
Expression of Cathepsins B, H, K, L, and S During HumanFetal Lung DevelopmentF. BUHLING,1* N. WALDBURG,2 S. KRUGER,3 C. ROCKEN,3 O. WIESNER,2 E. WEBER,4 AND T. WELTE2
1Institute of Immunology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany2Department of Pneumology and Critical Care, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany3Institute of Pathology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany4Institute of Physiological Chemistry, Martin-Luther-University Halle, Halle, Germany
ABSTRACT Cathepsins are involved in lyso-somal protein degradation, proenzyme activa-tion, antigen processing, and hormone matura-tion. They are secreted by tumor cells andmacrophages and catalyze the remodeling of ex-tracellular matrix proteins. To gain insight intothe expression pattern of cathepsins during fetallung development, the expression of cathepsinsB, H, K, L, and S at protein and mRNA levels wereevaluated by using immunohistochemistry andin situ hybridization. Early expression of ca-thepsins B, H, and K was found in epithelial cellsof the branching presumptive bronchi (<12thweek of gestation). The most intense cathepsinK–specific immunoreactivity was found in devel-oping airways with a lumen. Cathepsin K wasfound in epithelial cells only, whereas in con-trast, cathepsins B and H were detected both inepithelial and interstitial cells. During fetal mat-uration, interstitial cells displayed cathepsin Limmunoreactivity and, in the saccular phase(>26th week of gestation), both cathepsin L and Simmunoreactivities. A continuous decline in theproportion of cathepsin H-positive interstitialCD68-positive cells was observed. These discretetemporal and spatial variations in cathepsin ex-pression during organogenesis of the humanlung indicate different physiological roles for theindividual enzymes in different cell types anddevelopmental stages. © 2002 Wiley-Liss, Inc.
Key words: cathepsin; fetal development; hu-man; immunohistochemistry
INTRODUCTION
Mammalian cysteine proteases of the papain familyhave been implicated in protein degradation and turn-over within the endosomal/lysosomal system (Kirschkeand Wiederanders, 1987) and in limited proteolysisevents such as proenzyme activation (Samarel et al.,1989), antigen processing (Nakagawa et al., 1998;Riese and Chapman, 2000), and hormone maturation(Docherty et al., 1984). Evidence indicates that cys-teine proteases are involved in a variety of physiologi-cal and pathologic processes such as bone resorption(Inui et al., 1997), muscular dystrophy (Katunuma and
Kominami, 1987), arthritis (Mort et al., 1984), tumorinvasion and metastasis (Berquin and Sloane, 1996),and Alzheimer’s disease (Lemere et al., 1995). There-fore, these enzymes are of clinical interest as targetsfor the development of inhibitors (Smith and Ab-delmeguid, 1999). They are probably important duringontogenesis, because rapid tissue remodeling takesplace during embryonic and fetal development. Addi-tionally, various proteolytically processed hormonesand growth factors are involved in growth and differ-entiation. It has been shown consistently that inhibi-tion of cysteine proteases produces a concentration-dependent decrease in embryonic growth and anincrease in abnormalities (Daston et al., 1991).
Among the cysteine proteases, the papain family isby far the largest and includes the lysosomal cathep-sins B, C, H, L, and S, in addition to the more recentlydescribed cathepsins F, K, O, V, W, and X. Sequencehomologies of cathepsins indicate that these enzymesdiverged during early stages of eukaryotic evolution.They are synthesized as inactive preproproteins andare proteolytically processed and activated duringtransport to the lysosomes. Most cysteine proteases ofthe papain family are endopeptidases. Exopeptidases,such as cathepsin B (cathB) and cathepsin H (cathH),are likely to have evolved from the endopeptidase tem-plate by addition of structural elements capable of in-teracting with the C- or N-terminus of a substrate(Turk et al., 1997). In general, cathepsins show a highredundancy in their substrate specificity. Neverthe-less, single enzymes may play a crucial role in theprocessing of defined proteins in a given organ or celltype. Deficiency of cathepsin K (cathK) results in pyc-nodysostosis in man and osteopetrosis in mice (Saftiget al., 1998). Cathepsin L (cathL) deficiency leads to afurless phenotype in mice (Roth et al., 2000). Process-
Grant sponsor: Deutsche Forschungsgemeinschaft; Grant number:WE 1922/1-1.
*Correspondence to: Frank Buhling, Institute of Immunology, Otto-von-Guericke-University Magdeburg, Leipziger-Str. 44, 39120 Mag-deburg, Germany.E-mail: [email protected]
Received 24 August 2001; Accepted 7 June 2002DOI 10.1002/dvdy.10134Published online 19 July 2002 in Wiley InterScience (www.
interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 225:14–21 (2002)
© 2002 WILEY-LISS, INC.

ing of the invariant chain of the major histocompatibil-ity complex (MHC) class II complex is performed bydifferent cathepsins in macrophages, dendritic cells,and epithelial cells (Nakagawa et al., 1998; Driessen etal., 1999; Shi et al., 2000). The deficiency of the respec-tive proteases is not compensated for by up-regulatingan alternative enzyme, but instead leads to an alter-ation in MHC class II processing.
The development of human fetal lung during gesta-tion can be divided into five overlapping phases. Dur-ing the embryonic phase (4–7 weeks), the tubes thatresemble air passages are formed. During the pseu-doglandular phase (5–17 weeks), the pseudostratifiedepithelium of the lung buds is reduced to a columnarform and the epithelial tubes have no or little develop-ing lumen. After this phase, the canalicular phase(16–26 weeks) is characterized by the formation ofbronchioles. The first part of the respiratory gas ex-change unit is also formed, the mesenchymal tissuethins out, and there is capillary invasion into theseperipheral units. During the saccular phase (26–36weeks), a thinning of the interstitium and a flatteningof most of the epithelium results in a close link betweencapillaries and epithelial cells. Furthermore, differen-tiated type I- and type II-pneumocytes are found, andsurfactant production is induced. In parallel to epithe-lial cells, differentiation of mesenchymal cells intomyoblasts, fibroblasts, and macrophages occurs duringlate canalicular and saccular phases. The final matu-ration of the lung occurs in the alveolar phase (36weeks of gestation to 18 months postnatal) during lateprenatal and first postnatal period (Adamson, 1997).
To date, most of the detailed developmental studieshave been carried out on pregnant animals. To ourknowledge, there is no systematic study concerning thedynamics of the expression of lysosomal enzymes dur-ing human lung development. However, the recogni-tion of specific expression patterns of different enzymesin distinct cell types during various phases of fetaldevelopment is a precondition to understanding theirfunctional role in, for example, matrix remodeling, ly-sosomal degradation of foreign proteins, or growth fac-tor processing. This understanding may improve ourknowledge of the pathogenesis of dystrophic lung de-velopment. This study is the first to investigate theexpression pattern of cathepsins B, H, K, L, and S inhuman fetal lung tissue by using immunohistochemis-try. In selected samples, the expression pattern wasconfirmed at the mRNA level by using in situ hybrid-ization.
RESULTSExpression Pattern During Glandular Phase(<16th Week of Gestation)
In the early period of lung development, spreading ofthe bronchial tree was documented. Some of the epi-thelial tubes were developing or had a lumen duringthis phase of development (Fig. 1A, arrow). Betweenthe branches of the bronchial tree, a relatively compact
mass of interstitial tissue was found. Of all the ca-thepsins analyzed, only cathK and cathH were signif-icantly expressed in the earlier glandular phase. Bothenzymes were found in the cytoplasm of epithelial cellsof the branching presumptive bronchi with an expres-sion pattern similar to cytokeratin (Fig. 1B). From thisearly phase of development up to the canalicular phase,cathK immunostaining was most intense in bronchialtubes with columnar epithelial cells and a lumen (Fig.1C, arrow). Expression of cathK was less prominent indeveloping bronchial sacs with large numbers of undif-ferentiated epithelial cells (Fig. 1C, star). This findingindicates that cathK exhibits a different expression inimmature distal epithelial cells of respiratory tubulesin comparison with more differentiated proximal epi-thelial cells (see also Fig. 3B). In contrast to cathH,cathK was found in epithelial cells only. CathH (Fig.1D) and cathB (Fig. 1F,G) were detected at the begin-ning of the glandular phase (10th–12th week of gesta-tion) mainly in bronchial epithelial cells and in inter-stitial cells. The cathH and the cathB immunostainingwere similar in distal and proximal epithelial cells. Adistinct cathH immunostaining was present in somebasal epithelial cells (Fig. 1D, arrows) similar to thatobserved with the precursor of surfactant protein C(pro–SP-C) antiserum (Fig. 1E, arrows), which detectspro–SP-C–producing cells. CathL was expressed in in-terstitial cells but not in bronchial epithelial cells (Fig.1H,I). Immunostaining for cathS was not found in thisphase of fetal development.
For further analysis of cathepsin expression in inter-stitial cells, we analyzed the number of interstitial cellsthat displayed a cathB-, cathH-, cathL-, or cathS-spe-cific immunoreactivity in relation to the CD68-positivecells (Table 1). CD68 was shown to be a marker ofmyeloid cells. The coexpression of CD68 and differentcathepsins was documented by immunodoublelabeling(Fig. 2). We found no CD68-negative interstitial orintra-alveolar cells that displayed cathepsin B, H, L, orS immunoreactivity. Although cathB and cathL expres-sion were nearly unchanged during fetal development,we found a significant decrease in cathH immunoreac-tivity in later phases. In contrast, the expression ofcathS was increased in canalicular and saccular/earlyalveolar phases.
Canalicular Phase (16th–26th Week ofGestation)
The canalicular phase is characterized by furtherspreading of the presumptive bronchial tree and thedevelopment of smaller respiratory tubules while theinterstitial tissue thins (Fig. 3A). The development ofalveolar spaces begins at the end of this phase. Thestaining pattern of most cathepsins was unchanged,but cathK was expressed more ubiquitously in all sam-ples studied from this period of development. In addi-tion to staining of bronchial epithelial cells, cathK wasfound in the interstitium (Fig. 3B). In situ hybridiza-tion on selected tissue samples showed that cathK is
15CATHEPSINS IN FETAL LUNG DEVELOPMENT

synthesized by bronchial epithelial cells and by someinterstitial cells, which are not identical with theCD68-positive cells (Fig. 3C). Some cathS-positive cellswere detected in perivascular areas (Table 1).
Saccular Phase/Early Alveolar Phase (26th–37thWeek of Gestation)
This final phase of fetal development is characterizedby a further differentiation of the epithelium and by areduction in the interstitial mass (Fig. 3D). In thisphase, the staining pattern of most of the cathepsins
starts to resemble that of adult organs, which wasrecently described (Buhling et al., 1999a). The cathKstaining was limited to bronchial epithelial cells (Fig.3E). CathH (Fig. 3F,G), cathB (not shown), and cathL(not shown) were found predominantly in macrophagesand epithelial cells. Both cathH and pro–SP-C wereexpressed in single cells of terminal sacs, which mayrepresent type II pneumocytes (Fig. 3G,H, arrows).CathS was detected mainly in CD68-positive histio-cytic cells (Fig. 3I). Whereas the cathB- and cathL-expressing CD68-positive cells remained essentially
Fig. 1. Glandular phase: Epithelial cells of branching presumptivebronchi, identified by cytokeratin staining, with lumen (A,B, arrows) ex-press a stronger cathepsin (cath) K immunoreactivity (C, arrow) com-pared with less differentiated epithelial cells of developing airways with-out lumen (A–C, stars). CathH (D) and cathB (F,G, arrows) were found inbronchial epithelial cells and interstitial cells. Some interstitial cells ex-
pressed cathL (H,I, arrows). A: Hematoxylin and eosin, �200, case 5;B: anti-cytokeratin, �250, case 5; C: anti-cathK, �250, case 5; D:anti-cathH, �200, case 1; E: anti–pro-SP-C, �250, case 1; F: anti-cathB,�400, case 1; G: cathB antisense probe, �250, case 1; H: anti-cathL,�400, case 5; I: cathL antisense probe, �250, diaminobenzidine�nickelchloride, gray-black, case 5.
TABLE 1. Relative Number of Cells Expressing Immunoreactivity forCathepsins B, H, L, or Sa
Cathepsin
Gestational age10th–16th week
(n � 10)16th–26th week
(n � 9)�26th week
(n � 5)B 65 � 21% 58 � 19% 71 � 11%H 51 � 23% 28 � 11% 22 � 8 %L 54 � 18% 49 � 15% 48 � 21%S 0 � 0 % 6 � 3 % 13 � 4 %
aData presented as mean percent � SD. The number of cathepsin-positive interstitial cells wascounted as described in the Experimental Procedures section and was divided by the amount ofCD68-positive cells.
16 BUHLING ET AL.
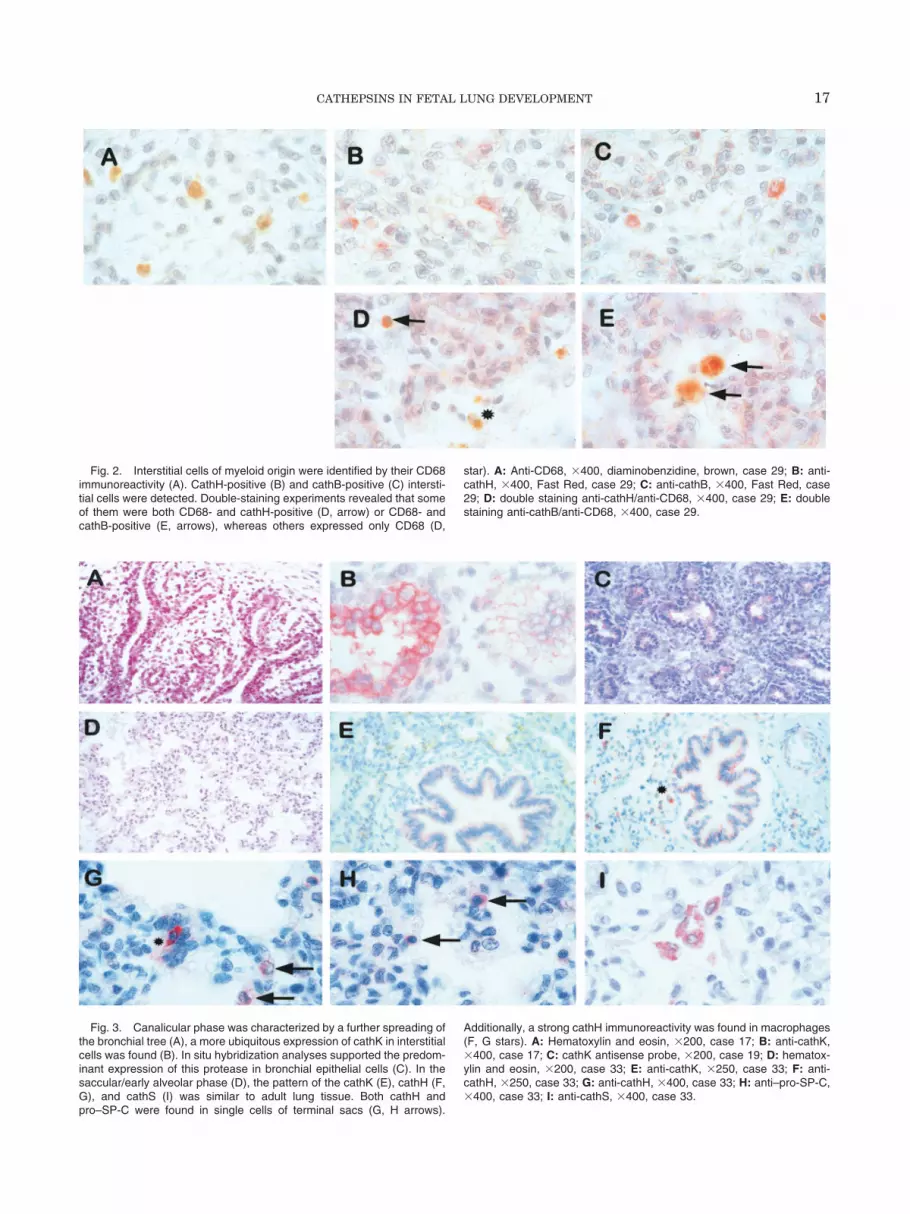
Fig. 2. Interstitial cells of myeloid origin were identified by their CD68immunoreactivity (A). CathH-positive (B) and cathB-positive (C) intersti-tial cells were detected. Double-staining experiments revealed that someof them were both CD68- and cathH-positive (D, arrow) or CD68- andcathB-positive (E, arrows), whereas others expressed only CD68 (D,
star). A: Anti-CD68, �400, diaminobenzidine, brown, case 29; B: anti-cathH, �400, Fast Red, case 29; C: anti-cathB, �400, Fast Red, case29; D: double staining anti-cathH/anti-CD68, �400, case 29; E: doublestaining anti-cathB/anti-CD68, �400, case 29.
Fig. 3. Canalicular phase was characterized by a further spreading ofthe bronchial tree (A), a more ubiquitous expression of cathK in interstitialcells was found (B). In situ hybridization analyses supported the predom-inant expression of this protease in bronchial epithelial cells (C). In thesaccular/early alveolar phase (D), the pattern of the cathK (E), cathH (F,G), and cathS (I) was similar to adult lung tissue. Both cathH andpro–SP-C were found in single cells of terminal sacs (G, H arrows).
Additionally, a strong cathH immunoreactivity was found in macrophages(F, G stars). A: Hematoxylin and eosin, �200, case 17; B: anti-cathK,�400, case 17; C: cathK antisense probe, �200, case 19; D: hematox-ylin and eosin, �200, case 33; E: anti-cathK, �250, case 33; F: anti-cathH, �250, case 33; G: anti-cathH, �400, case 33; H: anti–pro-SP-C,�400, case 33; I: anti-cathS, �400, case 33.
17CATHEPSINS IN FETAL LUNG DEVELOPMENT

unchanged during organogenesis, we found a continu-ous decrease in the number of cells expressing cathHand an increase in the level of cathS-positive cells(Table 1).
DISCUSSION
The aim of this study was to characterize the courseof the expression of different cysteine proteases in hu-man lung tissues during fetal development. The studywas retrospective and included specimen from 34aborted fetuses with a gestational age between 10 and37 weeks, which corresponds to the fetal developmentfrom glandular to late saccular/early alveolar phases.
Lysosomal proteases, such as cysteine proteases, cat-alyze the processing of endocytosed proteins. Thus,they are involved in the degradation of foreign sub-stances. Bronchial epithelial cells are the first cells ofthe lung to be in contact with the environment; there-fore, they play an important role in first-line defensemechanisms. Respiratory movements occur even dur-ing fetal development (Duenhoelter and Pritchard,1977) and lead to an aspiration and expiration of am-niotic fluid. This process was shown to be important fornormal lung development (Losa and Kind, 1998). Onthe other hand, aspiration of amniotic fluid involvesthe risk of exposing the lung tissue to infectious orhazardous agents. In animal models, it was shown thatthe intra-amniotic deposition of endotoxin leads to analtered lung development (Duenhoelter and Pritchard,1977; Bry and Lappalainen, 2001). The present studydemonstrates that cathepsins B, H, and K are ex-pressed primarily in bronchial epithelial cells; thisfinding may reflect their involvement in the processingof foreign proteins. Because immunostaining was de-tected in differentiated epithelial cells of developingairways with lumen, it remains to be clarified whethercathB, cathH, or cathK are involved in lumen forma-tion. Furthermore, the developmental pattern of theseenzymes illustrates the differentiation of the lysosomalapparatus and suggests that it first acquires functionalactivity in bronchial epithelial cells (Bertini et al.,1981).
With respect to cathK, recently we described its ex-pression in embryonic and fetal epithelial cells of par-enchymatous organs, including lung, kidney, and liver(Haeckel et al., 1999). Now we have shown that thisenzyme is one of the first cathepsins to be expressed inbronchial epithelial cells and that a strong correlationexists between the cathK expression and the presenceof differentiated epithelial cells in more central areas ofthe bronchial tree. These results support our recentreport that demonstrated a significant amount of theenzyme in bronchial epithelial cells of adult lung tissue(Buhling et al., 1999b). Additionally, we found thatbronchial carcinoma cells express significantly loweramounts of cathK in comparison to normal epithelialcells of the same tissue (Buhling, unpublished obser-vations). This finding suggests that the function of thisenzyme is related, at least in part, to environmental
components and its up-regulation corresponds to con-tact of the epithelial cells with the extracorporal space.In contrast to cathK, we found cathB and cathH immu-nostaining not only in epithelial cells, but also in mac-rophages. Therefore, it could be suggested that theseproteases have, independently of the cell type, more ofa “housekeeping” function probably in lysosomal pro-tein degradation. Previously, it was reported that thepromoter structure of the cathB gene exhibits “house-keeping-gene” characteristics that lead to a ubiquitous,weakly regulated protein expression (Berquin et al.,1999), whereas the cathK gene includes several puta-tive transcription regulatory elements (Li and Chen,1999). Continuous production of cathepsins in most ofthe cells was not unexpected, because it was shownthat up to 50% of the lysosomal protein turnover de-pends on cysteine proteases (Bohley and Seglen, 1992).
Lysosomal cysteine proteases may participate in deg-radation of extracellular matrix proteins. Embryonicand fetal development is characterized by rapid remod-eling of the lung tissue, and the most important cells inthese processes are interstitial macrophages; morethan 50% of the CD68-positive interstitial histiocyticcells expressed cathB, cathH, and cathL. Although theexpression of cathH declined during fetal development,increased expression of cathS was detected in the ter-minal saccular phase. The shift in the expression pat-tern of lysosomal proteases may reflect the change ofsubstrates that are degraded during development. Thisshift supports recent findings that indicate a specificrole for single lysosomal proteases in the breakdown ofdefined proteins, independent of the high redundancyin their substrate specificity. These results also agreewith previous reports that demonstrate functionalchanges in interstitial macrophages, for example, asignificant increase in both enzymatic activity and thecapacity to incorporate foreign particles at later phasesof gestation until the first week of life (Shin-Buehringet al., 1980; Bertini et al., 1981).
CathL and cathS were shown to be involved in theprocessing of the invariant chain of the MHC class IIcomplex (Nakagawa et al., 1998; Shi et al., 1999). Pre-vious studies have shown that the expression of MHCclass II molecules in human fetal tracheal tissues wasfound only in late phases of gestation or in postnatalspecimens (Stoltenberg et al., 1993). According to theseresults, the late appearance of cathS could direct theup-regulation of MHC class II expression and, thus,point to its specific role in the processing of these struc-tures.
Lysosomal proteases in lung epithelial cells may alsofunction in the processing of secreted proteins. Surfac-tant proteins are among the most important proteinsreleased by alveolar type II cells. The small surfactantproteins B and C (SP-B, SP-C) are released from largeprecursor molecules by proteolytic processing in themultivesicular and the lamellar bodies (Korimilli et al.,2000). In adult lung tissues, a significant expression ofcathH in type II cells was shown (Ishii et al., 1991), as
18 BUHLING ET AL.

well as a colocalization with surfactant proteins B andC (Brasch et al., 2002). In the fetal tissues analyzed inthis study, we observed distinct cathH immunostainingin several basal epithelial cells during glandular phaseand, in later phases, a distinct staining of single cells inthe terminal sacs. It was reported that type II pneu-mocytes develop from cells within the spreading bron-chial tree; therefore, we hypothesized that these cellscould be the precursors of type II cells and performedparallel immunostainings by using an antiserum spe-cific for pro–SP-C. Indeed, we found a similar stainingpattern. A similar pro–SP-C immunostaining of scat-tered epithelial cells in fetal lung tissues at the glan-dular phase of development and later of type II pneu-mocyte-like cells was described by Khoor et al. (Khooret al., 1994). However, they described a more generalimmunoreactivity in epithelial cells. Nevertheless,these results support our hypothesis that cathH maybe involved in the earlier steps of surfactant process-ing.
In conclusion, we describe the expression pattern ofcathepsins B, H, L, K, and S during the human fetallung development. The time course of the expression indifferent cell types during gestation and the specific
expression of single enzymes such as cathK and cathSin defined cells are highly suggestive of a defined rolefor these proteases. Further studies using cathepsin-deficient animals are required to investigate the func-tion of single cathepsins in the lung development. Al-though cathepsins are characterized by highredundancy in enzyme specificity and, therefore, trans-genic animals deficient in a single cathepsin did notexhibit any life-threatening defects of lung develop-ment (Deussing et al., 1998; Nakagawa et al., 1998;Saftig et al., 1998), this situation could be changedunder pathologic conditions, for example, intrauterineinfections.
EXPERIMENTAL PROCEDURESSelection of Fetal Material
Retrospectively, 34 fetuses with a gestational agebetween 10 and 37 weeks were selected from the ar-chive of the Institute of Pathology. These had beenlogged in the archive between 1996 and 1999. Wherepossible, two samples (Table 2) aborted during a par-ticular week of gestation were selected. The gestationalage of all the selected fetuses was determined indepen-dently by gynecologists and pathologists, and only
TABLE 2. Fetal Specimens Analyzed for Cathepsin Expressiona
Patient GA Sex Reason for pregnancy termination1 10 n.d. Induced abortion for psychosocial reasons2 10 f Induced abortion for psychosocial reasons3 12 m Induced abortion for psychosocial reasons4 12 m Induced abortion for psychosocial reasons5 13 f Induced abortion for medical reasons, intrauterine infection with Toxoplasma gondii6 13 m Induced abortion for medical reasons7 13 f Induced abortion for medical reasons8 14 m Induced abortion for medical reasons, cranioschisis9 14 m Induced abortion for medical reasons, psychiatric disease of the mother
10 15 m Induced abortion for for medical reasons11 15 m Induced abortion, rubella infection12 16 m Induced abortion for medical reasons, Potter sequence13 17 m Induced abortion for medical reasons, Potter sequence14 17 f Spontaneous abortion, placentitis15 18 f Spontaneous abortion, placentitis16 18 m Spontaneous abortion17 19 m Induced abortion, Klinefelter syndrome18 19 m Spontaneous abortion, premature placental separation19 20 m Induced abortion for medical reasons, Potter sequence20 20 m Induced abortion, Klinefelter syndrome21 21 m Induced abortion for medical reasons22 21 f Induced abortion for medical reasons23 21 m Spontaneous abortion, Trisomy 924 22 f Spontaneous abortion25 23 f Spontaneous abortion26 23 f Spontaneous abortion, trisomy 2127 24 f Spontaneous abortion, trisomy 1328 25 f Spontaneous abortion, preterm placental separation29 26 m Spontaneous abortion, preterm placental separation30 26 f Spontaneous abortion, pre-eclampsia31 28 f Spontaneous abortion, preterm placental separation32 33 f Preterm birth, infection with Klebsiella spp33 36 m Intrauterine death34 37 m Intrauterine death, premature placental separation, placental insufficiency
aGA, gestational age (weeks); n.d., not determined; f, female; m, male.
19CATHEPSINS IN FETAL LUNG DEVELOPMENT

those with matched ages in both analyses were in-cluded. Those cases with extensive autolysis, as deter-mined independently by two observers after hematox-ylin and eosin staining, were excluded. Fetuses withaberrations in lung development caused by genetic al-terations were also excluded, and twins were excludeddue to a possible delay in their intrauterine develop-ment. The cases that were included are described inTable 2.
Immunohistochemistry
Immunostaining was performed on serial sections offetal lung tissue by using monoclonal antibodies di-rected against cathepsin B (cathB; Calbiochem; IM27,80 ng/ml), cathepsin L (cathL; 33/2, 40 ng/ml) (Weber etal., 1997), CD68, anti-vimentin, anti-cytokeratin 19 (70ng/ml, all from DAKO, Denmark), and with polyclonalantibodies directed against cathepsin H (cathH;1:3,000), cathepsin S (cathS; 1:800; both were kindlyprovided by H. Kirschke, Turku, Norway), cathepsin K(cathK; 1:1,500; kindly provided by D. Bromme, NewYork) (Bromme et al., 1996), and pro-surfactant proteinC (pro–SP-C, kindly provided by J.J. Batenburg,Utrecht, The Netherlands) as described earlier (Buh-ling et al., 1999b). The antisera directed against cathe-psins recognized both pro- and mature enzymes. Anti-CD68 and anticytokeratin antibodies were used foridentification of myeloid and epithelial cells, respec-tively. Anti–pro-SP-C was used for detection of surfac-tant-producing cells. Biotin-conjugated anti-mouse andanti-rabbit sera (1:200, both from Vector Laboratories)were used as secondary antibodies. Before immuno-staining, the specimens were boiled in citrate buffer for5 min (cathB, cathL, and CD68, cytokeratin). Immuno-reactions were visualized with the avidin-biotin com-plex method applying a Vectastain ABC alkalinephosphatase kit or a Vectastain ABC peroxidase kit(Vector Laboratories). Fast red or 3,3�-diaminobenzi-dine (DAB) was used as a substrate. The sections werecounterstained with hematoxylin and mounted in gel-atin. The specificity of immunostaining was tested (1)by omitting the primary antibody, (2) by substitution ofthe first antibody with an irrelevant antibody or anonimmune serum, or (3) by preincubating the anti-serum with saturating amounts of purified antigen.The stained sections were examined independently bytwo observers using a conventional light microscope(Axioskop, Zeiss, Jena, Germany). The entire sectionswere examined thoroughly, including inspection of atleast 20 high-power fields (magnification, � 400) perslide, and representative areas were documented. Therelative amounts of cathepsin-expressing macrophageswere calculated as follows: number of interstitial cellswith cathepsin-specific immunoreactivity/number ofCD68-positive interstitial cells. If possible, 20 high-power fields were examined in the same region of serialsections. The whole section was examined if less than100 cells were found in this area or if the entire sectionwas smaller than the area of 20 high-power fields. This
was the case in some samples obtained from fetusesaborted during the earlier gestational period.
In Situ Hybridization
In three tissue samples, cathB, cathK, cathL, andcathS mRNA was detected by using in situ hybridiza-tion, which was performed as described previously(Haeckel et al., 1999). The following cDNA fragmentswere used for generation of RNA probes: cathB, 464 bplocalized at position 240-704 of the coding sequence;cathH, 300 bp localized at position 486-786; cathK, 361bp localized at position 172-533; cathL, 324 bp localizedat position 264-588; and cathS, 414 bp localized atposition 262-676 with no or minimal sequence homol-ogies to other cathepsins used. Sense and antisensemRNA probes were labeled by in vitro transcriptionwith digoxigenin-labeled UTP (DIG-labeling kit, RocheDiagnostics, Germany). Formalin-fixed, paraffin-em-bedded tissue sections were deparaffinized and rehy-drated under RNAse-free conditions, post-fixed inparaformaldehyde, and acetylated with acetic anhy-dride. Before prehybridization, the tissues were perme-abilized with 24 �g/ml proteinase K (Sigma) at 37°C for10 min. Prehybridization was performed by using hy-bridization buffer (4� saline-sodium citrate buffer,50% deionized formamide, 5% dextran sulfate, 1� Den-hardt’s solution, 10 mM dithiothreitol, and 0.5 mg/mlsalmon sperm; all from Sigma). For hybridization, 200ng of DIG-labeled probe (sense or antisense) and 1 �l oft-RNA was added and incubated for 18 hr at 55°C.Detection was performed by using anti-DIG antibody(Roche Diagnostics, Germany) and a commerciallyavailable avidin-biotin complex method according tothe manufacturer’s instructions (Vectastain, VectorLaboratories). Fast Red was used as a substrate. Fordetection of cathB, cathL, and cathS mRNA, a commer-cially available tyramine-based signal amplificationsystem (TSA, Perkin Elmer) and DAB with nickel chlo-ride (Vector Laboratories) as a substrate resulting in agray-black precipitate was used. The sections werecounterstained with Mayer’s hematoxylin and embed-ded in gelatin.
ACKNOWLEDGMENTS
The authors thank Ms. Marianne Blichmann andMs. Yvonne Peter for skilled technical assistance.
REFERENCES
Adamson IYR. 1997. Development of lung structure. In: Crystal RG,West BB, editors. The lung. Philadelphia: Lippinscottt, Raven. p993.
Berquin IM, Sloane BF. 1996. Cathepsin B expression in humantumors. Adv Exp Med Biol 389:281–294.
Berquin IM, Yan S, Katiyar K, Huang L, Sloane BF, Troen BR. 1999.Differentiating agents regulate cathepsin B gene expression inHL-60 cells. J Leukoc Biol 66:609–616.
Bertini F, Mayorga L, Gonzalez M. 1981. The development of lysoso-mal apparatus. II. Incorporation, subcellular distribution, and in-traparticulate hydrolysis of 131 I-albumin by liver of mice at peri-natal stages. J Cell Physiol 109:281–287.
20 BUHLING ET AL.

Bohley P, Seglen PO. 1992. Proteases and proteolysis in the lysosome.Experientia 48:151–157.
Brasch F, ten Brinke A, Johnen G, Ochs M, Kapp N, Muller KM,Beers MF, Fehrenbach H, Richter J, Batenburg JJ, Buhling F.2002. Involvement of cathepsin H in the processing of the hydro-phobic surfactant associated protein C in type-II-pneumocytes.Am J Respir Cell Mol Biol 26:659–670.
Bromme D, Okamoto K, Wang BB, Biroc S. 1996. Human cathepsinO2, a matrix protein-degrading cysteine protease expressed in os-teoclasts. Functional expression of human cathepsin O2 in Spodopt-era frugiperda and characterization of the enzyme. J Biol Chem271:2126–2132.
Bry K, Lappalainen U. 2001. Intra-amniotic endotoxin accelerateslung maturation in fetal rabbits. Acta Paediatr 90:74–80.
Buhling F, Gerber A, Ansorge S, Welte T. 1999a. Katheptische Zys-teinproteinasen in der Lunge. Pneumologie 53:400–407.
Buhling F, Gerber A, Hackel C, Kruger S, Kohnlein T, Bromme D,Reinhold D, Ansorge S, Welte T. 1999b. Expression of cathepsin Kin lung epithelial cells. Am J Cell Mol Biol 20:612–619.
Daston GP, Baines D, Yonker JE, Lehman-McKeeman LD. 1991.Effects of lysosomal proteinase inhibition on the development of therat embryo in vitro. Teratology 43:253–261.
Deussing J, Roth W, Saftig P, Peters C, Ploegh HL, Villadangos JA.1998. Cathepsins B and D are dispensable for major histocompati-bility complex class II-mediated antigen presentation. Proc NatlAcad Sci U S A 95:4516–4521.
Docherty K, Hutton JC, Steiner DF. 1984. Cathepsin B-related pro-teases in the insulin secretory granule. J Biol Chem 259:6041–6044.
Driessen C, Bryant RA, Lennon D, Villadangos JA, Bryant PW, ShiGP, Chapman HA, Ploegh HL. 1999. Cathepsin S controls thetrafficking and maturation of MHC class II molecules in dendriticcells. J Cell Biol 147:775–790.
Duenhoelter JH, Pritchard JA. 1977. Fetal respiration. A review.Am J Obstet Gynecol 129:326–338.
Haeckel C, Krueger S, Buehling F, Broemme D, Franke K, SchuetzeA, Roese I, Roessner A. 1999. Expression of cathepsin K in thehuman embryo and fetus. Dev Dyn 216:89–95.
Inui T, Ishibashi O, Inaoka T, Origane Y, Kumegawa M, Kokubo T,Yamamura T. 1997. Cathepsin K antisense oligodeoxynucleotideinhibits osteoclastic bone resorption. J Biol Chem 272:8109–8112.
Ishii Y, Hashizume Y, Kominami E, Uchiyama Y. 1991. Changes inimmunoreactivity for cathepsin H in rat type II alveolar epithelialcells and its proteolytic activity in bronchoalveolar lavage fluid over24 hours. Anat Rec 230:519–523.
Katunuma N, Kominami E. 1987. Abnormal expression of lysosomalcysteine proteinases in muscle wasting diseases. Rev Physiol Bio-chem Pharmacol 108:1–20.
Khoor A, Stahlman MT, Gray ME, Whitsett JA. 1994. Temporal-spatial distribution of SP-B and SP-C proteins and mRNAs indeveloping respiratory epithelium of human lung. J HistochemCytochem 42:1187–1199.
Kirschke H, Wiederanders B. 1987. Lysosomal proteinases. Acta His-tochem 82:2–4.
Korimilli A, Gonzales LW, Guttentag SH. 2000. Intracellular local-ization of processing events in human surfactant protein B biosyn-thesis. J Biol Chem 275:8672–8679.
Lemere CA, Munger JS, Shi GP, Natkin L, Haass C, Chapman HA,Selkoe DJ. 1995. The lysosomal cysteine protease, cathepsin S, isincreased in Alzheimer‘s disease and Down syndrome brain. Animmunocytochemical study. Am J Pathol 146:848–860.
Li YP, Chen W. 1999. Characterization of mouse cathepsin K gene,the gene promoter, and the gene expression. J Bone Miner Res14:487–499.
Losa M, Kind C. 1998. Dry lung syndrome: complete airway collapsemimicking pulmonary hypoplasia? Eur J Pediatr 157:935–938.
Mort JS, Recklies AD, Poole AR. 1984. Extracellular presence of thelysosomal proteinase cathepsin B in rheumatoid synovium and itsactivity at neutral pH. Arthritis Rheum 27:509–515.
Nakagawa T, Roth W, Wong P, Nelson A, Farr A, Deussing J, Vil-ladangos JA, Ploegh H, Peters C, Rudensky AY. 1998. Cathepsin L:critical role in Ii degradation and CD4 T cell selection in the thy-mus. Science 280:450–453.
Riese RJ, Chapman HA. 2000. Cathepsins and compartmentalizationin antigen presentation. Curr Opin Immunol 12:107–113.
Roth W, Deussing J, Botchkarev VA, Pauly-Evers M, Saftig P, HafnerA, Schmidt P, Schmahl W, Scherer J, Anton-Lamprecht I, VonFigura K, Paus R, Peters C. 2000. Cathepsin L deficiency as molec-ular defect of furless: hyperproliferation of keratinocytes and per-tubation of hair follicle cycling. FASEB J 14:2075–2086.
Saftig P, Hunziker E, Wehmeyer O, Jones S, Boyde A, RommerskirchW, Moritz JD, Schu P, Von Figura K. 1998. Impaired osteoclasticbone resorption leads to osteopetrosis in cathepsin-K-deficient mice.Proc Natl Acad Sci U S A 95:13453–13458.
Samarel AM, Ferguson AG, Decker RS, Lesch M. 1989. Effects ofcysteine protease inhibitors on rabbit cathepsin D maturation.Am J Physiol 257:C1069–C1079.
Shi GP, Villadangos JA, Dranoff G, Small C, Gu LJ, Haley KJ, RieseR, Ploegh HL, Chapman HA. 1999. Cathepsin S required for normalMHC class II peptide loading and germinal center development.Immunity 10:197–206.
Shi GP, Bryant RA, Riese R, Verhelst S, Driessen C, Li Z, Bromme D,Ploegh HL, Chapman HA. 2000. Role for cathepsin F in invariantchain processing and major histocompatibility complex class II pep-tide loading by macrophages. J Exp Med 191:1177–1186.
Shin-Buehring YS, Dallinger M, Osang M, Rahm P, Schaub J. 1980.Lysosomal enzyme activities of human fetal organs during develop-ment. Biol Neonate 38:300–308.
Smith WW, Abdelmeguid SS. 1999. Cathepsin K as a target for thetreatment of osteoporosis. Expert Opin Ther Patients 9:683–694.
Stoltenberg L, Thrane PS, Rognum TO. 1993. Development of im-mune response markers in the trachea in the fetal period and thefirst year of life. Pediatr Allergy Immunol 4:13–19.
Turk B, Turk V, Turk D. 1997. Structural and functional aspects ofpapain-like cysteine proteinases and their protein inhibitors. BiolChem 378:141–150.
Weber E, Bahn H, Gunther D. 1997. Monoclonal antibodies againstcathepsin L and procathepsin L of different species. Hybridoma16:159–166.
21CATHEPSINS IN FETAL LUNG DEVELOPMENT
Recommended

















