
NOTE
Do otolith increments allow correct inferences about ageand growth of coral reef fishes?
D. J. Booth
Received: 10 June 2013 / Accepted: 12 November 2013 / Published online: 23 November 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Otolith increment structure is widely used to
estimate age and growth of marine fishes. Here, I test the
accuracy of the long-term otolith increment analysis of the
lemon damselfish Pomacentrus moluccensis to describe
age and growth characteristics. I compare the number of
putative annual otolith increments (as a proxy for actual
age) and widths of these increments (as proxies for somatic
growth) with actual tagged fish-length data, based on a
6-year dataset, the longest time course for a coral reef fish.
Estimated age from otoliths corresponded closely with
actual age in all cases, confirming annual increment for-
mation. However, otolith increment widths were poor
proxies for actual growth in length [linear regression
r2 = 0.44–0.90, n = 6 fish] and were clearly of limited
value in estimating annual growth. Up to 60 % of the
annual growth variation was missed using otolith incre-
ments, suggesting the long-term back calculations of oto-
lith growth characteristics of reef fish populations should
be interpreted with caution.
Keywords Age validation � Coral reef damselfish �Growth � Otolith back calculation �Otolith increments � Pomacentrus moluccensis
Introduction
A key goal of ecological research is to understand how
species vary in their persistence. Performance measures
include survivorship and growth, and studies often assess
how these demographic parameters vary temporally and
spatially. For marine fishes, a structure often used to
hindcast survival and growth is the otolith or earstone,
which accretes rings that have been calibrated both as
annual (Fowler and Short 1998) and daily (Panella 1971).
Validation of these rings as annual can be done in the
laboratory by staining otoliths and holding fish for over a
year, or in the field using a marginal otolith increment
approach (Fowler and Short 1998) or by comparing with
known age of a tagged fish. The latter is the most reliable
but is rarely done since it requires a long-term (multiyear)
field monitoring program. Shorter-term field annual ring
validation consisted of marking the fish with a vital stain
such as tetracycline and recapturing the fish after 1–2 years
(e.g., Fowler and Short 1998) and thus cannot be applied to
the entire life span of longer lived fish.
More recently, the size of increments between otolith
rings has been used as a proxy for somatic growth,
allowing individual and population trajectories of growth
to be assessed (Campana 1990). Field time is also mini-
mized compared to mark-recapture, and a selection of
cohorts can be sampled at each site by sampling otoliths at
one point in time. However, assumptions of daily and
annual formation of rings and use of increments as growth
proxies have to be validated (Campana 1990). Regarding
the use of otolith increments as proxies for growth stanzas,
studies have shown that increments can decouple for
somatic growth, for short or longer periods (e.g., Wright
et al. 1990). While a number of statistical assumptions
must go into choice of back calculation model, validation
Communicated by Biology Editor Dr. Glenn Almany
D. J. Booth (&)
School of the Environment, University of Technology, Sydney,
PO Box 123, Broadway, NSW 2007, Australia
e-mail: [email protected]
123
Coral Reefs (2014) 33:255–258
DOI 10.1007/s00338-013-1105-2
of how otolith growth relates to somatic growth is often
lacking (Vigliola et al. 2000).
Here, based on a long-term study on the Great Barrier
Reef of a common coral reef damselfish (Pomacentrus
moluccensis), I ‘‘ground-truth’’ otolith-based estimates of
longevity and growth against actual data from tagged fish.
Materials and methods
I tagged fish at two sites in and adjacent to One Tree Island
lagoon, in the southern Great Barrier Reef (23�300S,
152�060E) as part of a long-term study of spatial and
temporal demographics. Cohorts of newly recruited fish
(size = 15 mm TL on average; 25–35 days post-hatching
after planktonic larval dispersal) were tagged with Visual
Implant Flourescent Elastomer paint and individuals mea-
sured (total length, TL mm) and followed annually. As a
part of the study, commenced in 2000, I recaptured fish and
recorded body length annually (mm TL). Six tagged fish
were recaptured and killed in 2006 (2 from one site and 4
from another) and measured in situ. Fish were collected
using SCUBA and with clove oil anaesthetic, and frozen
for transport to the University of Technology, Sydney,
where otoliths (sagittae) were removed by dissection and
stored dry until ready for processing. Otoliths were ground
to the primordium using 3 lm lapping film; wet polished
with a PMT 08A-10 multiflex 8’’ diameter polishing cloth
using 0.02–0.06 lm colloidal silica on a Kent 3—auto-
matic polishing unit; and then etched using a 5 % solution
of EDTA (ethylenedinitrilo tetra acetic acid). Prepared
otoliths were viewed at 40–809 on an Olympus BH2-
RFCA compound microscope, images stored, and putative
annual increments counted. The number of growth incre-
ments was counted along the most visible axis of the otolith
three times, with the mean taken as the age of the fish. If
counts differed by more than 5 %, otoliths were reexam-
ined and if subsequent counts again varied by over 5 %,
otoliths were rejected (sensu Campana and Neilson 1982).
Increment widths (primordium to putative Year One
Increment, Year One to Year Two Increment, etc.) were
measured along a consistent axis in microns, from the
digital images using Image J software.
Results and discussion
The same six fish that had been externally tagged soon after
settlement were monitored yearly for body size and col-
lected at year six for otolith age determination and mea-
surement of growth increments. First, putative annual age
rings on otoliths of P. moluccensis were validated, i.e., for
all six fish that were recaptured after 6 years, there were 6
opaque zones. Second, for each fish, the relationship
between annual actual somatic growth (mm TL) and otolith
increment width was plotted. If otolith increment width
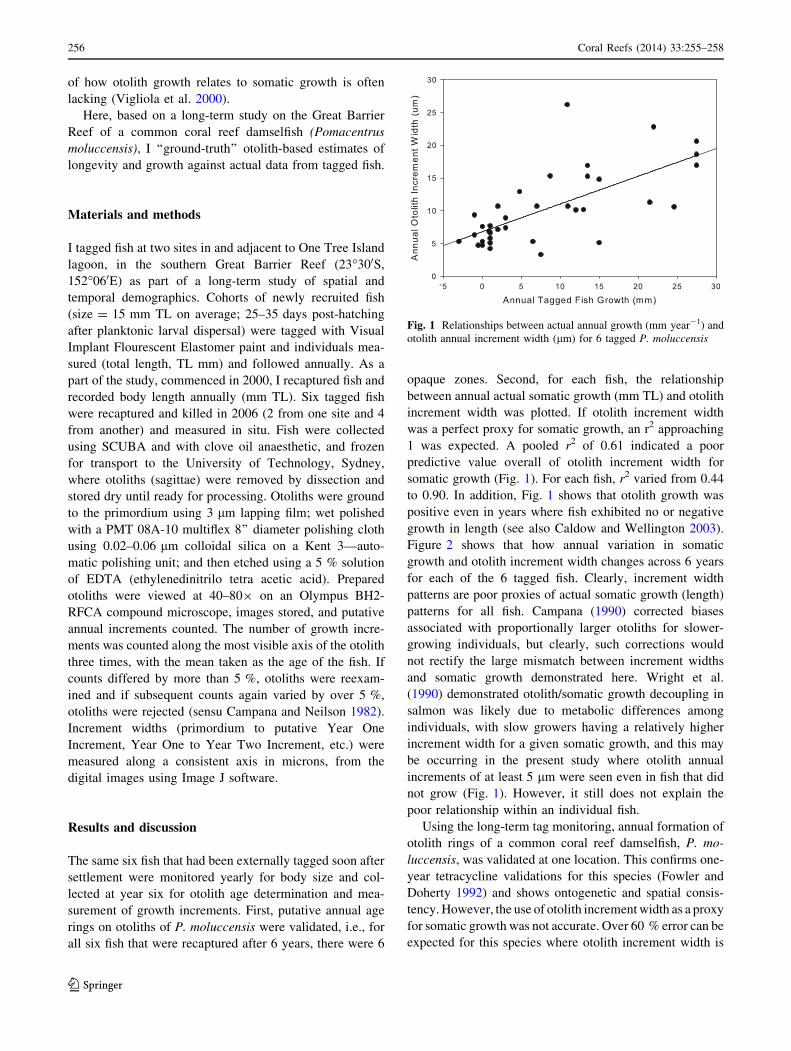
was a perfect proxy for somatic growth, an r2 approaching
1 was expected. A pooled r2 of 0.61 indicated a poor
predictive value overall of otolith increment width for
somatic growth (Fig. 1). For each fish, r2 varied from 0.44
to 0.90. In addition, Fig. 1 shows that otolith growth was
positive even in years where fish exhibited no or negative
growth in length (see also Caldow and Wellington 2003).
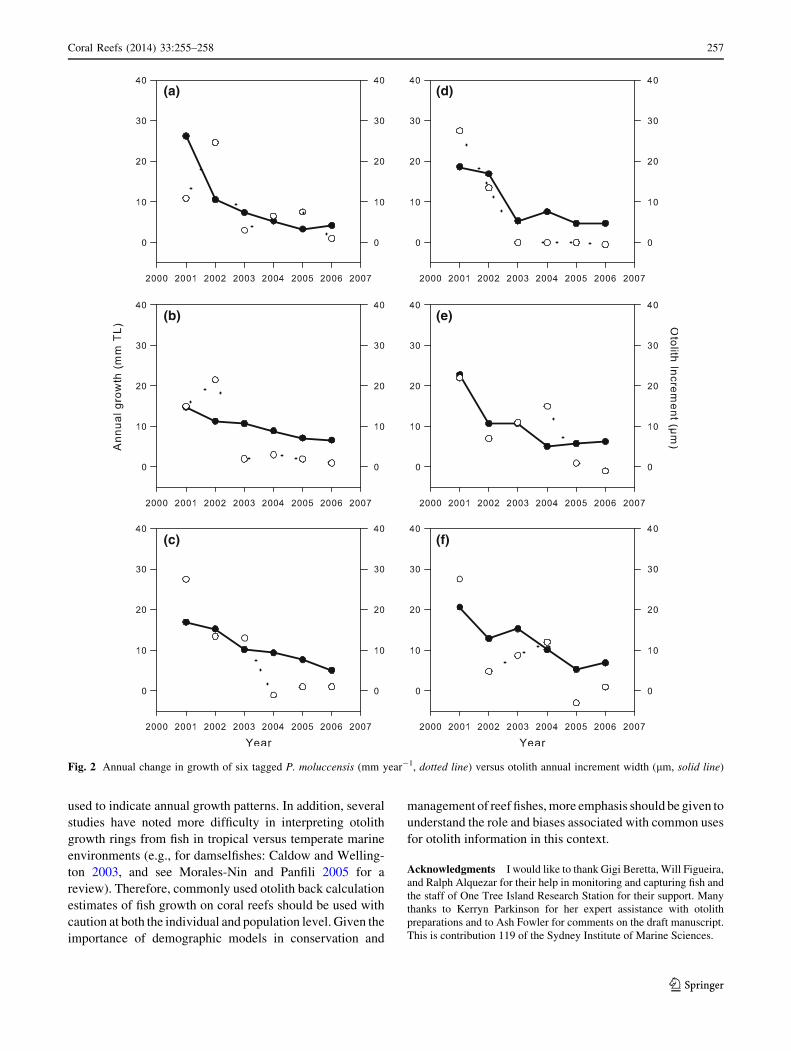
Figure 2 shows that how annual variation in somatic
growth and otolith increment width changes across 6 years
for each of the 6 tagged fish. Clearly, increment width
patterns are poor proxies of actual somatic growth (length)
patterns for all fish. Campana (1990) corrected biases
associated with proportionally larger otoliths for slower-
growing individuals, but clearly, such corrections would
not rectify the large mismatch between increment widths
and somatic growth demonstrated here. Wright et al.
(1990) demonstrated otolith/somatic growth decoupling in
salmon was likely due to metabolic differences among
individuals, with slow growers having a relatively higher
increment width for a given somatic growth, and this may
be occurring in the present study where otolith annual
increments of at least 5 lm were seen even in fish that did
not grow (Fig. 1). However, it still does not explain the
poor relationship within an individual fish.
Using the long-term tag monitoring, annual formation of
otolith rings of a common coral reef damselfish, P. mo-
luccensis, was validated at one location. This confirms one-
year tetracycline validations for this species (Fowler and
Doherty 1992) and shows ontogenetic and spatial consis-
tency. However, the use of otolith increment width as a proxy
for somatic growth was not accurate. Over 60 % error can be
expected for this species where otolith increment width is
Fig. 1 Relationships between actual annual growth (mm year-1) and
otolith annual increment width (lm) for 6 tagged P. moluccensis
256 Coral Reefs (2014) 33:255–258
123
used to indicate annual growth patterns. In addition, several
studies have noted more difficulty in interpreting otolith
growth rings from fish in tropical versus temperate marine
environments (e.g., for damselfishes: Caldow and Welling-
ton 2003, and see Morales-Nin and Panfili 2005 for a
review). Therefore, commonly used otolith back calculation
estimates of fish growth on coral reefs should be used with
caution at both the individual and population level. Given the
importance of demographic models in conservation and
management of reef fishes, more emphasis should be given to
understand the role and biases associated with common uses
for otolith information in this context.
Acknowledgments I would like to thank Gigi Beretta, Will Figueira,
and Ralph Alquezar for their help in monitoring and capturing fish and
the staff of One Tree Island Research Station for their support. Many
thanks to Kerryn Parkinson for her expert assistance with otolith
preparations and to Ash Fowler for comments on the draft manuscript.
This is contribution 119 of the Sydney Institute of Marine Sciences.
(a)
(b)
(c)
(d)
(e)
(f)
Fig. 2 Annual change in growth of six tagged P. moluccensis (mm year-1, dotted line) versus otolith annual increment width (lm, solid line)
Coral Reefs (2014) 33:255–258 257
123

References
Caldow C, Wellington GM (2003) Patterns of annual increment
formation in otoliths of pomacentrids in the tropical western
Atlantic: implications for population age-structure examination.
Mar Ecol Prog Ser 265:185–195
Campana SE (1990) How reliable are growth back-calculations based
on otoliths? Can J Fish Aquat Sci 47:2219–2227
Campana SE, Neilson JD (1982) Daily growth increments in otoliths
of Starry Flounder (Platichthys stellatus) and the influence of
some environmental variables in their production. Can J Fish
Aquat Sci 39:937–942
Fowler AJ, Doherty PJ (1992) Validation of annual growth
increments in the otoliths of two species of damselfish from
the Southern Great Barrier Reef. Aust J Mar Freshw Res
43:1057–1068
Fowler AJ, Short DA (1998) Validation of age determination from
otoliths of the King George whiting Sillaginodes punctata
(Perciformes). Mar Biol 130:577–587
Morales-Nin B, Panfili J (2005) Seasonality on the deep-sea and
tropics revisited: what can otoliths tell us? Mar Freshw Res
56:585–598
Panella G (1971) Fish otoliths: Daily growth layers and periodical
patterns. Science 173:1124–1126
Vigliola L, Harmelin-Vivien M, Meekan MG (2000) Comparison of
techniques of back-calculation of growth and settlement marks
from the otoliths of three species of Diplodus from the
Mediterranean Sea. Can J Fish Aquat Sci 57:1291–1299
Wright PJ, Metcalfe NB, Thorpe JE (1990) Otolith and somatic
growth rates in Atlantic salmon parr, Salmo salar L: evidence
against coupling. J Fish Biol 36:241–249
258 Coral Reefs (2014) 33:255–258
123
Recommended

















