Embed Size (px)
Citation preview
Multiscale model of the lac operon.
Villa et al. (2005) Structural dynamics of the lac repressor-DNA complexrevealed by a multiscale simulation. PNAS 102: 6783-6788.
Background:
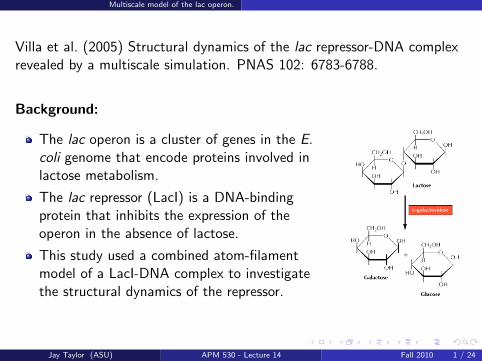
The lac operon is a cluster of genes in the E.coli genome that encode proteins involved inlactose metabolism.
The lac repressor (LacI) is a DNA-bindingprotein that inhibits the expression of theoperon in the absence of lactose.
This study used a combined atom-filamentmodel of a LacI-DNA complex to investigatethe structural dynamics of the repressor.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 1 / 24

Multiscale model of the lac operon.
The lac operon contains three protein-coding genes:
lacZ encodes β-galactosidase, which cleaves lactose into galactoseand glucose;
lacY encodes a lactose permease, which pumps lactose into the cell;
lacA encodes a thiogalactoside transacetylase.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 2 / 24

Multiscale model of the lac operon.
The lac genes are co-regulated.
The three lac genes share a common promoter and are transcribedas a single polycistronic mRNA.
The lac operon is transcribed at very low levels whenever lactose isabsent. (This is thought to prevent the wasteful production ofenzymes that aren’t needed.)
The operon is also transcribed at low levels when glucose is present,even if lactose is also present. (Glucose may be a preferred source ofenergy.)
The lac operon is transcribed at high levels if lactose is present andglucose is absent.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 3 / 24
Multiscale model of the lac operon.
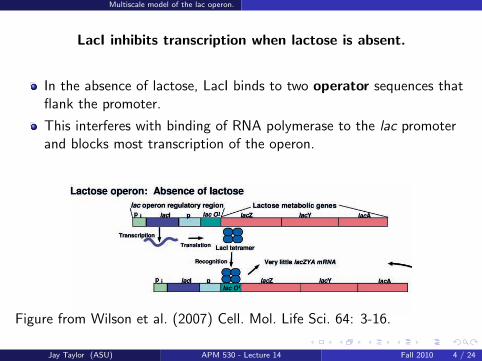
LacI inhibits transcription when lactose is absent.
In the absence of lactose, LacI binds to two operator sequences thatflank the promoter.
This interferes with binding of RNA polymerase to the lac promoterand blocks most transcription of the operon.
Figure from Wilson et al. (2007) Cell. Mol. Life Sci. 64: 3-16.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 4 / 24
Multiscale model of the lac operon.
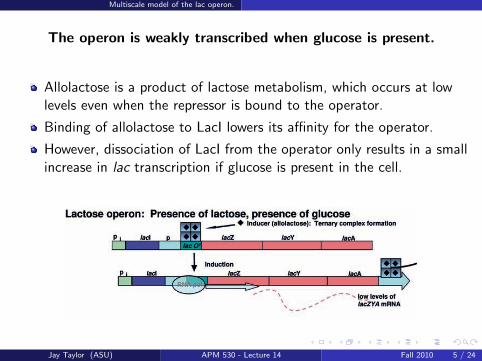
The operon is weakly transcribed when glucose is present.
Allolactose is a product of lactose metabolism, which occurs at lowlevels even when the repressor is bound to the operator.
Binding of allolactose to LacI lowers its affinity for the operator.
However, dissociation of LacI from the operator only results in a smallincrease in lac transcription if glucose is present in the cell.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 5 / 24
Multiscale model of the lac operon.
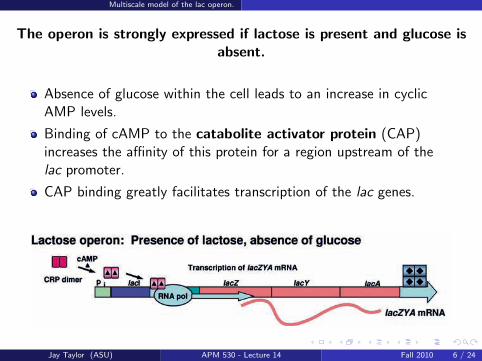
The operon is strongly expressed if lactose is present and glucose isabsent.
Absence of glucose within the cell leads to an increase in cyclicAMP levels.
Binding of cAMP to the catabolite activator protein (CAP)increases the affinity of this protein for a region upstream of thelac promoter.
CAP binding greatly facilitates transcription of the lac genes.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 6 / 24
Multiscale model of the lac operon.
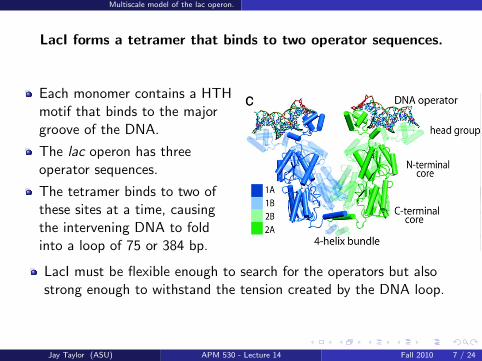
LacI forms a tetramer that binds to two operator sequences.
Each monomer contains a HTHmotif that binds to the majorgroove of the DNA.
The lac operon has threeoperator sequences.
The tetramer binds to two ofthese sites at a time, causingthe intervening DNA to foldinto a loop of 75 or 384 bp.
LacI must be flexible enough to search for the operators but alsostrong enough to withstand the tension created by the DNA loop.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 7 / 24

Multiscale model of the lac operon.
The lac operon functions as a metabolic switch.
Bistability occurs in thepresence of lactose: theoperon can be on or off.
Switching is triggered bydissociation of LacI fromboth operator sequences.
Fig. 4 from Choi et al. (2008), Science 322: 442-446.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 8 / 24

Multiscale model of the lac operon.
Multiscale model of a LacI-operator complex.
The aim of this study was to investigate the effects of the DNA loop onthe structural dynamics of the LacI-operator complex. However, MDsimulations face two problems:
The complete solvated LacI-operator system would contain between700,000 and two million atoms, mainly contributed by the solvent.
The loop dynamics occur on timescales of microseconds or longer,while all-atom MD models can be used to simulate nanosecond-scaleprocesses.
The authors’ approach is to use an all-atom model of the LacI boundto the recognition sites but model the intervening loop as an elasticribbon.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 9 / 24

Multiscale model of the lac operon.
Molecular Dynamics Protocol
The initial LacI-DNA complex was built from several crystalstructures and embedded in a periodic box of water and salt.
Two systems were simulated - one containing 226,314 atoms and theother containing 314,452 atoms. The difference is due to size of thewater box and the number of salt molecules.
Each system was simulated at constant temperature T = 300 K andpressure P = 1 atm.
The smaller system was simulated for 22.4 ns, while the larger wassimulated for 17 ns.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 10 / 24

Multiscale model of the lac operon.
Elastic Rod Model of DNA
The 75 bp loop was modeled as an elastic rod connected to theterminal base pairs of the LacI binding sites.
The equilibrium structure of the loop was determined by numericallysolving the Kirchoff equations of elasticity.
The Kirchhoff equations describe the kinematics of an elastic rod:each set of boundary conditions determines a single, equilibriumstructure which balances the elastic forces caused by bending andtwisting at every cross-section.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 11 / 24
Multiscale model of the lac operon.
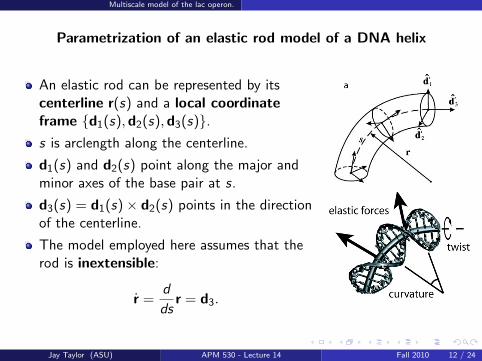
Parametrization of an elastic rod model of a DNA helix
An elastic rod can be represented by itscenterline r(s) and a local coordinateframe d1(s),d2(s),d3(s).s is arclength along the centerline.
d1(s) and d2(s) point along the major andminor axes of the base pair at s.
d3(s) = d1(s)× d2(s) points in the directionof the centerline.
The model employed here assumes that therod is inextensible:
r =d
dsr = d3.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 12 / 24

Multiscale model of the lac operon.
Curvature and Twist
The angular velocity of the local coordinate frame can be written as
di = k× di
where
k = (K1,K2,Ω) is the vector of strains;
K1(s) and K2(s) are the curvatures;
Ω(s) is the local twist.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 13 / 24

Multiscale model of the lac operon.
At equilibrium, the elastic forces N and torques M balance the externalforces f and torques g acting at each point of the centerline:
N + f = 0
M + g + r×N = 0
Here N =∑
i Nidi and M =∑
i Midi , where
N1,N2 are the shear forces;
N3 is the force of tension or compression;
M1,M2 are the bending moments of the principal axes;
M3 is the twisting moment.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 14 / 24

Multiscale model of the lac operon.
The Bernoulli-Euler approximation assumes that the elastic torquedepends linearly on the local curvatures and the twist:
M = A1κ1d1 + A2κ2d2 + Cωd3
where
A1,A2 are the bending rigidities;
C is the twisting rigidity;
κi (s) = Ki (s)− κ0i (s) is the deviation from the intrinsic curvature
of the rod;
ω(s) = Ω(s)− ω0(s) is the deviation from the intrinsic twist of therod.
The intrinsic curvatures and twist are the values adopted by anenergy-minimizing rod in the absence of any external forces.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 15 / 24

Multiscale model of the lac operon.
Kirchhoff’s equations characterize the geometry of an elastic rod atequilibrium under the Bernoulli-Euler approximation.
These can be written as a 13-dim system of nonlinear ODEs: seeeqns (2.39) - (2.49) in Villa et al. (2004); Multiscale Model. Simul.2: 527-553.
The equations can either be derived directly from the local balanceconditions on the force and torque, or from the Euler-Lagrangeequations for the energy functional:
U =1
2
∫ L
0
[β1(s)κ2
1(s) + β2(s)κ22(s) + γ(s)ω2(s)
]ds.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 16 / 24

Multiscale model of the lac operon.
Coupling of the MD and DNA Loop Model
The loop model was coupled to the MDsimulation by applying the forces generatedby the elastic stress and torque of loopingto the operator ends.
The equilibrium loop structure wasrecomputed every 10 ps using B.C.’sderived from the last MD structure.
Thermal fluctuations of the DNA loop weremodeled by subjecting the loop to acontinuously varying Gaussian force ofmagnitude 80 pN.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 17 / 24

Multiscale model of the lac operon.
Geometry of the LacI-Operator Complex
Several summary statistics were monitoredduring the simulations:
the cleft angle α measures the openingbetween the two dimer arms;
θi is the altitudinal angle of head group i ;
φi is the azimuthal angle of head group i .
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 18 / 24
Multiscale model of the lac operon.
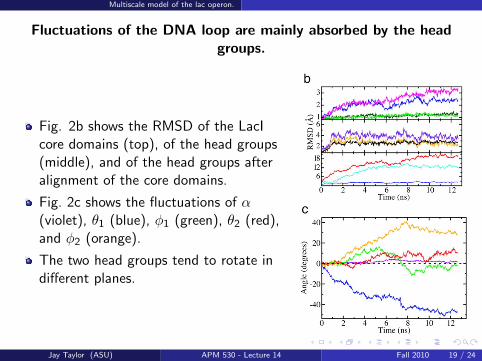
Fluctuations of the DNA loop are mainly absorbed by the headgroups.
Fig. 2b shows the RMSD of the LacIcore domains (top), of the head groups(middle), and of the head groups afteralignment of the core domains.
Fig. 2c shows the fluctuations of α(violet), θ1 (blue), φ1 (green), θ2 (red),and φ2 (orange).
The two head groups tend to rotate indifferent planes.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 19 / 24
Multiscale model of the lac operon.
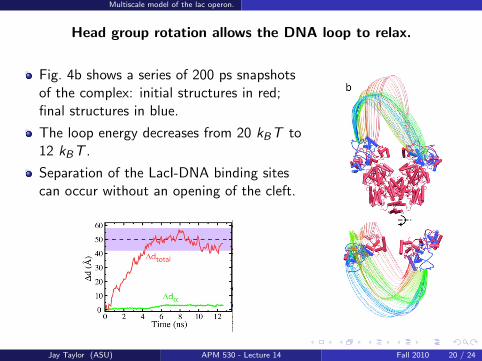
Head group rotation allows the DNA loop to relax.
Fig. 4b shows a series of 200 ps snapshotsof the complex: initial structures in red;final structures in blue.
The loop energy decreases from 20 kBT to12 kBT .
Separation of the LacI-DNA binding sitescan occur without an opening of the cleft.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 20 / 24
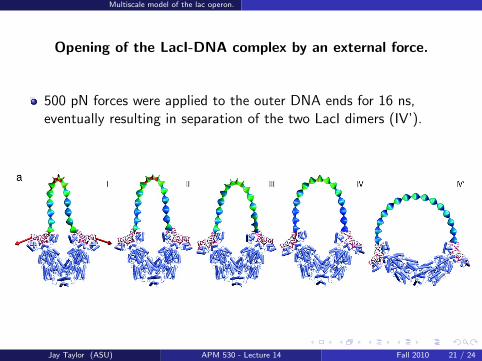
Multiscale model of the lac operon.
Opening of the LacI-DNA complex by an external force.
500 pN forces were applied to the outer DNA ends for 16 ns,eventually resulting in separation of the two LacI dimers (IV’).
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 21 / 24
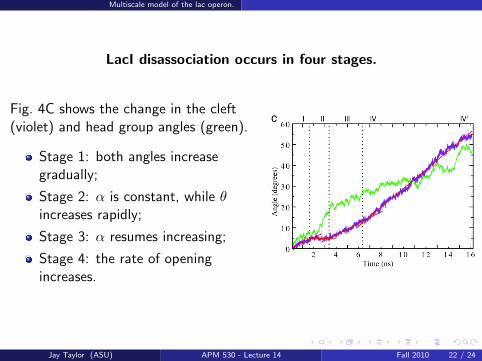
Multiscale model of the lac operon.
LacI disassociation occurs in four stages.
Fig. 4C shows the change in the cleft(violet) and head group angles (green).
Stage 1: both angles increasegradually;
Stage 2: α is constant, while θincreases rapidly;
Stage 3: α resumes increasing;
Stage 4: the rate of openingincreases.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 22 / 24

Multiscale model of the lac operon.
The LacI tetramer is stabilized by several intermolecularinteractions.
Salt bridges E2351B − R3262A and E2352B − R3261A form duringStage II and then rupture during Stage III.
A charge-dipole interaction between R3511A and the α-helix2472B − 2602B is disrupted during Stage III.
Hydrophobic interactions between two α-helices, 2221B − 2361B and2222B − 2362B disappear by Stage IV.
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 23 / 24

Multiscale model of the lac operon.
Summary
The mobility of the LacI head groups allows the enzyme to withstandthe substantial stresses generated by the DNA loop formed betweenthe two operator binding sites.
Several salt bridges and helix-helix interactions stabilize the Vconfiguration of the LacI tetramer during repression of the lac operon.
The dynamics of large protein-DNA complexes can be studied usingmultiscale models that couple atomic-level MD to mechanicaldescriptions of DNA loops.
However, the disparity of the timescales of these different processes isnot clearly addressed in this paper: can one simply accelerate themotion of the loop as done here?
Jay Taylor (ASU) APM 530 - Lecture 14 Fall 2010 24 / 24





























