
PhysicsLettersA169 (1992)134—144 PHYSICSLETTERS ANorth-Holland
A phasetransitionin humanbrain andbehavior
J.A.S.Kelso, S.L. Bressler,S. Buchanan1, G.C.DeGuzman,M. Ding,A. Fuchs2 and T. HoiroydProgramin ComplexSystemsandBrain Sciences,Centerfor ComplexSystems, Florida Atlantic University,BocaRaton,FL 33431-0991,USA
Received29April 1992; acceptedfor publication22July 1992Communicatedby A.P. Fordy
Using acircular 37-SQUID (superconductingquantuminterferencedevice) sensorarray,weobservespontaneoustransitionsin neuromagneticfield patternsinthehumanbrainwhichoccuratacritical valueof asystematicallyvariedbehavioralparameter.Coherentstatesof both brain andbehaviorarecapturedby thespatiotemporalpatternof phaserelationsamongparticipatingcomponents.Suchobservationssupportthethesisthat thebrain is a patternformingsystemthatcan switchflexibly from onecoherentstateto another.
At the beginning of this centuryCharles Scott to behavior.During the courseof a typical behav-Sherrington,thefatherof modernneurophysiology, ioral actmultiple anddistributedcorticalregionsaredescribedthebrainasanenchantedloomwheremil- active (see,e.g.,ref. [51).In contrastto approacheslionsof flashingshuttlesweavea dissolvingpattern, that emphasizesingle,noninteractingequivalentdi-alwaysa meaningfulpatternbut neveran abiding pole sources,herewe usethe spatialandtemporalone.This beautifulimage of the brain as a pattern resolutionof the devicealongwith computermeth-formingsystemis beginningto berealized.Onerea- odsto uncoverthe patterndynamicsof thebrain,asson is technological:only recently has technology seen,e.g.,in magnitudesandphasesof spectra.providedus with toolsof sufficientspatialandtem- A secondreason why Sherrington’svision mayporalresolutiontomeasuredynamicpatternsofbrain soonberealizedis theemergenceof theoreticalcon-cellassembliesandhowtheyaregenerated(see,e.g., ceptsof patternformationandcooperativephenom-refs. [1,2]). In the presentcontributionwe present enato handlelargeensemblesof neurons(or neu-some recentresults using one suchtool, a multis- ron-like elements)along with mathematicalandensor SQUID array. SQUIDs (superconducting computationaltools to model their behavior.Co-quantuminterferencedevices) allow accessto the operativephenomenain natureare typically inde-spatiotemporalpatterningof magneticfields gener- pendentof the particular molecularmachineryoratedby intracellulardendriticcurrentflow in thehu- materialsubstratethat instantiatesthem [6,71. Wemanbrain [3,4]. Becausethe skull and scalp are pursuethe possibility thatorganizationalprinciplestransparentto magneticfields generatedinside the may lie at the level of patternsthemselvesandthatbrainandbecausethesensorarrayis largeenoughto a focuson pattern(especiallythedynamicalfeaturescovera substantialportionofhumanneocortex,this of self-organization,stability and flexibility) maynewresearchtoolopensa (noninvasive)windowinto provide the conceptualtools necessaryto advancethe brain’sspatiotemporaldynamicsandits relation our understandingof how brain and behavioral
eventsare coordinated.Ourcollectiveansatzis that
l3iomagneticTechnologies,Inc., SanDiego,CA, USA. the linkagebetweeneventsat a “microscopic” level2 Also: Institute for TheoreticalPhysicsandSynergetics,Uni- (e.g. of neuronsandneuronalensembles)andthe
versityof Stuttgart,Stuttgart,Germany. “macroscopic”behaviorallevel isby virtue of shared
134 0375-96011921$05.00© 1992 ElsevierSciencePublishersB.V. All rightsreserved.

Volume 169, number3 PHYSICSLETTERSA 21 September1992
dynamics,not necessarilybecauseanysinglelevel has herentstateto another.An interestingaspectof ourontologicalpriority overanother.Twentyyearsago resultsis thatcoherentstatesandstatetransitionsinat the endof a specialNeurosciencesResearchPro- bothbrain andbehaviorare capturedby the samegramon Dynamicpatternsof braincell assemblies, observable,namely the spatiotemporalphaserela-it wasconcludedthat... “thepossibility [our italics] tionsamongparticipatingcomponents.of waves,oscillation, macrostatesemergingout of The SQUID sensorunit (BiomagneticTechnolo-cooperativeprocesses,suddentransitions, prepat- gies, Inc.) consistsof 37 axially symmetric first-terning etc.seemmadeto orderto assistin the un- order gradiometerpick-up coils designedto rejectderstandingof integrativeprocessesof the nervous uniform fields. The long axis of the niobium wiresystemthat remain unexplainedin contemporary coils (diameter20 mm, length 50 mm) are angledneurophysiology”(ref. [8], p. 152).With somenot- relativeto eachotherandpoint to the centerof aableexceptions,however,it is only recentlythat ob- sphereof radius 120 mm. By orienting all pick-upservationsof oscillationsandresonancesat several coils nearlyperpendicularto thesubject’shead,sen-levels of the nervoussystemhaveservedto rejuve- sitivity to the brain’sweak magneticfield is opti-nateinterestin dynamicalphenomenaandtheir im- mized.Eachcoil is attachedto a SQUID, thecoreofportancefor understandinghigher brain functions which consistsof a superconductingring with two(see,e.g., ref. [9J for a review), identicalJosephsonjunctions.TheSQUIDring and
On the behavioral side, cooperativeandcritical its associatedelectronicstransforma currentchangephenomenasuch as multistable states,hysteresis, in the input coil into a voltageproportionalto thedisorder—orderand order—orderphasetransitions intensityof the magneticfield. Thelow temperaturehavebeenshownto be characteristicof perception environmentrequired to maintain these compo-[10—12], sensorimotorcoordination [13—161and nentsin their superconductingstate is providedbylearning[171. Despitethe fact thatphasetransition a reservoirof liquid helium, containedin a dewar.phenomenahavebeendiscoveredin artificial neural Thedatafrom all SQUIDSaresentvia a fiber opticnetworks(see,e.g.,refs. [18—201)andplay acentral cableto a VME-bus computerlocatedoutsidetherole in some brain theories [6,21] no direct evi- magneticallyshielded room (built of mumetal)dence,to ourknowledge,hasbeenreportedforphase wherethe dataare acquired.EachSQUID signalistransitionsor bifurcationsin the humanbrain. Al- sampledat 862 Hz andbandpassfiltered from 0.1thoughbrainactivity “states” correspondingto var- Hz (12 dB peroctave) to 100 Hz (48 dB peroc-ious sleepstages,epilepticseizuresandsoforth are tave).Before theexperimentbegan,instrumentandwell-known,few, if any,experimentsexistthatstudy environmentalnoisemeasurementswere takenandbothbrainandbehaviorastheyevolvein timeunder foundto belessthan 10 if/\/ii~(fT = 10~tesla).the influenceof a controlparameter.To establisha Following a paradigm introduced by Kelso,phasetransition is important in such cases,even DelColleandSchöner[14] the subject’staskwastomethodologically,becausetransitionsallow a clear producea flexion movementwith the right (pre-distinctionbetweenpatternsandthusenablesus to ferred) indexfingerin responseto an auditorystim-find the dimension(s) on which patternsare de- ulus (2000Hz squarewave,100 msduration,36 dB)fined. Moreover, predictions about the dynamics deliveredthrougha loudspeakerinside the experi-(e.g.stability, lossof stability)canbe,andhavebeen, mentalchamber.Theresponseitselfwaspractically(in the caseof behavior)testednearcritical points isometric,sufficientto deforma piezoelectriccrystal(for a review, seeref. [22]). Using a circular 37- situatedundera responsekey.On eachexperimentalSQUIDarraywe reportherethat spontaneoustran- run, the frequencyof occurrenceof the auditorysitions in brain activity patternsoccur at a critical stimuluswas increasedafter 10 repetitionsin stepsvalueof a systematicallyvariedbehavioralparam- of 0.25 Hz from a startingfrequencyof 1.0Hz andeter.Ourobservationssupportthethesis[6] thatthe endingat 3.25 Hz. Instructionsemphasizedthat abrainis a self-organizing,patternformingsystemthat singleresponsebe placedbetweensuccessivestim-operatescloseto instability points,therebyallowing ulusrepetitions,i.e. thesubjectwastoperformasyn-it to switch flexibly andspontaneouslyfrom oneco- copatedmodeof coordination.Stresswas placedon
135
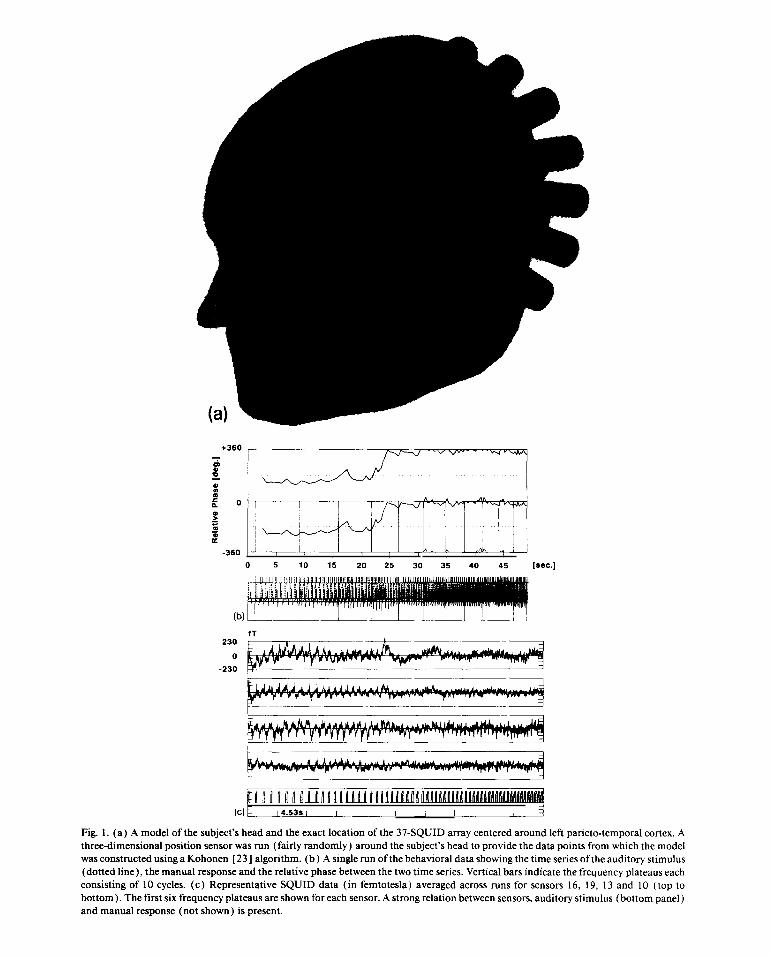
(a)
+360 ~ -~.,_dv-.. -. . ‘~.4
-36:~ L~TT
1i~P
0 5 10 15 20 25 30 35 40 45 lSec.1
fit! I! I I I11LJIfflILj~
(b) __________________________________________________
~(c)L~j4.53s~ I __j___ I I I
Fig. 1. (a) A modelof thesubject’sheadandtheexactlocationof the37-SQUIDarraycenteredaroundleft parieto-temporalcortex.Athree-dimensionalpositionsensorwasrun (fairly randomly)aroundthesubject’sheadto providethedatapointsfromwhichthemodelwasconstructedusinga Kohonen[23] algorithm.(b) A singlerunofthebehavioraldatashowingthetime seriesoftheauditorystimulus(dottedline),themanualresponseandtherelativephasebetweenthetwo timeseries.Vertical barsindicatethefrequencyplateauseachconsistingof 10 cycles. (c) RepresentativeSQUID data (in femtotesla)averagedacrossruns for sensors16, 19, 1 3 and 10 (top tobottom).Thefirst six frequencyplateausareshownfor eachsensor.A strongrelationbetweensensors,auditorystimulus(bottompanel)andmanualresponse(notshown) is present.

Volume169, number3 PHYSICSLETTERSA 21 September1992
maintaining a 1: 1 stimulus—responserelation be- theoreticallymodelledusingnonlinearlycoupledos-tweenmetronomeandfingermovement.Fortycon- cillators [14,15]. We note that someof thesefea-secutiverunswere carriedout in two blocks of 20 tures (e.g.phase-andfrequency-locking)havebeenruns,eachrun lastingabout1 mm. During theentire observedin the mammalianbrainbothwithin [27]experimentthe subjectlay in the right decubituspo- andacrosshemispheres[28], otherssuchasswitch-sition with headandnecksupportedby a specially ing at a critical parametervalue,havenot.constructedbrace.Head-basedcoordinatemeasures An exampleof the SQUID dataaveragedacrossallowedusto determinetheexactspatial locationof runsis shownin fig. 1 c alongwiththestimuluschan-theSQUIDarrayrelativeto the subject’shead.Three nel.Thedatacomefrom sensorsaroundtheanterioreasily defined anatomical landmarks (fiduciary (top) andposterior(bottom) portionsof the array.points),the left and right preauricularpointsand the Notice thatthebrainsignalsshowperiodicvariationnasion,weredigitized thusdefiningthe 3D coordi- at the stimulusfrequencyandthat the amplitudeofnatesof the headin relationto the array. Figure 1 a therecordedsignalsisgenerallylargerin theanteriorshowsa model of the subject’sheadwith respectto thanposteriorpart of the array.Fourierpowerspec-the position of the 37-SQUID array (to scale),as tra were calculatedfor the time series from eachdisplayedon a SILICON GRAPHICSworkstation. SQUID sensor.Figure 2a showsthe spectralpowerThis model was reconstructedfrom the actual3D for a time window spanningthe secondfrequencydatapointsusinga Kohonenalgorithm [231 to self- plateau(1.25Hz), i.e., beforethe behavioraltran-organizea best fit surface.Eachcylinder represents sition. On the lower left, two boxesare shown: theoneof thegradiometers,with a SQUID loop at each topbox is theFouriertransformof thestimulusandendofthecylinder. Thesensormontagewasinitially the bottom one is of the manualresponse.The fun-placedoverC3 (in theInternational10—20system). damentalfrequencyoccursat 1.25 Hz inbothboxes,Following studiesof auditory (see,e.g.,ref. [24]), coincidingwith the stimulusfrequency.Dottedlinessomatosensory(see,e.g., ref. [251)andvoluntary on all the remainingboxes, which representthemovement-relatedfields (see,e.g.,ref. [261), thear- SQUIDarray, correspondtothe majorFouriercorn-raywas moved2 cm posteriorlyto optimize record- ponentsof the stimulus.Obviously, thereis a dom-ing from sensoryandmotorbrainareas,thusplacing inant peak at the stimulus (and response) fre-the array approximatelyover left parieto-temporal quency,strongeston the anteriorpart of the array.cortex. On thefirst two plateaus,thedominantfrequencyof
A singlerun ofthebehavioraldatais shownin fig. thebrainsignalstrackedthestimulusfrequencyper-lb which plotsthe time seriesof the auditorystirn- fectly. On the third andfourth frequencyplateaus,ulus (dottedline), themanualresponse,which isbi- severalfrequencycomponentsemergedand powerphasicdue to the closureandreleaseof the switch, becamemorediffuse acrossthe spectrum.In fig. 2bandthe relativephasebetweenthe two time series, we show the power spectraon the fifth frequencyThenotablefeatureisthataroundtheendofthethird plateau.Clearly, after the behavioraltransition isfrequencyplateau(1.75Hz) therelativephasestarts over, the frequencyof the brain signalsonceagainto changeabruptly from a nearanti-phase,synco- tracksthe stimulus.patedmodeof coordinationto a nearin-phase,syn- Althoughthepowerspectrain figs. 2aand2b showchronizedmodeof coordination.In actual fact, in a remarkabletemporalcoherencefor most of thethelattercasethe manualresponseactuallyprecedes SQUID sensors,singlepower spectracannotrevealtheauditorystimulusby asmallamount.Thisspon- theinterrelationsamongcortical regionsbeneaththetaneouschangefroma syncopated,off-beatto anan- array. Wethereforecomputedcross-powerandphaseticipatory,synchronizedmodeof coordinationat a betweeneachsensorand theensemblemeanoverthecritical frequencyhasbeenobservedbehaviorallyin entirearray. Figure 2c showstopographicmaps ofsevenOthersubjects [141. Stable phase-and fre- cross-powerfrom 4 softherestperiodbetweenruns,quency-synchronizedpatterns,shifts from anti- to andthe first 4 s of the first six frequencyplateaus.in-phasemodes,hysteresisandoccasionallossof en- In therestperiodthespectralpeakmappedis at 1.25trainmenthavealsobeenfoundin thisparadigmand Hz, probablyreflectingtheheartrate.This interpre-
137

_______________ 33 34 ________________
~s2L~Lj
I __________ A~4JJL1i
__ _ ~ ___
~LiLJ[kLJiii I __ ___
I I A ______
Plateau2
_ ~(a) _________
12345f 12348 /1.234.5. 1.2.3.4.5. 1.2.3.4.5.
_______________ 33 . .
I
__ ~LLi L :i~~ ~~L: LL~
~i:.2L~C ‘.LJL~2’
A _________ Lt,~&J~—~i LLL5-~~J__ __ Plateau5(b) ~ .- 2,5 5.0 7,6 / . -. 2.5 5.0 7.5 /
2.5 5.0 73 / 25 5.0 4 5.0 4 /
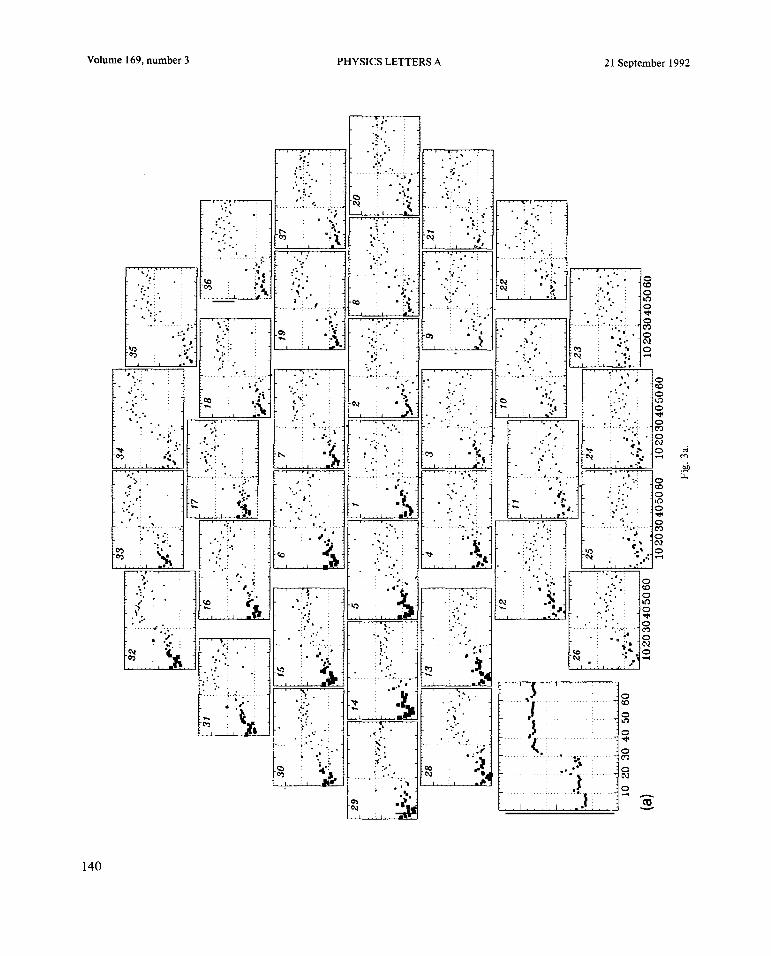
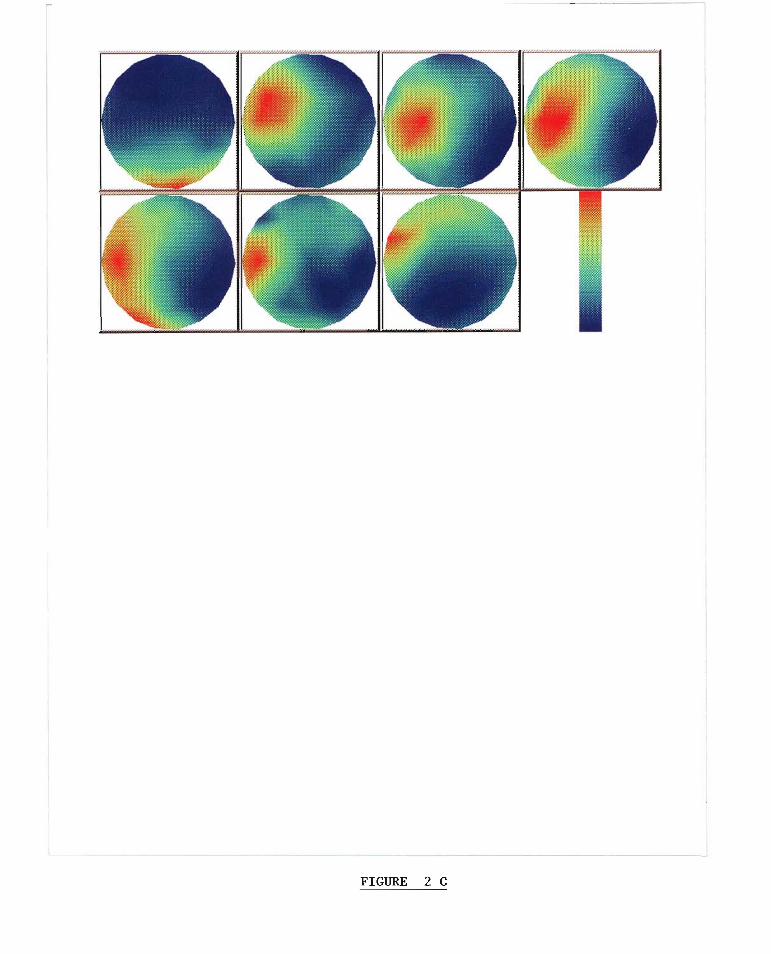
Fig. 2. (a) Powerspectrafor eachsensorin theentire SQUID array calculatedon the secondfrequencyplateau(1.25 Hz), i.e. pre-transition. (b) Powerspectrafor eachsensorcalculatedon thefifth frequencyplateau(2.0Hz), i.e. post-transition.(c) Topographicdistributionsof cross-powerfor therestperiod(upperleft) andthefirst six frequencyplateaus(left to right). Eachframeplotsthevalueof cross-powerfor eachsensoratthedominantpeakofthecross-spectrum,correspondingto theheartratefor therestperiod,andto thestimulusfrequencyfor thefrequencyplateaus.Thespatialmaximumof cross-powerin theanteriorportion ofthearray(absentfor therestperiod) shifts positiononly slightly from oneplateauto thenext.The cross-spectrumateachsensorwas computedbetweentheaveragewaveformof thatsensorandthegrandaveragewaveformcomputedoverall thesensors,wherethewaveformspanned4 sof therestperiodorthefirst4 sof thefrequencyplateaus.Eachdistributionhasbeennormalizedsothattherangeof valuesfrom minimumtomaximumcorrespondsto thefull colorscaleshown.

Volume169, number3 PHYSICSLETTERSA 21 September1992
,. .:-. .
(cil
Fig. 2. Continued.
tation is supportedby the maximumat the inferior abrupt increasein phaselead with respectto theedgeof the arrayandthe uniform fall-off in the su- stimulus.The observedtransitionis spatial as wellperior direction. In the first threefrequencypla- as temporal:scanningleft to right from sensor30 toteaus,there is an anteriormaximum in the cross- 37 the relativephasechangesfrom linear (30, 15)power,correspondingto thelocusof maximal power to diffuse (6) to nonlinear(7, 19,37). Similareventsin fig. 2a.On plateau4, theregionof maximalcross- are seenin adjacentrows. It is worth noting that apoweris extendedtocovermuchof theanteriorpor- significant increasein phaselag occursat the firsttion of the array. Following the transition,the max- cycle of eachplateau,andthat the phaseoftendriftsimum shrinksback to the approximatepre-transi- upwardandfluctuates(e.g. sensors6, 19, 37, 20)tionlocus,butwith thedistributionof elevatedpower before the switch, a typical sign of nonequilibriumshifted towardthe superiorpart of the array. phasetransitions.
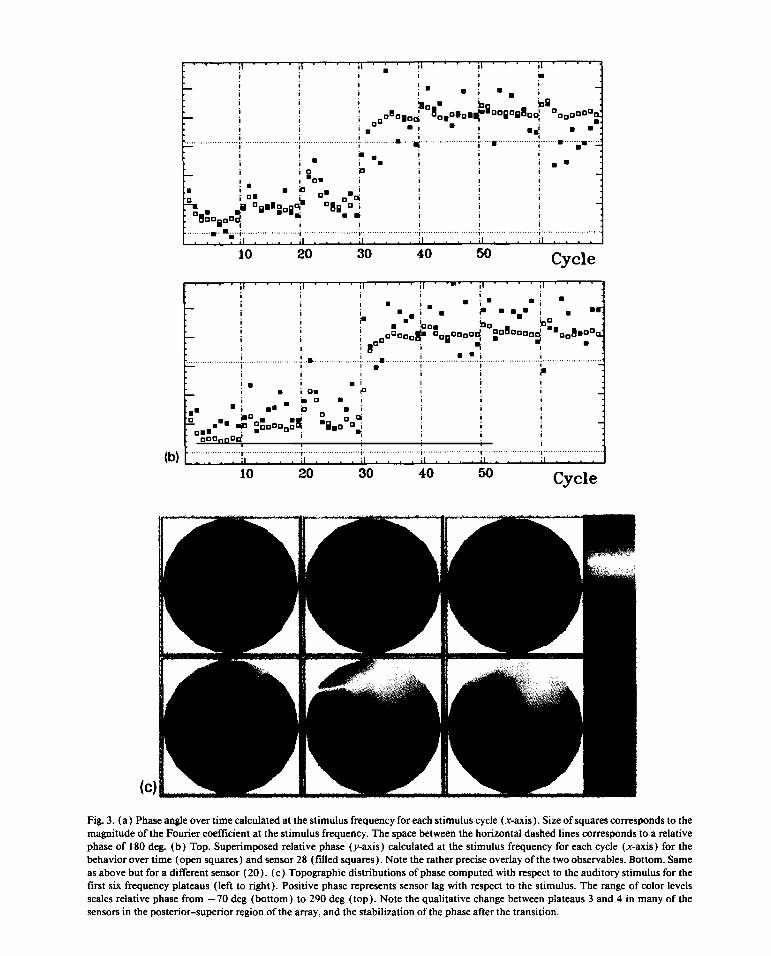
In fig. 3awe plot therelativephaseof eachsensor Predictedeffectsof systemsapproachinginstabil-with respectto the stimulusovertime for eachcycle ity such as critical fluctuationsandcritical slowingat the stimulusfrequency.The size of the squares down [6,15,29—31]canbe nicely seenby superim-correspondsto the powerof the Fouriercomponent. posingthe relativephasetimeseriesof the behaviorOn thelowerleft largerbox, the relativephaseofthe (opensquares)andtwo representativeSQUID sen-averagedresponsewith respectto the stimulus is sors (solidsquares).As shownin fig. 3b, theSQUIDplotted.In general,anabruptchangein coordinative data,as expected,are a little noisierthanthebehav-modeoccursneartheendofthethird frequencypla- ioral data. Nevertheless,it is easyto seethat bothteau,similar to the individual run shownin fig. lb. brain andbehaviorare perturbedmoreby the sameEvenmorestrikingare the relativephasedatafrom magnitudeof perturbation(a parameterchangeofthe SQUID sensorsthemselves.Over much of the 0.25 Hz) asthecritical pointapproaches.Moreover,sensorarraya remarkablechangein the phasere- critical slowing down is indicatedby the fact that itlation occurs. In certain posteriorsensors,for ex- takeslonger to returnto the pre-perturbationrela-ample,20,37 and36 the transitionparallelsthebe- tive phasevaluenearthetransition.Suchlossof sta-havioraltransition,i.e. at aboutcycle 25 thereis an bility appearsto bethe mainfactorunderlyingqual-
139
Volume 169,number3 PHYSICSLETTERSA 21 September1992
_____ ~ ~ ____
c
C’) C’,~
C’) ~, .~
• . . I I ~‘.• • ~ ‘.. .— ‘ .
1 ‘~ ‘ ,.~ ‘: , ~I~’’~ 0I ‘S:, . .1•
. ..~ .,.....~... ....... 0
• ., . . • 0S ‘~‘
14) •.. .. . . ____________________ -. . . I C’) ‘b 0C’) • ‘:‘ :‘ . . C’,~
0COI... ~ ..,
a C\~ • . •: - :~ • :.
_ _ _ _I
‘-. :~ _.~1! ~‘. .C’)
I, .
• . -.. ‘S. ._#‘ 0•-~ • ...t•% C”)
~ , - ~ •.“ .. .
I ..~ ~ 3 0
.-~ ..1.......L~ ~
140
‘‘‘‘II.’’’
— I i• • U.
I ~an
— : ~°0~Pl~ ~a.O!Q.~00U0~!aa~ aDaaoaa~
lila .~ U •~ . U
..f ..... .~. :—U • I i U.
0 I I -
— •0.• I• . C • •Io ~ a~ --. ~ ~ 0~ ~°~0D~O0d U I
________________________ I.
10 20 30 40 50 Cycle1
i • UI
— U ~ UU~ •I .• . . ~DD
U ~o. a • ~.•— ~ o~oo
0od 00 ODoa~ 00~UO°~i!0 .~ U
U— I I I •
I I I•i I
. iou— I • •D U i I
•. UI •U C Uia • . .& 0 I
— • ~ ~oo00~o~ •~Uo ~ ia.(b) :“~~:i~~ ~
10 20 30 40 50 Cycle
I I
F _(c)1 1 1.Fig.3. (a) Phaseangleovertimecalculatedat thestimulusfrequencyfor eachstimuluscycle (x-axis).Sizeof squarescorrespondsto themagnitudeoftheFouriercoefficientatthestimulusfrequency.Thespacebetweenthehorizontaldashedlines correspondsto a relativephaseof 180 deg. (b) Top.Superimposedrelativephase(y-axis)calculatedat the stimulusfrequencyfor eachcycle (x-axis) for thebehaviorover time (opensquares)andsensor28 (filled squares).Note theratherpreciseoverlayof thetwo observables.Bottom. Sameasabovebut for a differentsensor(20). (c) Topographicdistributionsof phasecomputedwith respectto theauditorystimulusfor thefirst sijC frequencyplateaus(left to right). Positivephaserepresentssensorlag with respectto thestimulus. The rangeof color levelsscalesrelativephasefrom —70 deg (bottom) to 290 deg (top).Note thequalitativechangebetweenplateaus3 and 4 in manyof thesensorsin theposterior—superiorregionof thearray,andthestabilizationofthephaseafterthetransition.
Volume 169, number3 PHYSICSLETTERSA 21 September1992
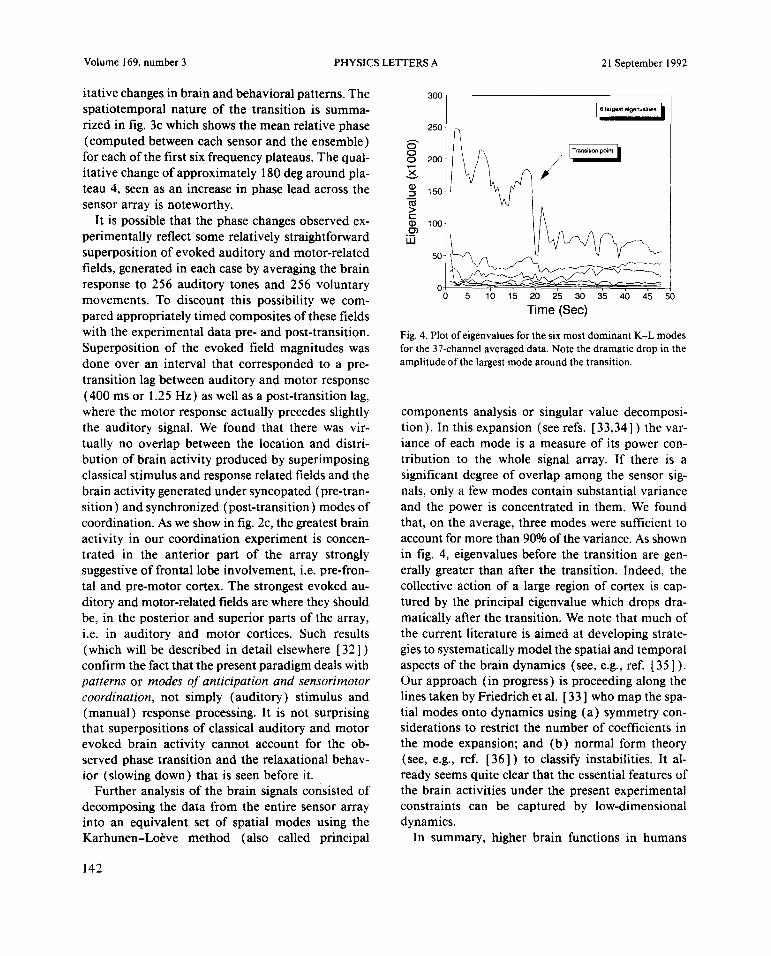
itativechangesin brainandbehavioralpatterns.The 300
spatiotemporalnatureof the transition is summa- 6 ~9e~t e~envaI~e,
rizedin fig. 3c which showsthe meanrelativephase 250
(computedbetweeneachsensorand theensemble)for eachofthe first sixfrequencyplateaus.The qual- 200 / Tr6fl6thon pOIflI
itative changeof approximately180 degaroundpla- ~.
teau4, seenas an increasein phaseleadacrossthe i~csensorarrayis noteworthy.
It is possiblethat the phasechangesobservedex- 100
~r~ystraightfbrwafields, generatedin eachcaseby averagingthe brainresponseto 256 auditory tones and 256 voluntary omovements.To discount this possibility we corn- 0 5 10 15 20 25 30 35 40 45 50
paredappropriatelytimedcompositesof thesefields Time (Sec)with the experimentaldatapre- andpost-transition. Fig.4. Plotof eigenvaluesfor thesix mostdominantK—L modes
Superpositionof the evokedfield magnitudeswas for the37-channelaverageddata.Notethedramaticdropin the
doneover an interval that correspondedto a pre- amplitudeof thelargestmodearoundthetransition.
transitionlag betweenauditoryandmotor response(400ms or 1.25 Hz) as well asa post-transitionlag,wherethe motor responseactuallyprecedesslightly componentsanalysisor singularvalue decomposi-the auditory signal. We found that therewas vir- tion). In thisexpansion(seerefs. [33,34]) thevar-tually no overlap betweenthe location and distri- ianceof eachmode is a measureof its power con-butionof brainactivity producedby superimposing tribution to the whole signal array. If there is aclassicalstimulusandresponserelatedfields andthe significant degreeof overlapamongthe sensorsig-brainactivity generatedundersyncopated(pre-tran- nals,only a few modescontainsubstantialvariancesition)andsynchronized(post-transition)modesof andthe poweris concentratedin them. We foundcoordination.As weshowin fig. 2c, thegreatestbrain that, on the average,threemodesweresufficient toactivity in our coordinationexperimentis concen- accountfor morethan90%of thevariance.As showntrated in the anterior part of the array strongly in fig. 4, eigenvaluesbefore the transitionare gen-suggestiveof frontal lobe involvement,i.e. pre-fron- erallygreaterthanafter the transition. Indeed, thetal andpre-motorcortex.The strongestevokedau- collective actionof a largeregion of cortexis cap-ditory andmotor-relatedfieldsarewheretheyshould tured by the principal eigenvaluewhich drops dra-be, in the posteriorandsuperiorpartsof the array, matically afterthe transition.We note thatmuch ofi.e. in auditory and motor cortices. Such results the current literatureis aimedat developingstrate-(which will be describedin detailelsewhere[321) giestosystematicallymodelthespatialandtemporalconfirmthefactthat thepresentparadigmdealswith aspectsof the brain dynamics(see,e.g.,ref. [35]).patternsor modesof anticipationand sensorimotor Our approach(in progress)is proceedingalongthecoordination, not simply (auditory) stimulusand linestakenby Friedrichet al. [33] who map thespa-(manual) responseprocessing.It is not surprising tial modesontodynamicsusing(a) symmetrycon-that superpositionsof classicalauditoryandmotor siderationsto restrictthe numberof coefficientsinevokedbrain activity cannotaccountfor the ob- the mode expansion;and (b) normal form theoryservedphasetransitionandthe relaxationalbehav- (see,e.g., ref. [361) to classify instabilities. It a!-ior (slowingdown) that is seenbefore it. readyseemsquite clearthat the essentialfeaturesof
Furtheranalysisof the brain signalsconsistedof the brain activities underthe presentexperimentaldecomposingthe datafrom the entiresensorarray constraints can be capturedby low-dimensionalinto an equivalentset of spatial modesusing the dynamics.Karhunen—Loève method (also called principal In summary,higherbrain functions in humans
142

Volume 169, number3 PHYSICS LETTERS A 21 September1992
suchasperception,learningandgoaldirectedmove- [5] L.P. Nunez, Electroencephalogr.Clin. Neurophysiol. 63
mentare often hypothesizedto dependon the col- (1986)75.
lective dynamics of large numbersof interacting [6] H. Haken,in: Synergeticsof thebrain,eds. E. Basar,H.Flohr,H.HakenandA.J. Mandell (Springer,Berlin, 1983)
neuronsdistributedthroughoutthe cortex.But typ-pp. 3—25.
ical signsof cooperativephenomenaare not acces- [7] G.Nicolis andI. Prigogine,Exploringcomplexity (Freeman,
sible through single neuroninvestigations.On the SanFrancisco,1989).
otherhand,from studiesof nonequilibriumsystems [8) A.K. Katchalsky,V. RowlandandR. Blumenthal,Neurosci.
it is well known that at critical points, spatialand ProVamBull. 12 (1974)1.[9] M. Barinaga,Science249 (1990)856.
temporalpatternsform in a so-calledself-organized [10] H.S. Hock, J.A.S. KelsoandG. Schöner,J. Exp. Psychol.
fashion [6]. A striking result of our experimentis Hum. Percept.Perf., in press.
thenearinstantaneousonsetof temporalandspatial [11] B. Julesz,Am. Sci. 62 (1974) 32.
coherencein the brain’smagneticfield, distributed [12] D.Williams, G. PhillipsandR. Sekuler,Nature324 (1986)
overthe entireSQUIDarraywhena meaningfulbe- 253.havioraleventis introduced.Suchcoherentstates, [13] J.A.S.Kelso,Am. J.Physiol.Reg.Int. Comput.Physiol. 15(1984)Rl000.seenas spatiallydistributedspectralpeaks,mustbe [14) J.A.S. Kelso,J.D.DelColleandG. Schöner,in: Attentionproducedby the cooperativeparticipationof very and performanceXIII, ed. M. Jeannerod(Erlbaum,
many neurons[37,38J. Our dataalso provideevi- Hillsdale, 1990)pp. 139—169.
dencefor cortical order—ordertransitionsas a be- [15) H. Haken,J.A.S. Kelso and H. Bunz, Biol. Cybern. 51(1985)347.
haviorallymanipulatedcontrolparameteris system- [161R.C.Schmidt,C. CarelloandM.T. Turvey, J. Exp.Psychol.
aticallyvaried.A spontaneousswitchin sensorimotor Hum. Percept.Perf. 16 (1990)227.
coordinationisdirectlymirroredby relativephasing [17] P.G. Zanone and J.A.S. Kelso, J. Exp. Psychol. Hum.
shiftsin partsof the SQUIDarray. Much recentat- Percept.Perf.,in press.
tentionhas focussedon synchronizedneuronalen- [18) J. Chover, in: Neural informationprocessingsystems,ed.D. Anderson (AmericanInstitute of Physics,New York,
sembleoscillationsand their potential significance 1988)pp. 192—200.
for understandingbrain function [39]. By demon- [191J.A. ReggiaandM. Edwards,NeuralComput.2 (1990)423.strating thatswitching amongrelativephasingpat- [20] J. Shrager,T. HoggandB. Huberman,Science235 (1987)
ternstakesthe form of a critical instability, our re- 1092.
sults suggest: (a) a bifurcation mechanismfor [21] V.1. Kryukov, in: Advancesin neuralinformationprocessingsystems1, ed. D.S. Touretzky(Morgan Kauffmann,Los
flexibility that is commonto bothbrainandbehav- Mtos, CA, 1989)pp. 610—617.
ior; and(b) a phasetransitionprinciplefor thecol- [22] 0. SchönerandJ.A.S.Kelso,Science239 (1988)1513.
lectiveactionof corticalneurons. [2~] T.Kohonen,Self-organizationandassociativememory,2ndEd. (Springer,Berlin, 1987).
Researchsupportedby NIMH (Neurosciences [24] M. Reite,J. Edich,J.T. ZimmermanandJ.E. Zimmerman,Electroencephalogr.Clin. Neurophysiol.45 (1978) 114.
ResearchBranch) Grant MH 42900, BRS Grant [25) R. HanandE.Kaukoranta,Prog.Neurosci.24 (1985)233.
RR07258and the U.S. Office of Naval Research [26] L. Deecke,Rev.Neurol. (Paris) 146 (1990)612.ContractN0001 4-88-J-119. [27] C.M. Gray,P. Konig, A.K. EngelandW. Singer,Nature338
(1989)334.[28) A.K. Engel,P. Konig, A.K. Kreiter andW. Singer,Science
252 (1991) 1177.References
[29] J.A.S.Kelso,J.P.ScholzandG.Schoner,Phys.Lett.A 118(1986)279.
[1] N.C. Andreasen,Science239 (1988) 1381. [30] G. SchOner,H. HakenandJ.A.S. Kelso, Biol. Cybern.53[2]P.S. Churchland,and T.J. Sejnowski,Science242 (1988) (1986)442.
741. [31] J.P.Scholz,J.A.S.KelsoandG. Schdner,Phys.Lett. A 123[3) Y. Okada,SJ.WilliamsonandL. Kaufman,Int.J.Neurosci. (1987)390.
17 (1982) 33. [32] A. Fuchs,J.A.S.KelsoandH.Haken,mt. J.Bifurc. Chaos,[4] S.J. Williamson and L. Kaufman, in: Handbook of to appear.
electroencephalographyandclinical neurophysiology,eds. [33) R. Friedrich,A. Fuchsand H. Haken,in: SynergeticsofA.S. GevinsandA. Redmond(Elsevier,Amsterdam,1987) rhythms, eds. H. Haken and HP. Köpchen (Springer,pp. 405—448. Berlin), in press.
143

Volume 169,number3 PHYSICSLETTERSA 21 September1992
[34] A. Fuchs,R. Friedrich, H. Hakenand D. Lehmann, in: [37] E. Basar,Am. J.Physiol. 245 (1983)R510.Computationalsystems:naturalandartificial, ed. H. Haken [381 E.R.John,in: Springerseriesin braindynamics,Vol. 1, ed.(Springer,Berlin, 1987)pp. 74—83. E. Basar (Springer,Berlin, 1988)pp.368—377.
1351 A. DestexheandA. Babloyantz,NeuralComput.3 (1991) [39] S. Bressler,TrendsNeurosci. 13 (1990) 161.145.
[36] VI. Arnold, Geometricalmethodsin thetheoryof ordinarydifferentialequations(Springer,Berlin, 1983).
144


Recommended

















