vasculaire causé par le FOC (Padwick) Matuo et Sato, est
une des majeurs facteurs limitant la production du pois chiche dans le monde (Sharma
et al., 2009; Pande et al., 2007; Jalali et Chande, 1992). Malgré qu'il existe un nombre
de facteurs biotiques et abiotiques qui participe dans l'abaissement de la production,
cette maladie reste une des épidémies les plus importantes (Farooq et al., 2005).
2.4.1. Dégâts causés par la fusariose du pois chiche
Le flétrissement vasculaire, causé par le Fusarium oxysporum Schlechtend.
Fr. f.sp. ciceri (Padwick) Matuo et Sato, cause des pertes annuelles de rendement
estimées entre 10 à 90% (Navas-Cortés et al., 1998; Jalali et Chand, 1992; Singh et
Reddy, 1991; Jimenez-Diaz et al., 1989; Trapero-Casas et Jimenez-Diaz, 1985) et
peut aller jusqu’à 100% de perte quand les conditions sont favorables (Sharma et
Muehlbauer, 2007; Navas-Cortés et al., 2000; Halila et Strange, 1996; Haware, 1990;
Haware et al., 1990; Haware et Nene, 1980). Les pertes annuelles du pois chiche dû
au flétrissement vasculaire ont été estimées à 10% en Inde (Singh et Dahiya, 1973),
50% au Pakistan (Ikramul et Farhat, 1992) et en Espagne (Trapero-Casas et Jiménez-
Díaz, 1985) et à 40% en Tunisie (Bouslama, 1980). La maladie peut apparaitre durant
tous les stades de la croissance du pois chiche (Navas-Cortés et al., 1998).

Synthèse bibliographique
33
2.4.2. Description des symptômes de la fusariose du pois chiche
Le flétrissement précoce causé par le FOC peut être observé sur les génotypes
sensibles 25 jours après le semis (Shah et al., 2009; Raju et al., 2008; Pande et al.,
2007; Navas-Cortés et al., 1998; Nene et al., 1978) et comme tardif au stade de
remplissage de gousses (Pande et al., 2007; Navas-Cortés et al., 1998). Le
flétrissement précoce cause plus de perte que le flétrissement tardif. Les jeunes
plantules infectées par le flétrissement vasculaire s'écroulent, s'aplaties et s'allongent
sur terre et gardent leur couleur verte sombre (Pande et al., 2007). Cependant, les
plantes adultes montrent des symptômes du flétrissement typique, par conséquent le
plant entier montre un abaissement soudain des feuilles (Shah et al., 2009; Raju et al.,
2008; Dubey et Singh, 2004), une couleur pale par rapport aux plantes saines (Shah et
al., 2009) et une décoloration du xylème de la tige (Raju et al., 2008), ensuite le plant
meurt. Les plantes du pois chiche attaquées par le FOC ne montre pas une pourriture
externe et apparaissent saines, lorsqu'on coupe verticalement la partie inférieur de la
région du collet, cette région montre une coloration marron des tissues internes (Shah
et al., 2009; Dubey et Singh, 2004; Nene et al., 1991). Certains auteurs rapportent la
présence d'une pourriture racinaire (Raju et al., 2008), cependant d’autres signalent
l'absence de cette pourriture (Dubey et Singh, 2004; Nene et al., 1991). Les gousses
des plantes du pois chiche malades apparaissent normales, mais les graines sont
généralement plus petites, froissées (pliées) et décolorées (Pande et al., 2007; Haware
et Nene, 1980).
Figure 5. Distribution typique de plantes du pois chiche infestées par le FOC
(Cunnington, 2009)

Synthèse bibliographique
34
2.4.3. Localisation de la fusariose du pois chiche
Le flétrissement vasculaire du pois chiche est répandu dans les pays tropicaux
et tempérés chauds en culture sous serre où en une culture intensive en plein champs
(Arroyo et al., 2003). Le flétrissement a été rapporté dans plusieurs pays dans le
monde (Nene et al., 1984), mais cette maladie est plus importante entre l'altitude 30N°
et 30S° où la saison de la culture est sèche et chaude (Sharma et al., 2009; Pande et
al., 2007), que les altitudes hautes (30-40N°) (Pande et al., 2007).
2.4.4. Répartition géographique et importance économique du flétrissement
vasculaire du pois chiche en Algérie
En Algérie, le flétrissement du pois chiche n’est connu que par quelques
données fragmentaires. Depuis 1970, des isolements effectués à partir des plants du
pois chiche présentant des symptômes du flétrissement et du jaunissement ont montré
la prédominance de Fusarium oxysporum (Si-Hassen, 1990). Durant la campagne
1989 – 1990, une prospection réalisée avec la participation de plusieurs spécialistes de
différentes structures a porté sur les principales cultures de légumineuses (pois chiche,
lentille et fèves) a montré que les pertes dues au F. oxysporum peuvent atteindre 50 à
100 % sur la variété locale RABAT (Bouznad, 1989). D’autres travaux réalisés au
cours de ces quatre dernières années ont montré l’importance des dégâts occasionnés
Figure 6. Symptômes de la fusariose du pois chiche: A, sur plant entier; B, sur coupe
longitudinale de la tige (Cunnington, 2009)
A B

Synthèse bibliographique
35
par F. oxysporum sur pois chiche (Mehrez et al., 2010; Ben Freha et al. 2010;
Merzoug, 2009; Zaim, 2007; Bekkar, 2007).
2.4.5. Facteurs influençant la maladie du flétrissement
Certains facteurs possèdent une très grande influence sur la croissance de
l’espèce fongique et principalement sur les possibilités de réalisation de l’interaction
hôte - parasite, parmi ces facteurs on distingue:
La température: le processus d'infection par le FOC est influencé par
l'environnement spécialement la température (Landa et al., 2001; Navas-Cortés et al.,
2000; Navas-Cortes et al., 1998; Gupta et al., 1987; Bhatti et Kraft, 1992; Westerlund
et al., 1974). Une température avoisinant 25°C est optimum pour le développement de
la fusariose du pois chiche (Sharma et Muehlbauer, 2007), et cela malgré une faible
densité d'inoculum initial du sol (Navàs-Cortès et al., 2000). Des observations de
l'incidence du flétrissement du pois chiche dans le champ ont montré que le facteur
chaleur augmente la sévérité de la maladie (Ben Freha et al., 2010; Trapero-Casas et
Jimenez-Diaz, 1985).
L’humidité: Des observations de l'incidence du flétrissement du pois chiche
dans le champ ont montré que les sols humides augmentent la sévérité de la maladie
(Trapero-Casas et Jimenez-Diaz, 1985).
Effet des exsudats racinaires: La majorité des interactions plante-pathogène
compte sur la libération des composés solubles ou volatiles à partir des grains ou des
racines qui stimulent la germination des propagules d'origine tellurique (Steinkllner et
al., 2005). La qualité et la quantité des substances libérées par les racines vers la
rhizosphère est conditionnées par les espèces de plantes, les conditions de croissance,
le stade de développement, les facteurs environnementales, les blessures mécaniques
ou dues aux bio-agresseurs et de l'activité microbienne (Curl et Truelove, 1986; Hale
et al., 1978). Ces substances peuvent être soit: des sucres, des acides aminées, des
acides organiques, des composés phénoliques, des flavonoïdes, des enzymes, des
acides gras, des régulateurs de croissance, des nucléotides, des tannins, des stéroïdes,
des terpenoides, des alcaloïdes, des poly acétylènes et des vitamines (Bertin et al.,
2003).

Synthèse bibliographique
36
La densité de l'inoculum: La sévérité de la maladie dépend également de la
densité d'inoculum. Ce facteur est rapporté par certains auteurs qui ont observé que
l'augmentation de la charge de l'inoculum augmente la sévérité et la rapidité de la
maladie, par contre sa diminution supprime l'expression de la maladie (Navas-Cortes
et al., 2000; Sugha et al., 1994; Gupta et al., 1987; Westerlund et al., 1974).
La sensibilité des cultivars: l'incidence de la maladie peut être influencé par le
génotype du pois chiche cultivé, cette incidence augmente lors de l'utilisation de
génotypes sensibles (Navas-Cortes et al., 2000).
3. Agent antagoniste (Actinomycètes)
3.1. Les actinomycètes
3.1.1. Présentation et caractéristiques des actinomycètes
Les actinomycètes sont des bactéries dont la croissance donne lieu à des
colonies circulaires (Eunice et Prosser, 1983) constituées d'hyphes qui irradient par
croissance centrifuge tout autour du germe qui leur a donné naissance (Eunice et
Prosser, 1983; Lechevalier et Lechevalier, 1981; Gottlieb, 1973), cela explique leur
dénomination actinomycètes, du grec aktis (à rayon), mykes (champignon):
champignons à rayons (Pandey et al., 2003; Gottlieb, 1973). Les actinomycètes sont
des bactéries filamenteuses, à Gram positive (Arifuzzaman et al., 2010; Ningthoujam
et al., 2009; Pandey et al., 2003; Lo et al., 2002) sporulant avec un ADN riche en GC
(57% -75%) (Pandey et al., 2003; Lo et al., 2002). 70% de ces microorganismes sont
producteurs de métabolites secondaires, parmi eux 95% appartiennent au genre
Streptomyces sp. (Bastide et al., 1986).
Figure 7. Observation microscopique des actinomycètes, A: au microscope optique (coloration de
Gram x100) (Todar, 2009); B et C: au microscope électronique montrant des chaines de spores
spirales et des surfaces de spores lisses (Atta et al., 2009; Yamamoto et al.,1998).
B C A

Synthèse bibliographique
37
Sur milieux solides, les actinomycètes forment en une semaine des colonies
souvent pigmentées (gris, vert, rouge, …) provenant de l'accumulation d'hyphes
ramifiés à contour lisse ou échancré à aspect compact, poudreux ou en chou-fleur
(Kitouni, 2007), quelques actinomycètes synthétisent des pigments foncées, la
mélanine ou mélanoide, qui est considéré comme un critère utile pour les études
taxonomiques (Venkatesan et al., 2008).
3.1.2. Ecologie des actinomycètes
Les actinomycètes sont le groupe de microorganismes le plus largement
répandu et distribué dans la nature (Oskay et al., 2004; Barakate et al., 2002).
Ils préfèrent un pH neutre ou peu alcalin, ils sont généralement mésophiles,
d'autres sont thermophiles tolérants à des températures allant jusqu'à 60°C (Omura,
1992). On retrouve les actinomycètes dans divers milieux écologiques, y compris les
sols désertiques et forestiers, des sols cultivés ou non cultivés, fertiles ou non fertiles
(Goodfellow et Simpson, 1987) et dans l'eau (Verma et al., 2009; Zitouni et al., 2005;
Sette et al., 2005; Lo et al., 2002). Les actinomycètes peuvent vivre à l'état libre ou
en association (Verma et al., 2009), les associations établies sont: la symbiose avec
les plantes non légumineuses comme le genre Frankia (fixation de l'azote
atmosphérique) (Benson et Silvester, 1993) ou l' endophytisme dans les tissus de
plantes où il stimulent leur croissance (Bradbury, 1986; Doumbou et al., 1998) ou
même dans les invertébrés marins (Salomon et al., 2004; Doumbou et al., 1998).
3.1.3. Importance des actinomycètes
La fonction majeure des actinomycètes est la décomposition des résidus
végétaux et animaux, à cause de leur diversité métabolique (Kang et al., 2010) et leurs
mécanismes spécifiques en rentrant dans les cycles de bio- remédiation (Sette et al.,
2005; Williams et Vickers, 1988). Les genres de la classe des actinomycètes et
spécialement Streptomyces spp. ont été reconnu depuis longtemps comme source
importante de métabolites bioactives utiles à grande valeur commerciale (Demain et
Sanchez, 2009; Zitouni et al., 2005; Berdy, 2005; Oskay et al., 2004; Sabaou et al.,
1998), comme les antibiotiques (Demain et Sanchez, 2009; Zitouni et al., 2005;

Synthèse bibliographique
38
Berdy, 2005; Oskay et al., 2004; Sabaou et al., 1998) et autres métabolites bioactives
comme les herbicides, les pesticides, les anti-parasitiques et les enzymes (Zitouni et
al., 2005; Oskay et al., 2004). Depuis la découverte de l'actinomycine, les
actinomycètes ont fournit plusieurs composés bioactives et sont continué d'être
sélectionner pour des nouvelles substances bioactives (Barakate et al., 2002).
Cependant leur utilisation intensive a eu pour conséquence l’adaptation des bactéries à
ces substances (Boughachiche et al., 2005).
Dans le domaine de la thérapie humaine et vétérinaire, de nombreux composés
intéressants sont produits industriellement, comme les antibiotiques, les antifongiques
systémiques ou topiques, les vitamines, les enzymes, l’antihistaminiques, les
vasodilatateurs et les immuno- stimulants (Rattanaporn et al., 2004; Oskay et al.,
2004; Lo et al., 2002; Nolan et Cross, 1988; Bastide et al., 1986; Tiraby et Etienne,
1983). La majorité des actinomycètes représentent un groupe fort et utile dans le
domaine agronomique (Sabaou et al., 1998), ils sont utilisées pour la prévention des
maladies de plantes (PGPR) et dans le domaine de la lutte biologique (Oskay et al.,
2004; Bastide et al., 1986). Une autre importance des actinomycètes, est leur
utilisation dans la fertilisation des sols à travers leur pouvoir fixateur d'azote (Sabaou
et al., 1998). Les enzymes des actinomycètes sont aussi utilisées dans l’industrie
alimentaires (conservateurs) et en alimentation animale (Bastide et al., 1986).
3.1.4. Taxonomie des actinomycètes
Le groupe des actinomycètes comprend actuellement plus de 40 genres et
quelques centaines d’espèces, les propriétés chimiques, physiologiques et
immunologiques des actinomycètes les rangent parmi les procaryotes (Williams et al.,
1973), leur paroi cellulaire renferme une glycoprotéine contenant de la lysine ou de
l'acide diaminopémilique (Mariat et Sebald, 1990; Lechevalier et Lechevalier, 1985).
Le diamètre des hyphes est plus petit que celui des champignons (Gottlieb, 1973). Ces
caractères s'ajoutent à d'autres (existence d'espèces anaérobies strictes, sensibilité à
des actino-phages, …) confirment leur classification parmi les bactéries (Mariat et
Sebald, 1990; Demain et Solomon, 1985). Les actinomycètes se classent dans le règne

Synthèse bibliographique
39
des procaryotes, à la division des Firmicutes, à la classe des Thalobacteria et l'ordre
des Actinomycetales (Bergey's, 1989).
3.1.5. Utilisation des actinomycètes en lutte biologique
Après Bacillus thuringiensis qui est le micro-organisme le plus utilisé comme
bipesticide, on rencontre des espèces appartenant au actinomycètes (Kavita et al.,
2010; El-Tarabily et al., 2006; Fourati- Ben Fguira et al., 2005; Doumbou et al.,
2002). Plusieurs travaux ont montré que ces derniers peuvent être utilisés dans ce
domaine. Ce groupe d’actinomycètes possède les principales propriétés d’un agent de
lutte biologique. Le tableau 9 résume quelques travaux publiés sur l'action antagoniste
des actinomycètes sur les champignons telluriques.
Tableau 9. Quelques publications sur l'utilisation des actinomycètes dans la lutte biologique contre les
pathogènes telluriques
Maladies Plantes Pathogènes Antagonistes Références
Fonte de
semi
Betterave
Concombre
Balsam
Pythium ultimum
Pythium
aphanidermatum
Phytophthora
aphanidermatum
Sclerotium rolfsii
Actinoplanes spp.-
Actinoplanes
Philippinensis -
Microbispara rosea-
Micromonospora chalcea-
Streptomyces sp.
(Khamna et al.,
2009; El-Tarabily,
2006; Khan et al.,
1997)
Pourriture
racinaire
Lupin
Banksia sp.
framboise
Plectosporium tabacinum
Phytophthora cinnamomi
Phytophthora fragariae
var. rubi
Actinoplanes spp.-
Missouriensis carbonacea-
Nocardioides sp.
(El-Tarabily,
2003; El-Tarabily
et al., 1996
;Valois et al.,
1996).
Cavity spot
Carottes
Pythium coloratum
Actinomadura rubra-
Actinoplanes
philippinensis- M.
carbonacea-
Streptosporangium
albidum -
Streptoverticillium
netropsis
(El-Tarabily et
al., 1997).
Piétin-
échandeage
Blé
Gaeumannomyces
graminis var. tritici
Streptomyces sp.-
Microbispara sp.
Micromonospora sp.-
Nocardioides sp.
(Coombs et al.,
2002 ; Coombs et
Franco., 2002).
Pourriture
grise
Vigne
Botrytis cinerea
Streptomyces analatus -
Micromonospora
flavogrisea- Streptomyces
sp.
(Loqman et al.,
2009).

Synthèse bibliographique
40
Flétrissement
vasculaire
bananier
Pois chiche
Palmier
dattier
Fusarium oxysporum
Streptomyces griseus-
Streptomyces sp.-
Streptomyces roseoflavus
(Getha et al.,
2005; Fourati -
Ben Fguira et al.,
2005; Dhedhi et
al., 1990).
Sclérotiniose laitue Sclerotinia minor M. carbonacea (El -Tarabily et
al., 2000).
lagale
argentée
Pomme de
terre
Helminthosporium solani Nocardia globerula (Elson et al.,
1997).
Alternariose pommier Alternaria brassicicola Streptomyces sp. (Khamna et al.,
2009).
Pourriture
sèche
Pomme de
terre
Colletotrichum
gloeosporioides
Streptomyces sp. (Khamna et al.,
2009).
Pourriture
verte
Oranger Penicillium digitatum Streptomyces sp. (Khamna et al.,
2009).
3.1.6. Mode d'action des Actinomycètes
les mécanismes responsables de la suppression de la maladie par les
actinomycètes sont: la compétition saprophytique des nutriments sur ou au contacte
de la surface racinaire (Alabouvette et Couteaudier, 1992; Lemanceau, 1989;
Alabouvette, 1986), compétition parasitique de sites d'infection sur les racines
(Schneider, 1984) et induction de la résistance systémique chez l'hôte (Liu et al.,
1995; Matta, 1989; Ogawa et Komada, 1985). Mandeel et Baker, (1991) ont démontré
que ces 03 mécanismes peuvent être actifs dans un même isolat à la fois. Plusieurs
auteurs ont rapporté que l'effet protecteur exercer par les Streptomyces sp. augmente
par la capacité de ces isolats a secréter des chitinases (Taechowisan et al., 2004), IAA
(Martinez-Noel et al., 2001), et des sidérphores (Hamdi et al., 2008; Matthijs et al.,
2007; Errakhi et al., 2007; Cao et al., 2002).
3.1.7. Possibilité d’utilisation des actinomycètes contre le FOC
Quelques travaux ont montré l’action antagoniste des espèces d’actinomycètes
vis-à-vis le FOC, aussi bien au champ qu’au laboratoire. Zaim et al., (2010) ont
obtenu une réduction de 83% du flétrissement vasculaire in vitro en utilisant
Streptomyces sp. Dhedhi et al., (1990) ont rapporté la réduction totale du flétrissement
vasculaire chez le pois chiche au champ en utilisant Streptomyces sp.

Synthèse bibliographique
41
3.2. Le genre Streptomyces
Vue l’importance du genre Streptomyces dans le domaine de la lutte
biologique, de plus, les souches bactériennes antagonistes de FOC isolées au
laboratoire responsables à la réduction du flétrissement vasculaire du pois chiche
appartient à ce genre d'actinomycètes, nous nous sommes efforcé d’apporter quelques
généralités sur ce groupe très particulier.
Plusieurs recherches ont montré que les Streptomyces sont des antagonistes
des champignons tellurique et ont la capacité de coloniser les racines de plantes hôtes
(El-Tarabily et Sivasithamparam, 2006; Cao et al., 2005; Xiao et al., 2002), ce qui
explique leur utilisation en agriculture, étant données que 60% des antimicrobiens
utilisé dans ce domaine sont isolées à partir de Streptomyces spp. (Ilic et al., 2007). La
plus part des composés issus des Streptomyces sp. utilisés en agriculture appartenant à
la famille des polyketides (Savic et Vasiljevic, 2006).
3.2.1. Caractéristiques du genre Streptomyces
Les Streptomyces ont pour niche écologique le sol (Pandey et al., 2003), la
majorité des Streptomyces jouent un rôle important dans la décomposition de
différents biopolymères et dans la minéralisation de la matière organique. Ces
bactéries saprophytes produisent diverses enzymes extracellulaires impliquées dans la
dégradation de nombreuses macromolécules issues des plantes et des animaux
(chitine, pectine, amidon, kératine, élastine, etc…) (Aghighi et al., 2004; Hirsh et
Christensen, 1983). Ce sont des eubactéries de l'ordre des Actinomycetales. Cette sous
division des bactéries à Gram+ (Lo et al., 2002; Sabaou et al., 1998) est caractérisée
notamment par leur haut pourcentage de bases guanine (G) et cytosine (C) dans leur
génome (Lo et al., 2002). Généralement, le pourcentage GC chez ces bactéries est de
72% contre 50% chez E. coli (Singh et al., 2006).
Les Streptomyces sont les actinomycètes les mieux connus. Ils sont très étudiés
pour leur propriété à synthétiser des antibiotiques (streptomycine, kanamycine,
chloramphénicol), des antifongiques (amphotéricine, candicine…), des antitumoraux
(mitomycine, actinomycine…), des antiparasitaires (hygromycine, salinomycine…),

Synthèse bibliographique
42
des antiviraux (Ara -C, tunicamycine…), des insecticides (avermectine,
milbémycine…) et des herbicides (bialphos, phosphinothricine…) (Rattanaporn et al.,
2004; Oskay et al., 2004; Lo et al., 2002; Nolan et Cross, 1988; Bastide et al.,
1986;Tiraby et Etienne, 1983).
3.2.3. Cycle de différentiation des Streptomyces
Le genre Streptomyces représente plus de 90% des bactéries filamenteuses
(Kurylowicz et al., 1975). Les Streptomyces se distinguent des autres bactéries par
leur cycle de différentiation. Très souvent, on considère qu'une colonie bactérienne est
un ensemble de bactéries toutes identique entre elles (puisque qu'elles dérivent par
scissiparité d'un clone isolé). Dans le cas des Streptomyces, on observe des colonies
formées de cellules qui n'ont pas la même morphologie, elles sont pourtant issues du
même clone initiale. Cette particularité est le résultat d'une différentiation
morphologique par étapes successives (Serrano et Trujillo, 2005).
La germination d'une spore, sur milieu solide donne d'abord naissance à un
mycélium basal. Celui-ci s'incruste dans le milieu de croissance par action d'enzymes
hydrolytiques synthétisées par les cellules. Ensuite, selon les souches et les conditions
environnementales, on observe le développement d'un mycélium aérien. Les hyphes
qui le composent peuvent être de forme variable selon les souches (droite, spiralée,
branchée,…) (Keulen et al., 2005; Shirling et Gottlieb, 1966). C'est à ce stade que
certaines colonies synthétisent les pigments dont la couleur et l'intensité varient selon
les souches et le milieu de culture solide. Les hyphes, contenant plusieurs copies de
l'ADN génomique, donnent naissance à une cinquantaine de spores haploïdes après
septation régulière. Ce développement sous forme de mycélium ressemble beaucoup
à celui de différentiation des champignons d'où le suffixe –myces- (Desjardin, 2002).
3.2.4. Classification des Streptomyces
Le genre Streptomyces appartient à l'ordre des Actinomycetales, la famille des
Streptomycetaceae, les espèces les plus connues sont : S. coelicor, S. lividans, S.
griseus, S. ambofaciens, S. albus, S. thermocarboxydus (Desjardin, 2002).

Synthèse bibliographique
43
3.2.5. Les avantages de l'utilisation des Streptomyces en lutte biologique
Notre choix des Streptomyces en vue d'une lutte biologique contre la fusariose
du pois chiche est basé essentiellement sur plusieurs propriétés associées au à ce genre
qui peuvent expliquer leur capacité d'agir comme agent de lutte biologique, ces
propriétés sont:
Leur adaptation écologique aux sols des régions arides leur attribuant une forte
compétitivité.
Leur capacité de coloniser la surface des plantes et les sites d'infections
(Dubey et al., 2007; Doumbou et al., 2002).
Leur capacité de produire différents types de métabolites secondaires et des
substances biologiquement actives d'une valeur hautement commerciale
comme les enzymes et les antibiotiques. Les enzymes dégradent directement
les parois des champignons et les phythotoxines (Lee et Hwang, 2002;
Doumbou et al., 2002; Grabley et Thiericke, 1999).
Les Streptomyces sont des contributeurs au tampon biologique des sols et ont
un rôle dans la décomposition de la matière organique (Lee et Hwang, 2002;
Grabley et Thiericke, 1999).
Certains Streptomyces améliorent la croissance des plantes (Merriman et al.,
1974; Brown., 1974).
Ce genre de microorganisme est bien adapté à la formulation et l'application
dans le champ (Landa et al., 1997).
Les produits issus des Streptomyces ne laissent pas des résidus toxiques à
l'opposition des produits chimiques (Dubey et al., 2007).
Cependant, plusieurs inconvénients sont associés à l'utilisation des
Streptommyces sp. en lutte biologique, parmi ces inconvénients plusieurs auteurs ont
rapporté que quelques composés antibiotiques produits par les Streptomyces sont
néfaste pour les microorganismes bénéfiques comme les Rizobium (Foo et verma,
1976; Mall, 1973; Von Schreven, 1964; Vintika et Rosa, 1961; Fravel, 1988).

Synthèse bibliographique
44
4. Moyens de lutte contre la fusariose du pois chiche
La protection des plantes contre les maladies représente une part importante
des interventions phytosanitaires sur de nombreuses cultures, tant en fréquence
d’application qu’en volume de produits utilisés. La demande de méthodes alternatives
dans ce domaine est considérable et a suscité une forte augmentation des efforts de
recherche ces dernières années (Nicot, 2002). La plupart des moyens utilisés pour
lutter contre les maladies et les ravageurs ne sont pas durables, les moyens de contrôle
de F. oxysporum, hormis la résistance génétique, sont assez peu efficaces du fait que
ce champignon peut survivre de nombreuses années dans le sol en absence d’hôte (au
moins 06 ans sous forme de chlamydospores) sans prendre en compte les capacités
saprophytiques du champignon qui augmentent ces chances de survie. (Haware et al.,
1996; Nene, 1980), et cela rend son contrôle difficile (Sharma et Muehlbauer, 2007),
différents moyens de lutte ont été préconisés en vue de réduire les pertes dues à la
fusariose du pois chiche.
4.1. Pratiques culturales
L'utilisation des semences non infectées ou traitées pour prévenir l’infection
du pois chiche durant la saison de croissance (Chérif et al., 2007), la destruction des
débris de récolte des plantes fusariées et la rotation culturale du pois chiche de plus de
06 ans avec des cultures non hôtes (Chérif et al., 2007; Gupta, 1991; Haware et Nene,
1982), peuvent agir sur la densité de l'inoculum présent dans le sol, mais l'éradication
du champignon par ces techniques ne parait pas très efficace, car le Fusarium
oxysporum, même en absence de son hôte, peut survivre à l'état saprophytique dans
les débris végétaux et la rhizosphère des plantes non hôtes (Edel et al., 1995; Booth,
1971). Le contrôle du flétrissement vasculaire du pois chiche peut être atteint par
l'ajustement des dates de semis (Chérif et al., 2007; Landa et al., 2004; Navas-Cortés
et al., 2000; Jimenez-Diaz et al., 1998; Navas-Cortés et al., 1998; Jalali et Chand,
1992; Jimenez-Diaz et al., 1991), l'avancement de la date de semi du pois chiche du
début de printemps vers la fin de l’hiver empêche le développement épidémique du
flétrissement et minimise la sévérité de la maladie (Navas-Cortes et al., 1998).
Cependant, l'efficacité de ce contrôle est influencée par la sensibilité des cultivars du

Synthèse bibliographique
45
pois chiche, la nature des races de FOC et sa densité d'inoculum (Navas-Cortes et al.,
2000).
L'ensoleillement des sols avant le semis pendant 06 semaines semble être
efficace pour réduire l'incidence de la fusariose du pois chiche (Mukerji et al., 2009).
4.2. Méthodes chimiques
Les produits chimiques sont efficaces pour lutter contre certaines maladies,
mais leur coût est trop élevé. En plus, de nombreuses études démontrent les risques
que représentent les pesticides sur la santé humaine et l’environnement (Ploetz, 2000;
Ploetz et al., 2003). L'inoculum de FOC présent dans les semences peut être éradiqué
par l'enrobage des graines à l’aide des fongicides tels que: Benlate -T, Benomyl 30%
+ Thiram 30% à 1.5% (Haware et al., 1978). Kaur et Mukhopadhyay, (1992) ont
signalé que le traitement des graines par les deux fongicides: Vitavax et Ziram réduit
l'incidence de la fusariose (Kaur et Mukhopadhyay, 1992). Pour lutter contre ce
parasite, les fongicides sont souvent d’un emploi facile mais ils présentent des
inconvénients: perte rapide d’efficacité, risque de résidus dans les plantes, coût
souvent trop élevé et aide à indiscriminé la microflore bénéfique du sol (Sharma et
Muehlbauer, 2007). En plus, l’utilisation de ces molécules chimiques est souvent
remise en cause en raison de ses impacts sur l’environnement et des problèmes de
santé humaine qu’elle entraîne (Ploetz et al., 2003; Ploetz, 2000). Face à une demande
croissante pour des alternatives à la lutte chimique, les efforts de la recherche ont
abouti à la mise sur le marché de plusieurs produits commerciaux à base de micro-
organismes. Ces produits représentent encore une très faible fraction du marché
mondial des fongicides, laissant entrevoir une marge confortable d’expansion. De ce
fait, la lutte biologique à l’aide d’agents biologiques (bio-pesticides) pourrait
représenter une solution alternative à la lutte chimique classique (Thakor, 2006).
4.3. Méthode biologique
Les systèmes de la lutte biologique n'élimine ni la pathogène ni la maladie
mais les mis en balance naturelle (Dhingra et Sinclair, 1995), cependant les avantages
sont la sécurité environnementale, le coût et le degré de protection (Dubey et al.,
2007).

Synthèse bibliographique
46
4.3.1. Utilisation des microorganismes
La lutte biologique avec les antagonistes est devenue une des plus
prometteuses alternatives à la lutte chimique contre les maladies fongiques. Les
produits d’origine naturelle, sont plus respectueux de l’environnement et de la santé
des utilisateurs et des consommateurs, pourront être introduits dans des programmes
classiques pour permettre de diminuer l’apport d’intrants chimiques. Différents
microorganismes (Bactéries, champignons) ont été signalé par plusieurs auteurs pour
lutter contre la fusariose du pois chiche, tel que les bactéries appartenant aux genres
Bacillus (Duijff et al., 1999; Kumar, 1999; Hervas et al., 1998; Hervàs et al., 1997;
Landa et al., 1997; Lemanceau et Alabouvette, 1993), Pseudomonas (Landa et al.,
2004; Landa et al., 2001; Duijff et al., 1999; Kumar, 1999; Hervas et al., 1998;
Hervas et al., 1997; Lemanceau et Alabouvette, 1993), Burkholderia spp. et
Paenibacillus spp. (Duijff et al., 1999; Hervas et al., 1998; Lemanceau et
Alabouvette, 1993), Rhizobium (Chérif et al., 2007; Arfaoui et al., 2006) et des
souches de FOC non pathogènes (FOC avirulent) (Hervàs et al., 1997), des
actinomycètes (Dhedhi et al., 1990) et d'autres genres appartenant au groupe des
champignons, tel que le champignon mycorhizien, Glomus spp. (Rao et Krishnappa,
1995), Trichoderma harizianum et T. viride (Kaur et Mukhopadhyay, 1992) et
Gliocladium virens (Raju et al., 2008; Chérif et al., 2007; Landa et al., 2004; Landa
et al., 2001; Hervas et al., 1997; Singh et al., 1993).
4.3.2. Méthode génétique
4.3.2.1. La résistance variétale
Cette résistance est dite soit spécifique soit partielle, selon que l’on se trouve
en présence d’un ou plusieurs gènes de résistance variétale. Le choix variétal peut être
intégré au raisonnement de la lutte contre le flétrissement du pois chiche. Certaines
variétés sont tolérantes et parfois, dans certaines situations (modes de contamination,
semences et/ou sol) témoigner des résistances. Ces dernières peuvent être valorisées
en agriculture. L'utilisation des cultivars résistants est une des stratégies les plus
pratiques et la moins couteuse pour le contrôle de la fusariose du pois chiche, mais le
déploiement des variétés résistantes n'a pas été extensive à cause des caractères
agronomiques parfois indésirables (Upadhyaya et al., 2007).

Synthèse bibliographique
47
En plus, leur résistance est non durable à cause de la présence des races
physiologiques de FOC (Sharma et Muehalbauer, 2007) et certains cultivars
présentent une sensibilité à d’autres maladies (Sharma et al., 2009; Jimenez-Gasco et
al., 2004; Ansari et al., 2001; Navas-Cortes et al., 2000; Singh et al., 1994; Singh et
al., 1994; Jalali et Chand, 1992; Nene et Reddy, 1987). Les variétés du pois de chiche
cultivées par les agriculteurs ont des niveaux de résistance très différents les unes des
autres. Cette résistance est fonction des gènes présents lors de la création de la variété.
Deux types de résistance sont mis en évidence au niveau du pois de chiche, la
résistance spécifique et la résistance partielle.
La résistance spécifique est déterminée par des gènes dominants aussi
appelés gènes R. Cinq de ces gènes ont été découverts à ce jour (R1 à R5) (Sharma et
al., 2009), ils déclenchent chez la plante une réaction d’hypersensibilité en présence
du parasite, ce qui limite l’extension du parasite aux tissus voisins. Cette voie de
résistance permet un retard du début de l’épidémie jusqu’à ce que la plante se trouve
en présence d’une race de fusariose possédant les gènes de virulence qui
correspondent à ses propres gènes d’hypersensibilité et peut alors se retrouver
rapidement détruite (Jimenez-Casco et al., 2004; Flor, 1971).
La résistance partielle est déterminée par un grand nombre de gènes.
Ce type de résistance va agir sur la pénétration, la dissémination du champignon dans
les tissus ou encore l’intensité de la sporulation. Cette complexité d’intervention rend
difficile les études sur ce type de résistance, mais la faible probabilité de son
contournement par le parasite en fait un support intéressant de recherche génétique
(Madrid et al., 2008).
4.3.2.2. Méthode de défense naturelle
La découverte de molécules secrétées par les micro-organismes ou présentes
dans leurs surfaces cellulaires appelées « éliciteurs », capables d’induire l’ensemble
des réactions de défense des plantes, a permis d’approfondir leur étude. Les gènes de
défense correspondants ont été isolés, ce qui a conduit à montrer que leur expression
est précocement stimulée lors de l’incompatibilité, alors qu’elle est beaucoup plus

Synthèse bibliographique
48
tardive en situation compatible c’est-à- dire lorsque les plantes sont sensibles (Adam,
2008).
L’acide salicylique et l’éthylène stimulent également la défense des plantes
(Chérif et al., 2007). Selon des voies dont l’importance relative reste à évaluer. L’état
actuel des connaissances permet d’apprécier la puissance et la diversité des
mécanismes d’adaptation des plantes aux agressions causées par les micro-organismes
pathogènes. Les gènes de résistances et de défense sont autant de cibles pour la
conception rationnelle de nouvelles approches de lutte contre les maladies parasitaires
(Adam, 2008).
4.3.2.3. Résistance induite
L’induction des mécanismes de défense chez le pois chiche par une
application des inducteurs biologiques est une nouvelle stratégie de protection des
plantes. Des études ont indiqué l'augmentation de la résistance contre la fusariose
dans les semences traitées par les composés naturels (acide salicylique, spermine) et le
FOC avirulent du pois chiche (Chérif et al., 2007). Les composés phénoliques
contribuent à l’amélioration de la force mécanique de la paroi cellulaire de la plante
hôte du pois chiche et peut aussi inhiber la croissance du champignon, car les
phénoliques sont de nature toxique vis-à-vis des champignons (Raju et al., 2008).
Plusieurs études ont révélé que la résistance induite à travers l’accumulation de
composés phénoliques et de phytoaléxines, ainsi que l’activation des peroxydases, des
polyphénol oxydases et des enzymes clés des voies des phénylpropanoides et des
isoflavonoides, peut jouer un rôle crucial dans la lutte biologique et la résistance du
pois chiche vis-à-vis des attaques pathogéniques (Chérif et al., 2007).
4.3.2.4. Utilisation des espèces sauvages de Cicer
42 espèces sauvages de Cicer rapportaient par (Van Der Maesan, 1987),
présentent des niveaux élevés de la résistance au flétrissement (Singh et al., 1998).
Ces espèces de Cicer ont montré également une multiple résistance aux stress biotique
et abiotique (Singh, 1990) (Tableau 11).

Synthèse bibliographique
49
Tableau 10. Résistance multiple chez Cicer sp. (Singh, 1990).
Winter et al., (2000) ont rapporté l'utilité des espèces sauvages dans la
localisation des gènes de résistance au flétrissement vasculaire, après avoir hybridé C.
arietinum x C. reticulatum. Une autre utilité est d'obtenir des hybrides résistants au
flétrissement (Singh et Ocampo, 1997), à coté de l'utilisation des espèces sauvages
d'autres chercheurs s'intéressent de l'amélioration de la résistance en utilisant des
hybridations entre les variétés de C. arietinum (Sudupak, 2004; Winter et al., 2000;
Singh et Ocampo, 1997).
Tableau 11. Principales espèces de Cicer utilisées dans les programmes d'amélioration de la
résistance du pois chiche contre le flétrissement vasculaire
Espèces Références
C. arietinum (Rao et al., 2007; Sethy et al., 2006; Sudupak, 2004; Iruela et al., 2002;
Winter et al., 2000; Singh et al., 1998; Singh et Ocampo, 1997;
Ladizinsky, 1975).
C. bijugum RECH (Van Der Maesan et al., 2007; Sethy et al., 2006; Sudupak, 2004;
Iruela et al., 2002; Singh et al., 1998).
C. chorassanicum (Bunge)
M popov
(Sethy et al., 2006; Singh et al., 1998; Tayyar et Waines, 1996).
C. cuneatum HOCHST ex.
Rich
(Berger et al., 2003; Sethy et al., 2006; Iruela et al., 2002; Singh et al.,
1998).
C. echinospermum DAVIS (Sethy et al., 2006; Sudupak., 2004; Berger et al., 2003; Iruela et al.,
2002; Singh et al., 1998; Singh et Ocampo, 1997).
C. judaicum BOISS (Sethy et al., 2006; Sudupak., 2004; Iruela et al., 2002; Singh et al.,
1998; Tayyar et Waines, 1996; Ladizinsky, 1975).
C. pinnatifidum JAUB et
SPACH
(Sethy et al., 2006; Sudupak., 2004; Iruela et al., 2002; Singh et al.,
1998; Tayyar et Waines, 1996).
C. reticulatum LADIZ (Rao et al., 2007; Sethy et al., 2006; Sudupak, 2004; Iruela et al.,
2002; Winter et al., 2000; Singh et al., 1998; Singh et Ocampo, 1997;
Ladizinsky, 1975).
C. yamashitae KIT (Sethy et al., 2006; Berger et al., 2003; Iruela et al., 2002; Singh et al.,
1998).
C. anatolicum (Sudupak, 2004; Iruela et al., 2002; Choumane et al., 2000; Kazan et
Cicer sp. anthracnose flétrissement Nématode cyst Froid
C. bijugum R R R R
C. echinospermum S R S R
C. judaicum S R S S
C. reticulatum S R S R
C. pinnatifidum S NI R R

Synthèse bibliographique
50
Muehlbauer, 1991).
C. incisum K. Maly Sudupak, 2004.
C. Multijugum Iruela et al., 2002.
C. macracanthum Iruela et al., 2002.
C. microphylum Iruela et al., 2002.
C. canariense Iruela et al., 2002.
C. oxydon Iruela et al., 2002.
4.4. La lutte intégrée
Selon Singh et al., (1993), le traitement des grains du pois chiche par une
suspension sporale d'un agent de lutte biologique Gliocladium virens et Foltaf à 2g/Kg
réduit considérablement le flétrissement vasculaire. D’autres auteurs, comme
Mukhopadhyay et al., (1992) ont proposé la Carboxine avec le même agent de lutte
biologique. La solarisation des sols pendant 6 semaines combinée avec l'inoculation
des mycorrhizes (Glomus fasciculatum) suivi d’un traitement avec du Carbosulfan
peuvent réduire le flétrissement du pois chiche (Mukerji et al., 2009). Enfin, d’autres
travaux ont montré que l'application de T. harizianum avec des traitements fongicides
(Vitavax et Ziram) sur des graines, réduit l'incidence de la fusariose (Kaur et
Mukhopadhyay, 1992).
4.5. Application du phénomène d’antagonisme et ses limites
La description des nombreux mécanismes d’antagonismes existant dans la nature
permet d’envisager des applications pratiques dans le cadre de la lutte biologique.
Cependant, il faut bien constater que l’application dirigée des antagonistes ne soit que
de très loin les résultats de la recherche car, pour passer de la démonstration de
l’existence d’antagonisme au laboratoire ou dans une parcelle expérimentale à
l’emploi à grande échelle en lutte biologique, l’antagoniste doit posséder certaines
propriétés.
Aptitude à la colonisation de la rhizosphère et à la conservation dans les sols
(Dubey et al., 2007).
Propriété technologique: multiplication de l’antagoniste, formulation simple à
l’emploi (Hjeljord et al., 2000; Collin et Jacobsen, 2003).

Synthèse bibliographique
51
propriété toxicologique: indemne de pathogénie pour l’homme, les animaux et
les végétaux traités, pas de risque lors de la fabrication (Ploetz et al., 2003).
propriété agronomique: l’apport doit pouvoir s’inscrire dans un itinéraire
technique souvent les meilleures stratégies visant à associer la lutte
biologique à une lutte chimique ou à une amélioration des plantes.
problème économique et psychologique: pas de perturbation importante de
l’organisation de la production, protection moins complète que la lutte
chimique d’où utilisation dans les cas où aucun moyen chimique n'existe.

30
Chapitre 2: Matériel et Méthodes

Matériel et méthodes
45
1. Caractérisation des symptômes de la fusariose et identification de l'agent
causal (FOC)
1.1. Les prospections
Les prospections ont été effectuées afin de caractériser les symptômes de la
fusariose du pois chiche au champ et déterminer l’importance de la maladie dans les
trois régions de culture du pois chiche. Il s’agit des régions de Mostaganem, Tlemcen
et Ain Témouchent. Les plantes présentant les symptômes de la maladie sont
prélevées pour être analysé au laboratoire. Le matériel fongique utilisé provient des
échantillons de plantes infectées, récoltées durant la campagne 2008-2009 entre les
mois de mai et juin (Stade post-floraison et formation de gousses). Les
caractéristiques des parcelles de la culture et les dates de prospections sont indiquées
dans le tableau 14. Ces régions ont été choisies en raison de l’importance des
superficies cultivées du pois chiche. Ces prospections avaient comme objectif:
Localiser les foyers du flétrissement du pois chiche.
Evaluer l’importance de la maladie et identifier les espèces responsables
ensuite évaluer la fréquence de leur répartition.
Confirmer l’identité de l’agent causal responsable de la fusariose
vasculaire de pois chiche sur la base des symptômes de la maladie et des
caractéristiques morphologiques des souches isolées à partir des
échantillons infectés.
Tableau 12. Températures en °C et pluviométries en mm mensuelles des chefs-lieux des
wilayas d’étude en 2009
Mois Tlemcen Ain Témouchent Mostaganem
T° min T° max T° moy Pluv. T° min T° max T° moy Pluv. T° min T° max T° moy Pluv.
Fév. 5.8 16.5 11.15 37 9.9 16.4 13.15 13 5.8 17 11.4 15
Mars 8.5 19.1 13.8 24 11.6 17.8 14.7 34 9.2 19.3 14.25 36
Avr. 8.1 21.6 9.9 42 12.9 20.2 16.55 35 9.6 19.5 14.55 48
Mai 12.7 26.9 19.8 6 16.4 23.8 20.1 5 14.1 26.4 20.25 20
Juin 16.7 31.2 23.95 2 20.1 27.7 23.9 00 17.6 29.3 23.45 1
T° min: Température minimale; T° max: Température maximale; Pluv. : Pluviométri
1.2. Estimation de l’incidence et de la gravité de la maladie au champ

Matériel et méthodes
46
Pendant la compagne agricole 2008/2009 et lors de chaque prospection,
l’incidence et la gravité de la maladie ont été évaluées selon le protocole utilisé par
Trapero-Casas, (1983).
L’incidence de la maladie nécessite uniquement le comptage du nombre
d’unités malades qui peut être exprimé par un pourcentage du nombre total d’unités.
Pour l’évaluation de l’incidence du flétrissement, nous nous sommes basés sur le
protocole adopté par Trapero-Casas, (1983). Ce protocole est le suivant: au niveau de
chaque parcelle, trois rangées d’environ 10 mètres de long chacune, ont été choisies
arbitrairement, les plantes de chaque rangée ont été examinées et le nombre de plantes
présentant des symptômes du flétrissement vasculaire a été relevé. L’incidence de la
maladie (IC) a été ensuite exprimée par le pourcentage de plantes infectées, comptées
dans les trois rangées par rapport au nombre total de plantes qui varie suivant
l’espacement entre les plantes adoptées par l’agriculteur (80 à 130 plantes par rangée).
L’estimation de cet indice a été évaluée selon l’échelle ci-dessous (Trapero-
Casas, 1983): 0 (nulle), 0.1 à 5% (peu élevée), 5.1 à 20% (moyennement élevée), 20.1
à 50% (élevée), > 50% (très élevée).
Tableau 13. Parcelles prospectées des 03 régions choisies: Tlemcen, Ain Témouchent et Mostaganem
Régions N° de la
parcelle
Sites
prospectées
Stade végétatif Date de
prospection
Mostaganem P1 Sidi Lakhdar Floraison 24 Mai 2009
P2 Ouled Bouziane Remplissage des gousses
P3 Zerefa Post floraison
Ain
Témouchent
P1 Terga Remplissage des gousses 02 juin 2009
P2 Ain el Tolba Remplissage des gousses
P3 Ain el Kihel Remplissage des gousses
P4 Ain el Kihel Remplissage des gousses
Tlemcen P1 Remchi Remplissage des gousses
P2 Hennaya Remplissage des gousses
P3 Hennaya Remplissage des gousses
P4 Sabaa chioukh Remplissage des gousses
La gravité de la maladie a été estimée en se basant sur l’évaluation visuelle de
la proportion des parties de la plante atteinte par le flétrissement ou le jaunissement.
Sur une superficie de 03 rangées, les plantes affectées ont été examinées et un indice
IC (%) = Nombre de plantes malades/nombre total de plantes x 100

Matériel et méthodes
47
de gravité leur a été attribué en fonction de l’importance des symptômes (Trapero-
Casas, 1983):
0: pas de symptômes, 1: jaunissement ou flétrissement du 1/3 de la plante,
2 : même symptômes mais affectant les 2/3 de la plante, 3: symptômes identiques affectant la
plante entière, 4: plante morte.
1.3. Obtention du matériel fongique
Le matériel fongique utilisé provient des échantillons de plantes infectées
présentant des symptômes caractéristiques de la maladie. Il s’agit d’échantillons de
tiges, du collet et de racines. Au niveau de chaque site prospecté, les différents
symptômes sont décrits ainsi que le stade végétatif. A partir de chaque champ, 10
plants malades âgés de 4 mois environ ont été entièrement arrachées puis mise dans
des sachets en plastique afin d’être analysés au laboratoire.
1.3.1. Isolement de l’agent causal
La technique d’isolement utilisée est celle décrite par Rapilly, (1968). Elle
consiste à préparer des petits fragments de tiges et de racines d’environ 0,6 cm. Les
fragments sont ensuite soigneusement désinfectés soit à l’aide d’hypochlorite de
Figure 8. Localisation des sites de prélèvement des isolats de FOC

Matériel et méthodes
48
sodium à 8%, soit à l’aide d’alcool à 95% pendant 03 minutes. Ils sont ensuite rincés
trois fois dans trois bains d’eau distillée stérile.
Après séchage entre deux papiers filtres stériles, les fragments de tiges, de
racines et des collets sont déposés dans des boites de Petri contenant du milieu PDA
gélosé (annexe 1) à raison de 6 fragments par boite. Les boites sont ensuite incubées à
l’obscurité et à 28 °C pendant 5 à 6 jours.
1.3.2. Purification et conservation de l’agent pathogène
Les champignons isolés sont fréquemment contaminés par des germes
bactériens ou des espèces fongiques indésirables. Pour éviter les risques de
contamination, des repiquages successifs de manière aseptique sont effectués par
prélèvement des explants choisis au niveau de la zone périphérique des thalles
correspondants à l’espèce Fusarium oxysporum. Selon plusieurs auteurs, le genre
Fusarium oxysporum se distingue nettement des autres champignons par la production
des microconidies et des macroconidies, structures caractéristiques de ce champignon.
Après purification et obtention des cultures monospores, les isolats obtenus et
identifiés selon leur aspect macroscopique et microscopique sont conservés à 4°C
dans des tubes à essai contenant du milieu PDA incliné, afin de constituer une
collection d'isolats qui devrait servir aux études ultérieures.
Préparation des cultures monospores
Le principe de cette méthode est d’obtenir des souches génétiquement
homogènes, afin d’éviter toute variation possible des souches. Selon Booth, (1971),
les cultures monospores sont obtenues de la manière suivante: une suspension de
microconidies est diluée dans de l'eau distillée stérile, de façon à obtenir une faible
concentration environ (100 microconidies / ml).
Cette concentration est ajustée à l’aide de la cellule de Malassez. Une goutte
de la suspension est déposée à l’aide d’une pipette Pasteur stérile, puis étalée en stries
à l’aide d’une anse à la surface d'une couche fine d'eau gélosée en boite de Petri.
Après 24h d'incubation à l'obscurité et à 25°C, les tubes germinatifs issus d'une
microconidie unique sont d'abord repérés au fort grossissement de la loupe

Matériel et méthodes
49
binoculaire, puis prélevés stérilement et déposés séparément sur milieu PDA en boite
de Petri. Pour chaque souche, 10 jeunes thalles issus chacun d’une seule conidie sont
repiqués.
Le comportement de chaque thalle issu de la germination des microconidies
est observé (croissance, aspect du mycélium et pigmentation). Si tous les thalles
présentent des caractères morphologiques identiques entre eux et à ceux de la culture
mère, un seul est choisi pour constituer le clone représentatif de l'isolat de départ.
En revanche, si différents types morphologiques apparaissent dans les clones
issus d'un même isolat, ils sont tous conservés comme tête de clone, chacun étant
représentatif d'une morphologie donnée et d'une éventuelle diversité génétique de la
souche de départ.
Tableau 14 .Répartition des isolats de FOC isolés à partir de tiges du pois chiche présentant les
symptômes du flétrissement vasculaire
1.4. Détermination de la forme spéciale ciceri
Le matériel végétal nécessaire à l’expérimentation nous a été aimablement
fourni par l'ITGC de Saida (Institut Technologique des Grandes Cultures). Dans le but
de confirmer la forme spéciale ciceri de 17 isolats de Fusarium oxysporum, isolées
des segments de la tige du pois chiche, ces isolats ont été testés sur la variété (ILC-
482) reconnue pour sa sensibilité au flétrissement (Harrabi et al., 1985).
1.4.1. Préparation du matériel fongique (l’inoculum)
Pour obtenir un mode d’inoculation qui puisse être valable, nous avons
employé une technique couramment utilisée pour tester les champignons du sol
(Rouhani, 1978). Pour cela, nous avons préparé l’inoculum constitué par 17 isolats de
F. oxysporum ciceri provenant chacun d’une région d’étude. Les isolats ont été
ensemencés chacun dans des boîtes de Petri contenant 15 ml de milieu PDA. Les
Régions Isolats
Tlemcen FOC1- FOC2- FOC3- FOC4- FOC5 -FOC6- FOC7- FOC8- FOC9- FOC10
Mostaganem FOC11- FOC12- FOC13- FOC14
Ain Témouchent FOC15- FOC16- FOC17

Matériel et méthodes
50
boites sont mises à incuber à 25°C pendant une semaine. Au bout de 08 jours, les
champignons ayant suffisamment poussé pour envahir toute la boîte.
L'inoculum est préparé de la manière suivante: le contenu de la boite de Petri
est mis en suspension dans de l’eau stérile, le tout étant ensuite bien agité. L’inoculum
est obtenu par lavage de la surface de culture avec de l’eau distillée stérile. Cette
suspension est recueillie, et la concentration de l’inoculum est ajustée à 106 spores par
millilitre après comptage à l’aide de la cellule de Malassez. L’inoculum est prêt à être
utilisé. D’après certains auteurs (Sharma et Muehlbauer, 2007; Westerlund et al.,
1974), cette concentration est suffisante pour reproduire les mêmes symptômes
observés au champ.
1.4.2. Préparation du matériel végétal
Les graines de la variété du pois chiche ILC-482 sont tout d’abord
désinfectées à l’aide d’une solution d’hypochloride de sodium à 12% pendant 2 à 3
minutes afin d’éliminer toutes traces d’infections superficielles préexistantes. Les
graines sont ensuite mises à germé pendant 4 à 5 jours à 25°C dans des boites de Petri
stériles sur papier filtre humidifié avec de l’eau stérile. Après germination, les graines
sont placées dans des petits pots en matière plastique, percés de quelques trous, afin
d’éviter des engorgements lors des arrosages. Ces pots ont un volume d’environ 100
ml et contenant du terreau. Ces pots sont espacés de 5cm environ, et sont laissées en
chambre de culture pendant 08 jours (température: 24 à 26°C, humidité relative: 90
%, photopériode: 12 heures). Les plantules sont arrosées 2 à 3 fois par semaine. Au
bout 8 jours, les jeunes plantules atteignant le stade de deux feuilles.
Ces jeunes plantules ont été soigneusement prélevées, leurs racines sont
ensuite délicatement sectionnées puis inoculées par une suspension sporale des 17
isolats de FOC. Ce test nous a servi comme test d'identification de la forme spéciale
ciceri.
1.4.3. Technique d’inoculation des plantules du pois de chiche
Les plantules du pois chiche, âgées de 8 jours sont inoculées par trempage des
racines dans une suspension conidienne (106
spores/ml) (El-Aoufir, 2001; Trapero-

Matériel et méthodes
51
Casas, 1983; Westerlund et al., 1974; Erwin, 1958). Les plantules sont délicatement
arrachées, leurs racines sont nettoyées par lavage à l’eau stérile et sectionnées puis
trempées pendant 30 minutes dans une suspension conidienne de FOC. Les plantules
témoins sont traitées de la même manière à l’exception qu’elles sont trempées dans de
l’eau stérile. Après inoculation, les plantules sont repiquées dans les mêmes pots à
raison de 3 par pot. Les pots sont placés dans une serre (environ 25°C et 90 %
d’humidité relative). Un arrosage régulier est effectué par la suite en veillant à ce que
de l’eau soit toujours présente dans la soucoupe. Les expérimentations en petits pots
offrent l’avantage d’être facilement manipulables.
1.4.4. Notation des symptômes
Les plantes inoculées correspondant à la variété ILC-482 ont été examinées au
bout de 40 jours. La sensibilité des plantules du pois chiche est appréciée par des
notations effectuées en cours de la végétation. L’efficacité est mesurée par des
comptages de plantes atteintes. L’étude est complétée par une observation de
noircissements des vaisseaux du xylème, après une coupe longitudinale de la tige. Ces
noircissements témoignent de la réaction de la plante à l’infection du champignon.
1.5. Caractérisation morphologique de l’agent pathogène
1.5.1. Caractérisation culturale et description des différents morphotypes
Après obtention d’une culture pure de l’agent causal, l’identification a été
réalisée par une étude macroscopique et microscopique (Rapilly, 1968), en se référant
à la clé d’identification des champignons (Peres, 1985).
Les cultures de 17 isolats pris de la collection sont effectuées sur milieu PDA.
Ce milieu est communément utilisé pour caractériser les Fusarium (Liddell et
Burgess, 1983; Tousson et Nelson, 1976). Pour chaque isolat, l'ensemencement a été
effectué au moyen d'un disque de 0.8 cm de diamètre prélevé aseptiquement à l’aide
d’un emporte-pièce stérile à partir d’une culture de FOC. Après une semaine
d’incubation à 25°C, l’étude macroscopique est basée sur les caractères suivants:
aspect de la colonie, couleur et pigmentation, développement du mycélium. La
variabilité de la morphologie est appréciée entre divers isolats obtenus. Les différents

Matériel et méthodes
52
morphotypes obtenus de l’agent causal sont ensemencés en déposant un fragment
mycélien dans des tubes à essai ou en boite de Petri sur milieu PDA. Après 5 jours
d’incubation à 25°C, les colonies sont photographiées. Ces cultures ont permis de
décrire les différents types morphologiques rencontrés chez F. pxysporum f. sp. ciceri.
L’étude microscopique a été basée sur l’aspect du mycélium coloré au bleu de
cotton, la forme et la taille des conidiophores, des phialides et enfin, les types de
conidies et leur position sur les conidiophores (Leslie et Summerell, 2006).
L’identification du genre Fusarium ne pose aucun problème, car il se distingue
nettement des autres champignons par ses macroconidies fusiformes, pointues aux
deux extrémités avec des cellules basales en forme de pied (Booth, 1971).
L’identification de l’espèce a été effectuée selon des clés de détermination établies par
Messiaen, (1959).
1.5.2. Etude biométrique des conidies
La caractérisation des 17 isolats retenus pour cette étude a été également
effectuée par une étude biométrique basée sur la mensuration des spores après 6 à 10
jours de culture. L’examen direct au microscope photonique mené d'un micromètre
permet donc d’effectuer ses mensurations (longueur et largeur) des macroconidies,
microconidies et la présence ou l'absence des chlamydospores.
En général, les observations microscopiques ont porté sur 40 conidies
(microconidies et macroconidies) par isolat. Toutes les observations ont été effectuées
au grossissement x100. Des observations microscopiques complémentaires sur le type
de conidies, la forme et le nombre de cloison ainsi que l’absence ou la présence des
chlamydospores permettent une meilleure caractérisation des isolats.
1.6. Etude physiologique des isolats de FOC
1.6.1 Recherches des conditions optimales de croissance mycélienne des isolats
1.6.1.1. Estimation de la croissance de FOC
La technique utilisée est celle décrite par (Kherbanda et Brewer, 1980;
Leach, 1962; Brewer, 1960). La mesure de la croissance mycélienne du champignon

Matériel et méthodes
53
consiste à ensemencer un explant d’environ 08 mm de diamètre du champignon sur
milieu PDA. La croissance mycélienne a été évaluée tous les deux jours par la mesure
de deux diamètres perpendiculaires de chaque colonie pendant 8 jours.
La croissance (moyenne de toutes les valeurs) a été ensuite déduite par
soustraction du diamètre de l’explant initial à partir des valeurs relevées, ainsi la
croissance mycélienne est déterminée selon la formule suivante:
L: croissance mycélienne ; D: diamètre de la colonie ; d: diamètre de l'explant.
1.6. 1.2. Effet de l’obscurité et la lumière sur le FOC
Pour l’étude de l’effet de la lumière et de l’obscurité sur la croissance, les
17 isolats ont été cultivés sur milieu PDA (pH= 5.6) pendant 8 jours, sous les deux
conditions suivantes:
A l’obscurité et à la température de 25°C.
Sous une lumière continue et à la température de 25°C.
1.6.1.3. Effet de la température
La détermination de la température optimale de la croissance mycélienne
de 17 isolats de FOC a été étudiée sur une gamme allant de 10 à 35°C, avec un
intervalle de 5°C. Pour cette expérience, nous avons utilisé le milieu PDA ajusté à un
pH= 5.6. L'incubation est réalisée à l'obscurité. La croissance mycélienne a été
évaluée tous les deux jours par la mesure de deux diamètres perpendiculaires de
chaque colonie pendant 8 jours.
1.6.1.4. Effet du pH
Dans le but d’apprécier l’action du pH sur la croissance mycélienne de 17
isolats de FOC, nous avons utilisé comme milieu de culture le PDA avec une gamme
du pH tamponnée afin d'éviter toute variation lors de la dégradation des éléments
nutritifs par le champignon. La gamme du pH choisie se situe entre 4.5 et 7.5. Les
tampons ont été préparés selon la méthode de Geigy, (1990). Pour réaliser ce test,
L= (D-d) /2

Matériel et méthodes
54
nous avons utilisé le milieu PDA en lui ajoutant des tampons décrits dans le tableau
en annexe 2. Chaque pH obtenu est contrôlé à l’aide d’un pH mètre pour une
éventuelle correction. Après autoclavage, les milieux de culture tamponnés sont
coulés dans des boites de Petri puis ensemencés à l’aide d’un explant mycélien de 8
mm de diamètre, déposé au centre des boites. Les boites sont incubées à 25°C et à
l’obscurité. L’action du pH est évaluée selon la méthode précédemment citée.
1.6.2. Influence des facteurs nutritionnels sur la croissance mycélienne de FOC
Les champignons ont besoin pour assurer leur croissance de composés
organiques notamment des sources carbonées comme source d’énergie et des sources
azotées pour la synthèse des protéines.
Afin de rechercher les sources de carbone et d’azote donnant la meilleure croissance
de FOC, nous avons étudié tout d’abord l’effet de quelques milieux de culture sur la
croissance mycélienne. Nous avons ensuite évalué l’action de quelques sources de
carbone et d’azote.
1.6.2.1. Effet des milieux de culture
Pour toutes les espèces fongiques, le choix d’un milieu de culture dépend
des exigences nutritionnelles du champignon (Bouznad, 1989). Ainsi pour toutes les
espèces de Fusarium spp. le problème se pose différemment puisqu’elles peuvent se
développer sur une large gamme de milieux de cultures qui peuvent être organiques,
minéraux ou semi organiques. Dans cette étude, un certain nombre de milieux de
culture communément utilisés en phytopathologie ont été étudiés:
03 milieux organiques: Malt, PDA, Pois chiche.
02 milieux minéraux: Richard et Czapeck.
02 milieux semi organiques: Sach et Waksman.
La composition respective de chacun de ces milieux est présentées en
annexe1, les cultures de FOC (sous forme d’un explant de 8 mm de diamètre) sont
ensemencées sur les différents milieux de culture puis incubées à 25°C et à l’obscurité

Matériel et méthodes
55
pendant 8 jours. La croissance mycélienne a été évaluée tous les deux jours par la
mesure de deux diamètres perpendiculaires de chaque colonie.
1.6.2.2. Influence de la source de carbone sur la croissance de FOC
Dans le but d’apprécier l’influence de la source de carbone sur la
croissance mycélienne de 17 isolats de FOC, nous avons utilisé comme milieu de
culture du Czapeck dox en modifiant la source de carbone pour chaque test. 03
sources de carbone ont été utilisées: Saccharose, glucose et galactose. Les cultures de
FOC ensemencées sur différentes sources de carbone ont été incubées à une
température de 25°C et à l’obscurité, pendant 8 jours. La croissance mycélienne a été
évaluée tous les deux jours par la mesure de deux diamètres perpendiculaires de
chaque colonie.
1.6.2.3. Influence de la source d'azote sur la croissance de FOC
Dans le but d’apprécier l’influence de la source d’azote sur la croissance
mycélienne des 17 isolats de FOC, nous avons utilisé le milieu Czapeck dox en
modifiant la source d'azote pour chaque test. 03 sources d'azote ont été utilisées:
KNO3, Peptone et NaNO3. Les cultures de FOC ensemencées sous forme d’un explant
de 08 mm de diamètre sur les différentes sources d’azote ont été ensuite incubées à
une température de 25°C et à l’obscurité, pendant 8 jours. La croissance mycélienne a
été évaluée tous les deux jours par la mesure de deux diamètres perpendiculaires de
chaque colonie.
1.7. Répétition et traitement statistique
03 répétitions sont retenues pour chaque paramètre étudié. L'analyse de la
variance des mensurations des isolats et l'étude physiologique de FOC a été réalisée à
l'aide du test de Newman et Keuls à 5 %. La méthode de Newman et Keuls est basée
sur la comparaison des amplitudes observées pour les groupes de deux, trois, ect…,p,
avec l'amplitude maximale attendue à un niveau de signification donnée (Newman,
1939; Keuls, 1952).

Matériel et méthodes
56
2. Recherches de moyens de lutte biologique
2.1. Obtention des bactéries actinomycétales
2.1.1. Prélèvement des échantillons du sol
Les prélèvements du sol ont été effectués en mars 2009, dans deux régions du
sud Algérien: Béchar et Timimoune. Les prélèvements ont été effectués à l’aide d’une
spatule stérile sur une profondeur de 5 à 10 cm. Le sol ainsi récolté est mis dans des
sachets en plastiques. Chaque échantillon est muni d’une étiquette indiquant la date, le
lieu et la profondeur. Les échantillons du sol ont été ensuite acheminés vers le
laboratoire où ils ont été séchés puis tamisés afin de procéder à leur analyse
microbiologique. Le tableau 15 présente les régions et les sites de prélèvement.
Tableau 15. Sites d'échantillonnage des sols pour l'isolement des actinomycètes
Régions Sites de prélèvement Nombre
d'échantillons
Béchar Wakda (à 2km de Béchar) 2
Knadssa (à 22 Km de Béchar) 5
Béni Abbés 1
Timimoune Kali 1
Massine 1
2.1.2. Isolement, purification et conservation des actinomycètes
2.1.2.1. Isolement
Après séchage des échantillons du sol, 50g ont été tamisés pour éliminer les
débris végétaux indésirables. Les échantillons ont été ensuite broyés dans un mortier
stérile. Après homogénéisation, 5g de chaque échantillon sont pesés puis introduits
dans un erlenmeyer contenant 50ml d’eau distillée stérile. Après agitation de la
solution pendant 10 min, une série de dilution allant de 10-1
à 10-4
a été préparée, en
prélevant à chaque opération 1 ml. Ainsi, les deux dernières dilutions (10-3
et 10-4
) ont
servit pour l’isolement des actinomycètes sur milieu ISP2. Pour réaliser cette
opération, nous avons utilisé deux méthodes d'ensemencement: en masse et en
surface.

Matériel et méthodes
57
Ensemencement en masse: Cette méthode consiste à déposer tout d’abord 0.1 ml de la
dilution 10-3
ou 10-4
dans une boite de Petri, puis coulé environ 15 ml de milieu ISP2
en surfusion. Les boites sont agitées par un mouvement circulaire lent, afin
d’homogénéiser le contenu des boites. Les boites sont ensuite incubées pendant 07
jours à 30°C.
Ensemencement en surface : Cette méthode consiste à déposer 0.1 ml de chaque
échantillon correspondant soit à la dilution 10-3
ou 10-4
à la surface du milieu de
culture ISP2 gélosé, puis étalé à l’aide d’un étaloir stérile. Les boites sont incubées
pendant 7 jours à 30°C.
2.1.2.2. Purification des isolats d’actinomycètes
Après une semaine d’incubation, les colonies d’actinomycètes apparues et
observées à l’aide d’un microscope photonique sont repérées d’après leur aspect
macroscopique et microscopique caractéristique. Ces colonies sont en générale des
colonies ronde parfois bombées présentant souvent des traits circulaires
concentriques, difficile à les prélever, présentant différentes couleurs, avec une odeur
caractéristique: la géosmine (odeur de la terre fraichement remuée). Enfin, en général,
toute colonie présentant des filaments courts autour d’elle est retenue, puis purifiée
sur milieu ISP2. Les colonies suspectées d’être des actinomycètes sont reprises
séparément sur boite de Petri contenant le même milieu de culture, elles sont ensuite
purifiées par repiquages en strie. L’étude macroscopique et microscopique a permis
de révéler plusieurs morphotypes d’actinomycètes.
2.1.2.3. Conservation des isolats d’actinomycètes
Après isolement et purification des actinomycètes sur milieu ISP2. Ces
dernières sont ensemencées dans des tubes contenant du milieu GLM incliné
(annexe1). Les tubes sont ensuite incubés pendant une semaine à 30°C puis conservés
à 4°C. Les souches d’actinomycètes sont repiquées régulièrement (tous les 03 mois).
2.2. Etude du phénomène d’antagonisme : F. oxysporum – actinomycètes in vitro
Le choix d’actinomycètes comme agent de lutte biologique contre la fusariose
du pois chiche est basé essentiellement sur deux arguments:

Matériel et méthodes
58
Leur adaptation écologique aux sols des régions arides ou semi-arides leur
attribuant une forte compétitivité.
Leur aptitude bien connue à produire des antibiotiques à large spectre d’action.
L’utilisation des souches d’actinomycètes comme agent de lutte biologique
contre les champignons phytopathogènes du sol, constitue une première étape, elle
permet d’apprécier l’aptitude de ces micro-organismes à inhiber ou à réduire la
croissance de FOC. Pour évaluer l’action antagoniste des isolats d’actinomycètes,
deux méthodes ont été choisies: méthode de confrontation directe et action des
métabolites secondaires.
2.2.1. Méthode de confrontation directe
Le but de cette méthode est de sélectionner des souches d’actinomycètes
capables d’inhiber ou de réduire la croissance mycélienne de 03 isolats de Fusarium
oxysporum ciceri, représentative de chaque région (Tlemcen, Ain Temouchent et
Mostaganem). Ce test est un critère de sélection important, il a été effectué in vitro,
par confrontation directe.
Dans des boites de Petri stériles contenant du milieu PDA (annexe1), un
explant de 8mm de diamètre d’une culture de F. oxysporum f .sp ciceri âgée de 7
jours est déposé, la souche d’actinomycète est ensuite ensemencée à l’aide d’un
écouvillon stérile sous forme d’un trait en tenant compte d’une distance d’environ
2.5cm entre les deux protagonistes.
Les boites sont ensuite incubées à 30 °C, la zone d’inhibition est mesurée
après 8 jours d’incubation. L’évaluation de l’inhibition exercée par les 03 souches
d'actinomycètes présélectionnées est estimée par le calcul du pourcentage d’inhibition
de la croissance mycélienne selon la formule de Wang et al., (2001).
Pourcentage d'inhibition= Diamètre de témoin – Diamètre de l'isolat testé/ Diamètre de témoin

Matériel et méthodes
59
2.2.2. Mise en évidence de l’action des métabolites d’actinomycètes
Cet essai, permet de mettre en évidence le pouvoir inhibiteur des filtrats de
culture des souches d’actinomycètes en inhibant ou en réduisant la croissance
mycélienne de FOC.
2.2.2.1. Test de filtrat de culture: Le but de ce test est de sélectionner les souches
dont les métabolites sécrétés sont les moins favorables à la croissance mycélienne de
FOC.
2.2.2.1.1. Préparation des filtrats de culture d’actinomycètes
Cette méthode consiste à ensemencer les souches d’actinomycètes
présélectionnées suite à la méthode de confrontation directe dans des erlenmeyers
contenant 50ml de milieu ISP2 liquide (annexe1). Les erlenmeyers sont incubés à
30°C. Après 8 jours d’incubation, 25 ml a été prélevés puis centrifugées pendant
10min à 4000 t/mn. Le surnageant a été récupéré à l’aide d’une seringue stérile puis
filtré par passage sur un filtre millipore de 0.20µm de diamètre. Le surnageant obtenu
est utilisé pour ce test.
2.2.2.1.2. Incorporation des filtrats de culture d’actinomycètes au milieu
Dans des boites de Petri, les filtrats de culture stériles sont incorporés volume
à volume (V/V) à un milieu de culture gélosé PDA à double concentration d’agar-agar
maintenu en surfusion à 45°C. Après homogénéisation et refroidissement du milieu,
un explant de 08mm de diamètre est déposé au centre de la boite.
Une boite contenant de l’eau distillée stérile et le même milieu de culture sert
de témoin. Les boites sont incubées à température ambiante pendant 6 jours. Après
incubation, les diamètres des colonies de Fusarium oxysporum ciceri en présence des
filtrats de culture sont mesurés et comparés aux diamètres des colonies des boites
témoins.
2.2.2.2. Technique de puits
Dans des boites de Petri contenant du milieu PDA semi solide (0.7%), 04 puits
ont été creusés à l’aide d’une cloche de Durham stérile. Dans chaque puit, 100µl de

Matériel et méthodes
60
filtrat stérile sont ensuite déposés. Un disque de 8 mm du champignon de FOC est
déposé au centre de la boite.
Les boites sont tout d’abord incubées à une température ambiante pendant 30
minutes, puis transférées dans une étuve réglée à 25°C pendant 5 jours. Le même
protocole a été adopté pour les boites témoins à l’exception des filtrats de culture qui
ont été remplacés par de l’eau distillée stérile. L’action des filtrats de culture sur la
croissance mycélienne de FOC est mesurée. Les deux diamètres perpendiculaires ont
été mesurés et comparé par rapport au témoin.
2.2.2.3. Technique des disques de papier Wattman
2.2.2.3.1. Préparation des filtrats de culture d’actinomycètes
Cette méthode consiste à ensemencer la souche d’actinomycète ACT2
présélectionnées après le test de confrontation directe, dans des erlenmeyers
contenant 50ml de milieu ISP2 liquide (annexe1). Les erlenmeyers sont incubés à
30°C. Après 8 jours d’incubation à température de 30°C, le milieu ensemencé a été
filtré à travers un papier filtre ordinaire.
2.2.2.3.2. Extraction de la molécule antifongique
Le surnageant filtré à travers le filtre ordinaire a fait l'objet d'une extraction
liquide - liquide à l'aide d'une ampoule à décanter, en utilisant le même volume du
solvant n-butanol qui est le meilleur extractant selon, Zitouni et al., (2005).le
surnageant filtré de la culture a été ensuite mélangé avec le n-Butanol volume à
volume. Aprés décantation du mélange 02 phases apparaissent, une phase organique
correspond au solvant organique et la molécule bioactive et une phase aqueuse
correspond au filtrat du milieu de culture ISP2. La couche organique formée a été
évaporé à l'aide d'un rotavapeur de marque BUCHI R-200. L'extrait obtenu sert par la
suite à imbiber les disques du papier Wattman stériles.
2.2.2.3.3. Test de l'activité de l'extrait
Dans des boites de Petri, contenant environ 15 ml de milieu PDA maintenu en
surfusion, 100µl d’une suspension sporale de FOC ajusté à une concentration de 106

Matériel et méthodes
61
conidies/ml sont déposés. Les boites sont ensuite agitées par des mouvements lents.
Après homogénéisation, 03 disques imprégnés préalablement d'extrait de la souche
ACT2 ont été déposés à raison de 03 disques par boite de Petri. Les boites ont été
ensuite incubées pendant 5 jours à 25°C.
L’évaluation de l’action de l’extrait brut de la souche d’actinomycète a été
estimée par la mesure des deux diamètres perpendiculaires de la zone d’inhibition
autour des disques.
2.3. Identification des isolats d'actinomycètes antagonistes
Dans le but d’identifier les 06 souches d’actinomycète isolées des différents
échantillons du sol, ces dernières sont soumises à une étude des différents caractères
morphologiques.
2.3.1. Etude macroscopique
Les caractères morphologiques et culturaux sont déterminés sur 03 milieux de
culture: ISP2, gélose nutritive et milieu Sabouraud (Shirling et Gottlieb, 1966). Les
milieux sont coulés dans des boites de Petri 36h avant leur utilisation afin de contrôler
leur stérilité. Ils sont ensuite ensemencés en stries à partir d'une goutte de suspension
d’actinomycète déposée en bordure de la gélose. L'étude de l’aspect macroscopique
est basée essentiellement sur l'importance de la croissance, le développement du
mycélium aérien, la couleur du mycélium aérien et de substrat ainsi que la production
de mélanine sur chaque milieu est observée après 15 jours d'incubation à 30°C.
2.3.2. Etude microscopique
Après 2 semaines d’incubation à 30°C, les cultures des différentes souches
d'actinomycètes antagonistes retenues après sélection ont été observées sous
microscope optique au grossissement (x100). Pour caractériser l'aspect des
sporophores, une observation in situ a été effectuée sur la morphologie des chaînes de
spores, le mycélium aérien et le mycélium de substrat.

Matériel et méthodes
62
2.4. Essai d'extraction et de séparation de la molécule bioactive
2.4.1. Préparation des filtrats de culture d’actinomycètes
Cette méthode consiste à ensemencer 2 L de milieu ISP2 liquide avec la
souche d’actinomycète présélectionnée. Les erlenmeyers sont incubés à 30°C, sous
agitation. Après 8 jours d’incubation, les cultures ont été prélevées puis centrifugées
pendant 10 min à 4000t/mn. Le surnageant est filtré à travers un disque de papier
filtre.
2.4.2. Extraction liquide - liquide: l'extraction à été faite dans une ampoule à
décanter, en utilisant le n-butanol (Zitouni et al., 2005) volume à volume. La phase
organique a été récupérée puis séché à l'aide d'un rotavapeur de marque BUCHI R-
200.
2.4.3. Chromatographie sur couche mince (CCM): Nous avons utilisé des
bandelettes d'une plaque de CCM de (2cm X 20cm). Le dépôt de l'extrait a été
réalisé sous forme de spot placé à 1cm du bord inférieur de la bandelette et au centre.
20µl de l'extrait utilisé a été déposés progressivement à l'aide d'une pipette pasteur, la
plaque ensuite était séchée sous courant d'air chaud. Dans des cuves en verre
contenant 50ml de système éluant et saturé préalablement par la vapeur du système
éluant utilisé n-butanol/acide acétique/eau (3:1:1) pendant 2h. La plaque de CCM a
été ensuite déposée. Après 15 cm de distance parcourue par le solvant à partir du point
de départ la chromatographie est arrêtée. Le solvant est éliminé.
La visualisation des spots de la plaque CCM a été faite par le révélateur
anisaldehyde /acide sulfurique (Fourati-Ben Fguira et al., 2005) puis à l'aide d'une
ampoule à UV.
2.5. Essai de la lutte biologique in vivo
D’après la littérature, très peu de travaux ont été consacrés à cette maladie et
au moyen de lutte, ce qui explique en partie l’inexistence de mesures de lutte
phytosanitaires efficaces et adéquates contre les attaques de cet agent pathogène. Pour
cette raison nous nous sommes intéressés à l’étude du comportement de F. oxysporum
ciceri en présence d’un agent de lutte biologique connu par ces propriétés

Matériel et méthodes
63
antagonistes, il s’agit d’une espèce appartenant au groupe des actinomycètes. Le
choix de la souche d’actinomycète ACT2 qui a fait l’objet de cette étude est basé sur
les résultats des tests in vitro.
2.5.1. Préparation de l'inoculum
L'actinomycète ACT2 sélectionnée a été ensemencé sur milieu ISP2 puis
incubée à 30°C, pendant 14 jours. 15 ml d'eau distillée stérile a été verser sur la
surface des colonies de l'actinomycète et à l'aide d'un cure dent stérile, les spores de
celle-ci ont été grattées et récupérées dans un tube stérile puis, ajustées à la
concentration 106 spores/ml.
La préparation de l'inoculum de l'agent pathogène a été obtenue par la
méthode décrite précédemment pour la détermination de la forme spéciale ciceri en
utilisant 03 isolats de FOC pris de la collection, il s'agit du FOC8, FOC13 et FOC 15
représentatives des 03 régions d'étude.
2.5.2. Obtention des plantules du pois chiche
L'obtention des plantules du pois chiche a été réalisée par la méthode décrite
prédemment pour la détermination de la forme spéciale ciceri. 03 variétés du pois
chiche ont été testées, une sensible (ILC-482) et les deux autres sont modérément
tolérantes (Col-27 et PPC-25). Au bout de 8 jours, les plantules arrivées au stade de
deux feuilles ont été inoculées par la suspension sporale de la souche d'actinomycète
ACT2, après 24h la suspension sporale de 03 isolats de FOC à été inoculés. Des
plantules non inoculées et d'autres inoculés uniquement par le champignon servent
comme témoin ont été ajoutées.
2.5.3. L'estimation des symptômes
Les plantes inoculées ont été examinées au bout de 40 jours et le nombre de
feuilles montrant des symptômes de jaunissement a été noté pour chaque plante. La
gravité de la maladie exprimé en indice de maladie (IM) a été évaluée par la formule
(Molot et Mas, 1975; Besri et al., 1984).
IM= Nombre de feuilles atteintes / nombre total de feuilles x 100

Matériel et méthodes
64
En comparant l'indice de la maladie chez les plantes du pois chiche inoculées
seulement par le champignon et ceux inoculées par le champignon et l'actinomycète
en déduit le degré de protection fournit par la souche d'actinomycète ACT2.
3. Détermination de l'agressivité et du comportement variétal
3.1. Détermination de l'agressivité
La détermination de l'agressivité a été réalisée sur la variété sensible ILC-482
au FOC. Les techniques de préparation de l'inoculum, la préparation du matériel
végétal sont les mêmes que celles procédé pour la détermination de la forme spéciale
ciceri. L'inoculation a été réalisée sans avoir sectionné les racines des jeunes plantules
obtenues après 8 jours de semi.
L'estimation des symptômes a été évaluée par la formule de Molot et Mas,
(1975) et Besri et al., (1984).
3.2. Etude du comportement variétal
Cette étude a été réalisée sur une gamme de variétés du pois chiche (11
variétés). Les 11 variétés du pois chiche utilisées sont: ILC 3279, ILC-482, Col-27,
PPC-25, Fedjoj, Haddache, 39-96, 2G-03, F10-31, F10-13, F7-43 offrit aimablement
par INRA de Sidi Bel-Abbés et l'ITGC de Saida.
Les techniques de préparation de l'inoculum, la préparation du matériel
végétale et la technique d'inoculation sont les mêmes que celles procédé pour la
détermination de la forme spéciale ciceri.
3.2.1. Estimation des symptômes
Les plantes des 11 variétés du pois chiche inoculées ont été examinées au bout
de 40 jours et le nombre de feuilles montrant des symptômes de jaunissement et de
flétrissement a été noté pour chaque plante. L'estimation de la réaction des cultivars
du pois de chiche inoculés a été réalisée suivant l'échelle adoptée par Haware et Nene
(1982). Cette échelle est établie en fonction de la gravité des symptômes évalués par

Matériel et méthodes
65
le pourcentage des feuilles jaunies ou nécrosées par rapport au nombre total des
feuilles 40 jours après l'inoculation:
0 à 20%: la réaction est qualifiée de tolérance ; 21 à 50%: la réaction est une sensibilité modérée.
> 50% : la réaction est une sensibilité élevée.
3.2.2. Ré isolement du champignon pathogène
Pour confirmer l’identité de l'agent pathogène, les plantes des variétés
inoculées ont été récupérées en vue de réisoler l'agent pathogène (vérification du
Postula de Koch). Le réisolement a été fait par incubation des fragments de tige de
toutes les plantes après désinfection à l’eau de javel, sur le milieu PDA, à 25 C°. Les
cultures fongiques obtenues sont comparées à leurs cultures mères. Le pourcentage de
réisolement positif (%RP) est exprimé par la formule suivante:
Ce test permet de mettre en évidence si le champignon est présent, s’il est
vivant et s’il est capable de développer les mêmes symptômes observés dans la nature.
%RP= Nombre de fragments colonisés/Nombre de fragments testés x 100
IM= Nombre de feuilles atteintes / nombre total de feuilles x 100

45
Chapitre 3: Résultats et discussion

Résultats et discussion
91
1. Caractérisation des symptômes de la fusariose et identification de l'agent
causal (FOC)
1.1. Importance du flétrissement du pois chiche dans les 03 régions d'études
1.1.1. Résultats
1.1.1.1. Observation des symptômes au champ
Les différentes prospections effectuées au champ nous ont permis d’observer
les symptômes caractéristiques de la fusariose sur culture du pois de chiche. Les
symptômes du flétrissement sont souvent plus graves entre 18 et 25°C. Les
symptômes provoqués par F. oxysporum ciceri s’observent régulièrement à peu près
chaque année dans les régions du nord ouest d’Algérie (Figure 9). L’importance de
l’épidémie varie d’une année à l’autre et d’une région à une autre.
Les symptômes observés aux champs correspondent à un relâchement des
pétioles, un flétrissement rapide des feuilles avec une couleur vert-terne, suivi du
desséchement de la plante entière avant même le stade de floraison. Pour d'autres
plantes, les symptômes se sont manifestés plus tard sous forme d'un jaunissement
vasculaire caractéristique, soit unilatéraux (Figure 10A), soit généralisés (Figure
10B). Le flétrissement du feuillage atteigne tout d’abord, les feuilles les plus basses et
progressent ensuite vers le haut (Figure 11). Ce dernier cas était le plus fréquent
observé dans les parcelles prospectées.
Figure 9. A: une culture du pois chiche à Ain Temouchent présentant un foyer malade; B: culture du
pois chiche présentant plusieurs foyer malades à Mostaganem.
A B

Résultats et discussion
92
Dans les deux manifestations pathologiques, une coupe longitudinale suivant
l'axe de la plante avait révélé un brunissement des tissus internes des tiges,
témoignant de leur colonisation par l'agent pathogène (Figure 10 C).
Le système racinaire des plantes malades présente une réduction des racines
secondaires (Figure 11). L'examen à l’œil nu des graines issues de plantes malades a
montré une réduction de leur taille par rapport aux graines saines, en plus leur couleur
était plus foncé que les graines normales.
Dans les parcelles de Mostaganem, la maladie ne s'est déclarée que
tardivement en post floraison, la manifestation pathologique observée était du type
jaunissement vasculaire caractéristique du flétrissement tardif. Dans la région de
Tlemcen et Ain Temouchent et surtout à Tlemcen, nous avons remarqué la présence
de jeunes plantules entièrement sèches à stade pré-floraison correspondant au
flétrissement précoce.
Figure 10. A: Plants du pois chiche, présentant une attaque de fusariose unilatérale observée en moi de mai;
B: Plants de pois chiche, présentant une attaque de fusariose généralisée; C: coupe longitudinale d'une tige
du pois chiche présentant un noircissement du xylème, observée dans une parcelle à Tlemcen.
A B C

Résultats et discussion
93
A l’exception de ces sujets présentant les manifestations épidémiques
précitées, dans l’ensemble les cultures étaient saines et indemnes de tous symptômes
de maladie.
1.1.1.2. Etude de l'incidence et de la gravité de la maladie
Les prospections effectuées dans les trois régions du nord ouest d’Algérie ont
permis d’observer les symptômes caractéristiques de la fusariose. La distribution de la
maladie varie d’une région à l’autre. Dans les régions d'Ain Témouchent et Tlemcen,
les prospections ont permis de constater que, dans plusieurs champs de culture du pois
chiche, la maladie était présente et a entraîné des dégâts importants. En revanche,
dans la région de Mostaganem, la maladie était rarement observée et parfois absente.
Les valeurs d'estimation de l'incidence et de la gravité de la maladie de la
fusariose du pois chiche sont présentées dans le Tableau 16. Ces deux paramètres ont
été évalués par analyse des racines des plantes d'apparence saines et qui montrent des
symptômes du flétrissement.
Figure 11. Une réduction des ramifications secondaires sur le système racinaire
des plantes malades de pois chiche par rapport aux racines des plantes saines
Bas
Plante
malade
Plante
saine
Haut

Résultats et discussion
94
Tableau 16. Incidence (I) et gravité (S) du flétrissement vasculaire dans les parcelles de culture du pois
chiche prospectées.
Régions Les parcelles Moyenne Intervalle Total
Incidence ou
gravité
P1 P2 P3 P4
Mostaganem Incidence% 24.33 21.28 7.43 --- 17.68 7.43-24.33 I=
53.90 Gravité 1.06 1.02 0.18 --- 0.75 0.18-1.06
Ain
Temouchent
Incidence% 65.89 66.66 30.76 84.61 61.98 30.76-84.61
Gravité 0.61 1.45 0.40 2.30 1.19 0.40-2.30 S= 1.82
Tlemcen Incidence% 92.30 94.87 74.35 66.66 82.04 66.66-94.87
Gravité 2.64 1.91 1.51 1.98 2.01 1.51-2.64
Incidence (I): est exprimée en % ; elle est nulle (0), peu élevée (0-1.5), moyennement élevée (5.1-20),
élevée (20.1-50), très élevée (>50). Gravité (S): la maladie est peu grave (0<S<1), moyennement grave
(1<S<2), grave (2<S<3), très grave (3<S<4).
L'incidence de la maladie varie de 7.4 à 94.87% avec une moyenne de 53.90%
pour toutes les parcelles prospectées, ainsi que, pour la gravité de la maladie, qui varie
de 0.18 à 2.64 avec une moyenne de 1.82 (Tableau 16).
A Mostaganem, les 03 parcelles prospectées ont montré un pourcentage de
plantes malades faible par rapport aux deux autres régions prospectées. Il est
d’environ 7.68% et un indice de gravité de 0.75.
A Ain Témouchent, les 04 parcelles prospectées montrent un pourcentage de
plantes infectées plus important par rapport à la région de Mostaganem, alors qu’il est
moins élevé que la région de Tlemcen. L'incidence de la maladie dans cette région est
de l'ordre de 61.98%, alors que, la gravité est à environ 1.19. Au niveau des plaines
intérieures, les 04 sites agricoles de la région de Tlemcen montrent des plantes
malades dont l'incidence et la gravité avoisine 82.04% et 2.02 respectivement.
Au niveau des régions prospectées, l'incidence de la maladie a témoigné une
gravité élevée de cette épidémie, les deux paramètres étudiés (l'incidence et la gravité

Résultats et discussion
95
de la maladie) induite par le FOC, s'avère augmenté au fur et à mesure du
développement de la culture.
1.1.1.3. Détection et isolement de l’agent pathogène
A travers les différentes prospections, plus d’une vingtaine d’échantillons ont
été analysés au laboratoire, ces échantillons ont pour la plupart été prélevés au hasard
pendant nos prospections. Des fragments d’organes (tiges, collets ou racines)
présentant des symptômes sont déposés sur milieu de culture dans des boîtes de Petri
puis incubées à 25°C pendant 4 à 5 jours.
Les analyses réalisées au laboratoire ont révélé la présence régulière de F.
oxysporum, champignon de la famille des Moliniacées. Suite aux différentes
caractéristiques des isolats et en tenant compte de la plante hôte, nous pouvons
supposer que cette espèce correspond à la forme spéciale F. oxysporum ciceri. Donc,
cette dernière est impliquée dans le développement de cette épidémie affectant
l’ensemble des régions productrices dans l’ouest algériens.
Néanmoins, il reste à tester son agressivité sur une variété du pois chiche
sensible, afin de pouvoir déclarer que cette espèce est strictement inféodée à ce
matériel végétal (vérifier le Postulat de Koch) et que cette dernière est bien pathogène
et responsable des symptômes sur culture du pois chiche observés au champ.
Figure 12. Colonies de F. oxysporum ciceri obtenues sur boite de Petri, contenant le milieu PDA
A: Face inférieure ; B: Face supérieure.
A B

Résultats et discussion
96
Tableau 17. Nombre des isolats obtenus par région et par partie de plantes infectées
Région Racines Collet Tige Total Nombre Total
Tlemcen 12 21 10 43
65 Mostaganem 03 03 04 10
Ain Temouchent 04 05 03 12
Les résultats de l'analyse au laboratoire ont permis d’isoler l’agent causal de
cette maladie. Les figures 12A et 12B représentent les colonies de F. oxysporum f. sp.
ciceri obtenues sur milieu PDA. Le champignon est facilement reconnaissable grâce à
son thalle souvent blanchâtre. La détection de ce parasite est donc aisé, même lorsque
les fragments sont contaminés par des bactéries, des champignons saprophytes ou
parasites de faiblesse comme les Alternaria, les Aspergillus ou les Penicillium. En ce
qui concerne l’isolement, c’est-à-dire l’obtention de F. oxysporum ciceri en culture
pure, il ne pose aucun problème particulier. En effet, la croissance initiale est très
rapide et il suffit donc d’opérer les prélèvements aussi tôt que possible. L’aspect de la
fusariose du pois chiche s’est révélé assez homogène d’un plant à un autre et d’une
région de culture à une autre.
Nom
bre d
es isolats
Figure 13. Histogrammes présentant la répartition des isolats de
FOC par région et par partie de plante.

Résultats et discussion
97
Au total une collection de 65 isolats de FOC, a été obtenue après isolement et
purification, dont 26.15 % à partir de tige, 44.61% à partir du collet et 29.23% à partir
des racines. La répartition du nombre d'isolats de la collection obtenus de FOC par
région est représentée par la Figure 13.
Parmi les 65 isolats de FOC obtenus à partir des différents échantillons
analysés, 43 ont été obtenus de la région de Tlemcen, 12 de la région d'Ain-
Temouchent et 10 provenant de la région de Mostaganem. Les résultats d’isolement
de l’agent pathogène ont montré que le pourcentage d’isolement était beaucoup plus
élevé à partir des fragments du collet par apport à ceux des tiges et de racines.
Le pourcentage de réussite d’isolement de F. oxysporum ciceri a varié de 60 à
80%, selon les fragments mis en culture sur milieu PDA.
1.1.1.4. Détermination de la forme spéciale ciceri
Les plantes du pois chiche de la variété ILC-482 connue par sa sensibilité au
flétrissement vasculaire ont été inoculées par 17 isolats de FOC (après blessure des
racines), en vue de déterminer la forme spéciale ciceri. Tous les isolats ont provoqués
les symptômes typiques de la fusariose similaires aux celles observés aux champs.
Ces symptômes ont été observés deux semaines après inoculation chez certains isolats
et ont progressé très rapidement en entrainant la destruction totale des plantules après
30 jours. Les plantes témoins maintenues dans les mêmes conditions n'ont manifesté
aucun symptôme. En tenant compte de ces résultats, nous pouvons conclure que nos
isolats testés appartiennent à la forme spéciale ciceri.
1.1.2. Discussion
1.1.2.1. Observation des symptômes
Le flétrissement vasculaire peut être accompagné d'une pourriture racinaire
(Raju et al., 2008) quand il est induit par F. solani (Rhrib, 1990; Westerlund et al.,
1974; Kraft, 1969) ou de nécroses des racines et du collet quand il est associé à
certains biotypes de l'espèce Fusarium oxysporum (Rhrib, 1990; Jimenez-Diaz et
Trapero-Casas, 1988).

Résultats et discussion
98
Dans d'autres cas les racines gardent une apparence saine et ne montrent ni
pourriture, ni dessèchement, un tel symptôme a été décrit comme étant la conséquence
de l'infection par le FOC (Shah et al., 2009; Cunnington et al., 2009; Dubey et Singh,
2004; Nene et al., 1991; Haware, 1988; Nene et Reddy, 1987). Les plantules du pois
chiche détruites par la maladie du flétrissement peuvent être confondues avec d’autres
maladies du complexe du flétrissement si on ne les examine pas avec précaution
(Pande et al., 2007). L'examen des racines des plants infectés peut différencier entre
le flétrissement vasculaire (la fusariose) et les autres maladies du complexe du
flétrissement chez le pois chiche. Les symptômes des deux maladies sont souvent
identiques mais la différence réside au niveau des racines.
Fusarium oxysporum f. sp. ciceri provoque d’abord un flétrissement du
feuillage progressant du bas de la plante vers le haut avec une ramification des racines
secondaire. Il pénètre dans les racines par des blessures et développe le plus souvent
un mycélium blanc. Les premiers symptômes de contamination par F. oxysporum f.
sp. ciceri sont des jaunissements et flétrissements unilatéraux des feuilles. On observe
également sur les tiges des stries longitudinales brunes. Ces symptômes du
flétrissement vasculaire observés dans les parcelles prospectées sont similaires à ceux
rapportés par Westerlund et al., (1974) en Californie, par Nene et Reddy, (1987) et
Haware, (1988) en Inde, par Jimenez-Diaz et Trapero-Casas, (1988) et Trapero-Casas
et Jimenez-Diaz, (1985) en Espagne, par El-Aoufir, (2001) au Maroc, par Halila et
Strange, (1996) en Tunisie et par Bekkar, (2007) en Algérie.
Le symptôme le plus fréquemment observé dans les champs prospectés était
le jaunissement vasculaire induit par le FOC tel qu'il a été rapporté en Espagne
(Trapero-Casas et Jimenez-Diaz, 1985) et qui correspond au flétrissement tardif décrit
par Nene et al., (1979). Etant donnée que les prospections effectuées ont été réalisé au
stade post floraison et remplissage de gousses, nous n’avons pas pu montré la
présence du flétrissement précoce qui correspond au Wilt décrit par divers auteurs (El
Aoufir, 2001; Haware, 1988; Trapero-Casas et Jimenez-Diaz, 1985; Nene et al., 1979;
Erwin, 1958). Cependant, nous avons remarqué la présence de jeunes plantules morte
(stade pré- floraison), ce type du flétrissement qui apparait au stade préfloraison et qui

Résultats et discussion
99
abouti à la mort des plantes avant qu'elles n'atteignent le stade floraison, cause plus de
pertes que le jaunissement vasculaire progressif qui apparaissant plus tardivement.
Le flétrissement précoce causé par le FOC peut être observé chez les
génotypes sensibles 25 jours après le semis (Shah et al., 2009; Raju et al., 2008 ;
Pande et al., 2007; Navas-Cortés et al., 1998 ; Nene et al., 1978), alors que le
flétrissement tardif s’observe au stade de remplissage des gousses (Pande et al.,
2007 ; Navas-Cortés et al., 1998).
D'après les observations faites par Pande et al., (2007), les jeunes plantules
infectées par le flétrissement vasculaire précoce s'écroulent, s'aplaties et s'allongent
sur terre en gardant leur couleur verte sombre. Cependant, les plantes adultes qui
montrent des symptômes du flétrissement tardif, se caractérise par un flétrissement
soudain des feuilles et un affaissement des pétioles, de rachis et de folioles, avec la
présence d’une couleur pale par rapport aux plantes saines et une décoloration du
xylème de la tige (Shah et al., 2009 ; Raju et al., 2008 ; Pande et al., 2007; Dubey et
Singh., 2004). La manifestation de ces deux types de symptômes avec une fréquence
plus grande du jaunissement vasculaire a été rapportée en Algérie par (Bekkar, 2007).
Néanmoins, l'observation de gousses avait souvent révélé la présence de graines
atrophiées, ce qui est en accord avec l'observation de Haware et Nene (1980).
Dans les 03 régions, les conditions de semis du printemps relatives à la
température et à l’humidité du sol sont souvent réunies pour que les spores et les
chlamydospores germent et pénètrent à l’intérieur des vaisseaux de la plante. Lorsque
l’infection est réalisée, le cryptogame se multiplie et progresse à l’intérieur des tissus
de la plante en direction de la partie aérienne. En effet, l’intensité d’une attaque du
flétrissement est en rapport avec l’importance du taux de contamination des variétés
du pois chiche qui peuvent être plus ou moins résistantes et également les diverses
races de la fusariose dont la virulence est plus ou moins importante (Bouznad, 1989).
Cette maladie est toujours prête à reprendre son extension si l’intérêt que l’on peut lui
accordé se relâche.

Résultats et discussion
100
1.1.2.2. Incidence et gravité de la maladie
En Algérie les études réalisées sur les pathogènes du pois chiche ont surtout
porté sur l'anthracnose provoqué par l'agent Aschochyta rabiei, véritable obstacle à la
culture auquel s'ajoute actuellement le complexe du flétrissement et de pourriture
radiculaire appelé syndrome du flétrissement (Rhrib, 1990).
Le flétrissement vasculaire s'est manifesté principalement par un jaunissement
dans toutes les parcelles de culture de printemps. Toutefois la comparaison des
résultats de la présente étude a montré, que les niveaux d'attaque étaient beaucoup
plus élevés (Incidence>50%) dans les parcelles de production du pois chiche
printanier à Tlemcen et à Ain Temouchent. On peut préjuger à ces niveaux d'attaque,
que les rendements seront considérablement réduits par cette maladie.
L'évaluation de l'incidence (pourcentage de plantes infestées) et de la gravité
(proportion de tissu infesté) habituellement employée dans l'étude des maladies
causées par les agents pathogènes telluriques (Campbell et Powel, 1990), a permis
l'estimation de la fréquence de la maladie au niveau des 03 régions prospectées. Son
importance en termes d'incidence et de gravité a varié entre ces régions et d’une
parcelle à une autre dans une même région. Les résultats de l'incidence et la gravité
obtenus par cette étude sont en accord avec autres recherches réalisées dans le nord
ouest de l'Algérie. Ben Freha et al., (2010) ont permis d'identifier une incidence de la
maladie de la fusariose du pois chiche allant de 37.7 à 100% et une gravité entre 1 et
2.79. Selon Mehrez et al., (2010) la moyenne de l'incidence est d’environ 63.77%,
avec une gravité avoisinant 2.97%.
Cette situation peut être la conséquence de certains facteurs qui possèdent une
très grande influence sur la croissance de l’espèce fongique et principalement sur les
possibilités de réalisation de l’interaction hôte – parasite, pour expliquer cette
situation, plusieurs facteurs entre en jeux. Parmi ces facteurs on distingue : les
conditions d'aménagement des parcelles, leur histoire, et surtout les conditions
environnementales: certains auteurs ont rapporté que la température joue un rôle clé
dans le processus d'infection (Landa et al., 2006; Landa et al., 2001; Navas-Cortés et

Résultats et discussion
101
al., 2000; Navas-Cortes et al., 1998; Bhatti et Kraft, 1992; Gupta et al., 1987;
Westerlund et al., 1974).
Plusieurs auteurs ont rapporté que la température affecte le processus
métabolique, la virulence et le développement de la plante, une température entre 20
et 30°C est favorable pour le développement de cette maladie (Sharma et Muehlbauer,
2007; Landa et al., 2006 ; Trapero-Casas et Jimenez-Diaz, 1985). Ce qui est le cas
dans ces régions étant donné que la température avoisine les 25°C pendant cette
saison (Tableau 12).
La température peut affecter les moyens de défense liés la résistance de
certains cultivars aux races de FOC sous les conditions naturelles (Landa et al., 2006).
D’autres observations ont montré que les sols secs durant cette saison, augmente la
sévérité de la maladie (Landa et al., 2004 ; Sugha et al., 1994; Bhatti et Kraft,
1992; Trapero-Casas et Jimenez-Diaz, 1985), ce qui corrèlent avec nos observations
qui indiquent que l'incidence et la sévérité de la fusariose a été maximale en post
floraison et pendant le remplissage des gousses. L'incidence de la maladie peut être
aussi influencé par le génotype du pois chiche cultivé, cette incidence augmente lors
de l'utilisation de génotypes sensibles (Shah et al., 2009; Raju et al., 2008; Pande et
al., 2007; Navas-Cortes et al., 2000; Navas-Cortés et al., 1998; Nene et al., 1978).
La sévérité dépend aussi de la densité d'inoculum (Navas-Cortes et al., 2000;
Sugha et al., 1994; Gupta et al., 1987; Westerlund et al., 1974). Ces derniers auteurs
ont observé que l'augmentation de la charge de l'inoculum augmente la sévérité et la
rapidité de la maladie, par contre sa diminution supprime l'expression de la maladie.
La sévérité élevée de la maladie à Tlemcen et à Ain Temouchent peut être due
probablement au système des monocultures. L'incidence peut être affectée aussi par
les sols infestés par les nématodes du genre Meloidogyne spp. et Pratylenchus
thornei. Les blessures provoquées par certains nématodes prédisposent les racines de
pois chiche à l'attaque par le FOC (Castillo et al., 1998; Uma Maheswari et al., 1997).
Autre facteur déterminant, est les races physiologiques de FOC (Navas-Cortes et al.,
1998), en Algérie, Bekkar, (2007) a proposé la présence de la race 5, considérée
comme la race la plus virulente.

Résultats et discussion
102
1.1.2.3. Détection et isolement de l’agent pathogène
Dans les conditions d'isolement sur milieu gélosé PDA, le FOC a été isolé à
partir des échantillons flétris et/ou jaunis, récoltés lors des prospections. Il est
probable que la fréquence d'isolement soit le résultat d'une augmentation avec l'âge et
de la sensibilité de la plante vis-à-vis de Fusarium telle qu'elle a été démontrée par
certains auteurs (Van Rheenen et al., 1989; Gupta et al., 1987).
Certains échantillons ayant fait l'objet de cette étude n'ont montré aucun
développement du champignon alors qu'au champ, ils avaient manifesté des
symptômes du flétrissement.
Ceci ne peut pas être dû au milieu de croissance qui était adéquat pour révéler
le Fusarium car le PDA est le milieu considéré d'être riche en nutriment comme les
carbohydrates. De plus, le PDA est principalement utilisé pour l'identification des
cultures de Fusarium car il induit la pigmentation et il est utilisé pour observer les
taux de croissance du champignon qui est un caractère important pour l'identification
des espèces (Summerell et al., 2003; Nelson et al., 1983). En effet, il est connu que
les symptômes peuvent se manifester avant l'installation de l'agent pathogène dans
l'organe. Ce cas a été rapporté pour Fusarium oxysporum f. sp. pisi (Winstead et
Walker, 1954) et le FOC (Farooq et al., 2005). Ces agents agissent à distance par
l'émission de métabolites (substances enzymatiques ou toxines). Ce résultat peut être
également expliqué par une absence de certains conditions d’incubation, étant donnée
que les cultures sont souvent maintenues à une température de 25°C jour/20°C la nuit
et incubées sous lumière journalière (Nelson et al., 1983) et n'ont pas à une obscurité
totale. D'autres facteurs indéterminés pourraient être impliqués dans le phénomène
car, il est connu que les affections causées par d'autres agents comme le Verticilium
dahliae (Haware, 1988), les symptômes sont difficilement discernables de ceux
induits par le FOC.
Le flétrissement pourrait aussi résulter de l'obstruction du système vasculaire
des racines qui constitue la voie de pénétration du Fusarium oxysporum dans la
plante, ce qui explique le pourcentage élevé de l'isolement de FOC à partir de racines
et de collet par rapport à la tige.

Résultats et discussion
103
Ce pathogène est donc le plus présent et a été déjà largement incriminé dans
les études antérieures (Ben Freha et al., 2010; Mehrez et al., 2010; Bekkar, 2007).
Cette espèce semble être implantée dans l’ensemble des zones prospectées ou cette
culture est pratiquée. En effet, le F. oxysporum isolé du sol ou à partir du végétal ne
peut pas être classé dans une forme spéciale qu’après inoculation artificielle sur l’hôte
sensible (Laterrot et al., 1978).
1.1.2.4. Détermination de la forme spéciale
Cette expérimentation a pour objectif de reproduire les symptômes du
flétrissement sur la plante du pois chiche en conditions de contamination artificielle.
Le résultat de l’inoculation a été positif sur la variété sensible du pois chiche.
Donc, cette espèce de Fusarium oxysporum est bien pathogène et responsable des
symptômes observés au champ. Cette expérimentation a permis de vérifier le test de
Postulat de Koch. La rapidité d'apparition des symptômes pour ce test est due
probablement aux blessures que nos avons provoquées sur les racines.
L'étude menée sur le comportement des isolats sur la variété sensible du pois
chiche, nous a permis de reproduire les symptômes. Ces symptômes causés par le
FOC sont en effet ceux décrits par plusieurs auteurs (Bekkar, 2007; Honnareddy et
Dubey, 2006; Jimenez-Gasco et al., 2004; Jimenez-Gasco et Jimenez-Diaz, 2003; El-
Aoufir, 2001; Navas-Cortes et al., 1998), il s'agit d'un flétrissement et d'un
jaunissement accompagné d'une décoloration vasculaire au niveau des tiges.
1.2. Etude morphologique de FOC
L'identification morphologique est basée sur les similarités des caractères
morphologiques observables physiques et physiologiques (Leslie et al., 2001). Les
caractères physiques correspondent à la forme et la taille des conidies, cependant les
caractères physiologiques correspondent aux taux de croissance (Taylor et al., 2000;
Leslie et al., 2001) et la sécrétion des métabolites primaires (Desjardins et al., 1992;
Nelson et al., 1983; Torp et Langseth, 1999; Thrane, 2001; Rheeder et al., 2002).
Pour la taxonomie du genre Fusarium la forme des macroconidies est le premier
caractère important pour l'identification des espèces (Gerlach et Nirenberg, 1982;

Résultats et discussion
104
Nelson et al., 1983; Summerell et al., 2003; Leslie et Summerell, 2006). Autres
caractères morphologiques comme les microconidies, les conidiophores et les
chlamydospores sont aussi important dans l'identification des espèces (Booth, 1971;
Gerlach et Nirenberg 1982; Nelson et al., 1983).
1.2.1. Résultats
1.2.1.1. Caractérisation physique
1.2.1.1.1. Caractéristiques culturales
L'étude des caractéristiques culturales de 17 isolats de FOC isolés à partir de
la tige a porté sur l'aspect du mycélium aérien et la pigmentation du thalle et de
mycélium. Dans le cadre de ce travail, l'évolution phénotypique de quelques isolats
de FOC a été suivie dans le but de préciser la part des caractéristiques culturales dans
l'identification de cet agent fongique à partir des cultures jeunes et âgées. Les
conditions d'incubation étaient stables et similaires. Le tableau 18 résume les
morphotypes et les pigments rencontrés chez les isolats de FOC.
1.2.1.1.1.1. Description des morphotypes rencontrés
Notons que l'aspect général d'une colonie, est déterminé par la couleur de la
colonie et sa texture. Sur milieu PDA gélosé le thalle de la plupart des isolats de F.
oxysporum f. sp. ciceri était formé de mycélium dense, aérien et d’aspect plus ou
moins cotonneux. Après 5 à 7 jours de culture, le thalle de certaines souches vire de la
couleur blanchâtre au violet, en raison de la production des pigments.
L’étude des caractéristiques des isolats issue d’une culture monospore,
prélevée sous la loupe binoculaire après étalement d’une suspension convenablement
diluée sur de l’eau gélosée en boîte de Petri de Fusarium oxysporum a révélé une
variabilité morphotypique (Tableau 18).

Résultats et discussion
105
Tableau 18. Caractéristiques culturales des morphotypes sauvages de 17 isolats de FOC
Isolats Morphotypes Pigments
FOC1 Ras muqueux Violet
FOC2 Duveteux Blanc
FOC3 Duveteux Blanc
FOC4 Duveteux Blanc
FOC5 Cotonneux Blanc
FOC6 Ras muqueux Blanc
FOC7 Ras muqueux Violet
FOC8 Duveteux Violet
FOC9 Duveteux Blanc
FOC10 Ras muqueux Blanc
FOC11 Ras muqueux Blanc
FOC12 Ras muqueux Blanc
FOC13 Duveteux Blanc
FOC14 Ras muqueux Blanc
FOC15 Ras muqueux Blanc
FOC16 Cotonneux Blanc
FOC17 Duveteux Blanc
Dans les essais réalisés seuls les morphotypes mycéliens ont été rencontrés,
nous n'avons pas trouvé dans les cultures des d'organes de fructification tels que les
sporodochies et les pinnotes ou d'organe massifs tels que des sclérotes. Ainsi, 04 types
de thalles mycéliens ont été décrits dans l'ensemble des essais:
Figure 14. Morphotype du thalle de FOC ; A: Morphotype blanc cotonneux sauvage; B: Sous
morphotype bouclé; C: Sous morphotype en mèche.
A B C

Résultats et discussion
106
Type cotonneux: le mycélium aérien est très abondant épais et dense, sa couleur
blanche (ou violette) est, du fait de l'abondance du mycélium, beaucoup plus franche
que dans le type duveteux. Ce mycélium cotonneux est très peu sporifère et ne produit
que des microconidies, tardivement et en faible quantité.
Le nombre d'isolats portant ce type est: 2. Après vieillissement de la culture (après 10
mois de conservation et de repiquage successifs), ce morphotype cotonneux a présenté
deux sous types, le premier sous type bouclé (Figure 14B), le second, le mycélium se
réunit en mèches (Figure 14C).
Type duveteux: il est représenté par un mycélium aérien assez court, mais dense,
portant de très nombreuses microconidies, les chlamydospores et les macroconidies se
forment tardivement. Le mycélium et le milieu peuvent être coloré ou non. Le nombre
d'isolats portant ce type est: 7. Après vieillissement de la culture (10 mois de
conservation et de repiquage successifs), le morphotype duveteux a présenté un sous
type bouclé.
Morphotype ras muqueux: la rareté ou l'absence totale de mycélium aérien donne à la
culture un aspect muqueux comme si elle était envahie par des bactéries (Figure 16A
et 16B). Les microconidies sont très abondantes, les macroconidies rares, les
chlamydospores abondantes, mais tardives, le nombre des isolats portant ce type est 8.
Après vieillissement de la culture (10 mois de conservation et de repiquage
successifs), le morphotype muqueux présente un sous type d'aspect arbusculeux
(Figure 16C).
Figure 15. Morphotype du thalle de FOC:
morphotype duveteux sauvage

Résultats et discussion
107
Morphotype ras sénescent: l'aspect du thalle est à peu prés le même que celui du ras
muqueux, mais la croissance radiale est faible et le front de croissance irrégulier et
festonné (Figure 17A). La culture est en général très fortement colorée (Figure 17B),
ce type a été rencontré uniquement chez les cultures âgées.
1.2.1.1.1.2. Pigmentation du thalle
A travers l'ensemble de 17 FOC, nous avons pu révéler une pigmentation très
diversifiée sur le milieu PDA. Après 06 jours d'incubation, le mycélium présente des
pigmentations blanchâtres et violettes. La pigmentation chez les isolats sauvages est
blanche chez la plupart et elle représente un pourcentage de 82,35 %. Elle est violette
chez uniquement 03 isolats avec un pourcentage à environ 17.65% (Tableau 18).
Il semble que la morphologie du thalle et la pigmentation, chez cette forme
spéciale, ne caractérisent pas l'origine géographique des isolats puisque la plupart des
morphotypes et des pigments ont été rencontrés quelle que soit l'origine géographique
des isolats.
Figure 16. Morphotype du thalle de FOC ras muqueux : A: Morphotype ras muqueux coloré en violet;
B: Morphotype ras muqueux coloré en orange; C: Sous morphotype arbusculeux
Figure 17. Morphotype du thalle de FOC ras Sénescent, A: Morphotype
ras sénescent blanc; B: Morphotype ras sénescent rose
C B A
A B

Résultats et discussion
108
1.2.1.1.2. Caractéristiques microscopiques et biométriques des spores
1.2.1.1.2.1. Etude microscopique de FOC
Les observations au microscope photonique ont montré la présence d’un
mycélium cloisonné et ramifié, des conidiophores très courtes ainsi que des
macroconidies et des microconidies caractéristiques du genre Fusarium (Figure18 A
et 18B).
Les microconidies sont produite en fausse tète sur des monophialides chez
quelques isolats mais chez la plupart, le mycélium produit une seule microconidie par
phialide collé à ce dernier. Tous les isolats produisent les microconidies en
abondance sauf l'isolat FOC5 et FOC16. Des macroconidies épaisses sont dispersées
et leur nombre varie de moyen à très faible.
Nous avons observé également la présence des chlamydospores (Figure19),
produit par certains FOC après 15 jours de culture. Alors que d'autres isolats les
produisent après 21 jours ou même après un mois. Les chlamydospores sont
généralement formés en position terminale ou intercalaire. Ils sont menés d'une paroi
rigide. Elles sont arrondies, en ballonnet, avec une double paroi très caractéristique.
Aucune structure de sclérotes n'a été produite par les 17 isolats de FOC après 15 jours
de culture sur PDA.
Figure 18. Observation microscopique du FOC; A: Observation microscopique au bleu de Cotton sous
microscope optique x2000 montrant des monophialides portant des conidies regroupées en fausse; B:
Observation microscopique d'une culture sur lame coloré au bleu de Cotton montrant des microconidies et
des macroconidies x2000.
B A

Résultats et discussion
109
L'identification morphologique des isolats a confirmé la vérification de
Postula de Kock.
1.2.1.1.2.2. Etude biométrique
Les mensurations des conidies varient d'un isolat à un autre, la longueur des
microconidies varie de 5.55 à 8.44µm et la largeur entre 2.28 à 2.72µm. De même,
pour les macroconidies, la longueur varie de 18.2 à 29.64µm et la largeur entre 2.81 et
4.03µm. Le tableau 19 présente les résultats obtenus des mensurations pour chaque
isolat.
Tableau 19. Mensurations (µm) des macroconidies et des microconidies de 17 isolats de FOC
Isolats Microconidies Macroconidies Chlamydo
spores longueur Largeur Longueur Largeur
FOC1 6.97µm 2.36µm 24.68µm 3.75µm -
FOC2 8.44µm 2.6µm 18.97µm 3.29µm +
FOC3 5.82µm 2.35µm 23.75µm 3.76µm -
FOC4 6.07µm 2.31µm 25.2µm 3.38µm -
FOC5 8.35µm 2.41µm 23.36m 2.81µm -
FOC6 8.27µm 2.72µm 20.32µm 4.03µm +
FOC7 6.4µm 2.27µm 26.75µm 3.77µm -
FOC8 5.93µm 2.47µm 21.2µm 3.58µm +
FOC9 7.36µm 2.65µm 23.92µm 3.00µm -
FOC10 6.03µm 2.44µm 21.42µm 2.89µm +
Figure 19. Observation microscopique des
chlamydospores colorés au bleu de cotton sous
microscope optique x2000

Résultats et discussion
110
FOC11 5.55µm 2.4µm 29.64µm 3.56µm +
FOC12 6.5µm 2.38µm 25.1µm 3.68µm -
FOC13 5.68µm 2.28µm 23.2µm 3.55µm +
FOC14 6.06µm 2.44µm 23.15µm 3.24µm +
FOC15 6.94µm 2.38µm 18.2µm 3.6µm +
FOC16 6.5µm 2.32µm 20.72µm 3.13µm +
FOC17 6.52µm 2.36µm 25.12µm 3.77µm +
Moyenne 6.67µm 2.42µm 23.21µm 3.45µm
L'analyse statistique portant sur la longueur, ainsi que sur la largeur, des
microconidies ont montré qu'il n'y a pas des différences significatives entre les 17
isolats de FOC. Cette homogénéité a donné de corrélation de r=0.636 entre les
mensurations de la largeur et de la longueur des microconidies de 17 isolats à origine
géographique différente. Les résultats de l'analyse statistique des mensurations de la
longueur et la largeur des macroconidies montrent des différences significatives. Cette
étude montre donc que les mensurations en longueur et largeur des macroconidies
sont hétérogènes.
L'étude statistique nous a permis de regrouper les 17 isolats de FOC dans 04
groupes:
Groupe a: comprend les deux isolats FOC 15 et FOC2.
Groupe b: comprend les isolats: FOC6, FOC16, FOC8, FOC10, FOC14, FOC13 et
FOC5.
Groupe c: comprend FOC3, FOC9, FOC12, FOC1, FOC17.
Groupe d: comprend FOC4, FOC7, FOC11.
1.2.2. Discussion
1.2.2.1. Caractérisation physique
1.2.2.1.1. Caractéristiques culturales
L’étude d’un organisme pathogène implique de maîtriser son identification, de
plus, dans un souci d’exactitude, il faut pouvoir vérifier l’identité du champignon

Résultats et discussion
111
utilisé tout au long des expériences. L’identification correcte d’un pathogène est
également un élément clé dans le suivi des épidémies sur le terrain (Hatsh, 2004).
Les variations morphologiques sont portées sur des caractères culturaux, des
organes fructifères qui leur donnent éventuellement naissance (sporodochies et
pinnotes) et enfin sur la présence ou l'absence de sclérotes. Les morphotypes dits
sporodochial, sclérotial et pinnotal sont caractérisés par la présence d'organes massifs
respectivement sporodochies, sclérotes et pinnotes, lorsqu'ils sont absent, on définit
les morphotypes mycéliens du thalle qui sont constitué d'un mycélium dont l'aspect
est duveteux, cotonneux et ras muqueux (Henni et al., 1994).
Certains champignons phytopathogènes présentent en culture pure des
caractères typiques relativement stables qui permettent leur identification (Waite et
Stover, 1960). Certaines formes spéciales de Fusarium oxysporum ainsi que des races
peuvent être identifiées par l'aspect morphologique des cultures (Snyder et Smith,
1972; Awam et Lorbeer, 1986; Wong, 1988), c'est le cas de la forme spéciale
albedinis qui se distingue par son aspect arbustif et sporodochial stable (Djerbi et al.,
1985; Cherrab, 1989). La variabilité morphologique a été étudiée dans différentes
formes specialisées du Fusarium oxysporum f. sp. appi race2 (Awuah et Lorbeer,
1988), f. sp. capae (Abawi et Lorbeer, 1965;1971), f. sp. cubense (Waite et Stover,
1960; Follin et Laville, 1966) f. sp. lycopercisi (Wellman et Blaidell, 1941; Henni et
al., 1994) f. sp. melonis (Miller, 1945;1946), f. sp. lentis (Belabid, 2003), f. sp. ciceri
(Bekkar, 2007).
Etant donné que les champignons montrent rarement tous les aspects
morphologiques nécessaires à leur identification sous microscope, il est nécessaire de
les cultiver. La culture des champignons sur milieu de culture sert alors à induire des
phénotypes recherchés (Hatsh, 2004). Le PDA est principalement utilisé pour
l'identification des cultures de Fusarium car il induit la pigmentation (Nelson et al.,
1983; Summerell et al., 2003), cependant les cultures des champignons sur PDA ne
complète pas l'identification des espèces de Fusarium, à cause de ces concentrations
élevées en nutriment induisant ainsi des mutations morphologiques chez les isolats de

Résultats et discussion
112
Fusarium (Leslie et Summerell., 2006). Ces résultats sont en accord avec nos isolats
de FOC après vieillissement de ces dernières.
Les résultats obtenus lors de cette étude, montrent en général une variabilité
importante de la morphologie de FOC et l'observation des caractères morphologiques
de 17 FOC ont révélé une hétérogénéité au sein de la forme spéciale ciceri, ce qui
rapproche nos résultats à ceux décrit par Chabasse et al., (2002) et Haware, (1990) qui
indiquent que la croissance de FOC sur PDA à 25°C et à l'obscurité montre des
colonies qui sont duveteuses à cotonneuse, blanches à pourpres. Ces colonies
deviennent ridées en culture âgée. 04 morphotypes sauvages mycéliens différents ont
été rencontrés, tels qu'ils sont décrits par Waite et Stover (1960); Messiaen et Cassini
(1968); Henni et al., (1994). El Aoufir, (2001) n'a observé aucune formation de
sporodochies chez le FOC, ce qui est en accord avec nos résultats étant donné que nos
n'avons pas observé le morphotype sporodochial chez cette forme spéciale. Cette
structure a été rapportée chez le FOC par Bekkar, (2007). De même, pour les autres
formes spéciales cubense (Waite et Stover, 1960) et albedinis (Sedra, 1985; Cherrab,
1989) et lycopersici (Henni et al., 1994) et lentis (Belabid, 2003). La variabilité de la
morphologie mycélienne a été également démontrée chez d'autres espèces fongiques
telles que Colletoricum graminicola, agent de l'anthracnose du sorgho (Kaboré et al.,
2001).
Une importante variabilité se manifeste chez les isolats de FOC, les
morphotypes rencontrés en début d'expérimentation sont constitués en majorité de
morphotypes (cotonneux et duveteux), après vieillissement des cultures le
morphotype ras muqueux domine.
Quelques soit la morphologie d'origine de l'isolat, nous avons observé en
culture âgées, une tendance très nette à l'apparition ou au maintien du morphotype ras
muqueux. Ce résultat est similaire a celui signalé par Henni et al., (1994) en utilisant
la forme spécial lycopercisi.
Les isolements pratiqués à partir du matériel naturellement infectés produisent
le plus souvent des morphotypes duveteux et cotonneux que l'on peut considérer
comme représentant le phénotype sauvage (Abawi et Lorbeer, 1965;1971; Messian et

Résultats et discussion
113
Cassini, 1981; Nelson, 1981; Awuah et Lorbeer, 1988). Le maintien en culture de ces
isolats conduit en général à des variations se manifestant par la raréfaction du
mycélium aérien (évolution vers le type ras) (Wellman et Blaisdell, 1941; Abawi et
Lorbeer, 1965; 1971; Awuah et Lorbeer, 1988), ce qui a été observé après
vieillissement des cultures obtenues dans cette étude. Cependant le type sauvage de
départ peut dans la plupart du temps être maintenu en culture dépourvu de repiquages
soient fait à partir de microconidies prélevées dans des thalles jeunes (Follin et
Laville, 1966). Par ailleurs certains morphotypes sont considérés comme
particulièrement stables et notamment le morphotype ras (Wellman et Blaisdell, 1941;
Miller, 1946).
Les résultats obtenus lors de cette étude montrent une variabilité importante de
la pigmentation de FOC, ces résultats se rapprochent de ceux obtenus par Bekkar,
(2007) ; Dubey et al., (2009) sur les isolats de FOC et chez d'autres formes spéciales
comme Fusarium oxysporum f. sp. vasinfectum (Assigbetsé, 1989), Fusarium
oxysporum f. sp. lycopersici (Henni et al., 1994) et Fusarium oxysporum f. sp. lentis
(Belabid, 2003). La pigmentation est un critère clé dans la détermination des
Fusarium élaborée par Messiaen et Cassini (1968; 1981). Cependant, si elle constitue
un critère d'identification au niveau de certaines espèces, elle ne permet pas de
distinguer les formes spéciales de Fusarium oxysporum, car plusieurs d'entre elles
peuvent manifester une même coloration comme chez les formes lycopersici, lini,
pisi (Cherrab, 1989). Si au sein de ces dernières et des formes spéciales albidinis et
elaeides peu de variabilité a été rapportée, il apparait que ce ne soit pas le cas chez le
FOC puisque les différents isolats incubés sur un même milieu et dans les mêmes
conditions, ont manifesté une gamme de coloration, blanche, violette, rose, orange ce
qui concorde avec l'hypothèse définie par Messiaen et Cassini (1968; 1981) chez le
Fusarium oxysporum.
L'idée qu'il convient de tirer est que la morphologie des isolats étudiés de FOC
est très instable. La variabilité se manifeste aussi bien entre les différents isolats que
chez la descendance issus du même clone, dans les morphotypes et la pigmentation.
Ce résultat est en accord avec celui obtenu par Henni et al., (1994) sur la forme
spéciale lycopersici. ces variations portées sur des caractères culturaux (aspect du

Résultats et discussion
114
mycélium aérien, pigmentation du thalle) et des organes fructifères (sporodochies)
créent des difficultés de maintien en culture du phénotype sauvage des isolats de
FOC. Cette diversité reste jusqu'à ce jour sans explication. D'après, Jimenez-Gasco et
al., (2002) la diversité phénotypique observée chez le FOC, suggère leur possibilité
d'être polyphylétique, mais la réalité est que le FOC est monophylétique. Selon
Messiaen et Cassini, (1968), la pigmentation s'exprime en fonction des conditions du
milieu et sa disparition se produit fréquemment au cours du vieillissement de la
culture (Henni et al., 1994).
Taylor et al., (1999) ont rapporté que le FOC et comme tous les champignons
à reproduction asexuée ne subit pas une recombinaison régulière et la variation
génétique résulte principalement de l'accumulation des mutations. Par conséquent, le
génome entier est lié et se transmettre en une seule unité d'une génération à une autre.
La variabilité phénotypique serait due probablement à des phénomènes de mutations
(d'origine cytoplasmique ou d'origine nucléaire) (Burnett, 1984). Par la suite, la
redistribution de cette variabilité serait faite au sein des individus et des populations
par des mécanismes de reproduction asexuée et de compatibilité végétative. La
variabilité liée à des mécanismes de compatibilité végétative est bien connue chez les
champignons à reproduction asexuée (Fusarium oxysporum).
Henni et al., (1994) ont rapporté que la variabilité chez la descendance de
même clone se manifeste dans des cultures âgées et n'ont pas dans des cultures jeunes,
ce qui a été observé au cours de notre expérimentation. Selon le même auteur du point
de vue morphologique, la culture est en quelques sortes constituées d'une mosaïque de
cellules ayant des potentialités différentes et c'est une propriété non constitutive qui se
développe au cours du vieillissement du thalle. Le thalle issu d'une microconidie est
homogène et donne naissance à des microconidies ayant toutes les mêmes
potentialités au niveau morphologique. Mais au cours du vieillissement, la
différentiation affecte différemment les cellules de sorte qu'une hétérogénéité peut
être détectée chez les thalles âgés.
La variabilité des espèces de Fusarium de produire plusieurs caractères
morphologiques signifie que chez les espèces oxysporum, les caractères

Résultats et discussion
115
morphologiques traditionnelles épanchés dans les systèmes taxonomiques ne sont pas
suffisante pour établir une identification de cette espèce (Leslie et Summerell, 2006).
D’une manière générale, on peut se demander pourquoi les phytopathologistes
s’intéressent d’aussi près à des variations morphologiques qui se produisent en
cultures essentiellement à partir des thalles âgés. La réponse est à rechercher dans
l’existence possible de relations entre le morphotype d’un isolat et son pouvoir
pathogène, elle varie selon les auteurs et les organismes étudiés. D’ailleurs, selon
Djerbi et al., (1986), la forme spécialisée albedinis, agent du bayoud du palmier
dattier en Afrique du nord, présente un aspect morphologique caractéristique par
rapport aux autres formes spécialisées et par rapport aux F. oxysporum saprophytes du
sol. Ce parasitisme serait lié dans ce cas précis, à un type morphologique. Wellman et
Blaisdell, (1941) ont montré que le pouvoir pathogène des F. oxysporum f. sp.
lycopersici est lié à la morphologie: le morphotype duveteux serait très pathogène
alors que le pathotype ras muqueux serait peu pathogène.
1.2.2.2. Caractéristiques microscopiques et biométriques
1.2.2.2.1. Etude microscopique
Nos observations microscopiques montrent que l'ensemble des isolats de FOC
produisent 03 types de spores asexuées: des microconidies, des macroconidies et des
chlamydospores portés sur des monophialides courtes en fausse tète. Cette
observation est en accord avec la description faite par plusieurs auteurs pour cette
espèce (Chabasse et al., 2002; Leslie et Summerell, 2006; Nelson et al., 1983;
Haware, 1990). Les monophialides de Fusarium oxysporum sont plus courts en
comparaison avec ceux produite par Fusarium moniliforme et Fusarium solani
(Mart.) Appel et Wollenn. Emend. Synd. &Hans. (Burgess et al., 1989).
L'observation des microconidies a révélé qu'ils se naissent sur des
conidiophores simples et courts et qui apparaissent latéralement sur l'hyphe. Ils sont
nombreuses, plus petite dans la taille que les macroconidies, d'aspect ellipsoïdales ou
cylindrique, droite ou légèrement courbées et uninuclées correspondent au
observations faite par Chabasse et al., (2002); Henni et al., (1994); Booth, (1971);

Résultats et discussion
116
Nelson et al., (1983); Gurro et al., (1999); Nelson et al., (1994); Buxton, (1956); El-
Ani, (1968). Henni et al., (1994) ont signalé que les noyaux uninuclée des
microconidies observées chez le Fusarium oxysporum provient tous d'un unique
noyau au départ.
L'observation des macroconidies a montré qui ils sont moins nombreuses que
les microconidies, discrètement incurvées avec une cellule basale en forme de pied
distinctive, et une cellule apicale accrochée. Ils sont multinucliées et ont plusieurs
septales transversales environs 3 à 7 septations (Chabasse et al., 2002, Nelson et al.,
1983, Burgess et al., 1989, Booth, 1971; Dubey et al., 2009).
Nous avons observé aussi de nombreuses chlamydospores isolées ou en paires,
mais occasionnellement en chaines courts ou en touffe, avec des parois épaisse. Ce
résultat est similaire à celui apporté par plusieurs auteurs (Booth, 1971; Nelson et al.,
1983; Gurro et al., 1999; Nelson et al.,1994; Chabasse et al., 2002). Les
chlamydospores sont des conidies d'accessoires qui servent principalement comme
structure de survie dans l'environnement (Booth, 1971; Nelson et al., 1983; Gurro et
al., 1999; Nelson et al.,1994).
Un autre groupe de champignon filamenteux, les Coelomycetes à
reproduction asexuée en formant des conidies sur les bourgeons des fruits et ces
conidies sont similaires en morphologie au macroconidies produites par Fusarium,
par exemple: Botrycerea, Heteropatella, Cylindrocarpon et Pycnofusarium (Seifert,
2001). Cependant le genre Cylindrocarpon à des ressemblances très étroites avec le
genre Fusarium (Seifert et Gams, 2001). Il est très important de différencier les
macroconidies produites par ces champignons avec ceux du genre Fusarium, la
cellule basale de forme de pied des macroconidies de Fusarium est le caractère
morphologique le plus important pour séparer le genre Cylindrocarpon de Fusarium
(Seifert, 2001; Seifert et Gams, 2001).
1.2.2.2.2. Etude biométrique
Les résultats présentés dans le tableau19 montrent une homogénéité de la
longueur et de la largeur des microconidies de FOC. Ce résultat est en contradiction

Résultats et discussion
117
avec ceux obtenus par Abbas, (1995) en Syrie chez le Fusarium oxysporum f. sp.
lentis. Par contre, les mensurations en longueur et largeur des macroconidies sont
héterogènes. Des travaux sur les dimensions des conidies de Fusarium oxysporum f.
sp. melonis races 1, 2, et 3 présentent des variations importantes, même à l'intérieur de
cette forme spéciale ou à l'échelle d'une race (Messiaen et Cassini, 1968).
D'après les résultats obtenus nous avons trouvé que la taille des microconidies
varie de (5.55-8.44 x 2.28-2.78µm), ces résultats se rapprochent à d'autres obtenues
par plusieurs auteurs (Dubey et al., 2009; Haware, 1990; Chabasse et al., 2002). Les
tailles des macroconidies obtenues au cours de cette étude varient entre (18.2-29.64 x
2.81-4.03µm). Ce résultat se rapproche avec celui trouvé par Chabasse et al., (2002);
Dubey et al., (2009); Haware, (1990).
Les différences dans les résultats obtenus au cours de cette étude et autres
travaux faite par plusieurs auteurs sont probablement les conséquences de la variation
dans les conditions de cultures, d'incubation et de l'âge des cultures. Généralement,
les mensurations des macroconidies, des microconidies, des conidiophores et les
chlamydospores sont mesurés sur milieu CLA (Carnation leaf Agar). De même, pour
les conditions d'incubation, les cultures sont souvent maintenu en une température de
25°C jour/20°C la nuit et incubées sous lumière journalière (Nelson et al., 1983). Le
milieu CLA favorise la sporulation que la croissance mycélienne des cultures de
Fusarium. Ce milieu a gagné cette popularité pour les espèces de Fusarium après que
Fisher et al., (1982) ont montré que ce dernier est favorable à la croissance et préserve
les cultures de Fusarium pendant une longue période. La capacité des espèces de
Fusarium à sporuler mieux sur CLA est attribuée à sa composition pauvre en
carbohydrate et a la présence des feuilles d'œillet, qui fournit le même complexe
naturel des substances comme dans l'environnement naturel (Leslie et Summerell,
2006), ce qui n'a pas été respecté dans cette étude, étant donné que nous avons utilisé
le PDA.
L'identification biométrique a des inconvénients chez le genre Fusarium,
étant donné que les similarités et les différences observées des macroconidies des
espèces de Fusarium dépendent des conditions de croissance et peuvent être difficile

Résultats et discussion
118
à décerner (Thrane et Seifert, 2000; Summerell et al., 2003; Lesli et Summerell,
2006). En plus, la croissance dépend des caractères morphologiques, les isolats qui
devrait être regroupé comme des espèces différents peuvent appartenir à la même
espèce, cette male identification et cette hétérogénéité a été observé chez F.
oxysporum (Snyder et Hansen, 1940; 1941).
Le nombre restreint des macroconidies en culture est due principalement au
milieu utilisé qui est le PDA. En effet le milieu communément utilisé pour la
production de macroconidies est le CLA utiliser par plusieurs auteurs (Ciamp et al.,
2008).
1.2.1.2. Résultats de la caractérisation physiologique
1.2.1.2.1. Recherche des conditions optimales
Comme tout micro-organisme, la croissance fongique est dépendante d’un
certain nombre de paramètres intrinsèques et extrinsèques du milieu (Tubac, 2007).
Certains facteurs possèdent une très grande influence sur la croissance de l’espèce
fongique et principalement sur les possibilités de réalisation de l’interaction hôte –
parasite, parmi ces facteurs on distingue les facteurs biotiques (interactions avec
d'autres organismes) et les facteurs abiotiques (température, pH,…). Les facteurs
abiotiques les plus importants sont la température et l'humidité (Landa et al., 2007;
Farooq et al. ,2005; Magan et Lacey, 1984).
1.2.1.2.1.1. Estimation de la croissance mycélienne de 17 FOC sur PDA
Les résultats de la croissance mycélienne sur milieu PDA de 17 isolats de
FOC sont présentés par les courbes regroupées dans les figures (20, 21, 22). D'après
les résultats obtenus la vitesse de croissance mycélienne est variable.
Les résultats de l'étude de la variance de cette croissance ont montré qu'il
existe des différences hautement significatives et sont regrouper dans 04 groupes
homogènes:
Groupe a: FOC8, FOC15, FOC1, FOC5.
Groupe b: FOC 6, FOC 4, FOC 7, FOC 9, FOC 3.
Groupe c: FOC10, FOC12, FOC16, FOC 2.
Groupe d: FOC13, FOC 17, FOC 11, FOC14.

Résultats et discussion
119
La croissance mycélienne est comprise entre 9.93cm et 9.46cm chez 04
souches (groupe a), soit 23.52% de la population. Ce groupe est caractérisé par une
vitesse de croissance très rapide. La croissance est par contre, faible chez 04 souches
(groupe b), soit 23.52% de la population. Les 9 souches restantes constituent deux
groupes intermédiaires dont la croissance est comprise entre 8.74 et 7.12 cm.
Toutes les souches originaires de Tlemcen ont le diamètre de croissance le
plus élevé (03 souches dans le groupe a, 5 souches dans le groupe b et 2 souches dans
le groupe c), aucune souche appartenant à cette région n'a été classée dans le groupe d
(diamètre de croissance faible).
Par contre, 75% de souches de Mostaganem ont été classée dans le groupe d
(groupe de croissance faible). Les 03 souches d'Ain Témouchent ont des vitesses de
croissance variables allant de plus élevées (groupe a) au plus faibles (Groupe d).
1.2.1.2.1.2. Effet de la température
Les résultats de l’expérience sont représentés par les figures (23, 24, 25), le FOC se
développe aux températures comprises entre 10 et 30°C, la température optimale est de
30°C, les températures inférieures à 10°C ralentissent la croissance mycélienne. A la
température 35°C, la croissance est considérablement ralentie voir inhibée. En revanche,
l'exposition des isolats à des températures de 10 et 35°C, ne s'avère pas létale, car le
champignon reste vivant en formant des chlamydospores. Si on l'expose à nouveau à une
température optimale, celui-ci reprend sa croissance normale.
L'analyse de la variance de la croissance mycélienne, a montré qu'il ya des
différences hautement significatives entre les températures étudiées et le
comportement des isolats de FOC (annexe 3). Les températures sont classées selon
l'ordre suivant, de l'ambiante à la défavorable: 30°C>25°C>20°C >15°C>35°C>10°C.
Le test de Newman-Keuls, nous a permis de classer les isolats en 03 groupes distincts:
Groupe a: composé de FOC 10, FOC15, FOC1, FOC3 et FOC4.
Groupe b: composé de FOC6, FOC11, FOC 5, FOC7, FOC13, FOC2.
Groupe c: composé de FOC9, FOC8, FOC17, FOC14, FOC12, FOC16.

Résultats et discussion
120
1.2.1.2.1.3. Effet du pH
Les résultats de l’expérience sont représentés par les figures (26, 27, 28), le pH
optimal est de 5.5. L'analyse de la variance de la croissance mycélienne des isolats, a
montré qu'il a des différences hautement significatives entre les pH étudiés et les
isolats de FOC (annexe 3). Les pH sont classés selon l'ordre suivant, du propice au
défavorable à la croissance mycélienne des 17 FOC: 5.5 > 6.5 > 4.5 > 7.5.
Ainsi le test de Newman-Keuls nous a révélé la présence de 03 groupes
homogènes:
Groupe a: composé de FOC6, FOC3, FOC7, FOC5, FOC9.
Groupe b: composé de FOC8, FOC11, FOC15, FOC4, FOC10, FOC12, FOC16.
Groupe c: composé de FOC2, FOC1, FOC13, FOC14, FOC17.
1.2.1.2.1.4. Effet de la lumière et de l'obscurité
Les résultats dans nos essais sont exprimées dans les figures (29, 30, 31).
D'après ces résultats la croissance du champignon montre une indifférence à la
lumière et à l'obscurité. Toutefois, la lumière semble influencer le type de production
de spores. Ainsi, elle semble favoriser la production des macroconidies, alors que leur
production en obscurité est faible.
1.2.1.2.2. Influence des facteurs nutritionnels sur la croissance de FOC
Les champignons ont besoin pour assurer leur croissance de composés organiques
notamment des sources carbonées comme source d’énergie et des sources azotées pour la
synthèse des protéines. Pour rechercher les sources de carbone et d’azote donnant la
meilleure croissance de FOC, nous avons étudié l’effet de quelques sources de carbone et
d’azote.
1.2.1.2.2.1. Effet du milieu de culture
Les résultats rapportés dans les figures (38, 39, 40) font apparaitre des
différences du comportement de 17 isolats de FOC sur les différents milieux de
culture. Après 8 jours d'incubation, un maximum de croissance mycélienne est obtenu
sur le milieu pois chiche. Quant aux milieux Richard, Malt et Sach, ils se sont montrés

Résultats et discussion
121
défavorables à la croissance de FOC. Le test de Newman-Keuls, nous a permis de
classer les milieux de culture selon l'ordre suivant: Pois chiche>PDA>Waksman>
Czapeck >Malt>Sach>Richards.
L'analyse de la variance de la croissance mycélienne, a montré qu'il y a des
différences hautement significatives entre le comportement des isolats vis à vis les
milieux de cultures étudiés (annexe3). Concernant le classement des différents isolats,
le test de Newman-Keuls nous a permis de les regrouper en 03 groupes homogènes:
Groupe a: composé de FOC8, FOC15, FOC5, FOC1, FOC6.
Groupe b: composé de FOC4, FOC9, FOC7, FOC3, FOC16, FOC10, FOC12.
Groupe c: composé de FOC2, FOC13, FOC11, FOC17, FOC14.
1.2.1.2.2.2. Effet de la source de carbone
Les résultats de ce test sont exprimés dans les figures (32, 33, 34). L'analyse de la
variance a montré qu'il y a des différences hautement significatives du comportement des
isolats vis à vis les sources de carbones testés (annexe 3).
Le test de Newman-Keuls nous a permis de classer les sources de carbone selon
l'ordre suivant: Glucose > Galactose> Saccharose. Cette étude statistique nous a permis
aussi, de regrouper les isolats testés dans 03 groupes:
Groupe a: FOC11, FOC15, FOC3, FOC4, FOC8, FOC10, FOC1, FOC7.
Groupe b: FOC13, FOC6, FOC16, FOC9, FOC12, FOC17.
Groupe c:FOC2, FOC5, FOC14.
1.2.1.2.2.3. Effet de la source d'azote
Les résultats de ce test sont exprimés dans les figures (35, 36, 37). L'analyse de la
variance a montré qu'il y a des différences hautement significatives du comportement des
isolats étudiés vis à vis les sources de carbones testés (annexe 3). Le test de Newman-
Keuls nous a permis de classer les sources d'azote selon l'ordre suivant: peptone > NaNO3>
KNO3. Cette étude statistique nous a permis aussi de regrouper les isolats testés dans 03
groupes:

Résultats et discussion
122
Groupe a: FOC6, FOC1, FOC8, FOC3, FOC9.
Groupe b: FOC15, FOC5, FOC7, FOC11, FOC2, FOC4, FOC16, FOC13.
Groupe c: FOC12, FOC14, FOC10, FOC17.

Résultats et discussion
123
Figure 20. Estimation de la croissance mycélienne en cm sur le milieu de culture des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de
Fusarium oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC1 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC2 sur milieu de
culture PDA Croissance mycélienne en cm de FOC3 sur milieu de
culture PDA
Croissance mycélienne en cm de FOC4 sur milieu
de culture PDA
Croissance mycélienne en cm de FOC5 sur milieu
de culture PDA Croissance mycélienne en cm de FOC6 sur les7
milieu de culture PDA
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
124
Figure 21. Estimation de la croissance mycélienne en cm sur le milieu de culture des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de
Fusarium oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC7 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC8 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC9 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC10 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC11 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC12 sur milieu
PDA de culture
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
125
Figure 22. Estimation de la croissance mycélienne en cm sur le milieu de culture des isolats (FOC13, FOC14, FOC15, FOC16, FOC17) de
Fusarium oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC13 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC14 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC15 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC16 sur milieu
PDA de culture
Croissance mycélienne en cm de FOC17 sur milieu
PDA de culture
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
126
Croissance mycélienne en cm de FOC1 sur la gamme
de température
Croissance mycélienne en cm de FOC2sur la
gamme de température
Croissance mycélienne en cm de FOC3sur la
gamme de température
Croissance mycélienne en cm de FOC4 sur la gamme de
température
Croissance mycélienne en cm de FOC5sur la gamme de
température
Croissance mycélienne en cm de FOC6sur la gamme
de température
Figure 23. Influence de la température sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de Fusarium
oxysporum f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
127
Croissance mycélienne en cm de FOC7sur la gamme de
température
Croissance mycélienne en cm de FOC8 sur la
gamme de température
Croissance mycélienne en cm deFOC9sur la
gamme de température
Croissance mycélienne en cm de FOC10 sur la gamme
de température
Croissance mycélienne en cm de FOC11 sur la
gamme de température
Croissance mycélienne en cm de FOC12 sur la
gamme de température
Figure 24. Influence de la température sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de Fusarium
oxysporum f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
128
Croissance mycélienne en cm de FOC13 sur la
gamme de température
Croissance mycélienne en cm de FOC14 sur la
gamme de température
Croissance mycélienne en cm de FOC15 sur la
gamme de température
Croissance mycélienne en cm de FOC16 sur la gamme
de température
Croissance mycélienne en cm de FOC17 sur la
gamme de température
Figure 25. Influence de la température sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC16, FOC17, FOC18) de Fusarium
oxysporum f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
129
Croissance mycélienne en cm de FOC1 sur les 04
différents PH du milieu PDA. Croissance mycélienne en cm deFOC2 sur les différents
PH du milieu PDA.
Croissance mycélienne en cm de FOC3 sur les
différents PH du milieu PDA.
Croissance mycélienne en de FOC 5 sur les différents
PH du milieu PDA. Croissance mycélienne en cm de FOC6 sur les
différents PH du milieu PDA.
Croissance mycélienne en de FOC 4 sur les différents PH
du milieu PDA.
Figure 26. Influence du pH sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de Fusarium oxysporum f. sp.
ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
130
Croissance mycélienne de FOC8 sur les différents PH du
milieu PDA.
Croissance mycélienne de FOC10 sur les différents PH du
milieu PDA.
Croissance mycélienne en cm de FOC11 sur les
différents PH du milieu PDA. Croissance mycélienne en cm de FOC12 sur les
différents PH du milieu PDA.
Figure 27. Influence du pH sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de Fusarium oxysporum f. sp.
ciceri
Croissance mycélienne de FOC7 sur les différents PH
du milieu PDA.
Croissance mycélienne de FOC9 sur les différents PH
du milieu PDA.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
131
Croissance mycélienne de FOC13 sur les différents PH du
milieu PDA.
Croissance mycélienne de FOC14 sur les différents PH
du milieu PDA. Croissance mycélienne en cm de FOC15 sur les
différents PH du milieu PDA.
Croissance mycélienne de FOC16 sur les différents PH
du milieu PDA. Croissance mycélienne de FOC17 sur les différents PH
du milieu PDA.
Figure 28. Influence du pH sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC16, FOC17) de Fusarium oxysporum f. sp.
ciceri.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
132
Croissance mycélienne en cm de FOC1 sous lumière et
obscurité.
Croissance mycélienne en cm de FOC2 sous lumière
et Obscurité.
Croissance mycélienne en cm de FOC3 sous
Lumière et Obscurité.
Croissance mycélienne de FOC4 sous Lumière et
Obscurité.
Croissance mycélienne en cm de FOC5 sous Lumière
et Obscurité
Croissance mycélienne en cm de FOC6 sous Lumière
et Obscurité.
Figure 29. Influence de la lumière et de l'obscurité sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5 et FOC6) de Fusarium
oxysporum f. sp. ciceri.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
133
Croissance mycélienne de FOC7 sous Lumière et
Obscurité.
Croissance mycélienne en cm de FOC8 sous lumière
et Obscurité.
Croissance mycélienne de FOC9 sous Lumière et
Obscurité.
Croissance mycélienne de FOC10 sous Lumière et
Obscurité.
Croissance mycélienne de FOC11 sous Lumière et
Obscurité.
Croissance mycélienne de FOC12 sous Lumière et
Obscurité
Figure 30. Influence de la lumière et de l'obscurité sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11 et FOC12) de Fusarium
oxysporum f. sp. ciceri.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
134
Croissance mycélienne de FOC13 sous Lumière et
Obscurité.
Croissance mycélienne de FOC14 sous Lumière et
Obscurité.
Croissance mycélienne de FOC15 sous Lumière et
Obscurité.
Croissance mycélienne en cm de FOC16 sous
Lumière et Obscurité. Croissance mycélienne de FOC17 sous Lumière et
Obscurité.
Figure 31. Influence de la lumière et de l'obscurité sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC17) de Fusarium oxysporum f.
sp. ciceri.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
135
Croissance mycélienne en cm de FOC1 sur
différents source d'azote.
Croissance mycélienne en cm de FOC2 sur
différents source d'azote.
Croissance mycélienne en cm de FOC3 sur différents
source d'azote.
Croissance mycélienne en cm de FOC5 sur les
différentes sources d’azote.
Croissance mycélienne en cm de FOC6 sur
différents source d'azote.
Figure 32. Influence de la source d'azote sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de Fusarium
oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC4 sur les
différentes sources d’azote.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
136
Croissance mycélienne en cm de FOC 9 sur
différents source d'azotes.
Croissance mycélienne en cm de FOC 10 sur différents
sources d'azote.
Croissance mycélienne en cm de FOC11 sur différents
sources d'azote.
Croissance mycélienne de FOC12 sur différents source
d'azote.
Figure 33. Influence de la source d'azote sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de Fusarium
oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC 7 sur
différents source d'azotes.
Croissance mycélienne en cm de FOC 8 sur différents
source d'azotes.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
137
Croissance mycélienne de FOC13 sur différents
sources d'azote.
Croissance mycélienne en cm de FOC14 sur
différents sources d'azote.
Croissance mycélienne en cm de FOC15 sur
différents sources d'azotes.
Croissance mycélienne de FOC16 sur différents
source d'azote.
Croissance mycélienne en cm de FOC17 sur
différents sources d'azote.
Figure 34. Influence de la source d'azote sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC16, FOC17) de Fusarium oxysporum
f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
138
Croissance mycélienne de FOC1 sur différents source de
carbone.
Croissance mycélienne de FOC2 sur différents source
de carbone. Croissance mycélienne de FOC3 sur différents source
de carbone.
Croissance mycélienne de FOC4 sur différents source de
carbone.
Croissance mycélienne de FOC5 sur différents
source de carbone.
Croissance mycélienne de FOC6 sur différents
source de carbone.
Figure 35. Influence de la source de carbone sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de Fusarium
oxysporum f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
139
Croissance mycélienne de FOC7 sur différents source de
carbone.
Croissance mycélienne de FOC8 sur différents source
de carbone.
Croissance mycélienne de FOC9 sur différents source
de carbone.
Croissance mycélienne de FOC10 sur différents source de
carbone.
Croissance mycélienne de FOC11 sur différents source
de carbone.
Croissance mycélienne de FOC12 sur différents source
de carbone.
Figure 36. Influence de la source de carbone sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de Fusarium
oxysporum f. sp. ciceri C
roissan
ce mycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
140
Croissance mycélienne de FOC13 sur différents
source de carbone.
Croissance mycélienne de FOC14 sur différents
source de carbone.
Croissance mycélienne de FOC15 sur différents
source de carbone.
Croissance mycélienne de FOC16 sur différents
source de carbone.
Croissance mycélienne de FOC17 sur différents
source de carbone.
Figure 37. Influence de la source de carbone sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC16, FOC17) de Fusarium oxysporum
f. sp. ciceris
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
141
Croissance mycélienne en cm de FOC1 sur les
07différents milieux de culture testés.
Croissance mycélienne en cm de FOC3 sur les 07
différents milieux de culture testés.
Croissance mycélienne en cm de FOC2 sur les 07
différents milieux de culture testés.
Croissance mycélienne en cm de FOC4 sur les 07
différents milieux de culture testés.
Croissance mycélienne en cm de FOC5 sur les 07
différents milieux de culture testés.
Croissance mycélienne en cm de FOC6 sur les7
différents milieux de culture testés.
Figure 38. Influence des milieux de culture sur la croissance mycélienne des isolats (FOC1, FOC2, FOC3, FOC4, FOC5, FOC6) de Fusarium
oxysporum f. sp. ciceri
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
142
Figure 39. Influence des milieux de culture sur la croissance mycélienne des isolats (FOC7, FOC8, FOC9, FOC10, FOC11, FOC12) de Fusarium
oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC7 sur les 7
différents milieux de culture testés.
Croissance mycélienne de FOC8 sur les 7 différents
milieux de culture testés.
Croissance mycélienne en cm de FOC9 sur les 7
différents milieux de culture testés.
Croissance mycélienne en cm de FOC10 sur les 07
différents milieux de culture testés.
Croissance mycélienne en cm de FOC11 sur les 7
différents milieux de culture testés.
Croissance mycélienne en cm de FOC12 sur les 7
différents milieux de culture testés.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion
143
Croissance mycélienne en cm de FOC 13 sur les 7
différents milieux de culture testés.
Croissance mycélienne de FOC14 sur les 7
différents milieux de culture testés.
Croissance mycélienne en cm de FOC 15 sur les 7
différents milieux de culture testés.
Figure 40. Influence des milieux de culture sur la croissance mycélienne des isolats (FOC13, FOC14, FOC15, FOC16, FOC17) de Fusarium
oxysporum f. sp. ciceri
Croissance mycélienne en cm de FOC 16 sur les 7
différents milieux de culture testés.
Croissance mycélienne en cm de FOC 17 sur les 7
différents milieux de culture testés.
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm
Cro
issance m
ycélien
ne en
cm

Résultats et discussion Moyens de lutte
124
1.2.1.3. Discussion des résultats de la caractérisation physiologique
La première observation qui convient de faire à cette étude physiologique est
que le comportement in vitro de 17 isolats étudiés de FOC, est très variable et
instable. La variabilité et l'instabilité se manifeste aussi entre les isolats provenant
d'une même région. L'observation du comportement des 17 FOC étudiés aux
différents facteurs a révélé une hétérogénéité au sein de cette forme spéciale.
1.2.1.3.1. Recherche des conditions optimales
1.2.1.3.1.1. Estimation de la croissance mycélienne
A cause de la richesse du milieu PDA en carbohydrates, ce milieu favorise la
croissance mycélienne que la sporulation (Leslie et Summerell, 2006), c'est pour cette
raison qu'il est utilisé pour l'observation des taux de croissance qui est un caractère
secondaire important pour l'identification des espèces (Nelson et al., 1983; Summerell
et al., 2003).
D'après les résultats obtenus la croissance mycélienne des isolats est
hétérogène, elle varie d'un isolat à un autre. Cette vitesse de croissance semble être
dépendante de la région géographique. Des résultats similaires ont été rapporté par
Belabid, (2003) sur Fusarium oxysporum lentis.
1.2.1.3.1.2. Effet de la température
Nos résultats obtenus au laboratoire indiquent que la température optimale de
17 FOC testé est à environ 30°C. Ce résultat se rapproche beaucoup aux travaux de
plusieurs auteurs qui ont rapporté qu'en boite de Pétri et sur milieu gélosé, la
croissance mycélienne de FOC est optimale à un intervalle de 25°C à 30°C (Walker,
1965; Molot et Mas, 1975; Molot et al., 1990; Bekkar.,2007; Farooq et al.,2005). Ces
mêmes auteurs ont montré que ce champignon ne se développe pas aux températures
voisines de 4°C et 36°C, ce qui confirme nos résultats. Gupta et al., 1986 ont rapporté
des résultats similaires en ce qui concerne la température demandée par le FOC. La
relation de la température du sol a indiqué que la température adéquate pour le

Résultats et discussion Moyens de lutte
125
développement du flétrissement vasculaire chez le pois chiche se situe entre 25 à
30C° (Chauhan, 1965).
Plusieurs auteurs ont signalé que la croissance de FOC est influencé par la
température (Bhatti et Kraft, 1992; Navas-Cortes et al., 1998; Navas-Cortés et al.,
2000; Landa et al., 2001; Gupta et al., 1987; Westerlund et al., 1974), une
température au tour de 25°C est optimal pour le développement de ce dernier (Sharma
et Muehlbauer, 2007) et à cette intervalle la quantité maximale de la maladie peut être
atteinte malgré une petite densité d'inoculum initial du sol (Navàs-Cortès et al., 2000).
De même, Molot et Mas, 1975, ont rapportés que le pouvoir pathogène était stimulé
par des températures moyennes (22°C).
De plus, le pois chiche est une plante à climat intermédiaire, et le
développement de cette plante a besoin d’une température minimale de 10°C et une
température maximale de 35 °C, au delà de 35 et 10°C sa croissance est ralentie
(Verret, 1982; Girrard, 1985). L’apparition des températures extrêmes entraine des
dégâts irréversibles sur cette culture. Les températures supérieures à 30C°qui se
manifestent durant une période de 3 à 4 jours occasionnent de lourdes pertes du
rendement (Ben Mbarek et al., 2009). Ceci est en parfaite concordance avec les
températures de croissance des souches de FOC isolées. Un climat intermédiaire et
une température au tour de 25°C, souvent le cas pendant le printemps en Algérie,
favorise fortement le développement des plantes des cultures printanières du pois
chiche et l'installation de la maladie de la fusariose. Donc la sélection des variétés
tardives parait judicieuse, ou encore mieux la sélection des variétés résistantes au
froid et à l'anthracnose pour un semi d'hiver.
1.2.1.3.1.3. Effet du pH
Le second facteur abiotique étudié, l'effet du pH s'est révélé comme un facteur
de grande importance pour le développement du champignon. La croissance de FOC
a été obtenue sur tous les niveaux du pH testés, mais elle est maximale sur le PH 5.5.
Ce résultat est en contradiction avec celui signalé par Farooq et al., (2005) qui a
trouvé un pH optimum à environ 7. Les résultats de cette présente étude est en accord

Résultats et discussion Moyens de lutte
126
avec autres faite par Hayes, (1978). Shaikh, (1974) a signalé que le FOC peut tolérer
une gamme du PH entre 5-6.5.
En (1997), Keller a étudié l’effet du pH sur la croissance de Fusarium
proliferatum, la croissance du champignon était maximale à un PH=5,6.
1.2.1.3.1.4. Effet de l'action de la lumière et de l'obscurité
Contrairement à certaines espèces très exigeantes en lumière pour la
sporulation (Leach, 1962), tous les résultats obtenus montrent que le FOC peut
pousser indifféremment en présence ou en absence de la lumière. La lumière favorise
la maturation des conidies et la germination des spores (Pfohl-Leszkowicz, 2001).
Carlile en (1956) a démontré que la lumière favorise la formation d'un pigment orange
et les microconidies. Cependant l'obscurité favorise la production des pigments rouges
et violettes et les chlamydospores chez les cultures âgées.
1.2.1.3.2. Influence des facteurs nutritionnels sur la croissance de FOC
1.2.1.3.2.1. Effet des milieux de culture
Les champignons sont des microorganismes ubiquistes qui peuvent se
développer sur une grande variété de substrats (Tabac, 2007). Des milieux de cultures
différentes synthétiques et non synthétiques, ont une influence profonde sur les
caractéristiques culturales et morphologiques des champignons (Shaikh, 1974).
Le premier facteur étudié est le milieu de culture, d'après nos résultats le
meilleur milieu de culture est le milieu pois chiche. Des résultats de l'expérimentation
faite par Farooq et al., (2005) ont révélé que le milieu solide Czapeck dox et CSMA
sont les meilleurs pour la croissance radiale de FOC. Khanzada et al., (2003) ont
aussi rapporté des résultats similaires avec M. phaseolina. Bekkar, (2007) a observé
que le milieu le plus favorable pour la croissance de FOC est le PDA alors que, le
milieu Richard semble être le moins favorable. Haware et al., 1986 ont modifié le
milieu Cape Do Agar en ajoutant PCNB, la streptomycine et le vert de malachite, ce
milieu est hautement effectif pour la croissance de Fusarium oxysporum.

Résultats et discussion Moyens de lutte
127
1.2.1.3.2.2. Effet de la source de carbone
D'après les résultats, le glucose semble être la meilleure source de carbone.
Des résultats faits par Farooq et al., (2005) ont montré que toutes les sources de
carbones testés (glucose, amidon, saccharose) sont convenables pour la croissance du
champignon.
Cependant le glucose semble être la meilleure pour la croissance du
champignon. Le FOC peut convertir certains forme de composé de carbone complexe
vers une forme simple, qui peut être prêt a être métabolisé (Bais et al., 1970).
1.2.1.3.2.3. Efffet de la source d'azote
La peptone était trouvée la meilleur source d'azote pour Fusarium oxysporum
f. sp. ciceri. Farooq et al., (2005) ont obtenu un résultat similaire. Tariq et al., (2003)
ont obtenus une croissance maximale de Botrytis glaiolorum lorsqu'ils ont utilisé le
glucose et le nitrate de potassium. Farooq et al., (2005) ont indiqué que le rôle du
ration C:N est très important. Le FOC colonise les substances organiques dans le sol.
L'augmentation de l'inoculum potentielle et la sévérité de la maladie corrèle
positivement avec la base d'alimentation des substances organiques. Les débris
végétaux servent comme base d'alimentation et peuvent aussi servir comme un pont
d'infection.
2. Recherche de moyens de lutte biologique
2.1. Isolement et étude de la diversité des actinomycètes
2.1.1. Résultats
2.1.1.1. Isolement des actinomycètes
Dans cette étude, les actinomycètes ont été isolés à partir de 10 échantillons
des sols provenant des régions arides du sud algérien.

Résultats et discussion Moyens de lutte
128
Tableau 20. Nombre des souches d'actinomycètes isolés sur milieu ISP2.
Les souches obtenues ont été isolé sur milieu ISP2 sans antibiotique
(Yamamura et al., 2007; Shirlling et Gottlieb, 1966). Après 7 jours d'incubation à
30°C Les actinomycètes apparaissent et se développent lentement. Elles sont repérées
d'après leur aspect macroscopique caractéristique, puis purifiées afin d'obtenir des
cultures pures pour la conservation. L'isolement des actinomycètes à partir des
échantillons de sols arides sur milieu ISP2 a conduit aux résultats rapportés dans le
Tableau 20. Les résultats présentés font apparaitre que le nombre total de colonies
d'actinomycètes isolés est de 61 souches dont, le plus grand nombre d'actinomycètes a
été isolé à partir de l’échantillon de sol provenant de la région de Knadssa " sol S4 ".
Le tableau 21 résume les pourcentages de l'isolement des actinomycètes à
partir des deux dilutions effectuées 10-3
et 10-4
.
Echantillons Lieu de
prélèvement
Nombre de souches
d'actinomycètes
isolées
Nombre
d'actinomycètes à
activité antifongique
1 Wakda " S11 " 02 00
2 Wakda " S10 " 05 00
3 Knadssa " S4 " 14 09
4 Knadssa " S5 " 02 01
5 Knadssa " S6 " 19 06
6 Knadssa " S7 " 02 00
7 Knadssa " S8 " 01 00
8 Béni-Abbés "S1" 08 02
9 Kali "S2 " 03 00
10 Massine " S3 " 05 00
Total 61 18

Résultats et discussion Moyens de lutte
129
Tableau 21. Pourcentages d'isolement des actinomycètes à partir des deux dilutions effectuées
Echantillons Régions Dilutions Nombre de
souches isolées
Total
1 Wakda" S11" 10-3
02 02
10-4
00
2 Wakda "S10" 10-3
04 05
10-4
01
3 Knadssa "S4" 10-3
12 14
10-4
02
4 Knadssa "S5" 10-3
02 02
10-4
00
5 Knadssa "S6" 10-3
11 19
10-4
08
6 Knadssa "S7" 10-3
02 02
10-4
00
7 Knadssa "S8" 10-3
01 01
10-4
00
8 Béni-Abbès
"S1"
10-3
08 08
10-4
00
9 Kali "S2" 10-3
02 03
10-4
01
10 Massine "S3" 10-3
04 05
10-4
01
D'après les résultats du tableau 21, le plus grand nombre des souches a été
isolé à partir de la dilution 10-3
environ 78.68 % des souches. Le reste des souches a
été isolé à partir de la dilution 10-4
.

Résultats et discussion Moyens de lutte
130
2.1.1.2. Caractéristiques des actinomycètes isolés
Les souches d'actinomycètes isolées présentent des caractéristiques
macroscopiques différentes: couleur des colonies, couleur du dos de la boite et aspect
des colonies (Figure 41). Ce résultat indique que les sols arides du sud algérien, bien
qu'étant sous l'influence d'un climat sec et aride, sont loin d'être des sols pauvres en
microorganismes et sont une source riche en actinomycète quantitativement et
qualitativement.
2.1.2. Discussion
Le résultat obtenu par cette étude qui montre que le plus grand nombre
d'actinomycètes a été isolé à partir de la dilution 10-3
est en accord avec celui obtenu
par Kitouni, (2007) lors de ces travaux sur la diversité des actinomycètes isolés à
partir des sols arides du sud algérien.
Figure 41. Quelques aspects des actinomycètes isolés à partir des sols arides du sud algérien
(Bechar et Timimoune)

Résultats et discussion Moyens de lutte
131
Des travaux de recherches ont été réalisés à partir du sol de Béni Abbès, prise
comme modèle d'étude, mais aussi, au niveau de plusieurs autres régions du sud
algérien ont montré la diversité et la richesse de cette région en actinomycètes
antagonistes, ces dernières sont stimulées par la rhizosphère du palmier dattier
(Sabaou et al., 1998). De même, Khamna et al., (2009) ont rapporté qu'il est possible
que les exsudats des plantes améliorent la croissance des Streptomyces sp., qui à leur
tour synthétisent des composés antimicrobiennes. Germida et al., (1998) ont rapporté
que la diversité d’une communauté microbienne dans un environnement spécifique
dépend des espèces des plantes trouvées.
D’autres facteurs importants ont été également cités par d’autres chercheurs
contrôlant ainsi l'abondance et l'activité des actinomycètes dans un sol. Parmi ces
facteurs, la disponibilité des éléments nutritifs, la nature et l'abondance de la matière
organique, la salinité, la composition en humidité relative, la température, le pH et la
végétation du sol (Goodfellow et Williams, 1983; McCarthy et Williams, 1990). Xu et
al., (1996) ont trouvé que la population des actinomycètes présents dans les régions
froides de la Chine est constituée de plus de 83% du nombre total de la microflore du
sol. Le nombre des actinomycètes a été relié au type du sol et aux conditions
édaphiques (Barakate et al., 2002). En effet, Crawford et al., (1993) ont rapporté que
les sols rhizosphériques présentent une population d’actinomycètes supérieure deux à
trois fois que celle des sols non rhizosphérique.
Cette diversité a été rapportée par plusieurs auteurs qui ont travaillé sur cette
région (Sabaou et al., 1980, 1992 et 1998; Kitouni, 2007; Zitouni et al., 2005).
D’ailleurs, Zitouni et al., (2005) ont rapporté que les sols de sud algérien sont exposés
a un climat aride et représente de ce fait un écosystème particulier. Ce type
d’écosystème peut représenter un réservoir important en microflore d’intérêt
biotechnologique. Sabaou et al., (1998) ont rapporté que les sols sahariens d’Algérie
constituent un écosystème assez particulier en raison de l'environnement aride et
mériteraient d'être exploités dans le domaine de la recherche des micro-organismes
capables d’être utilisé comme agents de lutte biologique contre les maladies fongiques

Résultats et discussion Moyens de lutte
132
des plantes, notamment contre l'agent responsable de la fusariose du palmier dattier
(Sabaou et Bounaga, 1987).
Des genres d’actinomycètes parfois assez rares dans le monde, ont été isolés
en nombre important et dont plusieurs sont producteurs d'antifongiques à large spectre
d'action (Boudjella, 1994; Zitouni, 1995). D’après, cette contribution ainsi que les
travaux de certains équipes algériennes, Il semble que l'Algérie présente beaucoup
d'environnement non exploités, comme la rhizosphère des plantes endémiques, les
sols de déserts, les sols des montagnes, l'eau de mer et l'eau de la sebkha.
2.2. Etude du phénomène d'antagonisme FOC/Actinomycètes
2.2.1. Résultats
2.2.1.1. Test de confrontation directe
L'ensemble de la collection d'actinomycètes a été testé afin de mettre en
évidence l'activité antifongique des souches isolées vis-à-vis de FOC, par la technique
de confrontation directe, cette technique est une méthode de diffusion en milieu
gélosé et elle a été choisie pour une première sélection car elle est plus simple et
rapide que le test de cylindre d'agar ou autres. Elle nous a permis de détecter l'effet
inhibiteur de toute la collection d'actinomycètes obtenue (61 souches) vis-à-vis des 03
isolats de FOC représentatives de 03 régions qui sont Tlemcen (FOC8), Ain
Temouchent (FOC15) et Mostaganem (FOC13).
Les souches d'actinomycètes qui n'ont pas présentées une activité antifongique
vis-à-vis le FOC et dont le pourcentage d'inhibition est inferieur à 20% ont été
éliminées. Les autres ont été retenues pour calculer le taux d'inhibition. Seulement 06
souches d'actinomycètes ont présenté un pourcentage d'inhibition non négligeable.
Ces dernières ont été retenues pour des tests ultérieurs. Il en ressort des résultats de
tableau 20 que sur les 61 isolats d'actinomycètes, seules 18 actinomycètes soit 29.50%
sont actives sur au moins un des isolats de FOC étudié. 70.5% des actinomycètes
isolés n'ont montré aucunes activités antifongiques vis-à-vis les 03 isolats de FOC en
utilisant le milieu gélosé PDA comme milieu de production.

Résultats et discussion Moyens de lutte
133
Tableau 22. Pourcentage d'inhibition des 06 isolats d'actinomycètes sur les 03 FOC
Souche Région Pourcentage d'inhibition en % Moy.
FOC15 FOC13 FOC8
ACT1 Knadssa "S4" 32.4% 37.5% 22.95% 30.95%
ACT2 Béni Abbés "S1" 46.66% 58% 21% 41.88%
ACT3 Knadssa "S4" 24% 56% 15.55% 31.85%
ACT4 Knadssa "S4" 33.33% 20% 11.11% 21.48%
ACT5 Knadssa "S4" 35.2% 20% 30.37% 28.52%
ACT6 Knadssa "S4" 58.1% 19.2% 18% 31.76%
D'après les résultats du tableau 22, la souche ACT6 isolée du sol de Knadssa
"S4" a montré une activité contre un seul isolat de FOC (FOC15). Les souches ACT3
et ACT4 isolées du sol de Knadssa S4 inhibent seulement 02 FOC (FOC13 et
FOC15).
Par contre les 03 souches ACT1 et ACT5 isolées du sol de Knadssa S4 et la
souche ACT2 isolée à partir du sol de Béni-Abbès S1 inhibent les 03 isolats de FOC
testés. La moyenne la plus grande du taux d'inhibition a été produite par la souche
ACT1 (58.1%). Cependant la moyenne la plus faible a été produite par la souche
ACT4 (11.11%).
Figure 42. Pourcentages d'inhibition par série de couleurs d'actinomycètes.
40%
30%
20%
10%
00%

Résultats et discussion Moyens de lutte
134
Les données présentées par la figure 42 montrent aussi que les pourcentages
d'inhibition par série de couleur sont presque égale, il est de 31.35% pour la série des
blanches, de 26.66% pour la série des grises et de 35.2% pour la série des beiges. Les
isolats qui ont des taux d'inhibition élevés sont les séries de beige suivi par la série des
blanches, ensuite la série des grises. En effet nous avons remarqué que les souches de
la même série présentent des pourcentages d'inhibition très différents
L'activité antagoniste des actinomycètes testés vis-à-vis des 03 isolats de FOC
est différente, nous avons observé une inhibition des 02 isolats de FOC provenant de
Mostaganem et de Ain Témouchent (FOC13 et FOC15), par contre, nous avons
observé une tolérance de l'isolat FOC8 provenant de la région de Tlemcen.
2.2.1.2. Test des filtrats
L'implication des composés antifongiques produits par les actinomycètes dans
l'inhibition in vitro de la croissance mycélienne de FOC (Fusarium oxysporum f. sp.
ciceri) a été confirmée ensuite par l'action des filtrats de culture et la technique des
puits. Les deux techniques nous ont permis de détecter l'effet inhibiteur des souches
testées vis-à-vis des 03 isolats de Fusarium oxysporum f. sp. ciceri utilisés.
Les 06 souches présentant les meilleurs taux d'inhibition, ont été ensuite
soumises au test de filtrat. Cette technique a été introduite dans ce screening afin de
rechercher l'action des filtrats des souches pré-sélectionnées sur l'inhibition des
isolats de FOC par les antifongiques élaborés par les actinomycètes du sol.
L'incorporation des filtrats dans le milieu PDA a pour but d'essayer de distinguer entre
les filtrats actifs et non actifs.
FOC13
ACT5
ACT6 FOC15 FOC8 ACT2
Figure 43. Résultats du test de confrontation entre les isolats de FOC et les souches d'actinomycètes :
A: FOC13/ACT5; B: FOC15/ACT6; C: FOC8/ACT2.
A B C

Résultats et discussion Moyens de lutte
135
Tableau 23. Diamètre d'inhibition de FOC par les filtrats des 06 actinomycètes sélectionnés
Diamètre en mm
du témoin
Diamètres en
mm de FOC15
Diamètres en mm
de FOC13
Diamètres en mm
de FOC8
5 5.1 6.2
ACT1 2.5 3.7 4.5
ACT2 2.8 3.5 4.1
ACT3 2.8 4 4.3
ACT4 2.8 4.2 4.3
ACT5 3.2 4.2 4.4
ACT6 2.4 4.3 3.9
Le tableau 23 montre que tout les filtrats inhibent la croissance des conidies
des 03 isolats de FOC ce qui n'est pas le cas pour le premier test. Les diamètres
varient de 2.4mm à 3.2mm pour FOC15 et de 3.5mm à 4.3mm pour FOC13 et de 3.9
mm à 4.5 mm pour FOC8 (figure 44).
L'ensemencement d'un inoculum d'un isolat de FOC inhibé par les filtrats de
culture des actinomycètes dans une autre boite contenant du PDA stérile, a montré
que l'isolat s'est développé normalement. Ce test nous a indiqué que la substance
diffusible a une action fongistatique.
ACT5 FOC13 FOC15 ACT4 ACT1 FOC8
Figure 44. Résultats du test de filtrat entre les isolats de FOC et les souches d'actinomycètes : A:
FOC13/ACT5; B: FOC15/ACT4; C: FOC8/ACT1.
A B C

Résultats et discussion Moyens de lutte
136
2.2.1.3. Test de puits
Cette technique est une méthode de diffusion en milieu gélosé. Les résultats
de l'inhibition sont regroupés dans la figure 46. L'emploi du test des puits n'ayant pas
donné une entière satisfaction, sauf quelques inhibitions après de dizaines de
répétions. Il a été nécessaire d'introduire une autre technique basée sur l'étude de
l'activité de l'extrait obtenue par la souche ACT2.
2.2.1.4. Test des disques
L'étude de l'extrait préparé après culture sur milieu liquide ISP2 de la souche
d'actinomycète ACT2 s'est révélée fiable. Il est préférable, en ce qui concerne
l'analyse des extraits et des surnageant, d'enregistré le pH et l'effet du solvant utilisé
afin d'éliminer les interférences dues au pH et au solvant. Les résultats sont présentés
par la figure 46.
L'extrait de filtrat inhibe les 03 isolats de FOC. Le diamètre d'inhibition varie
du 1.1cm pour l'isolat FOC13, de 1.2cm pour l'isolat FOC15 et de 0.9cm pour l'isolat
FOC8.
ACT3 FOC13 FOC15 ACT1 ACT6 FOC8
Figure 45. Résultats du test des puits entre les isolats de FOC et les 06 souches d'actinomycètes : A: FOC13/ACT1; B:
FOC15/ACT2; C: FOC8/ACT3.
A B C

Résultats et discussion
45
2.2.2. Discussion
2.2.2.1. Test de confrontation
Le pourcentage des souches d'actinomycètes inhibant le champignon est
confirmé par plusieurs auteurs qui ont trouvées des pourcentages presque égale a
celui cité par cette étude (Saadoun et Al-Momani, 1997; Ndonde et Semu, 2000;
Barakate et al., 2002), cependant autres auteurs ont trouvé des pourcentages moins
élevées (Rovasz, 1986; Saadoun et al., 1998). Les souches d'actinomycètes qui n'ont
pas montré une activité vis-à-vis le FOC, il est probable qu'elles produisent d'autres
composés notamment antibactériennes contre d'autres microorganismes non testés
(Barakate et al., 2002). En effet, Porter, (1971) a signalé que probablement tout les
Streptomyces possèdent des propriétés antibactérienne si leur conditions sont prise en
considération durant la culture des ces microorganismes. Dans la plupart des cas la
capacité antagoniste des bactéries utilisées dans la lutte est déterminé juste contre un
ou quelques isolats du pathogène (Landa et al., 1997).
Les pourcentages d'inhibition trouvé sur milieu PDA sont élevés, ce qui rend
le milieu PDA un milieu favorable pour la production et la diffusion d'antifongique
produit (Jayasinghe et Parkinson, 2008).
Les Streptomyces appartenant à la même série de couleur peuvent être de
même espèce mais produisent des molécules bioactives différentes ou bien sont des
espèces différentes (Barakate et al., 2002).
Dans cette présente étude les différences dans la réponse des isolats de FOC
aux produits élaborés par les actinomycètes sont similaires à ceux trouvé par Landa et
Figure 46. Résultats du test des disques imbibés entre les 03 isolats de FOC et la souche ACT2 d'actinomycète :
A: ACT2/FOC13; B: ACT2/FOC15; C: ACT2/FOC8
FOC8 FOC15 FOC13
ACT2 ACT2 ACT2
C
B
A

Résultats et discussion
46
al., (1997) en utilisant des bactéries Pseudomonas et des Bacillus et ils ont expliqué
ce résultat par la différence dans les races de FOC.
La tolérance de l'isolat de Tlemcen (FOC8) aux antifongiques des 06
actinomycètes testés semble être la cause de la virulence élevée de cet isolat. Des
résultats similaires ont été obtenus par Dubey et al., (2007) en utilisant Trichoderma
sp. contre le FOC race 5, cette race est la plus virulente de FOC et résiste à la plupart
des antifongiques (Hervas et al., 1995) ou peut être à cause de la croissance rapide de
cette isolat. Jayasinghe et Parkinson en 2008 ont signalé que les genres du
champignon qui ont une croissance rapide sont plus tolérants aux actinomycètes que
les lentes. Les actinomycètes sont généralement à croissance lente et les champignons
à croissance rapide peuvent bloquer l'inhibition (Jayasinghe et Parkinson, 2008). En
plus les champignons à croissance rapide et qui sont antagonistes développent une
résistance élevée pour se protégé de l'autolyse et cela peu expliquer leur résistance.
Les métabolites de différentes espèces d'actinomycètes ont été rapportés d'être
active contre le Fusarium spp. et autres phytopathogènes (Kavitha et al., 2010). Des
résultats in vitro ont indiqué que les actinomycètes inhibent la croissance du
champignon en utilisant des composés antifongiques ou par l'hyper parasitisme
(Jayasinghe et Parkinson, 2008) ou par la production de siderophores qui sont
impliqué dans la suppression de plusieurs formae speciales de F. oxysporum (Scher et
Baker, 1980;1982; Sneh et al., 1984; Elad et Baker, 1985; Kumar et Dube, 1992;
Duijff et al., 1993; Kumar,1990; 1996; 1998), ou à travers la production des enzymes
qui dégradent la paroi cellulaire du champignon (El-tarabily et al., 2000; Errakhi et
al., 2007; Getha et al., 2005; Goodfellow et Williams, 1983).
Ces résultats indiquent que peut être une production d'un antifongique seul est
impliquée dans l'inhibition de la croissance mycélienne de FOC ou peut être un des
mécanismes contrôlant ce pathogène, en se basant sur les arguments suivant:
- Pour les 06 souches sélectionnées aucun contact direct n'a été établi entre le
mycélium fongique et les colonies bactériennes des actinomycètes, donc

Résultats et discussion
47
l'inhibition de la croissance mycélienne de FOC a été à travers une substance
qui diffuse dans l'agar du milieu.
- Le milieu PDA utilisé dans ce test est riche en nutriment et une compétition
des nutriments est à exclure.
- L'antibiose est un mode très connus d'antagonisme observé chez les
Streptomyces.
Si on affirme suite aux arguments cités précédemment que les souches testées
cultivées sur PDA libèrent un métabolite extracellulaire qui inhibent la croissance des
03 isolats de FOC, le potentiel antagoniste, implique la production des composés
antifongiques, cette production d'antifongique a été rapporté par plusieurs auteurs
(Crawford et al., 1993; Ouhdouche et al., 2001). Yuan et Crawford , (1995) ont
rapporté l'activité antifongique in vitro de Streptomyces lydicus contre Pythium
ultimum et Rhizoctonia solani, Anitha et Rebeeth, (2009) ont rapporté l'inhibition des
pathogène: Fusarium oxysporum, Fusarium solani, et Rhizoctonia solani et Alternaria
alternata in vitro par Streptomyces griseus. Les Streptomyces inhibent le Fusarium
sp. (F. oxysporum et F. solani) (Sacramento et al., 2004). Paul et Banerjee, (1986) ont
montré que les antibiotiques solubles produits par Streptomyces galbus peuvent
inhiber la germination de Alternaria solani, Aspergillus niger, Curvularia pallescens,
Helminthosporium oryzae. De plus, la croissance mycélienne de C. eragrostides et C.
gloeosporioides a été aussi inhibé par les cultures d'actinomycètes lorsqu'elles se
développent en parallèle sur PDA (Soares et al., 2006). Autres résultats présentés par
Kathiresan et al., (2005) ont indiqué l'efficacité des souches d'actinomycètes marine
in vitro notamment du genre Streptomyces sur l'inhibition de Pyricularis oryzae qui
attaque le Riz. De même Landa et al., 1997 ont montré qu'environs 32% de bactéries
isolées à partir de la rhizosphère du pois chiche inhibent in vitro la croissance de
FOC.
Il existe plusieurs hypothèses qui peuvent expliquer la production
d'antibiotique, mais la plus acceptée est que la molécule antimicrobienne aide
l'organisme à la compétition avec d'autres vis à vis des nutriments dans le sol en
réduisant la compétition (Aghighi et al., 2004).

Résultats et discussion
48
2.2.2.2. Test des filtrats de culture
Les filtrats inhibent la croissance des conidies des 03 isolats de FOC ce qui
n'est pas le cas pour le premier test de confrontation. Cela peut être due à la
composition du milieu utilisé pour l'extraction qui est le ISP2 pour ce test qui a peut
être favorisé la production de métabolite.
les résultats de l'inhibition de FOC par les filtrats de culture sont similaires à
ceux trouvé par Landa et al., (1997) en utilisant des Pseudomonas et des Bacillus. Ces
résultats sont aussi en accord avec d'autres (Yang et al., 2007), qui ont étudié la
capacité in vitro des filtrats de culture de Penicillium oxalicum souche PY- dans le
contrôle de l'infection par Scertinia sclertiorum sur les plantes de Colza, et Khamna et
al., (2009) qui ont étudié la capacité in vitro des filtrats de Streptomyces sp. CMU-PA
dans le contrôle de l'infection par Alternaria porri.
D'après les résultats obtenus du test de filtrat et des puits les 06 souches
d'actinomycètes produisent des molécules bioactives sur le milieu ISP2, qui diffère du
PDA dans la composition en nutriment (PDA milieu naturelle/ ISP2 milieu
synthétique) et la contenance en agar (PDA gélosé, ISP2 liquide). Les résultats de ce
test montrent des différences dans l'activité par rapport au test de confrontation, cela
peut être à cause de la différence dans la composition et la période d'incubation et la
densité d'inoculum et la technique. Des résultats obtenus par plusieurs auteurs ont
indiqué que la composition en nutriments a un effet sur les activités des
microorganismes à cribler (Sivasithamparam et Parker, 1980; Whipps, 1987). Singh et
al., 2006 ont indiqué que quelques souches d'actinomycètes n'ont aucune activité
lorsqu'elles sont transférées du milieu gélosé vers un milieu liquide.
2.2.2.3. Test des disques
Le résultat du test de l'activité de l'extrait brut est similaire avec celui trouvé
par Khamna et al en (2009) qui ont rapporté que l'extrait brut de Streptomyces sp.
CMU-PA a la capacité d'inhiber la croissance mycélienne de Aspergillus porri .

Résultats et discussion
49
2.3. Identification des actinomycètes pré sélectionnées
2.3.1. Résultats
2.3.1.1. Etude des caractères culturaux
Les critères généraux utilisés pour l'identification des actinomycètes sont: la
morphologie du mycélium aérien et de substrat, la morphologie des spores, la
production des pigments diffusibles, la production de la mélanine et l'utilisation des
sources de carbone et d'azote (Simon et al., 1999).
Après ensemencement sur 03 milieux de culture différents (Sabouraud, ISP2
et gélose nutritive), les colonies des souches ACT1, ACT2, ACT3, ACT4, ACT5,
ACT6 apparaissent au bout de 4 jours d'incubation à 30°C et se développent sur les
trois milieux gélosés. Tous les isolats se développent sur les milieux de culture
gélosés montrant ainsi des morphologies typiques des Streptomyces. Ces souches sont
des bactéries à Gram+ (Figure 47), aérobies, rondes à contour irrégulier, opaque,
pliées et à croissance lente, à bonne sporulation, à grande compacité. Les colonies
adhèrent fermement au milieu de culture où elles forment une légère dépression et
sont difficiles à prélever et à mettre en suspension. Ils sont menées d'un mycélium
aérien et de substrat de couleurs différentes et sont d'aspect corné et de surface
poudreuse, en plus toutes les colonies possèdent une odeur de terre, fraîchement
remuée correspondant probablement à la géosmine, et sont productrices
d'antibiotiques. Les caractéristiques culturales, et morphologiques de différentes
souches de Streptomyces sont présentées dans le Tableau 24.
Figure 47. Coloration de Gram+
de la souche
ACT2 Observée au microscope optique x1800

Résultats et discussion
50
Toutes les souches se développent sur les 03 milieux de cultures utilisés avec
différents degrés de croissance, allant d'une croissance faible à une croissance
abondante. La couleur du mycélium de substrat et aérienne varie selon les isolats :
l'isolat ACT2 produit un pigment marron sur Sabouraud et ISP2. La croissance la plus
abondante, pour les 06 souches étudiées a été observée sur le milieu (ISP2), elle est
moyenne sur le milieu (sabouraud). Le milieu (GN) est le milieu le moins favorable
pour la croissance des 06 souches d'actinomycètes.
Sur le milieu ISP2, la souche Streptomyces ACT1 est de contour irrégulier, la
surface des colonies est poudreuse, de couleur blanche ce qui nous a permis de la
classer dans la série des blanches, proposée par Shirling et Gotlieb, (1966). L'envers
des colonies est beige sur milieu ISP2 et absence de production de pigments
diffusibles.
La souche Streptomyces ACT2 montre une croissance moyenne sur ISP2 et
GN, et une croissance faible sur Sabouraud. Elle est d'aspect bosselées, compactées et
très sèche. La couleur du mycélium aérien apparait beige sur ISP2, ce qui nous a
permis de la classer dans la série des beiges, proposée par Shirling et Gotlieb, (1966).
Cependant, le mycélium de substrat est vert olive, la souche produit un pigment
marron diffusible sur ISP2. Cette souche se développe très lentement sur milieu ISP2
par rapport aux autres souches (5 à 6 jours) après ensemencement.
Les colonies de la souche Streptomyces ACT3 sont à croissance importante
sur les milieux ISP2 et Sabouraud, avec des colonies très petites, elles se développent
sous forme de colonies détachables, opaques. Le mycélium de substrat est abondant,
bien développé de couleur grise au centre à blanche de côté, ce qui nous a permis de
la rattacher à la série des grises proposée par Shirling et Gotlieb, (1966). La couleur
de mycélium de substrat est vert olive sur milieu ISP2, absence de production de
pigments diffusibles.
Les colonies de la souche Streptomyces ACT4 sont compacte non détachables,
à croissance abondante sur ISP2 et GN, la couleur du mycélium aérien est grise, ce
qui nous a permis de la classer dans la série des grise proposée par Shirling et Gotlieb,

Résultats et discussion
51
(1966), et de substrat vert olive sur ISP2, absence de production de pigments
diffusibles.
Les colonies de la souche Streptomyces ACT5 sont des colonies petites
détachables, à croissance importante sur ISP2 et sabouraud, le mycélium aérien est de
couleur blanche, ce qui nous a permis de la classer dans la série des blanches proposée
par Shirling et Gotlieb, 1966; cependant le mycélium de substrat est de couleur beige
sur ISP2, absence de production de pigments diffusibles.
Tableau 24. Caractères culturaux des souches: ACT1, ACT2, ACT3, ACT5 et ACT6 après 14 jours
d'incubation à 30°C.
Milieux /Souches ACT1 ACT2 ACT3 ACT4 ACT5 ACT6
GN
Croissance moyenne faible moyenne abondante moyenne abondante
Couleur de
mycélium
aérien
Blanc beige gris gris beige blanc
Couleur de
mycélium
de substrat
beige beige Vert olive Vert olive beige blanc
Mélanine - - - - - -
Sab.
Croissance moyenne faible abondante moyenne abondante abondante
Couleur de
mycélium
aérien
beige beige gris beige blanc blanc
Couleur de
mycélium
de substrat
beige beige Vert olive beige Blanc blanc
Mélanine - + - - - -
ISP2
Croissance abondante moyenne abondante abondante abondante abondante
Couleur de
mycélium
aérien
blanche beige Gris et
blanc de
côté
gris blanc beige
Couleur de
mycélium
de substrat
beige beige Vert olive beige beige beige

Résultats et discussion
52
Mélanine - + - - - -
Type de sporophore crochet droit spirale spirale spirale spirale
La souche Streptomyces ACT6 est à croissance importante sur les 03 milieux
ISP2, Sabouraud et GN, elle présente des colonies très petites, non détachables,
opaques. Le mycélium de substrat est abondant, la couleur du mycélium aérien et de
substrat est beige et absence de production de pigments diffusibles, ce qui nous a
permis de la classer dans la série des beiges proposée par Shirling et Gotlieb, (1966).
Les isolats de Streptomyces sont reconnue par 05 séries de couleurs (Blanc,
gris, rouge, bleu, variable: du violet-orange ou rose). Selon la couleur du mycélium
aérien sporulé et mature (Shirling et Gottlieb, 1966).
Dans ce présent travail, les 06 isolats de Streptomyces ont été distribués en
séries selon la couleur de leur mycélium aérien mature. 02 isolats ont été trouvé
appartenant à la série des grises (ACT3 et ACT4), 02 isolats autres appartiennent à la
série des blanches (ACT1 et ACT5), et enfin les souches ACT2 et ACT6
appartiennent à la série des beiges.
2.3.1.2. Observations microscopiques
L'examen microscopique au microscope optique (x10) des 06 souches retenues
révèle l'appartenance des 06 souches à la famille des Streptomyceraceae. La
morphologie de sporophores des souches ACT3, ACT4, ACT5 et ACT6 (figure 48)
est de type spirale, ce qui les place dans le groupe des spires de la famille des
Streptomycetaceae et le genre de Streptomyces. La morphologie des sporophores de la
souche ACT1 est en crochet alors que le type de sporophore de la souche ACT2 est
droit (figure 48).

Résultats et discussion
53
2.3.2. Discussion
2.3.2.1. Etude des caractères culturaux
les caractéristiques culturales observées sur les 03 milieux de cultures sont les
mêmes décrits par plusieurs auteurs caractérisant le genre Streptomyces (Anderson et
Wellington, 2001; Locci, 1989; Wendisch et Kutzner, 1991; Williams et al., 1989; Lo
et al., 2002).
D'après les résultats du tableau 24, les 06 isolats présentent les mêmes
caractéristiques décrit par plusieurs auteurs (Shirlling et Gottlieb, 1966; Williams et
al., 1989) (Williams et al., 1983). Les différences de couleur du mycélium aérien des
isolats et la production ou non des pigments est une indication de la diversité des
Streptomyces isolées. Ces résultats sont en accord avec ceux obtenus par Barakate et
al., (2002).
La production du mélanine est conditionnée par la composition en éléments
nutritifs dans le milieu notamment, la source de carbone et d'azote (Venkatesan et al.,
2008). Parfois, il est difficile de déterminer ce pigment surtout quand il s’agit d’un
milieu organique complexe (Venkatesan et al., 2008; Dasger et al., 2006). La
pigmentation chez les Streptomyces permet une distinction entre les différentes
espèces de ce genre (Dasger et al., 2006).
2.3.2.2. Observation microscopique
Les résultats de l'observation microscopiques des sporophores se rapprochent
de ceux observés par (Pridham et al., 1958; Williams et al., 1983; Anderson et
C
Figure 48. Observation microscopique des colonies d'actinomycète à l'aide du microscope optique:
A: observation microscopique d'une colonie de Streptomyces (x250) montrant des filaments courts à
la surface des colonies; B: un sporophore de type droit (x250); C: des sporophores de type spirale
x250.
B A

Résultats et discussion
54
Wellington, 2001; Locci, 1989; Wendish et Kutzner, 1991; Williams et al., 1989;
Oskay et al., 2004). L’absence des autres genres d’actinomycètes dans nos isolements
est probablement due au milieu de culture utilisé, qui est le milieu ISP2, connue pour
leur utilisation pour l'isolement des Streptomyces (Shirling et Gotlieb, 1966). De plus,
selon la littérature disponible, la composition en espèces d'actinomycètes dans la
rhizosphère des plantes est principalement déterminée par les paramètres des
échantillons de sol. Cependant, l'abondance des espèces particulières dépend
largement des espèces de plantes (Kalakutskii et Sharaya, 1990). De même, plusieurs
auteurs ont rapporté que le genre Streptomyces compte environ 95% des
actinomycètes du sol (Lacey, 1973; Elander, 1987). Sabaou et al., (1998), ont
constaté que le genre Streptomyces prédomine dans la majorité des sols du sud
algérien (80 à 90% du total). De plus, les 06 souches sélectionnées ont été isolées en
utilisant des dilutions correspondant 10-3
et 10-4
, ce qui augmente les chances d'isoler
les espèces de Streptomyces (Preobrazhenskaya et al., 1978; Cross, 1982).
Il est possible que les excrétions des racines des plantes du site de
prélèvements des échantillons du sol ont induit la présence des Streptomyces
producteurs d'antibiotique, car différents espèces et variétés des plantes produisent
différents types d'exsudats cellulaires, qui peuvent stimuler l'activité des
microorganismes pour la production des antifongiques (Lemanceau et al., 1995;
Wiehle et al., 1996).
Les méthodes traditionnelles utilisées pour l'identification des actinomycètes
aérobies filamenteux est laborieuse, nécessite du temps et demandent souvent des
séries de tests spécialisés (Steingrube et al., 1995;1997; Wilson et al., 1998; Harvey et
al., 2001), en plus, cette détermination ne peut pas identifier le genre précis de l'isolat
(Lechevalier, 1989).
2.4. Essai d'extraction et de séparation de la molécule antibiotique produite par
la souche ACT2
2.4.1. Résultats
Le métabolite active a été extrait par le n-butanol qui apparait d'être le
meilleur extractant selon plusieurs auteurs (Gupte et Naik, 1999; Lazaro et al., 2002;

Résultats et discussion
55
Atta et al., 2009; Khamna et al., 2009; Boudjella et al., 2006). La phase organique a
été collectée et évaporé en utilisant un rotavapeur à 50°C. La séparation de l'agent
antimicrobien en composé individuelle a été réalisée par CCM, 05 taches ont été
apparues. Le rendement de l'extraction est de 0.683 gr à partir de 2.5l du milieu ISP2
liquide, et il est de couleur marron jaunâtre. Ce rendement est très faible pour séparer
la molécule par colonne.
2.4.2. Discussion
En effet, Kavitha et al., (2010) ont obtenue un rendement de 1.8 gr, en
utilisant (25l) du milieu de culture ce rendement est très faible en comparaison avec
celui obtenue par cette étude. Cependant, Khamna et al., (2009) ont obtenue 0.781g
en utilisant 500ml du milieu de culture, ce rendement apparait d'être plus élevé que
celui obtenue par la souche ACT2. Ce faible rendement est due peut être à la souche
elle-même qui ne produit pas beaucoup d'antibiotique, ou bien au milieu utilisé pour
la fermentation qui est le ISP2. En effet, plusieurs auteurs ont utilisé autres milieux
(Khamna et al., 2009; Ouhdouche et al., 2001). Ouhdouche et al., (2001) ont rapporté
que le milieu benett's est le plus approprié à la production de molécules bioactives par
les Streptomyces. Plusieurs auteurs ont rapporté que la température de croissance chez
les Streptomyces diffère de la température du production de métabolites (Iwai et
Omura, 1992; Hussain et al., 1986). Il nous reste a démontré la nature des 05 taches
obtenues si elles correspondent au 05 composés actives contre le FOC ou seulement
un ou plus sont actives.
Figure 49. Résultat de la séparation de l'extrait brut de
l'isolat ACT2

Résultats et discussion
56
2.5. Essai de la lutte biologique sous serre
2.5.1. Résultats
Les résultats obtenus après 40jours de l'inoculation de 03 cultivars (ILC-482,
Col-27 et PPC-25) par la souche Streptomyces ACT2, suivi par l'inoculation de 03
isolats de FOC (FOC8, FOC13 et FOC15) après 48h de la première inoculation sont
présentés dans le tableau 25. Les figures50, 51 et 52 montrent la réduction des
symptômes observés chez les 03 cultivars sous serre en utilisant la souche
Streptomyces ACT2 en comparaison avec les témoins inoculés uniquement par le
pathogène et les témoins non inoculés.
Tableau 25. Degrés de suppression de la maladie par la souche ACT2 en utilisant les 3 variétés du pois
chiche
Variétés du pois chiche
ILC-482 Col-27 PPC-25
T
inoculé
FOC
+ACT2
DSM
(%)
T
inoculé
FOC
+ACT2
DSM
(%)
T
inoculé
FOC+
ACT2
DSM
(%)
FOC8 74.95% 69.81% 05.14 39.49% 11.97% 27.78 38.70% 14.15% 24.55
FOC13 82.75% 61.55% 21.20 34.16% 36.30% 00 52.32% 39.74% 12.58
FOC15 38.76% 41.21% 00 51% 37.72% 13.28 31.38% 12.16% 19.22
DSM: Degré de la suppression de la maladie par la souche Streptomyces ACT2.
D'après les résultats obtenus, le degré de la suppression de la maladie par
ACT2 varie d'un cultivar à un autre et d'un isolat de FOC à un autre. Il varie de 00 à
21.2% chez le cultivar ILC- 482, de 00 à 27.78% chez le cultivar Col-27et de 12.58%
à 24.55% chez le cultivar PPC-25. Le maximum de la suppression de la maladie a été
obtenu chez le cultivar Col-27 en utilisant l'isolat FOC8. La souche ACT2 a présenté
des degrés de suppression de la maladie plus élevés chez les deux cultivars Col-27
(13.68%) et PPC-25 (18.78%) modérément sensibles que le cultivar sensible ILC-482
(8.76%). Le degré de la suppression de la maladie par isolat de FOC varie, il est de
19.15% pour FOC8, 11.26% pour FOC13 et 10.83% pour FOC15.

Résultats et discussion
57
2.5.2. Discussion
La lutte biologique est une alternative appropriée surtout pour le contrôle des
phytopathogènes d'origine tellurique comme le FOC (Dubey et al., 2007). Landa et
al., (1997) ont rapporté que le flétrissement vasculaire du pois chiche est mieux
contrôlé par l'utilisation des cultivars résistant race – spécifique.
L'efficacité de cette méthode de lutte contre les races virulentes du pathogène
de FOC peut être amélioré par l'utilisation des microorganismes antagoniste. Le but
de ce travail est de savoir si l'isolat Streptomyces ACT2 sélectionné suite à la
première étape de sélection in vitro est capable d'éliminer ou de réduire le
flétrissement vasculaire chez les 03 cultivars du pois chiche sensibles et modérément
sensibles envers 03 isolats de FOC testés sous les conditions de la serre (in vivo = in
planta). La suppression des maladies telluriques par les microorganismes antagonistes
(champignons, bactéries) in vivo a été rapporté par pusieurs recherches qui ont
confirmé l'efficacité de cette méthode de lutte dans la protection des culture à
importance économique.
A titre d'exemple des champignons utilisés en lutte biologique comme:
Talaromyces flavus, Ghocladum sp., Trichoderma spp., Aspergillus giganteus,
Penicillium chrysogenum et des bactéries comme: Bacillus subtilis, Pseudomonas
spp. et les Streptomyces sp. ont montré une activité contre plusieurs phytopathogènes
(De Boer et al., 2003; Whipps, 2001), notamment contre les flétrissements vasculaires
causés par les différentes formae speciale de Fusarium oxysporum . Kumar, (1999) a
étudié la suppression de FOC en utilisant deux isolats: Pseudomonas fluerescens
RBT13 productrice de siderophores et Bacillus subtilis AF1 productrice
d'antibiotique. De même, Kaur et al., (2007) ont montré que les Pseudomonas isolées
à partir de la rhizosphère d'une culture du pois chiche saine sont tous antagonistes de
FOC. Dijksterhuis et al., (1999) ont montré que Paenibacillus polymyxa est capable
de supprimer le Fusarium oxysporum sous serre. Da Silva et al., (2005) ont indiqué
l'efficacité du contrôle du flétrissement vasculaire de la tomate par l'utilisation de F.
oxysporum non pathogène contre le F. o. f. sp. lycopersici race 2. Saravanan et al.,
(2004) ont étudié l'effet de Pseudomonas fluorescens isolées de la rhizosphère du
bananier sur la réduction du flétrissement vasculaire chez le bananier causé par F. o. f.
sp. cubens. En effet, Irum, (2007) a signalé que les résultats de la suppression du

Résultats et discussion
58
flétrissement vasculaire du pois chiche en utilisant les microorganismes sont plus
efficaces qu'en utilisant le carbendazim ou le bénomyl, qui sont deux produits
chimiques très efficaces contre le Fusarium oxysporum f. sp. ciceri.

Résultats et discussion
141
FOC13+ACT2 T non inoculé T inoculé par
FOC13
Figure 50. Résultats de l'effet protectif de la souche Streptomyces ACT2 chez le cultivar ILC-482 inoculé par l'isolat FOC13.

Résultats et discussion
142
FOC8+ACT2 T inoculé par
FOC8 T non inoculé
Figure 51. Résultats de l'effet protectif de la souche Streptomyces ACT2 chez le cultivar Col-27 inoculé par l'isolat FOC8.

Résultats et discussion
143
FOC15+ACT2 T inoculé par
FOC15 T non inoculé
Figure 52. Résultats de l'effet protectif de la souche Streptomyces ACT2 chez le cultivar PPC-25 inoculé par l'isolat FOC15.

Résultats et discussion
1
Si on compare nos résultats de l'antagonisme de la souche Streptomyces ACT2
sur la réduction des symptômes causés par le FOC sous serre nous avons observé que
nos résultats obtenus des degrés de suppression sont moins élevés que ceux obtenues
par plusieurs auteurs sur autres plantes attaquées par le flétrissement vasculaire.
Khavitha et al., (2010) ont trouvé que le degré de la suppression du flétrissement
vasculaire chez Sorghum biocolor en utilisant la souche Streptomyces sp TK-VL-333
a réduit l'incidence de la maladie de plus de 53.3% en comparaison avec le témoin. De
même, dans l'étude de Sudrez-Estrella et al., (2007) ont obtenus une réduction du
flétrissement de Fusarium oxysporum f. sp. melonis plus de 90% in vivo en utilisant
Aspergillus fumigatus en comparaison avec le témoin de T. harizanum. Kaur et al.,
(2007) ont obtenu une réduction de maladie du flétrissement vasculaire du pois chiche
à 100% (plante non malade) après 30 jours d'inoculation en utilisant une combinaison
un d'isolat de FOC non pathogène et une souche Pseudomonas flueorescens. La
réduction du flétrissement vasculaire chez les tomates a été entre 30-65% en utilisant
des bactéries et 40-66% en utilisant des champignons (Fusarium sp.; Trichoderma sp.
et Gliocladium spp.) (Larkin et fravel, 1998). Zaim et al., (2010) ont obtenu une
réduction de 83% de FOC en utilisant 03 isolats de microorganismes (Bacillus,
Streptomyces et Trichoderma harizanum).
Plusieurs travaux ont rapportés que des études in vivo ont donné des résultats
satisfaisantes en utilisant des Streptomyces contre quelques pathogènes d'origine
tellurique (Aghighi et al., 2004; Doumbou et al., 2002; Mohammadi et Lahdenpera,
1992). Tahvonen, (1982a; 1982b) et Tahvonen et Avikainen, (1987) ont signalé
l'utilisation de Streptomyces griseoviridis contre plusieurs phytopathogènes:
Alternaria brassicola, Botrytis cinerea, Fusarium avenaceum, Fusarium culmorum,
Fusarium oxysporum f. sp. dianthi, Pythium debaryanum, Phomopsis sclerotiodes,
Rhizoctonia solani, Sclerotinia sclertiorum. De même, en chine Valois et al., (1996)
ont utilisé Streptomyces sp. (5406) pour protéger les cultures du coton contre les
phytopathogènes telluriques. Autres études ont rapporté l'activité antifongique in vivo
de Streptomyces rimosus contre Verticillium dahliae et V. albo-atrum (Chi et Hanson,
1965).

Résultats et discussion
2
Bressan et al., (2003) ont étudié la lutte biologique des champignons affectant
le maïs par l'utilisation des Streptomyces spp. Ils ont rapporté que ces isolats sont
efficaces dans la suppression du développement de Aspergillus spp., Curvularia
lunata et Drechslera maydis et réduisent l'incidence de Fusarium subglutinans et
Cephalosporium acremonium.
Singh et Mehrotra, 1980 ont étudié l'effet de 06 Streptomyces spp. sur la
réduction des symptômes causés par Rhizoctonia bataticola sur pois chiche, ils ont
observé que les souches de Streptomyces sp. réduisent les symptômes de la maladie et
améliorent la croissance et la production de la matière sèche chez le pois chiche. Il
existe aussi des rapports sur le contrôle des champignons pathogènes racinaires en
utilisant Streptomyces sp. (Tahvonen et Avikainen., 1987). Le produit commerciale
Mycostop, basé sur la souche K61 de Streptomyces griseoviridis et S. lydicus
WYEC108 est utilisé contre quelques maladies racinaires et le flétrissement causés
par Pythium spp., Fusarium spp., Rhizoctonia spp. et phytophthora spp. (Mahadevan
et Crawford, 1997). Streptomyces sp. présente le même potentiel à supprimer l'attaque
de la vigne causée par B. cinerea in planta en réduisant l'extension de la colonisation
du champignon dans les tissues de la plante (Loqman et al., 2009).
Le degré de suppression de la maladie est plus prononcé chez les cultivars
modérément tolérant (Col-27 et PPC-25) que le cultivar sensible (ILC-482). Ce
résultat est similaire à celui obtenu par Landa et al., (2001). En effet, les plantes
possèdent leurs propres mécanismes de résistance (Franceschi et al., 2005), mais les
microorganisme du sol peuvent contribuer à cette résistance en excrétant des
substances limitant la croissance des champignons phytopathogènes (Vassilev et al.,
2006; Jain et Jain, 2007) ou stimulant les défenses des plantes (PGPR) (Prévost et al.,
2006; Lehr et al., 2008).
L'observation d'un faible degré de suppression de la maladie du flétrissement
vasculaire par la souche ACT2 chez les 03 cultivars du pois chiche sous serre présenté
par cette étude peut être le résultat de plusieurs facteurs biotiques et abiotiques dans le
pathosystème cités par plusieurs auteurs (Kazmar et al., 2000; Raaijmakers et al.,
1995; Schisler et al., 1997; Smith et al., 1999).

Résultats et discussion
3
Parmi ces facteurs: les facteurs liés au pathogène (inoculation des isolats très
virulents, la concentration de la race 5 du FOC), les facteurs liés aux conditions de
l'environnement (température et pH) et les facteurs liés à l'agent de lutte biologiques
(température optimale, pH).
Les facteurs liés au pathogène: Landa et al., (1997); Harvas et al., (1995); Navas-
Cortes et al., (2000) ont montré que l'incidence et la sévérité du flétrissement du pois
chiche est influencée par la l'inoculation des isolats très virulents du FOC de la race 5.
Cette race est capable de présenter le même degré de la maladie provoqué par une
autre race même à faible quantité.
Les facteurs liés aux conditions de l'environnement: Etant donné que les conditions
de la serre où nous avons lancé le test n'était pas contrôlé. Landa et al., (1997);
Boland, (1997); Bull et al., (1991); Hannusch et Boland, (1996), Hervas et al., (1997);
Johnson, (1994); Kazmar et al., (2000); Larkin et Fravell, (1999); Montesinos et
Bonaterra, (1996); Raaijmakers et al., (1995); Schisier et al., (1997); Smith et al.,
(1999) ont rapporté que les conditions environnementales optimales pour le
développement de la maladie réduisent l'effet protecteur des antagonistes. Des études
sur la pourriture grise, la fonte de semi et le flétrissement vasculaire du pois chiche
ont étudiées cette effet (Landa et al., 2001; Ben –Yephet et Nelson, 1999; Harris et
al., 1997; Mukherjee et Raghu, 1997; Tedla et Stanghellini, 1992).
La température: est un facteur important influençant la réussite d'un agent de lutte
biologique (Landa et al., 2001), et la sévérité du flétrissement vasculaire est modulée
par la température (Gupta et al., 1987 ; Navas-Cortes et Jimenez-Diaz., 1998), mais
l'effet exacte de la température sur le développement de la maladie du flétrissement
vasculaire n'est pas encore connu (Landa et al., 2001). Plusieurs possibilités peuvent
expliquer l'effet de la température sur la suppression du flétrissement vasculaire du
pois chiche par les bactéries antagonistes. La première possibilité et que l'inoculum du
pathogène à 25°C est plus élevé à être neutraliser par l'agent de la lutte biologique, à
cause que la maladie se développe mieux à 25°C en comparaison avec 20°C et 30°C
(Landa et al., 2001), même si une densité plus petite a été inoculé, car à 25°C le
potentiel de la maladie apparait saturé.

Résultats et discussion
4
Landa et al., (2001) ont rapporté qu'en absence de l'agent de lutte biologique, des
nombres élevés de sites d'infections sont nécessaires à 20 et 30°C en comparaison
avec 25°C pour que le flétrissement vasculaire chez le pois chiche se développe tôt,
rapide et sévèrement. L'effet de la température sur la modulation de la suppression de
la fusariose du pois chiche par les bactéries introduites peut aussi résulter de
l'influence de la température sur leurs activités (Landa et al., 2001). Beaucoup de
travaux ont indiqué que l'efficacité d'un agent de lutte dans la suppression de la
maladie se réduit lorsque les conditions deviennent plus favorables au développement
de la maladie (Boland, 1997; Kazmar et al., 2000). Ben-yephet et Nelson, 1999 ont
trouvé que le degré de suppression de la maladie de la fonte de semi causée par
Pythium spp. et traiteé par les composts dépend de la température d'incubation, ils ont
suggéré que cet effet peut être due au différents niveau de l'activité microbienne dans
les différents composts à différentes températures d'incubation, ce type de réponse à la
température a été aussi observé dans la lutte biologique contre Pythium
aphanidermtum sur la canne à sucre par les rhizobactéries (Tedla et Stanghellini,
1992). Dans cette dernière étude la suppression du pathogène a été exprimé à 20°C
mais non pas à 27°C et cette effet a été directement corrélé avec l'activité des
bactéries dans la rhizosphère. Une colonisation rapide et persistante dans la
rhizosphère des agents de lutte biologiques est importante dans le contrôle biologique
des fusarioses à cause des infections motiles courtes dans ces maladies (Baker et
Paulitz, 1996). Parce que plusieurs bouts de racines se développent durant la
croissance racinaire, il y a une probabilité élevée des infections qui réussit sur eux aux
conditions optimum de la maladie, ainsi, la température adéquate pour une
suppression efficiente de la maladie doit réduire l'efficacité de l'inoculum du
pathogène et encore permettre aux rhizobactéries introduite de faire face avec la
croissance racinaire pour protéger le terrain de l'infection (Landa et al., 2001).
Hussain et al., (1986) ont rapporté que la température d'incubation des actinomycètes
joue un rôle important dans la détermination du composé d'antibiotique produit. En
effet une souche est capable de produire un antibiotique inhibant un pathogène à 25°C
produit un autre antibiotique qui inhibe un autre pathogène à 35°C sans aucun effet
sur le premier et le milieu de culture aussi joue un rôle important dans la
détermination du composé secrété et sa quantité.

Résultats et discussion
5
Les facteurs liés à la nature de l'agent de lutte biologique et la dose appliquée: la
quantité finale des bactéries qui ont adhéré aux racines et à la rhizosphère de la plante
hôte (pois chiche) joue un rôle essentiel dans la suppression de la maladie (Landa et
al., 1997). En effet, la quantité adhérée aux racines dépend de la nature du
microorganisme antagoniste (Hervas et al., 1997; Landa et al., 1997) et son intensité
de la colonisation (Colonisation partielle ou complète) (Jayasinghe et Parkinson,
2008),la dose appliquée de l'agent de lutte (Boland, 1997; Bull et al., 1991; Hannusch
et Boland, 1996, Hervas et al., 1997; Johnson, 1994; Kazmar et al., 2000; Larkin et
Fravell, 1999; Montesinos et Bonaterra, 1996; Raaijmakers et al., 1995; Schisier et
al., 1997; Smith et al., 1999) et la période d'incubation dans le sol (Jayasinghe et
Parkinson, 2008). Jayasinghe et Parkinson, (2008) ont montré que les plantules de
céréales inoculées après 5 jours de croissance dans le sol par les Streptomyces sont
complètement colonisées par plusieurs champignons. Cependant les plantules de
céréales inoculées par les phytopthogènes 14 jours après une inoculation par les
Streptomyces ont montré une absence totale des champignons phytopathogènes. Le
besoin de séparer la première inoculation avec le non pathogène par au moins
quelques jours (6 jours) de la deuxième inoculation avec le pathogène a été indiqué
par plusieurs auteurs chez plusieurs formae speciales du flétrissement vasculaire
(Matta, 1989; Hervas et al., 1997). La période du temps demandée entre la première
et la deuxième inoculation doit être nécessaire pour que l'antagoniste colonise
complètement les racines des plantes avant d'être attaquées (Hervas et al., 1997).
Simon et Sivasithamparam, 1989 ont démontré que la période d'incubation est
nécessaire pour une lutte efficace, elle joue un rôle dans la production d'antibiotiques
(Kathiresan et al., 2005).
Les facteurs liés au génotype de la plante hôte: Plusieurs auteurs ont signalé
que le génotype des plantes influence le degré de la suppression de la maladie (Landa
et al., 1997; Kazmar et al., 2000; Raaijmakers et al., 1995; Smith et al., 1999). Harvas
et al., (1995); Hervas et al., (1997) et Navas-Cortes et al., (2000) ont étudié cet effet
sur le pois chiche et ils ont constaté que les génotypes modérément sensibles sont
mieux protégés par les agents de lutte que les génotypes sensibles. Van Peer et al.,
(1991) ont signalé que la souche P. fluerecens WCS417 supprime le flétrissement

Résultats et discussion
6
vasculaire chez un cultivar modérément résistant de l'œillet (cv. Bllas) plus efficient
que chez le cultivar sensible (cv. Lena).
L'influence des génotypes du pois chiche sur la protection de la maladie n'est
pas attribuée aux différences dans la capacité de cette isolat à coloniser la rhizosphère
des cultivars testés (Hervas et al., 1997).
La méthode d'inoculation de l'agent de lutte biologique: le traitement des semences
par les antagonistes montre une efficacité plus élevé que le traitement du sol ou de
racine (Kaur et al., 2007). Nos résultats indiquent que le traitement des racines par les
Streptomyces réduit la maladie, mais il ne la supprime pas et que ce traitement n'est
pas suffisant. La recherche d'autres méthodes d'inoculation est mieux comme le
traitement des semences ou bien traitement du sol ou bien les deux combinés. Ce
résultat est similaire à celui obtenue par Landa et al., (1997) en utilisant des Bacillus
et des Pseudomonas contre le FOC (Landa et al., 1997).
Des résultats positifs du premier screening in vitro indiquent la production de
substances diffusibles dans le milieu n'indiquent pas que cette Streptomyces ACT2
protège les plantes du pois chiche par la production d'antibiotiques. Malgré que cette
production ait été observée in vitro contre le champignon aucune preuve n'indique
que la réduction de la maladie sous serre était par l'antibiose. Le mécanisme
spécifique d'action de la souche ACT 2 sur les 03 cultivars du pois chiche responsable
à la réduction de la croissance de FOC sous les conditions de la serre (sol,
température,…) reste inconnue (antibiose, compétition, hyper parasitisme,…)
(Jayasinghe et Parkinson, 2008).
3. Etude du comportement variétal
3.1. Résultats
3.1.1. Etude de l'agressivité
L'agressivité de 17 FOC choisis en fonction de l'organe d'isolement (tiges) et
de la région d’étude vis-à-vis du cultivar ILC-482 connu par sa sensibilité envers le
FOC (Jimenez-Diaz et al., 1988; Rhrib, 1990) a été également évaluée. Les résultats
présentés dans le Tableau 26, indiquent que l'ensemble des isolats inoculés à la variété

Résultats et discussion
7
ILC-482 ont provoqué des symptômes typiques de la fusariose vasculaire du pois
chiche. Les altérations foliaires des plantules inoculées ont débuté par un
jaunissement et une nécrose soudaine des folioles de la base. Ce jaunissement se
développe pour atteindre la totalité des feuilles.
Les premiers symptômes ont été apparus 17 jours après inoculation pour
quelques isolats, mais pour la plupart les premiers symptômes ont été apparus 23 jours
après inoculation. D'autres isolats ont provoqué les symptômes en 30 jours. Durant
toute la durée de l'essai, les plantes témoins maintenues dans les mêmes conditions
que les plantes inoculées, n'avaient manifesté aucun symptôme.
L'indice de la maladie (IM) a été calculé 40 jours après inoculation. Les degrés
d'agressivité des isolats sont regroupés en 5 classes: 0-20% (très peu agressif), 21-
40% (peu agressif), 41-60% (moyennement agressifs), 61-80% (agressifs), 81-100%
(très agressif).
Sur la base de cette échelle, les 17 isolats ont montré une variabilité de
l'agressivité (Tableau 26). Les indices de la maladie (IM) ont varié de 32.60% à
92.12%. Parmi ces isolats de FOC: 06 isolats se sont révélés peu agressif, 02
moyennement agressifs, 07 agressifs et 02 très agressifs.
Figure 53. Résultats des tests d'agressivité des isolats testés de FOC.

Résultats et discussion
8
Tableau 26. Classification des 17 isolats selon leur agressivité
Les résultats obtenus montrent que la population analysée, isolée de plants du
pois chiche cultivés dans les régions de Tlemcen, Ain Temouchent et Mostaganem,
sont variables, puisque le test de l'agressivité a montré une réaction hétérogène de
l'ensemble des isolats inoculés à la variété ILC-482.
3.1.2. Etude du comportement variétal vis-à-vis de FOC
Les réactions des 11 cultivars du pois chiche vis-à-vis des cultures
monosporées de 03 isolats testés sont résumées dans le tableau 27. Nous avons
constaté que tous les isolats testés ont manifesté une virulence vis – à- vis de ces 11
génotypes. Les résultats apportés par cette étude permettent d’effectuer un classement
Isolats IM (%) Evaluation de l'agressivité
FOC1 61.36 Agressif.
FOC2 92.12 Très agressif.
FOC3 39.44 Peu agressif.
FOC4 78.12 Agressif.
FOC5 67.37 Agressif.
FOC6 32.33 Peu agressif.
FOC7 89.29 Très agressif.
FOC8 74.95 Agressif.
FOC9 48.92 Moyennement agressif.
FOC10 76.26 Agressif.
FOC11 38.37 Peu agressif.
FOC12 32.60 Peu agressif.
FOC13 82.75 Agressif.
FOC14 55.07 Moyennement agressif.
FOC15 38.76 Peu agressif.
FOC16 70.27 Agressif.
FOC17 36.75 Peu agressif.

Résultats et discussion
9
des variétés en fonction de leur résistance au flétrissement selon trois catégories
(tolérantes, intermédiaires et sensibles). Les résultats obtenus nous ont permis
d’identifier une tolérance pour les variétés F7-43, F10-13 et 2G-03 pour les 03 isolats
de FOC. Une tolérance pour 02 isolats de FOC exprimée par les variétés ILC-2379,
Fedjoj, F10-31et seulement une variété tolérante à un seul isolat de FOC.
Le reste des génotypes présentent des réactions intermédiaires ou sensibles
vis-à-vis les trois FOC testés.
Les résultats de cet essai qui représente un effectif de 11 variétés, mettent en
évidence des niveaux de sensibilité différents et nous permettent d’établir un
classement en prenant compte des trois origines d’inoculum. Ce classement présente
une valeur supplémentaire, car il permet de donner une idée sur les gènes impliqués
dans la résistance ou la tolérance des variétés testées du pois de chiche.
Figure 54. Test de la résistance variétale montrant 02 variétés
tolérantes F10-31 et 2G-03.

Résultats et discussion
10
Tableau 27. Réponse des cultivars de pois chiche vis-à-vis de FOC
Variétés FOC15 FOC8 FOC 13
ILC-482 38.76% 74.95% 82.75%
ILC-2379 14.02% 26.4% 17.39%
39-96 45,55% 39,58% 27,67%
F7-43 00% 00% 00%
F10-13 00% 00% 00%
Fedjoj 4.10% 21.87% 1.91%
PPC-25 31.38% 38.70% 52.32%
Col-27 51% 39.49% 34.16%
haddache 48.53% 17.12% 56.05%
2G-03 19.86% 4.97% 9.12%
F10-31 34.15% 18.75% 12.43%
IM= nombre de feuilles atteintes/nombre de feuilles total
Chez tous les cultivars inoculés par les 03 isolats, les symptômes exprimés ont
consisté en un jaunissement vasculaire progressif (flétrissement tardif) qui s'est
développé dès le 23éme
jour après inoculation, en affectant d'abord les feuilles les plus
basses. Enfin les témoins, lots de cultivars non inoculés, non pas manifesté les
symptômes du flétrissement vasculaire.
Figure 55. Test de la résistance variétale montrant des symptômes de la
fusariose chez la variété sensible ILC-482
FOC15 T
ILC-482

Résultats et discussion
11
L'analyse statistique a montré qu'il ya des différences hautement significatives
dans le comportement de ces variétés testées. Ce test nous a permis de classer les
variétés du sensibles au tolérantes, comme suit: ILC-482>Col-27>PPC-25> Haddache
>39-96>F10-31>ILC-3279>2G-03>Fedjoj>F7-43>F10-13.Après l'évaluation de la
réaction des lignées du pois chiche, des réisolements ont été effectués dans le but de
confirmer la nature des symptômes et de préciser si le pourcentage de réisolement
positif (%RP), à partir des plantes hôtes inoculées, est corrélé à leur réaction de
sensibilité ou de tolérance.
Le tableau 28 présente les pourcentages de réisolement positif (%RP) du
pathogène à partir de fragments de ces cultivars 40 jours après l'inoculation.
L'analyse des résultats a montré que ce dernier (RP%) peut être élevé quand
l'indice de la maladie est faible (réaction de tolérance), c'est le cas pour les cultivars
2G-03 et F10-13. Ceci témoigne de la colonisation de la plante hôte même quand
celle ci est tolérante à l'infection.
Figure 56. Photo de ré isolement du FOC à partir des
fragments de plantes de pois chiche

Résultats et discussion
12
Tableau 28. Résultats du test de ré isolement des 03 FOC
NT: Non testée
Le FOC a été fortement réisolé à partir des variétés exprimant une sensibilité.
Par contre le pourcentage de réisolement peut être faible quand l'indice de la maladie
est élevé, c'est le cas des cultivars Haddache et Col-27. De plus, le pathogène n'a pas
été obtenu à partir des plantes témoins. Le réisolement de FOC à partir des lignées
tolérantes à l'infection, témoigne de leur colonisation par l'agent pathogène.
3.2. Discussion
3.2.1. Etude de l'agressivité:
Les degrés de l'agressivité obtenue par cette étude montrent que les isolats de
la population étudiée présentent différents degrés. Ce résultat se rapproche des
résultats d'El Aoufir, (2001); Jimenez-Casco et al., (2004); Bekkar , (2007) sur le
FOC et Belabid,2003 sur FOL.
FOC13 FOC8 FOC15 Variétés
NT NT NT ILC-482
NT NT NT ILC-2379
32.5% 33.45% 55.60% 39-96
4.54% 1.02% 2.66% F7-43
8.51% 3.44% 4.76% F10-13
40.17% 51.69% 55.76% Fedjoj
48.73% 59.84% 53.47% PPC-25
21.14% 59.84% 50.34% Col-27
21.14% 59.84% 29.34% haddache
46.21 11.31% 4.76% 2G-03
3.19% 10.89% 5.48% F10-31

Résultats et discussion
13
3.2.2. Etude du comportement variétal
Cette étude a permis de mettre en évidence que, parmi les variétés du pois
chiche, des tolérances variétales ainsi que des sensibilités à la fusariose ont été
observé.
La prise en compte par l’agriculteur de ce moyen alternatif de lutte par
l’utilisation de variétés ciblées obtenues par ce test peut permette de répondre de
façon préventive au risque potentiel de la présence de cette maladie sur son
exploitation. En raison de la grande disparité variétale observée, même si aujourd’hui
les 02 variétés testées (F7-43 et F10-13) ne sont pas utilisées malgré leur bon
comportement vis-à-vis du cryptogame étudié, d’autres variétés se sont montrées
intéressantes comme (ILC-2379, Fedjoj, F10-31). Une bonne connaissance de la
tolérance variétale est un élément déterminant dans une logique de lutte raisonnée
contre F. oxysporum ciceri, elle permet ainsi d’adapter les seuils de déclenchement
des modèles épidémiologiques. Il faut cependant garder à l’esprit que les tolérances
sont en constante évolution et sont vouées, à plus ou moins long terme, à être
contournées par l’agent causal. L’évaluation du comportement variétal, présente un
double intérêt:
Elle permet de connaître le niveau de résistance de ces variétés, pour pouvoir
ensuite les intégrer dans les stratégies de lutte raisonnée.
Elle permet de remettre à jour le classement des sensibilités variétales établis
les années précédentes.
L’apport de la résistance variétale dans la lutte contre le flétrissement
vasculaire du pois chiche peut être pris en compte. Nous avons observé des
différences entre les différentes variétés du pois chiche tant au niveau de la précocité
des attaques que de leur sensibilité à la maladie. En effet, un décalage d’au moins 1 à
2 semaines est observé au niveau des attaques entre les variétés sensibles et tolérantes.
La variété sensible ILC-482 a été touchée par la maladie plus rapidement que la
variété tolérante 2G-03. Enfin, Il apparait clairement que le critère tolérance variétale
doit être pris en compte dans la mise en place d’un programme de lutte intégrée. Les
informations issues de ce test variétal permettent de mieux utiliser le potentiel de

Résultats et discussion
14
chaque variété dans la mise en œuvre du raisonnement de la protection fongicide. Des
recherches doivent être poursuivies en utilisant le maximum de variétés cultivées en
Algérie dans le but de faire un screening des variétés résistantes ou tolérantes au
flétrissement. De même, l’identification des races physiologiques existantes a son
importance sur le comportement variétal.
Depuis longtemps dans de nombreux pays, des gènes de résistance au
flétrissement ont été sélectionnés (Muehlbauer et Rajesh., 2008; Gowda et al., 2009;
Halila et al., 2009), cette sélection se heurte malheureusement au grand potentiel
adaptatif de l’agent causal FOC, qui se manifeste par une sélection de races
virulentes, ce qui ruine assez rapidement les résistances incorporées aux cultivars.
4. Etude de la relation entre l'agressivité, les morphotypes, la pigmentation des
thalles, le diamètre de la croissance mycélienne, les mensurations des conidies
4.1. Résultats
Le tableau 29 représente la répartition des isolats de FOC en fonction des
morphotypes, la pigmentation du thalle, la croissance mycélienne, la mensuration des
conidies et le degré de l'agressivité. Le tableau 30, présente une matrice de corrélation
entre l'ensemble des paramètres étudiée.
Tableau 29. Répartition des isolats de FOC en fonction des morphotypes, la pigmentation du thalle, la
croissance mycélienne, la mensuration des conidies et le degré d'agressivité.
Degré d'agressivité Total
Peu
agressif
Moyennement
agressif
Agressif Très
agressif
Morphotype
Cotonneux
Duveteux
Ras muqueux
0
2
4
0
1
1
2
3
2
0
1
1
17
Pigmentation
Blanc
6
2
5
1
17

Résultats et discussion
15
Les groupes établis selon le groupement des isolats effectués par le test de Newman et Keuls à 5%.
4.1.1. Relation entre l'agressivité, la morphologie et la pigmentation du thalle
L'analyse des résultats de la morphologie des thalles de FOC n'a pas donné de
corrélation entre ce caractère et le degré d'agressivité des isolats (r=0.225) (Tableau
30). Il semble que la morphologie du thalle, chez cette forme spéciale, ne caractérise
pas le degré d'agressivité des isolats puisque la plus part des morphotypes ont été
rencontrée, à des pourcentages prés, quel que soit leur niveau d'agressivité.
Le coefficient de corrélation obtenu entre l'agressivité / pigmentation (r=-
0.203) est faible et qui démontre l'absence de corrélation. Ce résultat montre, chez le
FOC, que la pigmentation ne caractérise pas le degré de l'agressivité des 17 isolats.
4.1.2. Relation entre l'agressivité et le diamètre de croissance du thalle
Les résultats de diamètre de croissance du thalle n'ont pas donné une
corrélation entre ce caractère et le degré d'agressivité des isolats (r=0.23). La
variabilité de la croissance n'est pas associée à l'agressivité.
Violet 0 0 2 1
Croissance
Groupe a
Groupe b
Groupe c
Groupe d
1
2
1
2
0
1
0
1
3
1
2
1
0
1
1
0
17
Mensuration des
conidies
Groupe a
Groupe b
Groupe c
Groupe d
1
1
3
1
0
1
1
0
0
5
1
1
1
0
0
1
17

Résultats et discussion
16
4.1.3. Relation entre l'agressivité et la mensuration des macroconidies et des
microconidies
Les résultats des mensurations en largeur et la longueur des macroconidies
montrent qu'il n'existe pas de corrélation entre ces deux caractères et le degré
d'agressivité des isolats (r=-0.315 et -0.128) respectivement. Les deux caractères:
degré d'agressivité des isolats et la longueur des microconidies n'ont pas montré une
corrélation (r=0.144). De même, pour le degré d'agressivité et la largeur des
microconidies (r=-0.128) (Tableau 30). D'après les résultats du tableau 30, une
corrélation a été établie entre la longueur des microconidies et la largeur des
microconidies (r=0.636).
Tableau 30. Matrice de corrélation calculée sur 8 paramètres étudiés
A B C D E F G H
A 1
B 0.053 1
C -0.023 -0.485 1
D 0.05 0.088 0.361 1
E -0.084 0.025 0.112 0.636 1
F -0.192 0.028 -0.337 -0.456 -0.38 1
G -0.131 -0.219 0.233 -0.025 -0.062 0.121 1
H 0.225 -0.203 0.23 0.144 -0.128 -0.128 -0.315 1
A: Morphotype, B: Pigmentation, C: Diamètre de croissance mycélienne, D: Longueur des
microconidies, E: Largeur des microconidies, F: Longueur des macroconidies, G: Largeur des
macroconidies, H: Agressivité.
4.2. Discussion
Les résultats obtenus lors de cette étude montrent qu'il n'existe pas de
corrélation entre les morphotypes et les degrés d'agressivité. Des résultats similaires
ont été montrés par El-Aoufir, (2001).

Résultats et discussion
17
Nos résultats concordent aussi avec ceux de Follin et Laville, (1996) chez
Fusarium oxysporum f. sp. cubens, d'Assigbeste (1989) chez F. oxysporum
vasinfectum, de Henni et al., 1994 chez F. o. f. sp. lycopersici, de Belabid, (2003)
chez F. o f. sp. lentis et de Bekkar, (2007) chez le FOC. Par contre plusieurs auteurs
ont trouvés une corrélation entre le morphotype et le degré de l'agressivité (Chauhan,
1992; Claudius et Mehrotra, 1973; Abbas, 1995).
L'étude de la corrélation entre l'agressivité et les mensurations de la longueur
et de la largeur des microconidies et des macroconidies a montré qu'il n'existe pas de
corrélation entre eux. Ce résultat est en contradiction avec le résultat obtenu par
Bekkar, (2007) sur le FOC et Belabid, 2003 sur le FOL. Ces caractéristiques ont été
associées aux isolats agressifs et moyennement agressifs. Par contre, les résultats de
Abbas, (1995), sur la forme spéciale lentis, ont montré une corrélation entre le degré
d'agressivité et la largeur des microconidies seulement.
Une corrélation a été établie entre la longueur et la largeur des microconidies
(r=0.636), ce résultat est en accord avec celui obtenu par Bekkar, (2007) chez le FOC.

45
Conclusion et perspectives

Conclusions et perspectives
45
Conclusion générale et perspectives
En Algérie, la culture du pois chiche occupe une place importante parmi les
légumineuses alimentaires. Cette culture occupait en 2002 une superficie avoisinant
les 550 ha. En plus de son utilisation comme source de protéines, le pois chiche
participe à l’amélioration de la fertilisation du sol grâce à sa capacité à fixer l’azote
atmosphérique. Cependant, cette culture est exposée aux infections naturelles pouvant
conduire rapidement à la dégradation de la qualité sanitaire originelle. Parmi les
pathogènes les plus redoutables et qui méritent donc une attention particulière, le
flétrissement vasculaire du pois chiche, qui constitue une énorme contrainte pour la
culture, cela dans de nombreuses régions où elle est pratiquée. Cette maladie est
causée par un agent pathogène appelé F. oxysporum f.sp. ciceri. elle constitue une
menace autant plus grave que l’anthracnose, pour la culture du pois chiche, en raison
de son développement rapide et les dégâts qu’il occasionne.
Au cours des prospections effectuées dans les régions de Tlemcen, Ain
Temouchent et Mostaganem, nous avons constaté que les attaques du flétrissement du
pois chiche dû à l’agent causal FOC peuvent prendre une grande ampleur durant
certaines saisons lorsque l’apparition de la maladie est précoce et les conditions
climatiques sont favorables. Les symptômes du flétrissement vasculaire manifestés au
champ sont similaires à ceux rapportés dans d'autres pays, il s'agit soit d'un
flétrissement précoce entrainant la mort de la plante avant le stade de la floraison, soit
d'un jaunissement vasculaire progressif accompagné d'un brunissement vasculaire et
qui correspond au flétrissement tardif, qui a été le plus répandu.
La fusariose du pois chiche reste une maladie grave en attaque précoce et le
rendement peut être fortement diminué. En attaque tardive, c’est la qualité de la
récolte qui est mise en cause. De plus, le producteur doit faire face à de nouvelles
races du flétrissement dont l’agressivité semble plus forte. Cette évolution de
l’agressivité est d’ailleurs démontrée par diverses études. D’autres travaux se
rejoignent pour confirmer l’évolution du comportement des races physiologiques de
F. oxysporum. Leur agressivité, la précocité des attaques, la rapidité d’extension des
dégâts expriment cette évolution.

Conclusions et perspectives
46
Au terme de cette étude, les différents essais mis en place ont permis de
dégager les constatations suivantes: le Fusarium oxysporum est le genre le plus
dominant des isolements effectués à partir des fragments de tiges et de racines des
plantes infectées. Ce constat, nous a permis de supposer que le FOC, est l’agent
responsable du flétrissement observé sur culture du pois chiche dans les régions
prospectées. Nos observations laissent apparaître qu’il est indispensable de
sensibiliser les agriculteurs sur l’étiologie de cette maladie cryptogamique pour qu’ils
appliquent les mesures prophylactiques au moment opportun et d’encourager les
études sur la dynamique des populations de ce pathogène ainsi que sa relation avec la
plante-hôte. Au cours de ce travail, nous avons maîtrisé la technique d’isolement et de
caractérisation de l’agent causal, nous avons ainsi obtenu une collection de 65 isolats
de FOC. Cette étude a permis une meilleure connaissance de la dynamique de
population de F. oxysporum ciceri et a mis en évidence la présence de plusieurs
morphotypes de l’agent causal.
Il en ressort de l'étude physiologique que le milieu le plus favorable pour la
croissance de FOC est le milieu pois chiche, et que la température optimale pour ce
parasite est autour de 30°C et un pH de 5.5 est le plus favorable. Nous avons
démontré aussi que l'obscurité apparait adéquate pour le FOC que l'incubation sous
lumière continue et ceci induit la pigmentation. Concernant la détermination de la
source de carbone nous avons remarqué que le glucose est la meilleur source, en
revanche la peptone s'est révèle être la meilleur source d'azote.
Les diverses méthodes utilisées pour tester le phénomène d’antagonisme entre
Fusarium oxysporum ciceri et espèces d’actinomycète retenues pour cette étude, nous
a permis de conclure que les souches d’actinomycètes testées exercent une action
antibiotique contre F. oxysporum ciceri, celle-ci a été mise en évidence lors du test de
confrontation direct. L’action des métabolites diffusibles dans le milieu a montré une
réduction importante de la croissance mycélienne de F. oxysporum ciceri par rapport
au témoin. Les résultats de l’effet protecteur ont montré que le traitement des racines
avec des souches d’actinomycètes retardaient le début des symptômes et réduisaient
l’incidence de la maladie comparé avec les contrôles non traités. Ces résultats

Conclusions et perspectives
47
indiquent que le traitement des racines de plants du pois chiche modérément tolérants
avec les isolats d’actinomycètes sélectionnées peut aider dans le bio-contrôle de FOC.
Cette étude préliminaire basée sur ces techniques de base offre la possibilité
d’obtenir et de sélectionner un agent de bio-contrôle contre cet agent pathogène. Elle
mérite également d’être encouragée, développée et poursuivie dans le but de
sélectionner une ou plusieurs souches performantes. Il serait opportun de prévoir
d’une part, l’analyse des métabolites des souches d’actinomycètes performantes afin
d’identifier la nature chimique des substances antibiotiques mises en jeu dans le
phénomène d’antibiose et d’autre, pour mieux maîtriser leur dosage.
Les résultats des tests du comportement variétal a permis de bien montrer
l’intérêt que pourrait avoir l’intégration de la résistance variétale dans la lutte
raisonnée contre F. oxysporum ciceri. Sa prise en compte dans des systèmes d’aide à
la décision est particulièrement appropriée, car elle permet d’adapter au mieux la date
de premier traitement et les rythmes d’applications pendant la saison. Rappelons
cependant que cette résistance est susceptible d’évoluer d’année en année. Pour mettre
en évidence cette évolution, il est intéressant d’effectuer des essais et de comparer les
résultats obtenus avec ceux des années précédentes. Il faut noter que les résistances
variétales sont vouées, à plus ou moins long terme, à être contournées par l’agent de
flétrissement. Il est donc nécessaire de remettre à jour régulièrement les classements
de sensibilité en renouvelant les essais d’étude de la résistance variétale.
Compte tenu de l’évolution des populations de l’agent pathogène et des systèmes
de production agricole, il faut poursuivre les recherches afin de mettre au point de
nouvelles variétés du pois chiche possédant une résistance au champignon. Cette
résistance est importante pour les producteurs de cette culture. L’évaluation du niveau
de risque et le choix de mesures de prévention appropriées sont tributaires d’une
information précise et actuelle sur les lieux où le pathogène est présent. Vue la
mobilité du pathogène, cela suppose des programmes et des activités à la ferme et à
l’échelle du territoire. De plus, pour pouvoir empêcher la propagation du pathogène, il
faut poursuivre la recherche sur le dépistage de ces organes de résistance dans le sol et
dans les tissus végétaux. Enfin, une meilleure connaissance des populations de FOC,

Conclusions et perspectives
48
permettrait de mieux évaluer le risque du flétrissement, de choisir des mesures de
prévention judicieuses et de réduire au minimum l’usage de fongicides chimiques.
En conclusion, l’exploitation des capacités antagonistes des actinomycètes
dans une lutte biologique contre la fusariose du pois chiche semble prometteuse. La
mise au point d’une telle lutte est cependant difficile; nombreux sont les problèmes
qui se posent encore. A la fin de notre étude, nous pouvons conclure que le
flétrissement, dont l’agent causal est F. oxysporum f.sp ciceri constitue une menace
autant plus grave que l’anthracnose pour la culture du pois chiche.
Cette étude a été réalisée afin de mieux connaître le champignon, de savoir
l’identifier, pour ensuite envisager des études de physiopathologie (interaction hôte –
parasite) pour arriver à éliminer ou prévenir la maladie. Pour limiter les problèmes de
flétrissement, il faut éviter les implantations de culture du pois chiche en conditions
limites. Du fait des capacités de conservation de F. oxysporum ciceri et de son survie
dans le sol (chlamydospores en particulier), il existe différentes sources potentielles
de contamination. Or, la pratique d’une rotation ne peut malheureusement pas être le
garant de l’absence de la maladie. Face à des moyens de protection chimique assez
peu efficaces, il apparaît important de connaître le niveau de risques des parcelles
afin, d’éviter d’utiliser des variétés sensibles, si possible d’employer des variétés
résistantes ou tolérantes. Il est important d’éviter d’introduire ce pathogène dans les
sols encore indemnes. Un point important pour les plantations est l’utilisation de
plants sains. Compte tenu de son caractère épidémique, il est souhaitable de ne jamais
laisser ce parasite pénétrer dans les parcelles.
Des mesures prophylactiques s’imposent pour réduire l’inoculum primaire
présent dans le sol:
Réduire au maximum les sources d’infection primaire en détruisant les débris
végétaux, ces mesures prophylactiques sont essentielles.
Adopter une gestion correcte des déchets et des débris végétaux: aucun
déchets ou débris végétaux et même les résidus de cultures ne doit se trouver à
proximité de la parcelle cultivée, car ces résidus se présentent comme une
source majeure d’inoculum.

Conclusions et perspectives
49
Utiliser des plants du pois chiche sains.
Veiller à réaliser une implantation correcte et éviter de planter dans des
parcelles historiquement connues par la maladie.
Ce travail a fait l’objet d’une présentation scientifique lors d’un séminaire national en
phytopathologie, qui s’est déroulé en mai 2010 à Khemis Miliana.

45
Références bibliographiques

Références bibliographique
45
Références bibliographiques
Abawi G.S. and Lorbeer J.W. 1965. Cultural variability and pathogenicity of
Fusarium oxysporum f. sp. cepae. Phytopathology., 55: 1051.
Abawi . GS. and Lorbeer J.W., 1971. Populations of Fusarium oxysporum f. sp.
cepae in organic soils in New York. Phytopathology., 61: 1042-1048.
Abbas A., 1995. Variation in some cultural and physiological characters and
host/pathogen interaction of Fusarium oxysporum f. sp. lentis and inheritance of
resistance to lentil wilt in Syria. Ph. D thesis, Faculty of Agriculture, University of
Alleppo, Syria, 50-68.
Abdelguerfi L. M., Bouzid L., Zine F., Hamdi N., Bouzid H. et Zidouni F., 2001. Evaluation de quelques cultivars locaux de pois chiche dans la région de Bejaia.
Institue nationale de la recherche Agronomique d'Algérie., 9:31-42.
Abdelguerfi L.M., Zine F., Bouzid L., Laib M. et Kadri A., 2000. Caractérisation
préliminaire de quelques cultivars locaux de Cicer arietinum collectés dans la région
de Tizi-Ouzou. Recherche agronomique., 7:51-65.
Aboul-Soud M. A. M., Yun B-W., Harrier L. A. and Loake G. J., 2004.
Transformation of Fusarium oxysporum by particle bombardment and
characterization of the resulting transformants expressing a GFP. Mycopathologia.,
158:475-482.
Adam A., 2008. Elicitation de la résistance systémique induite chez la tomate et le
concombre et activation de la voie de la lipoxygénase par des rhizobactéries non
pathogens. Thèse de Doctoirat en Sciences., Université de liège, 166pp.
Aghighi A. S., Bonjar G. H. S., Rawashdeh R., Batayneh S. and Saadoun I., 2004.
First repport of antifungal spectra of activity of Iranian actinomycetes strains against
Alternaria solani, Alternaria alternate, Fusarium solani, Phytophthora megasperma,
Verticillium dahlia and Saccharomyces cerevisiae. Asian Journal of Plant Sciences.,
4:463-471.
Agrios G. N., 2005. Plant Pathology, 5ème
édition. Department of Plant Pathology
University of Florida; Elsevier Academic Press.pp.948.
Ahlawat I. P. S., 1990. Diagnosis and alleviation of mineral nutrient constraints in
chickpea. In Chickpea in the Nineties. Eds. Vanrheenen H. A. and Saxena M. C.pp.
93-99. ICRISAT Hyderabad, India.

Références bibliographique
46
Alabouvette C. and Couteaudier Y., 1992. Biological control of Fusarium wilts
with non pathogenic Fusaria. In: Tjamos, E. S. (Ed.). Biological Control of Plant
diseases. New York. Plenum Press. pp.415-426.
Alabouvette C., 1986. Fusarium wilt suppressive soil from the Chateauennard
region, a review of a 10 year study. Agronomic., 6:273-284.
Ali M. A., Nawab N.N., Abbas A., Zulkiffal M. and Sajjad M., 2009. Evaluation of
selection criteria in Cicer arietinum L. using correlation coefficients and path analysis.
Australian Journal of Crop Science., 3(2):65-70.
Allen D. J., 1983. The pathology of tropical food legumes. John Wiley and Sons,
New York, 413pp.
Alvarez A. M. and Briner G. C., 1987. Root rot of chickpea caused by Fusarium
solani. Agriculture Technica., 47:78-79.
Alvarez M. Y. and Moreno A., 1984. Identification de hongos causante de pudricion
radicular en garbanzo (Cicer arietinum L.). Agriculture Technica (Chile)., 44 (3):253-
258.
Anderson A. S. and Wellington E. M. H., 2001. The taxonomy of Streptomyces and
related genera. International Journal of Systematic Evolution of Microbiology.,
51:797-814.
Anitha A. and Rebeen M., 2009. In vitro antifungul activity of Streptomyces griseus
against phytopathogenic fungi of tomato field. Academic Journal of Plant Sciences.,
2:119-123.
Ansari S. A., Ginwal H. S, Kumar P. and Singh S., 2001. Ascorbic acid promotes
advantitions rhizogenesis in Teak (Tectona grandis). Indian Foreter., 127 (5):599-602.
Arifuzzaman M., Khatun M. R., and Rahman H., 2010. Isolation and screening of
actinomycetes from Sundarbans soil for antibacterial activity. African Journal of
Biotechnology., 29: 4615-4619.
Armstrong G. M. and Armstrong J. K., 1981. Formae specials and races in
Fusarium oxysporum causins wilt diseases. Pages 391-399 In: Fusarium Diseases.
Biology and Taxonomy. Nelson P. E., Tousson T. A. and Cook R. J. eds Pennsylvania
State University Park., 457pp.

Références bibliographique
47
Arroyo-Garcia R., Cenis J.L., Tello J., Martinez-Zapater JM. and Cifuentes D.,
2003. Genetic relationships among seven specialized forms and AFLPs. Spanish
Journal of Agricultural Research., 3:55-63.
Assigbeste., 1989. Etude de la variabilité spontannée chez Fusarium oxysporum f. sp.
vasinfectum, agent causal de la fusariose de cotonnier. Mémoire de DEA, Université
de Paris XI, INAPG., pp 39.
Athalye M., Lacey J. and Goodfellow M.,1981. Selective isolation and enumeration
of actinomycetes using rifampicin. Applied Journal of Bacteriology., 51:289-297.
Atta H. M., Dabour S. M. and Desoukey S. G., 2009. Sparsomycin Antibiotic
Production by Streptomyces sp. AZ-NIOFD1: Taxonomy, Fermentation, Purification
and Biological activities. American-Eurasian Journal of Agriculture and
Environemental Science., 3: 368-377.
Awam R. I. and Lorbeer J. W., 1986. A sorbose-based selective medium for
enumerating propagules of Fusarium oxysporum f. sp. appii race 2 in organic soil.
Phytopathology., 76:1202-1205.
Awuah R .T. and Lorbeer J.W.,198 8. Nature of cultural variability in Fusarium
oxysporum f. sp. api¿ Race 2. Phytopathology., 78: 385-389.
Aykroyd W. et Doughty J., 1982. Les graines de légumineuses dans l'alimentation
humaine. 2éme
édition, n°20 F.A.O Rome.17p.
Babar B. M., Shah T. M., Abbas G. and Ahsanul haq M., 2009. Genotype X
environment interaction for seed yield in Kabuli chickpea (Cicer arietinum L.)
genotypes developed through mutation breeding. Pakistan Journal of Botany., 4:1883-
1890.
Bacha F. et Ounane S. M., 2003. Etude de l'effet du stress hydrique sur les activités
des enzymes nitrate réductase et nitrogénase de la culture du pois chiche (Cicer
arietinum L.). Institut National de la Recherche agronomique d'Algérie., 13:1111-
1992.
Bais B. S., Singh S. B. and Singh D. V., 1970. Effect of different carbon and
nitrogen sources on the growth and sporulation of Curvularia pallescens. Indian
Phytopath., 23: 511–7
Baker R. and Paulitz T. C., 1996. Theoretical basis for microbial interactions
leading to biological control of soilborne plant pathogens.Pages 50-70 In: Principales
and Practice of managing soilborne plant pathogens. Hall R., ed. The American
Phytopathological Society, St. Paul,MN.

Références bibliographique
48
Bakhsh A., Iqbal S. M. and Haq I.K., 2007. Evolution of chickpea germplasm for
wilt resistance, Pakistan Journal of Botany., 39(2): 583-593.
Barakate M., Ouhdouche Y., Oufdou K. H. and Beaulieu C., 2002.
Characterization of rhizospheric soil Streptomyces from Moroccan habitats and their
antimicrobiol activities. World Journal of Biotechnology., 18: 49-54.
Barik B. P., Tayung K., Jagadev P. N. and Dutta S. K., 2010. Phylogenetic
Placement of an Endophytic Fungus Fusarium oxysporum Isolated from Acorus
calamus Rhizomes with Antimicrobial Activity. E.J.B.S., 2 (1).
Bastide A. et De Méo M., Andriantsao M., Laget M. et Duménil G., 1986.
Isolement et sélection de souches d'actinomycètes productrices de substances
antifongiques de structure non polyénique. MIRCEN Journal., 2: 453-466.
Beckman C. H., 1987. The nature of wilt of Plants.American Phytopathological
Society, St. Paul, Minn.
Bekkar A. A., 2007., Variabilité de la morphologie et du pouvoir pathogène chez
Fusarium oxysporum Schelcht. Emend. Snyd. & Hans. f. sp. Ciceri (Padwick), agent
du flétrissement vasculaire du pois chiche (Cicer arietinum L.). Mémoire de Magister,
Université de Mascara, Mostafa Istambouli, Algérie, pp 92.
Belabid., 2003. La fusariose vasculaire de la lentille (Lens culinaris Med.) dans le
nord-Ouest algérien: Morphologie et diversité génétique chez Fusarium oxysporum
(Schlecht.) Emend. S. and H. f. sp. lentis (Vasud. And Srini) en relation avec la
répartition géographique et le pouvoir pathogène. Thèse de Doctorat. Université
d'Oran, Es-sénia., pp.178.
Ben Freha F., Benyeless M. et Merzoug A., 2010. La fusariose vasculaire du pois
chiche (Cicer arietinum L) dans le nord oust algérien: extension et dégâts. 1er
séminaire National sur la protection des plantes cultivées, Centre Universitaire de
Khemis Miliana, Institut des Sciences de la Nature et de la terre. p.62.
Ben Mbarek K., Boujelben A., Boubaker M. et Hannachi C., 2009. Criblage et
performances agronomiques de 45 génotypes de pois chiche (Cicer arietinum L.)
soumis à un régime hydrique limité. Biotechnologies Agronomique et Sciences
Environnementales., 3:381-393.
Benson D. R. and Silvester W. B., 1993. Biology of Frankia strain, actinomycetes
symbiont of actinorrhizal plants. Microbiological Review., 57:293-319.

Références bibliographique
49
Ben-Yephet Y., and Nelson E. B., 1999. Differential suppression of damping-off
caused by Pythium aphanidermum, P. irregular, and P. myriotylum in composts at
different temperatures. Plant Disease., 83:356-360.
Benzohra I. E., 2009. Contribution à l'étude d'Ascochyta rabiei(Pass.) Labr., agent
causal de l'anthracnose du pois chiche (Cicer arietinum L.): Caractérisation
morphologique et étude du pouvoir pathogène. Thèse de Doctorat.
Berdy J., 2005. Bioactive microbiol metabolites. Journal of Antibiotics., 58:1-26.
Berger J., Abbo S. H. and Turner N. C., 2003. Ecogeography of annual wild Cicer
species: the poor state of the world collection. Crop Sciences., 43:1076-1090.
Bergey D. H., 1989. Bergey's manual of systematic Bacteriology, vol.4. Baltimore,
USA: Williams and Wilkins Company. ISBN0-683-09061-S.
Bertin C., Yang X. and Weston L. A., 2003. The role of root exudates and
allelochemicals in the rhizosphere. Plant and Soil., 256:67-83.
Besri M., Zrouri M. et Beye I., 1984. Appartenance raciale et pathogénique de
quelques isolats de Verticillium dahliae (Kleb) obtenus à partir de tomates résistantes
au Maroc. Phytopathology., 109: 289-294.
Bhatti M.A., and Kraft J.M., 1992. Influence of soil moisture on root rot and wilt of
chickpea. Plant Diseases., 76 :1259-1262.
Bock B., 2009. Cicer arietinum L.Tela botanica, Base de données Nomenclaturale de
la flore de France. BDNFFV4.02.http://www.tela-botanica.org.
Boisson C., 1991. Variabilité de la morphologie et du poivoir pathogène des
champignons appurtenant au genre Fusarium. Mise au point bibliographique,
ORSTOM, 24pp.
Boland G. J., 1997. Stability analysis for evaluating the influence of environment on
chemical and biological control of white mold (Sclerotinia sclerotium) of bean.
Biological Control., 9:7-14.
Booth C., 1971. The genus Fusarium. Commonwealth Mycological Institute, Kew,
Surrey, England., 237 p.
Boudjella H., 1994. Influence des milieu de culture,des antibiotiques et du pré-
traitement des échantillons à la chaleur sur la selection des genres et des espèces
d'actinomycètes rares de quelques sols sahariens. Magister de Microbiologie, ENS de
Kopuba, Algérie.

Références bibliographique
50
Boudjella H., Bouti K., Zitouni A., Mathieu F., Lebrihi A. and Sabaou N., 2006. Taxonomy and chemical characterization of antibiotics of Streptosporangium Sg 10
isolated from a Saharan soil. Microbiology and Research., 161: 288-98.
Boughachiche F., Reghioua S., Oulmi L., Zerizer H., Kitouni M., Boudemagh A.
et Boulahrouf A., 2005. Isolement d'actinomycétales productrices de substances
antimicrobiennes à partir de la Sebkha de Ain Mlila. Sciences et Technologie., 23: 5-
10.
Bouslama M., 1980. Chickpea improvement in Tunisia. In : Proceeding Workshop
Chickpea improvement, ICRISAT, Hyderabad, India, PP. 277-280.
Boutati E. I. and Anaissie E. J., 1997. Fusarium, a significant emerging pathogen in
patients with hematologic malignancy: ten years experience at a cancer center and
implications for management. Blood., 90: 999-1008.
Bouzerzour H., Abbas K., Benmahammed A., 2003. Les céreales, les
légumineuses alimentaires, les plantes fourrageres et Pastorales. Recueil des
Communications Atelier N°3 «Biodiversité Importante pour l’Agriculture» MATE-
GEF/PNUD Projet ALG/97/G31.pp.79.
Bouznad Z., 1989. Contribution à la connaissance du genre Ascochyta chez les
légumineuses en Algérie. Etude biologique, ultrastructure et cytochimique des
relations hote-pathogène chez le couple Ascochyta pisi/Pisium sativum. Thèse de
doctorat en Sciences naturelles; Université Pierre Marie Curie, 190 p.
Bradbury J. F., 1986. Guide to plant pathogenic bacteria. London, CAB
International Mycological Institute Publisher.
Braun P. H., Planquaert P. H., Wery J., 1988. Pois chiche : culture, utilisation;
Brauchure. Ed. ITCF 11p.
Bressan W., 2003. Biological control of Maize seed pathogenic fungi by use of
actinomycetes. Kluwer Academic Publishers Biocontrol., 48:233-240.
Brewer D., 1960. Studies of Ascochyta pisi. Cannadain Journal of Botany., 38:705-
717.
Brown M. E., 1974. Seed and root bacterization. Annual Review of Phytopathology.,
12:181-197.

Références bibliographique
51
Bull C. T., Weller D. M. and Thomashow L. S., 1991. Relationship between root
colonization and suppression of Gaeumannomyces graminis var. tritici by
Pseudomonas fluorescens strain 2-79. Phytopathology., 81:954-959.
Burgess L. W., Nelson P. F. and Summerell B. A., 1989. Variability and stability of
morphological characters of Fusarium oxysporum isolated from soil in Australia.
Mycology., 81:818-822.
Burnett J.H ., 1984. Aspects of Fusarium genetics. In : The applied mycology
ofFusarium. British Mycological Society Symposium, 7, (M.O. MOSS and J.E.
SMITH, ed itors), Cambridge University Press: 39-69.
Bushula V. S., 2008. Native Fusarium species from indigenous Fynbos soils of the
western cap. Master Thesis, University of Stellenbosch.pp.197.
Buxton E. W., 1954. Heterocaryosis and variability in Fusarium oxysporum f. sp.
gladioli (Snyder et Hansen). Journal of Gentics Microbiology., 10: 71-84.
Campbell C. L. and Powell N. T., 1990. Progression of diseases induced by
soilborne pathogens: Tobacco Black Shank. Protection and Ecology., 2:177-182.
Cao L., Qiu Z., You J., Tan H. and Zhou H., 2005. Isolation and characterization of
endophytic Streptomycetes antagonists of Fusarium wilt pathogen from surface-
sterilized banana roots. FEMS Microbiology Letters., 247:147-152.
Carbera de la Colina J., Trapero-Casas A. and Jimenez-Diaz R. M., 1985. Races
of Fusarium oxysporum f. sp. ciceri in Andalucia, Southern Spain. International
chickpea Newsletter., 13: 34-26.
Carlile M. J., 1956. A study of the factors influencing non-genetic variation in a
strain of Fusarium oxysporum. Journal of Genetics and Microbiology., 14:643-654.
Castillo P., Mora-Rodriguez Ma. P., Navas-Cortes J.A. and Jimenez-Diaz R.
M., 1998. Interactions between Pratylenchus thornei and Fusarium oxysporum f. sp.
ciceris on chickpea. Phytopathology., 88: 836-844.
Catwright R. Y., 1978. Use of antibiotics: Antifungals. British Medical Journal., 2:
108-111.
Chabasse D., Bouchara J-P. ,De Gentil L., Baum S., Cimon B. et Penn P., 2002.
Les moisissures d'intérêt médical. Ouvrage réalisé par le laboratoire de parasitologie-
Mycologie du CHU d'Agers-U, rue Larrey 49033 Angers cedex, Bioforma n°25.
Pp.159.

Références bibliographique
52
Chakraborti D., Sarkar A., Gupta S. and Das S., 2006. Small and large scale
genomic DNA isolation protocol for chickpea (Cicer arietinum L.), suitable for
molecular marker and transgenic analyses. African Journal of Biotechnology., 8: 585-
589.
Chauhan, S. K., 1965. The interaction of certain soil conditions in relation to the
occurrence of Fusarium wilt of gram. Indian Jornal of Agriculture., 35: 52
Chen C. Y., Wang Y. H. and Huang C. J., 2004. Enhancement of the antifungal
activity of Bacillus subtilis F29-3 by the chitinase encoded by Bacillus circulans chiA
gene. Canadian Journal of Microbiology.,50: 451-454.
Chérif M., Arfaoui A. and Rhaim A., 2007. Phenolic compounds and their rol in
Bio-control and resistance of chickpea to fungal pathogenic attacks. Tunisian Journal
of Plant protection., 2:7-12.
Cherrab M., 1989. Contribution à l'étude morphologique et biochimique de quelques
forms specials Fusarium oxysporum. Thèse de doctorat 3éme
cycle, Université Cadi
Ayad, Merrakech, Maroc, 104 p.
Chi C. C. and Hanson E. W., 1965. In Vitro effects of Streptomyces rimosus on
some soil-inhabiting pathogenic fungi. Plant Diseases Rep., 49: 159-163.
Choumane W., Winter P., Weigand F. and Kahl G., 2000. Conservation and
variability of sequence-tagged microsatellite sites (STMSs) from chickpea (Cicer
arietinum L.) within the genus Cicer. Theoritical and Applied Genetics., 101:269-
278.
Ciampi L P., Nissen J. M., Venegas E. G., Fuentes R. P., Costa M. L., Schöbitz
R. T., Alvarez E. D., and Alvarado P. A., 2008. Identification of two species of
Fusarium Link that cause wilting of colored callas (Zantedeschia aethiopica L.)
cultivated under greenhouse conditions in Chile. Chilean Journal of Agricultural
Research., 69(4):516-525.
Collin D. P. and Jacobsen B. J., 2003. Optimizing a Bacillus subtilis isolate for
biological control of sugar beet Cercospora leaf spot. Biology and Control., 26 (2):
153-161.
Conrath U., Pieterse C. M. J. and Mauch-Mani B., 2002. Priming in plant–
pathogen interactions. Trends in Plant Science 7: 210–216.

Références bibliographique
53
Coombs J. T., Franco C. M. M. and Loria R., 2002. Complete sequencing and
analysis of pEN2701, a novel 13-kb plasmid from an endophytic Streptomyces sp.,
49(1):86-92.
Covell S., Ellis R.H., Roberts E.H., Summerfield R.J., 1986. The influence of
temperature on seed germination rate in grain legumes .I. Acomparaison of chickpea,
lentil, Soybean and cowpea at constant temperature. Journal of experimental Botany
37:705-715.
Crawford D. I., Lynch J. M., Whipps J. M. and Ousley M. A., 1993. Isolation and
characterization of actinomycete antagonists of a fungal root pathogen. Applied of
Environemental Microbiology., 59:3899-3905.
Cross T., 1982. Invitational ONR lecture. Actinomycetes: A continuing source of
new metabolites. Developments in Industrial Microbiology., 23: 1-20.
Cross T., 1989. The actinomycete. II. Growth and examination of actinomycetes-
some guidelines. In: Williams S. T., Sharpe M. E. and Holt J. G., (eds), Bergey's
manual of Systematic Bacteriology. The Williams and Wilkins Company., Baltimore,
vol. 4.pp. 2340-2345.
Cunnington J., Lindbeck K., Rodney H. and Jones., 2009. Diagnostic methods for
Fusarium wilt of chickpea (Fusarium oxysporum f. sp. ciceris) Padil. Plant
Biosecurity Toolbox page 1-22.
Curl E. A. and Truelove B., 1986. The rhizosphere. Springer-Verlag. Berlin., 288pp.
Da Silva J. C. and Bettiol W., 2005. Potential of Non-Pathogenic Fusarium
oxysporum isolates for control of Fusarium wilt of Tomato. Fitopatologia bras., 30(4):
409-412.
Dastager S.G.1., Li W-J., Dayanand A., Tang S-K, Tian X-P., Zhi X-Y., Xu L-
H. and Jiang C-L., 2006. Seperation, identification and analysis of pigment
(melanin) production in Streptomyces African Journal of Biotechnology., 5 (8): 1131-
1134.
De Boer M. P., Boom F., Kindt J. J. B., Keurentjes I., Van der Sluis., Van Loon
L. C. and Bakker P. A. H. M., 2003. Control of Fusarium wilt of Radish by
combining Pseudomonas putida strains that have different disease-suppressive
mechanisms. Phytopathology., 93:626-632.
Delgado-jarana J., Martinez-Rocha A. L., Roldan-Rodriguez R., Roncero M. I.
G. and Di-Pietro A., 2005. Fusarium oxysporum G-protein B subunit Fgb1 regulates

Références bibliographique
54
hyphal growth, development, and virulence through multiple signalling pathways.
Fungal Genetics and biology.,42: 61-72.
Demain A. L. and Sanchez S., 2009. Microbiol drug discovery: 80 years of progress.
Journal of Antibiotics., 62:5-16.
Demain A. L. and Solomon N. A., 1985. Biology of industrial microorganisms. The
Benjamin/cummings publishing company, INC. California.
Desjardin V., 2002. Réduction du chrome IV par la souche Streptomyces
thermocarboxydus NH50 isolée à partir d'un sol pollué. Thèse de Doctorat, Institue
Nationale des Sciences Appliquée de Lyon, pp. 236.
Desjardins A. E., Hohn T. M., and McCormick S. P., 1992. Effect of gene
disruption of trichodiene synthase on the virulence of Gibberella pulicaris. Mol.
Plant-Microbe Interact. 5:214–222.
Dhedhi B. M., Gupta O., Patel V. A. and Gupta O., 1990. Antagonistic effect of
microorganisms to Fusarium oxysporum f. sp. ciceri. Indian Paecilomyces lilacinus
with oil cakes in the control of root rot of mungbean. Tropical Sciences., 35(3):294-
299.
Dhingra O. D. and Sinclair J. B., 1995. Basic Plant Pathology Methods. CRC Press
USA, pp: 287-296,390-391.
Di Pietro A., Garcia-Maceira F. L., Huertas-Gonzàlez D., Ruiz-Roldan M. C.,
Carcuel Z., Barbieri A.S. and Roncero I.G., 1998. Endopolygalacturonase PG1 in
different Formae Speciales of Fusarium oxysporum. Applied and envirronmental
microbiology., 64: 1967-1971.
Dick A. J. and Elad Y., 1999. Comparison of antagonists of Botrytis cinerea in
greenhouse cucumber and tomato under different climatic conditions. European
Journal of Plant Pathology., 105:123-137.
Dickenson S., 1932. The nature of saltation in Fusarium and Helminthosporium.
Minn. Agr. Exp. Sta. Tech. Bull., 88.
Dijksterhuis J., Sanders M., Gorris L.G. M., Smid E. J., 1999. Antibiosis plays a
role in the context of direct interaction during antagonism of Paenibacillus polymyxa
toward Fusarium oxysporum. Journal Applied of Microbiology., 86:13-21.

Références bibliographique
55
Di-Primo P., Cartia G. and Katan T., 2001. Vegetative compatibility and
heterokaryon stability in Fusarium oxysporum f. sp. radicis-lycopersici from Italy.
Plant Pathology., 50:371- 382.
Djerbi M., Aouad L., Filali H., Saaidi M., Chtioui A., Sedra M. S., Allaoui M.,
Hamdaoui T. and Ouhdouche M., 1986. Preliminary results of selection of high
quality bayoud-resistant clones among natural date palm population in Morroco. In
proceedings of the second symposium on the date Palm, Saoudi Arabia,383-399.
Djerbi M., Mlaiki A. and Booslarna M., 1978. Food legumes diseases in North
Africa. In: Hawtin G. C. et Chancellor Reds G. J. food legume improvement and
development . proceeding of workschop held at the university of Aleppo, Syria .
ICARDA et IDRC;pp: 103-105.
Djerbi M., Sedra M. H. and Idrissi M. A., 1985. Caractéristiques culturales et
identification du Fusarium oxysporum f. sp. albedinis, agent causal du bayoud.
Institut National de Recherche Agronomique; Tunisie., 58:1-8.
Doohan F. M., Parry D.W., Jenkinson P. and Nicholson P., 1998. The use of
species-specific PCR-based assays to analyse Fusarium ear blight of wheat. Plant
Pathology., 47: 197–205.
Doumbou C. L., Akimov V. and Beaulieu C., 1998. Selection and characterization
of microorganisms utilizing Thaxtomin A, a phytotoxin produced by Streptomyces
scabies. Applied Environement Microbiological., 64: 4313-4316.
Doumbou C. L., Salove M. K. H., Crawford D. L. and Beaulieu., 2002.
Actinomycetes, promising tools to control plant diseases and to promote plant growth.
Phytoprotection., 82:85-102.
Dubey R.C., Kumar H., and Pandey R. R., 2009. Fungistatic effect of Neem Extracts
on Growth and sclerotial survival of Macrophomina phaseolina in vitro. Journal of
American Sciences.,5(5): 17-24.
Dubey S. C. and Singh B., 2004. Reaction of chickpea genotypes against Fusarium
osysporum f. sp. ciceri causing vascular wilt. Indian Phytopathology., 57: 233.
Dubey S. C., Suresh M. and Singh B., 2007. Evaluation of Trichoderma species
against Fusarium oxysporum f. sp. ciceris for integrated management of chickpea
wilt. Biological Control., 40:118-127.
Duijff B. J., Meijer J. W., Bakker P. A. H. M. and Schipper B., 1993. Siderophore
mediated competition for iron and induced resistance in the suppression of Fusarium

Références bibliographique
56
wilt of carnation by fluorescent Pseudomonas spp. Neth Journal of Plant Pathology.,
99:277-291.
Duijff B. J., Recorbet G., Bakker P. A. H. M., Loper J. E., and Lemanceau P.,
1999. Microbial antagonism at the root level is involved in the suppression of
Fusarium wilt by the combination of non pathogenic Fusarium oxysporum Fo47 and
Pseudomonas putida WCS358. Phytopatology., 89:1073-1079.
Duschak., 1871. Zur Botanik des Talmud .I. Neur, Pest :105-106 In Singh ,F. et
Diwakar, B.1995.Chickpea Botany and Production Practices. Skill Development
Series n°.16; ICRISAT.
Edel V., Steinberg C., Gautheron N., Recorbet G. and Alabouvette C., 1995.
Comparison of 3 molecular methods for the characterization of Fusarium oxysporum
strains. Phytopathology., 85:579-585.
Edwards S. G., McKay T. and Seddon, B., 1994. Interaction of Bacillus species
with phytopathogenic fungi-Methods of analysis and manipulation for biocontrol
purposes. In: Blackman, J. P. and Williamson, B.(eds). Ecology of Plant
Pathogens.pp.101-118. CAB International,Wallingford, Oxon, UK.
El Guilli M., 1987. Production, contrôle, qualité, commercialisation des semences de
petits pois et étude de quelques aspects phytosanitaires de la culture au Maroc.
Mémoire de 3éme
cycle. Institut Agronomique Hassan II, Rabat, Maroc.
Elad Y. and Baker R., 1985. The role of competition for iron and carbon in
suppression of chlamydospore germination of Fusarium spp. by Pseudomonas spp.
Phytopathology., 75:1053-1059.
Elander R. P., 1987. Microbial screening, selection and strain improvement. In:
Basic Biotechnology,pp.217-251. Edited by J. Bu'Lock and B. Kristiansen. London:
Academic Press.
Elani S., 1968. The cytogenetics of the conidium in Microsporum gypseum and of pleomorphism and the dual phenomenon in fungi. Mycologia., 60: 999-1015.
El-Aoufir A., 2001. Etude du flétrissement vasculaire du pois chiche (Cicer
arietinum) cause par le Fusarium oxysporum f. sp. ciceri. Evaluation de la fiabilité de
l'analyse isoenzymatique et de la compatibilité végétative pour la caractérisation des
races physiologiques. Thèse de Doctorat, Université Laval, P161.

Références bibliographique
57
Elson M. K., Schisler D. A. and Bothast R. J., 1997. Selection of microorganisms
for biological control of Silver scurf (Helminthosporium solani) of potato tubers.
Plant Disease., 81:647-652.
El-Tarabily K. A. and Sivasithamparam K., 2006. Non-Streptomycete
actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant
growth promoters. Soil Biology and Biochemestry., 38: 1505-1520.
El-Tarabily K. A., 2003. An endophytic chitinase-producing isolate of Actinoplanes
missouriensis, with potential for biological control of lupine caused by Plectosporium
tabacinum. Australian Journal of Botany., 51:257-266.
El-Tarabily K. A., 2006. Rhizosphere competent isolates of Streptomycete and non
Streptomycete actinomycetes capable of producing cell-wall degrading anzymes to
control Pythium aphanidermatum damping off disease of cucumber. Cannadian
Journal of Botany., 84: in Press.
El-Tarabily K. A., Hardy G. E. St. J., Sivasithamparam K., Hussein A. M. and
Kurtboke I. D., 1997. The potential for the biological control of cavity spot disease
of carrots caused by Pythium coloratum by Streptomycetes and non-streptomycete
actinomycetes in Western Australia. New Phytologist., 137:495-507.
El-Tarabily K. A., Soliman M. H., Nassar A. H., Al-Hassani H. A.,
Sivasithamparam K., Mc Kenna F. and Hardy G. E. St. J., 2000. Biological
controlof Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant
Pathology., 49:573-583.
El-Tarabily K. A., Sykes M. L., kurtboke I. D., Hardy G. E. S. J., Barbosa A. M.
and Dekker R. F. H., 1996. Synergistic effects of a cellulose-producing
Micromonospora carbonacea and an antibiotic producing Streptomyces violascens on
the suppression of Phytophthora cinnamomi root-rot of Banksia grandis. Canadian
Journal of Botany., 74:618-624.
Erler F., Ceylan F., Erdemir T., and Toker C., 2009. Preliminary results on
evaluation of chickpea, Cicer arietinum, genotypes for resistance to the pulse beetle,
Callosobruchus maculatus. Journal of Insect Science., 9:58-72.
Errakhi R., Bouteau F., Lebrihi A. and Barakate M., 2007. Evidences of
biological control capacities of Streptomyces spp. against Sclerotium rolfsii
responsible for damping-off disease in sugar beet (Beta vulgaris L.). World Journal of
Microbiology and Biotechnology., 23:1503-1509.

Références bibliographique
58
Erwin D. C., 1958. Fusarium oxysporum f. sp. ciceri, incitant of Fusarium wilt of
Cicer arietinum. Phytopathology., 48:498-501.
Eunice J. A. and Prosser J. I., 1983. Mycelial growth and branching of
Streptomyces coelicolor A3 (2) on solid medium. Journal of genetic microbiology.,
129:2029-2036.
Fahim N. M., Osman A. R., El-Attar A. H. and Mabrouk M. S. M., 1987. Root rot
of common bean. Egyptian Journal of Phytopathology., 19:71-83.
FAOSTAT data., 2005. Food and Agriculture Organisation odf The United Nation,
Rome, Italy., http://faostat.fao.org.
FAOSTAT data., 2007. Food and Agriculture Organisation odf The United Nation,
http://faostat.fao.org/site/567/default.aspx.
FAOSTAT data., 2010. Food and Agriculture Organisation odf The United Nation,
http://faostat.fao.org/site/567/Desktop default.aspx.
Farooq S., Iqbal S. H. M. and Abdul Rauf C. H., 2005. Physiological studies of
Fusarium oxysporum f. sp. ciceri. International Journal of Agriculture and Biology.,
275-277.
Fisher N. L., Burgess N. W., Toussoun T.A. and Nelson P. A., 1982. Carnation
leaves as a substrate and for preserving cultures of Fusarium species.
Phytopathology., 72 :151-153.
Flandez-Galvez H., Ford R., Pang E. C. K. and Taylor P. W. J., 2003. An
interspecific linkage map of the chickpea (Cicer arietinum l.) genome based on
sequence tagged microsatellite site and resistance gene analog markers. Theoritical
Applied in Genetics., 1447-1456.
Flor H., 1971. Current status of the gene for gene concept. Annuals Review of
Phytopathology., 9: 275-296.
Follin J.C . et E. Lavell,1 966. Variations chez le Fusarium oxysporum f. cubense
(agent causal de la maladie de Panama du bananier). Fruits., 21: 261-268.
Foo E. L. and Varma A.K., 1976. Inhibitory effect of Streptomyces antibioticus and
other microorganisms on Rhizobium. Folia microbial., 21:315-319.
Fourati-Ben Fguira L., Serge R., Mehdi M., Lotfi M. and Hartmut L., 2005.
Purification and structure elucidation of antifungal and antibacterial activities of

Références bibliographique
59
newly isolated Streptomyces sp. strain US80. Research in Microbiology., 156:341-
347.
Franceschi V. R., Kroken P., Christiansen E., Krekling T., 2005. Anatomical and
chemical defenses of conifer bark against bark beetles and other pests. New
Phytopathology., 167:353-375.
Fravel D., 1988. Role of antibiosis in the biocontrol of plant diseases. Annual
Review of Phytopathology.,26: 75-91.
Garcia-Pedrajas M. D., Bainbridge B. W., Heale J. B., Perez-Artes E. and
Jimenez-Diaz R. M., 1999. A simple PCR-based method for the detection of the
chickpea wilt pathogen Fusarium oxysporum f. sp. ciceris in artificial and natural
soils. European Journal of Plant Pathology., 105: 251-259.
Gaur P.M., Pande S., Upadhyaya H.D. and Rao B.V., 2006. Extra-large Kabuli
chickpea with high resistance to Fusarium wilt. International chickpea and Pigeonpea
Newsletter., 13: 5-6.
Geigy., 1990. Préparation des réactifs et des prélèvements. 8éme édition, vol, 3,
Novartis Pharma A G, Basel, Suisse.
Gerlach et Nirenberg H. I. 1982. The Genus Fusarium-a Pictorial Atlas. Berlin-
Dahlem: Paul Parey. 406 p.
Germida J.J., Sicilliano S. D. et al., 1998. Diversity of root associated with field
grown canoi (Brasica napus L.) and wheat (Triticum aestivum L.). FEMS of
Microbiology and Ecology., 26(1):43-50.
Getha K., Vikineswary S., Wong W. H., Seki T., Ward A. and Goodfellow M.,
2005. Evaluation of Streptomyces sp. strain g10 for suppression of Fusarium wilt and
rhizosphere colonization in pot-grown banana plantlets. Journal of Industrial
Microbiology and Biotechnology., 32:24-32.
Girrard C., 1985. L'installation du pois chiche du printemps. In Bulltin FNAMS
semences: 25-27.
Goodfellow M. and Williams S. T., 1983. Ecology of actinomycetes. Annuals
Review of Microbiology., 37:189-216.
Goodfellow M., and Simpson K. E., 1987. Ecology of Streptomycetes. Front.
Applied in Microbiol., 2: 97-125.

Références bibliographique
60
Gordon T. R. and Martyn R. D., 1997. The evolutionary biology of Fusarium
oxysporum. Annual Review of Phytopathology., 35:111-128.
Gottlieb D., 1973. General consisderation and implication of the Actinomycetales. In:
Actinomycetales characteristics and practical importance. Edited by G. Sykes and F.
A. Skinner. Academic Press, London, New York.
Grabley S and Thiericke R., 1999. Drug Discovery from nature. Springer London,
pp: 5-7.
Grewal J. S., 1982. Control of important seed borne pathogens of chickpeas. Indian
Journal of Genetics and Plant Breeding., 42 :393-398.
Grewal J.S. and Pal M., 1970. Fungal diseases of gram and arhar. Proc. IV Annual
Workshop of Pulse Crops, p. 168. PAU, Ludhiana.
Guarro J., Gene J. and Stchigel A. M., 1999. Developments in fungal taxonomy.
Clinic Microbiology Review., 12:454-500.
Gupta O. M., Kotasthane S. R. and Khare M. M., 1986. Fusarium wilt of chickpea
(Cicer arietinum). Agricultural Review., 7:87-97.
Gupta O. M., Kotasthane S. R. and Khare M. M., 1987. Factors influencing
epidemiology of vascular wilt of chickpea. Proceeding National Academic Sciences,
India., 57: 86-91.
Gupte T. E. and Naik S. R., 1999. Isolation and taxonomic and fermentation studies
on a new strain of Streptomyces arena var ukainiana prodicing a tetrene antibiotic.
World Journal of Microbiology and Biotechnology., 15:545-552.
Gwata E. T., Silim S. N. and Mgonja M., 2006. Impact of a New Source of
Resistance to Fusarium Wilt in Pigeonpea. Journal of Phytopathology., 154: 62–64.
Hale M., Moore L. D. and Griffin G. J., 1978. Root exudates and exudation., In:
Dommergues Y. R., Krupa S. V., editors. Interactions between non pathogenic soil
microorganisms and plants. Amesterdam, Oxford, New York: Elsevier Scientific
publishing company, p 163-203.
Halila H. M. and Harrabi M. M., 1990. Breeding for dual resistance to Ascochyta
and wilt diseases in chickpea. Seminaire Méditeranéen n° 9, 11-13 july 1988,
Zaragoza, Spain.
Halila H. M., Grindley H. E. and Houdiard P., 1984. Sources of resistance to
Fusarium wilt in Kbuli chickpeas. International Chickpea Newsletter., 10:13-14.

Références bibliographique
61
Halila I., Cobos M. J., Rubio J. and Milan., 2009. Tagging and mapping a second
resistance gene for Fusarium wilt race 0 in chickpea. European Journal of pathology.,
124:87-92.
Halila M. H. and Strange R. N., 1996. Identification of the causal agent of wilt of
chickpea in Tunisia as Fusarium oxysporum f. sp. ciceri race 0. Phytopathologia
Mediterranea., 35: 67-74.
Hamdi Y. A., Ahmed D. and Al-Tai A. M., 1980. Genera and species of
actinomyceters isolated from Iraqi soil. Egyptian Journal of Microbiology., 15:7-22.
Hamilton-Miller J. M. T., 1973. Chemistry and biology of the polyene macrolide
antibiotics. Bacteriological Reviews ., 37: 166-196.
Hannusch D. J. and Boland G. J., 1996. Interactions of air temperature, relative
humidity and biological agents on grey mold of bean. European Journal of Plant
Pathology., 102:133-142.
Harlan., 1992. Crops and Man. Second Edition. Madison (WI): American Society of
Agronomy/Crop Science Society of America., Wisconsi,pp.63-262.
Harrabi M. M., Halila M. H. and Ben Brahim M., 1985. Evaluating sources of
resistance to Fusarium and Verticilium wilts in chickpea. Arabe Journal of Palm
Production., 3:91-93.
Harris A. R., Siwek K. and Wiseman B. M., 1997. Interactions between damping-
off fungi, antagonists and Capsicum seedlings. Applied Soil Ecology., 6:251-263.
Harris D. R., 1998. The spread of Neolithic agriculture from the levant to western
central Asia. In: Damania A. B., Valkoun J., Willocox G and Qualset,C. O. (eds). The
origine of Agriculture and Crop Domestication. ICARDA, Allepo, Syria, pp.54-64.
Harvey, I., Cormier, Y., Beaulieu, C., Akimov, V. N., Me´ riaux, A. & Duchaine,
C. (2001). Random amplified ribosomal DNA restriction analysis for rapid
identification of thermophilic actinomycete-like bacteria involved in hypersensitivity
pneumonitis. Syst Appl Microbiol 24, 277–284.
Hassan F., 2006. Heterologous expression of a recombinant chitinase from
Streptomyces olivaceoviridis ATCC 11238 in Transgénic Pea (Pisum sativum L.)
Doctorat thesis, University of Damas, Syria, pp.150.

Références bibliographique
62
Hatsh . D., 2004. Interaction hôte/pathogène : étude du modèle Humulus lupulus /
Fusarium graminearum. Identification, génomique et transcriptomique du pathogène.
Université Louis Pasteur Strasbourg I. pp.183.
Haware M. P., Nene Y. L. and Rajeshwari R., 1978. Eradication of Fusarium
oxysporum f. sp. ciceri transmitted in chickpea seed. Phytopathology., 68:1364-1367.
Haware M. P. and Nene Y. L., 1982. Races of Fusarium oxysporum f. sp. ciceri.
Plant Diseases., 66:809-810.
Haware M. p., 1988. Fusarium wilt and other important disease of chickpea in the
mediterraneen area. Proceedings of international Worcshop on present status anfd
future prospects on chickpea crop production and improvement in the Mediterranean
countries. CIHEAM/EEC, Agrimed/ ICARDA.11-13, Zaragoza, Spain.
Haware M. P., 1990. Fusarium wilt and other important diseases of chickpea in the
Mediterranean area. Option Mediterranean, series des siminaires.,9: 163-166.
Haware M. P., Jimenez-Diaz R. M., Amin K. S., Phillips J. C. and Halila H.,
1990. Integrated management of wilt and root rots of chickpea. In: Chickpea in the
nineties: Proc 2nd
Int. Workshop chickpea improvement. ICRISAT Center,
Patancheru, India ICRISAT Publication pp, 129-133.
Haware M. P., Nene Y. L. and Mathur S. B., 1986. Seed borne diseases of
chickpea. Technical Bulletin from the Danish government institute of Seed Pathology
for Developing Countries, Copenhagen Denmark.,1: 14.
Haware M. P., Nene Y. L. and Natarajan M., 1996. The survival of Fusarium
oxysporum f. sp. ciceri in the soil in the absebce of chickpea. Phytopathology
Mediteranean., 35: 9-12.
Haware M. P., Nene Y. L. and Rao N., 1981. Additional sources of resistance to
wilt and roots of chickpeas. International Chickpea Newsletter., 4:18.
Haware M. P.. and Nene Y. L., 1980. Sources of resistance to wilt and root rots of
chickpea. International Chickpea Newsletter., 4: 18.
Haware M.P. and Nene Y. L., 1980. Sources of resistance to wilt and root rots of
chickpea. International chickpea Newsletter., 4: 18.
Hayes W.A., 1978. Nutrition, substrates and principles of disease control. In: The
Biology and Cultivation of Edible Mushrooms, pp. 202–22. Academic press. New
York

Références bibliographique
63
Henni J., Boisson C et Geiger J. P., 1994., Variabilité de la morphologie chez
Fusarium oxysporum F. sp. lycopercisi. Phytopathology mediterranean., 33: 51-58.
Hervas A., Landa B. and Jimenez-Diaz R. M., 1997. Influence of chickpea
genotype and Bacillus sp. on protection from Fusarium wilt by seed treatment with
nonpathogenic Fusarium oxysporum. European Journal of Plant Pathology., 103:631-
642.
Hervas A., Landa B., Datnoff L. E. and Jimenez-Diaz R. M., 1998. Effects of
commercial and indigenous microorganisms on Fusarium developpement in chickpea.
Biological Control., 13:166-176.
Hervas A., Trapero-Casas J. L. and Jimenez-Diaz R. M., 1995. Induced resistance
against Fusarium wilt of chickpea by non pathogenic races of Fusarium oxysporum f.
sp. ciceris and non pathogenic isolates of Fusarium oxysporum. Plant Diseas.,
79:1110-1116.
Hirsch C. F. and Christensen D. L., 1983. Novel Method for selective isolation of
Actinomycetes. Applied and Environemental Microbiology., 46: 925-929.
Hjeljord L. G., Stensvand A. and Tronsmo A., 2000. Effect of temperature and
nutrient stress on the capacity of commercial Trichoderma products to control
Botrytis cinerea and Mucor piriformis in greenhouse strawberries. Biological control.,
19(2):149-160.
Honnareddy N. and Dubey SC., 2006. Pathogenic and molecular characterization
of Indian isolates of Fusarium oxysporum f. sp. ciceris causing chickpea wilt. Current
science., 91(5): 611-666.
Hussain A., Iqbal S. M., Ayub N. and Haqqani A. M., 2003. Physiological studies
of Sclerotium rolfsii. Pakistan Journal of Plant Pathology. 2: 102–6
Hussain H. A., Ibrahim M. A. K., Mahmood M. J. and Al-Hussainy W. A., 1986.
Temperature-dependent production of two macrolide-like antibiotics produced by an
actinomycete (isolate 69B200
). MIRCEN Journal., 2:319-326.
Ikramul H. and Farhat F. J., 1992. Screening of chickpea lines in the wilt sick plot
and effect of environmental temperature on wilt incidence. Proceedings of
COMSTECH-NIAB , International Workshop on Agroclimatology, pests and diseases
and their control, Faisalabad, Pakistan., 21-26.

Références bibliographique
64
Ilic S. B., Konstantinovic S. S., Todorovic Z. B., Lazic ML, Veljkovic V. B.,
Jokovic N. and Radovanovic B. C., 2007. Characterization and antimicrobial
activity of the bioactive metabolites in Streptomycete isolates. Microbiology., 76:421-
428.
Iruela M., Rubio J., Cubero J. I. and Millan T. G., 2002. Phylogenetic analysis in
the genus Cicer and cultivated chickpea using RADP and ISSR markers. Theoritical
Applied in Genetics., 104:643-651.
Irum M., 2007. Comparison of phytochemical and chemical control of Fusarium
oxysporum f. sp. ciceri. Mycopathology., 5 (2):107-110.
Iwai Y. and Omura S., 1992. Cultural conditions for screening of new antibiotics.
Journal of Antibiotics., 35: 123–141
Jain P. K. and Jain P. C., 2007. Isolation,characterization and antifungal activity of
Streptomyces sampsonii GS 1322. Indian Journal of Experiment Biology., 45:203-
206.
Jalali B. L. and Chand H., 1992. Chickpea wilt. In: plant Diseases of International
Importance. Vol.1 Diseases of cereals and Pulses (eds Singh, U. S. et al.,), Prentice
Hall, N. J,1992, pp. 420-444.
Jayasinghe D. B. A. T. and Parkinson D., 2008. Actinomycetes as antagonists of
litter decomposer fungi. Applied in Soil Ecology., 38:109-118.
Jimenez-Diaz R. M and Trapero-Casas A., 1990. Improvement of chickpea
resistance to wilt and root rot diseases. Options Mediterraneennes. Serie Seminaires 9:
65–72.
Jimenez-Diaz R. M. and Trapero-Casa A., 1988. Improvement of chickpea
resistance to wilt and root rot diseases. In: Proceeding on present status and future
prospects of chickpea crop production and improvement in the Mediterranean
countries,11-13 july, Zagaroza, Spain.
Jimenez-Diaz R. M., Basallote U. J. and Rapoport H., 1989. Colonization and
pathogenesis in chickpea infected by races of Fusarium oxysporum f. sp. ciceri.
Vascular Diseases of Plants., 28:113-918.
Jimenez-Diaz R. M., Crino P., Halila M. H., Masconi C. and Trapero-Casas A.
T.,1993. Screening for resistance to Fusarium wilt and Ascochyta blight in chickpea.
In: Singh K. B. and Saxena MC (eds) Breeding for stress tolerance in cool-season
food legumes. John Wiley and Sons, New York, pp 77-95.

Références bibliographique
65
Jimenez-Diaz R. M., Porta-Puglia A. and Tivoli B., 1998. New approaches in the
integrated management of legume diseases: Toward sustainable crop health. Pages 89
- 93. In: European Conference of Grains legumes, 3 ed. Opportunities for high
quality, Healthy and added-value crops to Meet European Demands. European
Association for grain legumes, ed. Valladolid, Spain.
Jimenez-Gasco M. D. M., Milgroom M. G. and Jimenez-Diaz R. M., 2004.
Stepwise evolution of races in Fusarium oxysporum f. sp. ciceris inferred from
fingerprinting with repetitive DNA sequences. Phytopathology., 94:228-235.
Jimenez-Gasco MM, Milgroom MG, Jimenez-Diaz RM., 2002. Gene genealogies
support Fusarium oxysporum f. sp. ciceris as a monophylectic groupe. Plant
pathology., 51,72-77.
Jiminez-Diaz R. M., Singh K. B. and Trapero-Casas A., 1991. Resistance in
Kabuli chickpea to Fusarium wilt. Plant Diseases., 75:914-918.
Jiminez-Gasco M. M. and Jiminez-Diaz R. M., 2003. Development of a specific
polymerase chain reaction-based assay for the identification of Fusarium oxysporum
f. sp. ciceris and its pathogenic races 0, 1A,5, and 6. The American Phytopathological
Society., 2: 200-209.
Johnson K. B., 1994. Dose-response relationships and inundative biological control.
Phytopathology., 84:780-784.
Kaboré B. K., Couture L., Dostaler D. et Bernier L., 2001. Variabilité phénétique
du Colletotricum graminicola du Sorgho. Cannadian Journal of Plant Pathology.,
23:138-145.
Kainer W. J. and Hannan R. M., 1983. Etiology and control of seed decay and
preemergence damping-off of chickpea by Pythium ultimum. Plant Diseases.,67 :77-
81.
Kaiser W. J., Alcala-Jimenez A. R., Hervas-Vergas A., Trapero-Casas J. L. and
Jimenez-Diaz R. M., 1994. Screening of wild Cicer species for resistance to races 0
and 5 of Fusarium oxysporum f. sp. ciceri. Plant Diseases., 78:962-967.
Kalakuskii L. V. and Sharaya L. S., 1990. Actinomycetes and higher plants. Usp
Mikrobiology., 2:26-65.

Références bibliographique
66
Kang M. J., Strap J. L. and Crawford D. L., 2010. Isolation and characterization
of potent antifungal strains of the Streptomyces violaceusniger clade active against
Candida albicans. Journal of Microbiology and Biotechnology., 37:35-41.
Kathiresan K., Balagurunathan R., Masilamin Selvam M., 2005. Fungicidal
activity of marine Actinomycetes against phytopathogenic fungi. Indian Journal of
Biotechnology., 4: 271-276.
Kaur N. P. and Mukhopadhayay A. N., 1992. Integrated control of chickpea wilt
complex by Trichoderma spp. and chemical methods in India. Tropical Pest
Management., 38:372-375.
Kaur R., Kaur J., Singh R. S. and Alabouvette C., 2007. Biological control of
Fusarium oxysporum f. sp. ciceri by nonpathogenic Fusarium and Fluorescent
Pseudomonas. International Journal of Botany., 3(1):114-117.
Kavitha A., Prabhakar P., Vijayalakshmi M. and Venkateswarlu Y., 2010.
Purification and biological evaluation of the metabolites produced by Streptomyces
sp. TK-VL-333. Research in Microbiology.,1-11.
Kazan K. and Muehlbauer F. J., 1991. Allozyme variation and phylogeny in annual
species of Cicer (Leguminosae). Plant Systematic Evolution.,175: 11-21.
Kazmar E. R., Goodman R. M., GGrau C. R., Johnson D. W., Nordheim E. V.,
Undersander D. J. and Handelsman J., 2000. Regression analyses for evaluating
the influence of Bacillus cereus on alfalfa yield inder variable disease intensity.
Phytopathology., 90:657-665.
Keller S. E., Sullivan T. M. and Chirtel S., 1997. Factors affecting the growth of
Fusarium proliferatum and the production of fumoisin B1: Oxygen and PH. Journal of
Indian Microbiology and Biotechnology., 19:305-309.
Kelly A., Alcala-Jimenez A. R., Bainbridge B. W., Heale J. B., Perez-Artes E.
and Jimenez-Diaz R. M., 1994. Use of genetic fingerprinting and Random Amplified
Polymorphic DNA to Characterize Pathotypes of Fusarium oxysporum f. sp. ciceris
Infecting Chickpea. The American Phytopathological Society., 84:1293-1298.
Keulen Van . G., Alderson J., White J. and Sawers R. G., 2005. Nitrate respiration
in the actinomycete Streptomyces coelicolor. Biochemical Society Transactions.,
33:210-212.
Keuls M., 1952. The use of the studentized range in connection with an analysis of
variance. Euphytica., 1:112-122.

Références bibliographique
67
Khamna S., Yokota A., Peberdy J. F. and Lumyong S., 2009. Antifungal activity
of Streptomyces spp. Isolated from rhizosphere of Thai medicinal plants.
International Journal of Integrative Biology., 6:143-147.
Khan H. R., Mc Donald G. K. and Rengel Z., 1998. Assessement of the Zn status
of chickpea by plant analysis. Plant and soil 198:1-9.
Khan H., Zeb A., Ali Z. and Shah S. M., 2009. Impact of five insecticides on
chickpea (Cicer arietinum L.) nodulation, yield and nitrogen fixing rhizospheric
bacteria. Soil and environement., 1: 56-59.
Khan N. J., Filonow A. B. and Singleton L. L., 1997. Augmentation of soil with
sporangia of Actinoplanes spp. For biological control of Pythium damping-off.
Biocontrol Science and Technology., 7:11-22.
Khanzada S. A., Iqbal S. M. and Haqqani A.M. 2003. Physiological studies on
Macrophomina phaseolina. Mycolpathology., 1: 31–4.
Kherbanda P. D. and Brewer C. C., 1980. Cultural and pathogenic variability
among isolates of Ascochyta fabea. Cannadian Journal of Plant Pathology., 2:139-
142.
Kitouni M., 2007. Isolement de bactéries Actinomycetales productrices
d'antibiotiques à partir d'écosystèmes extrême; identification moléculaire des souches
actives et caractérisation préliminaire des substances élaborées. Thèse de Doctora,
Université Mentouri Constantine; pp.175.
Klein K. K. and Correll J. C., 2001. Vegetative compatibility group diversity in
Fusarium. In Summerfield B. A. and Leslie J. F., Backhouse D, Dryden W. L. and
Burgess L.W. (eds). Fusarium, Pau,E. Nelson Memorial Symposium. ASP Press: St.
Paul, MN, USA, pp. 88:83-96.
Kobayashi G. S. and Medoff G., 1977. Antifungal agents. Recent developments.
Annual Review of Microbiology., 31: 291-308.
Kohl J., Lombaer-van der plas C. H., Molhoek W. M. L., Kessel G. J. and
Goosen-van der Geijn H. M., 1999. Competitive ability of the antagonists
Ulocladium atrum and Gliocladium roseum at temperatures favourable for Botrytis
spp. development. Biocontrol., 44:329-346.

Références bibliographique
68
Kraft J. M. Haware M. P., Jiménez-Díaz R. M., Bayaa B., Harrabi M., 1994.
Screening techniques and sources of resistance to root rots and wilts in cool season
food legumes. Euphytica, 73: 27–39.
Kraft J. M., 1969. Chickpea, a new host of Fusarium solani f. so. Pisi. Plant Disease
Report., 53:110-111.
Kumar B. S. D., 1999. Fusarial wilt suppression and crop improvement through two
rhizobacterial strains in chick pea growing in soils infected with Fusarium oxysporum
f. sp. ciceris. Biology of Fertile soils., 29:87-91.
Kumar D. B. S. 1998. Disease suppression and crop improvement through
fluorescent pesudomonads isolated from cultivated soils. World Journal of
Microbiology and Biotechnology., 14 (5): 735-741.
Kumar D. B. S. and Dube H. C. 1992. Seed bacterization with a fluorescent
Pseudomonas for enhanced plant growth, yield and disease control. Soil Biology and
Biochemestry., 24 (6): 539-542.
Kumar D. B. S., 1990. Studies on rhizobacteria of some crop plants with special
reference to plant growth-promoting rhizobacteria (PGPR).Ph.D thesis, Bhavnagar
University, India,pp. 250.
Kumar D. B. S., 1996. Crop improvement and disease suppression by a Bacillus sp.
SR 2 from pea nut rhizosphere. Indian Journal of Experiment Biology., 34 (8): 794-
798.
Kumar D. B. S., 1999. Fusarial wilt suppression and crop improvement through two
rhizobacterial strains in chickpea growing in soils infested with Fusarium oxysporum
f. sp. ciceris. Biology and Fertile soils., 29:87-91.
Kumar J. and Abbo S., 2001. Genetics of flowering time in chickpea and its bearing
on productivity in semiarid environments. Advances Agronomique., 72:107-138.
Kurylowicz M., Paszkiewicz A., Szulga T., Woznicka W. and Karzatkowski W.,
1975. Classification of Streptomyces by different numerical methods. Postepy Hig.
Med. Dosw., 29:281-355.
Labdi M., 1990. Chickpea in Algeria. Options méditerrannéennes ,Série séminaires
9:137-140.
Labdi M., Robertson L. D., Singh K. B. and Charrier A., 1996. Genetic diversity
and phylogenetic relationships among the annual Cicer species as reveald by isozyme
polymorphism. Euphytica 88:181-188.

Références bibliographique
69
Lacey J., 1997. Actinomycetes in composts. Annuals of Agriculture and
Environnement., 4:113-121.
Lacey, J., 1973. Actinomycetes in soils, composts and fodders. In Actinomycetales:
Characteristics and Practical Importance, pp. 231–251. Edited by G. Sykes & F. A.
Skinner. London: Academic Press.
Ladizinsky G. and Adler A., 1976. The origin of chickpea Cicer arietinum L.
Euphytica 25: 211-217.
Ladizinsky G., 1975. A New Cicer from Turkey. Notes Roy. bot. Gard. Edinb.(in
Press) In Muehlbauer F. J, Rajesh P. N., 2008. Chickpea, a common source of protein
and starch in the semi-arid tropics. PH. Moore, R Ming (eds.) Genomics of tropical
crop plants.
Landa B. B., Hervas A., Bettiol I. W. and Jimenez-Diaz R. M., 1997. Antagonistic
activity of bacteria from the chickpea rhizosphere against Fusarium oxysporum f. sp.
ciceris. Phytoparasitica., 4:305-318.
Landa B. B., Navas-Cortes J. A. and Jimenez-Diaz R. M., 2004. Integrated
management of Fusarium wilt of chickpea with sowing date, host resistance, and
biological control. Phytopathology., 94:946-960.
Landa B. B., Navas-Cortes J. A., Hervas A. and Jimenez-diaz M. R., 2001.
Influence of temperature and inoculum density of Fusarium oxysporum f. sp. ciceris
on suppression of Fusarium wilt of chickpea by Rhizosphere bacteria. Biological
control ., 8: 807-816.
Landa B. B., Navas-Cortes J. A., Jimenez-Gasco M. D. M., Katan J., Retig B.
and Jimenez-Diaz R. M., 2005. Temperature Response of chickpea cultivars to races
of Fusarium oxysporum f. sp. ciceris, causal agent of Fusarium wilt. Plant Disease.,
90:365-374.
Landa B. B., Navas-Cortes J. a., Jimenez-Gasco M. M., Retig B. and Jimenez-
Diaz., 2006. Temperature response of chickpea cultivars to races of Fusarium
oxysporum f. sp. ciceris, causal agent of Fusarium Wilt. The American
Phytopathological Society., 90:365-374.
Larkin R. P. and Fravel D. R., 1999. Mechanisms involved in biological control of
Fusarium wilt of cucumber with strains of non pathogenic Fusarium oxysporum.
Phytopathology., 81:462-469.

Références bibliographique
70
Larkin R. P. and Fravel R., 1998. Efficacity of various fungal and bacterial
biocontrol organisms for control of Fusarium wilt of Tomato. Plants Diseases.,
82:1022-1028.
Larkin R. P. and Fravell D. R., 2002. Effects of varying environnemental conditions
on biological control of Fusarium wilt of Tomato by non pathogenic Fusarium spp.
Phytopathology., 92:1160-1166.
Laterrot H., Bramd R. and Dunay M-C., 1978. La résistance a Corynebacterium
michiganeuse chez la tomate. Annuals Amelioration des Plantes., 28:579-591.
Lazaro E., San Felix A., Van Den L., Ottentheijm H. C. and Ballesta J. P., 2002.
Characterization of Sparsomycin resistance in Streptomyces sparsogens.
Antimicrobiol agents chemother., 46 (9): 2914-2919.
Leach C. M., 1962. The quantitative relationship of UV and Visible radiations to the
induction of reproduction in Ascochyta pisi. Cannadian Journal of Botany., 40:1577-
1602.
Lechevalier H. A., 1989. The Actinomycetes III, A Practical Guide to Generic
Identification of Actinomycetes. Bergey' s Manual of Systematic Bacteriology.
Williams & Wilkins Company, Baltimore. Vol. 4. pp. 2344-2347.
Lechevalier H. A., and Lechevalier M. P., 1981. Introduction to the order
Actinomycetales. In: The prokaryotes, Vol. 2 (Starr M. P., H. Stolp, H. G. Truper, A.
Ballows and H. G. Schlegel. Eds.), Springer-Verlag, Berlin.p.1915-1922.
Lechevalier M. P. and Lechevalier H., 1985. Biology of actinomycètes not
belonging to genus Streptomyces In: Biology of industrial microorganisms. The
Benjamen Cummings Publishing Company, INC.315-360.
Lee J. Y. and Hwang B. K., 2002. Diversity of antifungal actinomycetes in various
vegetative soils of Korea. Canadian Journal of Microbiology., 48:407-417.
Lehr N. A., Schrey S. D., Hampp R. and Tarkka M. T., 2008. Root inoculation
with a forest soil Streptomycete leads to locally and systemically increased resistance
against phytopathogens in Norway spruce. New Phytopathology. doi:10.1111/j.1469-
8137.2007.02322.x.

Références bibliographique
71
Lemanceau P. and Alabouvette C., 1993. Suppression of Fusarium wilts by
fluorescent Pseudomonas: Mechanisms and applications. Biocontrol Science and
Technology., 3:219-231.
Lemanceau P., 1989. Role of competition for carbon and iron in mechanisms of soil
suppressivens to Fusarium wilts. In : Tjamos E. C. and Beckman C. H. (eds) Vascular
Wilt Diseases of Plants (pp385-396),Springer-Verlag,Berlin.
Lemanceau P., Corberand T., Gardan L., Latour X., Laguerre G., Boeufgras J.
M. and Alabouvette C, 1995. Effect of two plant species,ax (Linum usitatissinum L.)
and tomato (Lycopersicon esculentum Mill.), on the diversity of soilborne populations
of Fuorescent pseudomonads. Applied Environnemental Microbiology., 61: 1004-
1012.
Leslie J. F. and Summerell B. A., 2006. The Fusarium Laboratory workshop: a
recent history. Mycotoxin Research., 22:73-74.
Leslie J. F., Zeller K. A., and Summerell B. A., 2001. Icebergs and specialization in
species of Fusarium. Physiology and Plant Pathology., 59:107-117.
Lidell C. M. and Burgess W. L., 1983. Loboratory manual for Fusarium research.
University of Sidney, Sideney,162 pp.
Lie T. A., 1971. Temprature dependent root nodule formation in pea cv. Tran. Plant
soil., 34: 751-752.
Ling L. Y. and Robinson R. J., 1976. Extracting and fractionating lipids from
chickpea. Cereals Food World, 21: 424.
Liu L., Kloepper J. W. and Tuzun S., 1995. Induction of systemic resistance in
cucumber by plant growth promoting rhizobacteria: Duration of protection and effect
of protection and effect of host resistance on protection and root colonization.
Phytopathology, 85: 1064-1068.
Lo C.W., Lai N. S, Cheah H. Y., Wong N. K. I. and HO C. C., 2002.
Actinomycetes isolated from soil samples from the crocker range sabah. ASEAN
Review of Biodiversity and environemental conservation (ARBEC).
Locci R., 1989. Streptomyces and related genera. In: Bergey's Manual of Systematic
bacteriology, vol.4. (ed., Williams, S. T.)Williams and Wilkins company: Baltimore,
USA, pp: 2451-2469.

Références bibliographique
72
Lopez-Bellido L., Lopez-Bellido R. J., Castillo J. E. and Lopez-Pellido F. J., 2004.
Chickpea response to tillage and soil residual nitrogen in a continuous rotation with
wheat I. Biomass and seed yield. Field Crops research 88: 191-200.
Loqman S., Ait Barka E., Clément C. and Ouhdouche Y., 2009. Antagonistic
actinomycetes from Moroccan soil to control the graevine gray mold. World Journal
of Biotechnology., 25:81-91.
Madrid E., Rubiales D., Moral A., Moreno M. T., Millan T., Gil J. and Rubio J.,
2008. Mechanism and molecular markers associated with rust resistance in a chickpea
interspecific cross (Cicer arietinum x Cicer reticulatum). European Journal of Plant
Pathology., 121:43-53.
Magan N. and Lacey J., 1984a. The effect of temperature and pH on the water
relations of field and storage fungi. Transactions of the British Mycological Society
82: 71–81
Magan N. and Lacey J., 1984b. Water relations of some Fusarium species from
infected wheat ears and grain. Transactions of the British Mycological Society 83:
281–285
Mahadevan B. and Crawford D. L., 1997. Properties of the chitinase of the
antifungal biocontrol agent Streptomyces lydicus WYEC108. Enzyme and
Microbiology and Technology., 20:489-493.
Mahler R. L., Saxena M. C., and Aeschllmann J., 1988. Soil fertility requirements
of pea, lentil, chickpea, and faba bean. Pages 279-289 in world Crops (Summerfield
R. J, ed). Dordrecht, The Netherlands: Kluwer academic publishers.
Malhotra R. S., Erskine W., Ergashev N., Beniwal S. P. S. and Shevtsov., 2000.
Chickpea: present status and future prospects in central Asia and the Caucasus. Grain
legumes n°29.
Malhotra R. S.., Pundir K. P. et Slinkard A. E., 1987. Genetic ressources of
chickpea.pp.11-34 In: Saxena H. C. and Singh K. B. (reds). The chickpea CAB
International, Walling ford, U.K.
Mall S., 1973. Studies on microbial antagonism in rhizosphere soils of potato
varieties differentially susceptible to black-scurf and wilt. Plant and Soil., 39: 547.

Références bibliographique
73
Mandeel Q. and Baker R., 1991. Mechanisms involved in biological control of
Fusarium wilt of cucumber with strains of non pathogenic Fusarium oxysporum.
Phytopathology., 81: 462-469.
Mani A. and Sethy C. L., 1984. Plant growth of chickpea as influenced by initial
inoculums levels of Meloidogyne incognita. Indian Journal of Nematodology., 14:41-
44.
Mantovani A., Maranghi F., Ricciardic., Marci C., Stazi A. V. and Attias L. and
Zappoui G. A., 1998.Developmental toxicity of carbendazim: comparison of no
observed-adversed affect level and benchmark dose approach, Food Chemistry and
Toxicology., 36 :37-45.
Mariat f. et Sebald M., 1990. Les actinomycètes Dans: Bactériologie médicale. Le
Minor. Edition Médecine-Science. Flammarion. France.
Markell S., Wise K., McKay K., Goswami R., Gudmestad N., 2008. Ascochyta
blight of chickpea. Plant Disease and management., pp.1362.
Martinez-Noel G. M. A., Madrid E. A., Bottini R. and Lamattina L., 2001. Indole
acetic acid attenuates disease severity in potato-Phytophtora infestans interaction and
inhibits the pathogen growth in vitro. Plant Physiology and Biochemistery.,39:815-
823.
Matta A., 1989. Induced resistance ti Fusarium wilt diseases. In: Tjamos E. C. and
Beckman C. H. (eds) Vascular Wilt Diseases of Plants (pp175-195),Springer-
Verlag,Berlin.
Matthijs S., Tehrani K. A., Laus G., Jackson R. W., Cooper R. M. and Cornelis
P., 2007. Thioquinolobactin, Pseudomonas siderophore with antifungal and anti-
Pythium activity. Environmental Microbiology., 9: 425-434.
Matuo T. and Ishigami K., 1958. On the wilt of Solanum melongenena L. and its
causal fungus Fusarium oxysporum f. sp. melongenase n. f. Annuals of
Phytopathology Sciety JPN., 23:189-192.
Mazur S., Nawrocki J. and Kucmieerz j., 2004. Disease symptoms on chickpea
(Cicer arietinum L.) and their causal agents. Folia Hirticulturae Ann., 16: 47-53.
McCarthy AJ, Williams ST (1990). Methods for Studying the Ecology of
Actinomycetes. Methods In Microbiology. Ed. by. R. Grigorova and JR Norris,
Academic Press Limited, London, 22: 533-363.

Références bibliographique
74
Mehrez K., Belabid L. et Ben Freha F., 2010. Détermination de races et recherché
des génotypes de pois chiche (Cicer arietinum) résistant a Fusarium oxysporum f. sp.
ciceri (agent de flétrissement vasculaire du pois chiche.1er séminaire National sur la
protection des plantes cultivées, Centre Universitaire de Khemis Miliana, Institut des
Sciences de la Nature et de la terre. p.62.
Melakhessou Z., 2007. Etude de la nuisibilité directe des adventices sur la culture de
pois chiche d'hiver (Cicer arietinum L.) variété ILC-3279, cas de Sinapsis arvensis L.
Mémoire du Magister Université El- Hadj Lakhdar -Batna pp.72.
Merriman P. R., Price R. D., Kollmorgen J. F., Piggott T. and Ridge E. H., 1974.
Effect of seed inoculation with Bacillus subtilis and Streptomyces griseus on the
growth of cereals and carrots. Australian Journal of Agricultural Research., 25: 219.
Merzoug A., Ben Freha F. et Taleb M., 2009. Les principales maladies fongiques
du petit pois (Pisum sativum) et Pois chiche (Cicer arietinum) dans le nord oust
algérien. Colloque International : Gestion des risques phytosanitaires, Marrakech,
Maroc.
Messiaen C. M. and Cassini R., 1981. Taxonomy of Fusarium. In: Fusarium
diseases, biology and Taxonomy. Nelson P. E., Tousson A et Cook (reds). The
Pennsylvania state, Press university, Park.
Messiaen C. M., 1959. La systématique du genre Fusarium selon Snyder et Hansen.
Revue de Pathologie végétale et d'entomologie Agricole Française., 38: 253-266.
Messiaen, C. M. & Cassini, R. 1968. Systématique des Fusarium. Annales de
Phytopathologie 3: 386-454.
Miller J. J., 1945. Studies on the Fusarium of muskmelon wilt. I - Pathogenic arid
cultural studies with particular reference to the cause and nature of variation in the
causal organism. Cannadian Journal of Research., 23: 16-43.
Miller J. J., 1946. Cultural and taxonomic studies on certain Fusaria. I - Mutationin
culture. Cannadian Journal of Research., 24: 188-212.
Mohammadi O. and Lahdenpera M. L., 1992. Mycostop biofungicide in practice.
Pages 1-7 in 10th
International Symposium on modern fungicides and antifungal
coumpounds. Thuringia, Germany.

Références bibliographique
75
Molot P. M. et Mas P., 1975. Influence de la température sur la croissance
mycèliènne et le pouvoir pathogène des quatres races physiologiques de Fusarium
oxysporum f. sp. melonis. Annuals of Phytopathology., 7: 175-178.
Molot P. M., Cornus M., Ferriere H. et Lombard D., 1990. Evolution de la
fusariose de l'asperge en conditions hivernales. Influence du froid sur la receptivité de
la plante, utilisation des greffes "frigo". Reviews Horticulture., 303:33-36.
Montesinos E. and Bonaterra A., 1996. Dose-response models in biological control
of plant pathogens :as empirical verification. Phytopathology., 86:464-472.
Moolani M. K. Y. and Chandra S., 1970. Gram cultivation in Haryana. Haryana
Agricultural University, Hisar , India,15pp.
Moreno M. T. and Cubero J. I., 1978. Variation in Cicer arietinum L. Euphytica
27: 465 - 485.
Muehlbauer F. J. and Kaiser N. J., 1964. Using host plant resistance to manage
biotic stress in cool season grain legumes. Euphytica 73:1-10.
Muehlbauer F. J. and Rajesh P. N., 2008. Chickpea, a common source of protein
and starch in the semi-arid tropics. PH. Moore, R Ming (eds.) Genomics of tropical
crop plants.
Muehlbauer F. J. and Singh K. B., 1987. Genetics of chickpea. In: Saxena M. C.
Singh K. B (eds). The chickpea. CAB International Publisher, UK,pp 99-126.
Mukerji K. G., Rajev H., Upadhyay O B., Chamola P. and Dubey P., 2009.
Integrated Pest and diseses management. Publishing corporation 4435-36/7, Ansar
Road, Darya Ganj, New Delhi-110002, 710 pages. Printed at Balaji offset Navis
Shahdara, Delhi, 32.
Mukherjee P. K. and Raghu K., 1997. Effect of temperature on antagonistic and
biocontrol potential of Trichoderma sp. on Sclerotium rolfsii. Mycopathologia.,
139:151-155.
Nakayama K., 1981. Sources of Industrial Microorganisms. In Biotech Ed. By H. J.
Rehm and G. Reed, VCH Verlag, Veinheim.Vol: 1: 355-410.
Namiki F., Shiomi T., Kayamura T and Tsuge T., 1994. Characterization of the
formae specials of Fusarium oxysporum causing wilt of cucurbits by DNA

Références bibliographique
76
fingerprinting with nuclear repetitive DNA sequences. Applied Environnemental
Microbiology., 60: 2684-2691.
Navas Cortes J. A., Alcala-Jimenez A. R., Hau B. and Jimenez-Diaz R. M., 2000.
Influence of inoculum density of races 0 and 5 of Fusarium oxysporum f. sp. ciceris
on development of Fusarium wilt in chickpea cultivars. European Journal of Plant
Pathology., 106:135-146.
Navas-Cortes J. A., Hau B. and Jimenez-Diaz R. M., 1998. Effect of sewing date,
host cultivar, and race of Fusarium oxysporum f. sp. ciceris on development of
Fusarium wilt of chickpea. Phytopathology., 88: 1338-1346.
Ndonde, M. J. M. and Semu E., 2000. Preliminary characterization of some
Streptomyces species from four Tanzanian soils and their antimicrobial potential
against selected plant and animal pathogenic bacteria. World Journal of Microbiology
and Biotechnology., 16:595-599.
Nelson P. E., Toussoun T. A. and Marasas W. F. O., 1983. Fusarium species: An
illustrated manual for identification. The Pennsylvania State University Press,
University Park., 193pp.
Nelson P. E., 1981. Life cucle and epidemiology of Fusarium oxysporum. In: Fungal
wilt diseases of plants. MACEM., Bell A et Beckman C., 1981-ed. Academic Press.,
51-78.
Nelson P. E., Dignani M. C., and Anaissie E. J., 1994. Taxonomy, Biology, and
clinical aspects of Fusarium species. Clinical Microbiology Reviews., 7(4):479-504.
Nene Y. L., Haware M. P. and Reddy M. V., 1978. ICRISAT Information Bulletin
3,44 pp.ICRISAT, Patancheru, Andhra Pradesh, India.
Nene Y. L. et Reddy M. V., 1987. Chickpea diseases and their control. Pages 233-
370 In : The chickpea Saxena M. C and Singh K. B. red. Walking Ford Oxfordshire,
UK: CAB International.
Nene Y. L., 1980. A world list of pigeonpea (Cajanus cajan) and chickpea (Cicer
arietinum) pathogens. ICRISAT Pulse Pathology Progress Report., 3: 1-44.
Nene Y. L., Haware M. P. and Reddy M. V., 1979. Diagnosis of some wilt-like
disorders of chickpea (Cicer arietinum). ICRISAT Information Bulletin., 3: 1-44.

Références bibliographique
77
Nene Y. L., Haware M. P. and Reddy M. V., 1981. Chickpea diseases: resistance
screening techniques. Information Bulletin n°, 10, International Crop Research
Institute for the semi Arid Tropics, Patancheru, pp.1-10.
Nene Y. L., Reddy M. V., Haware M. P., Ghanekar A. M. and Amin K. S., 1991.
Field diagnosis of chickpea diseases and their control. In: Information Bulletin n0.28,
ed. By International Crops Research Institute for the semi Arid Tropics, Patancheru,
India.
Nene Y. L., Sheila V. K. and Sharma S. B., 1984. A world list of chickpea (Cicer
arietinum L.) and pigeon pea (Cajanus cajan L.). Millsp. Pathogens. ICRISAT Pulse
Pathology. Prog. Rep. 32.19pp.
Nene Y. L., Sheila V. K. and Sharma S. B., 1989. A world list of chickpea (Cicer
arietinum L.) and pigeonpea (Cajanus cajan L.)Millsp. Pathogens. ICRISAT legumes
pathology progress report vol.7, Patancheru, India.
Nene Y. L., Sheila V. K. and Sharma S. B., 1996. A world list of chickpea and
pigeon pea pathogens. ICRISAT, India.
Nene Y.L., and Sheila V.K., 1992. Important disease problems of kabuli chickpea. In
Disease resistance breeding in chickpea. Edited by K.B. Singh and M.C. Saxena.
ICARDA, Aleppo, Syria. pp. 11-22.
Nene Y.L., Sheila V. K. and Sharma S.B., 1996. A world list of chickpea and
pigeonpea pathogens. ICRISAT. Patancheru. Andhra Pradesh. India. pp. 27.
Newman D., 1939. The distribution of range in samples from a normal population,
expressed in terms of an independent estimate of standarard deviation. Biometrika.,
31(1):20-30.
Nicot P. C., 2002. Lutte biologique avec des microrganismes contre les maladies
aériènne: situations actuelle et perspectives de développement. 2éme conférences
Internationale sur les moyens alternatifs de lutte contre les organismes nuisibles aus
végetaux., 4-7 Mars,Lille, France.,p.28-36.
Nielson D. C., 2001. Production functions for chickpea, field pea and lentil in the
Central Great Plains. Agronomic Journal., 93:563-569.
Ningthoujam D. S., Sanasam S. and Nimaichand S., 2009. Screening of
Actinomycete Isolates from Niche Habitats in Manipur for Antibiotic Activity.
American Journal of Biochemistry and Biotechnology., 4: 221-225.

Références bibliographique
78
Nolan R. D. and Cross T., 1988. Isolation and screening of actinomycetes. In :
Actinomycetes in Biotechnology, eds. Goodfellow M., Williams S. T. and Mordaarski
M. M. London: Academic Press.ISBN:0-12-289673-4.Academic Press: London, pp:1-
32.
Obaton., 1980. Activité nitrate réductase et nitrogénase en relation avec la
photosynthèse et les facteurs de l'environnement. Bulltin ASF., 55-60.
Odds F. C, Van Gerven F., Espinel-Ingroff A., Bartlett M. S., Ghannoum M. A.,
Lancaster M. V., Pfaller M. A., Rex J. H., Rinaldi M. G. and Walsh T. J., 1998.
Evaluation of possible correlations between antifungal susceptibilities of filamentous
fungi in vitro and antifungal treatment outcomes in animal infection models.
Antimicrobilogy Agents Chemother., 42:282-288.
Ogawa K. and Komada H., 1985. Biological control of Fusarium wilt of sweet
potato with cross protection by prior inoculation with non pathogenic Fusarium
oxysporum. JARQ., 19:20-25.
Ohri D. and Pal M., 1991. The origin of chickpea (Cicer arietinum L.): Karyotype
and nuclear DNA amount. The genetical society of great Britain Heredity., 66: 367-
372.
Omura S., 1992. The search for bioactive compounds from microorganisms.
Springer, Verlag, New York.
O'Neil T. M., Niv A., Elad Y., and Shtienberg D., 1996. Biological control of
Botrytis cinerea on tomato stem wounds with Trichoderma harzianum. European
Journal of Plant Pathology., 102:635-643.
Oskay A., Tamer U. and Azeri C., 2004. Antibacterial activity of some
actinomycetes isolated from farming soils of turkey. African Journal of
Biotechnology., 9: 441-446.
Ouhdouche Y., Barakate M. and Finace C., 2001. Actinomycetes from Maroccan
habitats: screening for antifungal activities. European Journal of Soil and Biology.,
37:69-74.
Padwick G. W., 1940. The genus Fusarium study of the fungus causing wilt of gram
(Cicer arietinum L.) and of the related species of the subsection Orthocera, with
special relation to the variability of key characteristics. Indian Journal of Agricultural
Sciences., 10.

Références bibliographique
79
Pande S., Narayana R. J. and Sharma M., 2007. Establishement of the chickpea
wilt pathogen Fusarium oxysporum f. sp. ciceris in the soil through seed transmission.
Plant Pathol. J., 1: 3-6.
Pande S., Siddique K. H. M., Kishore G.K., Baya B., Gaur P. M., Gowda C. L.
L., Bretag T. and Crouch J. H. 2005. Ascochyta blight of chickpea: biology,
pathogenicity, and disease management. Australian Journal of Agricultural Research
56:317–332.
Pandey B., Ghimire P. and Agrawal V. P., 2003. Studies on the antibacterial
activity of the Actinomycetes isolated from Khumbu Region of Nepal. Academy of
Science and Technology., 4pp.
Patankar A. G., 2000. Biochemical and Molecular analysis of the defense
mechanism in chickpea against biotic stress. Thèse de Doctorat en Biotechnologie,
université de Pune.
Paul A. K. and Banerjee A. K., 1986. In vitro effect of antifungal antibiotic
produced by Streptomyces galbus 5 ME-14. Hindustan Antibiotics ., 28:15–19
Pedrajas M. D. G., Bainbridge B. W., Heale J. B., Perez-Artes E. and Jimenez-
Diaz., 1999. A simple PCR-based method for the detection of the chickpea-wilt
pathogen Fusarium oxysporum f. sp. ciceri in artificial natural soils. European Journal
of Plant Pathology., 105:251-259.
Peres A., 1985. Symptomes et identification des maladies au laboratoire. Creation
CETIOM., 96pp.
Pfohl-Leszkowicz A., 2001. Définition et origine des mycotoxies in les mycotoxines
dans l'alimentation, évaluation et gestion du risque, Ed. Tech and Doc, 3-14.
Pieckova E. and Jesenska Z., 1999. Microscopic fungi in Dwellings and their health
implications in humains. Ann Agric Environ Med., 6:1-11.
Ploetz, R. C. 2000. Management of the most important disease of banana and
plantain, black Sigatoka. Pest. Outl., 11:19-23.
Ploetz, R. C., 2003. Diseases of Tropical Fruit Crops. ed. CAB International,
Wallingford, Oxon, UK.
Polhill R. M. and Raven P. H., 1981. Advances in legume systematic, Part 1 and 2,
eds, Royal Botanic Gardens, Kew, UK.

Références bibliographique
80
Porter J. N., 1971. Prevalence and distribution of antibiotic-producing actinomycets.
Advances in Applied Microbiology., 14:73-92.
Prakash S and Chowdhury J. B., 1992. Effect of Fusarium oxysporum f. sp. ciceri
culture filtrate on callus growth of different genotypes of Cicer arietinum L.
International Chickpea Newsletter., 26:1357-1363.
Preobrazhenskaya T. P., Sveshnikova M. A., Terekhova L. P. and Chormonova
N. T., 1978. Selective isolation of soil actinomycetes. In: Nocardia and Streptomyces,
ed. Mordarski, M., Kurylowicz W and Jeljaszewics J., pp.119-123. Stuttgart: Gustav
Fisher Verlag.
Prévost K., Couture G., Shipley B., Brzezinski R. and Beaulieu C., 2006. Effect of
chitosan and a biocontrol streptomycete on field and potato tuber bacterial
communities. Biocontrol., 51:533-546.
Pridham T. G., Hesseltine C. V. and Benedict R. G., 1958. Aguide for the
classification of Streptomyces according to selected groups placement of strain in
morphological sections. Applied Microbiology., 6:52-70.
Raaijmakers J. M., Leeman M. M. P., Van Oorshot I. V., Slius E., Schippers B.
and Bakker P. A. H. M., 1995. Dose-response relationships in biological control of
Fusarium wilt of radish by Pseudomonas spp. Phytopathology., 85:1075-1081.
Rahel M. A., 1983. Contribution à la verticilliose de la pomme de terre due à
Verticillium dahliae. Mémoire de 3éme
cycle agronomie, Institut Agroniomique et
Véterinaire Hassan II, Rabat, Maroc.
Rahman M., Haware M. P., Mian I. H. and Akanada A. M., 1998. Races of
Fusarium oxysporum f. sp. ciceri causing chickpea wilt in India. Bangladesh Journal
of Plant Pathology., 14:234-237.
Raju S., Jayalakshmi S. K. and Sreeramulu K., 2008. Comparative study on the
induction of defense related enzymes in two different cultivars of chickpea (Cicer
arietinum L.) genotypes by Salicylic acid, spermine and Fusarium oxysporum f. sp.
ciceri. Australienne Journal of Crop Science.,3 :121-140.
Rao L. S., Rani P. U., Deshmukh P. S., Kumar P. A. and Panguluri S. K., 2007.
RAPD and ISSR fingerprinting in cultivated chickpea (Cicer arietinum L.) and its
wild progenitor Cicer reticulatum Ladizinsky. Genetics resources of Crop Evolution.,
54:1235-1244.

Références bibliographique
81
Rapilly F., 1968. Les techniques de mycologie en pathologie végétale. Annueles des
Epiphytes, vol. 19. Institut National de la recherche Agronomique, Paris, 102pp.
Rattanaporn S., Kurakami K., Sukchotiratana M. and Tokuyama S., 2004.
Coastal soil actinomycetes: Thermotolerant strains producing N-Acylamino Acid
Racemase. Science Asia., 30:123-126.
Reddy M. V., Nene Y. L. and Singh K. B., 1980. Field screening of chickpeas for
resistance to Ascochyta blight. International Chickpea Newsletter., 2:13-15.
Rekha K. T. and Thiruvengadam M., 2009. An efficient Micropropagation of
chickpea (Cicer arietinum L.). Phillip Agriculture Scientist., 3: 320-326.
Rekha Y., Shtienberg D. and Katan J., 2000. Disease development following
infection of tomato in basil foliage by airborne conidia of the soilborne pathogens,
Fusarium oxysporum f. sp. radicis-lycopersicy and F. oxysporum f. sp. basili.
Phytopathology 90:1322-1329.
Rheeder J. P., Marasas W. F. O., and Vismer H. F., 2002. Production of
Fumonisin Analogs by Fusarium Species. Applied and Environemental
Microbiology., 2101–2105.
Rhrib A., 1990. Trachéomycoses et pourriture racinaires du pois chiche (Cicer
arietinum L.). Mémoire de 3éme
cycle. Institut Agronomique et Véterinaire Hassan II,
Rabat, Maroc.
Richerdson M. I., 1990. An annotated list of seed-borne diseases (4th
ed.).ISTA
Zurich, Switzerland.
Rouhani H., 1978. Etude de la microflore fongique pathogène sur raciness et tiges de
mais. Mémoire Ingenieur en Biologie appliquée à l'agriculture. ENSA-M(FRA).,
125p.
Rouibah M., 1989. The contribution to the study of the wilting of the chickpeas in
Algeria. Institut National Agronomique El-Harrach.Algérie, 51p.
Rovasz K. Zicsi A., Conteras E., Szell V. and Szabo I. M., 1986. Uber die
Darmaktionoinyceten Geineishaften einiger regenwurum. Opuscula Zoologica,
Budapest., 22:85-102.
Saadoun I. and Al-Momani F., 1997. Studies on soil streptomycetes from Jordan.
Actinomycetes., 8:42-48.

Références bibliographique
82
Saadoun I., Mohammadi H. J., Al-Momani F. and Meqdam A., 1998. Diversity of
soil streptomycetes in northern Jordan. Actinomycetes., 9:53-58.
Sabaou N. et Bounaga N., 1987. Actinomycètes parasites de champignons: étude des
espèces , spécificité de l'action parasitaire au genre Fusarium et antagonisme dans le
sol envers Fusarium oxysporum f. sp. albidinis (Killian et Maire) Gordon. Cannadian
Journal of Microbiology., 33:445-451.
Sabaou N., Amir H ., Bounaga N., 1980. Le palmier et la fusariose. X:
Dénombrement des Actinomycètes de la rhysosphère; leur antagonisme vis-à-vis de
Fusarium oxysporum f.sp. albedinis. Annuals Phytopathology., 12: 253-257.
Sabaou N., Boudjella H., Bennadji A., Mostefaoui A., Zitouni A., Lamari L. et
Bennaji H., 1998. Les sols des oasis du Sahara algérien, source d’actinomycètes,
rares producteurs d’antibiotiques. Sécheresse., 9: 147-153.
Sabaou N., Boudjella H., Mostefaoui A., Zitouni A., Lamari L., Bouti K., Nekhili
H., Nouasri A., Zebiri S., Badji B. et Bennadji A., 1998. Rétrospective sur les
actinomycètes des sols du Sahara algérien: Sélection, écologie, taxonomie et
antibiotiques. Séminaire CRSTRA: zones arides Rétrospectives, enjeux et Stratégies,
Adrar.
Sabaou N., Hacen H., Bennadji A., Bennadji H. and Bounaga N., 1992.
Distrubution quantitative et qualitative des actinomycètes dans les horizons de sol de
surface et profonds d'une palmeraie algériènne. Cannadian Journal of Microbiology.,
38:1066-1073.
Sacramento D. R., Coelho R. R. R., Wigg M. D., Linhares L. T. L., Dos Santos
M. G. M., Semedo L. A. S., and Da Silva A. J. R., 2004. Antimicrobiol and antiviral
activities of an actinomycete (Streptomyces sp.) isolated from a Brazilian tropical
forest soil. World Journal of Microbiology and Biotechnology., 20: 225-229.
Sakal R., Sinha R. B. and Singh A. P., 1988. Effect of born application on black
gram and chickpea production in calcareous soil. Fertiliser News., 33:27-30.
Salomon C. E., Magarvey N. A. and Sherman D. H., 2004. Merging the potential
of microbial genetics with biological and chemical diversity: an ever brighter future
for marine natural product drug discovery. National Production Rep., 21:105-121.
Saravanan T., Bhaskaran R. and Muthusamy M., 2004. Pseudomonas fluorescens
induced enzymological changes in banana roots (Cv. Rasthali) against Fusarium wilt
disease. Plant Pathology Journal., 3(2):72-80.

Références bibliographique
83
Saurez-Estrella F., Vargas-Garcia C., Lopez M. J., Capel C. and Moreno J.,
2007. Antagonistic activity of bacteria and fungi from horticultural compost againt
Fusarium oxysporum f. sp. melonis. Crop Protection., 26: 46-53.
Savic M. and Vasiljevic B., 2006. Targeting polyketide synhase gene pool within
actinomycetes: new degenerate primers. Journal of Microbiology and Biotechnology.,
33:423-430.
Saxena., 1987. Agronomy of chickpea. In : Saxena N., Johansen P. C. and Singh K.
B., eds. The chickpea. Wallingford, UK: CAB International, 207-232.
Scher F. A. M. and Backer R., 1982. Effect of Pseudomonas putida and a synthetic
iron chelator on incubation of soil suppressiveness to Fusarium wilt pathogen.
Phytopathology., 72:1567-1573.
Scher F. A. M. and Baker R., 1980. Mechanism of biological control in a Fusarium
suppressive soil. Phytopathology., 70:412-417.
Schisier D. A., Slininger P. J. and Bothast R. J., 1997. Effects of antagonist cell
concentration and two-strain mixtures on biological control of Fusarium dry rot of
potatoes. Phytopathology., 87:177-183.
Schneider R. W., 1984. Effects of non pathogenic strains of Fusarium oxysporum on
celery root infection by Fusarium oxysporum f. appi and a novel use of the
Lineweaver-Burk double reciprocal plot technique. Phytopathology., 74:646-653.
Schouten A., Van Der Berg G., Edel-Hermann V., Steinberg C., Gautheron N.,
Alabouvette C., De Vos C. H., Lemanceau P. and Raaijmakers J. M., 2004.
Defense Responses of Fusarium oxysporum to 2, 4-Diacetylphloroglucinol, a Broad-
Spectrum Antibiotic produced by Pseudomonas fluorecens. The American
Phytopathological Society., 11:1201-1211.
Sedra M. H., 1985. Potentiel infectieux et receptivité de quelques sols de palmeraie à
la fusariose vasculaire du palmier dattier causée par Fusarium oxysporum f. sp.
albedinis. Mémoire de 3éme cycle. Institut Agronomique et Véterinaire Hassan II,
Rabat, Maroc, 88p.
Seifert K. A. and Gams W., 2001. The taxonomy of anamorphic fungi. The Mycota
VII Part A, Systematics and evolution, Maclanghlin /Mclanglin /Lemke(Eds).
Springer-Verlag, Berlin. Heidelberg.

Références bibliographique
84
Seifert K. A., 2001. Fusarium and anamorphic generic concepts. In: Summerell B. A
and Leslie J. F, Backhouse D, Bryden W. L. and Burgess L. W (eds), Fusarium: Paul
E. Nelson Memorial Symposium American phytopathology Society Press, St. Paul,
Minnesota., pp.15-28.
Serrano J. A. and Trujillo A. H. S., 2005. Identificacion y diagnostic de
Actinonicetales pathogenos. Publicaciones del Vicerrectorado Académico Collecion
Ciencias de la Salud. pp 256.
Sethy N. K., Choudhary S., Shokeen B. and Bhatia S., 2006. Identification of
microsatellite markers from Cicer reticulatum molecular variation and phylogenetic
analysis. Theoritical Applied Genetics., 112:347-357.
Sette L. D., De Oliveira V. M. and Manfio G. P., 2005. Isolation and
characterization of alachlor - degrading actinomycetes from soil. Antonie Van
Leeuwenhoek., 87:81-89.
Shah T. S., Babar M. A., Iqbal J. M. and Ahsanul M. H., 2009. Screening of
chickpea (Cicer arietinum) ibduced mutants against Fusarium Wilt. Pakistan Journal
of Botany., 4: 1945-1955.
Shaikh M. H., 1974. Studies on wilt of gram (Cicer arietinum L.) caused by
Fusarium oxysporum f.sp. ciceri in Marathwada Region. Master Thesis, Marathwada
Krishi Vidyapeeth, Parbhani, India
Sharma D. K. and Muehlbauer F. J., 2007. Fusarium wilt of chickpea:
physiological specialization, genetics of resistance and resistance gene tagging.
Euphytica., 157:1-14.
Sharma K. D., Chen W. and Muehlbauer F. J., 2005. Genetics of chickpea
resistance to five races of Fusarium wilt and a concise set of race differentials for
Fusarium oxysporum f. sp. ciceris. Plant Diseases., 89: 385-390.
Sharma M., Kumar R. V., Narayan R. J., Kannan S., Holsington D. and Pande
S., 2009. Genetic diversity in Indian isolates of Fusarium oxysporum f. sp. ciceris,
chickpea wilt pathogen. African Journal of Biotechnology., 6: 1016-1023.
Sharma P. C. and Gupta P. K., 1983. Cytological studies in the genus Cicer L. In:
Proceedings of the XV International Congress of Genetics, New Delhi, 12-21
Dec.1983. Oxford and IBH Publishing Company, New Delhi, India, Abstract n°1257.
Shimizu M. Y., Nakagawa Y., Sato T., Furumai Y., Igaroshi Y., Onaka H.,
Yoshida R. Y. and Kunoh H., 2000. Studies on endophytic actinomycetes

Références bibliographique
85
Streptomyces sp. islated from Rododendron and its antifungal activity. Journal of
General Plant Pathology., 66: 360-366.
Shirling E. B. and Gottlieb D., 1966. Methods for characterization of Streptomyces
species. International Journal of systematic bacteriology.,16: 313-340.
Si-Hassen A., 1990. Contribution à l'étude du flétrissement du pois chiche (cicer
arietinum L.) en Algérie. Mémoire Ingenieur en agronomie. INA El-Harrach,
Algérie., 60pp.
Simon A. and Sivasithamparam K., 1989. Pathogen suppression: A case study in
biological suppression of Gaeumannomyces graminis var. tritici in soil. Soil Biology
and Biotechnology., 21:331-337.
Simon D. G., Rosana M. P., Martins Rosalie R. R., Nazareth C. L., Marta M. H.,
Branquinha A. and Beatriz V., 1999. Influence of growth medium in protease and
pigment production in Streptomyces cyaneus. Mem.Inst.Oswaldo.Cruz. 94 (2): 173-
177.
Singh B. P, Saikia R., Yadav M., Singh R., Chauhan V. S and Arora D. K.,2006.
Molecular characterization of Fusarium oxysporum f. sp. ciceri causing wilt of
chickpea. African Journal of Biotechnology ., 6: 497-502.
Singh F. and Diwakar B., 1995. Chickpea Botany and production Practices. Skill
Development series ICRISAT India., 16 : 502-324.
Singh G. and Bushan L. S., 1979. Water use, water use efficiency and yield of
dryland chickpea as influenced by fertilization, stored soil water and crop season
rainfall. Agricultural Water Management., 2:299-305.
Singh K. B. and Dahya B. S., 1973. Breeding for wilt resistance in chickpea. Pages
13-14 In: Symposium on wilt problem and breeding for wilt resistance in Bengal
gram, India, Agricultural resources Institute, (New Delhi).
Singh K. B. and Jimenez-Diaz R. M., 1996. Registration of six Fusarium wilt-
resistant chickpea germplasm lines. Crop Sciences., 36:817.
Singh K. B. and Ocampo B., 1997. Exploitation of wild Cicer species for yield
improveemnt in chickpea. Theor Appl Genet.,95: 418-423.
Singh K. B. and Reddy M. V., 1991. Advances in diseases resistance breeding in
chickpea. Advances in Agronomic., 45: 191-222.

Références bibliographique
86
Singh K. B., 1987. Chickpea breeding. In: Saxena M. C. and Singh K. B. (eds) the
chickpea. CAB International Publisher, UK, pp127-162.
Singh K. B., 1990. Prospects of developping new genetic material and breeding
methodologies for chickpea improvement. Options méditerranéennes, série
séminaires., 9:43-50.
Singh K. B., Malhotra., Halila M. H., Knights E. J. and Verma M. M.,1994.
Current status and future strategy in breeding chickpea for resistance to biotic and
abiotic stresses. Euphytica., 73:137-149.
Singh K. B., Ocampo B. and Robertson L. D., 1998. Diversity for abiotic and biotic
stress . Ressources and Crop Evolution 45:9-17.
Singh O. P. Raghavendra K., Nanda N., Mittal P. K. and Subbarao S. K., 2002.
Pyrthroid resistance in An. Culcifacies in surat district, Gujarat, West India. Current
Science., 82:547-550.
Singh O., Gowda C. L. L., Sethi S. C., Dasgupta T., Kumar J. and Smithson.,
1993. Genetic analysis of agronomic characters in chickpea .II. Estimates of genetic
variances from line X tester mating desingns. Theor Applied Genetics., 85:1010-1016.
Singh P. J., Mehrotra R. S., 1980. Biological control of Rhizoctonia bataticola on
gram by coating seed with Bacillus and Streptomyces spp. and their influence on plant
growth. Plant and soil., 56:475-483.
Singh U. and Singh B., 1992. Tropical grain legumes as important human foods.
Economic botany., 46:310-321.
Singh U., 1985. Nutritional quality of chickpea (Cicer arietinum l.): Current status
and future research needs. Quality plant foods Human Nutrition., 35: 339-351.
Singh, K.B. and Saxena M.C., 1999. Chickpeas. In R. Coste (ed). The Tropical
Agriculturist. CTA, ICARDA. MacMillan Education Ltd, London.
Sivasithamparam K. and Parker C. A., 1980. Interaction of certain isolates of soil
fungi with Gaeumannomyces graminis var. trici on agar media. Australian Journal of
Botany., 28: 411-419.
Smith T. L., Pearson M. L. and Wilcox K. R., 1999. Emergence of vancomycin
resistance in Staphylococcus aureus. New Eng. Journal of Mediteranean., 340: 493-
501.

Références bibliographique
87
Smithson J. B., Thompson J. A. and Summerfield R. J., 1985. Chickpea (Cicer
arietinum L.). In: Summerfield R. J. and Roberts E. H. (Eds), Grain Legume Crop ,
pp.312-390. Collins, London, UK.
Sneh B., Dupler M., Elad Y. and Baker R., 1984. Chlamydospore germination of
Fusarium oxysporum f. sp. cucumerinum as affected by fluorescent and lytic bacteria
from a Fusarium suppressive soil. Phytopathology., 74:1115-1124.
Snyder W. C and Hansen H. N., 1940. The species concept in Fusarium. Annual
Journal of Botany., 27: 64-67.
Snyder W. C. and Hansen H. N., 1941. The qspecies concept in Fusarium wilt
reference to section Martiella. Am. Journal of Botany., 28: 738-742.
Snyder W. C. and Smith S. N., 1972. Biology of bayoud disease organism Fusarium
oxysporum and Fusarium wilt populations in the soil in relation to the control of
bayoud. Proceeding of the first International Seminar and Worckshop. Ministry of
Agriculture and land reform Algeria, 169-176.
Soares A. C. F., Sousa C. S., Garrido M. S., Perez J. O. Almeida N. S., 2006. Soil
Streptomycetes with in vitro activity against the yam pathogens Curvularia
eragrostides and Colletotrichum gloeosporioides. Brazilian Journal of Microbiology.,
37:456-461.
Staginnus C., Winter P., Desel C., Schmidt T. and Kahl G., 1999. Molecular
structure and chromosomal localization of major repetitive DNA families in the
chickpea (Cicer arietinum L.) genome. Plant molecular Biology 39:1037-1050,1999.
Steingrube V. A., Brown B. A., Gibson J. L., 1995. DNA amplification and
restriction endonuclease analysis for differentiation of 12 species and taxa of
Nocardia, including recognition of four new taxa within the Nocardia asteroides
complex. Journal of Clinical Microbiology., 33: 3096–3101.
Steingrube V. A., Wilson R. W., Brown B. A., Jost K. C., Blacklock Z., Gibson J.
l., and Wallace R. J. J., 1997. Rapid identification of the clinically significant
species and taxa of aerobic actinomycetes, including Actinomadura, Gordona,
Nocardia, Rhodococcus, Streptomyces and Tsukamurella isolates by DNA
amplification and restriction endonuclease analysis. Journal of Clinical
Microbiology., 35: 817-822.
Steinkellner S., Mammerler R. and Vierheilig H., 2005. Microconidia germination
of the tomato pathogen Fusarium oxysporum in the presence of root exudates. Journal
of Plant Interactions., 1: 23-30.

Références bibliographique
88
Stevenson P. C., Pedgham D. E. and Haware., 1995. Root exudates associated with
the resistance of four chickpea cultivars (Cicer arietinum) to two races of Fusarium
oxysporum f. sp. ciceri. Plant Pathology., 44:686-694.
Sudupak M. A., Akkaya M. S. and Kence A., 2004. Genetic relationships among
perennial and annual Cicer species growing in turkey assessed by AFLP.
fingerprinting Theoritical Applied in Genetics.,108:937-944.
Sugha S. K., Kapoor S. K. and Singh B. M., 1994. Factors influencing Fusarium
wilt of chickpea (Cicer arietinum L.). Indian Journal of Mycology and Plant
Pathology., 24:97-102.
Summerbell R. C. and Schroers H. J., 2002. Analysis of phylogenetic relashionship
of Cylindrocarpon lichenicola and Acremonium falciforme to the Fusarium solani
species complex and a review of similarities in the spectrum of opportunistic
infections caused by these fungi. Journal of Clinical Microbiology., 40:2866-2875.
Summerell B. A., Salleh B. and Leslie J. F., 2003. A utilitarian approach to
Fusarium identification. Plant Disease., 87:117-128.
Summerfield R. J., Minchin F. R., Roberts E. H. and Hadley P., 1979. The effects
of photoperiod and air temperature on growth and yield chickpea (Cicer arietinum
L.). Proceedings International workshop on chickpea improvement. Ed.
ICRISAT:121-144.
Summerfield R. J., Vbrmani S. M., Roberts E. H. and Ellis R. H., 1990.
Adaptation of chickpea to agroclimatic constraints. Pages 61-72 in chickpea in the
nineties: Proceedings of the second International Workshop on chickpea
Improvement, 4-8 Desc., ICRISAT Center, India.
Susanne D. Wolfgang B., 1990. Elicitor-induced metabolic changes in cell cultures
of chickpea (Cicer arietinum L.) cultivars resistant and susceptible to Ascochyta
rabiei.Planta.,182 :279-286.
Taechowisan T., Peberdy J. F. and Lumyong S., 2004. PCR cloning and
heterologous expression of chitinase gene of endophytic Streptomyces aureofaciens
CMUAc130. Journal of Genetics Applied of Microbiology.,50: 177-182.
Tahvonen R. 1982b., Preliminary experiments into the use of Streptomyces spp.
Isolated from peat in the biological control of soil and seed borne diseases in peat
culture. Journal of Agriculture Sciences., 54: 357-369.

Références bibliographique
89
Tahvonen R. T. and Avikainen H., 1987. The biological control of seed borne
Alternaria brassicicola of cruciferous plants with a powdery preparation of
Streptomyces sp. Journal of Agricultural Science in Finland., 59:199-208.
Tahvonen R., 1982a. The suppressiveness of Finnish light coulerd Sphagnum peat.
Journal of Agriculture Sciences., 54:345-356.
Tanaka Y. and Omura S., 1993. Agroactive compounds of microbial origin.
Annuals Review of Microbiology., 47:57-87.
Tariq M., J. H. Mirza and A. B. Shakir., 1993. Physiological studies of Botrytis
gladiolorum and its in vitro sensitivity to fungicides. Pakistan Journal Phytopathol., 5:
89–92
Taylor J. W., Jacobson D. J., Kroken S., Kasuga T., Geiser D. M., Hibbert D. S.,
and Fisher M. C., 2000. Phylogenetic species recognition and species concepts in
fungi. Fungal Genetics of Biology., 31:21-32.
Taylor, J. W., D. J. Jacobson, and M. C. Fisher. 1999. The evolution of asexual
fungi: reproduction, speciation and classification. Annual Review of Phytopathology
37:197--246.
Tayyar R. I. and Waines J. G., 1996. Genetic relashionships among annual species
of Cicer (fabaceae) using isoenzyme variation. Theoritical and Applied Genetics.,
92:245-254.
Tedla T. and Stanghellini M. E., 1992. Bacterial population dynamics and
interactions with Pythium aphanidermatum in intact rhizosphere soil.
Phytopathology., 82:652-656.
Tello J. C. and Lacas A., 1990. Fusarium oxysporum en los cultivos intensivos del
litoral mediterraneo de Espana. Fases parasitica (Fusariosis vasculars del tomate y del
clavel) y no parasitaria. BolSan Veg no., 19-190 pp.
Thakor Y., 2006. The biopesticide market for global agricultural use. Industrial
Biotechnology. 2 (3): 294-208.
Thrane U. and Seifert K., 2000. Fusarium Link: Fr. In: Samson R. A., Hockstra, E.
S., Frisvad J. C. and Filtenborg, O. (eds). Introduction to food and airborne fungi.
Cebtraalburean Voor Schimmecultures, Utecht, pp. 120-157.
Thrane U., 2001. Developments of taxonomy of Fusarium species based on
secondary metabolites. In Summerell, B. A., Leslie J. F.., Backhouse D. Bryden W. L.

Références bibliographique
90
and Burgess L. W (eds), Fusarium: Paul N. Nelson Memorial Symposium. American
Phytopathology Society Press, St. Paul, Minnesota, pp.29-49.
Tiraby G. et Etienne G., 1983. L'avenir des Streptomyces. Biofuture., 15:33-37.
Todar K., 2009. The Normal Bacterial Flora of Humans. Lectures in Microbiology ,
University of Wisconsin-Madison Department of Bacteriology.
Torp M., Langseth W., 1999. Production of T-2 toxin by a Fusarium resembling
Fusarium poae. Mycopathologia., 147:89–96.
Toulaiti., 1988. L'agriculture algérienne: les causes de l'échec. Ed. Office des
publications universitaire, Alger, 550p.
Tousson T. A. and Nelson P. E., 1976. A pictorial guide to the identification of
Fusarium species. Pennsylvania State University Press, University Park, 51pp.
Trapero-Casa A., 1983. Wilt and Root rot of chickpea in the Guadelquivir valley:
Importance, distribution, etiology, epidemiology and control (Original in Spanish).
Doctorat Thesis , University of Cordoba, 295pp.
Trapero-Casas A. and Jimenez-Diaz R. M., 1985. Fungal wilt and root rot diseases
of chickpea in southern Spain. Phytopathology 75:1146-1151.
Trapero-Casas A., Kaiser W. J. and Ingram D. M., 1990. Control of Pythium seed
rot and preemergence damping-off of chickpea in the U. S. Pacific Northwest and
Spain. Plant Diseases., 74:563-569.
Tubac C., 2007. Flore fongique de different substrats et conditions optimales de
production des mycotoxines. Thèse de doctorat, Institue Nationale polytechnique de
Toulose et de l'université de Bucarest., 190p.
Uma Maheshwari T, Sharma SB, Reddy DDR, Haware MP., 1997.Interaction of
Fusarium oxysporum f. sp. ciceri and Meloidogyne javanica on Cicer arietinum.
Journal of nematology 29(1):117-126.
Upadhyaya H. D., Dwivedi S. L., Gowda C. L. L. and Singh S., 2007. Identification of diverse germplasm lines for agronomique traits in a chickpea (Cicer
aritinum L.) core colletion for use in crop improvement. Field Crops Research., 100:
153-158.

Références bibliographique
91
Valacilova K., Ohri D., Vrana J., Cihalicova J., Kubalakova M., Kahl G. and
Dolezel J., 2002. Development of flow cytogenetics and physical genome mapping in
chickpea (Cicer arietinum L.). Chromosome Research., 10:695-706.
Valois D., Fayad K., Barasubiye T., Garon M., Dery C., Brzezinski R. and
Beaulieu C., 1996. Glucanolytic actinomycetes antagonistic to Phytophthora
fragariae var. rubi, the causal agent of raspberry root rot. Applied and Environmental
Microbiology., 62:1630-1635.
Van Der Maesan L. J. G., 1972. Cicer L. a monograph on the genus with special
references to the chickpea (Cicer arietinum L.), its ecology and cultivation. Thesis,
Agricultural University Wageningen Medad Landbouwhogeschool,Wageningen., 72-
10.
Van Der Maesan L. J. G., 1987. Origine, history and taxonomy of chickpea.
Pages11-34 In Singh, F. et Diwakar, B.1995.Chickpea Botany and Production
Practices.Skill Development Series n°.16; ICRISAT.
Van Der Maesan L. J. G., Maxted N., Javadi F., Coles S. and Davies A. M. R.,
2007. Taxonomy of the genus Cicer revisited. In: Yadav S. S., Reeden B., Chen W,
and Sharma B (eds) Chickpea breeding and management. CAB International,
Wallingford, pp 14-46.
Van Der Plank J. C., 1968. Disease resistance in plants. Academic press, New York,
London: 206p.
Van Peer R., Niemann G. J. and Schippers B., 1991. Induced resistance and
phytoalexin accumulation in biological control of Fusarium wilt of carnation by
Pseudomonas spp. strain W. C. S. 417 r. Phytopathlogy., 81:728-734.
Van Rheenen H. A., Reddy M. V., Kumar J. and Haware M. P., 1989. Breeding
for resistance to soil borne disease in chickpea. Consultaney Meeting on breeding for
diseases resistance in Kabuli Chickpea. ICARDA, Allepo, Syria.
Van Schreven D. A., 1964. Antagonistic effect of actinomycetes on rhizobia. Plant
and Soil., 21:283.
Vankatesan S., Ganesan R. and Muthuchelian K., 2008. Isolation, Identification
and characterization of Melanin pigment production in Streptomyces from Megamalai
forest soil. Ethnobotanical Leaflets., 12(1): 827-831.

Références bibliographique
92
Vassilev N., Vassileva M. and Nikolaeva I., 2006. Simultaneous P-solubilizing and
biocontrol activity of microorganisms: Potentials and future trends. Applied in
Microbiology and Biotechnology., 71 :137-144.
Verma V. C., Gond S. K., Kumar A., Mishra A., Kharwar R. N. and Gange A.
C., 2009. Endophylitic Actinomycetes from Azadirachta indica A. Juss.: Isolation,
Diversity, and Anti-Microbiol activity. Microbiol Ecology., 57: 749-756.
Verret F., 1982. Etude de quelques légumineuses à gousses grains adaptées au semis
de printemps dans la zone méditerranéenne. Mémoire D.A.A. Montpellier,France,
72p.
Vincent C. and Gregory P. J., 1986. Differences in the growth and development of
chickpea seedling roots (C. arietinum). Experimental Agriculture., 22: 233-242.
Vintika J and Vintikova H., 1961. Effect of antibiotics on rhizobia. For Socialist
Agricultural Science., 7: 349.
Waite B . H. and Stover R. H. 1960. Studies on Fusarium wilt of Bananas. VI -
Variability and the cultivar concept in Fusarium oxysporum f. cubense. Cannadian
Journal of Botany., 38: 985-994
Waksman SA (1961). The Actinomycetes, Classification, Identification and
Description of Genera and Species. Baltimore: The Williams and Wilkins Company.
Vol. 2 pp. 61-292.
Walker J. C., 1965. Use of environmental factors in screening diseas resistance.
Annuals Review of Phytopathology., 3: 197-208.
Wang X., Bunkers G. J., Walters M. R., and Thomas R. S., 2001. Purification and
characterization of three antifungal proteins from cheesweed (Malva parviflora).
Biotechnology Research., 282: 1224-1228.
Warnock D.W. and Speller D. C. E., 1982 Antifungals today. Journal of
Antimicrobial Chemotherapy., 10: 164-167.
Wellman L . and Blaisdell D.J., 1941. Pathogenic and cultural variation
amongsingle-spores isolates from strain of the tomato-wilt Fusarium.
Phytopathology., 31: 103-120.
Wendisch F. K. and Kutzner H. J., 1991. The family Streptomycetaceae. In
Prokaryotes. Ed. by A. Ballows et al., Springer Verlag. Second edition. Vol. 1. pp.
922-995.

Références bibliographique
93
Westerlund J., Campbelle R. N. and Kimble K. A., 1974. Fungal root rots and wilt
of chickpea in California. Phytopathology., 64:432-436.
Whipps J. M. and Davis K. G., 2000. Success in biological control of plant
pathogens and nemathodes by microorganisms. In: Gurr G. and Wratten S. S., eds.
Measures of success in biological control. Dordrecht: The Netherlands: Kluwer
Academic Puplisher, 231-269.
Whipps J.M., 1987. Effect of media on growth and interactions between a range of
soil borne glasshouse pathogens and antagonistic fungi. New Phytopathologist., 107-
142.
Wiehle W., Scholoter M. and al., 1996. Detection of colonization by Pseudomonas
PsIA12 of inoculated roots of Lupinus albus and Pysum sativum in greenhouse
experiments with immunological techniques. Symbiosis., 20 (2): 129-145.
Williams S. T., Sharples G. P. and Bradshaw R. M., 1973. The fine structure of
Actinomycetales, p.113-130. In: Sykes G. and Skinner F. A. (ed). Actinomycetales:
Characteristics and practical importance. Academic Press. Inc., New York,N.Y.
Williams P. C. and Singh U., 1987. The chickpea nutritional quality and the
evaluation of quality in breeding programmes. In: Saxena M. C., Singh K. B. (eds),
The chickpea, CAB International, Wellingford, UK,pp.329-356.
Williams S. T. and Vickers J. C., 1988. Detection of actinomycetes in natural
environment, problems and perspectives. In : Biology of Actinomycetes 88,ed. Okami
Y., Beppu T. and Ogawara H.pp.265-270.Tokyo:Japan Scientifics societies
Press.ISBN:4-7622-1552-X.
Williams S. T., Goodfellow M. and Anderson G., 1989. Genus Streptomyces
Waksman and Henrici,1943,339AL. In: Williams ST, Sharpe M. E (eds) Bergey's
manual of systemic bacteriology, vol 4. Springer, New York, pp 2452-2492.
Williams S.T., Goodfellow M., Alderson G., Wellington E. M., Sneath P.H. and
Sackin M., 1983. Numerical classification of Streptomyces and related genera.
Journal of Genetics and Microbiology., 129:1743-813.
Wilson, R. W., Steingrube, V. A., Brown, B. A. & Wallace, R. J., Jr (1998). Clinical application of PCR-restriction enzyme pattern analysis for rapid
identification of aerobic actinomycete isolates. J Clin Microbiol 36, 148–152.
Winstead N. N. and Walker J. C., 1954. Toxic metabolites of the pathogen in
relation to Fusarium resistance. Ibid., 44:159-166.

Références bibliographique
94
Winter P., Benko-Iseppon A-M., Huttel B., Ratnaparkhe M., Tullu A., Sonnante
G., Pfaff T., Tekeoglu M., Santra D., sant V. J., Rajesh P. N., Kahl G. and
Muehlbauer F. J., 2000. A linkage map of the chickpea (Cicer arietinum L.) genome
based on recombinant inbred lines from a C. arietinum X C. reticulatum cross:
localization of resistance genes for Fusarium races 4 and 5. Theor Appl Genet.,101:
1155-1163.
Wong W. C., 1988. A differential medium for the identification of races 1 and 4 of
Fusarium oxysporum f. sp. cubense. Letters in Applied Microbiology., 6: 51-54.
Xiao K., Kinkel L. L. and Samac D. A., 2002. Biological control of Phytophthora
root rots on alfalfa and Soybean with Streptomyces. Biological Control., 23:285-295.
Xu L. H., Li Q. R. and Jiang C. L., 1996. Diversity odf soil actinomycetes in
Yunnan. China Applied in Environnemental and Microbiology., 62:244-248.
Yadav S. S., Redden R., Chen W. and Sharma B., 2007. Chickpea breeding and
management. Cambridge library of Congress. (Livre).
Yamamoto Y., Kouchiwa T., Hodoki Y., Hotta K., Uchida H. and Harada K-I.,
1998. Distribution and identification of actinomycets lysing cyanobacteria in a
eutrophic lake. Journal of Applied Phycology., 10: 391–397.
Yamamura H., Tamura T., Sakiyama Y. and Harayama S., 2007. Nocardia
amamiensis sp. nov., isolated from a sugar-cane field in Japan. International Journal
of Systematic Evolution of Microbiology., 57:1599-1602.
Yang L., Xie J. et al., 2008. Antifungal sustances produced by Penicillium oxalicum
strain PY-1 potential antibiotics against plant pathogenic fungi. World Journal of
Microbiology and Biotechnology., 24 (7): 909-915.
Yuan W. M. and Crawford D. L., 1995. Characterization of Streptomyces lydicus
WYE108 as potential biocontrol agent against fungal root and seed rots. Applied in
Environemental and Microbiology., 61:3119-3128.
Yusuf A. M., Krishnamurthy L., Saxena N. P., Rupela O. P., Kumar J. and
Johansen C., 2002. Scope for genetic manipulation of mineral acquisition in
chickpea. Plant and soil., 245:123-134.
Zaim S., 2007. Contribution à l'étude de Trichoderma spp. agent antagoniste de
Fusarium oxysporum f. sp. ciceri et Fusarium oxysporum f. sp. lentis, agents du

Références bibliographique
95
flétrissement vasculaire du pois chiche et de la lentille. Mémoire de magister,
Université de Mascara., 113pp.
Zaim S., Bekkar A. A., Bellahcen M. et Beabid L., 2010. Effets des antagonistes du
pois chiche sur l'infection par Fusarium oxysporum f. sp. ciceri. 1er séminaire
National sur la pritection des plantes cultivées, Centre universitaire de Khemis
Miliana, Institut des Sciences de la nature et de la terre 25-26 mai 2010.
Zitouni A., Boudjella H., Lamari L., Badji B., Mathieu F., Lebrihi A. and
Sabaou N., 2005. Nocardiopsis and Saccharothrix genera in Saharan soils in Algeria:
Isolation, biological activities and partial characterization of antibiotics. Research in
Microbiology., 156:984-993.
Zohary D. and Hopf M., 2000. Domestication of plants in the old world, 3rd
edn.
Oxford University Press, New York.

45
Annexes

Annexes
45
Annexe 1
Composition des milieux de cultures utilisés
Milieu PDA
Extrait de pomme de terre……………………………………………………........200g
Glucose………………………………………………………………………...........20g
Agar-agar……………………………………………………………………………15g
Eau distillée……………………………………………………………………..1000ml
pH……………………………………………………………………………………5.6
Milieu pois chiche
Extrait de pois chiche……………………………………………………………….70g
Saccharose…………………………………………………………………………..20g
Agar-agar……………………………………………………………………............20g
Eau distillée……………………………………………………………………..1000ml
pH……………………………………………………………………………............5.6
Gélose nutritive
Extrait de viande………………………………………………………………...........1g
Extrait de levure………………………………………………………………...........2g
Peptone……………………………………………………………………………….5g
Chlorure de sodium…………………………………………………………………..5g
Agar-agar……………………………………………………………………………15g
pH……………………………………………………………………………………7.4
Milieu TSP2
Extrait de malt………………………………………………………………………10g
Extrait de levure………………………………………………………………...........4g
Glucose……………………………………………………………………………….4g

Annexes
46
Agar-agar……………………………………………………………………............20g
Eau distillée…………………………………………………………………......1000ml
pH……………………………………………………………………………………7.3
Milieu Malt
Extrait de malt………………………………………………………………………20g
Agar-agar……………………………………………………………………………20g
Eau distillée……………………………………………………………………..1000ml
pH……………………………………………………………………………………5.6
Milieu Czapeck
NaNO3………………………………………………………………………….........2g
K2HPO…………………………………………………………………………..........1g
KCL………………………………………………………………………………...0.5g
MgSO4, 7H2O………………………………………………………………………0.5g
FeSO4, 7H2O……………………………………………………………………...0.01g
Saccharose…………………………………………………………………………..30g
Agar-agar………………………………………………………………....................15g
Eau distillée……………………………………………………………..............1000ml
pH……………………………………………………………………………………5.6
Milieu Richards
KH2PO4………………………………………………………………………………1g
MgSO4, 7 H2O……………………………………………………………………...0.5g
FeSO4, 7H2O……………………………………………………………….........0.005g
ZnSO4……………………………………………………………………………0.005g
MnCl2 …………………………………………………………………………...0.002g
Galactose……………………………………………………………………………20g

Annexes
47
Glycocolle……………………………………………………………………….2.250g
Agar-agar……………………………………………………………………………20g
Eau distillée……………………………………………………………………..1000ml
pH……………………………………………………………………………………5.6
Milieu Sabouraud
Dextrose…………………………………………………………………………….40g
Peptone……………………………………………………………………………...10g
Agar…………………………………………………………………………………20g
Eau distillée……………………………………………………………………..1000ml
pH……………………………………………………………………………………5.6
Milieu GLM
Extrait de levure……………………………………………………………………...3g
Extrait de malt………………………………………………………………………..3g
Peptone……………………………………………………………………………….5g
Glucose……………………………………………………………………………...10g
Agar…………………………………………………………………………………15g
Eau distillée………………………………………………………………………100ml
pH……………………………………………………………………………………7.2
Annexe 2
Préparation des tampons pour le test de la détermination du pH (selon Geigy, 1990)
pH Solutions-stocks Composition du
tampon A B
4.5 Citrate disodique 0.1M HCl 0.1M 74.9 ml A+ 25.1 ml B
5.5 Citrate disodique 0.1M NaNOH 0.1M 70.3 ml A+ 29.7 ml B

Annexes
48
6.5 Citrate disodique 0.1M NaNOH 0.1M 53.2 ml A+ 46.8 ml B
7.5 Phosphate
monopotassique 1/15 M
Phosphate disodique 1/15
M
12.8 ml A+ 87.2 ml B
Annexe 3
Résultats de l'analyse statistique
Analyse de la variance des mensurations (longueurs et les largeurs des conidies)
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 4796.11 19 252.42 918.73
Var. factorielle 4768.28 3 1589.42
Var. résiduelle 27.83 16 1.73
Analyse de la variance de la croissance mycélienne des 17 isolats de FOC sur PDA
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 129.84 17 7.63 138.52
Var. factorielle 116.36 1 116.36
Var. résiduelle 13.48 16 0.84
Analyse de la variance de l'influence de la température sur la croissance mycélienne
des 17 isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 460.51 21 21.92 65.52
Var. factorielle 439.02 5 87.80
Var. résiduelle 21.49 16 1.34

Annexes
49
Analyse de la variance de l'influence du pH sur la croissance mycélienne des 17
isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 58.45 19 3.07 4.5
Var. factorielle 26.73 3 8.91
Var. résiduelle 31.72 16 1.98
Analyse de la variance de l'influence de milieux de cultures sur la croissance
mycélienne des 17 isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 131.421 22 5.97 11.53
Var. factorielle 106.641 6 17.77
Var. résiduelle 24.78 16 1.54
Analyse de la variance de l'influence de la source de carbone sur la croissance
mycélienne de 17 isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 28.96 18 1.60 1.12
Var. factorielle 3.55 2 1.77
Var. résiduelle 25.41 16 1.58

Annexes
50
Analyse de la variance de l'influence de la source d'azote sur la croissance mycélienne
de 17 isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 52.46 18 2.91 8.07
Var. factorielle 26.33 2 13.16
Var. résiduelle 26.13 16 1.63
Analyse de la variance du comportement variétal de 11 cultivars de pois chiche vis-à-
vis 03 isolats de FOC
SCE ddl SCE/ddl Fc=Var f/Var r
Var. total 12835.65 12 1069.63 9868.55
Var. factorielle 12829.13 2 6414.56
Var. résiduelle 6.52 10 0.65

45
اىميخص
- عه ذمشىد- ذيمغان )مه خاله اىمغخ اىذقي ىمضاسع اىذمص ف اىشماه اىغشت ىيجضائش
نن مصش االورشاس عىذ ذفش اىمىار FOCاعرىرجىا أن مشض اىزته اىعائ اىزي غثث اىفطش (مغرغاوم
أعشاض . إر جذوا أن شذج اىمشض خطسذ مشذفعح خصصا ف مىطقح عه ذمشىد ذيمغان .ياىمالئم ه
اىمشض اىر شاذواا ف اىذقو ذمصيد ف اصفشاس مرأخش ظش عيى شنو اصفشاس عائ رطس مه األعفو
. إىى األعيى
عضه اىرعشف عيى زا اىفطش مخثشا أظشخ ىىا ها هاف وفظ اىقد،اىرقىاخ اىمخريفح اىر اعرمذن
دساعح اىممضاخ اىمسفىجح أظشخ جد . FOC عالىح مه اىفطش 17تعط اىماصفاخ اىمشفىجح ىــ
شالز أشناه اخرالفاخ اظذح ف اىشنو اىصثغح ته اىفطشاخ اىافعح اىثاىغح مزىل اىذساعح اىثمرشح
. أظشخ اخرالفاخ اظذح ته أتعاد اىجشاشم اىنوذح
تعذ أن أجشىا اخرثاس مخثشاStreptomyces تعذ أن اعرطعىا اخراس اىرعشف عيى عرح عالالخ مه
Streptomyces ، اىغالىح FOCقذسج زي اىغالالخ سشاداذا عيى ذصثط وم شالز عالالخ مه اىفطش
ACT2ف اىثد اىثالعرن ىذماح شالز ا اىر أظشخ وغة مؤح معرثشج ف ذصثط اىفطش اخرثشواا مجذد
اىىرائج اىمذصو عيا أظشخ قذسج زي PPC-25, Col27, ILC-482, :أصىاف مه وثاخ اىذمص
. FO شالز عالالخ مه اىـب دقىم األصىاف اىصالشح مه مشض اىزته اىعائ تعذدماحاىغالىح عيى
منىىا مه اىذصه عيى وغة ILC-482 اخرثاس اىششاعح عيى صىف مه اىذمص شذذ اىذغاعح
عالالخ قييح 06: ىيفطش إىى 17 عيى زا األعاط قغمىا اىغالالخ اىـ 92.12% إىى32.33% ذرشاح ته
. فقط شذذج اىششاعح02 ششعح 07, اىششاعح ، عالىره مرعطح اىششاعح
04 ىىا تاخراس عمخ FOC مهعالالخ 03صىف مه وثاخ اىذمص عىذ إصاتر تـ 11 دساعح عيك
-F7-43, F10-13, Fedjoj, 2G: مشض اىزته اىعائ م عيى اىراىهأصىاف مه وثاخ اىذمص مقامح
03 .
,’Fusarium oxysporum f. sp. ciceri’ اىرعاد’ اىششاعح’ مشض اىزته اىعائ:الكلمات المفتاحية
Actinomycètes, Cicer arietinum L.

46