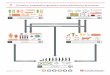
Evolutionary operator of the population.Selection

Two level populations
From the point of view of genetics population has two levels of organization, namely zygote and gamete levels, tied together by the processes of meiosis and fertilization. Usually we consider a population as consisting of zygotes or, more precisely, of organisms which are genetically identified with zygotes. In parallel with the population of zygotes there exists and evolves the population of gametes, or the gamete pool of a given population.

f
BBAA AB
m
AA
AB
AB
BB
½AA+½AB
½BB+½AB
½BB+½AB ¼AA+¼BB+½AB
AA
BB
AB ½AA+½AB
Let state of population is xAA,xBB,xAB
(AA,AA) - xAAxAA ; (AA,BB) – xAAxBB; (AA,AB) - xAA xAB; (BB,BB) - xBB xBB; (BB,AB) - xBB xAB; (AB,AB) - xAB xAB
(xAA ) ´ = (xAA)2 + xAAxAB + ¼(xAB ) 2
(xBB ) ´ = (xBB)2 + xBBxAB + ¼(xBB ) 2
(xAB ) ´ = 2xAAxBB + xAAxAB + xBBxAB + ½(xAB ) 2
Autosomal locus

(xAA )´ = (xAA)2 + xAAxAB + ¼(xAB ) 2 = (xAA+ ½xAB )2
(xBB )´ = (xBB)2 + xBBxAB + ¼(xBB ) 2 = (xBB+ ½xAB )2
(xAB )´ = 2xAAxBB + xAAxAB + xBBxAB + ½(xAB ) 2 = 2(xAA+ ½xAB )(xBB+ ½xAB )
p = (xAA+ ½xAB ); q = (xBB+ ½xAB );
p+q=1p and q is the frequencies of alleles A and B in the population.
(xAA )´ = p2; (xBB ) ´ = q2 ;(xAB )´ = 2pq;
p`=(xAA )´ + 1/2(xAB )´ = p2 +pq=p(p+q)=p
q`=(xBB )´ + 1/2(xAB )´ = q2 +pq=q(p+q)=q
p`= p2 +pq; q` = q2 +pq.

II. X-linkage
Let distributions genotypes A1A1, A2A2, A1A2 in female part of current generation are (x11,x22,x12) accordingly, and distributions genotypes A1, A2 in male part of current generation are (y1,y2). As usual x and y nonnegative and x11+x22+x12=1; y1+y2=1.
Evolutionary equations of male part of population
y1’=x11y1+x11y2+ ½x12y1+ ½x12y2
y2’=x22y1+x22y2+ ½x12y1+ ½x12y2

II. X-linkage
Evolutionary equations of female part of population
x11= x11y1+ ½x12y1
x22= x22y2+ ½x12y2
x12=x11y2+x22y1+ ½ x12y1+ ½ x12y2

II. X-linkage
Evolutionary operator of the population
y1’=x11y1+x11y2+ ½x12y1+ ½x12y2; y2
’=x22y1+x22y2+ ½x12y1+ ½x12y2
x11’= x11y1+ ½x12y1; x22
’= x22y2+ ½x12y2
x12’=x11y2+x22y1+ ½ x12y1+ ½ x12y2
Let pf= x11+ ½x12; qf= x22+ ½x12; pm=y1; qm=y2
Then y1’=pf, y2
’=qf genotype-gene
x11’=pfpm, x22
’=qfqm, x12’=pfqm+pmqf connection
pf’= x11
’+ ½x12’ = pfpm+ ½ (pfqm+pmqf)=
½ pf (pm+qm)+½ pm (pf+qf)=½ (pf + pm);
pf+qf=x11+x22+x12=1; pm+qm=y1+y2=1
pm’ = y1
’= pf.
pf, qf -frequencies A1 and A2 in female part of population; pm, qm -frequencies A1 and A2 in male part of population

II. X-linkage
Evolutionary operator of the population (on gene level)
pf’= ½ (pf + pm); qf
’= ½ (qf + qm); pm
’ = pf; qm’ = qf

Two level populations
Diploid organizms
Genotypes
AA, aa, Aa
Haplod hamete
Alleles
A, a
Diploid organizms
Genotypes
AA, aa, Aa
Generation N
Generation
N+
1

One-locus multiallele systemsLet A1, A2,…, As - set of alleles
Let p1, p2,…, ps –frequency alleles in current generation (of the gametes)(p1+ p2 + p3+… + ps=1)
?
(p1, p2,…, ps)
Set of possible zygotes {pipj}
p1p2 p1p1
p2p1 p5p2
p7p11

One-locus multiallele systems
Evolutionary equation
pi’ = pi p1+ pi p2 + pi p3+… + pi ps
pi’ = pi (p1+ p2 + p3+… + ps)
pi’ = pi
Zygote pipj produce gamete pi or pj
Zygote frequences: xij=pipj

Multiallele X-linkage system
pf1, pf2,…, pfspm1, pm2,…, pms
Let A1, A2,…, As - set of alleles by X-linkage loci
pf1, pf2,…, pfs –frequency gametes of the female originspm1, pm2,…, pms –frequency gametes of the male origins
(pY) Gametes level
pfi pmj pfiZygotes level
p’m1, p’
m2,…, p’ms Gametes levelp’
f1, p’f2,…, p’
fs
p’fi= ½(pfi+pmi)
p’mi= pfi
Equilibira conditions pfi=pmi (i=1,2,…,s)

An ideal population
1. Discrete non-overlapping generations
2. Allele frequencies are identical in males and females
3. Panmictic population: Mating of individuals is made at random
4. Population size is very large (infinite)
5. There is no migration (closed population)
6. Mutations can be ignored
7. Selection does not affect allele frequencies (neutral alleles)
Properties of an ideal diploid population studied at
a single autosomal locus with Mendelian inheritance

• Predictions from Hardy-Weinberg :• • IF… • – No selection • – No mutation • – No migration • – Random mating • • THEN… • – Allele frequencies remain constant • – Genotype frequencies predictable

HW for locus with dominant alleles

Blood groups
• A,B,O –alleles allel enzyme
• A O B dominance A A
• AA, AO, = A B B
• BB, BO, = B O -
• AB = AB
• OO = O


The ABO Blood Group
A, B, O –alleles A and B dominant to O
Blood type Genotypes Frequency
A AA,AO RA (= pA2 +2pApO)
B BB,BO RB (= pB2 +2pBpO )
AB AB RAB (= 2pApB )
O OO R0 (= pO2)
If RA, RB, RAB,and RO – are the observed frequencies of the blood type, we have pO= (RO) ½, (pA+ pO)2 = RA+
RO; …
HW

Selection

Example. Selection against recessive lethal gene

p`= p2 +pq; q` = q2 +pq.
p`= p2 +pq; q` = q2 +pq.
pqp
pqq
pqp
pqpp
2';
2'
22
2
22
12
2
)1(0122
1
2'
pppp
p
pqp
pqpppp pq

Trajectory calculator
p q generation
4 Go
reset
Current state (point) Next state (point)
pqp
pqq
pqp
pqpp
2';
2'
22
2

TRAJECTORY CALCULATION AND VIZUALIZATION
Letal1.exe

Dominant lethal allele
pqp
pqq
pqp
pqpp
2';
2'
22
2

Thalassemia A very large number of different mutations induce either a-thalassemia (a reduction in the synthesis rate of Hb a-chains) or b-thalassemia (a reduction in the synthesis rate of Hb b-chains). Both of these classes of mutations can induce malarial resistance in heterozygotes, but once again at the expense of anemia (which depending upon the exact nature of the thalassemia, can vary from virtually none to lethal) in the homozygotes. Thalassemia is found in high frequency in many historically malarial regions of the world in Africa, the Mediterranean, and Asia.

Selection in case Thalassemia
p`= p2 +pq; q` = q2 +pq
p`= 0.89p2 +pq; q` = 0.2q2 +pq

22
2
22
2
2.0289.0
2.0';
2.0289.0
89.0'
qpqp
pqqq
qpqp
pqpp
TRAJECTORY CALCULATION AND VIZUALIZATION
Equilibrium point
Talas1.exe

• Sravnit s nature population. Vibor coefficientov, chtobi poluchit nature chastotu. Odnoznachno li eto mozno cdelat- ved dva coefficient?
2 211 22
2 2 2 211 22 11 22
' ; '2 2
W p pq W q pqp q
W p pq W q W p pq W q
, 0 tif p q theninequilibria poin
11 22
11 22
11 22
11 22 22
22
11 22
; ;
;
(1 ) (1 )
( 2 ) 1
1
2
W W p q W W q p
W p q W q p
W p p W p p
p W W W
Wp
W W
0.2 1 0.80.879; 0.121
0.89 2 0.2 0.91p q

1 in 25 heterozygote alpha-thalassemia SE Asians,Chinese1 in 30 heterozygote beta-thalassemia Greeks, Italians
•B-thalassemia •1/20,000 in general population; ( )•1/100 in areas where malaria is endemic.
22 11
11 22 11 22
1 1; ;
2 2
W Wp q
W W W W
11 1?W
11
11 22
22 11
11/10 ;
2
9 8
Wq
W W
W W

Evolutionary operator with selection
2 2AA Aa aa AaW p W pq W q W pq
p ' ; q ' ;W W
2 2AA aa AaW W p W q 2W pq - mean fitness
Selection in case Thalassemia: WAA=0.89; WAa=1; Waa=0.2
Selection recessive lethal gene: WAA=1; WAa=1; Waa=0

2 2AA aa AaW W p W q 2W pq - mean fitness
2 2AA Aa aa AaW p W pq W q W pq
p ' ; q ' ;W W
WAA, WAa, Waa –individual fitnesses

Equilibria points
p=0, q=1 - population contains a allele only and on the zygote level the population consist of the homozygotes aa;
p=1,q=0 - population contains A allele only and on the zygote level the population consist of the homozygotes A A.
Homozygote equilibria states
2 2AA Aa aa AaW p W pq W q W pq
p ; q ;W W
2 2AA aa AaW W p W q 2W pq

Superdominance, when a heterozygote is fitter than both homozygotes
Superrecessivity , when a heterozygote is les fit than either homozygotes
In intermediate cases:
WAA Waa WAa (if WAA < WAa) or
WAa Waa WAA (if WAa < WAA)
The population has no polymorphic equilibria
aa Aa AA Aa
AA aa Aa AA aa Aa
W W W Wp ; q ;
W W 2W W W 2W
Heterozygote equilibrium states: p>0, q>0
AA aa Aamin(W , W ) W
AA aa Aamax(W , W ) W

Lethal allele
aa Aa AA Aa
AA aa Aa AA aa Aa
W W W Wp ; q ;
W W 2W W W 2W
Let WAA=0
AA aa Aamax(W , W ) W
AA aa Aamin(W , W ) W
If WAa > max(WAA,Waa) =Waa
Equilibrium point is polymorphic

Dominant selection
aa Aa AA Aa
AA aa Aa AA aa Aa
W W W Wp ; q ;
W W 2W W W 2W
AA aa Aamax(W , W ) W
AA aa Aamin(W , W ) W
Two different phenotypes
{AA, Aa}, {aa} WAA=WAa=1, Waa =1-s
No polymorphic equilibria point

Selection against a recessive allele.
WAA=WAa=1, Waa =1-s;
qWaa+pWAa=q(1-s)+p=1-sq; qWAa+pWAA=q+p=1;
2 2AA aa AaW W p W q 2W pq = p2+ q2+ 2pq- sq2 =1- sq2
When q is very small, a homozygotes are very rare.
When q is small, q2 is very small. So the recessive allele is hardly ever expressed.
The same logic applies to the case where the recessive allele is favored (1+s). The disfavored dominant can be eliminated easily even when scarce, but when the recessive is rare, even though it is favored it is very hard for selection to "see" it and build it up.

Example. Selection against recessive lethal gene

Fishers Fundamental Theorem of Natural Selection
Mean fitness increase along the trajectory

Convergence to equilibria2 2
AA aa AaW W p W q 2W pq
In intermediate cases:
Waa WAa WAA (or WAA WAa Waa)
The population has no polymorphic equilibria

Convergence to equilibria
Superdominance (overdominance), when a heterozygote is fitter than both homozygotes
Superrecessivity (underdominance), when a heterozygote is les fit than either homozygotes
AA aa Aamin(W , W ) W
AA aa Aamax(W , W ) W
2 2AA aa AaW W p W q 2W pq

One-locus multiallele autosomal systems

Fishers Fundamental Theorem of Natural Selection
2 211 1 12 1 2 nn nW W P W P P ... W P
Mean fitness
increase along the trajectory
Recommended