Embed Size (px)
Citation preview

Yes-associated protein (YAP) transcriptionalcoactivator functions in balancing growthand differentiation in skinHaiying Zhang, H. Amalia Pasolli, and Elaine Fuchs1
Laboratory of Mammalian Cell Biology and Development, Howard Hughes Medical Institute, The Rockefeller University, New York, NY 10065
Contributed by Elaine Fuchs, December 28, 2010 (sent for review November 18, 2010)
In mammals, skin begins as a single-layered epithelium, which,through a series of signals, either stratifies and differentiates tobecome epidermis or invaginates downward to make hair follicles(HFs). To achieve and maintain proper tissue architecture, keratino-cytes must intricately balance growth and differentiation. Here, weuncover a critical andhitherto unappreciated role for Yes-associatedprotein (YAP), an evolutionarily conserved transcriptional coactiva-tor with potent oncogenic potential. We show that YAP is highlyexpressed and nuclear in single-layered basal epidermal progeni-tors. Notably, nuclear YAP progressively declines with age andcorrelates with proliferative potential of epidermal progenitors.Shortly after initiation of HF morphogenesis, YAP translocates tothe cytoplasm of differentiating cells. Through genetic analysis, wedemonstrate a role for YAP in maintaining basal epidermal progen-itors and regulating HF morphogenesis. YAP overexpression causeshair placodes to evaginate into epidermis rather than invaginateinto dermis. YAP also expands basal epidermal progenitors, pro-motes proliferation, and inhibits terminal differentiation. In vitrogain-and-loss of function studies show that primary mouse kerati-nocytes (MKs) accelerate proliferation, suppress differentiation, andinhibit apoptosis when YAP is activated and reverse these featureswhen YAP is inhibited. Finally, we identify Cyr61 as a target of YAPin MKs and demonstrate a requirement for TEA domain (TEAD)transcriptional factors to comediate YAP functions in MKs.
Hippo signaling | gene expression | proliferation and cancer | skin cancer
During tissue and organ development, progenitors must bal-ance proliferation and differentiation in a temporally and
spatially controlled fashion. Understanding the molecular mech-anisms involved has attracted intense research interest, becausetoo little proliferation contributes to aging and too much predis-poses tissues to proliferative disorders, including cancers. Accu-mulating evidence has implicated an oncogene, Yes-associatedprotein (YAP), in this process. YAP is a transcriptional coac-tivator that lacks a DNA-binding motif but has a potent trans-activation domain at its C terminus (1). YAP’s N terminus has anassociation domain for the TEA domain (TEAD) family of DNA-binding proteins, which have been linked to YAP’s growth-pro-moting function (2). Depending on the splicing variant, one or twoprotein–protein interacting (WW) domains yield the potential foradditional interactions between YAP and other proteins. Theseinclude other possible transcriptional partners for YAP (ErbB,Runx2, and chromatin modeling proteins) as well as the negativeYAP regulator large tumor suppressor (LATS) kinase (3–6).YAP’s importance was discovered in Drosophila, where its ho-
molog, Yorkie, was shown to promote tissue growth by promotingcell proliferation and inhibiting apoptosis (6). Genetically, Yorkieis the ultimate effector of the evolutionarily conserved Hippo tu-mor suppressor pathway (6).UponHippo signaling inmice, YAP isphosphorylated at a key serine (S127), confining it to the cyto-plasm, where it can no longer function in target gene expression (7).Several lines of evidence suggest that YAP participates in
regulating stem cell (SC) behavior. In cultured ES cells, YAP ishighly expressed (8) and required for self-renewal and suppres-
sion of differentiation (9). In mouse intestine, YAP’s overex-pression causes an expanded pool of undifferentiated progenitorsin the crypt (10). In chick, YAP functions through TEAD to re-gulate neural progenitor cell number (11), and in Drosophila,Yorkie maintains neuroepithelial cells in an undifferentiatedstate (12). The mechanisms underlying YAP regulation in pro-genitors remain limited, however, particularly within the physio-logical context of tissues.In this study, we investigate YAP’s function during mouse
epidermal development. Skin begins as a single layer of multi-potent progenitors. In response to extrinsic cues, embryonic epi-dermal cells either stratify to produce the protective barrier at theskin surface or invaginate to form hair follicles (HFs) (13). Anunderlying basement membrane (BM) rich in extracellular matrixand growth factors maintains the epithelial progenitors that ad-here to it and also separates the epidermis and its appendagesfrom surrounding dermis. We show that YAP is highly expressedin the most proliferative of these progenitors, where it functionsin controlling their self-renewal and maintaining their un-differentiated state. We further implicate TEAD as a cofactor forYAP and uncover some of their possible targets in regulatingthe process.
ResultsNuclear YAP Marks the Progenitors in Developing Epidermis andCorrelates with Proliferation. Microarray analyses detected Yaptranscripts in embryonic epidermis (14). Using a pan-antibody(Ab) that recognizes all forms of YAP, we confirmedYAP proteinexpression as early as embryo day 12 (E12), when epidermis existsas a single layer. At this time, immunoreactivity was predom-inantly nuclear, suggesting that the majority of YAP is in its activeform (Fig. 1A).On stratification, YAP distributed differentially, concentrating
in the basal layer, where the majority of YAP was still nuclear(Fig. 1B). An Ab specific for the LATS phosphorylation site(Ser127) of YAP also showed some cytoplasmic staining at thistime, however, suggestive that the Hippo signaling pathway wasactive (Fig. S1A).As HFs initiated, YAP immunolabeling diminished but was
still nuclear in the early placode (Fig. S1B). During maturation,nuclear YAP was detected in the outer root sheath (ORS) andtransit-amplifying matrix (Mx) cells at the HF base (Fig. 1C).Notably, both of these populations are proliferative and adher-ent to the surrounding BM. Conversely, as shown by both pan-YAP and phospho-specific YAP Abs, YAP shifted mainly to the
Author contributions: H.Z., H.A.P., and E.F. designed research; H.Z. and H.A.P. performedresearch; H.Z., H.A.P., and E.F. analyzed data; and H.Z. and E.F. wrote the paper.
The authors declare no conflict of interest.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1019603108/-/DCSupplemental.
2270–2275 | PNAS | February 8, 2011 | vol. 108 | no. 6 www.pnas.org/cgi/doi/10.1073/pnas.1019603108
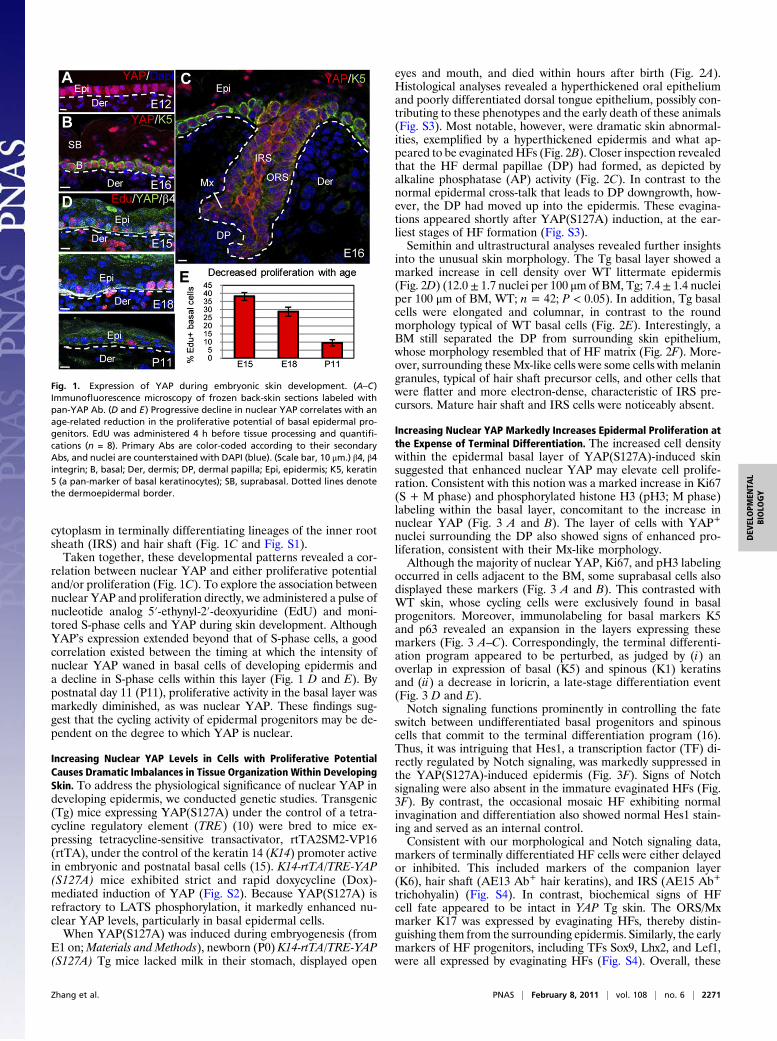
cytoplasm in terminally differentiating lineages of the inner rootsheath (IRS) and hair shaft (Fig. 1C and Fig. S1).Taken together, these developmental patterns revealed a cor-
relation between nuclear YAP and either proliferative potentialand/or proliferation (Fig. 1C). To explore the association betweennuclear YAP and proliferation directly, we administered a pulse ofnucleotide analog 5′-ethynyl-2′-deoxyuridine (EdU) and moni-tored S-phase cells and YAP during skin development. AlthoughYAP’s expression extended beyond that of S-phase cells, a goodcorrelation existed between the timing at which the intensity ofnuclear YAP waned in basal cells of developing epidermis anda decline in S-phase cells within this layer (Fig. 1 D and E). Bypostnatal day 11 (P11), proliferative activity in the basal layer wasmarkedly diminished, as was nuclear YAP. These findings sug-gest that the cycling activity of epidermal progenitors may be de-pendent on the degree to which YAP is nuclear.
Increasing Nuclear YAP Levels in Cells with Proliferative PotentialCauses Dramatic Imbalances in Tissue Organization Within DevelopingSkin. To address the physiological significance of nuclear YAP indeveloping epidermis, we conducted genetic studies. Transgenic(Tg) mice expressing YAP(S127A) under the control of a tetra-cycline regulatory element (TRE) (10) were bred to mice ex-pressing tetracycline-sensitive transactivator, rtTA2SM2-VP16(rtTA), under the control of the keratin 14 (K14) promoter activein embryonic and postnatal basal cells (15). K14-rtTA/TRE-YAP(S127A) mice exhibited strict and rapid doxycycline (Dox)-mediated induction of YAP (Fig. S2). Because YAP(S127A) isrefractory to LATS phosphorylation, it markedly enhanced nu-clear YAP levels, particularly in basal epidermal cells.When YAP(S127A) was induced during embryogenesis (from
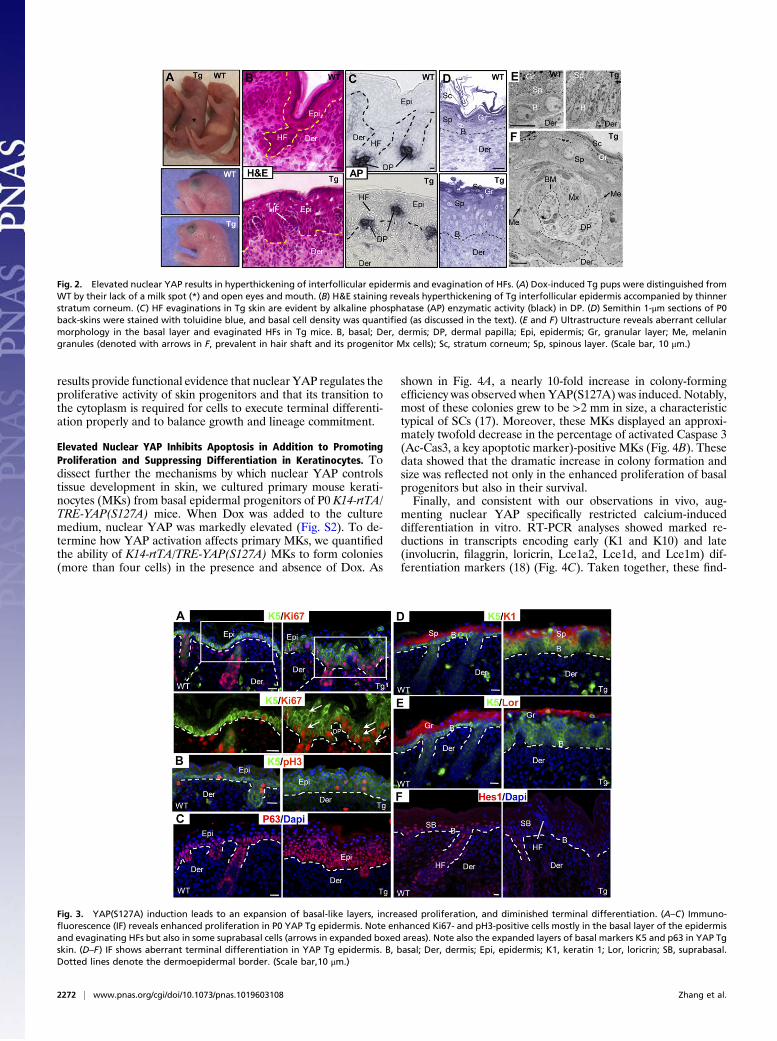
E1 on;Materials and Methods), newborn (P0) K14-rtTA/TRE-YAP(S127A) Tg mice lacked milk in their stomach, displayed open
eyes and mouth, and died within hours after birth (Fig. 2A).Histological analyses revealed a hyperthickened oral epitheliumand poorly differentiated dorsal tongue epithelium, possibly con-tributing to these phenotypes and the early death of these animals(Fig. S3). Most notable, however, were dramatic skin abnormal-ities, exemplified by a hyperthickened epidermis and what ap-peared to be evaginatedHFs (Fig. 2B). Closer inspection revealedthat the HF dermal papillae (DP) had formed, as depicted byalkaline phosphatase (AP) activity (Fig. 2C). In contrast to thenormal epidermal cross-talk that leads to DP downgrowth, how-ever, the DP had moved up into the epidermis. These evagina-tions appeared shortly after YAP(S127A) induction, at the ear-liest stages of HF formation (Fig. S3).Semithin and ultrastructural analyses revealed further insights
into the unusual skin morphology. The Tg basal layer showed amarked increase in cell density over WT littermate epidermis(Fig. 2D) (12.0± 1.7 nuclei per 100 μm of BM, Tg; 7.4 ± 1.4 nucleiper 100 μm of BM, WT; n = 42; P < 0.05). In addition, Tg basalcells were elongated and columnar, in contrast to the roundmorphology typical of WT basal cells (Fig. 2E). Interestingly, aBM still separated the DP from surrounding skin epithelium,whose morphology resembled that of HF matrix (Fig. 2F). More-over, surrounding theseMx-like cells were some cells with melaningranules, typical of hair shaft precursor cells, and other cells thatwere flatter and more electron-dense, characteristic of IRS pre-cursors. Mature hair shaft and IRS cells were noticeably absent.
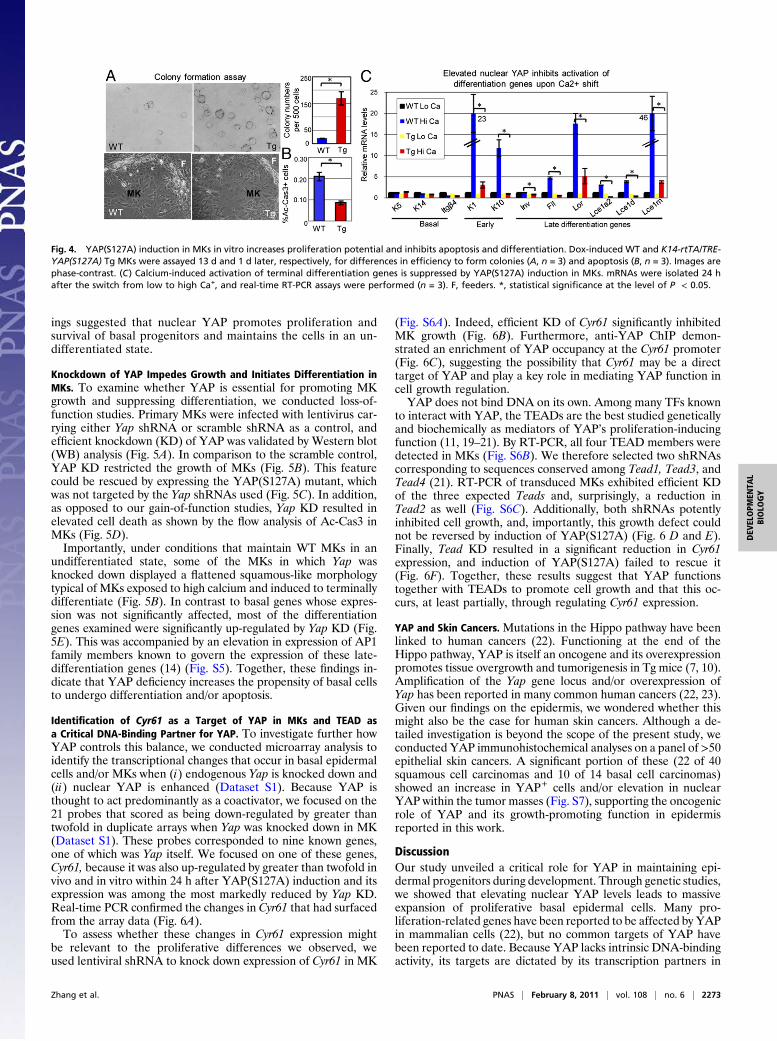
Increasing Nuclear YAP Markedly Increases Epidermal Proliferation atthe Expense of Terminal Differentiation. The increased cell densitywithin the epidermal basal layer of YAP(S127A)-induced skinsuggested that enhanced nuclear YAP may elevate cell prolife-ration. Consistent with this notion was a marked increase in Ki67(S + M phase) and phosphorylated histone H3 (pH3; M phase)labeling within the basal layer, concomitant to the increase innuclear YAP (Fig. 3 A and B). The layer of cells with YAP+
nuclei surrounding the DP also showed signs of enhanced pro-liferation, consistent with their Mx-like morphology.Although the majority of nuclear YAP, Ki67, and pH3 labeling
occurred in cells adjacent to the BM, some suprabasal cells alsodisplayed these markers (Fig. 3 A and B). This contrasted withWT skin, whose cycling cells were exclusively found in basalprogenitors. Moreover, immunolabeling for basal markers K5and p63 revealed an expansion in the layers expressing thesemarkers (Fig. 3 A–C). Correspondingly, the terminal differenti-ation program appeared to be perturbed, as judged by (i) anoverlap in expression of basal (K5) and spinous (K1) keratinsand (ii) a decrease in loricrin, a late-stage differentiation event(Fig. 3 D and E).Notch signaling functions prominently in controlling the fate
switch between undifferentiated basal progenitors and spinouscells that commit to the terminal differentiation program (16).Thus, it was intriguing that Hes1, a transcription factor (TF) di-rectly regulated by Notch signaling, was markedly suppressed inthe YAP(S127A)-induced epidermis (Fig. 3F). Signs of Notchsignaling were also absent in the immature evaginated HFs (Fig.3F). By contrast, the occasional mosaic HF exhibiting normalinvagination and differentiation also showed normal Hes1 stain-ing and served as an internal control.Consistent with our morphological and Notch signaling data,
markers of terminally differentiated HF cells were either delayedor inhibited. This included markers of the companion layer(K6), hair shaft (AE13 Ab+ hair keratins), and IRS (AE15 Ab+
trichohyalin) (Fig. S4). In contrast, biochemical signs of HFcell fate appeared to be intact in YAP Tg skin. The ORS/Mxmarker K17 was expressed by evaginating HFs, thereby distin-guishing them from the surrounding epidermis. Similarly, the earlymarkers of HF progenitors, including TFs Sox9, Lhx2, and Lef1,were all expressed by evaginating HFs (Fig. S4). Overall, these
Fig. 1. Expression of YAP during embryonic skin development. (A–C)Immunofluorescence microscopy of frozen back-skin sections labeled withpan-YAP Ab. (D and E) Progressive decline in nuclear YAP correlates with anage-related reduction in the proliferative potential of basal epidermal pro-genitors. EdU was administered 4 h before tissue processing and quantifi-cations (n = 8). Primary Abs are color-coded according to their secondaryAbs, and nuclei are counterstained with DAPI (blue). (Scale bar, 10 μm.) β4, β4integrin; B, basal; Der, dermis; DP, dermal papilla; Epi, epidermis; K5, keratin5 (a pan-marker of basal keratinocytes); SB, suprabasal. Dotted lines denotethe dermoepidermal border.
Zhang et al. PNAS | February 8, 2011 | vol. 108 | no. 6 | 2271
DEV
ELOPM
ENTA
LBIOLO
GY
results provide functional evidence that nuclear YAP regulates theproliferative activity of skin progenitors and that its transition tothe cytoplasm is required for cells to execute terminal differenti-ation properly and to balance growth and lineage commitment.
Elevated Nuclear YAP Inhibits Apoptosis in Addition to PromotingProliferation and Suppressing Differentiation in Keratinocytes. Todissect further the mechanisms by which nuclear YAP controlstissue development in skin, we cultured primary mouse kerati-nocytes (MKs) from basal epidermal progenitors of P0 K14-rtTA/TRE-YAP(S127A) mice. When Dox was added to the culturemedium, nuclear YAP was markedly elevated (Fig. S2). To de-termine how YAP activation affects primary MKs, we quantifiedthe ability of K14-rtTA/TRE-YAP(S127A) MKs to form colonies(more than four cells) in the presence and absence of Dox. As
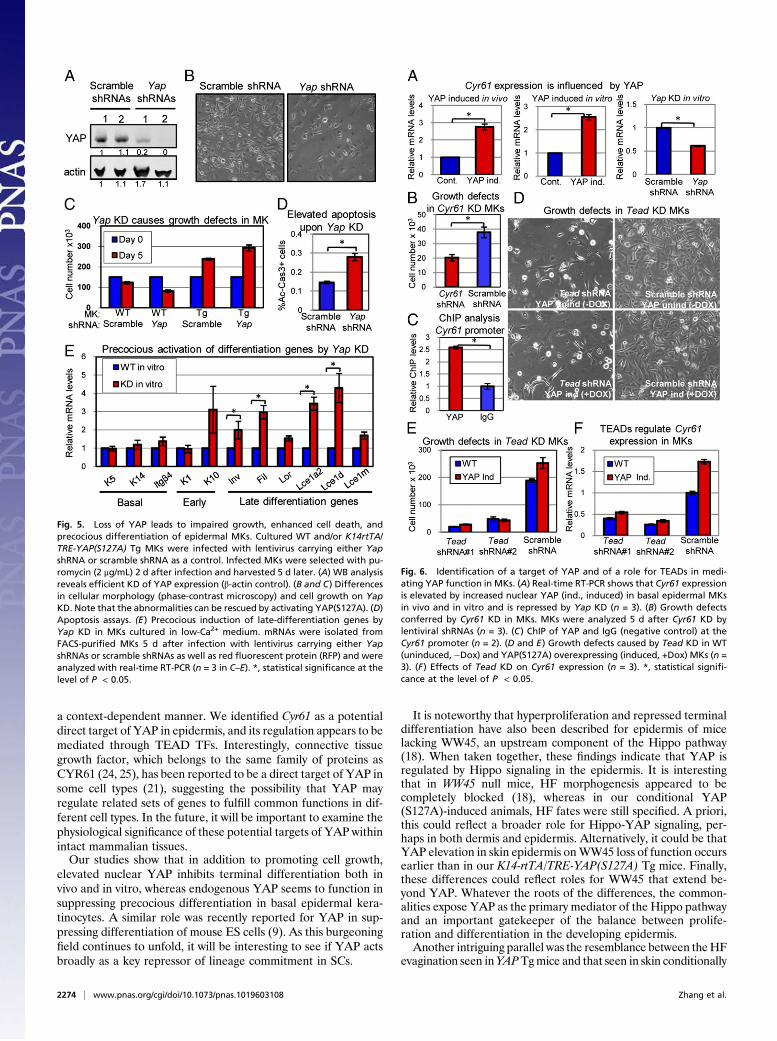
shown in Fig. 4A, a nearly 10-fold increase in colony-formingefficiency was observed when YAP(S127A) was induced. Notably,most of these colonies grew to be >2 mm in size, a characteristictypical of SCs (17). Moreover, these MKs displayed an approxi-mately twofold decrease in the percentage of activated Caspase 3(Ac-Cas3, a key apoptotic marker)-positive MKs (Fig. 4B). Thesedata showed that the dramatic increase in colony formation andsize was reflected not only in the enhanced proliferation of basalprogenitors but also in their survival.Finally, and consistent with our observations in vivo, aug-
menting nuclear YAP specifically restricted calcium-induceddifferentiation in vitro. RT-PCR analyses showed marked re-ductions in transcripts encoding early (K1 and K10) and late(involucrin, filaggrin, loricrin, Lce1a2, Lce1d, and Lce1m) dif-ferentiation markers (18) (Fig. 4C). Taken together, these find-
Fig. 2. Elevated nuclear YAP results in hyperthickening of interfollicular epidermis and evagination of HFs. (A) Dox-induced Tg pups were distinguished fromWT by their lack of a milk spot (*) and open eyes and mouth. (B) H&E staining reveals hyperthickening of Tg interfollicular epidermis accompanied by thinnerstratum corneum. (C) HF evaginations in Tg skin are evident by alkaline phosphatase (AP) enzymatic activity (black) in DP. (D) Semithin 1-μm sections of P0back-skins were stained with toluidine blue, and basal cell density was quantified (as discussed in the text). (E and F) Ultrastructure reveals aberrant cellularmorphology in the basal layer and evaginated HFs in Tg mice. B, basal; Der, dermis; DP, dermal papilla; Epi, epidermis; Gr, granular layer; Me, melaningranules (denoted with arrows in F, prevalent in hair shaft and its progenitor Mx cells); Sc, stratum corneum; Sp, spinous layer. (Scale bar, 10 μm.)
Fig. 3. YAP(S127A) induction leads to an expansion of basal-like layers, increased proliferation, and diminished terminal differentiation. (A–C) Immuno-fluorescence (IF) reveals enhanced proliferation in P0 YAP Tg epidermis. Note enhanced Ki67- and pH3-positive cells mostly in the basal layer of the epidermisand evaginating HFs but also in some suprabasal cells (arrows in expanded boxed areas). Note also the expanded layers of basal markers K5 and p63 in YAP Tgskin. (D–F) IF shows aberrant terminal differentiation in YAP Tg epidermis. B, basal; Der, dermis; Epi, epidermis; K1, keratin 1; Lor, loricrin; SB, suprabasal.Dotted lines denote the dermoepidermal border. (Scale bar,10 μm.)
2272 | www.pnas.org/cgi/doi/10.1073/pnas.1019603108 Zhang et al.
ings suggested that nuclear YAP promotes proliferation andsurvival of basal progenitors and maintains the cells in an un-differentiated state.
Knockdown of YAP Impedes Growth and Initiates Differentiation inMKs. To examine whether YAP is essential for promoting MKgrowth and suppressing differentiation, we conducted loss-of-function studies. Primary MKs were infected with lentivirus car-rying either Yap shRNA or scramble shRNA as a control, andefficient knockdown (KD) of YAP was validated by Western blot(WB) analysis (Fig. 5A). In comparison to the scramble control,YAP KD restricted the growth of MKs (Fig. 5B). This featurecould be rescued by expressing the YAP(S127A) mutant, whichwas not targeted by the Yap shRNAs used (Fig. 5C). In addition,as opposed to our gain-of-function studies, Yap KD resulted inelevated cell death as shown by the flow analysis of Ac-Cas3 inMKs (Fig. 5D).Importantly, under conditions that maintain WT MKs in an
undifferentiated state, some of the MKs in which Yap wasknocked down displayed a flattened squamous-like morphologytypical of MKs exposed to high calcium and induced to terminallydifferentiate (Fig. 5B). In contrast to basal genes whose expres-sion was not significantly affected, most of the differentiationgenes examined were significantly up-regulated by Yap KD (Fig.5E). This was accompanied by an elevation in expression of AP1family members known to govern the expression of these late-differentiation genes (14) (Fig. S5). Together, these findings in-dicate that YAP deficiency increases the propensity of basal cellsto undergo differentiation and/or apoptosis.
Identification of Cyr61 as a Target of YAP in MKs and TEAD asa Critical DNA-Binding Partner for YAP. To investigate further howYAP controls this balance, we conducted microarray analysis toidentify the transcriptional changes that occur in basal epidermalcells and/or MKs when (i) endogenous Yap is knocked down and(ii) nuclear YAP is enhanced (Dataset S1). Because YAP isthought to act predominantly as a coactivator, we focused on the21 probes that scored as being down-regulated by greater thantwofold in duplicate arrays when Yap was knocked down in MK(Dataset S1). These probes corresponded to nine known genes,one of which was Yap itself. We focused on one of these genes,Cyr61, because it was also up-regulated by greater than twofold invivo and in vitro within 24 h after YAP(S127A) induction and itsexpression was among the most markedly reduced by Yap KD.Real-time PCR confirmed the changes in Cyr61 that had surfacedfrom the array data (Fig. 6A).To assess whether these changes in Cyr61 expression might
be relevant to the proliferative differences we observed, weused lentiviral shRNA to knock down expression of Cyr61 in MK
(Fig. S6A). Indeed, efficient KD of Cyr61 significantly inhibitedMK growth (Fig. 6B). Furthermore, anti-YAP ChIP demon-strated an enrichment of YAP occupancy at the Cyr61 promoter(Fig. 6C), suggesting the possibility that Cyr61 may be a directtarget of YAP and play a key role in mediating YAP function incell growth regulation.YAP does not bind DNA on its own. Among many TFs known
to interact with YAP, the TEADs are the best studied geneticallyand biochemically as mediators of YAP’s proliferation-inducingfunction (11, 19–21). By RT-PCR, all four TEAD members weredetected in MKs (Fig. S6B). We therefore selected two shRNAscorresponding to sequences conserved among Tead1, Tead3, andTead4 (21). RT-PCR of transduced MKs exhibited efficient KDof the three expected Teads and, surprisingly, a reduction inTead2 as well (Fig. S6C). Additionally, both shRNAs potentlyinhibited cell growth, and, importantly, this growth defect couldnot be reversed by induction of YAP(S127A) (Fig. 6 D and E).Finally, Tead KD resulted in a significant reduction in Cyr61expression, and induction of YAP(S127A) failed to rescue it(Fig. 6F). Together, these results suggest that YAP functionstogether with TEADs to promote cell growth and that this oc-curs, at least partially, through regulating Cyr61 expression.
YAP and Skin Cancers. Mutations in the Hippo pathway have beenlinked to human cancers (22). Functioning at the end of theHippo pathway, YAP is itself an oncogene and its overexpressionpromotes tissue overgrowth and tumorigenesis in Tg mice (7, 10).Amplification of the Yap gene locus and/or overexpression ofYap has been reported in many common human cancers (22, 23).Given our findings on the epidermis, we wondered whether thismight also be the case for human skin cancers. Although a de-tailed investigation is beyond the scope of the present study, weconducted YAP immunohistochemical analyses on a panel of >50epithelial skin cancers. A significant portion of these (22 of 40squamous cell carcinomas and 10 of 14 basal cell carcinomas)showed an increase in YAP+ cells and/or elevation in nuclearYAP within the tumor masses (Fig. S7), supporting the oncogenicrole of YAP and its growth-promoting function in epidermisreported in this work.
DiscussionOur study unveiled a critical role for YAP in maintaining epi-dermal progenitors during development. Through genetic studies,we showed that elevating nuclear YAP levels leads to massiveexpansion of proliferative basal epidermal cells. Many pro-liferation-related genes have been reported to be affected by YAPin mammalian cells (22), but no common targets of YAP havebeen reported to date. Because YAP lacks intrinsic DNA-bindingactivity, its targets are dictated by its transcription partners in
Fig. 4. YAP(S127A) induction in MKs in vitro increases proliferation potential and inhibits apoptosis and differentiation. Dox-induced WT and K14-rtTA/TRE-YAP(S127A) Tg MKs were assayed 13 d and 1 d later, respectively, for differences in efficiency to form colonies (A, n = 3) and apoptosis (B, n = 3). Images arephase-contrast. (C) Calcium-induced activation of terminal differentiation genes is suppressed by YAP(S127A) induction in MKs. mRNAs were isolated 24 hafter the switch from low to high Ca+, and real-time RT-PCR assays were performed (n = 3). F, feeders. *, statistical significance at the level of P < 0.05.
Zhang et al. PNAS | February 8, 2011 | vol. 108 | no. 6 | 2273
DEV
ELOPM
ENTA
LBIOLO
GY
a context-dependent manner. We identified Cyr61 as a potentialdirect target of YAP in epidermis, and its regulation appears to bemediated through TEAD TFs. Interestingly, connective tissuegrowth factor, which belongs to the same family of proteins asCYR61 (24, 25), has been reported to be a direct target of YAP insome cell types (21), suggesting the possibility that YAP mayregulate related sets of genes to fulfill common functions in dif-ferent cell types. In the future, it will be important to examine thephysiological significance of these potential targets of YAP withinintact mammalian tissues.Our studies show that in addition to promoting cell growth,
elevated nuclear YAP inhibits terminal differentiation both invivo and in vitro, whereas endogenous YAP seems to function insuppressing precocious differentiation in basal epidermal kera-tinocytes. A similar role was recently reported for YAP in sup-pressing differentiation of mouse ES cells (9). As this burgeoningfield continues to unfold, it will be interesting to see if YAP actsbroadly as a key repressor of lineage commitment in SCs.
It is noteworthy that hyperproliferation and repressed terminaldifferentiation have also been described for epidermis of micelacking WW45, an upstream component of the Hippo pathway(18). When taken together, these findings indicate that YAP isregulated by Hippo signaling in the epidermis. It is interestingthat in WW45 null mice, HF morphogenesis appeared to becompletely blocked (18), whereas in our conditional YAP(S127A)-induced animals, HF fates were still specified. A priori,this could reflect a broader role for Hippo-YAP signaling, per-haps in both dermis and epidermis. Alternatively, it could be thatYAP elevation in skin epidermis onWW45 loss of function occursearlier than in our K14-rtTA/TRE-YAP(S127A) Tg mice. Finally,these differences could reflect roles for WW45 that extend be-yond YAP. Whatever the roots of the differences, the common-alities expose YAP as the primary mediator of the Hippo pathwayand an important gatekeeper of the balance between prolife-ration and differentiation in the developing epidermis.Another intriguing parallel was the resemblance between theHF
evagination seen inYAPTgmice and that seen in skin conditionally
Fig. 6. Identification of a target of YAP and of a role for TEADs in medi-ating YAP function in MKs. (A) Real-time RT-PCR shows that Cyr61 expressionis elevated by increased nuclear YAP (ind., induced) in basal epidermal MKsin vivo and in vitro and is repressed by Yap KD (n = 3). (B) Growth defectsconferred by Cyr61 KD in MKs. MKs were analyzed 5 d after Cyr61 KD bylentiviral shRNAs (n = 3). (C) ChIP of YAP and IgG (negative control) at theCyr61 promoter (n = 2). (D and E) Growth defects caused by Tead KD in WT(uninduced, −Dox) and YAP(S127A) overexpressing (induced, +Dox) MKs (n =3). (F) Effects of Tead KD on Cyr61 expression (n = 3). *, statistical signifi-cance at the level of P < 0.05.
Fig. 5. Loss of YAP leads to impaired growth, enhanced cell death, andprecocious differentiation of epidermal MKs. Cultured WT and/or K14rtTA/TRE-YAP(S127A) Tg MKs were infected with lentivirus carrying either YapshRNA or scramble shRNA as a control. Infected MKs were selected with pu-romycin (2 μg/mL) 2 d after infection and harvested 5 d later. (A) WB analysisreveals efficient KD of YAP expression (β-actin control). (B and C) Differencesin cellular morphology (phase-contrast microscopy) and cell growth on YapKD. Note that the abnormalities can be rescued by activating YAP(S127A). (D)Apoptosis assays. (E) Precocious induction of late-differentiation genes byYap KD in MKs cultured in low-Ca2+ medium. mRNAs were isolated fromFACS-purified MKs 5 d after infection with lentivirus carrying either YapshRNAs or scramble shRNAs as well as red fluorescent protein (RFP) and wereanalyzed with real-time RT-PCR (n = 3 in C–E). *, statistical significance at thelevel of P < 0.05.
2274 | www.pnas.org/cgi/doi/10.1073/pnas.1019603108 Zhang et al.

targeted for Dicer, encoding a core component of the microRNA(miRNA) processing machinery (26, 27). One possibility is thatYAP and certain miRNA(s) may antagonize each other’s functionin regulating expression of key proteins involved in HF morpho-genesis. Alternatively, Yap mRNA itself might be modulated bymiRNAs other than Hippo signaling. That said, hyperproliferationwas not a feature ofDicer-null epidermis; hence, such interactions,if they exist, would likely be tissue-specific.In searching for a possible mechanism underlying HF evagi-
nation, it is likely to be relevant that in WT embryos, endogenousYAP translocates from the nucleus to cytoplasm in the smallcluster of suprabasal cells that appear within the developing hairplacodes. It is tempting to speculate that nuclear YAP and ex-pression of its targets in these suprabasal cells may need to be shutdown to ensure the proper cell shape changes that adjacent basalcells need to invaginate. A role for YAP in regulating cytoskele-ton, polarity, and/or adhesive properties might also explain theelongated columnar shapes of YAP Tg basal cells. Although thedefects we observed could also be a reflection of cell crowdingwithin the basal layer, the underlying connection between YAPand these changes in cellular and tissue architecture merits at-tractive avenues for future investigations.
Materials and MethodsMice. Mice were housed and bred in the Association for Assessment andAccreditation of Laboratory Animal Care International-Accredited Compar-ative Bioscience Center animal facility at The Rockefeller University in ac-
cordance with university and National Institutes of Health guidelines. TRE-YAP(S127A) mice (10) were mated with K14-rtTA mice (14), and the geno-types were confirmed by PCR of toe DNA. Mice were fed Dox-containingfood (BIO-SERV) to induce YAP, or i.p. injections of Dox (100 μg) were ad-ministered for short-term induction (<3 d). EdU (Invitrogen) was adminis-tered to pregnant mice or to neonatal pups by i.p. injection (50 μg/g of EdU).
Lentivirus. Vesicular Stomatitis Virus G-pseudotyped lentivirus was pro-duced by calcium phosphate transfection of 293FT cells (Invitrogen) withpLKO.1 (carrying shRNA and either the puromycin or H2B-mRFP1 cassette)and helper plasmids pMD2.G and psPAX2 (Addgene plasmids 12259 and12260) (28). Infected MKs were selected with puromycin (2 μg/mL) 2 d afterinfection or FACS-isolated with RFP for subsequent experiments. shRNAsequences are listed in Table S1.
Statistics. For all graphs, data presented are mean value ± SE. P ≤ 0.05 wasaccepted for statistical significance, as determined by the Student’s t test(indicated by a bracket and asterisk in the figures).
ACKNOWLEDGMENTS. We thank F. D. Camargo for TRE-YAP(S127A) mice;N. Stokes, L. Polak, and D. Oristian for assistance in animal work; S. Williams,S. Beronja, and G. Livshits for reagents and advice on lentiviral experiments;the Bio-Imaging Resource Center, Flow Cytometry Resource Center, andGenomics Core Laboratory (Memorial Sloan–Kettering Cancer Center) fortechnical support; and members of the E.F. laboratory for advice and help.H.Z. was supported by National Research Service Award Training Grant CA09673 and is an American Cancer Society Postdoctoral Fellow (PF-09-042-01-DDC). This work was supported by a grant from the Starr Foundation (toE.F.). E.F. is an investigator at the Howard Hughes Medical Institute.
1. Yagi R, Chen LF, Shigesada K, Murakami Y, Ito Y (1999) A WW domain-containing yes-associated protein (YAP) is a novel transcriptional co-activator. EMBO J 18:2551–2562.
2. Vassilev A, Kaneko KJ, Shu H, Zhao Y, DePamphilis ML (2001) TEAD/TEF transcriptionfactors utilize the activation domain of YAP65, a Src/Yes-associated protein localizedin the cytoplasm. Genes Dev 15:1229–1241.
3. Sudol M, et al. (1995) Characterization of the mammalian YAP (Yes-associatedprotein) gene and its role in defining a novel protein module, the WW domain. J BiolChem 270:14733–14741.
4. Linn H, et al. (1997) Using molecular repertoires to identify high-affinity peptideligands of the WW domain of human and mouse YAP. Biol Chem 378:531–537.
5. Chen HI, Sudol M (1995) The WW domain of yes-associated protein binds a proline-rich ligand that differs from the consensus established for Src homology 3-bindingmodules. Proc Natl Acad Sci USA 92:7819–7823.
6. Huang J, Wu S, Barrera J, Matthews K, Pan D (2005) The Hippo signaling pathwaycoordinately regulates cell proliferation and apoptosis by inactivating Yorkie, theDrosophila Homolog of YAP. Cell 122:421–434.
7. Dong J, et al. (2007) Elucidation of a universal size-control mechanism in Drosophilaand mammals. Cell 130:1120–1133.
8. Ramalho-Santos M, Yoon S, Matsuzaki Y, Mulligan RC, Melton DA (2002) “Stemness”:Transcriptional profiling of embryonic and adult stem cells. Science 298:597–600.
9. Lian I, et al. (2010) The role of YAP transcription coactivator in regulating stem cellself-renewal and differentiation. Genes Dev 24:1106–1118.
10. Camargo FD, et al. (2007) YAP1 increases organ size and expands undifferentiatedprogenitor cells. Curr Biol 17:2054–2060.
11. Cao X, Pfaff SL, Gage FH (2008) YAP regulates neural progenitor cell number via theTEA domain transcription factor. Genes Dev 22:3320–3334.
12. Reddy BV, Rauskolb C, Irvine KD (2010) Influence of fat-hippo and notch signaling onthe proliferation and differentiation of Drosophila optic neuroepithelia. Develop-ment 137:2397–2408.
13. Fuchs E (2007) Scratching the surface of skin development. Nature 445:834–842.
14. Ezhkova E, et al. (2009) Ezh2 orchestrates gene expression for the stepwisedifferentiation of tissue-specific stem cells. Cell 136:1122–1135.
15. Nguyen H, Rendl M, Fuchs E (2006) Tcf3 governs stem cell features and represses cellfate determination in skin. Cell 127:171–183.
16. Blanpain C, Lowry WE, Pasolli HA, Fuchs E (2006) Canonical notch signaling functionsas a commitment switch in the epidermal lineage. Genes Dev 20:3022–3035.
17. Barrandon Y, Green H (1987) Three clonal types of keratinocyte with differentcapacities for multiplication. Proc Natl Acad Sci USA 84:2302–2306.
18. Lee JH, et al. (2008) A crucial role of WW45 in developing epithelial tissues in themouse. EMBO J 27:1231–1242.
19. Wu S, Liu Y, Zheng Y, Dong J, Pan D (2008) The TEAD/TEF family protein Scallopedmediates transcriptional output of the Hippo growth-regulatory pathway. Dev Cell14:388–398.
20. Zhang L, et al. (2008) The TEAD/TEF family of transcription factor Scalloped mediatesHippo signaling in organ size control. Dev Cell 14:377–387.
21. Zhao B, et al. (2008) TEAD mediates YAP-dependent gene induction and growthcontrol. Genes Dev 22:1962–1971.
22. Pan D (2010) The hippo signaling pathway in development and cancer. Dev Cell 19:491–505.
23. Steinhardt AA, et al. (2008) Expression of Yes-associated protein in common solidtumors. Hum Pathol 39:1582–1589.
24. Perbal B (2004) CCN proteins: Multifunctional signalling regulators. Lancet 363:62–64.25. Leask A, Abraham DJ (2006) All in the CCN family: Essential matricellular signaling
modulators emerge from the bunker. J Cell Sci 119:4803–4810.26. Yi R, et al. (2006) Morphogenesis in skin is governed by discrete sets of differentially
expressed microRNAs. Nat Genet 38:356–362.27. Andl T, et al. (2006) The miRNA-processing enzyme dicer is essential for the
morphogenesis and maintenance of hair follicles. Curr Biol 16:1041–1049.28. Beronja S, Livshits G, Williams S, Fuchs E (2010) Rapid functional dissection of genetic
networks via tissue-specific transduction and RNAi in mouse embryos. Nat Med 16:821–827.
Zhang et al. PNAS | February 8, 2011 | vol. 108 | no. 6 | 2275
DEV
ELOPM
ENTA
LBIOLO
GY





























