Upload
tomluigi
View
223
Download
0
Embed Size (px)
Citation preview
8/9/2019 Whiten Et Al (2009)
1/13
doi: 10.1098/rstb.2009.0069, 2417-24283642009Phil. Trans. R. Soc. B
Andrew Whiten, Nicola McGuigan, Sarah Marshall-Pescini and Lydia M. Hopper for child and chimpanzeeEmulation, imitation, over-imitation and the scope of culture
Supplementary datamlhttp://rstb.royalsocietypublishing.org/content/suppl/2009/07/10/364.1528.2417.DC1.ht
"Data Supplement"
Referenceshttp://rstb.royalsocietypublishing.org/content/364/1528/2417.full.html#ref-list-1
This article cites 49 articles, 6 of which can be accessed free
Rapid responsehttp://rstb.royalsocietypublishing.org/letters/submit/royptb;364/1528/2417
Respond to this article
Subject collections
(883 articles)evolution (172 articles)cognition (640 articles)behaviou r
Articles on similar topics can be found in the following collections
Email alerting service hereright-hand corner of the article or clickReceive free email alerts when new art icles cite this article - sign up in the box at the top
http://rstb.royalsocietypublishing.org/subscriptionsgo to:Phil. Trans. R. Soc. B To subscribe to
This journal is 2009 The Royal Society
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/content/suppl/2009/07/10/364.1528.2417.DC1.htmlhttp://rstb.royalsocietypublishing.org/content/suppl/2009/07/10/364.1528.2417.DC1.htmlhttp://rstb.royalsocietypublishing.org/content/364/1528/2417.full.html#ref-list-1http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.html#ref-list-1http://rstb.royalsocietypublishing.org/letters/submit/royptb;364/1528/2417http://rstb.royalsocietypublishing.org/cgi/collection/evolutionhttp://rstb.royalsocietypublishing.org/cgi/collection/evolutionhttp://rstb.royalsocietypublishing.org/cgi/collection/cognitionhttp://rstb.royalsocietypublishing.org/cgi/collection/behaviourhttp://rstb.royalsocietypublishing.org/cgi/alerts/ctalert?alertType=citedby&addAlert=cited_by&saveAlert=no&cited_by_criteria_resid=royptb;364/1528/2417&return_type=article&return_url=http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.pdfhttp://rstb.royalsocietypublishing.org/cgi/alerts/ctalert?alertType=citedby&addAlert=cited_by&saveAlert=no&cited_by_criteria_resid=royptb;364/1528/2417&return_type=article&return_url=http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.pdfhttp://rstb.royalsocietypublishing.org/cgi/alerts/ctalert?alertType=citedby&addAlert=cited_by&saveAlert=no&cited_by_criteria_resid=royptb;364/1528/2417&return_type=article&return_url=http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.pdfhttp://rstb.royalsocietypublishing.org/subscriptionshttp://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/subscriptionshttp://rstb.royalsocietypublishing.org/cgi/alerts/ctalert?alertType=citedby&addAlert=cited_by&saveAlert=no&cited_by_criteria_resid=royptb;364/1528/2417&return_type=article&return_url=http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.pdfhttp://rstb.royalsocietypublishing.org/cgi/collection/evolutionhttp://rstb.royalsocietypublishing.org/cgi/collection/cognitionhttp://rstb.royalsocietypublishing.org/cgi/collection/behaviourhttp://rstb.royalsocietypublishing.org/letters/submit/royptb;364/1528/2417http://rstb.royalsocietypublishing.org/content/364/1528/2417.full.html#ref-list-1http://rstb.royalsocietypublishing.org/content/suppl/2009/07/10/364.1528.2417.DC1.htmlhttp://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
2/13
Emulation, imitation, over-imitation and thescope of culture for child and chimpanzeeAndrew Whiten 1, * , Nicola McGuigan 2, Sarah Marshall-Pescini 1,3
and Lydia M. Hopper 1,41 Centre for Social Learning and Cognitive Evolution, and Scottish Primate Research Group,
School of Psychology, University of St Andrews, St Andrews KY16 9JP, UK 2 Department of Psychology, Heriot Watt University, Edinburgh EH14 4AS, UK
3 Department of Biomedical Science and Technology, University of Milan, Milan, Italy4 Department of Psychology, University of Durham, Durham DH1 3LE, UK
We describe our recent studies of imitation and cultural transmission in chimpanzees and children,which question late twentieth-century characterizations of children as imitators, but chimpanzees asemulators. As emulation entails learning only about the results of others actions, it has beenthought to curtail any capacity to sustain cultures. Recent chimpanzee diffusion experiments
have by contrast documented a signicant capacity for copying local behavioural traditions.Additionally, in recent ghost experiments with no model visible, chimpanzees failed to replicatethe object movements on which emulation is supposed to focus. We conclude that chimpanzeesrely more on imitation and have greater cultural capacities than previously acknowledged. However,we also nd that they selectively apply a range of social learning processes that include emulation.Recent studies demonstrating surprisingly unselective over-imitation in children suggest thatchildrens propensity to imitate has been underestimated too. We discuss the implications of these developments for the nature of social learning and culture in the two species. Finally, ournew experiments directly address cumulative cultural learning. Initial results demonstrate a relativeconservatism and conformity in chimpanzees learning, contrasting with cumulative cultural learn-ing in young children. This difference may contribute much to the contrast in these speciescapacities for cultural evolution.
Keywords: imitation; emulation; social learning; cultural transmission;cumulative culture; chimpanzees
1. INTRODUCTIONStudies of imitation in children and in our closest pri-mate relatives have now spanned a century ( Witmer1909 ), including over 60 experimental reports forgreat apes ( Tomasello & Call 1997 ; Whiten et al .2004 ) and countless others with children ( Hurley &Chater 2005 ).
The last quarter of the twentieth century saw an
upturn in the rigour and intensity of the ape research,stimulated by contentious theoretical and empiricalcontributions. New experimental studies and critiquesof previous research questioned the commonly acceptedand intertwined views that research had demonstratedthat apes ape (i.e. imitate) and exhibit cultures inthe wild ( Tomasello et al . 1987 ; Galef 1988 , 1992 ;Tomasello 1990 , 1996 ; Heyes 1993 ). Later studiesemploying more discriminating methodologies thanbefore generated a number of inuential ndings,several of which converged on a subsequently inuential
conclusion: that unlike children, apes are not true imita-tors but rather are emulators ( Tomasello 1996 ).Whereas in imitation an individual learns by copyingthe actions of another, in emulation they learn insteadabout the environmentfor example, about the resultsof what others do or about the relevant properties of tools and other objects involved. Tomasello & Call(1997) , reviewing experiments by Nagell et al . (1993) ,
Whiten et al . (1996) and others, concluded that It isnot likely that either monkeys or apes who have nothad extensive human contact imitatively learn theinstrumental behaviors of others. What are salient formonkeys and apes in such situations are the tool, thefood, and their spatial temporal causal relations.This is intelligent problem-solving behaviour, but it isnot attempting to reproduce the behavior or behavioralstrategies of another animate being (p. 294). Thatchildren imitate, by contrast, was not disputed.
In this paper, we describe more recent experimentsthat challenge this dichotomous view of the nature of social learning in apes and children. We conclude that
the evidence shows instead that both chimpanzees andchildren possess a portfolio of different social learningmechanisms, including both imitation and emulation,that are deployed selectively in different contexts. Thiswork indicates that copying what others do plays a
* Author for correspondence ( [email protected] ).Electronic supplementary material is available at http://dx.doi.org/10.1098/rstb.2009.0069 or via http://rstb.royalsocietypublishing.org .
One contribution of 13 to a Theme Issue Evolution, developmentand intentional control of imitation.
Phil. Trans. R. Soc. B (2009) 364 , 24172428doi:10.1098/rstb.2009.0069
2417 This journal is q 2009 The Royal Society
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
mailto:[email protected]://dx.doi.org/10.1098/rstb.2009.0069http://dx.doi.org/10.1098/rstb.2009.0069http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://dx.doi.org/10.1098/rstb.2009.0069http://dx.doi.org/10.1098/rstb.2009.0069http://dx.doi.org/10.1098/rstb.2009.0069mailto:[email protected]://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
3/13
more signicant role in both species than hithertoappreciated. We acknowledge, and indeed demonstrate,considerable differences in the social learning propensi-
ties of the two species, but we are led to conclusionsthat go beyond characterizing the difference as a simpleimitation/emulation dichotomy.
2. DEFINING, DISSECTING AND IDENTIFYINGIMITATION AND EMULATIONImitation and emulation are two processes withinlarger taxonomies of social learning ( Whiten & Ham1992 ; Heyes 1994; Whiten et al . 2004 ) but they areour foci here, so we dwell on their denitions. Doingso is important because the two processes are distin-guished in several quite different ways in the sociallearning literature. It is important to recognize a dis-tinction between debates about species differencesthat hinge on: (i) variations in the way a conceptsuch as imitation is dened versus (ii) for any givendenition, the adequacy of relevant empirical evidencein the species of interest. Only such evidence canprovide objective, scientic conclusions. Matters of denition, by contrast, cannot be resolved by research.The best we can hope for is some consensus in theterms we use; it would be naive to assume they havesome inherently correct meaning. Given the scopefor confusion, every study needs to dene its keyconcepts.
(a ) Dening and dissecting imitationFollowing Whiten & Ham (1992) , gure 1 offers a leandenition of imitation as copying the form of anaction. We follow this denition here. However,
elsewhere in the social learning literature, additionalcriteria are set. This means that imitation has ineffect been dissected into different subcategories.
For example, where the distinction noted above ismade, between imitation as copying an agentsactions and emulation as learning from the environ-mental results of what an agent does (emulation:Tomasello 1996 ), imitation becomes essentially bodilyimitation. The authors are free to restrict the scopeof imitation in this way, but we note at least two draw-backs in doing so, which can be illustrated by theexample of copying somebodys actions with a tool,such as a hammer. The rst concern is simply thatexcluding the movements of the hammer that arebeing copied from our conception of imitation seemsto lose touch with the way the term imitation is
applied in everyday life. The second, more importantscientic concern is that it remains unclear to whatextent copying the actions of objects that peoplemove (e.g. hammering, where the tool becomes anextension of the limb) is a fundamentally differentprocess than copying bodily actions per se (see Huberet al . 2009 ). Accordingly, in the present paper, whenwe discuss evidence for imitation, we may considerthis either in the restricted sense of bodily imitationor in the broader sense of copying the form of anaction that may go beyond the skin to include toolsand other objects. Such issues are analysed in depthby Whiten et al . (2004) .
An additional criterion for imitation required bysome authors is that the action copied be novel, orotherwise improbable ( Thorpe 1963 ). In this case,we are referring to imitative learning , rather than imita-tion in the broader, everyday sense that may include
sociallearning
copyingcopying what
anotherindividual does
affordance learninglearning about operating characteristics of
objects or environment
other forms of social learning
of properties
resultemulationgoalemulation
shapeimitationcopying the form of
an action
imitative
emulative
objectmovementre-enactmentcopying the form of a caused objectmovement
end-stateemulationcopying only the end or
outcome of an action sequence
sequential
structure
hierarchical
structure
causal links
intentional links
of relationships
of functions
Figure 1. A taxonomy of imitative and emulative learning processes. Here, senses of emulation distributed through the sociallearning literature are distinguished and fall under the headings of object movement re-enactment, end-state emulation andaffordance learning. Simpler forms of social learning, such as stimulus enhancement, are omitted here (see Whiten et al .2004 for a full taxonomy), as are related forms of social inuence, such as contagion (see Whiten & Ham 1992 ). For furtherexplanation, see text, particularly 2b.
2418 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
4/13
familiar actions in the copiers own repertoire. Thatsomething new is learned is an important requirementwhere interest focuses on the acquisition of elements of behaviour that constitute a local culture. Accordingly,we also recall this distinction in our analyses of imitation below.
( b ) Dissecting and dening emulationEmulation has come to be used to refer to processeseven more diverse than is the case for imitation(Byrne 2002 ), a subject also analysed in greaterdetail by Whiten et al . (2004) than there is scope forhere. At rst sight, this is confusing, but if the prolifer-ation reects interesting distinctions, real progress maybe made in charting the landscape of social learning.Three main senses of emulation are distinguished ingure 1 .
(i) Object movement re-enactment If imitation is limited to bodily movements only, copy-ing how a model makes an object such as a hammermove comes within the domain of emulation. Thissense of emulation was called object movementre-enactment (OMR) by Custance et al . (1999) ,underlining the similarity to copying the form of bodily actions. One can envisage a causal continuumfrom bodily motions to object movements directlymoved by the body (e.g. hammer use) to effects onthe environment caused by these (nuts cracked by thehammer). Copying could in principle pick out andre-enact elements anywhere along this causal se-quence. Ascription of imitation is most clearly apt at
the proximal end of this continuum, whereas theconcept of emulation is the clearest more distally.The category of OMR in gure 1 is accordingly anno-tated to indicate this continuum and the ambiguities itmay occasion, relevant in studies described below.
(ii) End-state emulationEmulation was originally distinguished from imitationby Wood (1989) , who noted instances where childrenachieve common goals to those modelled, but do so byusing idiosyncratic means that were never observed(p. 72). In gure 1 , this is classed within copying,but it is only the end-state(s) of what the model hasdone that is copied. As Wood suggested, copiers mayreach this end in their own way. However, they mighthappen to choose the same method as the model,which might thus be falsely recorded as imitation.
(iii) Affordance learning A nal alternative is that learning is instead aboutproperties of objects or other aspects of the environ-ment, which may help the learner in tackling the taskobserved without employing imitation. Such affor-dances of the physical context may include propertiesof objects such as brittleness, relationships between
objects such as containment or functions such aswhat a tool is good for ( Klein & Zentall 2003 ).Having recognized such affordances, the observermay then exploit them in their own behavioural strat-egies; these might happen to match the actions of
the model or not, but if they do, it would not bethrough imitation.
(c ) Chimpanzees as emulatorsThe reason that emulation has come to prominence insocial learning research and that chimpanzees havebeen characterized as emulators began with an early
study of Tomasello et al . (1987) . In this experiment,chimpanzees exposed to a model using a stick tool torake in food were more likely to acquire this behaviourthan controls that saw no model, but they did not copya technique the model had developed to gain foodfrom awkward places. Tomasello et al . interpretedthis as a failure to imitate. Because control chimpan-zees handled the sticks but did not apply them, theauthors suggested that the success of the observerchimpanzees depended on more than stimulusenhancement (attracting of attention to the relevantobjects); the subjects appeared to have learned byobservation that the stick afforded use as a tool.
Tomasello (1990) adopted Woods (1989) termemulation as a label for this. Reviews of severallater studies have tended to conrm this picture,with chimpanzees failing to copy details of a modelsactions (especially in comparison to childrens imita-tion), and performing instead in ways consistent withemulation ( Tomasello & Call 1997 ; Whiten et al .2004 ; Call et al . 2005 ). Such low-delity copyingwas suggested to severely constrain the capacity of chimpanzees for the cultural transmission of traditions(Tomasello 1999 ).
However, few attempts have been made to directlytest for emulation; instead, emulation has typically
been offered as a default explanation when there islittle or no evidence of detailed imitative copying.Accordingly, results can often be equally well des-cribed as consistent with imitation at a relativelycrude level. Thus, in the Tomasello et al . experimentoutlined earlier, the chimpanzees may have imitatedthe use of a stick as a rake, but without imitating cer-tain details of the technique. In the two sections thatfollow, we describe two sets of studies that challengethis characterization of chimpanzees as emulatorslikely to show little evidence of cultural transmission.First, we describe recent diffusion experimentsdesigned to track cultural transmission, then ghost
experiments that directly test for emulation.
3. COPYING FIDELITY IN SOCIALTRANSMISSION: DIFFUSION EXPERIMENTSThe majority of social learning experiments arerestricted to what a single observer learns fromwatching a single model. However, multiple culturaltransmissions are required to sustain traditions. Prop-erly investigating any animals cultural capacity thusrequires a different kind of experiment, in which de-lity of transfer is tracked over repeated transmissionevents. Just over 30 of these diffusion experiments
have now been completed in a variety of taxa includingsh and birds ( Whiten & Mesoudi 2008 ), but a corpusof such studies in non-human primates has been builtup only in the last few years ( Whiten et al . 2007 ).Likewise, only recently has a handful of such studies
Imitation and over-imitation A. Whiten et al. 2419
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
5/13
been completed with children ( Horner et al . 2006 ;Flynn 2008 ; Flynn & Whiten 2008 ; McGuigan &Graham in press ).
Six controlled diffusion studies have been publishedfor chimpanzees. Whiten et al . (2005 a ) trained a singlechimpanzee from one group to use one of the twotechniques to extract trapped food from an articialforaging device (the panpipes) and trained a chim-panzee from a different group to use an alternativetechnique. One technique ( lift ) involved using a stickto lift up an obstacle so that trapped food was releasedto the chimpanzee; in the second technique ( poke), thetool was instead inserted into an obscured opening,pushing the obstacle until the food fell onto a lowerlevel of the panpipes and rolled out to the chimpanzee.Each chimpanzee was then re-introduced to its 16-strong group and the task made available to all. Eachtechnique spread preferentially in the group in whichit was seeded, with a majority matching the techniqueused by the initial local model.
A further study involved an arbitrary convention posting plastic tokens in either of two differentreceptacles. Again, each method spread in the group inwhich it was seeded, with the exception of a single chim-panzee that discovered the method used in the othergroup ( Bonnie et al . 2007 ). This and the panpipe studyillustrate an open-diffusion design, in which whowatches whom and who tackles the task next is leftopen. These approaches were complemented by a trans-mission chain experiment in which transfer wasconstrained to a linear chain, A ! B ! C ! D, etc. Eachof the two techniques for opening a foraging box wastransmitted along a chain with high delity, such that
after six transfers, the last individual in each chain wasusing the same technique seeded at the start ( Horneret al . 2006 ).
Further diffusion experiments have examinedgroup-to-group transfer, an essential element of anyputative cultures that span hundreds of kilometres(and thus scores of communities) in Africa ( Whitenet al . 1999 ; Whiten 2005 ). For two different articialforaging tasks, each of the alternative techniquesseeded in different groups spread to a second andthird group, with high delity ( Whiten et al . 2007 ;see gure 2 ). For the most complex task, requiring asequence of two different operations, the original tech-
nique spread across up to 26 chimpanzees in threegroups, with none successfully performing thesequence customary in the other three groups.
An important conclusion of this set of studies is thatthe social learning processes at work are demonstrablyadequate to sustain multiple different traditions acrossdifferent communities, including quite complex vari-ations in foraging techniques. In relation to debatesabout the limits on imitation in apes and the impli-cations of these limits for culture, this is perhaps amore important conclusion than precisely what termsare agreed apt to describe the underlying learningprocesses.
That said, we can turn to a more focused analysis of the evidence for imitation in these studies, recalling thetwo supplementary criteria for imitation favouredby some authors and outlined above: (i) whether thecopied pattern is novel and thus truly acquired through
observation and (ii) whether bodily imitation isimplicated.
(a ) Are the techniques acquired novel or improbable?To experimentalists, the principal way of operationaliz-ing this question is through control conditions; couldchimpanzees succeed if no model were available?Such tests were incorporated into all of the exper-iments outlined above, with variable results thatplausibly correspond to the relative simplicity andlikely familiarity to the chimpanzees of the elementsof action involved. In the studies of Horner et al .(2006) and Bonnie et al . (2007) , the tasks weremotorically relatively simple, such as sliding or liftinga door to access food, and some chimpanzees solvedthe task in the no-model condition. These studiesoffered robust evidence of cultural transmission yetcalled on relatively familiar elements of action. Inthree diffusion studies that employed more challengingtasks (Whiten et al . 2005 a , 2007 ), chimpanzees inno-model conditions were not able to solve the taskthemselves. In different studies as many as 56chimpanzees have been presented with the panpipestask individually or in small groups, without solvingit: the techniques involved are thus to this extentinherently improbable (cf. Tennie et al . 2009 ).
In one larger group, a single chimpanzee didsucceed by her own efforts, using the poke method(Hopper et al . 2007 ). One solver in 56 chimpanzeesdoes not, however, amount to the technique beingwithin chimpanzees repertoire: to the contrary, a
ratio of independent solving as low as 1/56 isconsistent with a cultural scenario that includes rareinnovators, and a large majority that can only acquirethe innovation by copying it or through some otherbrand of social learning ( gure 3 ).
( b ) Is bodily imitation implicated?Most diffusion studies have employed only a singleinitial model and compared this with a control con-dition with no model ( Whiten & Mesoudi 2008 ).However, the chimpanzee studies described above allused a two-action design (e.g. poke versus lift in the
panpipes) that earlier played an important role in therising methodological sophistication of dyadic sociallearning studies ( Heyes 1996 ; Huber et al . 2009 ).Because each of the two different techniques spreadsin its respective group, they are by denition being dif-ferentially replicated or copied through the group, andin this sense can be described as broadly imitative.However, if we focus instead on the criterion of bodily imitation, then these results do not demonstratethis, because all the tasks involved object manipulationand it is the movements of these that may be beingemulated. The diffusion studies were not designed todistinguish such potential mechanisms; rather, the
goal was to examine the extent to which chimpanzeeswould sustain different traditions, whatever the under-lying mechanisms. To discriminate the latter, weconducted supplementary experiments, discussed in5. Before leaving the diffusion studies, however, we
2420 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
6/13
consider briey their broad implications for the scopeof cultural transmission in chimpanzees.
4. THE SCOPE FOR CULTURAL TRANSMISSIONIN CHIMPANZEES AND OTHER ANIMALSIn a previous paper ( Hopper et al . 2007 ), we concep-tualized the results of our diffusion experiments interms of learning thresholds, shown in simplediagrammatic form in gure 3 , partly to facilitateengagement with a partner paper 1 in the presentissue ( Tennie et al . 2009 ). In gure 3 , two thresholdsdivide a continuum expressing the degree of challengethat any particular task presents to the speciesconcerned. Below the lower threshold, the task issufciently easy that most individuals can acquire it
through their own efforts. Above this threshold, indi-vidual mastery becomes rarer, but so long as one ormore innovators achieve this, others can succeed bylearning from them. Above the upper threshold of dif-culty, the species simply cannot master the task by
individual effort, nor even by observing a competentmodel of a different species. Our various tasks mapto the three zones in several different ways. The one
innovator, 30
social learners of our panpipes studies(Whiten et al . 2005 a ; Hopper et al . 2007 ) serves as oneapt illustration of the zone conducive to the emergenceof traditions (Zone 2) hypothesized in gure 3 .
Tennie et al . (2009) conceive of a different kind of zone, a zone of latent solutions (ZLS) that theysuggest limits the social learning of which chimpanzeesare capable. This seems a potentially interesting con-cept, but unfortunately the authors do not offer aformal denition and the scope of the concept remainselusive. It is not clear how a ZLS will apply to otherspecies including humans and indeed how one wouldestablish its existence, scope and implications. Tennie
et al . appear to propose that chimpanzee traditionscan incorporate only acts that fall within a ZLSwhere in principle chimpanzees could discover thebehaviour themselves, given conducive conditions.But this seems to correspond simply to the conditions
stab
turn
slide
B2 B3B1
B4 B5 B6
0
50
100
0
50
100
MYKE CE MR TI HU PI ZO PE TA RA MA GE NI JA MO
MI HE BE CO JO UR KA ZIJU BE MO HO JE JA BI MI XE B0 PL AL MAMU SO BE AL TI LE GA
TO PU LY DO K U L U C A E M GL SA AB
lift
(b)(a )
(c)
(d )
Figure 2. Transmission of alternative foraging techniques within and across groups of chimpanzees (after Whiten et al . 2007 ). ( a )Stab technique applied to foraging box, ( b) alternative slide technique, ( c) spread of stab technique(dark grey bars)withingroupB1,followed by spread across groups B2 and B3, and ( d ) spread of slide technique (light grey bars) within and across groups B4B6.Each group was able to watch the performance of the preceding group through large windows. Fidelity of transmission was high.
Imitation and over-imitation A. Whiten et al. 2421
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
7/13
for our tradition zone ( gure 3 ): the acts have to bewithin the range of at least one chimpanzee that canact as innovator, yet above the lower threshold, orthere would be no need for social learning.
Tennie et al . offer an experiment they argue to sup-port their ZLS concept but we do not see how it can.Their study incorporates a loop technique that allchimpanzees failed to learn, when demonstrated by ahuman model. Tennie et al . propose that this is consist-ent with the ZLS hypothesis because if no ape couldmaster the task it is beyond their ZLS and so cannotbe socially learned. But the more parsimoniousexplanation is surely that the task is simply beyondchimpanzees capability (our Zone 3) as, of course,are so many others (likewise, we suspect that severalstudies in the social learning literature are not pro-ductive because at the opposite end in gure 3 , thetask is too elementary to require social learning: ourdiffusion studies have instead focused on discoveringthe true scope of Zone 2 for the species concerned).It appears to be Tennie et al .s sample of Homo sapiensthat conforms more clearly to their ZLS model, for
one young child showed that they could make a loopwithout the benet of a demonstration (so the task iswithin childrens ZLS) and most other childrencould learn it by observation (our Zone 2). The loopstudy thus does not appear to speak to the issue of chimpanzees and humans very different capacitiesfor cumulative cultural evolution. We return to thistopic in 8.
5. TESTING FOR EMULATION THROUGHGHOST EXPERIMENTSWe now shift from diffusion experiments to dyadic
experiments to further dissect underlying social learn-ing processes. One way of testing directly whetheremulation rather than imitation is involved is theghost experiment. The handful of existing studies isreviewed in Hopper et al . (2008) . The underlying
logic is exemplied in Tomasellos (1998) thoughtthat: if a mother chimpanzee rolls a log and eats theinsects underneath, her child will very likely followsuit . . . . (but) the youngster would have learned thesame thing if the wind, rather than the mother, hadcaused the log to roll over and expose the ants (p. 29).
One can experimentally create this impersonal kindof scenario by replacing the model with some hidden(ghostly) means of making the objects of interest dowhat they would if the model were moving them(like the wind in Tomasellos example). Hopper et al .(2007) did this using shing line to discretely pullthe obstacle in our panpipes up, releasing the foodwith no chimpanzee model present. In a secondghost condition, the stick tool was also attached, sothat it too moved up as if a ghostly agent were liftingit. If chimpanzees are emulators, it is precisely theseaspects of the scene they should attend to and learnfrom. Hopper et al . focused on the lift techniquebecause chimpanzees had hitherto acquired this tech-nique only after watching a chimpanzee performing it.
No chimpanzee learned from the ghost demon-strations. This was a startling result, especially in thecase of the ghost with tool presentation, which to ahuman adult, at least, appears rather clearly to givethe game away about what is required. Observer chim-panzees were documented as attending well to thedisplays, yet they failed to learn from them. Such resultsquestion whether chimpanzees are principally or exclu-sively emulators, rather than imitators. Thorndike(1898) dened imitation as learning to do an actfrom seeing it done (p. 50). It seems this is exactlywhat the ghost experiment implies chimpanzees need
to dolearn an act from seeing it done by a conspecicor other adequate animate model.However, the story is more complex. A further
ghost experiment has revealed subtly different results.In this study, the scenario observed was much simpler,because it was based on the one existing positive resultfrom a ghost experiment with a non-human species pigeons. In this, Klein & Zentall (2003) had providedone condition in which pigeons watched a conspecicslide a small panel to left or right to gain food, and aghost condition in which the panel moved with nomodel present. In an additional condition, the panellikewise moved automatically but a pigeon model
was present but merely fed on the food released anddid not push the panel. In this study, pigeonsshowed strong evidence of copying in the activemodel condition and pushed the door in the directionthey had seen it move in the ghost condition. Accord-ingly, the authors concluded emulation learning was atwork. Surprisingly, perhaps, there was no matching inthe condition that had included the pigeon alongsidethe ghost door move.
Hopper et al . (2008) replicated this study as closelyas feasible with both chimpanzees and children. Threemain results are of interest. First, in the chimpanzeemodel condition, subjects matched the direction of
the models pushing in virtually all trials (on average99%). Given how easy it is to push the panel eitherway, one might expect that subjects would explorethis, discover that either approach reveals food, andshow no side preference. The degree of delity to the
Zone 1learning zone
self-learningtypical
task easy task challenging
Zone 3no-go zone
* no innovatorsand/or
* social learningtoo difficult
Zone 2tradition zone
* rare innovators
* remainder learnsocially
Figure 3. A simple model of tradition and adjacent learningzones. The tradition zone (2) is proposed to lie between twothresholds of task complexity as experienced by the speciesof interest. Below the rst threshold, all or most individualscan master the task by their own efforts. Above the higherthreshold, the task is beyond the species capability. In thepart of the tradition zone nearest Zone 3, only rare innova-tors can master the task but others may acquire it throughsocial learning: such traditions will be relatively robust. Inthe part of the tradition zone near Zone 1, traditions maybe relatively short-lived as more individual-level learningoccurs. The transition between Zones 1 and 2 will therefore
be graded.
2422 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
8/13
model is therefore striking. Children responded with100 per cent delity in this condition ( gure 4 ).
In the ghost condition, both children and chimpan-zees initially showed a signicant tendency to matchthe direction of the panel movement. This is the rstconrmation of emulation in chimpanzees (or indeedany non-human primate) in a ghost condition. How-ever, in further trials, chimpanzees (but not children)tended to explore pushing in both directions and nolonger showed matching to what they had seen. This
result is more consistent with the results of the pan-pipes ghost experiment of Hopper et al . (2007) andcontrasts quite starkly with the delity observedwhen a chimpanzee had acted as a model, pushingthe panel. Our conclusion is that where a task is verysimple for a chimpanzee, as in the panel-pushing,emulation may occur, yet be eeting, and where thetask is more complex as in the panpipes, chimpanzeesneed to be able to see the fuller action display of amodel, in order to learn through observation. Thisseems consistent with the demonstration by Voelkl &Huber (2007) of detailed imitative matching by mar-mosets in a functional context where such matching
made a signicant contribution to successful taskcompletion ( Huber et al . 2009 ).The third and nal result that deserves mention is
that in the condition where the panel moved automati-cally but the conspecic model was passive, children,unlike both chimpanzees and pigeons, copied almostas faithfully as with the active model ( gure 4 ). Whychildren did this remains mysterious. Perhaps, it rep-resents a symptom of the extreme cultural conformityof children, to be discussed further below. One cannotstrictly describe the response as imitation, insofar asthe model child actually did nothing!
The results of these ghost experiments have thus
generated several implications for the nature of theimitative and emulative wings of chimpanzees andchildrens social learning. In the next two sections, weconsider further recent experiments bearing on theseprocesses in chimpanzees and children, respectively.
6. THE NATURE OF IMITATION AND SCOPEOF SOCIAL LEARNING IN CHIMPANZEESIf the ghost experiments speak against the simplenotion that chimpanzees are emulators, should wethink that in the diffusion experiments they were imi-tating? In the broad sense, yesinsofar as we showedchimpanzees traditions can replicate , spreading withhigh delity ( gure 2 ). However, if the criterion isbodily imitation, the answer is different. Althoughthe ghost experiments underline the importance tochimpanzees of the involvement of an agent, chimpan-zees may nevertheless be attending preferentially toevents in the neighbourhood of the tool and otherobjects, rather than limb congurations. Alternatively,they might be copying aspects of the form of the actionthat include both body parts and external objects.Below we discuss evidence that bears on these differentpossibilities. We begin, however, with evidence thatchimpanzees have the essential capacities for bodilyimitative matching.
(a ) Bodily imitationIn the Do-as-I-do (DAID) paradigm, the subject rstlearns to match a training set of bodily actions (e.g.touch chin) on the request to Do this!. If thisbasic routine is clearly grasped, testing proceeds to abattery of relatively novel actions. In both chimpanzees(Custance et al . 1995 ) and an orangutan ( Call 2001 ), abattery of 48 such novel actions showed that these apescould be identied in blind coding as copying asignicant number of actions such as touch shoulder/elbow/stomach/thigh/top-of-head; interlink ngers,
clap back of hands, lip smack and hug self. Matchingwith respect to out-of-sight body parts by the chim-panzee subjects was done as well as matching toin-sight parts ( Custance et al . 1995 ). These studiesdemonstrate a capacity for fairly elaborate, if approxi-mate, matching, although they can show only whatchimpanzees can imitate rather than what they willcopy spontaneously.
These results have recently been complemented byremarkable episodes in which, during the course of anexperimental study of the social learning of nut-cracking using a stone hammer ( Marshall-Pescini &Whiten 2008 a ), a young chimpanzee watching another
nut-cracking made repeated and moderately synchro-nous matching actions, but involving no nut orhammer. 2 Such matching or mirroring behaviour israre (we noted seven such episodes), but graphicallydemonstrated a spontaneous ability to match thebodily actions of a conspecic.
In some species, a two-action experimental designhas been used explicitly to test such matching, withmodels trained to act on a manipulandum usingeither of two body parts, such as beak versus foot.In both pigeons ( Zentall et al . 1996 ) and marmosets(Voelkl & Huber 2000 ), signicant matching wasrecorded (true imitation in these papers titles).
Relevant ape data have since become availablethrough an experiment designed to answer the morecomplex question of whether apes imitate rationally(Buttelmann et al . 2007 ). Chimpanzees witnessedhuman models pressing panels using unusual body
1.0
p r o p o r
t i o n o f m a t c h
i n g r e s p o n s e s
0.90.80.70.60.50.40.30.20.1
0active model passive model ghost
condition
Figure 4. Matching to observed scenarios in full modelversus ghost conditions (adapted from Hopper et al . 2008 ).Mean proportion ( s.e.) of matches to direction of doorslide witnessed by children (grey bars), chimpanzees (whitebars) and pigeons (black bars) in active model, passivemodel with ghost manipulation and full ghost (no model)conditions. Mean values and s.e. for pigeon data courtesy
of Thomas Zentall (2008, personal communication).
Imitation and over-imitation A. Whiten et al. 2423
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
9/13
parts: forehead, foot or bottom. The chimpanzeesmatched the odd body part used in 38 per cent of trials, even after their initial tendency to try a normalbody part like the hand had produced the desiredeffect, such as switching on a light.
There thus seems ample evidence that chimpanzeeshave a capacity for bodily copying and may apply thisin rational imitation. Combining this conclusion withthe results of the ghost studies reviewed above, thereseems less reason now why the default hypothesisabout what mechanism underlies a socially learnedtradition such as panpipe lifting versus panpipepoking should assume emulative rather than imitativelearning. Indeed, the broader context for consideringthese alternatives has changed much over recentyears. It is not long since it was argued that onlyhumans were capable of true imitation, and not chim-panzees and other animals ( Tomasello, 1996 ). In theintervening years, much evidence for imitation hasaccumulated for a diversity of animals includingbirds ( Zentall 2004 ), with particularly detailedcopying being implied in species as diverse as dogs,marmosets and archer sh in the studies describedby Huber et al . (2009) in this issue.
( b ) Imitation/emulation switchingAnother possibility is that chimpanzees will selectivelyopt to imitate or emulate according to one or severalfactors that make the choice adaptive. Horner &Whiten (2005) explored this by presenting to bothchimpanzees and children a task that a human modeldealt with using a mixture of causally relevant and irre-
levant actions, some performed on an opaque version of the task and some on a transparent equivalent, wherethe causal irrelevance of some actions was plain tosee. Horner and Whiten predicted that if chimpanzeesare able to switch adaptively between different formsof social learning, the opaque version would be tackledby more thorough imitative copying of all actions, thetransparent one instead being handled through a moreemulative response, omitting visibly irrelevant actions.The irrelevant actions involved stabbing a stick toolinto a hole in the test box, which in the transparent ver-sion could be seen merely to hit a false ceiling, with nocausal connection to the compartment beneath, from
which the food reward was later extracted. The youngchimpanzees tested did tend to stab the stick in thetop hole, signicantly more in the opaque than thetransparent condition. In this way, they demonstratedcontext-sensitive, exible social learning.
More recently, exibility has been demonstrated inrelation to social context. In Buttelmann et al .s(2007) study of chimpanzees imitation of a modelthat used unusual body parts to press a panel toswitch on a display, the model performed under twoimportantly different conditions: in one, their handswere free, whereas in the second condition, theirhands were unavailable (e.g. wrapped inside a blanket)
so there was good reason to use their forehead or footto press the panel. Human infants just 14 months oldhad earlier demonstrated rational imitation inthis paradigm, copying the bizarre actions of theadult in the hands-free condition, but rarely in the
hands-occupied condition ( Gergely et al . 2002 ). Inan ingenious study in which dogs watched a conspeci-cs either holding a ball in their mouth or not, whileusing their paw to operate a foraging device, Rangeet al . (2007) demonstrated a parallel effect. Likewise,Buttelmann et al . found that the chimpanzees theytested expressed this kind of selectivity, copyingthe unusual actions in 38 per cent of hands-free trials,versus only 19 per cent in the hands-occupied condition.
The Horner and Whiten, and Buttelmann et al .studies illustrate selective and functionally appropriateuse of either imitation or emulation in relation tovariations in both physical and social contexts. Theseresults are consistent with the portfolio theory of Whiten et al . (2004) ; that chimpanzees possess a port-folio of social learning processes (including imitation,emulation and others) that may be applied selectivelyaccording to context.
7. IMITATION AND OVER-IMITATION IN HUMANSHorner & Whiten (2005) discovered that unlikechimpanzees, which, as noted above, copied less of amodels actions when they appeared causally irrelevant,young children, surprisingly, copied these with highdelity. These results have been replicated for a largersample of 3-year-old children by McGuigan et al .(2007) . Moreover, McGuigan et al . extended thestudy to include 5-year olds, suspecting that as childrenmature cognitively, the susceptibility to blanket copyingwould decline. Results were to the contrary: 5-year oldswere even more likely than the 3-year olds to copy allthey saw, even with the transparent box.
Lyons et al . (2007) have replicated these results witha similar task, as well as others that incorporate bothcausally relevant and irrelevant actions, the lattertransparent to view. Lyons et al . checked that thechildrens responses are not merely to please the exper-imenter, both by allowing the child to complete thetask while the experimenter was out of the room (seealso Horner & Whiten 2005 ), and then asking thechild to check the reward object had been put inplace for the next child. A strong tendency to copythe irrelevant modelled actions remained evidenteven in this more rushed, real-world context(gure 5 ). Lyons et al . further demonstrated how sur-
prisingly inexible is young childrens conformity,which the authors dubbed over-imitation. Childrenwere exposed to a training programme in which theexperimenter extracted reward objects from eighttransparent containers, in each case using bothrelevant and irrelevant actions, and asking the childwhich actions the experimenter had to do andwhich were silly and unnecessary (e.g. tapping theside of a container with a feather). Children were effu-sively praised for correctly identifying irrelevantactions. Nevertheless, these children continued toover-imitate when tested later. With Lyons et al .s ver-sion of the Horner and Whiten test box, children
copied the irrelevant actions in over 90 per cent of cases versus under 10 per cent in baseline tests.Lyons etal . (2007) even repeated the experiment with
explicit instructions to avoid irrelevant acts. Before theusual test trials, each child was reminded of the silly
2424 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
10/13
extra thing that had not helped earlier, like the feather-tapping, then explicitly told I want you to watch reallycarefully, because when I open this (puzzle object),I might do something thats silly and extra, just like thefeather, later adding Remember, dont do anythingsilly and extra, okay? Only do the things you have todo. All such efforts failed to diminish over-imitation inthe 3- and 5-year-old children tested.
Whiten et al . (2005 b) described such effects as over-copying rather than over-imitation (the term of Lyonset al . 2007 ) to acknowledge the fact that for those whodene imitation as bodily imitation, there is no evi-dence here that childrens copying is faithful in this
respect. Indeed, Lyons et al . showed that copyingoccurred principally in terms not so much of specicactions, as of the transformations of object move-ments, a closer t to one or more of the classes of emulation distinguished in 2b and gure 1 . Perhaps,then, Lyons et al . documented over-emulation,following some authors terminology. However, sinceother studies have recorded very detailed imitationby human subjects down to the level of the digitused for a task ( Custance et al . 2006 ), over-copyingoffers a more neutral umbrella term pending furtherinvestigation of the underlying mechanisms.
Why do such apparently blinkered child responses
occur, making performance less efcient ( McGuiganet al . 2007 ) than with the more selective chimpanzees(Horner & Whiten 2005 )? Whiten et al . (2005 b)suggested that a plausible explanation . . . is simplythat we are such a thorough-going cultural speciesthat it pays children, as a kind of default strategy, tocopy willy-nilly much of the behavioral repertoirethey see enacted before them. Children have the long-est childhoods of any primate, much of which is spentin play, practice and exploration, so there is plenty of opportunity to weed out wrongly assimilated aspectsof the actions observed (p. 280). Given the massivecultural knowledge and skill reservoir of adult
humans and the relative ignorance of young children,blanket copying may only occasionally lead childreninto superstitionsas in experiments where sillycomponents are inserted on purpose. This will laterbe corrected by childrens direct interaction with
reality, so it is a copy-all, rene/correct-later strategy,an effect recently conrmed in a diffusion study usingthe Horner and Whiten boxes by Flynn (2008) . Lyonset al . (2007) suggest that this strategy enables a childparticularly to learn about causal factors that are sooften opaque in human skills and tasks, although theautomaticity of the system makes it re even whenthe circumstances appear transparently inappropriate,as in our experiments. 1
A copy-all, correct-later strategy may be far fromlimited to childhood. In our ongoing research in thisparadigm, we have recently tested adults and foundthat the trend we earlier documented, of increasingover-copying with age, is extended into adulthood,with adults showing an even higher tendency to actin this way than our 5-year-old sample (unpublisheddata). Although such apparent irrationality may seeminitially surprising, it may again by adaptive in thelong run, because adults too can easily be faced withhighly novel tasks within which there are causal linksinvisible to them, at least initially. Interestingly, severalpapers in the present issue echo this emerging issue of automaticity in human imitation ( Brass et al . 2009 ;Catmur et al . 2009 ; Massen & Prinz 2009 ).
However, the picture of humans as irrational (over-)copiers contrasts with another set of striking ndingsthat have documented what appears to be just the oppo-site. The work of Gergely et al . (2002) , in which even14-month olds avoided copying an adult performingunusual actions when the adult was constrained to actin this way, was described by the authors as rationalimitation. Other studies have shown that childrenscopying can be selective, discounting accidents
(Carpenter et al . 1998 ) and showing sensitivity tosigns of pedagogic engagement ( Csibra & Gergely2006 ) even in infancy. Just where the boundaries fall(and why) between, on the one hand, mindless orblinkered over-copying and, on the other hand, sophis-ticated and rational selectivity, in both child and adult,now emerges as an exciting and challenging territory toresearch more deeply.
8. CUMULATIVE CULTURAL TRANSMISSION:AN EXPERIMENTAL APPROACHStudies reviewed above, and others, have repeatedly
demonstrated high delity imitation by children,which contrasts with the rough and patchy copyingefforts of chimpanzees. Nevertheless, we have shownthat chimpanzees have sufcient copying capacity tosustain traditions incorporating quite complex behav-ioural techniques. The hypothesis that it is the lackof copying ability that prevents chimpanzees from gen-erating the cumulative cultures so evident in thehuman case ( Boyd & Richerson 1996 ; Tomasello1999 ) now sits less comfortably with these results.What, then, is the true explanation for the massivedifference between the two species in cumulativeculture ( Whiten et al . 2003 )?
Earlier we critiqued the loop experiment of Tennieet al . (2009) that purported to tackle this issue. Inour own approach to the topic ( Marshall-Pescini &Whiten 2008 b), juvenile chimpanzees rst learned,through observation of a human model, to slide a
puzzle box cage dome
100
p e r c e n t p r o d u c
i n g
i r r e
l e v a n
t a c
t i o n
80
60
40
20
0
Figure 5. Over-imitation in young children. Percentage of children copying highly visible, causally irrelevant actionsof model in three different tasks, compared with baselineperformance of comparable actions (after Lyons et al .2007 , with permission). See text for discussion. Black bars,observed adult; white bars, baseline. P , 0.001.
Imitation and over-imitation A. Whiten et al. 2425
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
11/13
cover that revealed a hidden hole in an articial fora-ging box and insert a probe to extract honey. Theythen saw the same model using the probe to push inan obscured catch so that, by inserting the probe inthe dipping hole, the hinged lid could be leveredopen to gain all the honey inside, plus several nuts.
Instead of adopting this potential step-up in cumu-lative culture, the chimpanzees stuck to their dipping,gaining only a few licks of honey each time. This wasnot because the task was too difcult, because some
control youngsters that saw no model did exploreand discover the more complex technique. We havenow tested 3- and 4-year-old childrens responses ina similar test ( gure 6 : see electronic supplementarymaterial, methods). Despite the fact that the second,more productive method was made slightly more chal-lenging for the children by the addition of a slidingcover over the crucial catch holding the lid closed,eight of 12 children already using the dipping tech-nique evidenced cumulation by upgrading to thesecond technique and another three children appearedto attempt the beginning of it: only one persisted indipping ( table 1 ).
These results suggest that although chimpanzeescan observationally learn quite complex techniques,these quickly become habitual, restricting whatalternative methods can be switched to, even wherethe benets appear manifest. Children, by contrast,demonstrated cumulative cultural learning. Hrubeschet al . (2008) have since demonstrated a similar resultto that we found in chimpanzees, which they describedas conservatism. Chimpanzees failed to upgrade toan improved technique for gaining food discoveredby a few members of the group; moreover, prac-titioners of this technique later prevented from usingit failed to copy the simpler method of others, which
remained productive. Although the inexibility of chimpanzees social learning in these situations mayappear surprising, there is a remarkable consistencyin these studies. What appears to be revealed is asocial learning propensity that is initially capable of
the sophisticated levels of copying described in earliersections of this paper, but in any one context quicklybecomes canalized or crystallized, producing aroutine resistant to cumulative or other change.
9. CONCLUSIONSOur results and conclusions bear on each of the threeaspects of imitation addressed in this theme issue:evolution, development and control.
On the evolutionary question, we have established sev-eral points of similarity between chimpanzee and humancultural transmission that indicate the characteristicslikely to have existed in our common ancestor of approxi-
mately 6 million years ago. These include capacities(i) to sustain multiple-tradition local cultures, (ii) tocopy others with sufcient delity to transmit culturallyvariant behavioural techniques within and between com-munities, and (iii) to call on both imitative and emulative
(a ) (b) (c) (d )
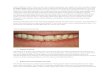
(e) ( f ) (g) (h)
Figure 6. Honey-dip task. ( a d ) model; ( e h) child participant. ( a ) Dip technique: model holds door open with one hand whiledipping tool into the box. ( b) Retrieval of star rewards. ( c) Poke technique: model slides the door with left hand while poking boltwith right. ( d ) Model levers lid open using tool. ( e) Child performing dip technique. ( f ) Child retrieving rewards. ( g ) Child
performing poke technique. ( h) Child levering lid open with tool.
Table 1. Response of each child in baseline andexperimental phases. M, male; F, female (with age inmonths). (1), probed but used hand to open the lid, withno levering; (2), slid panel open but failed to insert sticktool in hole to push the bolt.
child baselinephase 1dip model
phase 2 probe lever model
M36 no dip dip 3 probe open (1)M38 no dip dip 3 probe lever
F39 no dip dip 3 attempt (2)M40 no dip dip 3 dip 3F42 no dip dip 3 attempt (2)F42 no dip dip 3 probe leverF44 no dip dip 3 probe leverM45 dip 3 probe leverF47 no dip dip 3 probe leverF50 no dip dip 3 probe leverF50 no dip dip 3 probe leverF55 no dip dip 3 attempt (2)
2426 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
12/13
forms of learning within a portfolio of different sociallearning processes. Conversely, since the human andchimpanzee lineages diverged, our species has acquiredmore rened capacities for both higher delity imitationand cumulative cultural learning.
Turning to imitative development , an intriguingpicture is emerging in which children exhibit quitesurprising susceptibility to over-copying of others inthe face of apparently obvious perceptual evidence of causal irrelevance. These ndings appear paradoxicalin light of other studies showing sophisticated imitativeselectivity. However, we note that the latter ndingsconcern infants and, again perhaps surprisingly, it isin the older and more cognitively mature children(and even adults) that we nd more blanket copying.These intriguing, apparent paradoxes should drivefurther research to resolve them in coming years.
Finally, there is the question of control of imitation.As the prior paragraph indicates, one new and interest-ing set of issues here concerns ontogenetic changesalessening of control, towards more mindless blanketcopying as children develop. However, in bothhuman infants and apes, other studies are mapping awidening range of conditions in which individualsexpress selectivity in their social learning strategies.These include social factors such as the intentionsof, and constraints on, the model and physical factorssuch as principles of causality. In addition, our prelimi-nary results suggest that chimpanzees may berestricted in their capacity to modify and upgrade therst approaches to a task they learn, in contextswhere children show a readiness to upgrade, facilitat-ing a process of cumulative cultural change.
We conclude by noting that the overview we presentin this paper underlines how this theme issue comes ata time of rich discoveries about the evolution, develop-ment and control of imitation in both human andnon-human species and that, moreover, these threeperspectives are becoming linked in exciting andproductive ways.
The principal research ndings reported here result fromsupport gratefully received from BBSRC, ESRC, BritishAcademy, Leverhulme Trust and Royal Society. We aregrateful to David Buttelmann, Cecilia Heyes, LudwigHuber and Derek Lyons for comments on an earlierversion of the manuscript.
ENDNOTES1 Additional comments appraising the studies of Lyons et al . (2007)and Tennie et al . (2009) are in the electronic supplementary material.2 A video clip of the behaviour of the two chimpanzees accompaniesMarshall-Pescini & Whiten (2008 b) in electronic supplementarymaterial at http://dx.doi.org/10.1037/0735-7036.122.2.186.supp .The observer chimpanzee had already attempted cracking using ahammer stone, but not successfully.
REFERENCESBonnie, K. E., Horner, V., Whiten, A. & de Waal, F. B. M.
2007 Spread of arbitrary customs among chimpanzees:a controlled experiment. Proc. R. Soc. B. 274 , 367372.(doi:10.1098/rspb.2006.3733 )
Boyd, R. & Richerson, P. J. 1996 Why culture is common, butcultural evolution is rare. In Evolution of social behaviour patterns in primates and man (eds W. G. Runciman,
J. Maynard Smith & R. I. M. Dunbar), pp. 7793.Oxford, UK: Oxford University Press.
Brass, M., Ruby, P. & Spengler, S. 2009 Inhibition of imitativebehaviour and social cognition. Phil. Trans. R. Soc. B . 364 ,23592367. ( doi:10.1098/rstb.2009.0066 )
Buttelmann, D., Carpenter, M., Call, J. & Tomasello, M.2007 Enculturated chimpanzees imitate rationally. Dev.Sci. 10 , 3138.
Byrne, R. W. 2002 Emulation in apes: verdict not proven.Dev. Sci. 5, 2022. ( doi:10.1111/1467-7687.00198 )Call, J. 2001 Body imitation in an enculturated orangutan
(Pongo pygmaeus ). Cybern. Syst. 32 , 97119. ( doi:10.1080/019697201300001821 )
Call, J., Carpenter, M. & Tomasello, M. 2005 Copyingresults and copying actions in the process of social learn-ing: chimpanzees ( Pan troglodytes ) and human children(Homo sapiens ). Anim. Cogn. 8, 151163. ( doi:10.1007/s10071-004-0237-8 )
Carpenter, M., Akhtar, N. & Tomasello, M. 1998 Fourteen-through 18-month-old infants differentially imitateintentional and accidental actions. Inf. Behav. Dev. 21 ,315330. ( doi:10.1016/S0163-6383(98)90009-1 )
Catmur, C., Walsh, V. & Heyes, C. 2009 Associativesequence learning: the role of experience in the develop-ment of imitation and the mirror system. Phil. Trans. R.Soc. B . 364 , 23692380. ( doi:10.1098/rstb.2009.0048 )
Csibra, G. & Gergely, G. 2006 Social learning and socialcognition: The case for pedagogy. In Processes of changein brain and cognitive development. Attention and perform-ance XXI (eds Y. Munakata & M. H. Johnson),pp. 249274. Oxford, UK: Oxford University Press.
Custance, D. M., Whiten, A. & Bard, K. A. 1995 Can youngchimpanzees ( Pan troglodytes ) imitate arbitrary actions?Hayes and Hayes 1952 revisited. Behaviour 132 ,837859. ( doi:10.1163/156853995X00036 )
Custance, D. M., Whiten, A. & Fredman, T. 1999 Sociallearning of articial fruit processing in capuchin monkeys(Cebus apella ). J. Comp. Psychol. 113 , 1323. ( doi:10.1037/0735-7036.113.1.13 )
Custance, D., Prato-Previde, E., Spiezio, C. & Rigamonti,M. M. 2006 Social learning in pig-tailed macaques( Macaca nemestrina ) and adult humans ( Homo sapiens )on a two-action articial fruit. J. Comp. Psychol. 120 ,303313. ( doi:10.1037/0735-7036.120.3.303 )
Flynn, E. 2008 Investigating children as cultural magnets: dochildren transmit redundant information along diffusionchains? Phil. Trans. R. Soc. B 363 , 35413551. ( doi:10.1098/rstb.2008.0136 )
Flynn, E. & Whiten, A. 2008 Cultural transmission of tool-usein young children: a diffusion chain study. Soc. Dev. 17 ,699718. ( doi:10.1111/j.1467-9507.2007.00453.x )
Galef Jr, B. G. 1988 Imitation in animals: history, de-nitions, and interpretation of data from the psychologicallaboratory. In Social learning: psychological and biological perspectives (eds T. R. Zentall & B. G. Galef), pp. 328.Hillsdale, NJ: Erlbaum.
Galef, B. G. 1992 The question of animal culture. Hum. Nat. 3, 157178. ( doi:10.1007/BF02692251 )
Gergely, G., Bekkering, H. & Kiraly, I. 2002 Rational imita-tion in preverbal infants. Nature 415 , 755.
Heyes, C. M. 1993 Imitation, culture and cognition. Anim.Behav. 46 , 9991010. ( doi:10.1006/anbe.1993.1281 )
Heyes, C. M. 1994 Social learning in animals: categoriesand mechanisms. Biol. Rev. 69 , 207231. ( doi:10.1111/j.1469-185X.1994.tb01506.x )
Heyes, C. M. 1996 Genuine imitation? In Social learning inanimals: the roots of culture (eds C. M. Heyes & B. G.Galef), pp. 371389. London, UK: Academic Press.
Hopper, L. M., Spiteri, A., Lambeth, S. P., Schapiro, S. J.,Horner, V. & Whiten, A. 2007 Experimental studies of
Imitation and over-imitation A. Whiten et al. 2427
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://dx.doi.org/10.1037/0735-7036.122.2.186.supphttp://dx.doi.org/doi:10.1098/rspb.2006.3733http://dx.doi.org/doi:10.1098/rstb.2009.0066http://dx.doi.org/doi:10.1111/1467-7687.00198http://dx.doi.org/doi:10.1080/019697201300001821http://dx.doi.org/doi:10.1080/019697201300001821http://dx.doi.org/doi:10.1007/s10071-004-0237-8http://dx.doi.org/doi:10.1007/s10071-004-0237-8http://dx.doi.org/doi:10.1016/S0163-6383(98)90009-1http://dx.doi.org/doi:10.1098/rstb.2009.0048http://dx.doi.org/doi:10.1163/156853995X00036http://dx.doi.org/doi:10.1037/0735-7036.113.1.13http://dx.doi.org/doi:10.1037/0735-7036.113.1.13http://dx.doi.org/doi:10.1037/0735-7036.120.3.303http://dx.doi.org/doi:10.1098/rstb.2008.0136http://dx.doi.org/doi:10.1098/rstb.2008.0136http://dx.doi.org/doi:10.1111/j.1467-9507.2007.00453.xhttp://dx.doi.org/doi:10.1007/BF02692251http://dx.doi.org/doi:10.1006/anbe.1993.1281http://dx.doi.org/doi:10.1111/j.1469-185X.1994.tb01506.xhttp://dx.doi.org/doi:10.1111/j.1469-185X.1994.tb01506.xhttp://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://dx.doi.org/doi:10.1111/j.1469-185X.1994.tb01506.xhttp://dx.doi.org/doi:10.1111/j.1469-185X.1994.tb01506.xhttp://dx.doi.org/doi:10.1006/anbe.1993.1281http://dx.doi.org/doi:10.1007/BF02692251http://dx.doi.org/doi:10.1111/j.1467-9507.2007.00453.xhttp://dx.doi.org/doi:10.1098/rstb.2008.0136http://dx.doi.org/doi:10.1098/rstb.2008.0136http://dx.doi.org/doi:10.1037/0735-7036.120.3.303http://dx.doi.org/doi:10.1037/0735-7036.113.1.13http://dx.doi.org/doi:10.1037/0735-7036.113.1.13http://dx.doi.org/doi:10.1163/156853995X00036http://dx.doi.org/doi:10.1098/rstb.2009.0048http://dx.doi.org/doi:10.1016/S0163-6383(98)90009-1http://dx.doi.org/doi:10.1007/s10071-004-0237-8http://dx.doi.org/doi:10.1007/s10071-004-0237-8http://dx.doi.org/doi:10.1080/019697201300001821http://dx.doi.org/doi:10.1080/019697201300001821http://dx.doi.org/doi:10.1111/1467-7687.00198http://dx.doi.org/doi:10.1098/rstb.2009.0066http://dx.doi.org/doi:10.1098/rspb.2006.3733http://dx.doi.org/10.1037/0735-7036.122.2.186.supphttp://dx.doi.org/10.1037/0735-7036.122.2.186.supphttp://rstb.royalsocietypublishing.org/8/9/2019 Whiten Et Al (2009)
13/13
traditions and underlying transmission processes inchimpanzees. Anim. Behav. 73 , 10211032. ( doi:10.1016/j.anbehav.2006.07.016 )
Hopper, L. M., Lambeth, S. P., Schapiro, S. J. & Whiten, A.2008 Observational learning in chimpanzees and childrenstudied through ghost conditions. Proc. R. Soc. B 275 ,835840. ( doi:10.1098/rspb.2007.1542 )
Horner, V. K. & Whiten, A. 2005 Causal knowledge and
imitation/emulation switching in chimpanzees ( Pantroglodytes ) and children. Anim. Cogn. 8, 164181.(doi:10.1007/s10071-004-0239-6 )
Horner, V., Whiten, A., Flynn, E. & de Waal, F. B. M. 2006Faithful replication of foraging techniques along culturaltransmission chains by chimpanzees and children. Proc. Natl Acad. Sci. USA 103 , 13 87813 883. ( doi:10.1073/pnas.0606015103 )
Hrubesch, C., Preuschoft, S. & van Schaik, C. 2008 Skillmastery inhibits adoption of observed alternative sol-utions among chimpanzees ( Pan troglodytes ). Anim.Cogn. 12 , 209216. ( doi:10.1007/s10071-008-0183-y )
Huber, L., Range, F., Voelkl, B., Szucsich, A., Viranyi, Z. &Miklosi, A. 2009 The evolution of imitation: what do thecapacities of non-human animals tell us about themechanisms of imitation? Phil. Trans. R. Soc. B . 364 ,2299 2309. ( doi:10.1098/rstb.2009.0060 )
Hurley, S. & Chater, N. (eds) 2005 Perspectives on imitation: from neurosciences to social science . Boston, MA: MIT Press.
Klein, E. D. & Zentall, T. R. 2003 Imitation and affordancelearning by pigeons ( Columbia livia ). J. Comp. Psychol.117 , 414419. ( doi:10.1037/0735-7036.117.4.414 )
Lyons, D. E., Young, A. G. & Keil, F. C. 2007 The hiddenstructure of overimitation. Proc. Natl Acad. Sci. USA 104 ,1975119 756. ( doi:10.1073/pnas.0704452104 )
Massen, C. & Prinz, W. 2009 Movements, actions and tool-useactions: an ideomotor approach to imitation. Phil. Trans. R.Soc. B . 364 , 23492358. ( doi:10.1098/rstb.2009.0059 )
Marshall-Pescini,S. & Whiten, A. 2008 a Social learningof nut-cracking behaviour in East African sanctuary-living chim-panzees ( Pan troglodytes schweinfurthii ). J. Comp. Psychol.122 , 186194. ( doi:10.1037/0735-7036.122.2.186 )
Marshall-Pescini, S. & Whiten, A. 2008 b Chimpanzees ( Pantroglodytes ) and the question of cumulative culture: anexperimental approach. Anim. Cogn. 11 , 449456.(doi:10.1007/s10071-007-0135-y )
McGuigan, N. & Graham, M. In press. Cultural trans-mission of irrelevant tool actions in diffusion chains of 3- and 5-year-old children. Eur. J. Dev. Psychol. (doi:10.1080/17405620902858125 )
McGuigan, N., Whiten, A., Flynn, E. & Horner, V. 2007Imitation of causally-opaque versus causally-transparenttool use by 3- and 5-year-old children. Cogn. Dev. 22 ,353364. ( doi:10.1016/j.cogdev.2007.01.001 )
Nagell, K., Olguin, R. S. & Tomasello, M. 1993 Processes of sociallearninginthetooluseofchimpanzees( Pantroglodytes )and human children ( Homo sapiens ). J. Comp. Psychol. 107 ,174 186. ( doi:10.1037/0735-7036.107.2.174 )
Range, F., Viranyi, Z. & Huber, L. 2007 Selective imitationin domestic dogs. Curr. Biol. 17 , 15. ( http://dx.doi.org/10.1016/j.cub.2007.04.026 )
Tennie, C., Call, J. & Tomasello, M. 2009 Ratcheting up theratchet: on the evolution of cumulative culture. Phil Trans.R. Soc. B . 364 , 24052415. ( doi:10.1098/rstb.2009.0052 )
Thorndike, E. L. 1898 Animal intelligence: an experimentalstudy of the associative process in animals. Psychol. Rev. Monogr. 2 , 551553.
Thorpe, W. H. 1963 Learning and instinct in animals.London, UK: Methuen.
Tomasello, M. 1990 Cultural transmission in the tool useand communicatory signaling of chimpanzees? InLanguage and intelligence in monkeys and apes:
Comparative developmental perspectives (eds S. T. Parker& K. Gibson), pp. 274311. Cambridge, UK: CambridgeUniversity Press.
Tomasello, M. 1996 Do apes ape? In Social learning inanimals: the roots of culture (eds C. M. Heyes & B. G.Galef), pp. 319346. London, UK: Academic Press.
Tomasello, M. 1998 Emulation learning and cultural learn-ing. Behav. Brain Sci. 21 , 703704. ( doi:10.1017/
S0140525X98441748 )Tomasello, M. 1999 The cultural origins of human cognition.Cambridge, MA: Harvard University Press.
Tomasello, M. & Call, J. 1997 Primate cognition . Oxford,UK: Oxford University Press.
Tomasello, M., Davis-Dasilva, M., Camak, L. & Bard, K. 1987Observational learning of tool-use by young chimpanzees.Hum. Evol. 2, 175183. ( doi:10.1007/BF02436405 )
Voelkl, B. & Huber, L. 2000 True imitation in marmo-sets. Anim. Behav. 60 , 195202. ( doi:10.1006/anbe.2000.1457 )
Voelkl, B. & Huber, L. 2007 Imitation as faithful copying of a novel technique in marmoset monkeys. PLoS ONE 2 ,e611. ( doi:10.1371/journal.pone.0000611 )
Whiten, A. 2005 The second inheritance system of chimpan-zees and humans. Nature 437 , 5255. ( doi:10.1038/nature04023 )
Whiten, A. & Ham, R. 1992 On the nature and evolution of imitation in the animal kingdom: reappraisal of a centuryof research. Adv. Study Behav. 11 , 239283. ( doi:10.1016/S0065-3454(08)60146-1 )
Whiten, A. & Mesoudi, A. 2008 An experimental scienceof culture: animal social diffusion experiments. Phil. Trans.R. Soc. B 363 , 34773488. ( doi:10.1098/rstb.2008.0134 )
Whiten, A., Custance, D. M., Gomez, J.-C., Teixidor, P. &Bard, K. A. 1996 Imitative learning of articial fruitprocessing in children ( Homo sapiens ) and chimpanzees(Pan troglodytes ). J. Comp. Psychol. 110 , 314. ( doi:10.1037/0735-7036.110.1.3 )
Whiten, A., Goodall, J., McGrew, W. C., Nishida, T.,Reynolds, V., Sugiyama, Y., Tutin, C. E. G., Wrangham,R. W. & Boesch, C. 1999 Cultures in chimpanzees. Nature 399 , 682685. ( doi:10.1038/21415 )
Whiten, A., Horner, V. & Marshall-Pescini, S. R. J. 2003Cultural panthropology. Evol. Anthropol. 12 , 92105.(doi:10.1002/evan.10107 )
Whiten, A., Horner, V., Litcheld, C. A. & Marshall-Pescini,S. 2004 How do apes ape? Learn. Behav. 32 , 3652.
Whiten, A., Horner, V. & de Waal, F. B. M. 2005 a Confor-mity to cultural norms of tool use in chimpanzees. Nature437 , 737740. ( doi:10.1038/nature04047 )
Whiten, A., Horner, V. & Marshall-Pescini, S. 2005 b Selec-tive imitation in child and chimpanzee: a window on theconstrual of others actions. In Perspectives on imitation: From neuroscience to social science (eds S. Hurley &N. Chater), pp. 263283. Cambridge, MA: MIT Press.
Whiten, A., Spiteri, A., Horner, V., Bonnie, K. E., Lambeth,S. P., Schapiro, S. J. & de Waal, F. B. M. 2007 Trans-mission of multiple traditions within and betweenchimpanzee groups. Curr. Biol. 17 , 10381043. ( doi:10.1016/j.cub.2007.05.031 )
Witmer, L. 1909 A monkey with a mind. Psychol. Clin. 3 ,179205.
Wood, D. 1989 Social interaction as tutoring. In Interactionin human development (eds M. H. Bornstein & J. S.Bruner), pp. 59 80. Hillsdale, NJ: Lawrence ErlbaumAssociates, Publishers.
Zentall, T. 2004 Action imitation in birds. Learn. Behav. 32 ,1523.
Zentall, T. R., Sutton, J. E. & Sherburne, L. M. 1996 Trueimitative learning in pigeons. Psychol. Sci. 7, 343346.(doi:10.1111/j.1467-9280.1996.tb00386.x )
2428 A. Whiten et al. Imitation and over-imitation
Phil. Trans. R. Soc. B (2009)
on 3 August 2009rstb.royalsocietypublishing.orgDownloaded from
http://dx.doi.org/doi:10.1016/j.anbehav.2006.07.016http://dx.doi.org/doi:10.1016/j.anbehav.2006.07.016http://dx.doi.org/doi:10.1098/rspb.2007.1542http://dx.doi.org/doi:10.1007/s10071-004-0239-6http://dx.doi.org/doi:10.1073/pnas.0606015103http://dx.doi.org/doi:10.1073/pnas.0606015103http://dx.doi.org/doi:10.1007/s10071-008-0183-yhttp://dx.doi.org/doi:10.1098/rstb.2009.0060http://dx.doi.org/doi:10.1037/0735-7036.117.4.414http://dx.doi.org/doi:10.1073/pnas.0704452104http://dx.doi.org/doi:10.1098/rstb.2009.0059http://dx.doi.org/doi:10.1037/0735-7036.122.2.186http://dx.doi.org/doi:10.1007/s10071-007-0135-yhttp://dx.doi.org/doi:10.1080/17405620902858125http://dx.doi.org/doi:10.1080/17405620902858125http://dx.doi.org/doi:10.1016/j.cogdev.2007.01.001http://dx.doi.org/doi:10.1037/0735-7036.107.2.174http://dx.doi.org/http://dx.doi.org/10.1016/j.cub.2007.04.026http://dx.doi.org/http://dx.doi.org/10.1016/j.cub.2007.04.026http://dx.doi.org/doi:10.1098/rstb.2009.0052http://dx.doi.org/doi:10.1017/S0140525X98441748http://dx.doi.org/doi:10.1017/S0140525X98441748http://dx.doi.org/doi:10.1007/BF02436405http://dx.doi.org/doi:10.1006/anbe.2000.1457http://dx.doi.org/doi:10.1006/anbe.2000.1457http://dx.doi.org/doi:10.1371/journal.pone.0000611http://dx.doi.org/doi:10.1038/nature04023http://dx.doi.org/doi:10.1038/nature04023http://dx.doi.org/doi:10.1016/S0065-3454(08)60146-1http://dx.doi.org/doi:10.1016/S0065-3454(08)60146-1http://dx.doi.org/doi:10.1098/rstb.2008.0134http://dx.doi.org/doi:10.1037/0735-7036.110.1.3http://dx.doi.org/doi:10.1037/0735-7036.110.1.3http://dx.doi.org/doi:10.1038/21415http://dx.doi.org/doi:10.1002/evan.10107http://dx.doi.org/doi:10.1038/nature04047http://dx.doi.org/doi:10.1016/j.cub.2007.05.031http://dx.doi.org/doi:10.1016/j.cub.2007.05.031http://dx.doi.org/doi:10.1111/j.1467-9280.1996.tb00386.xhttp://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://rstb.royalsocietypublishing.org/http://dx.doi.org/doi:10.1111/j.1467-9280.1996.tb00386.xhttp://dx.doi.org/doi:10.1016/j.cub.2007.05.031http://dx.doi.org/doi:10.1016/j.cub.2007.05.031http://dx.doi.org/doi:10.1038/nature04047http://dx.doi.org/doi:10.1002/evan.10107http://dx.doi.org/doi:10.1038/21415http://dx.doi.org/doi:10.1037/0735-7036.110.1.3http://dx.doi.org/doi:10.1037/0735-7036.110.1.3http://dx.doi.org/doi:10.1098/rstb.2008.0134http://dx.doi.org/doi:10.1016/S0065-3454(08)60146-1http://dx.doi.org/doi:10.1016/S0065-3454(08)60146-1http://dx.doi.org/doi:10.1038/nature04023http://dx.doi.org/doi:10.1038/nature04023http://dx.doi.org/doi:10.1371/journal.pone.0000611http://dx.doi.org/doi:10.1006/anbe.2000.1457http://dx.doi.org/doi:10.1006/anbe.2000.1457http://dx.doi.org/doi:10.1007/BF02436405http://dx.doi.org/doi:10.1017/S0140525X98441748http://dx.doi.org/doi:10.1017/S0140525X98441748http://dx.doi.org/doi:10.1098/rstb.2009.0052http://dx.doi.org/http://dx.doi.org/10.1016/j.cub.2007.04.026http://dx.doi.org/http://dx.doi.org/10.1016/j.cub.2007.04.026http://dx.doi.org/http://dx.doi.org/10.1016/j.cub.2007.04.026http://dx.doi.org/doi:10.1037/0735-7036.107.2.174http://dx.doi.org/doi:10.1016/j.cogdev.2007.01.001http://dx.doi.org/doi:10.1080/17405620902858125http://dx.doi.org/doi:10.1080/17405620902858125http://dx.doi.org/doi:10.1007/s10071-007-0135-yhttp://dx.doi.org/doi:10.1037/0735-7036.122.2.186http://dx.doi.org/doi:10.1098/rstb.2009.0059http://dx.doi.org/doi:10.1073/pnas.0704452104http://dx.doi.org/doi:10.1037/0735-7036.117.4.414http://dx.doi.org/doi:10.1098/rstb.2009.0060http://dx.doi.org/doi:10.1007/s10071-008-0183-yhttp://dx.doi.org/doi:10.1073/pnas.0606015103http://dx.doi.org/doi:10.1073/pnas.0606015103http://dx.doi.org/doi:10.1007/s10071-004-0239-6http://dx.doi.org/doi:10.1098/rspb.2007.1542http://dx.doi.org/doi:10.1016/j.anbehav.2006.07.016http://dx.doi.org/doi:10.1016/j.anbehav.2006.07.016http://rstb.royalsocietypublishing.org/