
of May 18, 2015.This information is current as
) A1-GP-G2-G4-ELocus (del Common Deletion of the Ig Heavy Chain A Recurrent Breakpoint in the Most
Bottaro and Mario DeMarchiAlfredo Brusco, Silvia Saviozzi, Fernanda Cinque, Andrea
http://www.jimmunol.org/content/163/8/43921999; 163:4392-4398; ;J Immunol
Referenceshttp://www.jimmunol.org/content/163/8/4392.full#ref-list-1
, 11 of which you can access for free at: cites 49 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

A Recurrent Breakpoint in the Most Common Deletion of theIg Heavy Chain Locus (delA1-GP-G2-G4-E)1,2
Alfredo Brusco,3* Silvia Saviozzi,* Fernanda Cinque,* Andrea Bottaro,† and Mario DeMarchi ‡
Human Ig heavy chain constant regions are encoded by a cluster of genes, theIGHC locus, on 14q32.3. Several forms ofIGHCdeletions and duplications spanning one to five genes have been described in different populations, with frequencies of 1.5–3.5%and 4.5–44%, respectively. Despite the common occurrence of these gene rearrangements, little is known about the breakpointsites; evidence obtained from deletions in theIGHC locus and in other regions of the human genome suggests that they prefer-entially occur in highly homologous regions and might be favored by a variety of recombinogenic signals. We present here adetailed study of three homozygotes for the most common type ofIGHC multiple gene deletion, spanning theA1-GP-G2-G4-Egenes. Using a combination of Southern blotting, long-range PCR, and automated sequencing, the unequal crossover events of allof the six studied haplotypes have been mapped to a region of;2 kb with almost complete homology betweenEP1-A1and E-A2,flanked by two minisatellites. These results are consistent with the hypothesis that segments of complete homology may be requiredfor efficient homologous recombination in humans. The possible role of minisatellites as recombination signals is inferred, inagreement with current knowledge. The Journal of Immunology,1999, 163: 4392–4398.
T he human Ig heavy chain constant region locus (IGHC)4
is a multigene family composed of 11 genes, in the orderM, D/G3,G1,EP1,A1/GP/G2,G4,E, A2on chromosome
14q32.3 (1–4). In humans, the locus is very polymorphic bothat the protein (Gm and Am allotypes) and DNA (i.e., restrictionfragment length polymorphism (RFLP), switch regions) level. Itsconsiderable genetic variation is also highlighted by the highfrequency of haplotypes carrying gene deletions or duplications(4–21). An extreme example is theG4 gene duplication, which ispresent in Caucasians with a gene frequency of;45% (22). Seventypes of deletions and 10 types of duplications have been reportedso far, spanning from 20 to 150 kb. The most frequent deletion/duplication encompasses theA1, GP, G2, G4, andE genes (;150kb). Analysis of the overall structure of the deleted/duplicated hap-lotypes suggests that they probably originated through unequalcrossing-over during meiosis, due to mispairing between highlyhomologous regions, especially of the two largeG-G-E-Aclusters.
Haplotypes with deletions or duplications are present with avariable frequency in all populations analyzed thus far, with a sig-nificantly higher frequency of duplications over deletions (Refs. 20and 21 and the references therein). According to the unequal cross-over model, duplications and deletions should originate at the samerate. Therefore, the higher frequency of duplicated vs deleted hap-lotypes suggests that selective pressures may be operating on thelocus to eliminate deletions and/or favor duplications.
Our understanding of the origins of this remarkable genetic in-stability and our interpretation of the role that selection may playin determining the distribution of the variant haplotypes in thepopulation depend upon establishing the molecular mechanismsthat lead to the individual recombination events. So far, the re-combination sites of only two deletions have been mapped. Thefirst study, based on a Southern blot approach followed by libraryconstruction and sequencing, succeeded in locating the breakpointof aG1-G4gene deletion to a highly homologous region located 39of G3 andG4, outside of the switch sequences (23). More limitedanalyses of anA1-E deletion mapped the breakpoint to a largeregion between theE genes and the switchA regions (24, 25).
In this report, we use Southern blotting, long-range PCR, andsequencing to define the deletion breakpoint in three individualshomozygous for a deletion fromA1 to E, which is the most com-mon type of deletion in the Italian population (20).
Materials and MethodsSubjects
Peripheral blood samples were obtained from three unrelated subjects(BON, DEM, and MON) and 10 controls from different peninsular Italianregions. These three subjects are homozygous for a deletion of;150 kbencompassing theA1, GP, G2, G4, andE genes. Analysis ofIGHC RFLPmarkers and Gm allotypes suggests a different origin of the deleted hap-lotypes in MON and DEM (10, 13); BON is homozygous for all testedmarkers (26). Informed consent was obtained from all subjects.
Southern blot analysis
A total of 5 mg of genomic DNA, extracted using standard phenol/chlo-roform methods (27), was digested withSacI,PstI, PvuII, andSacI1PstIrestriction endonucleases (Roche Diagnostics, Monza, Italy) and run in0.8% TBE (0.045 M Tris-borate, 0.002 M EDTA)-agarose. DNA wastransferred onto Hybond N1 nylon filters (Amersham Pharmacia, Little
*Dipartimento di Genetica, Biologia e Biochimica, Universita di Torino, Torino, It-aly; †Departments of Medicine and Microbiology and Immunology, and Cancer Cen-ter, University of Rochester School of Medicine and Dentistry, Rochester, NY 14642;and ‡Dipartimento di Scienze Cliniche e Biologiche, Universita di Torino, Torino,Italy
Received for publication June 7, 1999. Accepted for publication August 3, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Ministero dell’Universita e della Ricerca Scientifica eTecnologica (60%).2 Electronic database information: Biocomputing Service Group, http://www.genius.embnet.dkfz-heidelberg.de:8080/menu/; Lalign 2.0 DNA analysis software,http://vega.igh.cnrs.fr/bin/lalign-guess.cgi. EMBL accession numbers are as follows:intergenic EP1-A1 region, AJ238957 and AJ238959; intergenicE-A2 region,AJ238958 and AJ238960; breakpoint region in BON, AJ238961; breakpoint region inDEM, AJ238962; breakpoint region in MON, AJ238963; 39of the 33-mer VNTR forall of the subjects, AJ238964.3 Address correspondence and reprint requests to Dr. Alfredo Brusco, Dipartimento diGenetica, Biologia e Biochimica, via Santena 19, 10126 Torino, Italy. E-mail address:[email protected] Abbreviations used in this paper: IGHC, Ig heavy chain constant region locus;RFLP, restriction fragment length polymorphism; VNTR, variable number of tandemrepeats.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Chalfont, U.K.) and hybridized to the following genomic probes usingstandard techniques:
pHSa2-2.6 is a 2.6-kbSacI/PstI fragment (see Fig. 1A) subcloned fromthe l-TOU e-a phage (kindly provided by M. P. Lefranc, University ofMontpellier II, Montpellier, France).
pHSa1-0.9 is a 0.9-kbPstI-PvuII fragment (see Fig. 1A) subclonedfrom pH-Iga11 (kindly provided by T. Honjo, Kyoto University, Kyoto,Japan).
These probes cross-hybridize with two regions betweenEP1-A1 andE-A2 genes.
Cloning and sequencing
The subcloning ofl-TOU e-a and pH-Iga11 was performed using stan-dard procedures (27). The clones obtained were sequenced using a primerwalking strategy and dye-terminator chemistry on a model 377 ABI Prismautomatic sequence analyzer (Applied Biosystems, Perkin-Elmer, Monza,Italy). Data were elaborated using the Navigator program (Perkin-Elmer)and compared with GenBank with BLAST.
PCR
To amplify the regions downstream ofEP1andE, specific forward primers886 (59-acactgggttcagctgacactacactgggttc-39) and 889 (59-ggcctcacacagc-cctccggtgtaccacag-39) were used with the same reverse primer (891, 59-tgtgtccacagatgtggctggatgctgagg-39) located 9 kb downstream of each gene.The reaction was performed using the “Expand Long Template PCR Sys-tem” (Roche Diagnostics) according to the manufacturer’s instructionswith the following cycling parameters: 2 min at 93°C; 10 cycles of 93°Cfor 10 s, 65°C for 30 s, and 68°C for 12 min; 20 cycles of 93°C for 10 s,
65°C for 30 s, and 68°C for 12 min with an increase of 20 s per cycle; anda final extension of 10 min at 68°C.
Hemi-nested PCRs of regions A (Fig. 1B; primers 886 or 889 and 399:59-agtgaccggagacccaggggcctg-39) and B (primers 562 (59-tgggcaaaacca-gaagaggcc-39) and 891) were amplified from one-fiftieth of the first PCRusing the same long-range PCR conditions described above and the fol-lowing parameters: 94°C for 2 min; 10 cycles of 94°C for 10 s, 65°C for30 s, 68°C for 6 min; 20 cycles of 94°C for 10 s, 65°C for 30 s, 68°C for6 min with a 20-s increase per cycle; and a final extension of 10 min at68°C. The two products were digested using frequent-cutting restrictionendonucleases and separated on a 3% low melting point agarose or 3:1Nusieve agarose (FMC, Rockland, ME) in 13 TBE buffer.
Restriction sites were localized on the sequence using the mapping fa-cilities at the Biocomputing Service Group (Heidelberg, Germany). Thecritical region C was amplified in three overlapping fragments, using one-fiftieth of the 9-kb product as a template. Primer pairs were: 1817 (59-ctcccaggcccctggtctccgggtcatcc-39)/1843 (59-ggggagtcagctttgtctgtgctcagt-ggg-39), 562/2265 (59-cgatggaattttctgtgcgtgcgtgaggacgcctgcc-39), and E05(59-tttctctgcccgcaccctga-39)/891.
Sequence comparison
Region C was screened for the following sequences involved in hot spotsand chromosomal rearrangements using Lalign 2.0 DNA analysis software(52): thex consensus sequence GCTGGTGG ofEscherichia coli, whichstimulates generalized recombination (28); the human hypervariable mini-satellite consensus sequences GGGCAGGARG (29) and AGAGGTGGGCAGGTGG (30); the consensus sequences for Translin (ATGCAG
FIGURE 1. Structure of the two homologousE-A subregions within the humanIGHC locus.A, theEP1-A1region and theE-A2 region. The restrictionsites are shown above, and the published sequences are shown beneath the map, with their GenBank accession numbers. The pHSa1-0.9 and pHSa2-2.6plasmid clones, used as probes, are indicated. Circled restriction sites are subregion-specific. VNTR sequences are indicated by hatched boxes. Switchaand Ia are indicated by gray boxes.B, Rationale for hemi-nested, long-range PCR. The primers are positioned with respect to the maps inA. Primers 886and 889 are specific for theEP1andE regions, respectively, whereas primers 399, 1817, 562, and 891 are common for both regions. To amplify the regionsdownstream ofEP1andE, the forward primers 886 and 889 were used in distinct reactions with the same reverse primer 891, located;9 kb downstreamof each gene. This PCR product was used in a hemi-nested reaction to obtain regions A, B, and C distinctive of theEP1-A1or theE-A2 regions.
4393The Journal of Immunology
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
and GCCCWCCT), which are found at chromosomal translocation break-points in lymphoid malignancies (31); the ATGACGT sequence present inthe recombinogenicM26 allele ofSchizosaccharomyces pombe ade6 (32);the tetranucleotide repeats TCTG4–6 and CAGG7–9 and the 289-bp con-sensus sequence identified in two recombinational hot spots of mouseMHC, between theAb 3 andAb 2 genes, and in theE b gene, respectively(33–36); and the retrotransposon long terminal repeat sequence TCATA-CACCACGCAGGGGTAGAGGACT, located at the recombinational hotspot of theE b gene in mouseMHC (37).
ResultsMapping the crossover breakpoint by Southern blotting
The structure of the two homologousEP1-A1andE-A2regions asreported in the literature (2) and integrated by present data isshown in Fig. 1A.
Southern blot analyses of BON, DEM and MON had previouslydemonstrated aBamHI-RFLP containingEP1at the 59 end of thedeleted region (i.e., absence of theBamHI site circled in Fig. 1)and aPstI-RFLP containingA2 at its 39 end (Refs. 10, 13, and 26;and data not shown); therefore, we assumed that the breakpointwas located in the intergenic region betweenEP1andA2 in all sixdeleted chromosomes.
In normal chromosomes, three restriction sites distinguish theEP1-A1from theE-A2 region (Fig. 1A): a PvuII site upstream oftheA1switch absent in the corresponding position upstream ofA2,a SacI site present only in the middle of the switchA2 sequence,and the above-citedBamHI site present within theE gene but notwithin EP1.
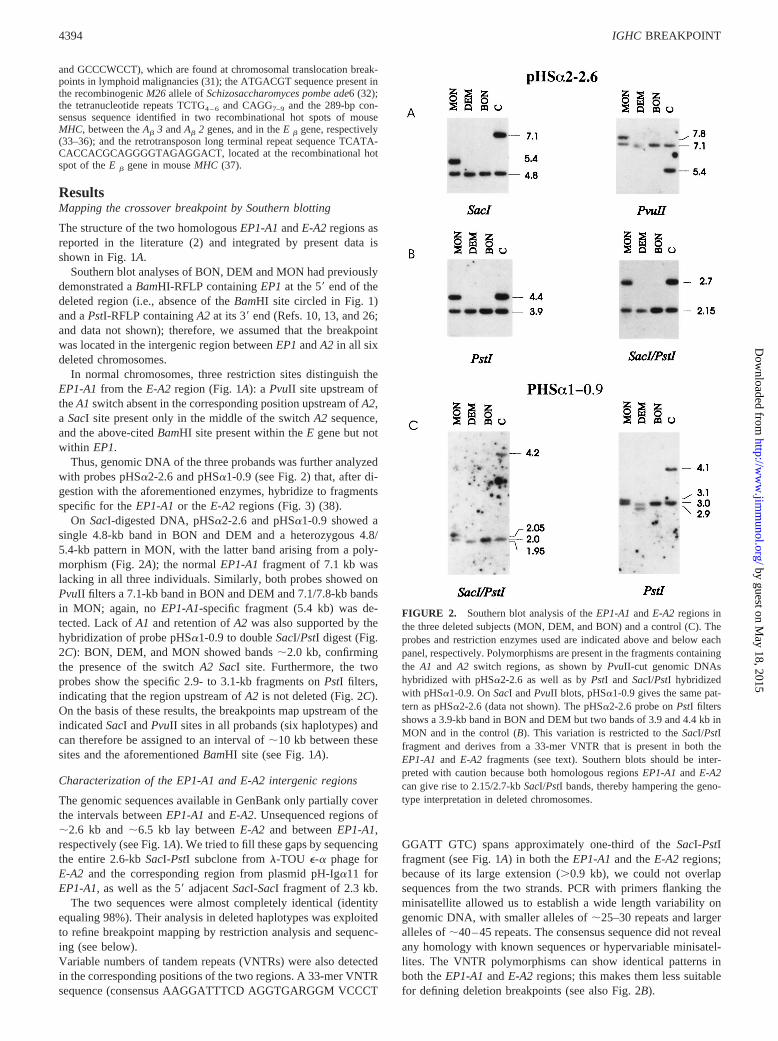
Thus, genomic DNA of the three probands was further analyzedwith probes pHSa2-2.6 and pHSa1-0.9 (see Fig. 2) that, after di-gestion with the aforementioned enzymes, hybridize to fragmentsspecific for theEP1-A1or theE-A2 regions (Fig. 3) (38).
On SacI-digested DNA, pHSa2-2.6 and pHSa1-0.9 showed asingle 4.8-kb band in BON and DEM and a heterozygous 4.8/5.4-kb pattern in MON, with the latter band arising from a poly-morphism (Fig. 2A); the normalEP1-A1fragment of 7.1 kb waslacking in all three individuals. Similarly, both probes showed onPvuII filters a 7.1-kb band in BON and DEM and 7.1/7.8-kb bandsin MON; again, noEP1-A1-specific fragment (5.4 kb) was de-tected. Lack ofA1 and retention ofA2 was also supported by thehybridization of probe pHSa1-0.9 to doubleSacI/PstI digest (Fig.2C): BON, DEM, and MON showed bands;2.0 kb, confirmingthe presence of the switchA2 SacI site. Furthermore, the twoprobes show the specific 2.9- to 3.1-kb fragments onPstI filters,indicating that the region upstream ofA2 is not deleted (Fig. 2C).On the basis of these results, the breakpoints map upstream of theindicatedSacI andPvuII sites in all probands (six haplotypes) andcan therefore be assigned to an interval of;10 kb between thesesites and the aforementionedBamHI site (see Fig. 1A).
Characterization of the EP1-A1 and E-A2 intergenic regions
The genomic sequences available in GenBank only partially coverthe intervals betweenEP1-A1andE-A2. Unsequenced regions of;2.6 kb and;6.5 kb lay betweenE-A2 and betweenEP1-A1,respectively (see Fig. 1A). We tried to fill these gaps by sequencingthe entire 2.6-kbSacI-PstI subclone froml-TOU e-a phage forE-A2 and the corresponding region from plasmid pH-Iga11 forEP1-A1, as well as the 59adjacentSacI-SacI fragment of 2.3 kb.
The two sequences were almost completely identical (identityequaling 98%). Their analysis in deleted haplotypes was exploitedto refine breakpoint mapping by restriction analysis and sequenc-ing (see below).Variable numbers of tandem repeats (VNTRs) were also detectedin the corresponding positions of the two regions. A 33-mer VNTRsequence (consensus AAGGATTTCD AGGTGARGGM VCCCT
GGATT GTC) spans approximately one-third of theSacI-PstIfragment (see Fig. 1A) in both theEP1-A1and theE-A2 regions;because of its large extension (.0.9 kb), we could not overlapsequences from the two strands. PCR with primers flanking theminisatellite allowed us to establish a wide length variability ongenomic DNA, with smaller alleles of;25–30 repeats and largeralleles of;40–45 repeats. The consensus sequence did not revealany homology with known sequences or hypervariable minisatel-lites. The VNTR polymorphisms can show identical patterns inboth theEP1-A1andE-A2 regions; this makes them less suitablefor defining deletion breakpoints (see also Fig. 2B).
FIGURE 2. Southern blot analysis of theEP1-A1andE-A2 regions inthe three deleted subjects (MON, DEM, and BON) and a control (C). Theprobes and restriction enzymes used are indicated above and below eachpanel, respectively. Polymorphisms are present in the fragments containingthe A1 and A2 switch regions, as shown byPvuII-cut genomic DNAshybridized with pHSa2-2.6 as well as byPstI andSacI/PstI hybridizedwith pHSa1-0.9. OnSacI andPvuII blots, pHSa1-0.9 gives the same pat-tern as pHSa2-2.6 (data not shown). The pHSa2-2.6 probe onPstI filtersshows a 3.9-kb band in BON and DEM but two bands of 3.9 and 4.4 kb inMON and in the control (B). This variation is restricted to theSacI/PstIfragment and derives from a 33-mer VNTR that is present in both theEP1-A1 and E-A2 fragments (see text). Southern blots should be inter-preted with caution because both homologous regionsEP1-A1and E-A2can give rise to 2.15/2.7-kbSacI/PstI bands, thereby hampering the geno-type interpretation in deleted chromosomes.
4394 IGHC BREAKPOINT
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

In addition, within the 0.5-kbSacI-PstI fragment of theSacI-SacI subclone from both theEP1-A1andE-A2regions, sequencingrevealed a VNTR with repeats of 40–41 bp (consensus TCCCTTCACC CACCCCTCCT GCAYCTATC YDCAGCCCCC A),each possessing a canonical minisatellite core (CNTCCTGCCC)and a sequence (CCCACCCC) perfectly matching Nakamura’sconsensus (CCCACNNC) (39). Two repeats also showed ax-likesequence (CCACAGC) with a single nucleotide missing from theconsensus (CCACCAGC) (28).
Restriction analysis of long-range PCR products
Most of the critical interval in which the breakpoints are locatedwas amplified by long-range PCR. Two specific forward primers(886 for EP1 or 889 for E) were designed where that the two
sequences diverge (i.e.,EP1 becomes a truncated pseudogene); acommon reverse primer (891) was tailored 9 kb downstream. Foreach region, two hemi-nested reactions (primers indicated in Fig.1B) produced fragment A (;3.5 kb) and fragment B (;4 kb).
The two amplification products from four control subjects (eighthaplotypes) were then tested with several frequent-cutting en-zymes to detect patterns specific for theEP1-A1 and theE-A2regions. Four enzymes (DdeI, AluI, MaeIII, andSmaIII) in frag-ment A and four (AluI, DdeI, NlaIII, and MaeIII) in fragment Bwere found to differentiate theEP1-A1region from theE-A2 re-gion (Fig. 4A).
In region A, both chromosomes of probands BON and MONshowed onlyEP1-A1-specific sites (Fig. 4B); proband DEM wasshown to be homozygous forEP1-A1sites withSmaI andMaeIII,but EP1-A1/E-A2was heterozygous withDdeI andHinfI, whichmight indicate a gene conversion event on one chromosome(DEM2 in Fig. 4B). In fragment B, all sites showed anE-A2-typical pattern in the three probands. Thus, the breakpoints couldbe restricted to a region comprising the two VNTRs and theirflanking regions (region C in Fig. 1), which was further analyzedby direct sequencing of overlapping PCR products.
Fine mapping of the breakpoint
The sequencing of region C in four control subjects inl-TOU e-aand in pH-Iga11 permitted us to draw the map shown in Fig. 5.Only positions differing betweenEP1-A1and E-A2 in all of thechromosomes studied are reported.
The differences used to map the breakpoint sites in deleted sub-jects are summarized in Fig. 5. The first seven base differenceslocate the breakpoint 39 of the 41-mer VNTR. Two informativebase differences are present 59of the second VNTR, whereas thereare 10 single base differences and one deletion at the 39. Thus, the
FIGURE 3. Observed alleles forA1 andA2 RFLPs. Multiple fragmentsizes within the same box indicate allelic variants.
FIGURE 4. Analysis of regions A and B amplified by hemi-nested, long-range PCR.A, The EP1-A1 or E-A2 discriminatory restriction sites areindicated.B, The results for BON, DEM, and MON are indicated below. Numbers 1 and 2 refer to individual chromosomes. Black dots indicateEP1-A1region sites, whereas white dots indicate theE-A2-associated restriction sites. In region B,DdeI andMaeIII (data not shown) display the same differencesasAluI.
4395The Journal of Immunology
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

crossover event could be mapped to the 2-kb region of high ho-mology between the two VNTRs in all subjects. The two nucleo-tide differences in the middle of the 2-kb region of homology arepolymorphic, and were not considered for breakpoint mapping.
Known signals favoring recombination were searched for in re-gion C (seeMaterials and Methods). Onex sequence and threebinding sites for Translin showed complete identity with their con-sensus; in addition, a number of sites showed a single nucleotidedifference from other putative recombinogenic sequences (Fig. 5and Table I).
DiscussionUnequal crossing-over between mispaired homologous regions isthe cause of many genetic disorders and of evolutionary variabilityin multigene families (reviewed in Ref. 40). One notable exampleis the IGHC locus, where deletions/duplications of single or mul-tiple genes reach a polymorphic frequency. Because homology isoften as elevated in intergenic regions ofIGHC as in the codingsequences, intergenic exchanges giving rise to hybrid genes arerare among recombinant haplotypes (41). The first recombinational
breakpoint within theIGHC, described by Keyeux et al. (23) in aG1-G4 homozygous deleted individual, occurred in a highly ho-mologous region, outside of the switch sequences, which the au-thors named “hsg”. These regions were subsequently shown tocontain repetitive MER-4 elements (42). Hsg homology regionswere also located downstream of eachG gene exceptG2, suggest-ing that even other deletions (e.g., ofEP1-GPandEP1-G4) couldderive from intrachromosomal recombinations between these re-gions. By analogy, theA1-E and GP-A2 deletions might arisethrough recombinations between homologous regions downstreamof the E or A genes but different from hsg. In general, it could beargued that extensive stretches of high sequence homology arenecessary for the generation of deletions and duplications in theIGHC locus.
The most common deletion/duplication, involving the genesfrom A1 to E, depends upon mispairing between twoG-G-E-Ablocks. Recently, a homozygousA1-E deletion (T17) was inde-pendently investigated by Wiebe et al. (24) and Chen et al. (25).Using a Southern blot approach, the deletion breakpoint was lo-cated in two different fragments due to the different interpretations
Table I. Homologies to recombinogenic consensus sequences in region Ca
Sequence Designation Consensus Species Sequence Foundb
1 Translin consensus GCCCWCCT Homo sapiens GCCGACCT2 Ade6sequence ATGACGT Seccaromyces pombe ATGACCT3 x sequence GCTGGTGG Escherichia coli GCTGGTGG in EP1-A1
GCTGGCGG in E-A24 Ade6sequence ATGACGT S. pombe AGGACGT5 Hypervariable minisatellite GGGCAGGARG H. sapiens GTGCAGGAGG6 Hypervariable minisatellite GGGCAGGARG H. sapiens GGGAAGGAAG7 Translin consensus ATGCAG H. sapiens ATGCAG8 x sequence GCTGGTGG E. coli GCTGATGG9 Translin consensus GCCCWCCT H. sapiens GCCCACCA
10 Translin consensus GCCCWCCT GCCCACCT rev11 Translin consensus ATGCAG H. sapiens ATGCAG12 Translin consensus GCCCWCCT H. sapiens GCCCAGCT13 Ade6sequence ATGACGT S. pombe ATGCCGT
aPosition numbering refers to Fig. 5.
b Sequence positions differing from consensus are boldface and underlined.
FIGURE 5. Sequence analysis of region C. Nucleotide differences betweenEP1-A1and E-A2 regions and polymorphisms are indicated. VNTR se-quences are shown as gray boxes. The double slash in the 33-mer VNTR indicates that it has not been completely sequenced. Numbered arrows indicatethe position and orientation of sequences homologous to known recombinogenic signals, reported in detail in Table I. The asterisk indicates a match of 29/32nucleotides, with a low complexity region in theIGLC (Ig light chain constant region) locus, pinpointed by BLAST searching (ctcattcacagacacacacgcag-gcacacactc). Below, the analysis in BON, DEM, and MON is represented as black and white dots; these dots indicate the presence of anEP1-A1or anE-A2nucleotide at a given position.
4396 IGHC BREAKPOINT
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

of the RFLP pattern. One group had assigned the breakpoint to a5.2-kbSacI/SacI fragment containing the switchA2. However, thisregion is highly polymorphic because of both the switch and the33-mer VNTR; therefore, the fragment was likely a normal poly-morphism unrelated to the deletion. Chen et al. (25) restricted thebreakpoint region to a 2.0-kbBamHI-SacI fragment just down-stream of theEP1 gene. Unfortunately, this conclusion is alsoincorrect because Southern blotting can only place the T17 break-point between the discriminatory sites reported in Fig. 1. It shouldbe noted that thePstI digestion used by these authors cannot dis-tinguish theEP1-A1from theE-A2derived fragments. Using long-range PCR, we have evidence that fragments of the same lengthcan originate from either region, likely due to the 33-mer VNTR(see Fig. 3 and the legend to Fig. 2).
Our study localizes the recombination for theA1-E deletion inthree unrelated homozygotes within the intergenic region betweenEP1/E and A1-A2 genes, which show;98% sequence identity.More precisely, the breakpoint was pinpointed in all individuals bysequencing to a;2-kb region of absolute homology (only twonucleotide differences) between two VNTRs. Furthermore onehaplotype (DEM2) also shows an intermixed restriction site pat-tern for EP1-A1 and E-A2, compatible with a gene conversionevent in proximity of the exchange site. Gene conversion eventshave been reported by several authors in association with meioticcrossing-over and other homologous recombination events. Thesefindings are in agreement with current double-strand break modelsof recombination (43).
Our data suggest that in humans, long stretches of identity favorrecombination between two sequences. Indeed, sequence homol-ogy has been demonstrated in recombinational hot spots such asthose of the mouseMHC locus (44), pseudoautosomal region ofsex chromosomes (45), hereditary neuropathy with liability topressure palsies, and Charcot-Marie-Tooth 1A disease (46).
In addition to sequence identity, other specific factors or signalsmay have targeted the recombination event between the twoG-G-E-A blocks. In fact, the same mispairing could theoretically alsogenerate other deleted/duplicated haplotypes involving theGP-A2,EP1-G4, andG1-G4genes: however, these are significantly rarer.Variable selective pressures are one potential explanation for thisdifference. However, it seems unlikely for a deletion of theA1-GP-G2-G4-Egenes to be significantly favored in terms of fitnessover one spanning, for instance,GP-G2-G4-E-A2. Alternatively,specific recombination-prone sequences betweenEP1-A1 and/orbetweenE-A2may have favored recombination in the interval. Thelow linkage disequilibrium level observed in particular within theE-A2 region (47–49) supports this model.
Repetitive DNA sequences have been proposed as preferredsites of recombination from microorganisms to humans. In theIGHC locus, the repetitive “switch” elements upstream of eachfunctional CH gene are good candidates in light of their pro-grammed involvement in somatic switch recombination. In an in-sertional mutagenesis assay, the Sm region was shown to representa recombinogenic hot spot with a 100-fold higher insertion fre-quency than the average genomic region (50).
In this respect, the two VNTRs found in the surroundings of therecombinational region may be particularly important. The con-sensus core sequence of human hypervariable minisatellites showssimilarity to theE. coli x sequence (39), a hot spot that promotesgeneralized recombination catalyzed by RecA-RecBC (51). Directexperimental evidence has also been obtained showing that mini-satellite sequences enhance the ability of a DNA molecule to un-dergo recombination at a distance, perhaps acting as recognitionsites for a specific enzyme or macromolecular complex (30).
In our study, the breakpoint critical region in all six investigatedhaplotypes lay in the same;2-kb sequence containing twoVNTRs and several consensus sequences that are known to beinvolved in recombination in humans, yeast, and bacteria. Thisstrongly suggests, although it does not directly prove, that theserecombinogenic elements may represent the site of the unequalcrossing-over, and may even coincide with an alleged recombina-tion hot spot of theIGHC locus.
AcknowledgmentsWe thank Profs. M. P. Lefranc and T. Honjo for providing clones, A.Plebani and M. G. Guerra for patients, Prof. N. Migone for criticaldiscussion, and C. Boccazzi for technical support. We are also very gratefulto Dr. Enza Ferrero for critical revision of the manuscript.
References1. Ravetch, J. V., U. Siebenlist, S. Korsmeyer, T. Waldman, and P. Leder. 1981.
Structure of the human immunoglobulinm locus: characterisation of embryonicand rearranged D and J genes.Cell 27:583.
2. Flanagan, J. G., and T. H. Rabbitts. 1982. Arrangement of the human immuno-globulin heavy chain constant region genes implies evolutionary duplication of asegment containingg, e, anda genes.Nature 300:709.
3. Lefranc, M. P., G. Lefranc, and T. H. Rabbitts. 1982. Inherited deletion of im-munoglobulin heavy chain constant region genes in normal human individuals.Nature 300:760.
4. Hofker, M. H., M. A. Walter, and D. W. Cox. 1989. Complete physical map ofthe human immunoglobulin heavy chain constant region gene complex.Proc.Natl. Acad. Sci. USA 86:5567.
5. Lefranc, M. P., G. Lefranc, G. G. DeLange, T. A. Out, P. J. van den Broek,J. van Nieuwoop, J. Radl, A. N. Helal, H. Chaabani, E. van Loghem, et al. 1983.Instability of the human immunoglobulin heavy chain constant region locus in-dicated by different inherited chromosomal deletions.Mol. Biol. Med. 1:207.
6. Migone, N., S. Oliviero, G. G. DeLange, D. L. Delacroix, D. Boschis, F. Altruda,L. Silengo, M. DeMarchi, and A. O. Carbonara. 1984. Multiple gene deletions inthe human immunoglobulin heavy chain cluster.Proc. Natl. Acad. Sci. USA 81:5811.
7. Chaabani, H., N. T. Bech-Hansen, and D. W. Cox. 1986. A multigene deletionwithin the immunoglobulin heavy chain region.Am. J. Hum. Genet. 37:1164.
8. Bech-Hansen, N. T., and D. W. Cox. 1986. Duplication of the human immuno-globulin heavy chaing2 gene.Am. J. Hum. Genet. 38:67.
9. Bottaro, A., M. DeMarchi, N. Migone, and A. O. Carbonara. 1989. Pulsed-fieldgel analysis of human immunoglobulin heavy-chain constant region gene dele-tions reveals the extent of unmapped regions within the locus.Genomics 4:505.
10. Bottaro, A., M. DeMarchi, G. G. DeLange, C. Boccazzi, F. Caldesi, R. Gallina,and A. O. Carbonara. 1989. New types of multiple and single gene deletions inthe IGHC locus.Immunogenetics 29:44.
11. Bottaro, A., M. DeMarchi, G. G. DeLange, and A. O. Carbonara. 1989. Genedeletions within the human immunoglobulin heavy chain constant region genecluster.Exp. Clin. Immunogenet. 6:55.
12. Bottaro, A., U. Cariota, G. G. DeLange, M. DeMarchi, R. Gallina, A. Vlug, andA. O. Carbonara. 1990. Multiple levels of analysis of an IgG4 gene deletion.Hum. Genet. 86:191.
13. Bottaro, A., U. Cariota, M. DeMarchi, and A. O. Carbonara. 1991. Pulsed-fieldelectrophoresis screening for the immunoglobulin heavy chain constant region(IGHC) multigene deletions and duplications.Am. J. Hum. Genet. 48:745.
14. Hendriks, R. W., M. van Tol, G. G. DeLange, and R. K. Shuurman. 1989. In-heritance of a large deletion within the human immunoglobulin heavy chainconstant region gene complex and immunological implications.Scand. J. Immu-nol. 29:535.
15. Smith, C. I. E., L. Hammarstrom, J. I. Henter, and G. G. DeLange. 1989. Mo-lecular and serological analysis of IgG1 deficiency caused by new forms of theconstant region of the Ig H chain gene deletions.J. Immunol. 142:4514.
16. Walter, M. A., C. A. Chambers, B. Zimmerman, and D. W. Cox. 1990. A mul-tigene deletion in the immunoglobulin heavy chain region in a highly atopicindividual. Hum. Genet. 85:643.
17. Olsson, P. G., M. H. Hofker, M. A. Walter, S. Smith, L. Hammarstro¨m,C. I. E. Smith, and D. W. Cox. 1991. Ig heavy chain variable and constant regiongenes in common variable immunodeficiency.J. Immunol. 147:2540.
18. Olsson, P. G., H. Rabbani, L. Hammarstrom, and C. I. E. Smith. 1993. Novelhuman immunoglobulin heavy chain constant region gene deletion haplotypescharacterised by pulsed-field electrophoresis.Clin. Exp. Immunol. 94:84.
19. Brusco, A., U. Cariota, A. Bottaro, C. Boccazzi, A. Plebani, A. G. Ugazio,R. Galanello, A. M. van Leeuwen, G. G. DeLange, S. Depelchin, et al. 1994.Structural and immunologic analysis of gene triplications in the Ig heavy chainconstant region locus.J. Immunol. 152:129.
20. Brusco, A., U. Cariota, A. Bottaro, C. Boccazzi, A. Plebani, A. G. Ugazio,R. Galanello, M. G. Guerra, and A. O. Carbonara. 1995. Variability of the im-munoglobulin heavy chain constant region locus: a population study.Hum.Genet. 95:319.
4397The Journal of Immunology
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

21. Rabbani, H., Q. Pan, N. Kondo, C. I. E. Smith, and L. Hammarstrom. 1996.Duplications and deletions of the humanIGHC locus: evolutionary implications.Immunogenetics 45:136.
22. Brusco, A., F. Cinque, S. Saviozzi, C. Boccazzi, M. DeMarchi, andA. O. Carbonara. 1997. TheG4 gene is duplicated in 44% of the human immu-noglobulin heavy chain constant region haplotypes.Hum. Genet. 100:84.
23. Keyeux, G., G. Lefranc, and M. P. Lefranc. 1989. A multigene deletion in thehuman IGH constant region locus involves highly homologous hot spots of re-combination.Genomics 5:431.
24. Wiebe, V., A. Helal, M. P. Lefranc, and G. Lefranc. 1994. Molecular analysis ofthe T17 immunoglobulin CH multigene deletion (del A1-GP-G2–G4-E).Hum.Genet. 93:520.
25. Chen, Z.-Q., M. H. Hofker, and D. W. Cox. 1995. Defining the breakpoint of amultigene deletion in the immunoglobulin heavy chain gene cluster.Immunoge-netics 41:69.
26. Bottaro, A., R. Gallina, A. Brusco, U. Cariota, C. Boccazzi, M. R. Barilaro,A. Plebani, A. G. Ugazio, A. M. van Leeuwen, G. G. DeLange, et al. 1993.Familial clustering ofIGHC deletions and duplications: functional and molecularanalysis.Immunogenetics 37:356.
27. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989.Molecular Cloning: A Lab-oratory Manual.2nd Ed. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
28. Smith, G., S. Kunes, D. Schultz, A. Taylor, and K. Triman. 1981. Structure ofxhot-spot of recombination in the murine major histocompatibility complex.Sci-ence 234:173.
29. Jeffrey, A., V. Wilson, and S. Lay Thein. 1985. Hypervariable “minisatellite”regions in human DNA.Nature 314:67.
30. Wahls, W., L. Wallace, and P. Moore. 1990. Hypervariable minisatellite DNA isa hot-spot for homologous recombination in human cells.Cell 80:95.
31. Aoki, K., K. Suzuki, T. Sugano, T. Tasaka, K. Nakahara, O. Kuge, A. Omori, andM. Kasai. 1995. A novel gene, Translin, encodes a recombination hot-spot bind-ing protein associated with chromosomal translocations.Nat. Genet. 10:167.
32. Schuchert, P., M. Langsford, E. Kaslin, and J. Kohli. 1991. A specific DNAsequence is required for high frequency of recombination in theade6gene offission yeast.EMBO J. 10:2157.
33. Kobori, J., E. Strauss, K. Minard, and L. Hood. 1986. Molecular analysis of thehot-spot of recombination in the murine major histocompatibility complex.Sci-ence 234:173.
34. Steinmetz, M., D. Stephan, and K. Fischer-Lindahl. 1986. Gene organisation andrecombinational hot-spot in the murine major histocompatibility complex.Cell50:719.
35. Uematsu, Y., H. Kiefer, R. Schulze, K. Fischer-Lindahl, and M. Steinmetz. 1986.Molecular characterisation of a meiotic recombinational hot-spot enhancing ho-mologous unequal crossing-over.EMBO J. 5:2123.
36. Shiroishi, T., N. Hanzawa, T. Sagai, M. Ishiura, T. Gojobori, M. Steinmetz, andK. Moriwaki. 1990. Recombination hot-spot specific to female meiosis in themouse major histocompatibility complex.Immunogenetics 31:79.
37. Zimmerer, E., and H. Passmore. 1991. Structural and genetic properties of the Ebrecombinational hot-spot in the mouse.Immunogenetics 33:132.
38. Keyeux, G., and J. E. Bernal. 1996. New allele variants of the immunoglobulinswitch (Sa) regions.Hum. Genet. 97:695.
39. Nakamura, Y., M. Carlson, K. Krapcho, M. Kanamori, and R. White. 1988. Newapproach for isolation of VNTR markers.Am. J. Hum. Genet. 43:854.
40. Lupski, J. R. 1998. Genomic disorders: structural features of the genome can leadto DNA rearrangements and human disease traits.Trends Genet. 14:417.
41. Natvig, J. B., and H. G. Kunkel. 1974. A hybrid IgG4-IgG2 Ig.J. Immunol.112:1277.
42. Jurka, J. 1990. Novel families of interspersed repetitive elements from the humangenome.Nucleic Acids Res. 18:137.
43. Lichten, M., and A. S. Goldman. 1995. Meiotic recombination hot-spots.Annu.Rev. Genet. 29:423.
44. Steinmetz, M., Y. Uematsu, and K. Fischer-Lindahl. 1987. Hot-spots of homol-ogous recombination in mammalian genomes.Trends Genet. 3:7.
45. Rappold, G. A. 1993. The pseudoautosomal regions of the human sex chromo-somes.Hum. Genet. 92:315.
46. Lopes, J., N. Ravise, A. Vandenberghe, F. Palau, V. Ionasescu, M. Mayer, N.Levy, N. Wood, N. Tachi, P. Bouche, et al. 1998. Fine mapping of de novoCMT1A and HNPP rearrangements within CMT1A-REPs evidences two distinctsex-dependent mechanisms and candidate sequences involved in recombination.Hum. Mol. Genet. 7:141.
47. Bech-Hansen, N. T., P. S. Linsley, and D. W. Cox. 1983. Restriction fragmentlength polymorphisms associated with immunoglobulin Cg genes reveal linkagedisequilibrium and genomic organization.Proc. Natl. Acad. Sci. USA 80:6952.
48. Migone, N., G. G. DeLange, A. Piazza, and L. L. Cavalli-Sforza. 1985. Geneticanalysis of eight linked polymorphisms within the human immunoglobulin heavychain region.Am. J. Hum. Genet. 37:1146.
49. Bottaro, A., R. Gallina, M. DeMarchi, and A. O. Carbonara. 1989. Genetic anal-ysis of new restriction fragment length polymorphisms (RFLP) in the human IgHconstant gene locus.Eur. J. Immunol. 19:2151.
50. Baar, J., and M. J. Shulman. 1995. The Ig heavy chain switch region is a hot-spotfor insertion of transfected DNA.J. Immunol. 155:1911.
51. Smith, G. R. 1983.x hot-spots of generalised recombination.Cell 34:709.
4398 IGHC BREAKPOINT
by guest on May 18, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
Recommended

















