
www.elsevier.com/locate/ijdevneu
Int. J. Devl Neuroscience 25 (2007) 133–139
Effects of maternal hyperhomocysteinemia induced by high methionine
diet on the learning and memory performance in offspring
Giyasettin Baydas a,*, Sema T. Koz a, Mehmet Tuzcu b,Victor S. Nedzvetsky c, Ebru Etem d
a Department of Physiology, Faculty of Medicine, Firat University, 23119 Elazig, Turkeyb Department of Biology, Faculty of Science, Firat University, 23119 Elazig, Turkey
c Department of Biophysics and Biochemistry, Faculty of Biology, Dnepropetrovsk National University, Dnepropetrovsk, Ukrained Department of Medical Biology, Faculty of Medicine, Firat University, 23119 Elazig, Turkey
Received 14 July 2006; received in revised form 6 February 2007; accepted 2 March 2007
Abstract
In this study, we suggest that chronic maternal hyperhomocysteinemia results in learning deficits in the offspring due to delayed brain
maturation and altered expression pattern of neural cell adhesion molecule. Although the deleterious effects of hyperhomocysteinemia were
extensively investigated in the adults, there is no clear evidence suggesting its action on the developing fetal rat brain and cognitive functions of the
offspring. Therefore, in the present work we aimed to investigate effects of maternal hyperhomocysteinemia on the fetal brain development and on
the behavior of the offspring. A group of pregnant rats received daily methionine (1 g/kg body weight) dissolved in drinking water to induce
maternal hyperhomocysteinemia, starting in the beginning of gestational day 0. The levels of glial fibrillary acidic protein, S100B protein, and
neural cell adhesion molecule were determined in the tissue samples from the pups. Learning and memory performances of the young-adult
offsprings were tested using Morris water maze test. There were significant reductions in the expressions of glial fibrillary acidic protein and S100B
protein in the brains of maternally hyperhomocysteinemic pups on postnatal day 1, suggesting that hyperhomocysteinemia delays brain maturation.
In conclusion, maternal hyperhomocysteinemia changes the expression pattern of neural cell adhesion molecule and therefore leads to an
impairment in the learning performance of the offspring.
# 2007 ISDN. Published by Elsevier Ltd. All rights reserved.
Keywords: Hyperhomocysteinemia; Cognitive functions; Brain development; NCAM; GFAP
1. Introduction
Homocysteine (Hcy) is a sulfur containing amino acid
derived from the metabolism of methionine (Clarke et al., 1991;
Reis et al., 2002). Dietary methionine contributes to a rise in
circulating homocysteine concentration (Hirche et al., 2006).
Thus, many pathogenic effects of hypermethioninemia may
be related to the metabolic link between methionine and
homocysteine. Accumulating evidence suggest that there is an
association between plasma Hcy levels and various vascular
diseases including coronary, cerebral and peripheral arterial
and venous thrombosis (Clarke et al., 1991; Kruman et al.,
* Corresponding author. Tel.: +90 424 237 00 00; fax: +90 424 237 91 38.
E-mail addresses: [email protected], [email protected]
(G. Baydas).
0736-5748/$30.00 # 2007 ISDN. Published by Elsevier Ltd. All rights reserved.
doi:10.1016/j.ijdevneu.2007.03.001
2000; Ho et al., 2002). More recently, clinical studies implicate
increased levels of Hcy and reduced levels of folate with
neurodegenerative conditions including Alzheimer’s disease
(Shea et al., 2002). Increased levels of Hcy are associated with
structural brain changes like cerebral infarcts, white matter
lesions and cerebral atrophy (Vermeer et al., 2002; den Heijer
et al., 2003).
Hcy is an excitatory amino acid and markedly enhances the
vulnerability of neuronal cells to excitotoxic and oxidative
injury in vitro and in vivo (Hankey and Eikelboom, 1999; Streck
et al., 2003; Baydas et al., 2006). Moreover, Hcy has ability
to inhibit the expression of antioxidant enzymes such as
glutathione peroxidase, superoxide dismutase, which might
potentate the toxic effects of free radicals (Ho et al., 2002;
Baydas et al., 2003a, 2006). Recently, Rosenquist and Finnell
(2001) have reported that an increase in the level of maternal
homocysteine induces abnormal development, inhibiting the

G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139134
function of N-methyl-D-aspartate (NMDA) receptors in the
neural epithelium.
Evidence from literature indicates that Hcy is an important
risk factor for cognitive dysfunctions (Miller, 2003). Recently,
our group (Baydas et al., 2005a) and others (Streck et al., 2004)
have suggested that chronic administration of Hcy to rats
impairs both short-term and long-term memory, implying that
hyperhomocysteinemia causes memory impairment through
the enhancement of oxidative stress (Baydas et al., 2005a).
Development of the nervous system requires numerous
factors including neural cell adhesion molecules (NCAM).
NCAM are likely to be involved in synaptogenesis and neuronal
plasticity. Furthermore, there is also evidence suggesting
strongly that they also participate in synaptic changes
underlying memory formation in adult individuals (Schachner,
1997). In addition, NCAM is involved in cellular migration,
axonal growth, and regeneration of peripheral axons (Walsh
and Doherty, 1997; Tzeng et al., 2001). One of the maturational
events in the brain is the alteration in the pattern of expression
of the polysialylated form of the NCAM (PSA-NCAM). PSA-
NCAM is expressed in immature neurons and is important for
morphological plasticity in the hippocampus (Seki and Arai,
1993). PSA-NCAM plays a critical role in axonal growth and
fasciculation during development. The level of PSA-NCAM
decreases dramatically during development in some brain areas
(Doherty et al., 1990).
Intermediate filament proteins comprise a major portion of
the brain’s cytoskeleton with nestin, vimentin, and glial
fibrillary acidic protein (GFAP) differentially expressed with
respect to neurologic development (Hutchins and Casagrande,
1989; Hewicker-Trautwein and Trautwein, 1993). Immature
astrocytes of the CNS usually express vimentin as the major
intermediate filament. Maturation of astrocytes is followed by a
switch between vimentin and GFAP expression, with the latter
being recognized as an astrocyte maturation marker (Gomes
et al., 1999). It is also demonstrated that glial S100B is involved
in neuronal differentiation and maturation (Yang et al., 1996).
The effects of Hcy were extensively investigated in postnatal
life, however, little is known about its deleterious action in
prenatal life. Thus, in the present work we investigated, for the
first time, effects of maternal hyperhomocysteinemia on brain
development by evaluating the expression of NCAM and
intermediate filaments in the brain and behavioral changes of
offsprings.
2. Materials and methods
2.1. Animals and treatments
Female Wistar rats were obtained from the Animal Research Unit, Firat
University, Elazig, and housed in standard plastic cages. Animal room was
maintained on a 12-h light, 12-h dark photoperiod, and all animals were
permitted free access to food and tap water. Daily vaginal smear was performed
on all animals. Pro-estrous animals were left overnight with male rats and the
following day was recorded as gestational day 0 when spermatozoa were
observed in a smear of the vaginal contents.
Pregnant rats were randomly divided into two groups (n = 5 animals/
group) as follows: one group of animals was assigned as control and the second
group was assigned as Hcy group and received daily methionine (1 g/kg body
weight) dissolved in drinking water, starting the first day of gestation through-
out pregnancy. Based upon our (Baydas et al., 2005a) and other published
reports (Bagi et al., 2003) the orally administered methionine dose in this study
increase plasma Hcy levels approximately three to six times compared with
the control values. This is an adequate methionine dose to induce chronic
hyperhomocysteinemia. Due to unattractive option of sulfur group, rats
drinked less water including methionine on the first day, however, from the
second day and onwards they drank adequate water as control rats did. All rats
were housed in individual cages until delivery. All animal procedures were
done according to the Guide for the Care and Use of Laboratory Animals, and
were reviewed by the Institutional Animal Care and Use Committee of the
Firat University.
After delivery, the litter size and the number of live offspring were recorded.
On postnatal day (PND) 1, the body weight and gender of the offsprings of each
litter were recorded. Total Hcy levels in plasma of mothers were determined
with an enzyme immunoassay kit (Axis-Shield AS, Oslo, Norway). Fifteen pups
per group (three from each litter) were dissected and their brains were stored at
�70 8C for the analysis of glial and neuronal markers on PND1. The remaining
pups from each experimental litter were weaned from their dams when they
were 21 days old. After weaning, pups were separated into male and female
cages. Behavioral testing commenced at approximately postnatal day 75.
Twenty-four hours after the behavioral testing, young-adult offsprings from
both groups were decapitated. Brain tissues were removed and hippocampus
were dissected and stored at �70 8C.
2.2. Immunoblotting
Briefly, frozen tissues were thawed; hand-homogenized using glass-glass
homogenisator on ice 1:10 (w/v) in a buffer containing 50 mM Tris (pH 7.4),
0.1 mM NaCl, 1% Triton X-100, 5 mM EDTA, 1.0 mM phenylmethylsulfonyl
fluoride, 10 mg/mL aprotinin and 10 mg/mL leupeptin. Tissue homogenates
were then centrifuged at 40,000 � g at 4 8C for 60 min, and supernatants were
collected into fresh tubes. Protein determinations in the homogenates were
performed according to the Lowry procedure using a protein assay kit (Sigma,
St. Louis, MO, USA). Equal quantities of proteins were separated by SDS-
polyacrylamide gel electrophoresis on 7.5–17.5 polyacrylamide gradient gels as
described previously (Baydas et al., 2002a,b). Separated proteins were trans-
ferred to nitrocellulose membranes (Schleicher and Schuell Inc., Keene, NH,
USA). Nitrocellulose blots were washed twice for 5 min each in phosphate-
buffered saline and blocked with 1% bovine serum albumin in phosphate-
buffered saline for 1 h prior to application of the primary antibody. Blots were
then incubated overnight at 4 8C with primary antibodies including rabbit
polyclonal antibody (Santa Cruz Biotechnology, Inc., USA) against NCAM
(diluted 1:1000), GFAP (diluted 1:2000) and S100B (diluted 1:1500), and
mouse monoclonal antibody (Chemicon Inc., CA, USA) against PSA-NCAM
(diluted at 1:500). The blots were washed and incubated for 1 h with a
secondary antibody, goat anti-rabbit or anti-mouse Ig peroxidase conjugated
(Santa Cruz Biotechnology, Inc., USA) at a dilution of 1:500. Specific binding
was detected using diaminobenzidine and H2O2 as substrates. The relative
amount of immunoreactive bands on Western blots was quantified in arbitrary
units by scanning blots using a computerized software program (LabWorks 4.0;
UVP, Inc., Cambridge, UK).
2.3. Morris water maze
Male (n = 4) and female (n = 4) rat offsprings (8 per group), on approxi-
mately PND75 from the two groups were tested in the Morris water maze test as
we described previously (Baydas et al., 2005b). Briefly, a circular galvanized
steel tank (120 cm diameter, 50 cm height) was filled to a depth of 30 cm with
water at 25 � 1 8C and the tank was divided virtually into four equal quadrants,
labeled N–S–E–W. The water was made opaque by the addition of semi-
skimmed milk. An escape platform (10 cm diameter) was placed in one of the
four maze quadrants (the target quadrant) and submerged 1.5 cm below the
water surface. For the spatial training, the platform was placed in a fixed
quadrant. The rats were required to find the platform using only distal spatial
cues available in the testing room. Cues were maintained constant throughout
testing. The rats were trained to locate and escape onto the platform for four

Fig. 1. Effects of chronic maternal hyperhomocysteinemia on the acquisition of
spatial learning of rats in the Morris water maze test (n = 8 per group). Mean
swimming times of the four trials per day for 5 days for each of the two groups are
shown. Escape latencies required by rats to find the hidden platform along five
consecutive days. Average latency time (days 3 and 5) to find hidden platform was
prolonged by hyperhomocysteinemia (ANOVA; P < 0.05 vs. control).
Fig. 2. Effects of chronic hyperhomocysteinemia on the mean percentage time
spent in the target quadrant in which the platform had previously been located
during acquisition (n = 8 per group). There is no significant difference between
the groups.
G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139 135
trials per day for five consecutive days. A different starting position was used on
each trial (in a quadrant not containing the platform). A trial began by placing
the rat into the water facing the wall of the pool at one of the starting points. The
animals were allowed to swim freely to find the hidden platform within 60 s and
after reaching the platform they were allowed to stay on it for 30 s and then
returned to the cage, which was warmed with a heating pad, to await its next
trial. There was a 30 s intertrial interval. If the rat had failed to escape, then the
rat was directed to the platform by the experimenter and allowed to remain there
for the same amount of time. Data collection was automated by a video tracking
system. Tracking was achieved by a system consisting of a video camera
mounted over the center of the pool. The tracker’s digitized coordinate values
were sampled in turn using a computer.
2.3.1. Probe trial
Twenty-four hours after the last training trial, a probe test was performed
wherein the extent of memory consolidation was assessed. The time spent in the
target quadrant indicates the degree of memory consolidation that has taken
place after learning. In the probe trial, the rat was placed into the pool as in the
training trial, except that the hidden platform was removed from the pool. The
time of crossing the former platform quadrant and the total time of crossing all
quadrants were recorded for 1 min.
Following the memory probe trial, a visible target task (cued trial) was
conducted using a raised platform of a contrasting color to confirm the tested
animals were not visually impaired and to test possible deficits in sensorimotor
processes (Baydas et al., 2003b). Latency times to reach the platform were
recorded for each trial.
2.4. Statistics
Data are reported as the mean � S.E. The data from spatial learning were
analyzed by two-way analysis of variance (ANOVA) for repeated measures. If a
main effect was found, post hoc analyses were done with the Tukey test.
Densitometric results of Western blotting and probe test were analyzed by
Mann–Whitney U test.
3. Results
Daily methionine administration via drinking water to the
pregnant rats significantly increased the plasma Hcy level
measured at the end of pregnancy (control: 6 mM/L and
Hcy group: 26 mM/L; P < 0.001). There was no significant
difference in the number of pups per litter between groups, as
follows (mean number per litter � S.E.): control 11.5 � 1.0 and
Hcy 10.2 � 0.6. The brain and body weights of pups from Hcy
group were slightly but not significantly lower than the control
values. During behavioral test, the mean weights of rats were as
follows: control 195 � 10 g and Hcy 188 � 8 g.
3.1. Morris water maze test
No significant differences occurred between male and female
rats, and the data were therefore pooled for subsequent analysis.
Although the latencies to reach the submerged platform
decreased gradually in both groups during the 5 days of training
in the Morris water maze test, the mean latency was significantly
prolonged in Hcy group compared to control group, indicating a
poorer learning performance due to maternal hyperhomocys-
teinemia (Fig. 1). Significant differences between groups were
observed in the time spent to find the platform on acquisition days
3 and 5. Hcy group showed longer swim latencies than those in
the control groups. ANOVA revealed a significant effect of Hcy
on escape latency (P < 0.05). During probe trials, all the rats
spent the most of the times searching in the quadrant where the
platform had been located (Fig. 2). Hcy offsprings showed no
significant impairment in spatial memory compared with the
controls. The control animals spent a slightly greater percentage
of swimming time in the target quadrant in which the platform
had previously been located compared to Hcy group, however
this greater percentage of time was statistically insignificant
(P > 0.05). Furthermore, the performances of control and Hcy
groups were similar in the trial with the visible platform
(latencies, 10.8 � 0.5 s in control and 11.0 � 0.5 s in Hcy
group), indicating that swimming motivation and ability were
similar and that the observed differences in spatial performance
were not due to sensorimotor disturbances. However, the number
of target crossings was significantly reduced in the rats from
hyperhomocysteinemic mother suggesting a spatial navigation
deficit (Fig. 3; P < 0.05).

Fig. 3. The mean number of platform region crossings from the hidden platform
(n = 8 per group). The number of the target crossings was significantly reduced
in the rats with chronic hyperhomocysteinemia (Mann–Whitney U test;
P < 0.05 vs. control group).
Fig. 5. The levels of S100B in total brain of PND1 pups from control and
hyperhomocysteinemic dams. Brain homogenates were electrophoresed, blotted,
and stained with S100B monoclonal antibody. A representative Western blot of
S100B protein is shown in (A). Computerized densitometry was performed and
the averages (�S.E.) for the two groups (n = 15 per group) are shown in (B). A
significant reduction in S100B contents was observed in the Hcy group of PND1
pups compared to the control pups (Mann–Whitney U test P < 0.001).
G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139136
3.2. Immunoblots
GFAP, the predominant intermediate filament protein of
mature astrocytes, exhibited decreased abundance in Hcy pups
on PND1 relative to the control pups (Fig. 4). The main band of
GFAP protein (49 kDa) is expressed in very low levels in Hcy
pups on PND1 (Fig. 4; P < 0.001). Similar results were also
found in the expression levels of S100B protein. The amount of
this protein was less in Hcy pups compared to the control values
on PND1 (Fig. 5). Western blotting of total brain homogenates
from pups of control and maternally hyperhomocysteinemic
rats revealed differences in the expression of NCAM. On
PND1, a decreased PSA-NCAM expression was observed in
Fig. 4. Effects of maternally induced hyperhomocysteinemia on the level of
GFAP in total brain of pups (n = 15 per group). Hyperhomocysteinemia was
induced by giving daily methionine (1 g/kg body weight) dissolved in drinking
water. Supplementation with methionine was continued throughout their preg-
nancies. Brain homogenates were electrophoresed, blotted, and stained with
GFAP polyclonal antibody. A representative Western blot of GFAP protein is
shown in (A). Computerized densitometry was performed and the averages
(�S.E.) for the two groups (control and Hcy) are shown in (B). A significant
reduction in GFAP contents was observed in the Hcy group of PND1 pups
compared to the control pups (Mann–Whitney U test; P < 0.001).
whole homogenates from Hcy pups compared to the control
values (Fig. 6; P < 0.001). On PND82, the amounts of whole
brain PSA-NCAM in both control and treated animals were
below the level of immunodetection. Thus, on PND82, the
amount of PSA-NCAM was measured by immunoblotting in
hippocampus. Densitometric comparison of immunoblots from
control and treated rats showed a non-significant decrease in the
level of PSA-NCAM expression in the hippocampal homo-
genates from Hcy rats (Fig. 7; P < 0.05).
Densitometric comparison of NCAM blots from Hcy group
showed a significant reduction in NCAM-140 expression in
whole homogenates of Hcy pups (Fig. 8; P < 0.001). No gender
differences in the expression of PSA-NCAM or other NCAM
isoforms were observed in control or Hcy rat pups at any time-
point. On PND82, while the level of NCAM-180 was markedly
decreased in the hippocampus of Hcy group (P < 0.001),
NCAM-120 and NCAM-140 isoforms appeared unaffected
(Fig. 9).
Fig. 6. Mean amount of PSA-NCAM in total brain of PND1 pups (n = 15 per
group) from the control and hyperhomocysteinemic mothers. Mouse mono-
clonal antibody against PSA-NCAM (diluted at 1:500) was used to stain blots.
PSA-NCAM expression was lower in Hcy group compared to the control values
(Mann–Whitney U test; P < 0.001).
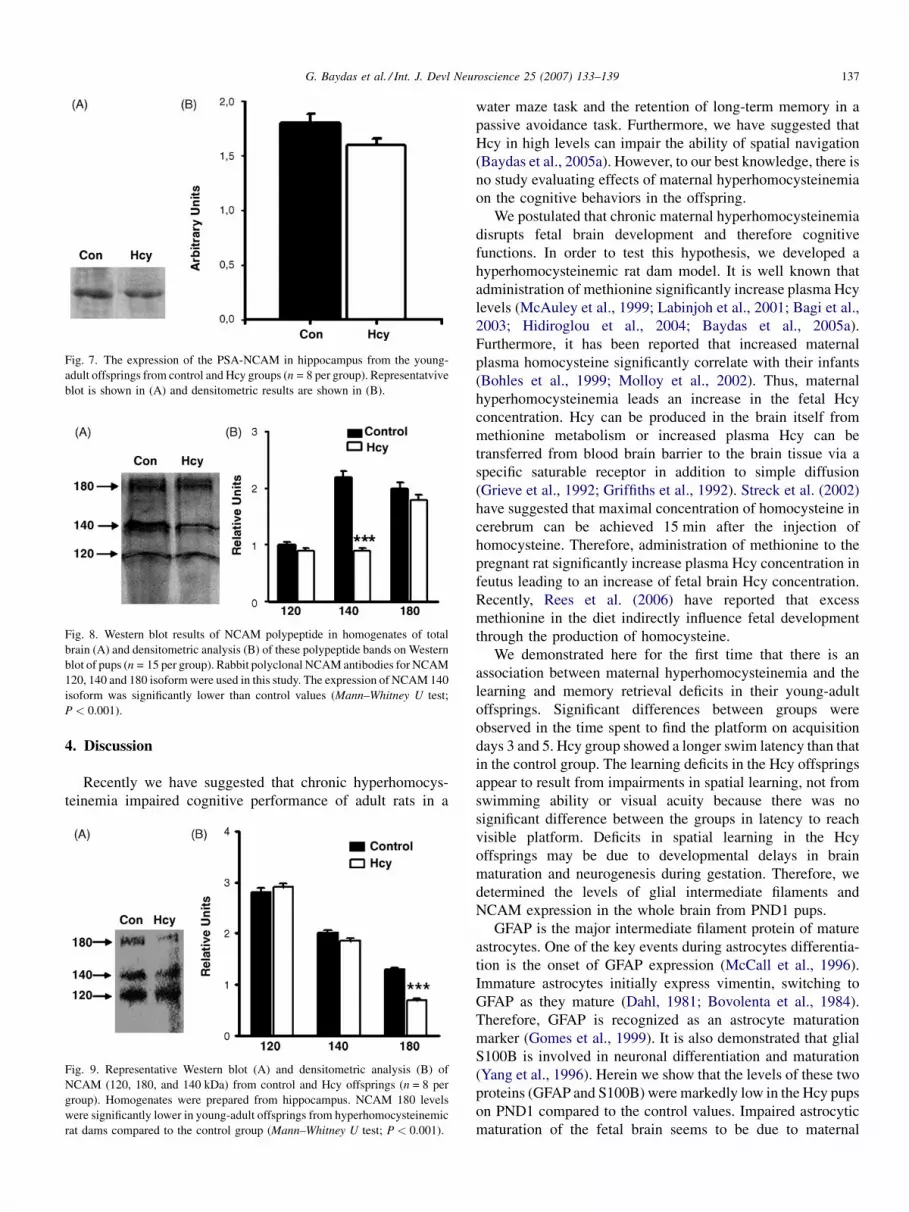
Fig. 7. The expression of the PSA-NCAM in hippocampus from the young-
adult offsprings from control and Hcy groups (n = 8 per group). Representatvive
blot is shown in (A) and densitometric results are shown in (B).
Fig. 8. Western blot results of NCAM polypeptide in homogenates of total
brain (A) and densitometric analysis (B) of these polypeptide bands on Western
blot of pups (n = 15 per group). Rabbit polyclonal NCAM antibodies for NCAM
120, 140 and 180 isoform were used in this study. The expression of NCAM 140
isoform was significantly lower than control values (Mann–Whitney U test;
P < 0.001).
G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139 137
4. Discussion
Recently we have suggested that chronic hyperhomocys-
teinemia impaired cognitive performance of adult rats in a
Fig. 9. Representative Western blot (A) and densitometric analysis (B) of
NCAM (120, 180, and 140 kDa) from control and Hcy offsprings (n = 8 per
group). Homogenates were prepared from hippocampus. NCAM 180 levels
were significantly lower in young-adult offsprings from hyperhomocysteinemic
rat dams compared to the control group (Mann–Whitney U test; P < 0.001).
water maze task and the retention of long-term memory in a
passive avoidance task. Furthermore, we have suggested that
Hcy in high levels can impair the ability of spatial navigation
(Baydas et al., 2005a). However, to our best knowledge, there is
no study evaluating effects of maternal hyperhomocysteinemia
on the cognitive behaviors in the offspring.
We postulated that chronic maternal hyperhomocysteinemia
disrupts fetal brain development and therefore cognitive
functions. In order to test this hypothesis, we developed a
hyperhomocysteinemic rat dam model. It is well known that
administration of methionine significantly increase plasma Hcy
levels (McAuley et al., 1999; Labinjoh et al., 2001; Bagi et al.,
2003; Hidiroglou et al., 2004; Baydas et al., 2005a).
Furthermore, it has been reported that increased maternal
plasma homocysteine significantly correlate with their infants
(Bohles et al., 1999; Molloy et al., 2002). Thus, maternal
hyperhomocysteinemia leads an increase in the fetal Hcy
concentration. Hcy can be produced in the brain itself from
methionine metabolism or increased plasma Hcy can be
transferred from blood brain barrier to the brain tissue via a
specific saturable receptor in addition to simple diffusion
(Grieve et al., 1992; Griffiths et al., 1992). Streck et al. (2002)
have suggested that maximal concentration of homocysteine in
cerebrum can be achieved 15 min after the injection of
homocysteine. Therefore, administration of methionine to the
pregnant rat significantly increase plasma Hcy concentration in
feutus leading to an increase of fetal brain Hcy concentration.
Recently, Rees et al. (2006) have reported that excess
methionine in the diet indirectly influence fetal development
through the production of homocysteine.
We demonstrated here for the first time that there is an
association between maternal hyperhomocysteinemia and the
learning and memory retrieval deficits in their young-adult
offsprings. Significant differences between groups were
observed in the time spent to find the platform on acquisition
days 3 and 5. Hcy group showed a longer swim latency than that
in the control group. The learning deficits in the Hcy offsprings
appear to result from impairments in spatial learning, not from
swimming ability or visual acuity because there was no
significant difference between the groups in latency to reach
visible platform. Deficits in spatial learning in the Hcy
offsprings may be due to developmental delays in brain
maturation and neurogenesis during gestation. Therefore, we
determined the levels of glial intermediate filaments and
NCAM expression in the whole brain from PND1 pups.
GFAP is the major intermediate filament protein of mature
astrocytes. One of the key events during astrocytes differentia-
tion is the onset of GFAP expression (McCall et al., 1996).
Immature astrocytes initially express vimentin, switching to
GFAP as they mature (Dahl, 1981; Bovolenta et al., 1984).
Therefore, GFAP is recognized as an astrocyte maturation
marker (Gomes et al., 1999). It is also demonstrated that glial
S100B is involved in neuronal differentiation and maturation
(Yang et al., 1996). Herein we show that the levels of these two
proteins (GFAP and S100B) were markedly low in the Hcy pups
on PND1 compared to the control values. Impaired astrocytic
maturation of the fetal brain seems to be due to maternal

G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139138
hyperhomocysteinemia. It is reported that GFAP is involved
neuronal–glial interaction (McCall et al., 1996). Thus,
alterations in the levels of GFAP may result in disturbed
neuron–neuron and neuron–glia connectivity.
Development of the nervous system is a complex process.
Evidence suggests that NCAM participate in several aspects
of neural development (Edelman and Crossin, 1991). NCAM
mediates cell–cell interaction, modulates developmental pro-
cesses including neuronal migration and neurite extension, and
participates in neuronal regeneration and hippocampal neuro-
genesis in adult brain (Seki and Arai, 1993; Walsh and Doherty,
1997). One of the latest maturational events in the brain is the
alteration in the pattern of expression of the PSA-NCAM. This
immature form of NCAM plays a critical role in axonal growth
and fasciculation during brain development (Doherty et al.,
1990). In the present study, although we found that PSA-
NCAM was highly expressed in the whole brain homogenates
from the PND1 pups, it was undetectable in the whole brains of
mature rats. However, on the PND82, PSA-NCAM was
detected in hippocampal homogenates. In the adults, PSA-
NCAM remains expressed in the regions exhibiting a robust
synaptic plasticity such as hippocampus, which is well
known to retain a high degree of plasticity in adult rat
(Hoffman et al., 2001).
In the present study, the expression pattern of NCAM
isoforms in adult rats was different from that in the PND1 pups.
On PND1, the level of NCAM 180 was higher than that of other
isoforms, whereas the level of NCAM 120 was higher in mature
brain. This is in accordance with the previous findings that
NCAM 120 is the latest isoform to appear during development
(Chuong and Edelman, 1984; Nagata and Schachner, 1986).
There seems to be an association between maternal hyperho-
mocysteinemia and the decrease in the expression of PSA-
NCAM both in the whole brain from the PND1 pups and in the
hippocampus from the young-adult offsprings. The addition of
PSA to NCAM polypeptides seems to be a critical functional
feature for NCAM-mediated cell–cell interaction and function.
High expression of PSA-NCAM during early development
appears to play a permissive role, allowing structural remo-
deling by decreasing cell adhesion mediated by NCAM and
thereby facilitating the guidance and targeting of axons and
migration of neuronal and glial precursors (Rutishauser, 1996;
Minana et al., 2000). Decreased PSA-NCAM expression in
the developing brain might create a suitable environment to
reduce the rate of axonal growth at the target, making NCAM
available in its highly adhesive state to modulate glial–neuronal
interaction-dependent processes (Minana et al., 2000). Thus,
maternal hyperhomocysteinemia seems to impair the expected
increase in NCAM polysialylation, possibly leading to
persisting neuroplastic deficits, which may cause impairments
of learning and memory performance.
In the present study, there was an imbalance in the expression
of NCAM isoforms with a significant decrease in the expression
of NCAM 140 in the brains of pups from the hyperhomocys-
teinemic mothers on PND1. However, NCAM 180 isoform was
expressed less in the hippocampus from the offsprings of Hcy
group on PND82 compared to the control ones. NCAM 140 is
expressed on both pre- and post-synaptic membranes and plays a
pivotal role in cell–cell adhesion and neurite outgrowth
(Schachner, 1997). Therefore, we have proposed that the
reduction in NCAM 140 expression might be implicated in
the structural alterations of brain during development. However,
the causal involvement still needs to be established.
The fact that NCAM 180 is the isoform particularly affected in
the hippocampus of offspring from mother with hyperhomo-
cysteinemia supports a key role for this isoform on cognitive
processing-associated synaptic plasticity in this brain region.
NCAM 180 has been suggested to be an essential determinant in
synaptic plasticity and to be critically implicated in the
stabilization of synaptic strength (Schuster et al., 1998; Dityatev
et al., 2000). Given the importance of NCAM 180 in synaptic
function, the decreased level of NCAM 180 expression in the
hippocampus of the offspring from hyperhomocysteinemic dams
could lead to a synaptic destabilization in neural circuits relevant
to the storage of information.
In conclusion, present results suggest that maternal hyperho-
mocysteinemia results in the learning deficits in offspring due to
delayed brain maturation and altered expression pattern of
NCAM.
Acknowledgement
This work was supported by the Firat University Research
Foundation (FUBAP-1148).
References
Bagi, Z., Cseko, C., Toth, E., Koller, A., 2003. Oxidative stress-induced
dysregulation of arteriolar wall shear stress and blood pressure in hyperho-
mocysteinemia is prevented by chronic vitamin C treatment. Am. J. Physiol.
Heart Circ. Physiol. 285, 2277–2283.
Baydas, G., Kutlu, S., Naziroglu, M., Canpolat, S., Sandal, S., Ozcan, M.,
Kelestimur, H., 2003a. Inhibitory effects of melatonin on neural lipid
peroxidation induced by intracerebroventricularly administered homocys-
teine. J. Pineal Res. 34, 36–39.
Baydas, G., Nedzvetskii, V.S., Nerush, P.A., Kirichenko, S.V., Yoldas, T.,
2003b. Altered expression of NCAM in hippocampus and cortex may
underlie memory and learning deficits in rats with streptozotocin-induced
diabetes mellitus. Life Sci. 73, 1907–1916.
Baydas, G., Nedzvetsky, V.S., Nerush, P.A., Kirichenko, S.V., Demchenko,
H.M., Reiter, R.J., 2002b. A novel role for melatonin: regulation of the
expression of cell adhesion molecules in the rat hippocampus and cortex.
Neurosci. Lett. 326, 109–112.
Baydas, G., Ozer, M., Yasar, A., Koz, S.T., Tuzcu, M., 2006. Melatonin prevents
oxidative stress and inhibits reactive gliosis induced by hyperhomocystei-
nemia in rats. Biochemistry (Mosc) 71, 91–95.
Baydas, G., Ozer, M., Yasar, A., Tuzcu, M., Koz, S.T., 2005a. Melatonin
improves learning and memory performances impaired by hyperhomocys-
teinemia in rats. Brain Res. 1046, 187–194.
Baydas, G., Ozveren, F., Tuzcu, M., Yasar, A., 2005b. Effects of thinner
exposure on the expression pattern of neural cell adhesion molecules, level
of lipid peroxidation in the brain and cognitive function in rats. Eur. J.
Pharmacol. 512, 181–187.
Baydas, G., Reiter, R.J., Nedzvetskii, V.S., Nerush, P.A., Kirichenko, S.V.,
2002a. Altered glial fibrillary acidic protein content and its degradation in
the hippocampus, cortex and cerebellum of rats exposed to constant light:
reversal by melatonin. J. Pineal Res. 33, 134–139.
Bohles, H., Arndt, S., Ohlenschlager, U., Beeg, T., Gebhardt, B., Sewell, A.C.,
1999. Maternal plasma homocysteine, placenta status and docosahexaenoic

G. Baydas et al. / Int. J. Devl Neuroscience 25 (2007) 133–139 139
acid concentration in erythrocyte phospholipids of the newborn. Eur. J.
Pediatr. 158, 243–246.
Bovolenta, P., Liem, R.K., Mason, C.A., 1984. Development of cerebellar
astroglia: transitions in form and cytoskeletal content. Dev. Biol. 102,
248–259.
Chuong, C.M., Edelman, G.M., 1984. Alterations in neural cell adhesion
molecules during development of different regions of the nervous system.
J. Neurosci. 4, 2354–2368.
Clarke, R., Daly, L., Robinson, K., Naughten, E., Cahalane, S., Fowler, B.,
Graham, I., 1991. Hyperhomocysteinemia: an independent risk factor for
vascular disease. N. Engl. J. Med. 324, 1149–1155.
Dahl, D., 1981. The vimentin-GFA protein transition in rat neuroglia
cytoskeleton occurs at the time of myelination. J. Neurosci. Res. 6, 741–
748.
den Heijer, T., Vermeer, S.E., Clarke, R., Oudkerk, M., Koudstaal, P.J., Hofman,
A., Breteler, M.M., 2003. Homocysteine and brain atrophy on MRI of non-
demented elderly. Brain 126, 170–175.
Dityatev, A., Dityateva, G., Schachner, M., 2000. Synaptic strength as a
function of post- versus presynaptic expression of the neural cell adhesion
molecule, NCAM. Neuron 26, 207–217.
Doherty, P., Cohen, J., Walsh, F., 1990. Neurite outgrowth in response to
transfected N-CAM changes during development and is modulated by
polysialic acid. Neuron 5, 209–219.
Edelman, G.M., Crossin, K.L., 1991. Cell adhesion molecules: implications for
molecular histology. Annu. Rev. Biochem. 60, 155–190.
Gomes, F.C., Paulin, D., Moura Neto, V., 1999. Glial fibrillary acidic protein
(GFAP): modulation by growth factors and its implication in astrocyte
differentiation. Braz. J. Med. Biol. Res. 32, 619–631.
Grieve, A., Butcher, S.P., Griffiths, R., 1992. Synaptosomal plasma membrane
transport of excitatory sulphur amino acid transmitter candidates: kinetic
characterisation and analysis of carrier specificity. J. Neurosci. Res. 32,
60–68.
Griffiths, R., Grieve, A., Allen, S., Olverman, H.J., 1992. Neuronal and glial
plasma membrane carrier-mediated uptake of L-homocysteate is not selec-
tively blocked by beta-p-chlorophenylglutamate. Neurosci. Lett. 147, 175–
178.
Hankey, G.J., Eikelboom, J.W., 1999. Homocysteine and vascular disease.
Lancet 354, 407–413.
Hewicker-Trautwein, M., Trautwein, G., 1993. An immunohistochemical study
of the fetal sheep neocortex and cerebellum with antibodies against nervous
system-specific proteins. J. Comp. Pathol. 109, 409–421.
Hidiroglou, N., Gilani, G.S., Long, L., Zhao, X., Madere, R., Cockell, K.,
Belonge, B., Ratnayake, W.M., Peace, R., 2004. The influence of dietary
vitamin E, fat, and methionine on blood cholesterol profile, homocysteine
levels, and oxidizability of low density lipoprotein in the gerbil. J. Nutr.
Biochem. 15, 730–740.
Hirche, F., Schroder, A., Knoth, B., Stangl, G.I., Eder, K., 2006. Methionine-
induced elevation of plasma homocysteine concentration is associated with
an increase of plasma cholesterol in adult rats. Ann. Nutr. Metab. 50, 139–
146.
Ho, P.I., Ortiz, D., Rogers, E., Shea, T.B., 2002. Multiple aspects of homo-
cysteine neurotoxicity: glutamate excitotoxicity, kinase hyperactivation and
DNA damage. J. Neurosci. Res. 70, 694–702.
Hoffman, K.B., Murray, B.A., Lynch, G., Munirathinam, S., Bahr, B.A., 2001.
Delayed and isoform-specific effect of NMDA exposure on neural cell
adhesion molecules in hippocampus. Neurosci. Res. 39, 167–173.
Hutchins, J.B., Casagrande, V.A., 1989. Vimentin: changes in distribution
during brain development. Glia 2, 55–56.
Kruman, I.I., Culmsee, C., Chan, S.L., Kruman, Y., Guo, Z., Penix, L., Mattson,
M.P., 2000. Homocysteine elicits a DNA damage response in neurons that
promotes apoptosis and hypersensitivity to exitotoxicity. J. Neurosci. Res.
20, 6920–6926.
Labinjoh, C., Newby, D.E., Wilkinson, I.B., Megson, I.L., MacCallum, H.,
Melville, V., Boon, N.A., Webb, D.J., 2001. Effects of acute methionine
loading and vitamin C on endogenous fibrinolysis, endothelium-
dependent vasomotion and platelet aggregation. Clin. Sci. (Lond.)
100, 127–135.
McAuley, D.F., Hanratty, C.G., McGurk, C., Nugent, A.G., Johnston, G.D., 1999.
Effect of methionine supplementation on endothelial function, plasma homo-
cysteine, and lipid peroxidation. J. Toxicol. Clin. Toxicol. 37, 435–440.
McCall, M.A., Gregg, R.G., Behringer, R.R., Brenner, M., Delaney, C.L.,
Galbreath, E.J., Zhang, C.L., Pearce, R.A., Chiu, S.Y., Messing, A.,
1996. Targeted deletion in astrocyte intermediate filament (Gfap) alters
neuronal physiology. Proc. Natl. Acad. Sci. U.S.A. 93, 6361–6366.
Miller, A.L., 2003. The methionine-homocysteine cycle and its effects on
cognitive diseases. Altern. Med. Rev. 8, 7–19.
Minana, R., Climent, E., Barettino, D., Segui, J.M., Renau-Piqueras, J., Guerri,
C., 2000. Alcohol exposure alters the expression pattern of neural cell
adhesion molecules during brain development. J. Neurochem. 75, 954–964.
Molloy, A.M., Mills, J.L., McPartlin, J., Kirke, P.N., Scott, J.M., Daly, S., 2002.
Maternal and fetal plasma homocysteine concentrations at birth: the influ-
ence of folate, vitamin B12, and the 5,10-methylenetetrahydrofolate reduc-
tase 677C-T variant. Am. J. Obstet. Gynecol. 186, 499–503.
Nagata, I., Schachner, M., 1986. Conversion of embryonic to adult form of the
neural cell adhesion molecule (NCAM) does not correlate with pre- and
postmigratory states of mouse cerebellum granule neurons. Neurosci. Lett.
63, 153–158.
Rees, W.D., Wilson, F.A., Maloney, C.A., 2006. Sulfur amino acid metabolism
in pregnancy: the impact of methionine in the maternal diet. J. Nutr. 136,
1701S–1705S.
Reis, E.A., Zugno, A.I., Franzon, R., Tagliari, B., Matte, C., Lammers, M.L.,
Netto, C.A., Wyse, A.T., 2002. Pretreatment with vitamins E and C prevent
the impairment of memory caused by homocysteine administration in rats.
Metab. Brain Dis. 17, 211–217.
Rosenquist, T.H., Finnell, R.H., 2001. Genes, folate and homocysteine in
embryonic development. Proc. Nutr. Soc. 60, 53–61.
Rutishauser, U., 1996. Polysialic acid and the regulation of cell interactions.
Curr. Opin. Cell Biol. 8, 679–684.
Schachner, M., 1997. Neural recognition molecules and synaptic plasticity.
Curr. Opin. Cell Biol. 9, 627–634.
Schuster, T., Krug, M., Hassan, H., Schachner, M., 1998. Increase in proportion
of hippocampal spine synapses expressing neural cell adhesion molecule
NCAM180 following long-term potentiation. J. Neurobiol. 37, 359–372.
Seki, T., Arai, Y., 1993. Highly polysialylated neural cell adhesion molecule
(NCAM-H) is expressed by newly generated granule cells in the dentate
gyrus of the adult rat. J. Neurosci. 13, 2351–2358.
Shea, T.B., Lyons-Weiler, J., Rogers, E., 2002. Homocysteine, folate depriva-
tion and Alzheimer neuropathology. J. Alzheimers Dis. 4, 261–267.
Streck, E.L., Matte, C., Vieira, P.S., Rombaldi, F., Wannmacher, C.M., Wajner,
M., Wyse, A.T., 2002. Reduction of Na(+),K(+)-ATPase activity in hippo-
campus of rats subjected to chemically induced hyperhomocysteinemia.
Neurochem. Res. 27, 1593–1598.
Streck, E.L., Vieira, P.S., Wannmacher, C.M., Dutra-Filho, C.S., Wajner, M.,
Wyse, A.T., 2003. In vitro effect of homocysteine on some parameters of
oxidative stress in rat hippocampus. Metab. Brain Dis. 18, 147–154.
Streck, E.L., Bavaresco, C.S., Netto, C.A., Wyse, A.T., 2004. Chronic hyper-
homocysteinemia provokes a memory deficit in rats in the Morris water
maze task. Behav. Brain Res. 153, 377–381.
Tzeng, S.F., Cheng, H., Lee, Y.S., Wu, J.P., Hoffer, B.J., Kuo, J.S., 2001.
Expression of neural cell adhesion molecule in spinal cords following a
complete transaction. Life Sci. 68, 1005–1012.
Vermeer, S.E., van Dijk, E.J., Koudstaal, P.J., Oudkerk, M., Hofman, A., Clarke,
R., Breteler, M.M., 2002. Homocysteine, silent brain infarcts, and white
matter lesions: the Rotterdam scan study. Ann. Neurol. 51, 285–289.
Walsh, F.S., Doherty, P., 1997. Neural cell adhesion molecules of the immu-
noglobulin superfamily: role in axon growth and guidance. Annu. Rev. Cell
Dev. Biol. 13, 425–456.
Yang, Q., Hamberger, A., Wang, S., Haglid, K.G., 1996. Appearance of
neuronal S-100 beta during development of the rat brain. Brain Res.
Dev. Brain Res. 91, 181–189.
Recommended

















