Embed Size (px)
Citation preview

Environmental Hypotheses of Hominin EvolutionRICHARD POTTSHuman Origins Program, National Museum of Natural History,Smithsonian Institution, Washington, DC 20560-0112
KEY WORDS climate; habitat; adaptation; variability selection;fossil humans; stone tools
ABSTRACT The study of human evolution has long sought to explainmajor adaptations and trends that led to the origin of Homo sapiens.Environmental scenarios have played a pivotal role in this endeavor. Theyrepresent statements or, more commonly, assumptions concerning the adap-tive context in which key hominin traits emerged. In many cases, however,these scenarios are based on very little if any data about the past settings inwhich early hominins lived. Several environmental hypotheses of humanevolution are presented in this paper. Explicit test expectations are laid out,and a preliminary assessment of the hypotheses is made by examining theenvironmental records of Olduvai, Turkana, Olorgesailie, Zhoukoudian, CombeGrenal, and other hominin localities. Habitat-specific hypotheses have pre-vailed in almost all previous accounts of human adaptive history. The rise ofAfrican dry savanna is often cited as the critical event behind the develop-ment of terrestrial bipedality, stone toolmaking, and encephalized brains,among other traits. This savanna hypothesis has been countered recently bythe woodland/forest hypothesis, which claims that Pliocene hominins hadevolved in and were primarily attracted to closed habitats. The ideas thathuman evolution was fostered by cold habitats in higher latitudes or byseasonal variations in tropical and temperate zones also have their propo-nents. An alternative view, the variability selection hypothesis, states thatlarge disparities in environmental conditions were responsible for importantepisodes of adaptive evolution. The resulting adaptations enhanced behav-ioral versatility and ultimately ecological diversity in the human lineage.Global environmental records for the late Cenozoic and specific records athominin sites show the following: 1) early human habitats were subject tolarge-scale remodeling over time; 2) the evidence for environmental instabil-ity does not support habitat-specific explanations of key adaptive changes;3) the range of environmental change over time was more extensive and thetempo far more prolonged than allowed by the seasonality hypothesis; and4) the variability selection hypothesis is strongly supported by the persistenceof hominins through long sequences of environmental remodeling and theorigin of important adaptations in periods of wide habitat diversity. Earlybipedality, stone transport, diversification of artifact contexts, encephaliza-tion, and enhanced cognitive and social functioning all may reflect adapta-tions to environmental novelty and highly varying selective contexts. YrbkPhys Anthropol 41:93–136, 1998. r 1998 Wiley-Liss, Inc.
TABLE OF CONTENTSA Paleoenvironmental Primer .............................................................................................. 95
Marine oxygen isotope analysis ........................................................................................ 96Ocean dust records ............................................................................................................ 98Ice cores .............................................................................................................................. 99
YEARBOOK OF PHYSICAL ANTHROPOLOGY 41:93–136 (1998)
r 1998 WILEY-LISS, INC.

Sedimentary environment ................................................................................................ 99Lake sediments .................................................................................................................. 99Loess sequences ................................................................................................................101Fossil pollen sequences .....................................................................................................101Stable isotope analysis of soils .........................................................................................101Faunal analysis ................................................................................................................104Summary ...........................................................................................................................104
A Framework for Analyzing Adaptation ..............................................................................104Environmental Hypotheses of Hominin Evolution .............................................................106
A concise history ...............................................................................................................106Challenges of the savanna ............................................................................................106Intrinsic and extrinsic approaches ...............................................................................106Turnover hypotheses .....................................................................................................108
Adaptive evolution in hominins .......................................................................................109Habitat-specific hypotheses ..........................................................................................109Short-term variability hypothesis ................................................................................111Long-term (variability selection) hypothesis ................................................................111
Test implications ...............................................................................................................112Habitat-specific hypotheses ..........................................................................................112Short-term variation (seasonality) hypothesis ............................................................113Variability selection hypothesis ....................................................................................113
Environmental Records at Early Human Localities ...........................................................1146.0 to 2.5 million years ago ...............................................................................................114
Eastern and southern Africa .........................................................................................114Discussion ......................................................................................................................116
2.3 to 1.7 million years ago ...............................................................................................118Turkana and Olduvai basins .........................................................................................118Discussion ......................................................................................................................119
1 million to 250,000 years ago ..........................................................................................122Olorgesailie basin ..........................................................................................................122Zhoukoudian, the Loess Plateau, and Bose basin, China ............................................122Discussion .....................................................................................................................125
140,000 to 50,000 years ago ..............................................................................................125Combe Grenal, France ..................................................................................................125Discussion .....................................................................................................................127
Environmental Perspectives on Adaptive Change ..............................................................128Conclusions ...........................................................................................................................130Acknowledgments .................................................................................................................131Literature Cited ...................................................................................................................132
Regarding the study of human evolution,it may be remarked that everybody talksabout the environment, but nobody doesmuch about it. The environment of humanevolution has been a source of speculationsince Darwin. Analyses of animal bones,pollen, and sediments have rendered a gen-eral portrait of Pliocene and Pleistocenehabitats. Yet field projects are rarely de-signed specifically to recover high-resolutiondata on the ecological settings inhabited byearly humans. Thus, there is a paucity offine-scale spatial and temporal data that
can be linked to in situ evidence of homi-nins.1 Usually, finding fossils and datingthem have overshadowed any effort to deter-
1Although it will take getting used to, the term hominin is nowpreferred in place of hominid to indicate the group of bipedal apesto which humans belong (genera Homo, Australophitecus, Paran-thropus, and Ardipithicus) (Begun, 1992; Andrews, 1992; An-drews et al., 1996, Delson and Tattersall, 1997). The systematicsof apes, including humans, now recognizes the growing body ofevidence that humans and chimpanzees are sister taxa. Withinthe superfamily Hominoidea (apes) is the family of great apes(Hominidae). The hominid family includes the subfamily ofAfrican apes (Homininae), or hominines. The tribe rank ofHominini, then, is reserved for humans and related bipeds sincethe split from the chimpanzee lineage. Because early Pliocenebipeds no longer comprise a valid subfamily, I use the informalterm australopiths to refer to them.
94 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

mine habitat contexts. A related problem isthat most projects still focus on surfacecollecting, with unavoidably poorer tapho-nomic and contextual control than obtain-able by digging. It is, however, only withexact stratigraphic relating of environmen-tal data, fossils, and artifacts that answersmay emerge about the range of habitatsassociated with particular species, when keyexpansions of habitat use occurred, or othersubstantive questions related to homininadaptive change.
As fossil discoveries have sparked morethan a century of ideas on the phylogenetichistory of hominins, so research on the con-texts of fossils and artifacts may come tostimulate a long phase of careful work on theecological history of hominins. The immedi-ate goal is to make this endeavor as system-atic, empirically based, and comparative aspossible. Clear definition of hypotheses andtheir test implications can help serve thisgoal.
The first purpose of this paper is to definepivotal hypotheses about the context of earlyhuman evolution and to lay out their testimplications. Second, I will present some ofthe best-known environmental sequencesfrom hominin sites as a preliminary series oftests. Environmental hypotheses can be di-vided into two types: those concerning adap-tation and others concerning species turn-over. Although the latter will be discussedbriefly, our focus is on environmental hypoth-eses as they relate to adaptive evolution.This topic is of marked interest becauseadaptive explanations relate directly to theprocesses of natural selection and thereforehelp to discern how important evolutionarytransformations took place.
Historically, the search for a convincingnarrative of human evolution has stimu-lated strong interest—and guesswork—about the adaptive history of hominins. Thus,essentially every account of hominin evolu-tion describes or assumes something aboutthe milieu of human ancestors and the envi-ronmental problems that uniquely humanfeatures supposedly solved or overcame. Ourfocus here on environmental hypotheses withan adaptive flavor requires a framework foranalyzing adaptation. So a third goal of thispaper is to outline such a framework, whichwill show how paleoenvironmental studies
may contribute decisive data to the study ofhominin evolution.
A final aim is to offer a brief primer to helpanthropologists comprehend the barrage ofpaleoenvironmental methods, data, and in-terpretations published almost weekly injournals such as Nature, Science, Quater-nary Research, and other scientific periodi-cals. A basic familiarity with isotope data,deep-sea cores, pollen analysis, loess climatecurves, orbital parameters, and the mainsources of paleoenvironmental evidence andhabitat change is essential for understand-ing the contexts in which hominins evolved.To the extent that context is important—andDarwinian and later evolutionary biologyestablished its fundamental role in organis-mal evolution—the field of paleoanthropol-ogy is, in this view, on the brink of novelideas and data sets concerning the how andwhy of hominin evolution. Tremendous ad-vances in the environmental sciences areforcing these developments to occur.
A PALEOENVIRONMENTAL PRIMER
An intricate system of heat and moisturecirculation determines Earth’s climate andhas strong influence over the distribution ofenvironmental zones and gradients. Thisenormous heating and plumbing systemlinks the atmosphere, oceans, and land. It isaffected by orbital cycles that determine theplanet’s variable dose of solar radiation.Volcanic activity and uplift of the crust alsoimpact it. The basic processes and factorsthat therefore contribute to this circulationsystem are
● the amount and yearly pattern of solarradiation, determined largely by the plan-et’s distance, orbital shape, and axial tiltrelative to the sun;
● the amount of solar radiation absorbedand reflected by the atmosphere and bythe ground surface;
● atmospheric processes, including evapora-tion, cloud formation, and rainfall;
● ocean and wind currents, two related pat-terns of circulation that redistribute heatand moisture in a biased manner aroundthe globe;
● the size and distribution of ice caps, landmasses, and major bodies of water, all of
95ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

which create complex feedback with atmo-spheric processes;
● land uplift, which may interrupt or deflectocean and atmospheric circulation cur-rents and may create rain shadows thatbias the distribution of rainfall near high-lands; and
● volcanism, which alters climate by theeruption of airborne particles, and re-gional landscapes by the deposition oflava or tephra (e.g., ash, pumice).
A series of methods has been developed tomeasure the interactions and effects of theseprocesses on Earth’s environments. All ofthese methods are applicable to the period ofhuman evolution; indeed, some of their mostfruitful applications have been in the con-text of paleoanthropological research.
Marine oxygen isotope analysis
Interaction between the ocean and atmo-sphere lies at the heart of oxygen stableisotope analysis, a technique that has revolu-tionized paleoclimate studies since the 1950s.During evaporation, the stable isotope 18O isenriched in ocean water as the lighter 16O ispreferentially released into the atmosphere.2In building their carbonate skeletons, cer-tain marine microorganisms, such as fora-minifera, absorb oxygen isotopes in propor-tion to the surrounding seawater. An 18O-enriched ocean will thus result in a higherratio of 18O to 16O in the microscopic shells.This transfer from ocean to foraminifera isalso affected, though, by temperature. Forevery 1°C drop in water temperature, theproportion of 18O to 16O within the foramshells is increased by about 0.22 parts permil (‰). Thus the oxygen isotope ratio—d18O (delta 18-O)—in foraminifera is a mea-sure, or proxy, of both temperature andevaporation.3
During glacial periods, oxygen (biased to-ward 16O) that has evaporated from the seabecomes locked up in ice sheets as precipita-tion falls on land. Thus, foram d18O fur-
nishes a record of glacial ice volume (whichcontains the sequestered 16O) and tempera-ture.4 Planktonic species of forams give infor-mation about sea-surface conditions, whilebenthic forams provide data about the deep-ocean environment, which is considered toreflect a wider record of ice (evaporation)and temperature history. This history isreconstructed by measuring d18O in foramskeletons found in cores drilled from thedeep sea. d18O curves are often thought toprovide a measure of global climate, but thisidea only refers to ocean temperature andice volume, a history that may be decoupledfrom environments on land (e.g., deMenocalet al., 1993).
Figure 1 depicts the average curve forbenthic forams from 70 Ma to the present.According to this curve, substantial globalcooling of oceans took place during thisperiod, with especially marked ice-sheet for-mation over the past 3 myr or so. By lookingat specific ocean cores, however, one seesthat this average cooling/drying trend isactually composed of many oscillations, theamplitude of which has risen over the past 6myr (Fig. 2A). Sets of oscillations occurredwithin warmer and colder periods, forming asequence of oxygen isotope stages (odd-numbered stages represent warm times;even refer to cold). Isotope stages 1–100cover the past 2.5 myr. As depicted in Figure2A, oscillation was particularly extreme overthe past 700 ka. In this period, single excur-sions of the d18O curve (about 100 kyr long)encompassed 1.5‰ or more, which exceededthe total average change since the lateMiocene (,1.4‰). Foram records back to thelate Oligocene (see Fig. 2B) indicate that thetotal range of d18O variation within 1 myrspans was fairly stable up through theMiocene. The total variation in each spanconsistently ranged between 0.3 and 0.8‰up until 5–6 million years ago. At that time,the amount of variation increased signifi-cantly and continued to rise to the present(Potts, in press a). As we will see, differenthypotheses about hominin evolution are sup-
2This process leading to isotope enrichment is called fraction-ation, in which the ratio of stable isotopes is altered during thetransfer of an element (in this case, oxygen) from one medium toanother.
3d18O measurements reflect the relative concentration of thetwo isotopes and are expressed as ‰ variation from a standardisotopic reference, measured on a belemnite from the Pee DeeFormation (Cretaceous age), South Carolina.
4For periods in Earth’s history when ice sheets were notpresent, the isotope ratio is primarily a measure of temperature.
96 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998
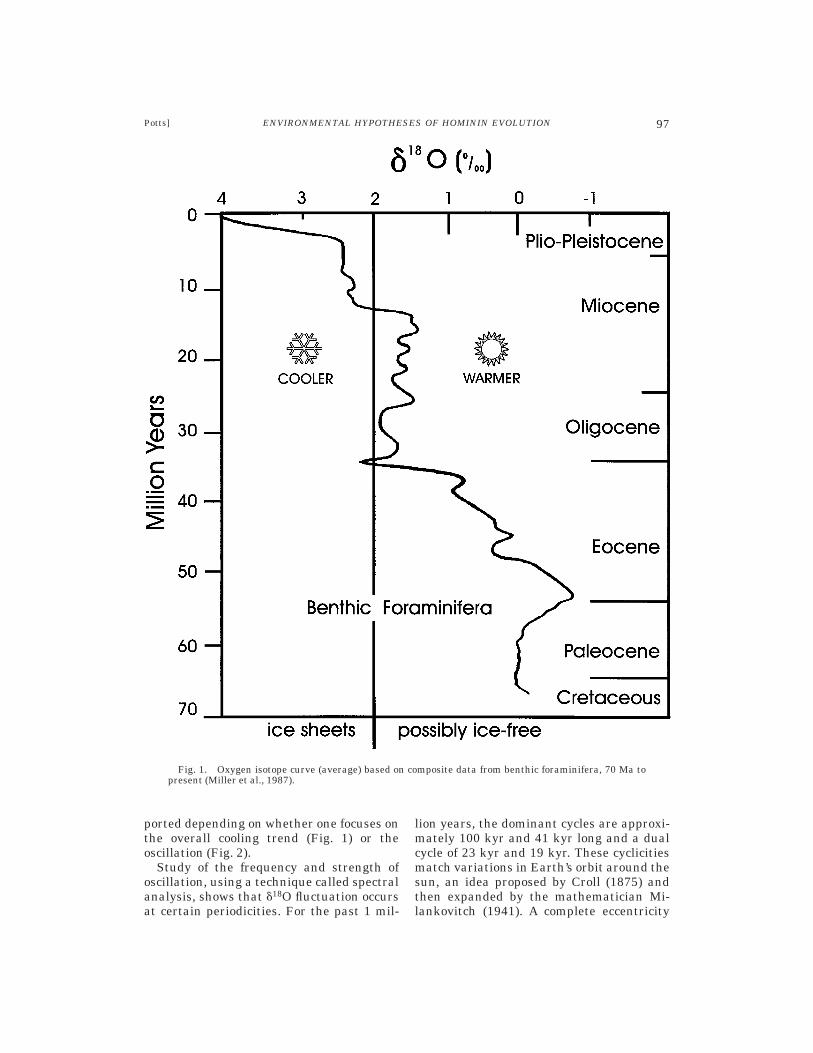
ported depending on whether one focuses onthe overall cooling trend (Fig. 1) or theoscillation (Fig. 2).
Study of the frequency and strength ofoscillation, using a technique called spectralanalysis, shows that d18O fluctuation occursat certain periodicities. For the past 1 mil-
lion years, the dominant cycles are approxi-mately 100 kyr and 41 kyr long and a dualcycle of 23 kyr and 19 kyr. These cyclicitiesmatch variations in Earth’s orbit around thesun, an idea proposed by Croll (1875) andthen expanded by the mathematician Mi-lankovitch (1941). A complete eccentricity
Fig. 1. Oxygen isotope curve (average) based on composite data from benthic foraminifera, 70 Ma topresent (Miller et al., 1987).
97ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]
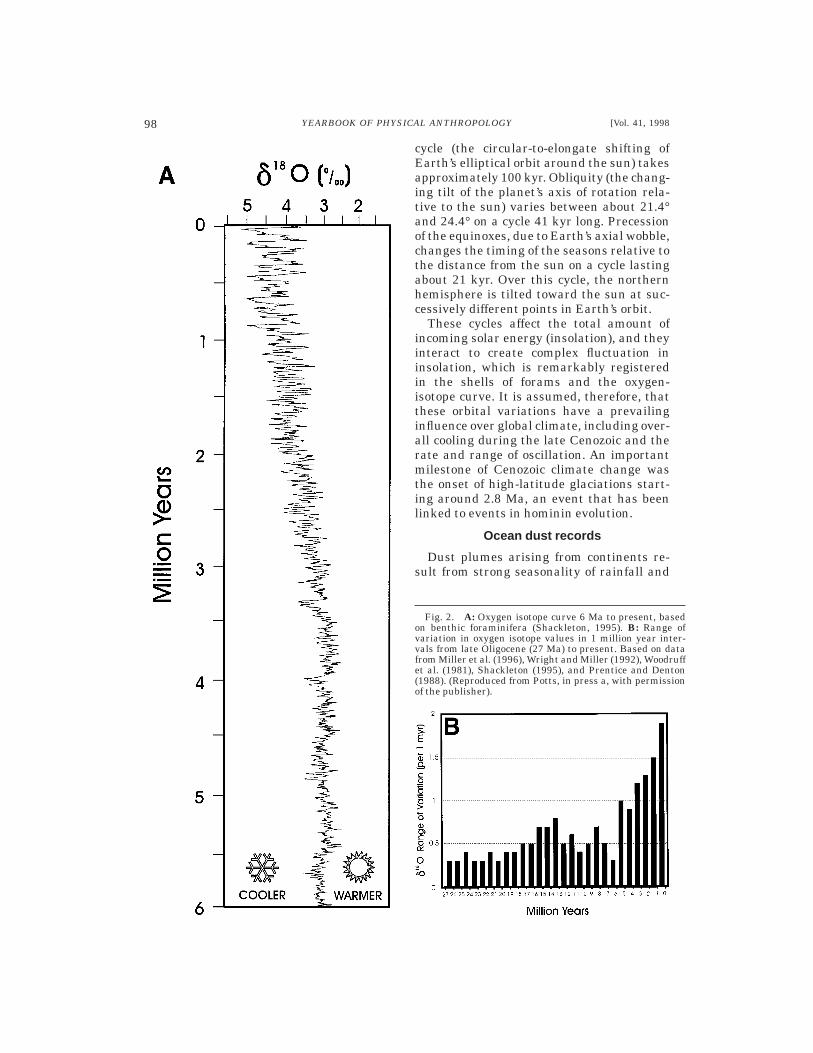
cycle (the circular-to-elongate shifting ofEarth’s elliptical orbit around the sun) takesapproximately 100 kyr. Obliquity (the chang-ing tilt of the planet’s axis of rotation rela-tive to the sun) varies between about 21.4°and 24.4° on a cycle 41 kyr long. Precessionof the equinoxes, due to Earth’s axial wobble,changes the timing of the seasons relative tothe distance from the sun on a cycle lastingabout 21 kyr. Over this cycle, the northernhemisphere is tilted toward the sun at suc-cessively different points in Earth’s orbit.
These cycles affect the total amount ofincoming solar energy (insolation), and theyinteract to create complex fluctuation ininsolation, which is remarkably registeredin the shells of forams and the oxygen-isotope curve. It is assumed, therefore, thatthese orbital variations have a prevailinginfluence over global climate, including over-all cooling during the late Cenozoic and therate and range of oscillation. An importantmilestone of Cenozoic climate change wasthe onset of high-latitude glaciations start-ing around 2.8 Ma, an event that has beenlinked to events in hominin evolution.
Ocean dust records
Dust plumes arising from continents re-sult from strong seasonality of rainfall and
Fig. 2. A: Oxygen isotope curve 6 Ma to present, basedon benthic foraminifera (Shackleton, 1995). B: Range ofvariation in oxygen isotope values in 1 million year inter-vals from late Oligocene (27 Ma) to present. Based on datafrom Miller et al. (1996), Wright and Miller (1992), Woodruffet al. (1981), Shackleton (1995), and Prentice and Denton(1988). (Reproduced from Potts, in press a, with permissionof the publisher).
98 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

in the winds that carry the dust to the ocean.Aeolian dust accumulations found in oceansediment cores thus provide a record oflong-term change in seasonality and prevail-ing wind patterns. These records may reflectthe history of continental vegetation cover,since large areas of open vegetation anddesert enhance the chances of erosion andwind transport of land detritus.
In a detailed study of cores off the westand east coasts of Africa, deMenocal (1995;deMenocal and Bloemendal, 1995) has shownthat African climate (and vegetation cover)was highly variable from early Pliocene tothe present (Fig. 3). Spectral analysis (rightside of Fig. 3) indicates a change in oscilla-tion at around 2.8 Ma, from a dominantfrequency of 23 to 19 kyr (precession) to oneof 41 kyr (obliquity). Other shifts occur at1.7 and 1.0 Ma, at which point the 100 kyrfrequency begins to dominate. These find-ings suggest that African climate variationwas tuned to orbit-related changes in insola-tion. These shifts represent periods of mark-edly increased aridity at 2.8, 1.7, and 1.0Ma; the origin of certain hominin species inAfrica after 2.8 Ma, therefore, may reflectthe rise of an arid-adapted biota (deMeno-cal, 1995). However, as noted by deMenocaland Bloemendal (1995), these dates primar-ily signal substantial rises in the overallvariance, or amplitude of fluctuation, ratherthan a permanent shift toward a drier, moreopen environment (Fig. 3).
Ice cores
Accumulation of ice in glaciers, ice sheets,and ice shelves results from precipitation ofwater evaporated largely from the ocean.Thus, d18O analysis of cores drilled frommajor ice bodies yields a high-resolutionrecord of climate, which is the inverse ofdeep-sea d18O. Particularly informative re-cords into the middle Pleistocene have beenobtained from Greenland and Antarctica.One significant finding from the Greenlandcores is evidence of extremely rapid oscilla-tions during certain time intervals. Knownas Dansgaard-Oeschger events, these oscilla-tions last around 1,000 years; during theseevents, fluctuations between interglacialwarmth and severe cold take place over acentury or even a decade in some cases
(Broecker and Denton, 1990; Taylor et al.,1993; Dansgaard et al., 1993). Based onthese findings, it has been argued (e.g.,Calvin, 1996) that hominin adaptabilityevolved in response to large, short-termfluctuations in middle or high latitudes.
Evidence of such abrupt variation, how-ever, is absent in the Antarctic ice cores(Jouzel et al., 1993), suggesting that theoscillation events may be largely a northernhemisphere phenomenon. A partial explana-tion is that rapid cooling may correspondwith periods of iceberg discharges (or raft-ing) into the North Atlantic (known as Hein-rich events); by adding fresh water to thesea, these discharges may suddenly reducethe heat-conveying effect of one of the mostimportant ocean circulation systems, theNorth Atlantic Deep Water, thereby causingabrupt cooling of 5–8°C (Bond et al., 1992;Stauffer et al., 1998). Deep-sea data fromthe North Atlantic show that such abruptcooling-warming cycles occurred regularlyback to at least 1 Ma (Raymo et al., 1998).
Sedimentary environment
Basic geologic analysis at hominin sitesincludes field recording and mapping of sedi-ments, describing major depositional envi-ronments (lithofacies), and logging strati-graphic change. Certain types of deposits,such as caliches, and the presence of particu-lar minerals are indicative of specific envi-ronmental conditions (e.g., aridity, rainfall).Other deposits, such as widely distributedtephra, permit ready correlation betweendistant locations. Correlation of stratigraphicsections over several kilometers enables thereconstruction of paleogeographic settingson a basinwide scale. Such reconstructionshave proved powerful in interpreting theadaptive contexts of hominins at Olduvai,Turkana, and other localities (Hay, 1976a;Brown and Feibel, 1991).
Lake sediments
Lakes may provide habitats for diatoms orother environmentally sensitive organisms(e.g., ostracods). Analysis of these organismscan yield a precise history of lake depth,salinity, and alkalinity—factors that arestrongly influenced by climatic and tectonicvariations. Study of diatom species in the
99ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]
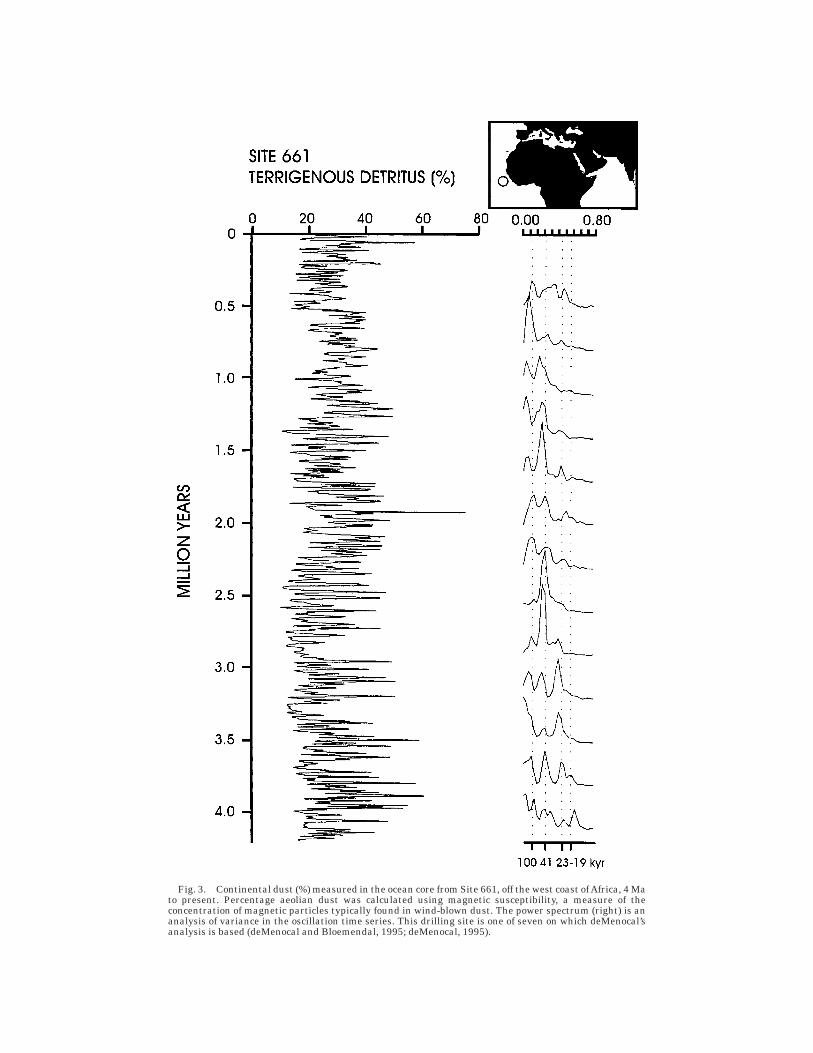
Fig. 3. Continental dust (%) measured in the ocean core from Site 661, off the west coast of Africa, 4 Mato present. Percentage aeolian dust was calculated using magnetic susceptibility, a measure of theconcentration of magnetic particles typically found in wind-blown dust. The power spectrum (right) is ananalysis of variance in the oscillation time series. This drilling site is one of seven on which deMenocal’sanalysis is based (deMenocal and Bloemendal, 1995; deMenocal, 1995).

Olorgesailie Formation, for example, hashelped to establish the history of an ancientlake basin inhabited by hominins (Owenand Renaut, 1981; Potts, 1994) and to cali-brate rates of environmental change facedby the toolmakers (see below).
Loess sequences
Thick sequences of alternating loess (wind-blown sand) and soil (paleosols) occurthroughout vast regions of Asia and Europe.Loess layers represent times when vegeta-tion cover was sufficiently low to allowbuildup of aeolian material. The soils, on theother hand, are often organic-rich due tothick vegetation cover at the time of soilformation. Loess sequences of north-centralChina have been especially well studied andprovide a detailed record of environmentalchange. Over the past 2.5 myr, at least 44major shifts from cold to warm conditionshave been documented.
Environmental interpretations are basedon the degree of loess and soil weathering,fossil pollen and fauna, carbonate content,and magnetic susceptibility. The lattermethod measures the concentration of mag-netic grains, which is especially high in finedust blown from distant sources. This finecomponent dominates during warm, wet pe-riods, while relatively coarse grains fromlocal sources tend to be deposited duringcold, dry eras. Thus, high susceptibility mea-surements are found in the soils and lowones in the loess layers. These measure-ments offer a record of oscillation in climateand vegetation. Figure 4 illustrates a sec-tion of the Luochuan loess sequence, inwhich strong fluctuations between cold, opensteppe and warmer, wooded to forested con-ditions have been documented (Liu, 1988;Kukla and An, 1989). Because they coverhuge continental areas, loess-soil depositsoffer a potentially powerful method of deter-mining environmental sequences in temper-ate latitudes occupied by early hominins.
Fossil pollen sequences
Pollen is deposited as ‘‘pollen rain,’’samples of which can be characterized bythe percentages of different vegetation com-ponents, known as a pollen spectrum. Thebasic components are usually arboreal (trees
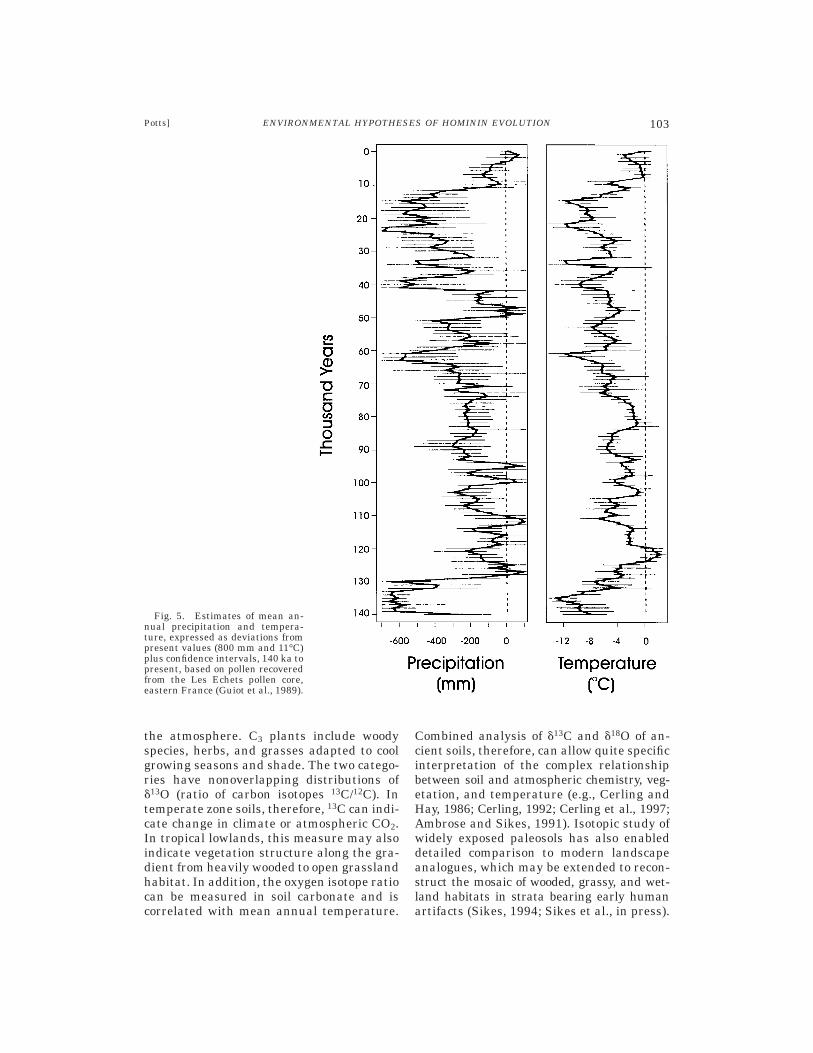
and shrubs) and nonarboreal (herbs) pollen(abbreviated AP and NAP, respectively). Per-centage AP reflects the density of tree cover,though this provides only a rough measurein low latitudes since tropical trees havelower pollen production than grasses andother NAP. Divisions of NAP often includegrasses (Gramineae), aquatics (such as Ty-pha and Cyperaceae), and the C/A group(Chenopodiaceae/Amaranthaceae). The firstindicates the relative density of grass cover,especially when aquatic taxa are factoredout. The last group can be a good indicator ofaridity in African settings, since most of theC/A plants come from saline soils and drystreams. Environmental interpretation ofpollen spectra is based on modern analoguesand thus requires careful comparison of pastand present samples and preservation condi-tions. The lack of accurate analogues forpast climates, particularly glacial condi-tions, can sometimes be offset by incorporat-ing other evidence (e.g., in Europe, the repre-sentation of fossil beetles, which areclimatically sensitive). The sum of certainAP taxa is often used as a climate index,although the validity of this approach hasbeen questioned. In East Africa, fossil pollenanalysis has yielded a sequence of generalvegetation and climate reconstructions forlocalities such as Olduvai, Turkana, andHadar; while in Europe the coring of peatbogs has enabled high-resolution analysis ofprecipitation and temperature based on pol-len (Bonnefille, 1995; Guiot et al., 1989,1993; Woillard, 1978). Figure 5 illustrates adetailed series of climatic fluctuations indi-cated by pollen spectra from the sedimentcore at Les Echets, France.
Stable isotope analysis of soils
Under certain conditions, buried soils, orpaleosols, preserve organic residues and car-bonate deposits that bear a stable-isotopicsignal of past vegetation. Interpretation ofvegetation is based on the photosyntheticchemistry of plants and the signature itleaves in soils. Most plant species undergoone of two distinct photosynthetic pathways;one involves three carbon atoms and theother four (thus C3 and C4 plants). C4 plantsare virtually all grasses adapted to hot, dryconditions and low CO2 concentrations in
101ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]
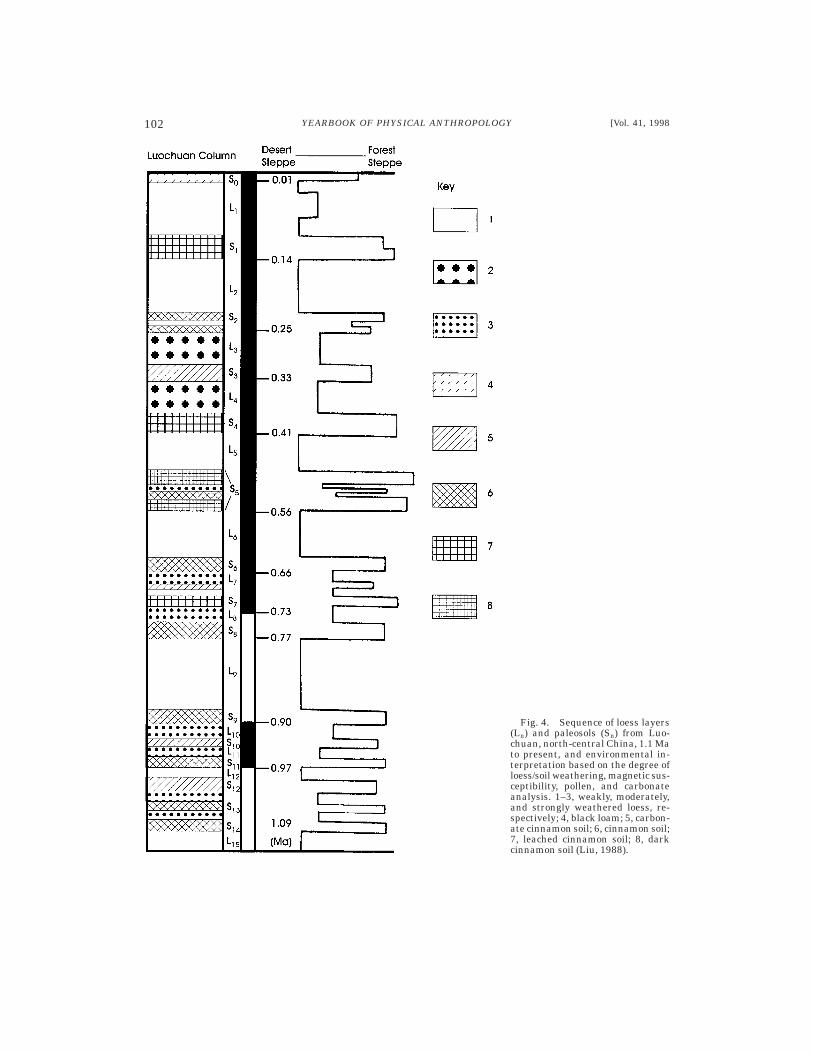
Fig. 4. Sequence of loess layers(Ln) and paleosols (Sn) from Luo-chuan, north-central China, 1.1 Mato present, and environmental in-terpretation based on the degree ofloess/soil weathering, magnetic sus-ceptibility, pollen, and carbonateanalysis. 1–3, weakly, moderately,and strongly weathered loess, re-spectively; 4, black loam; 5, carbon-ate cinnamon soil; 6, cinnamon soil;7, leached cinnamon soil; 8, darkcinnamon soil (Liu, 1988).
102 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998
the atmosphere. C3 plants include woodyspecies, herbs, and grasses adapted to coolgrowing seasons and shade. The two catego-ries have nonoverlapping distributions ofd13O (ratio of carbon isotopes 13C/12C). Intemperate zone soils, therefore, 13C can indi-cate change in climate or atmospheric CO2.In tropical lowlands, this measure may alsoindicate vegetation structure along the gra-dient from heavily wooded to open grasslandhabitat. In addition, the oxygen isotope ratiocan be measured in soil carbonate and iscorrelated with mean annual temperature.
Combined analysis of d13C and d18O of an-cient soils, therefore, can allow quite specificinterpretation of the complex relationshipbetween soil and atmospheric chemistry, veg-etation, and temperature (e.g., Cerling andHay, 1986; Cerling, 1992; Cerling et al., 1997;Ambrose and Sikes, 1991). Isotopic study ofwidely exposed paleosols has also enableddetailed comparison to modern landscapeanalogues, which may be extended to recon-struct the mosaic of wooded, grassy, and wet-land habitats in strata bearing early humanartifacts (Sikes, 1994; Sikes et al., in press).
Fig. 5. Estimates of mean an-nual precipitation and tempera-ture, expressed as deviations frompresent values (800 mm and 11°C)plus confidence intervals, 140 ka topresent, based on pollen recoveredfrom the Les Echets pollen core,eastern France (Guiot et al., 1989).
103ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

Faunal analysis
Relative abundance of different animalspecies permits a general characterizationof habitat, assuming that certain taxonomicgroups correlate with specific habitats inboth the past and present (e.g., Vrba, 1975;Potts, 1988; Shipman and Harris, 1988). Adifferent array of methods uses functionalmorphology to test for definitive anatomicalindications of environment. This approachallows an ecomorphic (as opposed to a taxo-nomic) characterization of the biota andassociated habitats without assuming par-ticular taxon-habitat correlations. Methodsof ecomorphic analysis have increasinglybeen applied to the faunas of early homininsites (e.g., Andrews, 1989; Plummer andBishop, 1994; Kappelman et al., 1997; Reed,1997).
Summary
Development of these and other methodsof environmental interpretation has pro-gressed impressively over the past severaldecades. The contexts in which these meth-ods can be applied include deep-sea cores,peat bogs, soils, and many diverse types ofsediment. The methods can in many cases gobeyond a general history of climate andhabitat to offer a quite detailed picture of theadaptive contexts faced by early hominins.
A FRAMEWORK FOR ANALYZINGADAPTATION
The process of adaptation, natural selec-tion, involves the relationship between anorganism and its surroundings. An organ-ism’s environment (at least aspects thataffect survival and reproduction) influencesthe traits that persist, increase, and spreadthrough a population. Traits positively af-fected by the process of biased reproductionare called adaptations. The analysis of adap-tations thus may afford insights into boththe process of natural selection and theenvironmental context in which evolutionhas occurred. The purpose of this section isto define an analytical framework that showshow paleoenvironmental data from homininsites may contribute to the study of adaptiveevolution in early humans.
The field of biology is divided between twomain approaches to the concept of adapta-
tion (Amundson, 1996; Endler, 1986). Thefirst is a nonhistorical approach, which de-fines an adaptation solely in terms of thecurrent, observed benefits that a particulartrait offers to an organism (Bock, 1980). Theprocess of origin (natural selection) and thusthe history of a given trait are, by thisdefinition, less relevant to deciding whethera trait is an adaptation. A nonhistoricaldefinition of adaptation, then, concerns sim-ply the contribution of a trait to currentfitness (Fisher, 1985).
The second is a historical approach, whichfocuses not only on fitness but also on theprocess by which adaptation occurred. Ac-cordingly, an adaptation is a trait generatedby natural selection due to its fitness ben-efits. This approach focuses, then, on thehistory of a trait. It allows the possibility ofstudying and citing the selective causes thatunderlie a particular adaptation. With homi-nin evolution, we are very much interestedin history. Thus, the second approach isadopted here. For one thing, current benefit(the basis for defining an adaptation in thenonhistorical approach) has no meaningwhen looking at characters of extinct homi-nins. In an evolutionary perspective, more-over, current benefit even in living humansdoes not always give a true picture of atrait’s original advantage or the conditionsunder which it evolved. In a historical analy-sis, then, adaptation is the result of naturalselection, and the analysis of adaptationentails an assessment of how the prevalenceof a trait in the fossil record may haveresulted from this process.
But how does one recognize natural selec-tion, or at least its results, in the fossilrecord? The first order of business is todefine a trait, feature, phenotype, or charac-ter (all considered synonyms here) or even acomplex of traits or a definable morphologi-cal or behavioral pattern. The definition of afeature or overall phenotype provides thebasis for examining function, benefit, andevolutionary setting.
In a historical approach, an analysis ofadaptation must also be based on the pro-cess of natural selection itself. As summa-rized by Lewontin (1970), the process hasthree necessary and sufficient conditions:variation, heritability, and differential fit-
104 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

ness. For selection to operate, the units ofevolutionary change (e.g., specific pheno-typic features) must vary. The variationsmust be inherited (e.g., the phenotypes mustrelate to specific genetic variations). And thevariations must differ in their representa-tion in later generations.
These three conditions furnish an initialframework, but many other factors enterinto an analysis of adaptation. Differentialfitness occurs presumably due to certainbenefits that one variant offers compared toothers (alternative phenotypes). So it is es-sential to determine what those benefitswere (or are), especially how a trait’s struc-tural design or pattern of expression pro-vided a selective advantage. Each beneficialfeature may also have its share of costs,which cannot outweigh the benefits if afeature is to increase via natural selection.The adaptive evolution of a feature is oftenassumed to take place in a competitive set-ting, and its increase may have seriousrepercussions for other individuals withinthat setting, which could affect the increaseand spread of the feature.
Furthermore, benefits, costs, and competi-tion all take place in relation to particularenvironments. This is one reason the analy-sis of adaptive evolution involves environ-mental reconstruction. The stability or dy-namics of environmental change may alsohave an impact on natural selection. Thedifferent time scales of environmental fluc-tuation may affect natural selection and, aswe will see, may determine the kind of traitsthat are ultimately represented in a popula-tion.
Finally, a growing number of evolutionarybiologists believe that lineages themselvespossess adaptations, referring to character-istics (e.g., geographic range, habitat vari-ety, dietary versatility) that go beyond themakeup of an individual organism (Vrba,1989; Jablonski, 1987). Since these charac-teristics may impact the survival of popula-tions over time, they may also affect thesuccess of particular features originated byconventional, individual-level natural selec-tion.
A basic framework for the analysis ofadaptation, then, can be conveyed by a se-ries of questions (Table 1) that take into
account phenotype definition, the operationof natural selection, and the factors thataffect differential fitness.
The analysis of adaptation is tricky largelybecause the questions posed in Table 1 aredifficult to answer and any answers putforth are usually controversial. As debate
TABLE 1. A 13 part framework for the analysisof adaptation
1. How is a particular feature (morphological struc-ture, behavior, phenotypic system or complex)manifested? What is its structure, or what are itscharacteristics?
2. What is the function of a particular feature?What is it designed to do? What selective advan-tages does the feature offer to an organism?
3. What ‘‘currencies’’ of advantage or success doesthe feature provide to the organism? An advan-tage in survivorship? A direct reproductiveadvantage? Does the feature raise the organism’sefficiency or economy (Steudel, 1994)? Ulti-mately, how does the feature contribute to thegenetic fitness of the individual?
4. Besides its beneficial function, does the featurehave any costs to the organism? What are thosecosts?
5. What is known about the feature’s heritability?6. What is known about the feature’s development
(growth and expression during the life cycle)?What are the necessary environmental param-eters for proper development?
7. Does the feature (e.g., a particular behavior)have any repercussions (costs or benefits) forother organisms? In other words, what was thecompetitive and/or social context in which thefeature evolved or currently functions?
8. In the context of natural selection, when did thefeature originate? In what environmental con-text or conditions did it evolve? What advantagesdid the feature offer within that original context?Did the feature represent a response to a specificenvironment or to diverse surroundings? Do theoriginal advantages and environmental contextstill apply to the organism that possesses thefeature?
9. What were the options or alternative phenotypes(e.g., the ancestral condition) available to theorganism when the specific feature evolved?
10. Historically, is the feature a by-product of adap-tive change in another trait (in which case it isnot an adaptation), or did the feature evolve dueto specific benefits of a task it performs or con-tributes to?
11. Does the feature serve its original function, or isit an exaptation (co-opted for some other function[Gould and Vrba, 1982])?
12. Over what time scale does the feature assist theorganism? Daily? Seasonally? Does it offer resil-ience to environmental variations faced over alifetime? Does it offer flexibility to a populationover longer periods of environmental change?
13. Are there lineage-level adaptations—that is, fea-tures of a lineage (rather than of an individual)that enhance its ability to spread, persist, ordiversify when compared with other, related lin-eages? If natural selection can take place at thespecies level, what kinds of features might beinvolved in this process?
105ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

about the evolution of the human chin dem-onstrates (Gould, 1977), even the first step—defining a trait subject to natural selection(Table 1, questions 1–3)—can create contro-versy, partly because traits can emerge as aby-product of adaptive modification (ques-tion 10). Besides these correlated effects,developmental (e.g., embryological) con-straints are ubiquitous, which place bound-aries on trait variation, the raw materialacted on by natural selection, and thus onadaptation itself. Structural design, the ben-efits, and the costs of morphological traitscan be considered with some success byfunctional anatomists. But specialists inter-ested in past behavior (e.g., australopithbipedality, late Pliocene stone flaking) usu-ally have greater difficulty evaluating de-sign properties, frequency of occurrence,heritability, and the advantage over alterna-tive behaviors. In fact, very little is knownabout the heritability of even simple struc-tures and behaviors unique to humans. Theheritability of characters (e.g., megadontia)that distinguish extinct hominins, more-over, is largely inaccessible except perhapsby analogy to living mammals that possesssimilar traits.
Given these difficulties, the study of earlyhominin adaptation is largely left with theanalysis of context (Table 1, question 8). Thequestions of time, place, and environmentalsetting are essential to all evolutionaryanalyses of adaptation. The selective historyof a trait is reflected in part by when, where,and the conditions or surroundings underwhich that trait first became apparent in thefossil record and began to characterize aparticular lineage or clade. The precise defi-nition of hypotheses relating past environ-ments to the emergence of unique hominintraits, therefore, helps to galvanize researchon early human evolution.
ENVIRONMENTAL HYPOTHESESOF HOMININ EVOLUTION
A concise historyChallenges of the savanna. Over muchof the twentieth century, scientists thoughtthat human evolution entailed a simple tra-jectory from apelike to humanlike and thatthis process was promoted by the challengesof an open savanna. In his book The Evolu-
tion of Man, Elliot Smith (1924:40) de-scribed the terrestrial domain of early hu-man ancestors as ‘‘the unknown worldbeyond the trees’’—a place of novel foodsources, quite distinct from the ‘‘land ofplenty’’ of the apes. Dart (1925) took thisidea to dramatic heights. To convey theselection pressures that drove human evolu-tion, Dart drew an analogy to the southernAfrican veldt. The early australopiths, heargued, confronted many dangerous preda-tors, a relative scarcity of water, and fiercecompetition for food, all of which he ulti-mately encapsulated in the killer-ape sce-nario (Dart, 1953). Later views, such as thedietary hypotheses of Robinson (1954,1961a,b) and Jolly (1970), also explicitlyrelated hominin origin to a shift from for-ested/wooded ‘‘apelike’’ habitat to an opensavanna setting.Intrinsic and extrinsic approaches. Ifindeed human evolution followed a progres-sive path from apelike to humanlike, it couldalso be reconstructed by extrapolating upfrom the anatomical, behavioral, and ecologi-cal characteristics of apes and other nonhu-man primates and extrapolating down fromethnological data on human foragers. Thepaper ‘‘Ecology and the Protohominids’’ byBartholomew and Birdsell (1953) made ex-plicit this approach to modeling human evo-lution. This paper was the first to drawattention to key theoretical aspects of earlyhuman ecology, including population den-sity and dynamics, territoriality, foraging,energetics, body size, and sociality. Yet noenvironment was reconstructed, and no ideaswere offered about the interaction of earlyhumans with specific environmental vari-ables. Implicit in their discussion was thatthe environment of hominin evolution wasalready generally known. It involved, atfirst, increasingly arid, savanna conditionsin Africa, followed by increasingly cool, gla-cial conditions in northern latitudes.
Despite their emphasis on ecology, Bartho-lomew and Birdsell (1953) provided an out-line for an intrinsic account of homininevolution whereby adaptive change resultedlargely from responses within populationsover time without regard to change in exter-nal conditions. All they required was thebasic scene, a relatively open savanna. Their
106 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

argument followed Darwin’s (1871) basicscenario: bipedality (ground living) and smallcanine teeth implied that the earliest homi-nins used tools, which permitted an ex-panded diet, including animal foods; thus,hunting in bands and changes in socialityoccurred as one adaptive change led to oth-ers. Washburn (1960) refined this type ofexplanation, and placed tools at the center ofan evolving system in which each noveladaptation fostered others in an intricateseries of feedback loops. ‘‘Tools, hunting,fire, complex social life, speech, the humanway and the brain evolved together to pro-duce ancient man of the genus Homo’’ (Wash-burn, 1960:63). Again, Washburn’s intrinsicfeedback model was not without context. Ifbipedal tool use initiated the cause-and-effect process of hominin evolution, life inthe open plains provided the crucial, initialsetting (Washburn, 1960).
Washburn’s influence and interest in in-trinsic accounts (as opposed to extrinsicaccounts, which interpret evolutionaryevents in relation to environmental change)may explain why many paleoanthropolo-gists in the 1950s to 1970s paid little atten-tion to environmental context. In two classicvolumes on hominin evolution (Washburn,1961, 1963), none of the 35 papers focusedprimarily on environmental settings. andonly seven considered the subject.5 Thisapproach was echoed in the influential fourvolume set Perspectives on Human Evolu-tion published between 1968 and 1978(Washburn and Jay, 1968; Washburn andDolhinow, 1972; Isaac and McCown, 1976;Washburn and McCown, 1978). The onlyarticles on paleoenvironmental context werein Volume 3 (six of its 21 chapters). Twodealt with Miocene faunas and floras (VanCouvering and Van Couvering, 1976; An-drews and Walker, 1976), two others consid-ered environmental evidence from specifichominin-bearing basins (Olduvai, Omo)(Hay, 1976b; Howell, 1976), and two chap-ters highlighted the general context of the
East African rift (Isaac, 1976; Bishop, 1976).It is interesting to note that Bishop’s paperdemolished the ‘‘myth of a Pliocene drought’’(p. 40) championed by Dart, which hadfueled the view that early human evolutionwas a response to a sharp shift from moistforest to arid savanna. Moreover, the papersconcerning Miocene sites showed that earlyapes lived in a complex mosaic of environ-ments, including closed and relatively openvegetation. These papers were among thefirst to hint at the intricate environmentalhistory related to ape and human evolution.Isaac (1976:133) ended his article by notingthe paucity of contextual studies and com-pared the existing information base aboutEast African paleoenvironments to an ‘‘ex-posed tip of an iceberg.’’
While most anthropologists took an intrin-sic approach, geologists and vertebrate pale-ontologists showed strong interest in paleo-environmental data and the reconstructionof early hominin settings. Thus, in Howelland Bourliere’s (1963) volume African Ecol-ogy and Human Evolution, almost all of the19 papers discussed the importance of con-textual data (the main exception was that byDeVore and Washburn). The papers (e.g., byButzer) included cautions about simplisticlinkages between higher latitude glacial se-quences and lower latitude environmentalhistory. Other papers surmised that com-plex tectonic and climatic events were inter-woven in creating the environmental historyof Africa (e.g., papers by Bishop and by deHeinzelin). Several papers showed evi-denced that Pleistocene Africa underwent acomplex series of habitat changes betweenmoist and dry and between forested andopen (e.g., papers by Moreau, Monod, Groveand Pullan, and de Heinzelin). Articles byBiberson and by Howell and Clark wereparticularly successful in placing Pleis-tocene stone technologies and behavior inspecific sedimentary, vegetational, and cli-matic contexts.
Later volumes by Bishop and Clark (1967),Butzer (1971), and Bishop (1978) continuedto emphasize the importance of environmen-tal (sedimentary, geomorphological, tapho-nomic, and ecological) settings for under-standing hominin evolution. More than halfof the articles published in Butzer and Isaac’s
5Four papers (by Chance, Oakley, Hallowell, and Sauer in the1961 volume) reiterated the general view that the earliestancestors inhabited the African savanna while later homininslived in an ice-age context. Three others (by Mayr, Dobzhansky,and Harrison and Weiner in the 1963 volume) discussed environ-ments as a theoretically important determinant of naturalselection, adaptive change, and speciation in hominins.
107ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

(1975) After the Australopithecines andnearly one-third of the contributions to Jol-ly’s (1978) Early Hominids of Africa focusedspecifically on the climatic, geologic, andpaleoecological contexts of hominin sites.
By the early 1980s, a wide range of envi-ronmental data was becoming available dueto a surge in paleoclimate studies and de-tailed work by field geologists at fossil andarcheological sites. Laporte and Zihlman(1983) made a strong case for the idea thatenvironment was a major driving force inearly human evolution. In their view, adap-tive change in apes, including Pliocene homi-nins, represented responses to either newenvironments or physical access to existingenvironments (e.g., via land bridge). Theyargued that global cooling reduced tree coverover large regions, and African uplift andrifting created a rain shadow effect (drying)in the eastern part of the continent. Thesefactors caused the spread of the savannamosaic, marked by strongly seasonal rain-fall, discontinuous distribution of trees, andheat-adapted, drought-resistant vegetation.Echoing earlier authors, Laporte and Zihl-man (1983:106–107) posited that the ‘‘chal-lenges of the savanna-mosaic’’ causedground-dwelling in a prehominin ape andbipedality in the earliest hominins. Two-legged walking enabled mobility and carry-ing of food and water over long distances asresources became more widely (patchily)distributed. Finally, they noted that the‘‘origin of hominids [sensu stricto] is not anisolated evolutionary event, but rather ispart of the overall radiation of African homi-noids; nor is it an unusual ecological occur-rence. The hominids are only one of severalmammalian groups that include the pigsand bovids that were able to exploit thesavanna-mosaic habitat that had becomewidespread by the end of the Miocene’’(Laporte and Zihlman, 1983:108).
Turnover hypotheses. At about the sametime, Vrba (1980, 1985, 1988; also Brain,1981) began to present an important idea,known as the turnover pulse hypothesis,which expanded these same themes—cli-matic forcing of evolution, a Pliocene aridityshift, and coincident change in homininsand other mammalian lineages. Turnover
pulse refers to a concentration of speciationand extinction events (i.e., turnover) in abrief period of time as a result of environmen-tal change. According to Vrba’s original hy-pothesis, many different African mamma-lian clades were involved in a turnoverepisode at 2.5 Ma. Synchronous change inmultiple groups was caused by a shift fromwarm, moist conditions to a cooler, drier, andmore open habitat. This environmental eventwas forced by global cooling, manifested as asharp transition (18O enrichment) in thedeep-sea isotope record. The hypothesis thusposits a strong causal link between homininevolution and global climatic events. Accord-ingly, the origins of Homo and of the robustaustralopiths at about 2.5 Ma were consid-ered part of the turnover pulse. Since aspeciation pulse in grazing bovids occurredat this same time (e.g., Vrba 1985, 1988),Homo and Paranthropus were also deemedpart of the Pliocene arid, open-grasslandbiota of Africa. Vrba et al. (1989) posited twoother turnover pulses, about 5 Ma and 0.9Ma, which also appeared to coincide withperiods of global cooling and continentaldrying. The origin of the Hominini, on theone hand and the extinction of robust austra-lopiths, coupled with the dispersal of H.erectus into Eurasia, on the other werethought to be correlated with these environ-mental shifts.
The original hypothesis has been re-vamped in two main ways. Since the mid-1980s, paleoclimatologists have broadenedthe age estimate of Pliocene global environ-mental change. Based on the deep-sea oxy-gen isotope record (Fig. 2A), for example, amajor shift in global ice sheets began asearly as 2.8–3.0 Ma, with the most signifi-cant change between 2.8 and 2.4 Ma (e.g.,Prentice and Denton, 1988). The deep-seadust record (see Fig. 3) also indicates amajor change in cyclicity beginning around2.8 Ma. Thus, the pulse in the late Plioceneturnover pulse hypothesis has been broad-ened; the target interval has been extendedfrom a point in time (2.5 Ma) to a span aswide as 400 kyr. According to this broaderpulse model, key events in hominin evolu-tion and concerted turnover in other mam-mals should be evident from 2.8 to 2.5 (or2.4) Ma rather than randomly distributed
108 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

throughout the entire 3.5 myr span of thePliocene (Prentice and Denton, 1988). InVrba’s (1995b) view, the turnover pulse inAfrica began 2.8 Ma but became apparent inthe fossil record only between 2.7 and 2.5Ma. This is the first way in which thehypothesis has been altered.
The second way involved refining the testimplications of the turnover pulse modeland broadening its theoretical basis. Build-ing on a synthetic view called habitat theory,Vrba (1992, 1995a) posits that the record of aturnover pulse should depend on the breadthof resources and habitats used by organ-isms. First and last appearances will besynchronized or spread out according to theecological tolerances of different lineages. Aglobal cooling event causing the spread ofsavanna between 2.8 to 2.5 Ma thus shouldhave caused turnover in specialized organ-isms first and in more generalized organ-isms later. Furthermore, it should cause alarger number of first appearances in arid-adapted taxa (e.g., hominins and grazingbovids) and more last appearances in warm,woodland-adapted species.
Vrba has been assiduous in testing theturnover pulse idea. Many appearances anddisappearances of bovid lineages in the Afri-can fossil record are clustered around 2.5Ma (Vrba, 1985, 1988, 1992, 1995b). Grazingbovids (alcelaphines such as wildebeest,hartebeest, and topi and antilopines such asgazelles) contributed strongly to the firstappearances. This supports the idea thataridity and grassland expansion were theenvironmental causes of bovid lineage turn-over. Vrba (1985, 1988) has also emphasizedWesselman’s (1984) conclusion (based on astudy of micromammals from Omo, Ethio-pia) that forest taxa prevalent at 3 Ma gaveway to xeric-adapted, open vegetation taxabetween 2.4 and 2.5 Ma. In the volumePaleoclimate and Evolution, with Emphasison Human Evolution (Vrba et al., 1995),papers concerning paleoclimate and vegeta-tion tend to confirm that large environmen-tal change occurred between 2.8 and 2.5 Ma.Many of these papers, however, stress theevidence for strong fluctuation at this timerather than a simple cooling or aridity shift.Conclusions drawn from African bovids, mi-cromammals, and hominins are said to sup-
port the turnover pulse idea (Vrba, 1995b;Wesselman, 1995), though an analysis ofsuids and hominins argues strongly againstthe hypothesis (White, 1995).
The pulse hypothesis has been questionedon other grounds, however, and counteredby what may be called the prolonged turn-over hypothesis (Behrensmeyer et al., 1997;McKee, 1996). Building on earlier observa-tions (Feibel et al., 1991), a recent study byBehrensmeyer et al. (1997) focused on theTurkana-Omo basin since this area offersthe richest and best calibrated record ofAfrican fossil mammals in the crucial spanbetween 3 and 1.8 Ma. Taking samplingvariation (rises and falls in fossil samplesover time) into account, this study found nostatistically significant pulse in either firstor last appearances at 2.5 Ma or between 2.8and 2.5 Ma. Instead, turnover in this inter-val was unremarkable, and mammalian lin-eages displayed a prolonged period of turn-over especially between 2.5 and 1.8 Ma.Moreover, a combination of xeric and mesicspecies was found to persist from 3 to 2 Ma.Behrensmeyer et al. (1997) argued that acomplex vegetation mosaic coupled with widefluctuation over time would explain both thepersistence of species of diverse ecologicaltolerances and the prolonged period of turn-over. Whether similar results might apply tothe whole of Africa has yet to be determined.
Adaptive evolution in hominins
Although the turnover pulse idea has at-tracted much attention, environmental hy-potheses pertaining to adaptive change re-main poorly developed. Hypotheses of thistype differ from turnover models. They at-tempt to describe how environmental con-text impinged on survival and natural selec-tion and in turn influenced the emergence ofkey adaptations. Since adaptive change maybe only loosely coupled to speciation (El-dredge, 1989), these hypotheses do not neces-sarily relate directly to the pattern of speciesturnover.
Habitat-specific hypotheses. The mostprominent narratives of hominin adaptiveevolution are habitat-specific. Each of thesescenarios points to a particular type of envi-ronment and sets forth the reason certain
109ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

adaptations arose to meet the special de-mands of that setting. Some scenarios sim-ply name the particular type of habitat inwhich hominins were assumed to live (e.g.,the savanna), while others may specify aparticular direction of environmental change(e.g., the onset of dry or cold conditions). Inboth cases, it is implicit that natural selec-tion had a consistent effect over time (i.e.,directional selection). This process served toimprove adaptive opportunities and favoredcertain innovations as ways of solving thechallenges posed by the special habitat ortrend.
The discovery of human fossils and arti-facts with ice-age fauna and strata, for ex-ample, led to the idea that advances inhuman evolution were forged in the harshsettings of Pleistocene Europe. This viewmay be termed the ice-age hypothesis. Anupdated version of this view is as follows:cool steppe and cold periglacial conditions inmid-latitude Eurasia posed a very challeng-ing environment to early human popula-tions, and advances in hominid social andcognitive abilities helped to solve these prob-lems of survival. Accordingly, the onset ofice-age habitats (due to global cooling) re-sulted in the evolution of key human adapta-tions.
As already noted, the savanna hypothesisis another habitat-specific standard and hasdominated thinking about the earliest phasesof hominin evolution. According to this idea,adaptation to drier and increasingly openenvironments was the hallmark of homininevolution from late Miocene through at leastthe early Pleistocene. Savanna was the con-text that incited the emergence of fundamen-tal human traits—terrestrial bipedality,larger brains, stone toolmaking, meat-eat-ing, and associated foraging behaviors suchas hunting. Essentially all textbooks prior to1996 explained the origin of these traits interms of adaptation to relatively open sa-vanna, defined by the presence of grass,discontinuous trees, overall aridity, and sea-sonal rainfall (e.g., Klein, 1989; Wolpoff,1980).
Recent support for the savanna hypoth-esis has risen from the ranks of paleoclima-tologists and paleontologists (Table 2). Theshared view is that emergence of dry, open
savanna was caused by steplike episodes ofglobal cooling starting in the late Miocene.Vrba (1995b:406), for example, refers to theonset of ‘‘the massive overall cooling trend’’of the late Pliocene. Intense global coolingled to pulses of aridification in Africa, whichcaused savanna expansion. The shift fromdense woodland to savanna had a strongeffect on adaptive characteristics, includingbipedality in early Australopithecus, hyper-trophy of the chewing apparatus in Paran-thropus, and various developments in Homo,such as encephalization and dependence ontoolmaking (Stanley, 1992; deMenocal, 1995;Prentice and Denton, 1988; Vrba et al., 1989;Vrba, 1988).
Although the savanna hypothesis hasgained recent support, it has also beenstrongly contested. Some authorities advo-cate a contradictory model—the woodland/forest hypothesis—which argues for the im-portance of closed vegetation in earlyhominin evolution. Early australopiths, ac-cording to some interpretations, were closelyassociated with wooded environments, exhib-ited significant arboreal activity, and shouldbe considered adapted to closed habitats(Clarke and Tobias, 1995; Berger and To-bias, 1996). Fossil pollen from Makapans-gat, for example, suggests that A. africanusinhabited conditions approximating a tropi-cal forest (Rayner et al., 1993). At Aramis,the oldest known hominin, Ardipithecusramidus, is associated with evidence of arelatively tree-dominated setting (WoldeGab-riel et al., 1994). Stable isotope studies,moreover, show that the shift to C4-domi-nated environments (open, grassy, heat-adapted vegetation) occurred later than pre-viously thought, as late as 1.7 Ma, with littleevidence of consistently open savanna untilafter 1.0 Ma (Cerling et al., 1991; Cerling,
TABLE 2. Recent advocates of the savanna hypothesis
Who What and when Result
VrbaS. StanleyG. DentonM. Prentice
Pulses of savannaexpansion =
> 2Aridification
>Global cooling (,5.0,
3.0, 2.8 to 2.4, 1.7,and 1.0 Ma)
Turnover (in Homoand robust austra-lopiths)
Novel adaptations toopen conditions(e.g., terrestri-ality, stone tool-making)
110 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

1992; Kingston et al., 1994; Sikes et al., inpress). This means that open grassland, akey ingredient of the savanna hypothesis,was by no means a regular feature of theEast African landscape by the late Plioceneor even the early Pleistocene. Comparison offorest and open-habitat chimpanzees offersanother interesting perspective. Tool use,hunting, food-sharing, greater social coopera-tion, and other traits usually associatedwith human evolution appear to be morestrongly displayed by closed-habitat chimps.On this basis, Boesch-Achermann and Boe-sch (1994) have suggested that woodland orforest may have provided the initial settingin which hominins evolved. The savannahypothesis has thus been criticized by pri-mate researchers, paleontologists, and envi-ronmental scientists, many of whom favorreplacing it with a woodland/forest-adapta-tion model.
A related idea—the riparian-woodlandscavenging model—also underlines the im-portance of closed habitat to early humans.In the modern Serengeti, animal carcassesare found to be seasonally abundant inwooded areas near water. This observationled Blumenschine (1986, 1987; Cavallo andBlumenschine, 1989) to suggest that a scav-enger niche could exist in riparian wood-lands. Thus, during Oldowan times, marrow-eating toolmakers may have occupied thewooded zones near water, where they couldexploit scavenging opportunities, which werefar less available in open settings. The ripar-ian-woodland model, like other versions ofthe woodland/forest hypothesis, supposesthat woodlands offered a vital challenge oropportunity—in this case, carcasses thatcould be exploited with the help of stonetools—which grasslands did not.
Short-term variability hypothesis. Theideas considered thus far see adaptation as aresponse to the consistent challenges ortendencies posed by a specific type of habi-tat. According to a different type of explana-tion, important adaptive changes may arisein response to environmental variation. Vari-ability hypotheses can be divided into short-term (e.g., seasonal variation) and long-term(i.e., variability selection).
The short-term perspective pays attentionto variations experienced by an organismover its lifetime, especially seasonal fluctua-tion, disease, and change from year to year.This view, which we may call the seasonalityhypothesis, places emphasis on how an or-ganism adjusts to alternative environmentsduring its lifetime, such as warm-cold orrainy-dry periods. Short-term variation isespecially marked in certain regions, particu-larly savannas, higher-latitude biomes, orany other habitat prone to large moisture ortemperature seasonality. Strong seasonal-ity, furthermore, brings about interannualvariation. The rainy season may be extremein one year but fail in another; the differencebetween winter and summer temperaturemay be large in some years but much less inothers.
The influence of short-term habitat varia-tion on savanna organisms, including earlyhumans, is discussed by Foley (1987). Foleynotes that although early hominins occupiedwooded-to-open savanna, the most signifi-cant aspect of this type of habitat is itsmarked seasonality. Driven by monthly vari-ance in rainfall, significant shifts occurwithin each year in the abundance andavailability of resources. Savanna organ-isms are adapted not to the variation per sebut to the pattern of variation. It is theexpected cyclicity of resources and theiravailability that is important (Foley, 1987).Short-term hypotheses of adaptive evolutionthus underline repeatable variations, whichrepresent regularities in the annual (or life-time) adaptive setting of an organism.
The difference between seasonality andother hypotheses, then, can be subtle. Habi-tat-specific views lay stress on the ever-present, or consistent, adaptive problems ina given setting. The short-term variabilityidea emphasizes habitat variation that anorganism is likely to experience and adapt toin every generation.
Long-term (variability selection) hypoth-esis. In contrast to seasonality and habitat-specific ideas is the variability selectionhypothesis. According to this view, environ-mental variation is accentuated over thelong term and plays a critical role in theadaptive process. Environmental change in-
111ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

volves large, episodic shifts in adaptive set-tings over many hundreds of thousands ofyears. As a result, a lineage of organismsmay face multiple, substantial disparities inselective environment over time. This ideafollows from the previously coined term vari-ability selection (Potts, 1996a,b, in press a).The variability selection hypothesis thusstates that certain adaptations have evolveddue to large environmentally caused incon-sistencies in selective conditions. This dispar-ity in the Darwinian optimum favors thebuildup of complex mechanisms for dealingwith unexpected, episodic change. This pro-cess of adaptive evolution therefore en-hances an organism’s capacity to thrive innovel conditions.
Variability selection (VS) calls attentionto the large remodeling of landscapes, vegeta-tion, animal communities, and regional hy-drology over long time frames. The long-term sum of habitat variability far exceedsseasonal or other types of variations encoun-tered by individuals within their lifetime.This recurrent, wide-scale revision of envi-ronment can thus have an unusual impacton the adaptive properties of a lineage. Theseasonality hypothesis implies that sea-sonal (or, more broadly, generation-scale)change can explain the emergence of adap-tive flexibility. The VS hypothesis, by con-trast, seeks the evolutionary cause of versa-tility in longer intervals of more dramaticchange in an organism’s survival regime.
The VS hypothesis requires that differentindividuals in different generations of thesame lineage experience marked inconsisten-cies in adaptive setting. As it is magnifiedover time, the disparity in adaptive milieushapes an overall selective environment thatcannot be predicted from the more consis-tent selective environments of shorter peri-ods. Selective disparity ultimately favorsgenes that build mechanisms of adaptiveversatility. It promotes a process of adapta-tion to novelty, including settings never pre-viously encountered. The resulting adapta-tions do not anticipate future conditions (aviolation of natural selection theory); rather,their evolution stems from a prior history ofmajor, periodic revision of the adaptive set-ting.
In VS, therefore, the selective effect isintegrated over time. This sets up the situa-tion in which habitat-specific traits end upeventual losers to features that in the pastproved successful in novel surroundings.This unusual process thus improves adapta-tion to long-term environmental dynamics.(See Potts (in press a) for a more detaileddiscussion and response to challenges.)
By contrast, habitat-specific and seasonal-ity explanations of evolution usually assumethe operation of directional selection, inwhich the fitness results tend to be consis-tent over the long term. They focus on aspecific setting, trend, or properties of varia-tion that recur from generation to genera-tion. The VS hypothesis is thus easily distin-guished from both habitat-specific andseasonality hypotheses.
Test implications
Each of these hypotheses has testableimplications—that is, a set of expectationsregarding
● the general paleoenvironmental record ofthe late Cenozoic;
● the precise association of hominins andhabitats;
● change in the breadth of habitats in whichhominins lived;
● the context in which hominin adaptationsfirst became apparent and continued tothrive; and
● the functions or utility of particular adap-tive features and trait complexes.
Habitat-specific hypotheses. Global andregional environmental records should showthat a particular type of habitat (e.g., wood-land, savanna, glacial cold) was either main-tained over a prolonged period or developedover time as a trend. Hominin fossils orartifacts, moreover, should be associatedconsistently with that particular type ofpast environment or at least a relativelynarrow habitat range. The influence of aparticular habitat type on adaptation canbest be demonstrated in a situation in whichalternative habitats were also present (andin which hominins were rare or absent).
The main expectation therefore is thathominins lived in and were especially at-tracted to a particular set of environmental
112 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

conditions. Accordingly, adaptive featuresshould be well suited to those conditions andshould have first appeared in that specifictype of habitat. Functional analysis shouldtherefore show that those features had prop-erties (e.g., shape, size, biomechanical, be-havioral, or ecological properties) that wereparticularly beneficial in the environmentspecified by the hypothesis (e.g., arid sa-vanna, woodlands, glacial cold).
Short-term variation (seasonality) hy-pothesis. The main expectations of thishypothesis, distinguishing it from the oth-ers, are that 1) hominins lived in settings ofhigh seasonality, 2) key adaptations reflectthe scale of habitat variation typically facedover a lifetime, and 3) large-scale fluctua-tions have occurred on seasonal, decade, andcentury time scales during spans of majoradaptive change.
Short-term variation may be quite ex-treme. According to climatologists Crowleyand North (1991:11), ‘‘the seasonal cycle isthe largest climate change we know of—thechange in temperatures over North Americafrom winter to summer is far greater thanglacial-interglacial changes in mean annualtemperatures of the Pleistocene.’’ By thisreasoning, seasonal cycles represent the wid-est range of environmental change to whichorganisms adapt in both temperate (Calvin1996) and tropical latitudes (Foley 1987).
Greenland ice-core records further indi-cate that huge deviations in climate mayoccur abruptly on a decade-to-century timescale—thus within the lifetimes of humansand other long-lived organisms. It is pos-sible that such rapid oscillation—and dra-matic shifts on an even shorter cycle, such asEl Nino—characterized the period of humanevolution (Calvin, 1996).6 Thus, short-termvariation can account for the range of set-tings to which hominins had to adapt. Ac-cordingly, large interannual variations inhigh seasonality regions should dominatethe environmental record of hominin evolu-tion.
Variability selection hypothesis. The VShypothesis suggests a series of well-defined
observations that differentiate it from otherhypotheses. Long-term global and regionalrecords should show large-scale environmen-tal fluctuation. The degree of environmentalchange should become larger over long timeperiods, greatly exceeding the scale of sea-sonal or lifetime fluctuation. The emergenceof key hominin adaptations, moreover,should coincide with spans of enhanced envi-ronmental fluctuation. In addition, theseadaptations should promote versatile behav-iors and responsiveness to habitat diversityand change.
The VS hypothesis also implies a set ofexpected observations regarding hominin-environment associations in the geologicrecord. These expectations are best pre-sented in a comparison with habitat-specificexplanations, as summarized in Table 3.
Comparison of these two sets of expecta-tions leads to an important point. Paleocli-mate records clearly demonstrate that dra-matic environmental oscillation occurredover the past 6 million years (Figs. 2–4), afinding that appears to support the VS hy-pothesis. Change does not mean, however,that the selective environment of hominins(or other organisms) was similarly altered. Agiven species may track the geographic shift-ing of key resources as its environment ismodified. In this way, an organism can stabi-lize its selective milieu rather than adapt tonovel variations.7 Thus, the external environ-ment (such as climate) and the selectiveenvironment of an organism may sometimesvary independently (Brandon, 1990). Forthis reason, habitat-specific explanations ofhominin evolution are quite plausible, forthey imply that early human populationscontinued to exploit certain favored habitatsdespite overall shifts in global and regionalenvironment (Table 3, expectation 1). Alter-natively, hominins may have mainly beenaffected by the overall trend, the averagechange in environment, and the trend was amore important selective factor on homininsthan the variance (range and pace of fluctua-tion) (Table 3, expectation 2).
6The El Nino cycle, however, may be only 5,000 years old(Sandweiss et al., 1996).
7Conversely, change in an organism’s selective environment(e.g., competitors, predators, disease vectors) may occur indepen-dent of climate, tectonics, or other geologic factors that alterhabitats.
113ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

The VS hypothesis differs here. It claimsthat external variance strongly impactedthe selective environment of hominins. Itasserts that variable conditions created awide disparity in adaptive settings overtime. This disparity then created an overallselective environment that fostered adap-tive change in hominins via the process ofvariability selection.
The most important factor in testing theseexpectations or any environmental hypoth-esis is the detail and precision of data re-lated to paleoenvironments and their asso-ciation with hominins (fossils or artifacts).With this in mind, I turn to four timeintervals in which evidence of homininsand/or paleoenvironments is reasonably de-tailed in certain localities or regions, suffi-cient at least for an initial test of the hypoth-eses defined here.
ENVIRONMENTAL RECORDS AT EARLYHUMAN LOCALITIES
6.0 to 2.5 million years agoEastern and southern Africa. Table 4summarizes environmental reconstructionsof ten hominin localities older than 2.5 Ma.These represent the likely contexts of earlyaustralopiths. A diversity of species, how-ever, inhabited these locales: in eastern Af-
rica, Ardipithecus ramidus (Aramis), Austra-lopithecus anamensis (Kanapoi, Turkana),and Au. afarensis (Hadar, Laetoli, Turkana);and in southern Africa, Au. africanus(Makapansgat, Sterkfontein). Specimenssometimes attributed to the Hominini areknown from Lothagam and Tugen Hills (Hilland Ward, 1988) but lack diagnostic charac-ters related to bipedality or canine reduc-tion. No hominins have yet been recoveredfrom late Miocene and early Pliocene beds atKanam (Kanam and Homa Formations),though new work provides an outline of theenvironmental sequence there. Taking allthe localities into consideration, Table 4shows that no single environmental recon-struction applies to the oldest recognizedmembers of the hominin clade or to thegeographic regions where they have beenfound.
Kanapoi, Tugen Hills, Lothagam, Aramis,and Kanam possess sediments between 6and 4 million years old. Large mammals andisotopic data from Kanapoi, Tugen Hills,and Lothagam suggest that a mosaic of openand closed vegetation typified East Africaduring this interval and that a relativelystable mosaic might have persisted for along time.According to Kingston et al. (1994),a heterogeneous setting composed of a mix
TABLE 3. Test expectations concerning the geologic association between hominins and paleoenvironments:Habitat-specific versus variability selection hypotheses
Habitat-specific hypotheses Variability selection hypothesis
Expectation 1
Expectation 2
Expectation 3
Over long periods, hominins were tied toa specific habitat type—for example,open savanna (savanna hypothesis),relatively closed habitat (woodland/forest hypothesis), cold steppe or peri-glacial setting (ice-age hypothesis)
Over long periods, hominins became asso-ciated with a new type of habitat (i.e.,habitat use changed in a specific direc-tion following the environmental trendof the period) (e.g., increasing homininpresence in open conditions supportsthe savanna hypothesis, even if earlierhominins were in woodland settings)
In a given stratigraphic interval (within abasin), hominins are correlated with aspecific habitat type (requires demon-strating the contemporaneity of dif-ferent habitat types); over time, thishominin-habitat correlation was eithermaintained or permanently shifted to anew habitat type
Expectation 4
Expectation 5
Expectation 6
The oldest hominins occupied a diversearray of habitats (responsible for theemergence of initial adaptations); overtime, hominins occupied an ever-widerdiversity of environments as theyevolved new means of adaptive versa-tility (correlated with widening envi-ronmental extremes manifested overthe long term)
Early in time, habitat use by homininsfocused on a specific vegetational set-ting (or other well-defined environ-ment) but became more diverse later onas the range of environmental fluctua-tion increased
In any given stratigraphic interval, homi-nins are found in a specific environ-mental setting; but in a long strati-graphic section (over long periods ofhabitat change), the sum total ofhominin environments is highlydiverse (i.e., hominins persistedthrough widely fluctuating environ-ments within a locality)
114 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

of C3 and C4 plants prevailed in centralKenya over the past 15.5 myr. These datasuggest long-term environmental stability,and that spatial habitat diversity (ratherthan long-term fluctuation) played a strongrole in the origin of bipedality.
With regard to Kanapoi (4.2 to 4.0 Ma),Leakey et al. (1995) similarly suggest thatthe oldest known bipeds inhabited a spatialmosaic of vegetation. Interpretation of themammal microfauna, however, suggests thatKanapoi’s environment was open and rela-tively dry around 4 Ma. Although trees werepresent, extensively wooded tracks did notapparently occur (Winkler, 1995).
The bovid, elephant, and equid fauna fromLothagam (7.9 to 4.7 Ma) supports the mo-saic interpretation. The fossils suggest amixture of riverine forest, woodland, andopen grassy patches, although the assem-blage is time-averaged over a lengthy pe-riod, making it difficult to discern the vari-ance between stratigraphic units. Stableisotopic study of soil carbonates also impliesa mixed habitat at Lothagam, a prevalenceof C3 plants but with C4 grasses also present(Leakey et al., 1996; Harris and Cerling,1995).
At Kanam (6.12–3.5 Ma), fossil verte-brates relate to a diverse, spatially mosaicenvironment. Isotopic data plotted with fine
stratigraphic resolution give evidence of atemporal oscillation between 30 and 70% C4grasses. The data suggest a mixed C3/C4plant community but with nonoverlappingvariations through time (Potts et al., 1997).Since hominin fossils have yet to be found inthis sequence, it is unclear whether bipedsever encountered the range of habitat changeimplied by the isotopic data.
At the Ethiopian site of Aramis (4.4 Ma),fossil wood, abundant seeds of the woodland/forest-dwelling Canthium, and the mix offossil vertebrates (rich in primates andtragelaphine bovids) all point to a moreclosed habitat (woodland or forest) than isgenerally recognized at the Kenyan sites(WoldeGabriel et al., 1994). The paleobotani-cal evidence, which seems particularly im-portant, derives from a carbonate horizonjust beneath the pedogenically altered siltsthat contain Ardipithecus ramidus and otherfossil vertebrates.
Later, at Laetoli (3.8 to 3.5 Ma), thegeologic clues (aeolian tuffs, calcite-rich soils,soil-weathering products) largely point to adry savanna, where vegetation was at leastseasonally insufficient to prevent wind trans-port of sand-sized ash particles (Hay, 1981,1987). Interpretation of the fossil pollensamples agrees with this open savanna-grassland interpretation (Bonnefille and Ri-
TABLE 4. African environments from late Miocene to Pliocene, prior to 2.5 million years ago
Locality Age (Ma) InterpretationHypothesis (conditions
of early bipedality) References1
Sterkfontein 3.5 to 2.5 Closed vegetation Climbing adaptation (forest) 1, 2Makapansgat 3.3 to 3.0 Many competing views
Fossil mammals and arthropods Savanna, woodlands, or forest 3–7Pollen: forest/bush = grass/shrub Fluctuation 8Pollen: forest (earliest australopiths) Adaptation to forest 9
Omo-Turkana 4.0 to 2.5 Dramatic floral change in gallery forest Forest mosaic or fluctuation 10–13Dry to wooded savanna plus gallery forest Adaptation to the mosaic 14
Hadar 3.9 to 3.0 Forest/bush = open country = grassy/plain = forest/bush
Fluctuation 15–17
Laetoli 3.8 to 3.5 Open savanna with seasonal aridity Adaptation to savanna 18–21Woodlands and wooded savanna Adaptation to woodland 22
Kanapoi 4.2 to 4.0 Mixed vegetation Adaptation to the mosaic 23Dry, open vegetation (micromammals) Adaptation to savanna 24
Aramis 4.4 Forest-woodland Adaptation to forest 25Tugen Hills 5.0 to 2.5 Mixed grassy woodland Adaptation to the mosaic 26Kanam 6.1 to 3.5 Closed = open = closed vegetation Fluctuation 27Lothagam 7.9 to 4.7 Mixed open woodland Adaptation to the mosaic 28, 291 References: 1, Clarke and Tobias, 1995; 2, Berger and Tobias, 1996; 3, deGraff, 1960; 4, Cooke, 1978; 5, Kitching, 1980; 6, Vrba, 1988;7, Reed, 1997; 8, Cadman and Rayner, 1989; 9, Rayner et al., 1993; 10, Bonnefille and DeChamps, 1983; 11, Bonnefille, 1984; 12,Bonnefille and Letouzey, 1976; 13, Williamson, 1985; 14, Brown, 1981; 15, Johanson et al., 1982; 16, Gray, 1980; 17, Bonnefille, 1995;18, Hay, 1981; 19, Hay, 1987; 20, Leakey and Harris, 1987; 21, Bonnefille and Riollet, 1987; 22, Andrews, 1989; 23, Leakey et al., 1995;24, Winkler, 1995; 25, WoldeGabriel et al., 1994; 26, Kingston et al., 1994; 27, Potts et al., 1997; 28, Leakey et al., 1996; 29, Harris andCerling, 1995.
115ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

ollet, 1987). However, a different reading ofthe evidence, largely the faunal remains,suggests a more closed, wooded communityof mammals, similar to woodland habitats ofthe Serengeti region today (Andrews, 1989).
Around the same time, 4 to 3 million yearsago, Hadar and the Omo-Turkana basinoffer evidence of a diverse range of habitats.Stratigraphic analysis of fossil wood, fruit,and pollen (Bonnefille, 1984, 1995; William-son, 1985) and large mammals (Gray, 1980;Johanson et al., 1982) suggest that consider-able fluctuation in climate, vegetation, andavailable water sources (basin hydrology)occurred over the long term in these areaswithout clear sign of an aridity trend.
The wide range of environmental interpre-tations of Pliocene sites, both in eastern andsouthern Africa (Table 4), depends on thetype of geologic evidence, time period, andeven the taxonomic group of fossil animalsand plants that are assessed in one study oranother. This range includes apparent sup-port for a savanna interpretation and for itsantithesis, in which tropical forest or wood-land is considered the favored environmentof early bipeds. At Makapansgat, for ex-ample, pollen data examined by Rayner etal. (1993) suggest that A. africanus wasadapted specifically to forested habitat. Yetan earlier study of fossil pollen from thissame site emphasized the ‘‘fluctuating cli-matic and vegetational conditions’’ in whichthis species lived (Cadman and Rayner, 1989:112).
Discussion. Late Miocene and earlyPliocene environments in Africa providedthe context of the oldest bipedal apes. Infor-mation about these environments is ulti-mately pertinent to the origin of bipedalityitself. This issue continues to be one of themost problematic in the study of homininevolution.
The largest problem in testing environ-mental hypotheses with data from theselocalities concerns the poor connection be-tween hominin fossils and paleoenviron-ments. Linkage in time and space betweenthese two types of data is imprecise. Rarelydo samples for paleoenvironmental analysiscome from the exact loci and sedimentaryenvironment in which hominin fossils were
originally buried or, based on taphonomicanalysis, in which the hominins may actu-ally have lived and died. At a locality likeMakapansgat, paleoenvironmental data maycome from the same stratigraphic memberas fossils of Australopithecus, but the amountof time-averaging represented by any mem-ber may be very long. Analysis of a singlestratigraphic unit may yield an averageenvironment largely irrelevant to the actualenvironments (and their variance) encoun-tered by early humans. At Laetoli, surfacecollecting of nearly all of the fossils meansthat 300,000 years of fossil animals aretreated as if they reflect a single communityor environment. Arguments about the bestor most accurate reconstruction of the Laetolipaleoenvironment are useless without bet-ter stratigraphic and analytical control overthe data. This same problem applies toHadar, Kanapoi, Tugen Hills, and most lo-calities.
The evidence reviewed here nonethelesssuggests a preliminary interpretation. Thebody of data does not consistently favoreither the savanna or forest adaptationmodel, even though data from individuallocalities or stratigraphic levels have beenused to defend both interpretations. Thepresence of Ar. ramidus in closed vegetation(at Aramis) coupled with Au. afarensis inmore open settings later in time (at Hadarand Laetoli) might suggest that the latterevolved the capability of inhabiting moreopen environments, while Ardipithecus wasconfined to closed woodlands (WoldeGabrielet al., 1994). This interpretation would favorthe savanna hypothesis (Table 3, expecta-tion 2). This view can be valid, however, onlyif eventually more than one Ar. ramiduslocality is found and closed habitat is deter-mined to have been the domain of otherpopulations of this species.
The combined evidence from all localitiesindicates that in general the oldest homi-nins were associated with a diverse range ofhabitats—forest, woodland, open savanna,and mosaic habitats. Lacking stratigraphicresolution in most of these sequences, it isstill evident that hominins experienced awide range of adaptive conditions at differ-ent sites and at different times. Notwith-standing the comparison between Aramis
116 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

and Hadar/Laetoli, there does not seem to bea dominant environmental trend in the data.At some sites, there are hints of long-termfluctuation in vegetation and climate. Com-bining all the localities also seems to sup-port this idea on a broad regional scale.
Hence, I have argued previously that earlyhominin bipedality may best be explained asa response to variability selection (Potts,1996a,b, in press a). Beyond the terrestrialevidence, the deep-sea isotopic record showsa dramatic rise in global environmentalvariability between 6 and 5 Ma relative tothe variability in each 1 million year inter-val back to 27 Ma (Potts, in press a) (see Fig.2B). The rise is enhanced even more be-tween 4 and 3 Ma. As Table 4 shows, habitatreconstructions are also sufficiently variedto be consistent with a rise in environmentalfluctuation, essential to the VS hypothesis.
The key to this interpretation, though,rests on the original functions of bipedalbehavior over the period of its origin. Ofwhat use was australopith bipedality in theenvironments in which it evolved and waslater maintained? Based on utility, it makessense that a novel form of terrestrial locomo-tion arose during forest reduction and sa-vanna expansion. The function of early aus-tralopith locomotor structure has, however,been controversial. According to one camp,early bipeds were committed to terrestrialbipedality; their skeletal structure was de-signed to conduct this form of locomotioneffectively (e.g., Lovejoy et al., 1973; Latimerand Lovejoy, 1990; Ohman et al., 1997).According to the other camp, the overallskeletal structure of Pliocene hominins indi-cates the importance of arboreal activity.Terrestrial bipedality was not particularlyeffective, and the behavior of these homininswas largely geared to life in the trees (e.g.,Stern and Susman, 1983; Susman and Stern,1991). The debate largely centers, then, onthe degree to which early australopiths werecommitted to either arboreal climbing orterrestrial bipedal striding.
That this controversy persists, however,suggests a different adaptive explanation.The two viewpoints can be reconciled to acertain degree by challenging the either/orquestion. Environmental fluctuation led, atdifferent times and places, to settings with
dense stands of trees and larger tracts ofopen vegetation. Responsiveness to this typeof environmental disparity was, over time, avital aspect of locomotor function. The loco-motor system of the earliest hominins, bythis interpretation, enabled them to accom-modate to the habitat variance. The originalfunctional benefit of bipedality resided inthe versatility it afforded as local, prehomi-nin populations faced a changing series ofvegetation types over time. As a result, theearly australopiths possessed a locomotorsystem that persisted for 2 million years ormore, adaptable to the diverse settings thatarose in the hominin geographic range be-tween 6 and 2.5 Ma.
In brief, expectations 1 and 3 (Table 3)concerning a habitat-specific interpretationseem to be contradicted even by the poor-resolution data available. Temporal and spa-tial resolution of hominin-environment asso-ciations is too poor either to support or rejectexpectation 2 (Table 3). The savanna hypoth-esis (as an explanation of bipedality) wouldbe favored if it could be shown, eventually,that 1) prehominins tended to occupy rela-tively closed vegetation, 2) the earliest homi-nins tended to live (and die) in less closedvegetation and, later still, in fairly openhabitats, and 3) locomotor and positionalfunctions of early australopiths were (incontrast to the argument above) largelydedicated to terrestrial two-legged walking.
The VS hypothesis, by contrast, does notappear to be contradicted by any aspect ofthe evidence. However, the sample size ofsites, the resolution of time, and the preci-sion of fossil-environment associations arenot adequate to test this hypothesis effec-tively. The oldest bipeds may have inhabiteda wide range of habitats (Table 3, expecta-tion 4). In any one time or place, they mayhave occupied a fairly specific habitat whichvaried according to the available vegetation(Table 3, expectation 6). The data, though,do not appear to support expectation 5 (Table3), the expansion of habitat diversity overtime.
Despite global and some local evidence offluctuation, it is quite conceivable that amosaic of open and closed vegetation (pro-moted by seasonal variation) favored theorigin of bipedality, including a mixture of
117ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

terrestrial and arboreal activities in earlyPliocene hominins. The key question hereconcerns the stability of the open/closedmosaic over time. Was the mosaic itselfadequate to account for the range of environ-ments to which early Pliocene homininswere adapted? Or was a more extensiverange of variability, expressed over longertime periods, crucial to the evolutionaryprocess? This, of course, is not necessarilyan either/or issue, as both spatial and tempo-ral variability may have played importantroles. The existing data from the 6 to 2.5 Matime span, however, do not appear to implyhabitat stability.
2.3 to 1.7 million years agoTurkana and Olduvai basins. Domi-nated by fluvial settings for most of itshistory, the Turkana basin offers a richdataset for assessing environmental changeassociated with early humans of the latestPliocene. In this period, Turkana (includingthe Omo region) underwent a series of majorpaleogeographic changes (Feibel, 1988; Fei-bel et al., 1991; Brown and Feibel, 1991).During the same time, hominin toolmakersconsiderably altered their landscape interac-tions. This is evident in the archeologicalrecord from the growing diversity of paleoen-vironments in which stone tools were dis-carded (Rogers et al., 1994).
About 2.3 Ma, a large, meandering river,like the Omo today, occupied the axis of thebasin. The river was fed by braided streamsthat descended from the eastern and west-ern highlands. Archeological sites, whichconsisted of low-density artifact clusters,were focused where the braided and mainriver systems met. Proximity to both aspectsof the fluvial system was apparently criticalin inducing hominins to produce and depositstone tools. Rocks, which were obtainedfrom the braided, marginal drainages, neednot have been transported by toolmakersany great distance, as stream conglomerateswere available within several tens of metersup to a few kilometers from the artifact sites(e.g., Lokalalei site: Kibunjia, 1994).
By about 1.9 Ma, the landscape hadchanged dramatically. The center of theTurkana basin was occupied by a sizable yetfluctuating lake fed from the north by the
proto-Omo River. Artifact concentrationswere as low as before, and the choice of rawmaterial was still quite localized, with trans-port distances possibly much less than 25km (Rogers et al., 1994). The main differenceis that archeological sites were stationed inat least three different contexts. One set-ting, most similar to earlier times, was atthe intersection between braided and axialdrainages. The main river itself, some dis-tance from the braided tributaries, was usedas a second setting; unlike the earlier pe-riod, the proto-Omo river itself was appar-ently a useful source of lithic raw material.The third setting occurred at the lake mar-gin, where alluvial fans from the westernhighlands drained into the lake. Accordingto Rogers et al. (1994), archeological re-mains in this context are the only knownPlio-Pleistocene lake-margin sites in the Tur-kana basin. The fluctuating lake persistedto about 1.7 Ma (Brown and Feibel, 1991).Within the interval 1.8 to 1.7 Ma, a majorshift in the isotopic composition of paleosolshas been noted, which signifies a likelyexpansion of open C4 plant habitat withinthe Turkana basin (Cerling et al., 1988;Cerling, 1992). This vegetation shift is re-corded at the top of the Olduvai geomagneticsubchron, currently estimated to be 1.757Ma (Cande and Kent, 1992).
By 1.7 Ma, another major paleogeographicshift had occurred. The central lake disap-peared, replaced by an unstable fluvial sys-tem. A very dynamic series of landscapesensued. Environments dominated by an axialriver alternated with others consisting ofintricate expanses of braided streams. Thispattern of fluctuation, filling in of channels,followed by further carving of the flood-plains, was influenced by massive volcanicactivity in the region. Dramatic change oc-curred as volcanic ash blanketed the terrainoccupied by toolmakers. Archeological sitesoccur in channel, gravel bar, overbank, andfloodplain settings—a far greater diversitythan in any earlier period, which seems tofollow from the temporal fluctuation andgreater spatial division of the landscape(Rogers et al., 1994).
The Olduvai basin offers a second impor-tant dataset for the latest Pliocene. Duringthis period, the basin was lake-dominated;
118 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

many archeological remains are preservedin lake-margin settings, a point of consider-able contrast to Turkana. The sediments ofBed I and lower Bed II at Olduvai (Fig. 6)are considered to span perhaps 50–100 kyr,bracketed between dates of 1.9 and 1.7 Ma(Hay, 1976a; Walter et al., 1991). A smallalkaline lake (7–25 km wide) persistedthroughout this time; thus, hydrologicalchange was not as dramatic as in the longerTurkana sequence. Sedimentary, isotopic,faunal, and pollen studies nonetheless pointto impressive shifts in climate and vegeta-tion (Hay, 1976a; Cerling, 1992; Cerling andHay, 1986; Sikes, 1994; Butler and Green-wood, 1973, 1976; Jaeger, 1976; Potts, 1988;Bonnefille and Riollet, 1980; Bonnefille,1995).
Three significant transitions are apparentin the environmental record of Bed I andlower Bed II (Fig. 6). Wet, marshy, andwooded conditions low in Bed I (approxi-mately 1.8 Ma) gave way to aridity and amore open setting near the top of this unit.In basal Bed II, there was a return to moremoist and wooded habitats. Finally, the Le-muta Member at the top of lower Bed IIsignals the prevalence of dry conditions withsparse vegetation coverage. Near the end ofthe Olduvai geomagnetic event (approxi-mately 1.76 to 1.75 Ma), recorded near thebase of the Lemuta Member, there was amarked shift in soil d13C and d18O (Cerlingand Hay, 1986). Evidently, this reflects adramatic change in the percentage of C4plants—from 20–40% up to 60–80%, repre-senting an open, grassy environment. Thisshift may be correlated with the similar oneat Turkana (Cerling, 1992). The aridity con-ditions of the Lemuta Member are esti-mated to have lasted perhaps 50 kyr (Hay,1976a).
Stone tool sites—both concentrations andscatters of artifacts—occur throughout thesequence. This suggests that Oldowan tool-makers had the means to adjust to thechanges they faced. Averaged over the se-quence, there was one major environmentaltransition every 33–50 kyr, which is withinthe range of the 41 kyr periodicity related toorbital obliquity. Later, in upper Bed II toBed IV times, the lacustrine system wasdisrupted, and numerous changes in fluvial
hydrology and terrain took place in responseto tectonic and climatic influences (Hay,1976a).
Discussion. Two important developmentsin this time span were the origin of Homoergaster (early African H. erectus), appar-ently by 1.88 Ma,8 and expansion of theranging patterns of the toolmakers. Withbody size and limb proportions similar tomodern humans, H. ergaster was an endur-ance-oriented biped whose skeleton ab-sorbed high mechanical stresses probablyrelated to locomotion (Ruff et al., 1993).With the Nariokotome skeleton (KNM-WT-15000) as a guide, body shape was linearand adapted to heat stress, typical of mod-ern humans from hot, dry, tropical areas.Since it was not well adapted to relativelyclosed, wet environments, H. ergaster mayhave marked a transition in the climaticadaptation of early hominins (Ruff andWalker, 1993; Ruff, 1991).
The Nariokotome skeleton (approximately1.50–1.55 Ma) postdates the dramatic shiftto an arid, hot climate signaled in the Tur-kana and Olduvai sequences at about 1.75Ma. In this regard, the climate-sensitiveshape of this particular individual makessense. Stature estimates based on otherfossils from Turkana further suggest thatthe transition to tall, linear body form (x 5170 cm, n 5 6) had occurred by 1.70 Ma,whereas individuals who lived about 1.88Ma (represented by fossils ER 1472, 1481,and 3728) were significantly shorter (x 5147 cm, n 5 3) (Ruff and Walker, 1993). Ingeneral, then, evidence for an adaptive shiftin body form is well correlated with orimmediately follows evidence of a majorshift to hot, open habitat.
The first appearance of H. ergaster, how-ever, is perhaps 130 kyr older than thisclimatic event. An arid interval slightly olderthan the major environmental shift seems tobe recorded in upper Bed I Olduvai (Fig. 6).Pollen analysis suggests that this environ-ment supported mainly small herbaceousplants rather than trees (Bonnefille andRiollet, 1980). Yet this environment is no
8Based on occipital specimen KNM-ER-2598 from the KBSMember of the Koobi Fora Formation (Rightmire, 1991; Wood,1991).
119ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]
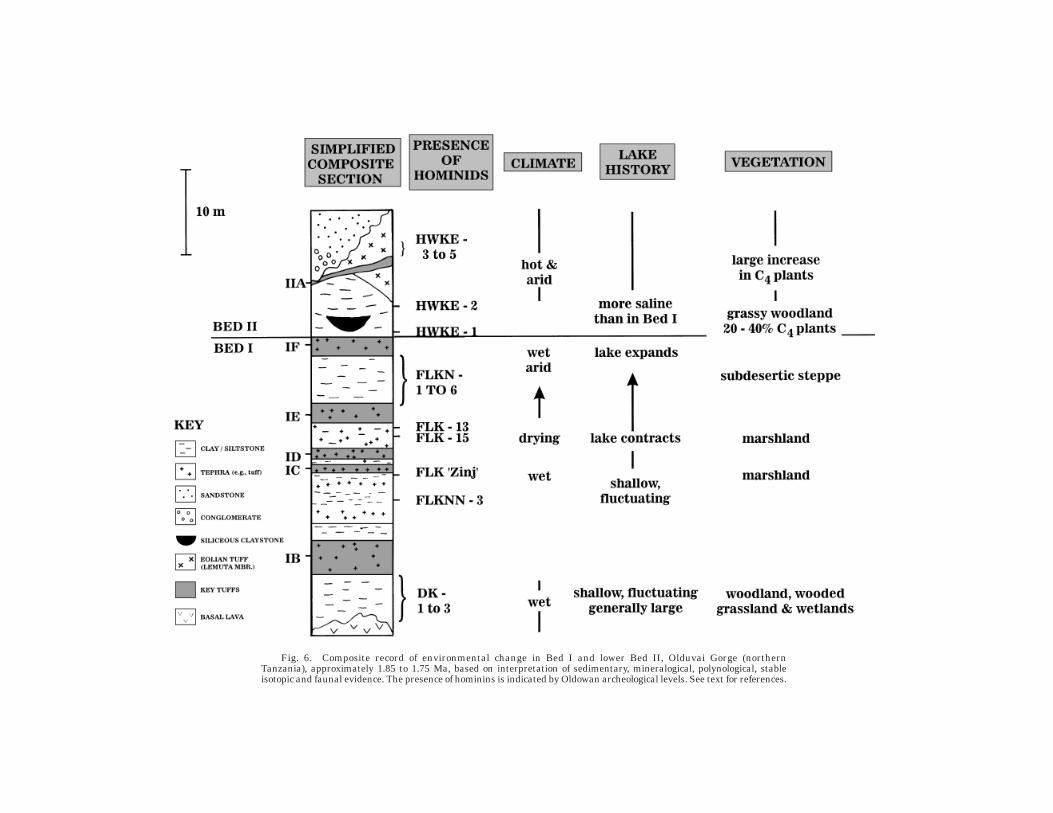
Fig. 6. Composite record of environmental change in Bed I and lower Bed II, Olduvai Gorge (northernTanzania), approximately 1.85 to 1.75 Ma, based on interpretation of sedimentary, mineralogical, polynological, stableisotopic and faunal evidence. The presence of hominins is indicated by Oldowan archeological levels. See text for references.

older than 1.8 Ma (Walter et al., 1991), stillafter the first appearance datum of H. ergas-ter. One explanation is that the adaptiveshift in body form was decoupled from specia-tion, evident also in the climatic adaptationof modern humans. If, on the other hand,further discoveries of postcranial bones indi-cate that body form changed in this speciesbefore 1.8 Ma, the adaptive shift precededthe most notable grassland expansion discov-ered so far in the East African Plio-Pleis-tocene record.
The second key development in the period2.3 to 1.7 Ma is reflected by archeologicallyvisible behaviors related to habitat use andstone transport. A significant problem hereconcerns the temporal and geographic over-lap of potentially multiple species of toolmak-ers. It is not possible, except by specialpleading to unsupported assumptions, tosay which species was/were responsible forthe stone tools deposited in any given stra-tum or place. Overlap of possibly three lin-eages of Homo, as well as the presence ofParanthropus, underscores the problem, andat present there is no obvious solution.Thus, in this period of the Plio-Pleistocene,paleoanthropologists can do no more thanmake observations about generic stone tool-makers—overall shifts in archeological pat-terns independent of how specific behaviorswere apportioned to lineages. This state ofaffairs is less than ideal for evolutionaryanalysis. But observations of this sort doallow important changes in behavioral strat-egies to be identified, which may be correlat-able with the rise of new taxa. Even whenthey are not correlated, generic behavioralshifts are important to identify in an analy-sis of hominin evolutionary history.
During the late Pliocene, Oldowan flakingwas conservative. The main behavioralchanges concerned the environments inwhich stone tools were used and left. In theTurkana and Olduvai basins, stone trans-port distances increased slightly, and arti-facts were left in a much greater diversity ofenvironments by the end of this period com-pared to its beginning (Rogers et al., 1994;Potts, in press b). In addition, the processingof animal tissues intensified. Bones fromarcheological sites manifest obvious damageas a result of hominins obtaining meat and
marrow from large animals (.30 kg) (e.g.,Bunn and Kroll, 1986; Potts, 1988; Blumen-schine, 1995; Monahan, 1996).
As noted above, these changes took placeat Turkana in a dynamic series of environ-ments. During a period of landscape instabil-ity (approximately 1.9 to 1.5 Ma), toolmak-ers became less tethered to local rock sourcesand particular habitats. They carried stonesfarther and made clusters of tools in morediverse areas of the landscape. The favoredareas included both old and new habitats(e.g., near the axial channel and on fluvialfloodplains). The evidence thus points to anexpansion in landscape use rather than adirectional shift.
The toolmakers of Bed I Olduvai tended touse stone sources within the confines of thelake floodplain. Transport distances rarelyexceeded 3 km (Leakey, 1971; Hay, 1976a).After a series of environmental fluctuations,the toolmakers of middle Bed II expandedtheir range to depositional environmentsboth within and beyond the immediate lake-margin area. Stone sources outside the lakemargin were also newly exploited. Thesedevelopments at Olduvai, then, parallelthose in the Turkana basin.
At both localities, toolmakers were activethrough substantial shifts in environmentand were present in arid, moist, warm, cool,more wooded, more open, lake-dominated,and fluvial-dominated phases. This suggeststhat Oldowan toolmakers could accommo-date to periodic shifts in climate, basinhydrology, and vegetation. The persistenceof hominin toolmakers and their eventualexpansion into a greater range of habitatsare consistent with the VS hypothesis. Inthis light, the transport of resources overlonger distances, occupation of a wider habi-tat range, and intensification of animal foodexploitation may represent means of dealingwith large changes in the distribution ofhabitats, water, food, and other necessities.If this interpretation is correct, these behav-ioral changes can be considered adaptationsto environmental instability rather than toany one specific type of habitat (e.g., wood-land) or directional trend (e.g., increasinglyarid savanna).
121ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

1 million to 250,000 years agoOlorgesailie basin. Located in the riftvalley of southern Kenya, Olorgesailie is aPleistocene lake basin well known for denseaccumulations of Acheulean stone bifaces(Isaac, 1977) and a well-calibrated record offossil mammals and environmental changefrom about 1.2 Ma to 49 ka (Isaac, 1978;Owen and Renaut, 1981; Potts, 1989, 1994,1996a; Deino and Potts, 1990; Tauxe et al.,1992; Potts et al., in press). The main por-tion of the Olorgesailie Formation, compris-ing 14 members, is dated between .992 kaand 493 ka, a span of about 500,000 years.The geologic sequence consists of lake diat-omites, associated floodplain and fluvial sedi-ments, and tephra.
Figure 7 depicts a composite stratigraphicsection of the Olorgesailie Formation (basedon observations by R.M. Shackleton, G.L.Isaac, A.K. Behrensmeyer, and R. Potts). Italso indicates the stratigraphic position andrelative concentration of stone artifacts asdetermined by recent research in the region.The illustration shows, in general terms, theperiods of stability and change in the Olorge-sailie basin and enables an estimate of therate of major environmental revisions indifferent parts of the stratigraphic section.The datable sequence can be divided intofive main intervals. Several significantchanges are obvious in each interval (seeFig. 7 for details).
1. Members 1–5 (992 to 974 ka). Coveringapproximately 18 kyr, five major remodel-ings of the landscape can be documentedafter the lake became established (Mem-ber 1). These changes affected basinwidehydrology in ways that would have al-tered the distribution of water-depen-dent plants and animals.
2. Member 6/7 (974 to 780 ka). This part ofthe section records three major landscapechanges. Toolmakers interacted with aseries of fluvial, fluctuating lake-marginand stable, vegetated environments. Fromthe dated pumice in Member 5 to theBrunhes/Matuyama boundary at the topof Member 7, a span of approximately194 kyr is represented. Most of this timeis taken by upper Member 7 soil forma-tion (Potts et al., in press).
3. Members 8 and 9 (780 to 662 ka). Threemajor shifts in the depositional and hydro-logical regime of the basin are recorded inthis period of about 118 kyr.
4. Members 10–12 (662 to 601 ka). In a spanof 61 kyr, three dramatic environmentalchanges occurred involving the disappear-ance, reappearance, and expansion of thelake.
5. Members 13 and 14 (601 to 493 ka). Inthis section, covering an estimated 108kyr, two major environmental shifts areinferred, which correspond to the twomembers originally recognized by Shack-leton (1978).
Artifacts occur throughout the entire sec-tion. There is no consistent association be-tween the density of stone artifacts (a par-tial measure of the level of hominin activity)and specific kinds of depositional environ-ment or hydrological regime. Toolmakersdid not occupy the basin continuously, butthey left evidence of their activity in fluvialand lacustrine settings and at times whenthe lake was large, small, or not apparent atall. Analysis of paleosol stable isotopes by N.Sikes (Sikes et al., 1997, in press, and inpreparation) has also documented change invegetation, including a grass-dominatedopen-savanna setting in upper Member 1and more closed, bushland conditions inlater members. Overall, vegetation was moreopen than in the period prior to 1 Ma, asdocumented at other East African sites.Based on isotopic data from Olorgesailie andOlduvai, Cerling (1992) infers that grass-lands became a persistent part of East Afri-can environments sometime after 1 Ma,perhaps as late as 600 ka. Even within thismore open setting, extensive and recurrentalteration of hydrology and landscape fea-tures was the dominant environmental sig-nal of the Olorgesailie Formation. Modifica-tions occurred in episodes several thousandsto tens of thousands of years apart, elicitedby climatic, tectonic, and volcanologicalcauses.
Zhoukoudian, the Loess Plateau, andBose basin, China. Outside of Africa,hominin localities with lengthy stratigraphicrecords (.500,000 years) and well-cali-
122 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

brated environmental sequences are ex-tremely rare. One of the few that come closeto meeting these two criteria is Zhoukou-dian, the famed Peking Man (Homo erectus)cave, southwest of Beijing in northern China.A section over 40 m thick at ZhoukoudianLocality 1 has been divided into 18 strati-
graphic units, though sedimentation ap-pears to have been largely uninterrupted.This section ranges from older than 780 kaat the base (layers 13–18) to about 230 ka(based on uranium series analysis) near thetop (layers 1–3). Between 1927 and 1982,hominin fossils and stone artifacts were
Fig. 7. Major environmental changes recorded in 14 members of the Olorgesailie Formation (southernKenya), 992 to 493 ka, based on lithology and diatoms. The concentration of stone tools within particularstrata is indicated in relative terms: high, medium, and low.
123ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

recovered from layers 10 through 2, coveringan estimated time range of 460 to 230 ka. H.erectus fossils are known from layers 2, 4, 8,and 10. Definite stone artifacts derive fromlayers 3–10, with the exception of layer 5(Black et al., 1933; Jia and Huang, 1990;Binford and Ho, 1985).
A variety of views has been offered aboutthe paleoenvironments of Zhoukoudianbased on Locality 1. Mammalian fossils exca-vated from each layer provide a rudimen-tary dataset but demonstrate convincinglythat substantial environmental change oc-curred during the period of H. erectus’spresence in or near the cave. According to Xuand Ouyang (1982), species that signal acool, grassland habitat outnumber foresttaxa in layers 11 and 10. Layers 9 and 8 aredecidedly different: grassland rodents areabsent, and forest mammals dominate theassemblage, indicating a change to a warm,humid environment. Slightly colder condi-tions prevailed in the time of layers 7 and 6,and a warm, humid fauna reappeared in thespan of layer 5. Layer 4 contains a cold-weather mammalian fauna, with a predomi-nance of grassland taxa. Finally, a cool,grassland-forest mosaic prevailed in layer 3times. Xu and Ouyang (1982) consider thissequence to match up with d18O stages 12through 8, which implies that the conditionsat Zhoukoudian were affected by the phasesof global cooling-warming and evaporationreflected in the deep-sea oxygen isotopecurve. Based on Zhoukoudian sediments,fossil pollen, and fauna, Liu (1983) supportsthis idea and notes that cold-warm fluctua-tions intensified during the middle Pleis-tocene. Jia and Huang (1990), however, dis-agree with this conclusion. Their view of themammalian fauna is that a single cool-warm-cool cycle is indicated between layers 11 and1 and that in general all levels included amixture of warm- and cool-adapted taxa.According to Jia and Huang (1990), most ofthe mammalian remains from Zhoukoudianbelong to species that were very adaptable toclimatic change.
To the south and west of Zhoukoudian isthe Loess Plateau, on which thick loess depos-its are distributed over an area of 274,000km2 (Liu, 1988). Although no hominin re-mains have, to my knowledge, been recov-
ered from the loess deposits, hominin locali-ties such as Lantian, Dali, and Dingcun aresituated in the loess region.
As reviewed earlier, China’s loess sequencefurnishes a record of environmental changeover the past 2.4 myr. Extensive fluctuationin vegetation, moisture, and temperaturecharacterized this region of north-centralChina (Fig. 4). Over the past 1 million yearsor so, loess accumulated uninterrupted for40 kyr or more on at least six occasions,beginning approximately 1,020 ka, 830 ka,650 ka, 480 ka, 180 ka, and 75 ka (Kukla andAn, 1989). (According to data illustrated inFig. 4, the earliest of these episodes mayhave entailed considerable fluctuation.) Theloessic intervals, which represented dry, rela-tively treeless habitat, were interspersedwith soil formation periods in which dry tomoist forest prevailed.
By contrast, southern China is generallybelieved to have had an extremely stablePleistocene climate. Red lateritic soils occurthroughout much of this region and intoSoutheastAsia and India. The laterites rangefrom a few meters to over 10 m in thickness,and their rather uniform appearance hasbeen taken to imply a static Pleistoceneenvironment (e.g., Teilhard de Chardin etal., 1935; Movius, 1969).
This characterization, however, appearsto be incorrect. Based on recent work in theBose basin, Guangxi Autonomous Region,an 8 m thick laterite containing stone tools(dated approximately 730 ka) exhibits strongcyclicity of clay, heavy minerals, and othersediments (possibly loess) when examinedon a microstratigraphic scale (10 cm inter-vals) (Huang and Potts, in preparation). ThePleistocene red soils of south China, in fact,appear to have formed as part of a climaticcycle linked to the loess-paleosol cycles tothe north (Zhu et al., 1995). Furthermore,analysis of fossil pollen obtained from coresdrilled in the South China Sea indicatesrecurrent north-south migration of tropicaland subtropical vegetation zones throughsouthern China during the Quaternary(Wang et al., 1991). Together, these lines ofevidence suggest that middle and late Pleis-tocene inhabitants of South China, like homi-nin populations in other parts of the world,
124 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

faced a significant range of environmentalvariability over time.
Discussion. Several evolutionary changesoccurred in hominins over the past 1 millionyears. An increase in relative brain size wasamong the most prominent from an adaptivestandpoint, and it is interesting to examinehow environmental change might relate toit. Rapid encephalization in hominins oc-curred during the middle Pleistocene, be-tween 780 and 130 ka (Ruff et al., 1997;Aiello and Wheeler, 1995). There is growingevidence, moreover, that cognitive and be-havioral competence expanded in homininsduring this same interval. This evidencemay include hearths and control of fire(Kretzoi and Dobosi, 1990), manufacture ofwell-designed spears (Thieme, 1997), neuro-logical control over speech (Kay et al., 1998),occupation of high latitudes (Waters et al.,1997), and possibly ways of enabling islandhabitation (Morwood et al., 1998). It is un-clear what parts of this cognitive/behavioralpackage were possessed by early humans ofOlorgesailie, Zhoukoudian, or central China.But as these localities offer among the beststratigraphic records for this period, theyprovide a way to assess the environmentsfaced by certain hominin populations asbrain size and behavior evolved.
None of these records lends support tohabitat-specific explanations of homininadaptive change. Substantial environmen-tal shifts, local to global in scale, occurredover the past 1 million years. Large-scalechanges were manifested over periods of104–105 years. At Olorgesailie, 16 majorrevisions of basin hydrology, paleogeogra-phy, and depositional environment occurredbetween 992 and 493 Ma, an average ofabout 31 kyr between each transition. Thedistribution is uneven, however, with meanintervals of about 3.6, 20, 39, 54, and 65 kyrbetween transitions for different parts of thestratigraphic section. Since depositionalrates were neither constant nor continuous(e.g., erosional phases at the base of M6/7and M9), these estimates do not representactual periodicities. Yet they do imply thatlarge-scale revamping of the terrain andenvironment, greatly exceeding seasonaltypes of oscillations, took place over lengthy
periods of time. In each record examinedhere, long-term modifications presented amuch more varied, complexly changing se-ries of adaptive conditions at different timesthan any one population would have facedover seasonal or interannual periods. Thesepoints are at odds with the expectations ofthe seasonality and habitat-specific hypoth-eses and are consistent with the idea ofvariability selection (Potts, in press a,b).
140,000 to 50,000 Years AgoCombe Grenal, France. The rock shelterat Combe Grenal, excavated by Bordes (Bordeset al., 1966), provides one of the longest latePleistocene sequences in Europe and themost complete record of environmentalchange in southwestern France between 140and 50 ka (Laville et al., 1980; Mellars,1996). Nearly 13 m thick, the sedimentsrepresent the fill of a small cave, which weredivided during excavation into 64 archeologi-cal layers. Figure 8 illustrates two datasets,the percentage of arboreal pollen and rela-tive abundance of two cervid species, thatserve as measures of climatic conditions.These are displayed in relation to the lithicindustries deposited in the shallow cave byMiddle Paleolithic toolmakers.Although Bor-des (1961) believed the diversity of indus-tries reflected different cultural traditions,others interpret the variations in terms ofdifferent activities, different phases of stonereduction, or temporal phases of a singlecultural tradition. It is generally believedthat populations of Neanderthals were re-sponsible for these varied stone tool assem-blages.
The Combe Grenal record documents ex-tensive changes in environment, which aresummarized by Mellars (1996).
1. Corresponding to d18O stage 6, layers 64to 56 reflect severe glacial conditions. Anextremely cold, dry, open climate is sig-naled by sedimentary frost cracking withlittle chemical weathering, large percent-ages of reindeer (92–97% of all ungulates[not represented in Fig. 8B]), and consis-tently low levels of tree pollen, most ofwhich is pine.
2. A hiatus is then represented by heavyweathering and soil formation of earlierdeposits. As no new sediments were laid
125ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]
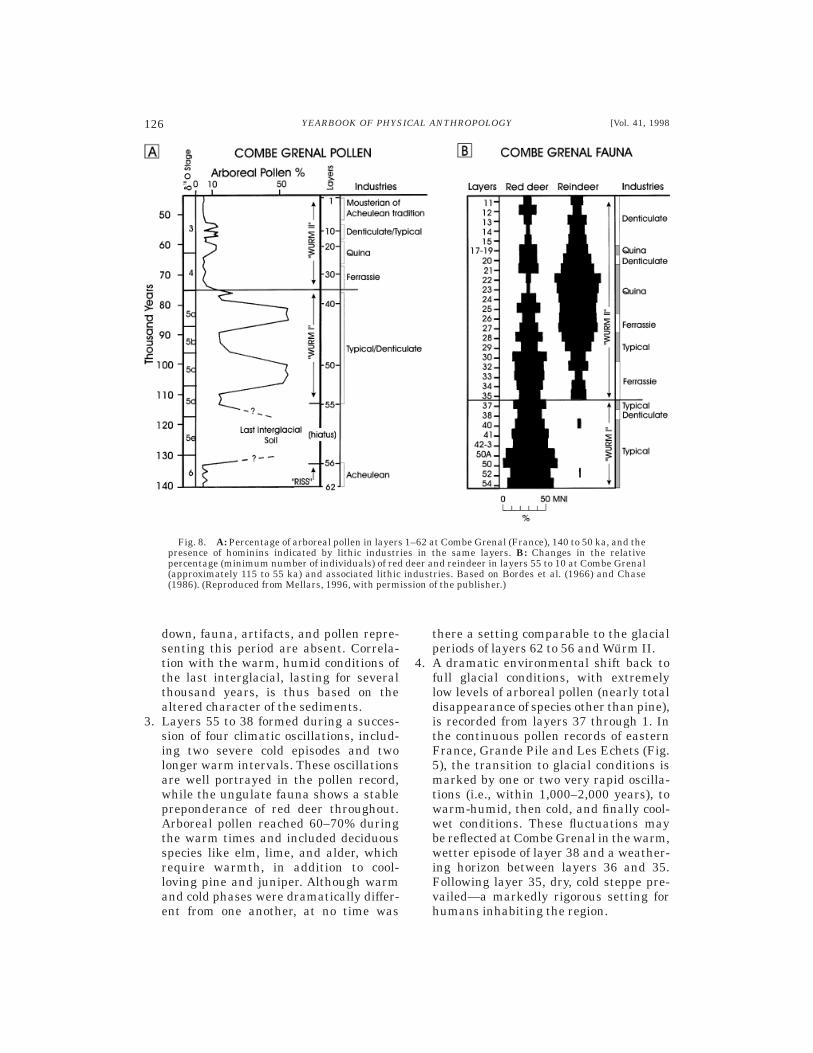
down, fauna, artifacts, and pollen repre-senting this period are absent. Correla-tion with the warm, humid conditions ofthe last interglacial, lasting for severalthousand years, is thus based on thealtered character of the sediments.
3. Layers 55 to 38 formed during a succes-sion of four climatic oscillations, includ-ing two severe cold episodes and twolonger warm intervals. These oscillationsare well portrayed in the pollen record,while the ungulate fauna shows a stablepreponderance of red deer throughout.Arboreal pollen reached 60–70% duringthe warm times and included deciduousspecies like elm, lime, and alder, whichrequire warmth, in addition to cool-loving pine and juniper. Although warmand cold phases were dramatically differ-ent from one another, at no time was
there a setting comparable to the glacialperiods of layers 62 to 56 and Wurm II.
4. A dramatic environmental shift back tofull glacial conditions, with extremelylow levels of arboreal pollen (nearly totaldisappearance of species other than pine),is recorded from layers 37 through 1. Inthe continuous pollen records of easternFrance, Grande Pile and Les Echets (Fig.5), the transition to glacial conditions ismarked by one or two very rapid oscilla-tions (i.e., within 1,000–2,000 years), towarm-humid, then cold, and finally cool-wet conditions. These fluctuations maybe reflected at Combe Grenal in the warm,wetter episode of layer 38 and a weather-ing horizon between layers 36 and 35.Following layer 35, dry, cold steppe pre-vailed—a markedly rigorous setting forhumans inhabiting the region.
Fig. 8. A: Percentage of arboreal pollen in layers 1–62 at Combe Grenal (France), 140 to 50 ka, and thepresence of hominins indicated by lithic industries in the same layers. B: Changes in the relativepercentage (minimum number of individuals) of red deer and reindeer in layers 55 to 10 at Combe Grenal(approximately 115 to 55 ka) and associated lithic industries. Based on Bordes et al. (1966) and Chase(1986). (Reproduced from Mellars, 1996, with permission of the publisher.)
126 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

Although data are lacking for the lastinterglacial (hiatus), it appears that Nean-derthal populations occupied the cave andits surroundings essentially during the en-tire sequence, leaving late Acheulean andMousterian artifacts in all settings.
Discussion. The environmental condi-tions of Mousterian toolmakers in westernEurope ranged from glacial steppe/tundra tointerglacial temperate forests. The last inter-glacial period (d18O stage 5e) represented anespecially warm period that contrastedstarkly with intense, long-lasting glacialconditions immediately before. In the initial5 kyr of the last interglacial, global meanannual temperature increased 10–15°C (asdid local temperature) (Fig. 5), and sea levelrose 5–6 m above that of the present (com-pared with 100 m lower during isotope stage6). Interglacial conditions (and interstadialperiods during the last glacial) established atemperate climate that included cold win-ters, to which trees of the European decidu-ous forests were adapted. Hominin popula-tions in the vicinity of Combe Grenal,therefore, lived in either seasonally or peren-nially colder surroundings than populationsin most other parts of the world.
Western European Neanderthals were ob-viously equipped to deal with large changesin vegetation, fauna, and climate, such asthose documented at Combe Grenal. Thesehominins produced a more complex stonetechnology and diverse series of tool kitsthan their predecessors, which may haveserved a wider variety of special tasks. Thepattern of faunal remains at Combe Grenal(Fig. 8B), moreover, is informative aboutforaging behavior. The shift from red deer toreindeer matches the shift from more tem-perate habitat (layers 55 to 37) to cold, dryconditions (layers 35 to 10). This suggeststhat the foragers of Combe Grenal were ableto adjust to major alterations by changingtheir focus from a climatically tolerant yetmainly temperate species (red deer) to a coldsteppe species (reindeer). The tremendousstability of red deer representation in layers55 to 37 (approximately 115 to 75 ka), how-ever, further implies that Mousterian tool-makers in this area also stabilized theiraccess to certain focal resources even as
climate and vegetation varied over time.The possible strategies for accomplishingthis might have involved shifting to sea-sonal (e.g., summer) use of the site duringcold phases (isotope stages 5a and 5c), whichmight have enabled continued access to reddeer herds, or long-distance tracking of reddeer (and transport of parts back to CombeGrenal) as its range shifted in response toclimatic oscillation.
Both patterns—initial stability and laterchange in faunal exploitation—suggest theconsiderable adaptability of Mousterian tool-makers at Combe Grenal. The same conclu-sion is warranted by the nearly continuouspresence of toolmakers through a period ofvast environmental alteration. As we haveseen in other places and times, this body ofevidence best fits the VS hypothesis (e.g.,Table 3, expectation 6). The humans ofCombe Grenal certainly were not tied to asingle type of habitat. Nor did they appear tobe more successful as the environmentchanged in one particular direction com-pared with another. Moreover, thousands totens of thousands of years passed betweenmajor remodelings of the adaptive setting.Long-term shifting of vegetation and cli-mate far exceeded seasonal ranges of varia-tion. This observation conflicts with anyattempt to explain the continuous presenceof hominins in this region over time in termsof adaptation to seasonality.
Yet on the basis of limb-bone proportions(Trinkaus, 1981; Holliday and Falsetti, 1995)and nasal and facial morphology (Wolpoff,1980; Coon, 1962), the skeletal structure ofEuropean Neanderthals appears to havebeen adapted specifically to colder environ-ments. This body of evidence implies thatNeanderthal (or ancestral) populations un-derwent directional selection adapting themto habitats ranging from cool to glacial cold.This view is, of course, supported by theenvironmental evidence from western Eu-rope. From an adaptive perspective, then,the Neanderthals (Mousterian toolmakersof the early Wurm) were an intriguing com-posite. They evidently had behavioral strat-egies that permitted them to persist throughwide changes in habitat. Yet the one consis-tency was a relatively cold to cool temperate
127ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

(i.e., seasonally cold) climate; their bodiesmanifested adaptations to this regularity.
Despite evidence of behavioral adaptabil-ity, there is also evidence that Mousteriantoolmakers were somewhat conservative intheir responsiveness to changing circum-stances. As Chase (1986) points out, forexample, faunal exploitation patterns atCombe Grenal did change with climate butwere slow to do so. Even though the humansat this site ultimately switched to huntingreindeer during the period beginning aroundlayer 37, they did not do so quickly (Fig. 8B).Compared with modern humans who popu-lated the same region later, Mousterianhunters did not specialize nearly as much byexploiting different animals when climatechanged. In many Upper Paleolithic sites ofthe last glacial period, for example, reindeerrepresented up to 95–99% of the faunalremains, suggesting that hunting was moredistinctly tuned to climatic conditions thanevidenced at Combe Grenal. This is just oneof several lines of evidence indicating thatlater occupants of western Europe weremore responsive to environmental dynamicsthan the Neanderthals (Mellars, 1996).
ENVIRONMENTAL PERSPECTIVESON ADAPTIVE CHANGE
From this review, there appears to be nosingle, all-encompassing environmental ex-planation of hominin evolution. Diverse se-lective processes operated on early humanpopulations in different lineages, times, andplaces. In the anatomy of Homo ergaster andNeanderthals, for example, the results ofadaptation to specific conditions are mani-fested: in H. ergaster, heat stress in an opentropical African habitat; in Neanderthals,adaptation to perennially and seasonallycold habitats in ice-age Europe and Asia.Seasonality, moreover, was a consistent partof the contexts in which hominin evolutionoccurred. Seasonal variations are critical tomaintaining the savanna mosaic of Africaand the temperate habitats of Eurasia (e.g.,deciduous forests and steppe). For this rea-son, it is often impossible to distinguishbetween evolutionary responses to specifictypes of habitat and to seasonal aridity orcold.
Although the recurrent opportunities andchallenges of savannas, woodlands, and ice-age habitats all had an effect on adaptation,the analysis here adds a different dimensionto the picture of hominin evolution. Directstudy of past strata highlights the inconsis-tency of adaptive settings over long timeframes and indicates that certain key homi-nin adaptations emerged during periods ofintense habitat instability.
The results of this analysis can be summa-rized as answers to three questions:
1. Did the origin of major hominin adapta-tions usually coincide with the appearance ofa particular type of habitat or with a span ofrising environmental variability?
On the basis of oxygen isotope data, thepast 6 million years were a time of risingenvironmental variability. The range of cli-matic fluctuation increased in successiveperiods. Variability continued to rise to thepoint that, by the middle Pleistocene, thevariation in single isotopic oscillations (cov-ering 100 kyr) usually equaled or exceededthe average change in d18O over the past 6million years. Variability in global climatetherefore swamped the overall trend towardcooling and drying. There is no doubt thatenvironments did become colder and morearid, but reversals in the trend had arguablya more significant impact on Earth’s cli-matic history since the late Miocene. Theemergence of hominins coincided with theoldest intervals of rising variability. Com-plex adaptations unique to humans origi-nated as climatic variability became magni-fied. Furthermore, virtually all other proxiesof past environments—continental dust, pol-len, and loess sequences—confirm that habi-tat variability increased over time in terres-trial settings.
2. How were early humans and environ-ments correlated over the course of homininevolution? In regions occupied by early hu-mans, were their remains consistently associ-ated with a particular type of habitat, astable mosaic of settings, or a highly fluctuat-ing range of environments?
All four time spans—and limited numberof hominin localities—explored here mani-fest a high degree of habitat remodeling overtime. In long stratigraphic sequences, theamount of variation greatly exceeded sea-
128 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

sonal types of variation; the tempo and kindof environmental alteration, moreover, dif-fered qualitatively from seasonal oscillation.Evidence of hominins occurred through es-sentially the full diversity of paleoenviron-ments that ensued over time, as reflected inlengthy stratigraphic sections. This sug-gests that hominin populations actually en-countered numerous alterations of the land-scape, climate, and biota in the localities wehave examined. These findings violate theexpectations of habitat-specific hypotheses—expectations 1–3 (Table 3)—and the season-ality hypothesis. But they are not usually indisagreement with expectations 4 to 6 (Table3), implied by the VS hypothesis.
The data available for most hominin locali-ties, though, are still inadequate to fully testthese three latter expectations. It is unclear,for example, whether African hominins ofthe early Pliocene encountered more or lesshabitat fluctuation than middle Pleistocenehominins. Whether a wider degree of habi-tat change was represented by 500 kyr atHadar or by an equivalent time span atOlorgesailie cannot yet be answered. Thus,the available data cannot distinguish be-tween expectations 4 and 5 (Table 3) relativeto the VS hypothesis. Very few studies haveattempted to address the habitat specificityof hominins within narrow stratigraphic in-tervals and across time (Table 3, expectation6) (Potts et al., in press). The oxygen isotoperecord gives a hint that environmental diver-sity, although already magnified in the pe-riod between 6 and 3 Ma, was further exag-gerated in successive intervals over the spanof hominin evolution (Fig. 2B). It remains tobe seen whether the sum of environmentalrecords from hominin sites during this spanwill show the same pattern.
3. What were the functions of key adapta-tions? Were they designed to best meet theregular, consistent challenges of one maintype of environment or directional change orto best respond to large-scale habitat incon-sistency over the long term?
We have briefly assessed the functions ofcertain hominin adaptations (or adaptivetrends), including bipedality, encephaliza-tion, stone tool transport, and growing cogni-tive sophistication. These fundamental char-acteristics are included in virtually every
scenario of human evolution. In each case,these features first appeared and becameelaborated within very dynamic environmen-tal sequences.
Obviously, ground bipedality makes sensein a landscape with few trees. On this logic,bipedal striding evolved as an adaptation toincreasingly open settings, in line with ahabitat-specific view. By studying the paleo-environmental records of hominin sites, wefind, however, that bipedal behavior emergedand was maintained over a long time in awide variety of contexts—in relatively closed,open, mixed/mosaic, and (where sequenceshave sufficient time depth) fluctuating envi-ronments. This finding seems to resolve thedebate about bipedal vs. arboreal compe-tence in A. afarensis: namely, this and otherearly australopiths possessed a locomotor/positional system that enabled the com-bined competence of both. A single anatomi-cal complex oriented toward versatility(climbing and walking) evolved as land-scapes were remodeled over many thou-sands of years. If this interpretation is cor-rect, it means that the locomotor system ofthe earliest hominins was adapted to theinstability of late Miocene and early Pliocenehabitats. Although very few records exist inAfrica from the period prior to the earliestaccepted australopiths, the global ocean re-cord (e.g., Fig. 2) suggests that environmen-tal instability was in fact on the rise duringthe latest Miocene.
A similar conclusion applies to other ma-jor adaptive features when analyzed in anenvironmental context. Enlargement of thebrain relative to body size during the middlePleistocene is a case in point. Our analysisreveals that habitats of this period under-went large revisions and that populationswere subject to strong inconsistencies inselective environment. Considering the ac-tual evidence, the view often propounded byevolutionary psychologists, who have madea large impact on the study of human cogni-tive evolution, needs to be reassessed. It ispatently incorrect to characterize the hu-man ancestral environment as a set of spe-cific repetitive elements, statistical regulari-ties, or uniform problems which the cognitivemechanisms unique to humans are designedto solve (e.g., Tooby and Cosmides, 1989,
129ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

1990, 1993; Pinker, 1994). This portrait ofthe Pleistocene environment should be dis-carded and with it the view that the humanmind is composed mainly of innate special-purpose devices or algorithms tied to a par-ticular array of past adaptive possibilities(Potts, in press a).
A variability-selection perspective, by con-trast, implies that the cognitive mecha-nisms evolved in Pleistocene humans fos-tered the input, analysis, and mentalrepresentation of highly varied external in-formation and the output of versatile, novelresponse. The archeological record of thepast 700 kyr strongly suggests that brainfunctions and actual behavior were designedto accommodate a diverse series of adaptivesettings, also evidenced in the archives ofPleistocene environmental history.
It is unlikely that the emergence andspread of modern H. sapiens, in its ecologicaldiversity, was preceded by a long process ofhabitat-specific adaptation. The apparentreplacement of Neanderthals in western Eu-rope by populations more tropically adapted(Trinkaus, 1981; Holliday and Falsetti, 1995)is especially significant, for it implies prioradaptation to environmental diversity on agrand scale, which yielded populations fit toaccommodate to the hot tropics, the coldregions where Neanderthals had lived, andeven more extreme environments. The so-cial adaptations of these late Pleistocenehumans included sophisticated means ofbuffering environmental uncertainty andthriving in novel settings (Potts, in press b).
To summarize, there is very little basis forthinking that adaptive settings were consis-tent over many generations or changed in aconsistent direction during the course ofhominin evolution. Habitat-specific sce-narios, which have prevailed in paleoanthro-pology, furnish a poor framework for explain-ing the origin of most complex humanadaptations. Short-term habitat variationalso offers a limited basis on which to ac-count for key adaptive changes. This hypoth-esis, as we noted earlier, relies on the claimthat short-term variation, such as seasonal-ity, was sufficiently large to have led to theadaptive flexibility observed in humans. Theenvironmental records examined here con-tradict this claim. The time frame of environ-
mental variability in hominin localities wasnot both rapid and extreme (i.e., unlike theGreenland ice cap). Rather, variability inlandscape structure, vegetation, and re-sources became greatly extended over spansof 104–105 years and involved a substan-tially different type and range of environ-mental remodeling than exhibited on a sea-sonal or lifetime scale. Adaptation to short-term patterns of variation does not thereforeresult in adaptation to the longer time frameof habitat remodeling.
As a final point, it is important to notethat the episodic revision of landscapes in-habited by hominins would also have alteredthe social context in which individuals com-peted for resources. Resource restructuringand uncertainty, according to the VS hypoth-esis, fostered novel strategies of competitionand social cooperation. Given the environ-mental dynamics we have discovered, thereis little basis for dismissing ecological fac-tors while embracing social competition asan explanation of cognitive evolution andbehavioral complexity (contra Humphrey,1976). The two surely went hand in hand.Indeed, the case can be made that a stronglycooperative dynamic within social groupsproved highly beneficial over the long termas a means of accommodating to unexpectedecological variability.
CONCLUSIONS
Three main kinds of environmental expla-nation can be considered in scenarios ofhominin adaptive evolution. My goal hasbeen to present these ideas as testable hy-potheses. Habitat-specific hypotheses focuson the adaptive opportunities or challengesof open savanna, densely wooded, or ice-age(cool temperate and periglacial) environ-ments. The seasonality hypothesis empha-sizes the impact of short-term environmen-tal variation on the adaptive process. Thevariability selection (VS) hypothesis postu-lates that wide variability in adaptive set-tings over time ultimately favored andhelped build complex adaptations whichwere responsive to novel conditions. Basedon preliminary tests of environmental re-cords from various hominin sites, the mainfindings of this review can be summarized asfollows.
130 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

1. Although usually ignored in explana-tions of human evolution, dramatic envi-ronmental fluctuation became magnifiedover long periods of time and had substan-tial impact on adaptive evolution in homi-nins. According to the VS hypothesis, thefunctions of key adaptations and adap-tive trends—Pliocene bipedality, stonetool transport and use, and Pleistocenebrain enlargement—primarily served thepurposes of behavioral versatility. Thesetraits enabled ways of adapting to envi-ronmental novelty—that is, to the dy-namic properties of environments, thusincluding settings that had not been pre-viously encountered. The selective pro-cess that brought about such traits musthave operated over broad time scales ofhabitat disparity.
2. Environment-specific adaptations didcharacterize certain lineages of homi-nins. These features, however, may havelargely involved anatomical structuresand related physiological functions ratherthan complex behavioral, cognitive, andsocial functions, which typically are cen-tral to accounts of human evolution. Habi-tat-specific scenarios do not adequatelyexplain the key adaptations they wereoriginally meant to explain. The savannahypothesis, for example, does not seem tooffer a sufficient account of the locomotorversatility that arose in early Pliocenehominins.
3. Short-term environmental variation wasubiquitous in the regions occupied byearly human populations, though season-ality or generational-scale variation usu-ally left no mark in the geologic record. Amuch more impressive archive of environ-mental fluctuation was left over spans ofmany thousands of years. Stratigraphicrecords at hominin sites show that rela-tively stable environmental periods wererepeatedly interrupted by large shifts inclimate, basin hydrology, and landscapestructure. The origin of important hu-man characteristics emerged as homininpopulations confronted a widely dispar-ate range of selective environments. Inother words, the general trends of humanadaptive evolution are better portrayedas a response to the dynamic (unstable)
properties than to the consistent proper-ties of environments since the lateMiocene.
4. It is important to note that most organ-isms are tied to a narrow range of adap-tive conditions—that is, they are habitatspecialists. In the face of environmentalfluctuations, these species were appar-ently sufficiently mobile or dispersed toenable geographic tracking of requisitehabitats and resources. Other lineagesbecame extinct, a frequent occurrence inmost clades of Pleistocene large mam-mals. In still other cases, populations orspecies lineages became adapted overtime to a growing diversity of ecologicalsettings. These lineages underwent a pro-cess of adaptive change that decoupledthem from any one specific habitat. Thisprocess seems to have occurred repeat-edly in hominins over the past 5 millionyears (Potts, in press a).
5. Further understanding of the adaptivehistory of hominins requires well-cali-brated data on paleoenvironments andtheir exact association with hominins.Tests of environmental hypotheses restheavily on correlation. The strength ofthese tests depends on improved strati-graphic resolution of the fossil, artifac-tual, and environmental records at homi-nin localities. Raising the density ofpaleoenvironmental data will prove espe-cially valuable. Future progress cannotrely on data solely from individual locali-ties. Instead, comparative study ofsamples of hominin localities from par-ticular time spans is required. This typeof research therefore leans heavily onintegrated work by teams of geologists,paleoanthropologists, environmental sci-entists, and geochronologists. Withoutthis integrated approach, paleoanthropol-ogy is relegated to sketching the phyloge-netic and technological outline of humanevolution while missing the substance ofthe story, the contexts and processes ofadaptive change.
ACKNOWLEDGMENTS
Funding for field research (Olorgesailie,Bose, Olduvai) was provided by the Smithso-nian’s National Museum of Natural History,
131ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

the Scholarly Studies Program, the NationalScience Foundation, and the Boise Fund.For discussions on variability selection andadaptation, I especially thankAlison Brooks,John Fleagle, Bernard Wood, and Kay Be-hrensmeyer. The manuscript benefited fromcomments by Peter deMenocal and ChrisRuff. I am grateful to Jennifer Clark forpreparing the figures and to Brian Rich-mond for corrections. This is a publication ofthe Smithsonian’s Human Origins Program.
LITERATURE CITED
Aiello LC, and Wheeler P (1995) The expensive-tissuehypothesis: The brain and the digestive system inhuman and primate evolution. Curr. Anthrop. 36:199–221.
Ambrose SH, and Sikes NE (1991) Soil organic carbonisotopic evidence for vegetation change in the KenyaRift Valley. Science 253:1402–1405.
Amundson R (1996) Historical development of the con-cept of adaptation. In MR Rose and GV Lauder (eds.):Adaptation. San Diego: Academic Press, pp. 11–53.
Andrews P (1989) Paleoecology of Laetoli. J. Hum. Evol.18:173–181.
Andrews P (1992) Evolution and environment in Miocenehominoids. Nature 360:641–646.
Andrews P, and Walker A (1976) The primate and otherfauna from Fort Ternan, Kenya. In GL Isaac and ERMcCown (eds.): Human Origins: Perspectives on Hu-man Evolution, Vol. 3. Menlo Park, CA: WA Benjamin,pp. 279–304.
Andrews P, Harrison T, Delson E, Bernor RL, andMartin L (1996) Distribution and biochronology ofEuropean and Southwest Asian Miocene catarrhines.In RL Bernor, V Fahlbusch, and H-W Mittmann (eds.):The Evolution of Western Eurasian Neogene MannalFaunas. New York: Columbia University Press, pp.168–207.
Bartholomew GA, and Birdsell JB (1953) Ecology andthe protohominids. Am. Anthropol. 55:481–498.
Begun DR (1992) Miocene fossil hominids and thechimp-human clade. Science 257:1929–1933.
Behrensmeyer AK, Todd NE, Potts R, and McBrinn GE(1997) Late Pliocene faunal turnover in the TurkanaBasin, Kenya and Ethiopia. Science 278:1589–1594.
Berger LR, and Tobias PV (1996) A chimpanzee-liketibia from Sterkfontein, South Africa and its implica-tions for the interpretation of bipedalism in Australo-pithecus africanus. J. Hum. Evol. 30:343–348.
Binford LR, and Ho CK (1985) Taphonomy at a distance:Zhoukoudian, ‘‘the cave home of Beijing Man’’? Curr.Anthrop. 26:413–442.
Bishop WW (1976) Pliocene problems relating to humanevolution. In GL Isaac and ER McCown (eds.): HumanOrigins: Perspectives on Human Evolution, Vol. 3.Menlo Park, CA: WA Benjamin, pp. 139–153.
Bishop WW (1978) Geological Background to FossilMan. Edinburgh: Scottish Academic Press.
Bishop WW, and Clark JD (1967) Background to Evolu-tion in Africa. Chicago: University of Chicago Press.
Black D, Teilhard de Chardin P, Young CC, and Pei WC(1933) Fossil Man in China. Geological Memoirs Se-ries A, Number 11. Peiping: Geological Survey ofChina.
Blumenschine RJ (1986) Early Hominid ScavengingOpportunities: Implications of Carcass Availability inthe Serengeti and Ngorongoro Ecosystems. Oxford:
British Archaeological Reports International Series283.
Blumenschine RJ (1987) Characteristics of an earlyhominid scavenging niche. Curr. Anthrop. 28:383–407.
Blumenschine RJ (1995) Percussion marks, tooth marks,and experimental determinations of the timing ofhominid and carnivore access to long bones at FLKZinjanthropus, Olduvai Gorge, Tanzania. J. Hum.Evol. 29:21–52.
Bock WJ (1980) The definition and recognition of biologi-cal adaptation. Amer. Zool. 20:217–227.
Boesch-Achermann H, and Boesch C (1994) Hominiza-tion in the rainforest: The chimpanzee’s piece of thepuzzle. Evol. Anthropol. 3:9–16.
Bond G, Heinrich H, et al. (1992) Evidence for massivedischarges of icebergs into the North Atlantic oceanduring the last glacial period. Nature 360:245–261.
Bonnefille R (1984) Cenozoic vegetation and environ-ments of early hominids in East Africa. In RO Whyte(ed.): The Evolution of the East Asian Environment,Vol. 2. Hong Kong: Centre of Asian Studies, pp.597–612.
Bonnefille R (1995) A reassessment of the Plio-Pleis-tocene pollen record of East Africa. In ES Vrba, GHDenton, TC Partridge, and LH Burckle (eds.): Paleocli-mate and Evolution, With Emphasis on Human Ori-gins. New Haven: Yale University Press, pp. 299–310.
Bonnefille R, and DeChamps R (1983) Data on fossilflora. In J de Heinzelin (ed.): The Omo Group: Ar-chives of the International Omo Research Expedition.Tervuren: Musee Royal de l’Afrique Centrale, pp.191–207.
Bonnefille R, and Letouzey R (1976) Fruits fossilesd’Antrocaryon dans la vallee de l’Omo (Ethiopie).Adansonia 16:65–82.
Bonnefille R, and Riollet G (1980) Palynologie, vegeta-tion et climat de Bed I et Bed II a Olduvai, Tanzanie.In Proceedings of the Eighth Pan-African Congress ofPrehistory and Quaternary Studies. Nairobi. Nairobi:TILLMIAP, pp. 123–127.
Bonnefille R, and Riollet G (1987) Palynological spectrafrom the Upper Laetolil Beds. In MD Leakey and JMHarris (eds.): Laetoli: A Pliocene Site in NorthernTanzania. Oxford: Clarendon, pp. 52–61.
Bordes F (1961) Mousterian cultures in France. Science134:803–810.
Bordes F, Laville H, and Paquereau MM (1966) Observa-tions sur le Pleistocene superior du gisement deCombe-Grenal (Dordogne). Actes de la Societe Lin-neenne de Bordeaux 103:3–19.
Brain CK (1981) The evolution of man in Africa: Was it aconsequence of Cainozoic cooling?Annex. Transv. Geol.Soc. S. Afr. 84:1–19.
Brandon RN (1990) Adaptation and Environment. Prin-ceton: Princeton University Press.
Broecker WS, and Denton GH (1990) The role of ocean-atmosphere reorganisations in glacial cycles. Quatern.Sci. Rev. 9:305–343.
Brown FH (1981) Environments in the Lower OmoBasin from one to four million years ago. In G Rapp Jrand CF Vondra (eds.): Hominid Sites: Their GeologicSettings. AAAS Symposium 63. Boulder: Westview,pp. 149–164.
Brown FH, and Feibel CS (1991) Stratigraphy, deposi-tional environments and palaeogeography of the KoobiFora Formation. In JM Harris (ed.): Koobi ForaResearch Project, Vol. 3. Oxford: Clarendon, pp. 1–30.
Bunn HT, and Kroll EM (1986) Systematic butchery byPlio-Pleistocene hominids at Olduvai Gorge, Tanza-nia. Curr. Anthrop. 27:431–452.
Butler PM, and Greenwood M (1973) The early Pleis-tocene hedgehog from Olduvai Gorge, Tanzania. In L
132 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

Leakey, R Savage, and S Coryndon (eds.): FossilVertebrates of Africa, Vol. 3. London: Academic Press,pp. 7–42.
Butler PM, and Greenwood M (1976) Elephant-shrews(Macroscelididae) from Olduvai and Makapansgat. InR Savage and S Coryndon (eds.): Fossil Vertebrates ofAfrica, Vol. 4. London: Academic Press, pp. 1–56.
Butzer KW (1971) Environment and Archeology: AnEcological Approach to Prehistory. Chicago: Aldine-Atherton.
Butzer KW, and Isaac GL (1975) After the Australopithe-cines. The Hague: Mouton.
Cadman A, and Rayner RJ (1989) Climatic change andthe appearance of Australopithecus africanus in theMakapansgat sediments. J. Hum. Evol. 18:107–113.
Calvin WH (1996) How Brains Think. New York: BasicBooks.
Cande SC, and Kent DV (1992) A new geomagneticpolarity time scale for the Late Cretaceous and Ceno-zoic. J. Geophys. Res. 97:13917–13951.
Cavallo JA, and Blumenschine RJ (1989) Tree-storedleopard kills: Expanding the hominid scavengingniche. J. Hum. Evol. 18:393–399.
Cerling TE (1992) Development of grasslands and savan-nas in East Africa during the Neogene. Palaeogeogr.Palaeoclimat. Palaeoecol. 97:241–247.
Cerling TE, and Hay RL (1986) An isotopic study ofpaleosol carbonates from Olduvai Gorge. Quatern.Res. 25:63–78.
Cerling TE, Bowman JR, and O’Neil JR (1988) Anisotopic study of a fluvial-lacustrine sequence: ThePlio-Pleistocene Koobi Fora sequence, East Africa.Palaeogeogr. Palaeoclimat. Palaeoecol. 63:335–356.
Cerling TE, Quade J, Ambrose SH, and Sikes N (1991)Fossil soils, grasses, and carbon isotopes from FortTernan: Grassland or woodland? J. Hum. Evol. 21:295–306.
Cerling TE, Harris JM, MacFadden BJ, Leakey MG,Quade J, Eisenmann V, and Ehleringer JR (1997)Global vegetation change through the Miocene/Pliocene boundary. Nature 389:153–158.
Chase PG (1986) The Hunters of Combe Grenal. Oxford:British Archaeological Reports International Series286.
Clarke RJ and Tobias PV (1995) Sterkfontein Member 2foot bones of the oldest South African hominid. Sci-ence 269:521–524.
Cooke HSB (1978) Faunal evidence for the biotic settingof early African hominids. In CJ Jolly (ed.): EarlyHominids of Africa. New York: St. Martin’s Press, pp.267–284.
Coon C (1962) The Origin of Races. New York: Knopf.Croll J (1875) Climate and Time. New York: Appleton.Crowley TJ, and North GR (1991) Paleoclimatology.
New York: Oxford University Press.Dansgaard WS, Johnsen SJ, et al. (1993) Evidence for
general instability of past climate from a 250-kyricecore. Nature 364:218–220.
Dart R (1925) Australopithecus africanus: The man-apeof south Africa. Nature 115:195–199.
Dart R (1953) The predatory transition from ape to man.Int. Anthrop. Ling. Rev. 1:201–218.
Darwin C (1871) The Descent of Man and Selection inRelation to Sex. London: Murray.
deGraff G (1960) A preliminary investigation of themammalian microfauna in Pleistocene deposits ofcaves in the Transvaal system. Palaeont. Afr. 7:59–118.
Deino A, and Potts R (1990) Single crystal 40Ar/39Ardating of the Olorgesailie Formation, southern Kenyarift. J. Geophys. Res. 95:8453–8470.
Delson E, and Tattersall I (1997) Primates. In: Encyclo-pedia of Human Biology, Vol. 7. New York: AcademicPress, pp. 93–104.
deMenocal PB (1995) Plio-Pleistocene African climate.Science 270:53–59.
deMenocal PB, Ruddiman WF, and Pokras EM (1993)Influences of high- and low-latitude processes onAfrican terrestrial climate. Palaeoceanography 8:209–242.
deMenocal PB, and Bloemendal J (1995) Plio-Pleis-tocene climatic variability in subtropical Africa andthe paleoenvironment of hominid evolution. In ESVrba, GH Denton, TC Partridge, and LH Burckle(eds.): Paleoclimate and Evolution, With Emphasis onHuman Origins. New Haven: Yale University Press,pp. 262–288.
Eldredge N (1989) Macroevolutionary Dynamics. NewYork: McGraw-Hill.
Elliot Smith GE (1924) The Evolution of Man. London:Oxford University Press.
Endler JA (1986) Natural Selection in the Wild. Prince-ton: Princeton University Press.
Feibel CS (1988) Paleoenvironments of the Koobi ForaFormation, Northern Kenya. Ph.D. dissertation, Uni-versity of Utah.
Feibel CS, Harris JM, and FH Brown (1991) Palaeoenvi-ronmental context for the late Neogene of the TurkanaBasin. In JM Harris (ed.): Koobi Fora Research Project,Vol. 3. Oxford: Clarendon, pp. 321–370.
Fisher DC (1985) Evolutionary morphology: Beyond theanalogous, the anecdotal, and the ad hoc. Paleobiology11:120–138.
Foley R (1987) Another Unique Species. Essex: Long-man.
Gould SJ (1977) Ontogeny and Phylogeny. Cambridge,MA: Harvard University Press.
Gould SJ, and Vrba ES (1982) Exaptation—a missingterm in the science of form. Paleobiology 8:4–15.
Gray T (1980) Environmental Reconstruction of theHadar Formation (Afar, Ethiopia). Ph.D. dissertation,Case Western Reserve University.
Guiot J, Pons A, de Beaulieu JL, and Reille M (1989) A140,000-year continental climate reconstruction fromtwo European climate records. Nature 338:309–313.
Guiot J, de Beaulieu JL, Cheddadi R, David F, Ponel P,and Reille M (1993) The climate in western Europeduring the last glacial/interglacial cycle derived frompollen and insect remains. Palaeogeog. Palaeoclimat.Palaeoecol. 103:73–93.
Harris JM, and Cerling TE (1995) Dietary preferences ofLothagam mammals (late Miocene, northern Kenya).J. Vert. Paleontol. 15(Supplement to no. 3):33A.
Hay RL (1976a) Geology of the Olduvai Gorge. Berkeley:University of California Press.
Hay RL (1976b) Environmental setting of hominid activi-ties in Bed I, Olduvai Gorge. In GL Isaac and ERMcCown (eds.): Human Origins: Perspectives on Hu-man Evolution, Vol. 3. Menlo Park, CA: WA Benjamin,pp. 209–225.
Hay RL (1981) Paleoenvironment of the Laetolil Beds,northern Tanzania. In G Rapp Jr and CF Vondra(eds.): Hominid Sites: Their Geologic Settings. AAASSymposium 63. Boulder: Westview, pp. 7–24.
Hay RL (1987) Geology of the Laetoli area. In MDLeakey and JM Harris (eds.): Laetoli: A Pliocene Sitein Northern Tanzania. Oxford: Clarendon, pp. 23–47.
Hill A, and Ward S (1988) Origin of the Hominidae: Therecord of African large hominoid evolution between 14my and 4 my. Yrbk. Phys. Anthropol. 31:49–83.
Holliday TW, and Falsetti AB (1995) Lower limb lengthof European early modern humans in relation tomobility and climate. J. Hum. Evol. 29:141–153.
133ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

Howell FC (1976) An overview of the Pliocene andearlier Pleistocene of the Lower Omo Basin, southernEthiopia. In GL Isaac and ER McCown (eds.): HumanOrigins: Perspectives on Human Evolution, Vol. 3.Menlo Park, CA: WA Benjamin, pp. 227–268.
Howell FC, and Bourliere F (1963) African Ecology andHuman Evolution. New York: Wenner-Gren Founda-tion.
Humphrey N (1976) The social function of intellect. InPPG Bateson and RA Hinde (eds.): Growing Points inEthology. Cambridge: Cambridge University Press,pp. 303–317.
Isaac GL (1976) East Africa as a source of fossil evidencefor human evolution. In GL Isaac and ER McCown(eds.): Human Origins: Perspectives on Human Evolu-tion, Vol. 3. Menlo Park, CA: WA Benjamin, pp.121–137.
Isaac GL (1977) Olorgesailie: Archaeological Studies of aMiddle Pleistocene Lake Basin in Kenya. Chicago:University of Chicago Press.
Isaac GL (1978) The Olorgesailie Formation: Stratigra-phy, tectonics and the palaeogeographic context of theMiddle Plestocene archaeological sites. In WW Bishop(ed.): Geological Background to Fossil Man. Edin-burgh: Scottish Academic Press, pp. 173–206.
Isaac GL, and McCown ER (1976) Human Origins:Perspectives on Human Evolution, Vol. 3. Menlo Park,CA: WA Benjamin.
Jablonski D (1987) Heritability at the species level:Analysis of geographic ranges of Cretaceous molluscs.Science 238:360–363.
Jaeger JJ (1976) Les rongeurs (Mammalia, Rodentia) duPleistocene inferior d’Olduvai Bed I (Tanzania). In RSavage and S Coryndon (eds.): Fossil Vertebrates ofAfrica, Vol. 4. London: Academic Press, pp. 58–120.
Jia L, and Huang W (1990) The Story of Peking Man.Beijing: Foreign Language Press.
Johanson DC, Taieb M, and Coppens Y (1982) Pliocenehominids from the Hadar Formation, Ethiopia (1973–1977). Am. J. Phys. Anthropol. 57:373–402.
Jolly CJ (1970) The seed eaters: A new model of hominiddifferentiation based on a baboon analogy. Man 5:5–26.
Jolly CJ (1978) Early Hominids of Africa. New York: St.Martin’s Press.
Jouzel J, Barkov NI, et al. (1993) Extending the Vostokice-core record for palaeoclimate to the penultimateglacial period. Nature 364:203–207.
Kappelman J, Plummer T, Bishop L, Duncan A, andAppleton S (1997) Bovids as indicators of Plio-Pleistocene paleoenvironments in East Africa. J. Hum.Evol. 32:229–256.
Kay RF, Cartmill M, and Balow M (1998) The hypoglos-sal canal and the origin of human vocal behavior. Proc.Natl. Acad. Sci. U. S. A. 95:5417–5419.
Kibunjia M (1994) Pliocene archaeological occurrencesin the Lake Turkana basin. J. Hum. Evol. 27:159–172.
Kingston JD, Marino BD, and Hill A (1994) Isotopicevidence for Neogene hominid paleoenvironments inthe Kenya Rift Valley. Science 264:955–959.
Kitching JW (1980) On some fossil Arthropoda from theLimeworks, Makapansgat, Potgietersrus. Palaeont.Afr. 23:63–68.
Klein RG (1989) The Human Career: Human Biologicaland Cultural Origins. Chicago: University of ChicagoPress.
Kretzoi M, and Dobosi VT (1990) Vertesszolos: Site, Manand Culture. Budapest: Akademiai Kiado.
Kukla G, and An Z (1989) Loess stratigraphy in centralChina. Palaeogeog. Palaeoclimat. Palaeoecol. 72:203–225.
Laporte LF, and Zihlman AL (1983) Plates, climate andhominoid evolution. S. Afr. J. Sci. 79:96–110.
Latimer B, and Lovejoy CO (1990) Hallucal tarsometa-tarsal joint in Australopithecus afarensis. Am. J.Phys. Anthropol. 82:125–133.
Laville H, Rigaud J-P, and Sackett J (1980) Rock Shel-ters of the Perigord. New York: Academic Press.
Leakey MD (1971) Olduvai Gorge, Vol. 3. London:Cambridge University Press.
Leakey MD, and Harris JM (1987) Laetoli: A PlioceneSite in Northern Tanzania. Oxford: Clarendon.
Leakey MG, Feibel CS, McDougall I, and Walker AC(1995) New four-million-year-old hominid species fromKanapoi and Allia Bay, Kenya. Nature 376:565–571.
Leakey MG, Feibel CS, Bernor RL, Harris JM, CerlingTE, Stewart KM, Storrs GW, Walker A, Werdelin L,and Winkler AJ (1996) Lothagam: A record of faunalchange in the late Miocene of East Africa. J. Vert.Paleontol. 6:556–570.
Lewontin R (1970) The units of selection. Annu. Rev.Ecol. Syst. 1:1–14.
Liu T (1988) Loess in China. Beijing: China OceanPress.
Liu Z (1983) The palaeoclimatic changes inferred fromPeking Man Cave deposits in comparison with theclimatic sequence of other formations. Acta Anthropo-logica Sinica 2:172–183.
Lovejoy CO, Heiple KG, and Burstein AH (1973) Thegait of Australopithecus. Am. J. Phys. Anthropol.38:757–780.
McKee JK (1996) Faunal turnover patterns in thePliocene and Pleistocene of southern Africa. S. Afr. J.Sci. 92:111–113.
Mellars P (1996) The Neanderthal Legacy. Princeton:Princeton University Press.
Milankovitch MM (1941) Canon of Insolation and theIce-Age Problem. Belgrade: Koniglich Serbische Acad-emie.
Miller KG, Fairbanks RG, and Mountain GS (1987)Tertiary oxygen isotope synthesis, sea level history,and continental margin erosion. Paleoceanography2:1–19.
Miller KG, Mountain GS, et al. (1996) Drilling anddating New Jersey Oligocene-Miocene sequences: Icevolume, global sea level, and Exxon records. Science271:1092–1095.
Monahan CM (1996) New zooarchaeological data fromBed II, Olduvai Gorge, Tanzania: Implications forhominid behavior in the Early Pleistocene. J. Hum.Evol. 31:93–128.
Morwood MJ, O’Sullivan PB, Aziz F, and Raza A (1998)Fission-track ages of stone tools and fossils on the eastIndonesian island of Flores. Nature 392:173–176.
Movius H (1969) Lower Paleolithic archaeology in south-ern Asia and the Far East. In WW Howells (ed.): EarlyMan in the Far East. Studies in Physical Anthropol-ogy No. 1. New York: Humanities Press, pp. 17–77.
Ohman JC, Krochta TJ, Lovejoy CO, Mensforth RP, andLatimer B (1997) Cortical bone distribution in thefemoral neck of hominoids: Implications for the locomo-tion of Australopithecus afarensis. Am. J. Phys. Anthro-pol. 104:117–131.
Owen RB, and Renaut R (1981) Paleoenvironments andsedimentology of the Middle Pleistocene OlorgesailieFormation, southern Kenya rift valley. In J Coetzeeand EM van Zinderen Bakker (eds.): Palaeoecology ofAfrica. Rotterdam: Balkema, pp. 147–174.
Pinker S (1994) The Language Instinct. New York:Morrow.
Plummer TW, and Bishop LC (1994) Hominid paleoecol-ogy at Olduvai Gorge, Tanzania as indicated by ante-lope remains. J. Hum. Evol. 27:47–75.
Potts R (1988) Early Hominid Activities at Olduvai. NewYork: Aldine de Gruyter.
134 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998

Potts R (1989) Olorgesailie: New excavations and find-ings in Early and Middle Pleistocene contexts, south-ern Kenya rift valley. J. Hum. Evol. 18:477–484.
Potts R (1994) Variables vs. models of early Pleistocenehominid land use. J. Hum. Evol. 27:7–24.
Potts R (1996a) Humanity’s Descent: The Consequencesof Ecological Instability. New York: Morrow.
Potts R (1996b) Evolution and climate variability. Sci-ence 273:922–923.
Potts R (in press a) Variability selection in hominidevolution. Evol. Anthropol.
Potts R (in press b) Behavioral response to variablePleistocene landscapes. In A Nowell (ed.): The Archae-ology of Intelligence. New York: Cambridge Univer-sity Press.
Potts R, Ditchfield P, Hicks J, and A Deino (1997)Paleoenvironments of late Miocene and early Pliocenestrata of Kanam, western Kenya. Am. J. Phys. Anthro-pol. 24(Suppl.):188–189.
Potts R, Behrensmeyer AK, and Ditchfield P (in press)Paleolandscape variation in early Pleistocene homi-nid activities: Members 1 and 7, Olorgesailie Forma-tion, Kenya. J. Hum. Evol.
Prentice ML, and Denton GH (1988) The deep-seaoxygen isotope record, the global ice sheet system andhominid evolution. In FE Grine (ed.): EvolutionaryHistory of the ‘‘Robust’’Australopithecines. New York:Aldine de Gruyter, pp. 383–403.
Raymo ME, Ganley K, Carter S, Oppo DW, and McMa-nus J (1998) Millennial-scale climate instability dur-ing the early Pleistocene epoch. Nature 392:699–702.
Rayner RJ, Moon BP, and Masters JC (1993) TheMakapansgat australopithecine environment. J. Hum.Evol. 24:219–231.
Reed K (1997) Early hominid evolution and ecologicalchange through the African Plio-Pleistocene. J. Hum.Evol. 32:289–322.
Rightmire PG (1991) The Evolution of Homo erectus.Cambridge: Cambridge University Press.
Robinson JT (1954) The genera and species of theAustralopithecinae. Am. J. Phys. Anthropol. 12:181–200.
Robinson JT (1961a) The australopithecines and theirbearing on the origin of man and of stone tool-making.S. Afr. J. Sci. 57:3–13.
Robinson JT (1961b) Australopithecines and the originof man. Annu. Rept. Smiths. Instit., pp. 479–500.
Rogers MJ, Feibel CS, and Harris JWK (1994) Changingpatterns of land use by Plio-Pleistocene hominids inthe Lake Turkana Basin. J. Hum. Evol. 27:139–158.
Ruff CB (1991) Climate, body size and body shape inhominid evolution. J. Hum. Evol. 21:81–105.
Ruff CB, and Walker A (1993) Body size and body shape.In Walker A and Leakey RE (eds.): The NariokotomeHomo erectus Skeleton. Cambridge: Harvard Univer-sity Press, pp. 234–265.
Ruff CB, Trinkaus E, Walker A, and Larsen CS (1993)Postcranial robusticity in Homo. I: Temporal trendsand mechanical interpretation. Am. J. Phys. Anthro-pol. 91:21–53.
Ruff CB, Trinkaus E, and Holliday TW (1997) Body massand encephalization in Pleistocene Homo. Nature387:173–176.
Sandweiss DH, Richardson JB III, Reitz EJ, Rollins HB,and Maasch KA (1996) Geoarchaeological evidencefrom Peru for 5000 years B.P. onset of El Nino. Science273:1531–1533.
Shackleton NJ (1995) New data on the evolution ofPliocene climatic variability. In ES Vrba, GH Denton,TC Partridge, and LH Burckle (eds.): Paleoclimateand Evolution, With Emphasis on Human Origins.New Haven: Yale University Press, pp. 242–248.
Shackleton RM (1978) A geologic map of the OlorgesailieFormation. In WW Bishop (ed.): Geological Back-ground to Fossil Man. Edinburgh: Scottish AcademicPress (in pocket).
Shipman P, and Harris JM (1988) Habitat preferenceand paleoecology of Australopithecus boisei in EasternAfrica. In FE Grine (ed.): Evolutionary History of the‘‘Robust’’ Australopithecines. New York: Aldine deGruyter, pp. 343–382.
Sikes NE (1994) Early hominid habitat preferences inEast Africa: Paleosol carbon isotopic evidence. J.Hum. Evol. 27:25–45.
Sikes NE, Potts R, and Behrensmeyer AK (1997) Isoto-pic study of Pleistocene paleosols from the Olorge-sailie Formation, southern Kenya rift. J. Hum. Evol.32:A20–A21.
Sikes NE, Potts R, and Behrensmeyer AK (in press)Early Pleistocene habitat in Member 1 Olorgesailiebased on paleosol stable isotopes. J. Hum. Evol.
Stanley SM (1992) An ecological theory for the origin ofHomo. Paleobiology 18:237–257.
Stauffer B, Blunier T, Dallenbach A, Indermuhle,Schwander J, Stocker TF, Tachumi J, Chappellaz J,Raynaud D, Hammer CU, and Clausen HB (1998)Atmospheric CO2 concentration and millennial-scaleclimate change during the last glacial period. Nature392:59–62.
Stern JT Jr and Susman RL (1983) Locomotor anatomyof Australopithecus afarensi. Am. J. Phys. Anthropol.60:279–317.
Steudel KL (1994) Locomotor energetics and hominidevolution. Evol. Anthrop. 3:42–48.
Susman RL, and Stern JT (1991) Locomotor behavior ofearly hominids: Epistemology and fossil evidence. InY Coppens and B Senut (eds.): Origine(s) de la Bipediechez les Hominides. Paris: Editions du CNRS, pp.121–131.
Tauxe L, Deino AD, Behrensmeyer AK, and Potts R(1992) Pinning down the Brunhes/Matuyama andupper Jaramillo boundaries: A reconciliation of orbitaland isotopic time scales. Earth Planet. Sci. Lett.109:561–572.
Taylor KC, Lamorey GW, et al. (1993) The ‘‘flickeringswitch’’ of late Pleistocene climate change. Nature361:432–436.
Teilhard de Chardin P, Young CC, Pei WC, and ChangHC (1935) On the Cenozoic formations of Kwangsiand Kwangtung. Bull. Geolog. Soc. China 14:179–205.
Thieme H (1997) Lower Palaeolithic hunting spearsfrom Germany. Nature 385:307–310.
Tooby J, and Cosmides L (1989) Evolutionary psychol-ogy and the generation of culture, part I. Ethol.Sociobiol. 10:29–49.
Tooby J, and Cosmides L (1990) The past explains thepresent: Emotional adaptations and the structure ofancestral environments. Ethol. Sociobiol. 11:375–424.
Tooby J, and Cosmides L (1993) The psychologicalfoundations of culture. In JH Barkow, L Cosmides,and J Tooby (eds.): The Adapted Mind. New York:Oxford Press, pp. 19–136.
Trinkaus E (1981) Neanderthal limb proportions andcold adaptation. In CB Stringer (ed.): Aspects ofHuman Evolution. London: Taylor and Francis, pp.187–224.
Van Couvering JAH, and Van Couvering JA (1976) EarlyMiocene mammal fossil from East Africa. In GL Isaacand ER McCown (eds.): Human Origins: Perspectiveson Human Evolution, Vol. 3. Menlo Park, CA: WABenjamin, pp. 155–207.
Vrba ES (1975) Some evidence of chronology and pal-aeoecology of Sterkfontein, Swartkrans, and Krom-draai from the fossil Bovidae. Nature 254:301–304.
135ENVIRONMENTAL HYPOTHESES OF HOMININ EVOLUTIONPotts]

Vrba ES (1980) Evolution, species and fossils: How doeslife evolve? S. Afr. J. Sci. 76:61–84.
Vrba ES (1985) Environment and evolution: Alternativecauses of the temporal distribution of evolutionaryevents. S. Afr. J. Sci. 81:229–236.
Vrba ES (1988) Late Pliocene climatic events and homi-nid evolution. In FE Grine (ed.): Evolutionary Historyof the ‘‘Robust’’ Australopithecines. New York: Aldinede Gruyter, pp. 405–426.
Vrba ES (1989) Levels of selection and sorting withspecial reference to the species level. In PH Harveyand L Partridge (eds.): Oxford Surveys in Evolution-ary Biology, Vol. 6. Oxford: Oxford University Press,pp. 111–168.
Vrba ES (1992) Mammals as a key to evolutionarytheory. J. Mammalogy 73:1–28.
Vrba ES (1995a) On the connection between paleocli-mate and evolution. In ES Vrba, GH Denton, TCPartridge, and LH Burckle (eds.): Paleoclimate andEvolution, With Emphasis on Human Origins. NewHaven: Yale University Press, pp. 24–45.
Vrba ES (1995b) The fossil record of African antelopes(Mammalia, Bovidae) in relation to human evolutionand paleoclimate. In ES Vrba, GH Denton, TC Par-tridge, and LH Burckle (eds.): Paleoclimate and Evolu-tion, With Emphasis on Human Origins. New Haven:Yale University Press, pp. 385–424.
Vrba ES, Denton GH, and Prentice ML (1989) Climaticinfluences on early hominid behavior. Ossa 14:127–156.
Vrba ES, Denton GH, Partridge TC, and Burckle LH(1995) Paleoclimate and Evolution, With Emphasis onHuman Origins. New Haven: Yale University Press.
Walter RC, Manega PC, Hay RL, Drake RE, and CurtisGH (1991) Laser-fusion 40Ar/39Ar dating of Bed I,Olduvai Gorge, Tanzania. Nature 354:145–149.
Wang K, Zhang Y, and Jian H (1991) Migration ofpaleovegetational zone and paleoclimatic changes ofQuaternary in the coastal region of South China.Chinese Science Bulletin 36:1721–1724.
Washburn SL (1960) Tools and human evolution. Sci.Am. 203(3):63–75.
Washburn SL (1961) Social Life of Early Man. Chicago:Aldine.
Washburn SL (1963) Classification and Human Evolu-tion. Chicago: Aldine.
Washburn SL, and Dolhinow P (1972) Perspectives onHuman Evolution, Vol. 2. New York: Holt, Rinehartand Winston.
Washburn SL, and Jay PC (1968) Perspectives on Hu-man Evolution, Vol. 1. New York: Holt, Rinehart andWinston.
Washburn SL, and McCown ER (1978) Human Evolu-tion: Biosocial Perspectives. Perspectives on HumanEvolution, Vol. 4. Menlo Park, CA: Benjamin/Cum-mings.
Waters MR, Forman SL, and Pierson JM (1997) DiringYuriakh: A Lower Paleolithic site in central Siberia.Science 275:1281–1284.
Wesselman HB (1984) The Omo Micromammals: System-atics and Paleoecology of Early Man Sites from Ethio-pia. Basel: S. Karger.
Wesselman HB (1995) Of mice and almost-men: Re-gional paleoecology and human evolution in the Tur-kana Basin. In ES Vrba, GH Denton, TC Partridge,and LH Burckle (eds.): Paleoclimate and Evolution,With Emphasis on Human Origins. New Haven: YaleUniversity Press, pp. 356–368.
White TD (1995) African omnivores: Global climaticchange and Plio-Pleistocene hominids and suids. InES Vrba, GH Denton, TC Partridge, and LH Burckle(eds.): Paleoclimate and Evolution, With Emphasis onHuman Origins. New Haven: Yale University Press,pp. 369–384.
Williamson PG (1985) Evidence for an early Plio-Pleistocene rainforest expansion in East Africa. Na-ture 315:487–489.
Winkler AJ (1995) Small mammals from the earlyPliocene at Kanapoi, West Turkana, northern Kenya.J. Vert. Paleontol. 15(Supplement to no. 3):60A.
Woillard GM (1978) Grande Pile peat bog: A continuouspollen record for the last 140,000 years. Quatern. Res.9:1–21.
WoldeGabriel G, White TD, Suwa G, Renne P, deHeinzelin J, Hart WK, and Helken G (1994) Ecologicaland temporal placement of early Pliocene hominids atAramis, Ethiopia. Nature 371:330–333.
Wolpoff MH (1980) Paleoanthropology. New York: Knopf.Wood BA (1991) Koobi Fora Research Project, Vol. 4.
Oxford: Clarendon.Woodruff F, Savin SM, and Douglas RG (1981) Miocene
stable isotope record: A detailed deep Pacific Oceanstudy and its paleclimatic implications. Science 212:665–668.
Wright JD, and Miller KG (1992) Miocene stable isotopestratigraphy, Site 747, Kerguelen Plateau. Proceed-ings of the Ocean Drilling Program, Scientific Results120:855–866.
Xu Q, and Ouyang L (1982) Climatic changes duringPeking Man’s time. Acta Anthropologica Sinica 1:80–90.
Zhu Z, Wang J, et al. (1995) Red soil, loess and globalchange. Quatern. Sci. 3:267–277.
136 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 41, 1998





























