Embed Size (px)
Citation preview

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
3 4 0 R. T S C H E S C H E , F. K O R T E U N D L. R E I C H L E
Zur Biogenese der Folsäure und des Coenzyms F 2. Mitt.:1 Weitere Untersuchungen über die Natur der Wachstumswirkung
verschiedener Pteridin-Derivate bei Streptococcus faecalis R
V o n R U D O L F T S C H E S C H E , F R I E D H E L M K Ö R T E u n d L I E S E L O T T E R E I C H L E
Aus der Biodiemischen Abteilung des Chemischen Staatsinstituts der Universität Hamburg (Z. Naturforschg. 10 b, 34&—350 [1955]; eingegangen am 1. April 1955)
Es wird festgestellt, daß bei der Umsetzung gewisser Pteridin-Derivate mit Zuckern Dihydro-N-glykoside entstehen bei gleichzeitiger Oxydation eines Teil des Zuckers, z. B. bei D-Glucose zu D-Gluconsäure. Bei Strept. faecalis R erwies sich nur das Glucose-Derivat als wachstums-wirksam, nicht das D-Ribosid. 7-Oxy-pteroylglutaminsäure ist nicht Zwischenstufe der Fol-säure-Biogenese; es wird die Frage diskutiert, ob Pteroyl-L-glutaminsäure überhaupt auf dem Wege zum Aufbau des Coenzyms F liegt. Leucovorin und 10IV-Formyl-tetrahydro-fol-säure können ebenso wie Folsäure die Sulfonamidhemmung bei E. coli nicht aufheben. Es wird auf die Bedeutung der Permeabilität bei derartigen Versuchen hingewiesen.
Streptococcus faecalis R und Lactobazillus casei wer-den seit der Entdeckung der Folsäure (Pteroyl-L-glutamin-säure) zur mikrobiologischen Bestimmung dieses Wachs-tumsfaktors verwendet, 0,0003 y/ccm bzw. 0,00007 y/ccm sind für ein halbmaximales Wachstum der beiden Test-organismen ausreichend. Beide Organismen können nach heutiger Anschauung diese Verbindung nidit selbst bil-den und sind auf Zufuhr aus der Nährlösung angewie-sen. Nun läßt sich aber dieses Bedürfnis herabsetzen oder aufheben, wenn man die Folgeprodukte der durch Fol-säure biodiemisch katalysierten Reaktionen dem Nähr-medium zusetzt. Als solche gelten Purine, Thymin, Serin und Methionin, wobei jedoch noch Unterschiede in der Ansprechbarkeit bei beiden Organismen für diese Stoffe auftreten. So kann z. B. Lact, casei mit Thymin an Stelle von Folsäure nidit wachsen, wohl aber Strept. faec. In den letzten Jahren hat sich nun gezeigt, daß die Folsäure nidit selbst Biokatalysator bei der Bildung der genannten Bausteine ist, als der eigentliche Wirkstoff wird jetzt das Coenzym F angesehen, für das die Klärung der chemi-schen Konstitution nodi aussteht2. Als weitere Schritte auf dem Weg von der Folsäure zum Coenzym F sind noch Leucovorin und Tetrahydro-10iV-formyl-folsäure zu erwä-gen. Für alle 3 Pteridin-Derivate ist nodi unentschieden, ob sie stets ganz, nur teilweise oder überhaupt nicht obli-gate Zwischenstufen beim biochemischen Aufbau des Co-enzyms F sind, wobei audi noch die Verhältnisse bei den einzelnen Organismen große Untersdiiede aufweisen kön-nen. Für Escherichia coli wird von W a c k e r , G r i s e -b a c h , T r e b s t und W e y g a n d 3 angenommen, daß Folsäure nicht Intermediärprodukt ist. Die Autoren sehen darin die Ursadie, daß die Sulfonamidhemmung bei dem genannten Bakterium durch Folsäure nicht aufgehoben wird. Sulfonamide stören den biochemisdien Einbau von
1 R. T s c h e s c h e u. F. K ö r t e , Z. Naturforsdig. 8 b, 87 [1954].
- J. M. B u c h a n a n u. D. W. W i l s o n , Fed. Proc. 12, 647 [1953]; G. R. G r e e n b e r g , Fed. Proc. 13, 221 [1954]; J. Amer. diem. Soc. 75, 1458 [1954],
p-Aminobenzoesäure zu Folsäure bzw. Coenzym F. 2 Be-funde spredien nach diesen Autoren dafür, daß Folsäure nicht Zwischenstufe für den Aufbau des Coenzyms F sein kann: Einmal wird diese Verbindung von E. coli nicht resorbiert, und dann kann in den Bakterien nach Zufuhr von radioakt. p-Aminobenzoesäure keine radioaktive Fol-säure papierehromatographisch aufgefunden werden. Da-gegen wird eine andere Substanz festgestellt, die als Co-enzym F angesprochen wird. Beide Befunde sind jedoch nicht unbedingt beweisend, da die Außenwand von E. coli vielleidit für Folsäure nicht permeabel sein kann, und die Nichtauffindung im Papierchromatogramm könnte auch damit erklärt werden, daß diese Verbindung eine so sdmelle Weiterveränderung erfährt, daß nur das End-produkt der Reaktionskette, nämlidi Coenzym F, faßbar wird.
T s c h e s c h e und K ö r t e 1 haben 1953 Vorstellun-gen über einen möglichen Syntheseweg der Folsäure ver-öffentlicht, als dessen früheste nachweisbare Stufe das Pyrimidin-Derivat Vicin angesehen wurde. Dieses Gly-kosid sollte über Isoxanthopterin-carbonsäure, den glyko-sidierten 2-Amino-4.7-dioxy-pteridinaldehyd-6 und 7-Oxy-pteroylglutaminsäure in Folsäure übergehen können. Die-ser Reaktionsweg war aber nur für Strept. faecalis R de-monstrierbar, da das Folsäurebedürfnis dieses Organismus ansdieinend auf einer mangelnden Bildungsfähigkeit für Pteridin-Derivate beruht. Lact, casei war nicht in der Lage, mit Substanzen aus unserem Schema, angefangen vom Vicin, eine Vermehrung zu zeigen. Dagegen ist Xanthopterin nadi Erhitzen mit dem glucose-haltigen Nährmedium sowohl für Strept. faec. wie für Lact, casci ein Wachstumsfaktor, allerdings in ganz anderer Größen-ordnung wie Folsäure, nämlich mit etwa 30—50 //ccm. Es wurde vermutet, daß dieses Pteridin-Derivat in grund-sätzlich anderer Weise in den Weg zur Folsäure oder zum Coenzym F eingeschleust wird als die zuerst genann-
3 A. W a c k e r , H. G r i s e b a c h , A. T r e b s t u. F. W e y g a n d , Angew. Chem. 66, 326 [1954],

Z U R B I O G E N E S E D E R F O L S Ä U R E U N D D E S C O E N Z Y M S F 3 4 7
ten Verbindungen. Dazu kommt, daß der Wachstums-effekt des glyk. Xanthopterins nicht durch Sulfonamide in-hibierbar i s t w i e z. B. dies für den glykosidierten 2-Amino-4.7-dioxy-pteridinaldehyd-6 der Fall ist. Die Ver-hältnisse sind also bei den verschiedenen Organismen un-terschiedlich und nur schwer durchschaubar. Dies drückt sich auch in dem bekannten Verhalten gegen Sulfon-amide aus sowie in der Aufhebung der dadurch gesetz-ten Wachstumshemmung durch so verschiedene Stoffe wie p-Aminobenzoesäure, p-Aminobenzoylglutaminsäure, Thy-min, Pteroylglutaminsäure u. a. Faktoren 4.
Ein Ansatzpunkt, um weiteres Licht in diese kompli-zierten Verhältnisse zu bringen, schien uns die Aufklä-rung der Reaktionen, welche bei der Umsetzung von ein-fachen Pteridinen mit Zuckern beim Sterilisieren der Nährlösung eintreten. Die Beobachtung, daß Xantho-pterin erst beim Erhitzen im glucose-haltigen Milieu Wachstumsaktivität für Lact, casei erlangt, geht auf die Untersuchungen von E l i o n , H i t c h i n g s , S h e r -w o o d und V a n d e r W e r f f 5 zurück. Danach sollen L-Arabinose, D-Fructose, D-Ribose und Dioxyaceton den gleichen Effekt wie D-Glucose ergeben, nicht aber Form-aldehyd, Acetaldehyd, Rohrzucker und Glycerin. So be-handeltes Xanthopterin sollte auch für Strept. faecalls R wachstums-wirksam sein.
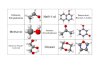
Wir konnten die Versuchsergebnisse der amerika-nischen Autoren im wesentlichen bestätigen und ha-ben jetzt geklärt, welche Umsetzung beim Erhitzen mit Zuckern bei einigen Pteridinen eintritt. Wir hatten schon früher angenommen, daß es sich dabei um eine N-Glykosid-Bildung handeln könnte, da nach saurer Hydrolyse solcher Umsetzungsprodukte der Wachstumseffekt wieder verschwand. Darüber hinaus zeigte sich aber nun, daß unter Oxydation eines Teils der angewandten Glucose Gluconsäure in der Reaktionslösung auftrat, die papierchromatographisch nachgewiesen werden konnte. In einem Vergleichs-ansatz ohne reaktionsfähige Pteridine konnte diese Säure nicht beobachtet werden. Dadurch wurde das gebildete N-Glykosid unter Aufnahme von Wasser-stoff zum Dihydroderivat reduziert. Wir geben die-sen Verbindungen die nachfolgende Konstitution, z. B . Dihydro-xanthopterin- und Dihydro-isoxanthopterin-A^-D-Glucosid (I und II) *. Während die Verbindung I I in wäßriger Lösung relativ stabil ist, tritt beim ent-sprechenden Xanthopterinderivat beim Stehen all-mählich Zersetzung unter Hydrolyse und Reoxydation
4 J. O. L a m p e n u. M. J o n e s , J. biol. Chemistry 164, 485 [1946]; 166, 435 [1946]; 170, 133 [1947]. R. T s c h e s c h e , K. S o e h r i n g u. K. H a r d e r , Z. Naturforschg. 2 b , 244 [1947]; E. A u h a g e n , Hoppe-Seyler's Z. physiol. Chem. 283, 195 [1948]; E. F. M ö l -l e r , F. W e y g a n d u. A. W a c k e r , Z. Naturforschg. 4 b, 100 [1949],
5 G. B. E l i o n , G. H. H i t c h i n g s , M. B . S h e r -w o o d u. H. V a n d e r W e r f f , Arch. Biochemistry 26, 337 [1950].
zum Xanthopterin ein. Besonders leicht bildet sich das entsprechende Glykosid der Dihydro-isoxantho-pterin-carbonsäure (III). Hier tritt die gefundene Re-aktion schon beim Stehen im Brutschrank bei 37° zu einem beträchtlichen Ausmaß ein, so daß nunmehr verständlich wird, warum Isoxanthopterin-carbon-säure schon ohne Erhitzen im glucose-haltigen Nähr-boden auf 100° Wachstumsaktivität zeigt.
Glucose
HoN—
Glucose II
OH H COOH
Glucose III
Wir hatten früher eine Tabelle von Pteridinen an-gegeben, die bei Strept. faec. beim Sterilisieren im glucose-haltigen Nährmedium wachstums-wirksam werden, und für solche, bei denen dies nicht der Fall ist 6 . Es schien nun möglich, daß der Unter-schied beider Gruppen darin zu suchen sein könnte, daß die einen unter den gewählten Umständen gly-kosidierbar wären und die anderen nicht. Nur die ersteren sollten eventuell einen Wachstumseffekt zei-gen. Die Untersuchung ergab, daß dies in der Tat so ist. Die merkwürdige Erscheinung, daß das 2-Amino-4.7-dioxy-6-methyl- und das 6-Aldehyd-Deri-vat des Pteridins wirksam werden und die entspre-chende 6-Oxymethyl-Verbindung nicht, findet darin ihre Erklärung. Worin allerdings die chemische Ur-sache des Unvermögens bei der 6-Oxymethyl-Verbin-dung besteht, muß vorerst dahingestellt bleiben.
Eine weitere Merkwürdigkeit bei den Versuchen von E l i o n und Mitarbb. 5 war die Unspezifität hin-
6 R. T s c h e s c h e , C.-H. K o e h n c k e u. F. K ö r t e , Z. Naturforschg. 5 b , 132 [1950]; R. T s c h e s c h e , F . K o r t e u. I. K o r t e , Z. Naturforschg. 5 b, 313 [1950]; dieselben, ebenda 6 b, 305 [1951].
* Über Herstellung und Konstitutionsbestimmung die-ser Verbindungen verweisen wir auf eine demnächst er-scheinende Arbeit in den Chemischen Berichten.

3 4 8 R. T S C H E S C H E , F. K Ö R T E U N D L. R E I C H L E
sichtlich der Zucker bei der Bildung der wachstums-wirksamen Derivate. Nun ist aber Xanthopterin we-gen der Unbeständigkeit seines Dihydro-Glykosides in wäßriger Lösung ein wenig geeignetes Untersu-chungsobjekt, wir haben daher die Versuche zunächst einmal mit D-Ribose und 6-Methyl-isoxanthopterin bei Strept. faec. wiederholt. Diese Anordnung hat den Vorzug, daß man ganz ohne Glucose im Nähr-medium auskommt, da dieser Organismus auch mit D-Ribose zu wachsen vermag. Dabei zeigte sich, daß Dihydro-6-methyl-isoxanthopterin-D-ribosid ganz ohne Wachstumseffekt ist. Wir halten es daher für möglich, daß die Unspezifität in den Versuchen von E 1 i o n
50 %
HO
30
20
70
0 20 10 60 80 700 y/cm3 —*-
Abb. 1. Wachstumseffekt in %, bezogen auf den Leerwert. a) Isoxanthopterin in 2% glucose-haltigem Nährmedium
auf 90° C erhitzt. b) Dihydro-isoxanthopterin-N-glucosid in 2% glucose-hal-
tigem Nährmedium auf 90° C erhitzt. Wachstum nach 5 Stdn. im folsäure-freien Nährmedium
nach T e p 1 e y und E 1 v e h j e m.
und Mitarbb. nur vorgetäuscht ist. Möglicherweise tritt in der Nährlösung relativ leicht ein Austausch der Ribose gegen Glucose auf. Wenn auch unsere Versuche in dieser Richtung noch nicht abgeschlos-sen sind, so möchten wir doch annehmen, daß nur Glucose-Derivate direkt wachstums-wirksam sind. Es scheint so, daß nur D-Glucose-Glykoside resorbiert werden, was vielleicht darauf hindeutet, daß enzy-matische Vorgänge bei der Aufnahme ins Innere des Testorganismus beteiligt sind.
Für eine solche Annahme spricht auch der Ver-gleich der Wachstumskurven (s. Abb. 1 und 2) der reinen Dihydro-pteridin-D-glucoside mit denen, die man erhält, wenn man Isoxanthopterin bzw. Xantho-pterin im glucose-haltigen Nährboden auf 100° er-hitzt. Unter diesen Umständen bleibt ein erheblicher Anteil der beiden Pteridine unverändert. Man sieht, daß besonders beim Isoxanthopterin mit steigender Konzentration die Wachstumswirkung immer mehr abnimmt, während das Dihydro-N-Glucosid einen stetigen Anstieg der Bakterienmenge verbürgt. Dies kann wohl nur so gedeutet werden, daß das freie
Isoxanthopterin einen hemmenden Einfluß auf die Resorption ausübt. Eine andere Möglichkeit wäre, daß es nach durchgeführter Aufnahme beider Stoffe ins Innere des Bakteriums zu einer Störung der Ver-wertung des Dihydro-N-glucosides in Gegenwart grö-ßerer Mengen Isoxanthopterin kommt. Das gleiche gilt auch für Xanthopterin, nur daß bei der man-gelnden Beständigkeit des Dihydro-N-glucosides die Wachstumskurve schon bald einen Knick erfährt. Diese Frage könnte allein durch Verwendung radio-aktiver Pteridin-Derivate geklärt werden.
Nachdem sich gezeigt hatte, daß offensichtlich nur die Dihydro-Derivate gewisser Pteridin-xV-glucoside
50 o/o HO
30
20
70
0 20 10 60 80 , 100 y/cm 3 —*-
Abb. 2. Wachstumseffekt in %, bezogen auf den Leerwert a) Xanthopterin in 2 % glucose-haltigem Nährmedium aul
90° C erhitzt. b) Dihydro-xanthopterin-lV-glucosid bei Zimmertempera-
tur steril zugegeben. Wachstum nach 5 Stdn. im folsäure-freien Nährmedium
nach T e p 1 e y und E 1 v e h j e m.
von Strept. faec. verwertet werden können, bei Lact, casei ist es nur das Xanthopterin-Derivat, erhob sich die Frage nach dem Einbau dieser Verbindungen in das von uns gegebene Schema der Folsäure-Bio-genese. Dabei soll das Problem des Xanthopterin-Derivates zunächst zurückgestellt werden, da es sich wahrscheinlich um einen Sonderfall handelt, wie schon ausgeführt worden ist. Unter diesen Umständen müßte ein glykosidierter und im Pteridinkern hydrier-ter 6-Aldehyd Zwischenstufe sein, doch sind unsere Untersuchungen gerade in diesem Punkte noch nicht abgeschlossen. Ein Glykosid tritt beim Erhitzen des 2-Amino-4.7-dioxy-pteridinaldehyd-6 (IV) mit Glu-cose auch auf, doch ist die Frage nach der Konstitu-tion der entstehenden Verbindung noch ungeklärt.
• /
/ /
S S
/ / / /
/ / T / 1 1 1
aJ

Z U R B I O G E N E S E D E R F O L S Ä U R E U N D D E S C O E N Z Y M S F 3 4 9
OH
COOH I
-CO-NH-CH I
(CH2)2-COOH
Anders liegen die Verhältnisse bei der 7-Oxy-pteroylglutaminsäure (V). Schon früher war aufgefal-len, daß diese Verbindung im Vergleich zu Pteroyl-L-glutaminsäure in Konzentrationen gleicher Größen-ordnung nur einen vergleichsweise geringen Wachs-tumseffekt entfaltete6 . J . O. L a m p e n * hat dieses Derivat jetzt gegen einen anderen Stamm von Strept. faec. untersucht und dabei als nicht wachstums-wirk-sam befunden. Wir konnten seine Ergebnisse mit dem gleichen Stamm bestätigen, während der unsrige mit derselben Substanz wieder etwa den alten Effekt zeigte. Daraus muß der Schluß gezogen werden, daß unser Stamm zu einer gewissen Verwertung der 7-Oxyfolsäure fähig ist, daß dies aber keine allgemeine Erscheinung für Strept. faec. darstellt. Da aber nun-mehr nachgewiesen ist, daß die Verwertung der Iso-xanthopterin-Verbindungen über Dihydro-N-gluco-side erfolgt, ist in unserem Schema auch kein rechter Platz mehr für diese Verbindung. Die neuen Befunde sprechen eher dafür, daß bei Strept. faec. Pteroyl-'L-glutaminsäure nicht unbedingt Zwischenstufe der Co-enzym-F-Bildung sein muß und daß die Biogenese sehr wohl direkt zu hydrierten Derivaten der Fol-säure führen kann. Auf welcher Stufe die Heraus-nahme des Sauerstoffs in der 7-Stellung in den 7-Oxypteridin - glykosiden erfolgt, bleibt eine offene Frage.
H,N—
C-CH 3
HC^ ^ C - C H 2 - C H 2 - O H S
Im Jahre 1951 teilte W o o 11 e y 7 die Beobachtung mit, daß 2-Amino-4-oxy-6-pteridylmethyl-(4'-methyl-5' - oxyaethyl - thiazolium) - Salze (VI) mit p-Amino-benzoyl-L-glutaminsäure in Gegenwart von Karpfen-Thiaminase Pteroylglutaminsäure bilden, die aller-dings nur mikrobiologisch nachgewiesen wurde.
7 D. W. W o o 11 e y , J. Amer. ehem. Soc. 75, 1898 [1951].
* Wir möchten Herrn Dr. J. O. L a m p e n , New Brunswick, auch hier für seine Versuche und die Über-lassung seines Stammes vielmals danken.
W o o 11 e y sah in dieser Umsetzung einen möglichen Weg der Folsäure-Biogenese. Es schien uns nicht ausgeschlossen, daß sich hier eine Möglichkeit bot, die folsäure-ähnliche Wirksamkeit von Dihydro-xanthopterin-D-glucosid bei Strept. faec. und Lact, ca-sei zu verstehen, die in grundsätzlich anderer Weise zustande zu kommen scheint als die der anderen einfachen Pteridin-Derivate bei Strept. faec. So könnte eventuell unter Einschiebung eines C-Atoms aus Xanthopterin mit dem Thiazolteil des Aneurins die W o o l l e y sehe Verb in dnug intermediär aufge-baut werden, um dann mit Thiaminase in Folsäure überzugehen. Um diese Frage zu untersuchen, haben wir die Verbindung (VI) hergestellt und geprüft, wie-weit sie bei den erwähnten Testbakterien ein Wachs-tum im folsäure-freien Nährmedium hervorbringen kann. Sie erwies sich als völlig wirkungslos in Men-gen von 0,0005—10 y/ccm, das gleiche gilt auch für ihren Phosphorsäure-Ester an der OH-Gruppe im Thiazolteil des Moleküls. Wir möchten damit nicht ausschließen, daß der W o o l l e y s c h e Bildungsmecha-nismus für Folsäure bei unseren Bakterien keine Be-deutung hat, da die Ursache des Mißerfolges viel-leicht durch die mangelnde Resorbierbarkeit dieser Verbindungen hervorgerufen sein mag. Auffällig bleibt nämlich die Tatsache, daß bei Strept. faec. mit Aneurin allein (20—40 y/ccm) eine etwa 10-proz. Wachstumssteigerung erzielbar ist. Der Thiazolteil des Aneurins war dagegen völlig wirkungslos.
Daß man bei Wachstums- und Enthemmungsver-suchen die Permeabilität für die zu prüfenden Ver-bindungen doch stärker berücksichtigen sollte, als das oft bisher geschehen ist, zeigen auch noch die beiden folgenden Versuche. Nachdem die Sulfonamidhem-mung des Wachstums bei E. coli durch Folsäure nicht aufgehoben werden kann 4 und W a c k e r u. a . 3 ge-zeigt haben, daß die Ursache darin zu suchen ist, daß diese Verbindung von dem Bakterium nicht aufge-nommen wird, haben wir uns der Prüfung von Leuco-vorin* und Tetrahydro-1(W-formyl-folsäure in diesem Versuch zugewandt. Es interessiert die Frage, ob mit diesen Verbindungen eine Enthemmung sulfonamid-vergifteter E. coli zu erzielen ist. Dies erwies sich ebenfalls nicht als möglich. Ob man daraus den Schluß ziehen darf, daß auch diese Verbindungen im Stoffwechsel von E. coli keine Rolle spielen, erscheint uns zweifelhaft. Wir sind eher geneigt anzunehmen, daß auch hier ein Permeabilitäts-Phänomen vorliegt. Angewandt wurde der Stamm E. coli Nr. 1883 auf
* Herrn Dr. Z. Z a k r z e w s k i , New Haven, danken wir sehr für die Überlassung einer Probe Leucovorin.

3 5 0 W. B R O S E R
Teply-Elvehjem-Nährmedium, als Sulfonamid wurde Sulfathiazol in Mengen von 12,5—25 7/ccm verwen-det. Leucovorin kam in Mengen von 0,05—0,0005 7/ccm und Tetrahydro-10N-formylfolsäure mit 0,01 bis 0,00005 y/ccm zur Anwendung.
B e s c h r e i b u n g d e r V e r s u c h e
Die mikrobiologisdien Teste wurden wie bereits be-schrieben durchgeführt1'6. Die Vorpassage der Bakterien wurde jeweils im optimalen Nährmedium angesetzt und die Versuchsröhrchen daraus mit der Öse beimpft.
2 - A m i n o - 4 - o x y - 6 - p t e r i d y l m e t h y l -( 4 ' - m e t h y l - 5 ' - o x y a e t h y l - t h i a z o l i u m ) -b r o m i d 2,8 g 2-Amino-4-oxy-pteridin-a-bromessigsäure-6 8 wer-
den in 10 ccm gesättigter HBr-Eisessig-Lösung 5 Min. erhitzt und die Lösung dann unter Feuchtigkeits-Aus-schluß im Vakuum eingedampft. Man versetzt jetzt mit 50 ccm wasserfreiem Glykol und 2 g 4-Methyl-5-/?-oxy-aethylthiazol und erhitzt unter Rühren und Feuchtig-keitsabsdiluß 1 Stde. auf 110°. Beim Eingießen in 500
8 R. T s c h e s c h e , F. K ö r t e u. R. P e t e r s e n , Chem. Ber. 84, 579 [1951].
ccm Eiswasser fallen 1,5 g des Pteridin-Derivates aus, das durch Umfällen aus n/lO-NaOH und Kristallisation aus Wasser gereinigt wird.
C1 3H1 5N602BrS (396,3) Ber. S 8,09 Br 25,39. Gef. S 7,49 Br 25,10.
P h o s p h o r s ä u r e - E s t e r
1 g des vorstehenden Pteridin-Derivates wird mit 20 g einer im Vakuum bei 150° entwässerten 85-proz. Phos-phorsäure 20 Min. auf 120° erhitzt. Nach dem Abkühlen wird mit konz. Ammoniaklösung vorsichtig neutralisiert und mit einer Lösung von AgN03 das Silbersalz gefällt. Man filtriert ab und zerlegt das Salz unter Rühren mit Schwefelwasserstoff. Nach dem Abfiltrieren löst man das Pteridinderivat in n/lO-NaOH und fällt bei pH 4 um. Beim Umkristallisieren aus Wasser enthält man 10 mg eines teilweise kristallisierten Produktes, das für die mi-krobiologischen Versuche verwendet wurde.
C13H1605N6S Br P (476,3) Ber. P 6,50; Gef. 5,80.
Wir danken der D e u t s c h e n F o r s c h u n g s -g e m e i n s c h a f t , Bad Godesberg, auch an dieser Stelle für die finanzielle Unterstützung dieser Arbeit.
Zur Prototropie des Kristallvioletts V o n W . B R O S E R
Aus dem Institut für Organische Chemie der Freien Universität Berlin, Berlin-Dahlem (Z. Naturforschg. 10 b, 350—353 [1955]; eingegangen am 28. Februar 1955)
Es werden die Prototropie-Gleichgewichte zwischen den einzelnen Ionisationsstufen des Kristallvioletts spektrophotometrisch gemessen. Die mesomeren ein-, zwei- und dreisäurigen Kationen sind miteinander durch Prototropie-Gleichgewichte verbunden, die sich momentan einstellen. Daneben entsteht in einer langsamen Reaktion ein farbloses Carbinolkation, für das eine dreifache positive Ladung ermittelt wurde. Die Gleichgewichtskonstanten für die einzelnen Reaktionen betragen KS l = 1,09 • 10-2 Mol/Z; KS2 = 1,56 • 10- 1 Mol II; KS s = 1,73 • 10-5 (MolII)
In einer früheren Arbeit1 wurden die Säure-Basen-
Gleichgewichte — die Prototropie-Gleichgewichte — zwi-schen den einzelnen lonisationsstufen des Fuchsins un-tersudit. Dabei wurde gefunden, daß ein sich unmeßbar schnell einstellendes Gleichgewicht zwischen dem 1-säu-rigen und dem 2-säurifen Kation besteht, und daß in einer langsamen Reaktion ein 3-säuriges Kation gebildet wird. Die einzelnen Ionisationsstufen F+, FH++ und FH_>3+ des Fuchsins wurden dabei als mesomere Kationen angenommen (vgl. Tab. 1). Nun erscheint es aber merk-würdig, daß sich das Gleichgewicht zwischen den meso-meren F + und F H + + momentan, zwischen FH++ und dem ebenfalls als mesomer angesehenen FH23+ langsam einstellt. Es liegt daher die Vermutung nahe, daß es sich bei dem in langsamer Reaktion gebildeten 3-säurigen
1 W. B r o s e r u. R. H a v e m a n n , Z. Naturforsdig. 5 a, 385 [1950].
Ion nicht um das mesomere Kation FHa3+, sondern um das Carbinolkation FHs • OH3+ handelt und daß sich in-folge ungünstiger spektroskopischer Bedingungen das mesomere 3-säurige Kation FH23+ der Beobachtung ent-zogen hat.
Günstiger liegen die experimentellen Bedingungen bei dem analog gebauten Kristallviolett. Hier beobachtet man nämlich beim Übergang zu kleineren p^-Werten ein momentanes Umschlagen der Farbe von Violett über Grün in Gelb, wobei die grüne und gelbe Farbe jedoch nicht beständig sind, sondern allmählich verblaßt. Da das Kristallviolett somit geeignet erscheint, Aufschlüsse über das Verhalten des Fudisins zu geben, und da anderer-seits die Prototropie-Verhältnisse des Kristallvioletts ge-nau bekannt sein mußten, ehe wir2 die Untersudiung
2 W. B r o s e r u. W. L a u t s c h , Z. Naturforsdig. 8 b, 711 [1953].

















![Synonymaverzeichnis 6. Synonymaverzeichnis Acetaldehyd [Acetaldehyd] Acetan [Ethylen] Aceton [Aceton] Acetoxylsäure [Essigsäure] Acetylentrichlorid [Trichlorethylen] Acetylhydrür](https://img.pdfslide.net/doc/110x75/5e1d817c0b1a5723a07c9249/synonymaverzeichnis-6-synonymaverzeichnis-acetaldehyd-acetaldehyd-acetan-ethylen.jpg)










