
Histochem Cell Biol
DOI 10.1007/s00418-007-0372-9ORIGINAL PAPER
Expression of glucose transporters GLUT-1, GLUT-3, GLUT-9 and HIF-1� in normal and degenerate human intervertebral disc
S. M. Richardson · R. Knowles · J. Tyler · A. Mobasheri · J. A. Hoyland
Accepted: 9 December 2007© Springer-Verlag 2007
Abstract The glucose transporters GLUT-1 and GLUT-3are targets of the hypoxia-inducible transcription factorHIF-1� and it has been shown that nucleus pulposus (NP)cells in rat intervertebral discs (IVD) express both HIF-1�and GLUT-1. However, there is limited data on the expres-sion of HIF-1� and GLUTs in human IVD. The aim herewas to (1) determine whether, like articular chondrocytes,human IVD cells express GLUT-1, 3 and 9 and whetherthere was any co-expression with HIF-1�; and (2) to local-ise expression of the GLUT isoforms in the disc and iden-tify any changes during degeneration. Real-time PCR wasused to identify expression of GLUT1, 3, 9 and HIF-1�mRNAs and immunohistochemistry was used to analyseprotein expression and localisation of GLUTs in normaland degenerate IVD biopsies. Results conWrmed HIF-1�,GLUT1, 3 and 9 mRNA expression in NP and AF and co-expression of each GLUT isoform with HIF-1� in the NP,but not the AF. Immunohistochemistry demonstratedregional diVerences in GLUT expression, with the highestexpression being in the NP. GLUT expression also changedas degeneration progressed. This study demonstrates that
NP and AF cells have diVerent GLUT expression proWlesthat suggest regional diVerences in the metabolic nature ofthe human IVD and that this environment changes duringdegeneration.
Keywords Intervertebral disc · Degeneration · Hypoxia · HIF-1� · GLUT-1 · GLUT-3 · GLUT-9 · Immunohistochemistry · Real-time PCR
Introduction
Degenerative disorders of the intervertebral disc (IVD) andarticular cartilage are generally characterised by disequilib-rium between extracellular matrix repair and degradativeprocesses (Le Maitre et al. 2004; Freemont et al. 2002).Molecular alterations include elevated matrix metallopro-teinase (Goupille et al. 1998; Crean et al. 1997) and aggre-canase activity (Pockert et al. 2006), increased expressionof (and sensitivity to) catabolic cytokines (Le Maitre et al.2005) and disc cell senescence (Le Maitre et al. 2007) andapoptosis (Gruber and Hanley 1998). These changes lead tonarrowing of the disc space (Yasuma et al. 1990) andmechanical failure of the disc (Thompson et al. 2000; Iatri-dis et al. 1999). Cell density in the fully developed, healthyIVD is very low (approximately 4,000 mm¡3 in the NP)(Maroudas et al. 1975) and the poor diVusion of nutrients(i.e. O2 and glucose) and accumulation of metabolic wasteproducts such as lactate (Ohshima and Urban 1992; Bartelset al. 1998) present disc cells with further environmentalchallenges because of the absence of microvasculature(Repanti et al. 1998). Normally the cells of the outer annu-lus Wbrosus (OAF) are supplied with nutrients by the bloodvessels within the outer surface of the AF, but the cells ofthe inner annulus Wbrosus (IAF) and nucleus pulposus (NP)
S. M. Richardson · R. Knowles · J. Tyler · J. A. Hoyland (&)Tissue Injury and Repair Group, Clinical and Laboratory Sciences, Faculty of Medical and Human Sciences, The University of Manchester, Manchester M13 9PT, UKe-mail: [email protected]
S. M. Richardsone-mail: [email protected]
A. MobasheriDivision of Veterinary Medicine, School of Veterinary Medicine and Science, University of Nottingham, Sutton Bonington Campus, College Road, Sutton Bonington, Leicestershire LE12 5RD, UK
123

Histochem Cell Biol
may be up to 8 mm away from the nearest capillary andtheir nutrients must be supplied by diVusion through thedisc matrix from blood vessels that terminate in the verte-brae above and below the disc (Urban et al. 2001; Bibbyet al. 2001).
Studies on the metabolism of the IVD and articular carti-lage suggest that these connective tissues are metabolicallyand bioenergetically diVerent (Lee and Urban 1997 and2002). In the IVD, anoxia and the use of glycolysis inhibi-tors results in a progressive “positive Pasteur eVect” sug-gesting that a large proportion of the IVD’s energy isderived from oxidative phosphorylation (Ishihara andUrban 1999). This situation is the opposite of that seen inarticular cartilage in which anoxia severely inhibits glucoseuptake and lactate production and the decrease in lactateformation correlates well with decreased glucose uptake bychondrocytes. This reduction in the rate of glycolysis inanoxic conditions is seen as evidence for a “negative Pas-teur eVect” in cartilage (Lee and Urban 1997). These stud-ies have led to suggestions that cell metabolism iscompromised by nutrient and oxygen deprivation aVectingenergy transduction in the disc.
Recent studies on the basic cell biology of rat disc cellshave constructed a “metabolic” and “phenotypic signature”for cells of the NP based on the hypothesis that in responseto oxygen and nutrient deprivation NP cells express threekey phenotypic markers: hypoxia-inducible factor alpha(HIF-1�), glucose transporter 1 (GLUT-1) and matrixmetalloproteinase-2 (MMP-2) (Rajpurohit et al. 2002)which enables them to be distinguished from surroundingtissues such as the IAF and OAF. HIF-1� has also beenrecently demonstrated in the NP of normal human IVD (Haet al. 2006). The presence of these proteins in the NP hashinted that cells in avascular and hypoxic regions of thedisc are capable of adaptive responses to low oxygen ten-sions and nutrient deprivation. An integral part of the meta-bolic adaptation to hypoxia is activation of genes involvedin promoting anaerobic glycolysis (i.e. hypoxia responsiveGLUT-1 and GLUT-3 facilitative glucose transporters)(Vannucci et al. 1998), lactate dehydrogenase (LDH) andphosphofructokinase (PFK) (Semenza et al. 1994, 1996;Richardson et al. 2003).
Based on the above observations and our recent studiesof glucose transport in articular chondrocytes (Richardsonet al. 2003; Mobasheri et al. 2002a) we hypothesised thathypoxia responsive glucose transporters GLUT-1 andGLUT-3 are expressed in IVD cells and their expression isincreased in disc degeneration via activation of the HIF-1�transcription factor. We also postulated that other GLUTisoforms (glucose transporters that have not yet been pro-posed to be regulated by hypoxia, i.e. GLUT-9) might benecessary for disc cell hexose transport as has been shownto be the case for articular chondrocytes (Mobasheri et al.
2002a, b; Shikhman et al. 2001, 2004). The overall objec-tives of the present study were to test these proposedhypotheses by: (1) identifying expression of HIF-1� and theglucose transporter isoforms in normal human NP and AFby quantitative, real-time RT-PCR; (2) determiningwhether HIF-1� mRNA expression correlates with GLUT-1, 3 or 9 mRNA expression in human NP and AF by quanti-tative real-time PCR; (3) localising expression of GLUT-1,3 and 9 protein within regions of normal human IVD usingimmunohistochemistry; (4) determining whether GLUTprotein expression changes with degeneration in eachregion of the human IVD.
Materials and methods
Human tissues for real-time PCR study
Ten NP and eight AF biopsies were taken from IVDs ofpatients with chronic lower back pain undergoing surgery.Informed consent and local medical ethics committeeapproval were obtained. For each patient, the disc removedhad been demonstrated to be the origin of their pain by dis-cography. Two normal NP and AF samples (which showedno evidence of degeneration based on the system describedby Sive et al. 2002) were obtained from IVD tissueremoved during post-mortem examination. Informed con-sent from relatives and local Ethical Committee approvalwas obtained for the use of the samples for research.
Quantitative real-time PCR
To investigate expression of GLUT and HIF-1�, mRNAwas analysed using quantitative real-time PCR. RNA wasextracted from homogenised biopsy material using a stan-dard TRIzol (Invitrogen) protocol and cDNA generatedusing the Superscript II (Invitrogen) protocol as previouslydescribed (Richardson et al. 2006a, b). Gene expressionwas analysed by real-time PCR using an ABI Prism 7,000sequence detection system (Applied Biosystems). Primersets (Sigma-Genosys) were designed for GLUT-1 (F: ACCATTGGCTCCGGTATCG; R: GCTCGCTCCACCACAAACA), GLUT-3 (F: GGAAAGGGCAGGAAGAAGGA; R: ACAGTCATGAGCGTGGAACAAA), GLUT-9(F: GTCTTCACTGGGCAGCTTCTG; R: CAAACAGGTATGGCCAGGTACTC) and HIF-1� (F: GTCGCTTCGGCCAGTGTG; R: GGAAAGGCAAGTCCAGAGGTG). Reactions were performed in duplicate as previouslydescribed (Richardson et al. 2006a, b). A melt-curve analy-sis was also conducted for each primer set to conWrm a sin-gle PCR product. Results were analysed using the 2¡�Ct
method and presented as relative expression normalised toGAPDH (Applied Biosystems).
123

Histochem Cell Biol
Statistical analysis of real-time PCR data
Real-time PCR results were analysed using non-parametriclinear regression analysis to investigate correlationsbetween expression of each GLUT and HIF-1� in both NPand AF tissues.
Human tissues for immunohistochemistry
Thirty-one biopsies were taken from IVDs of patients withchronic lower back pain with informed consent and localmedical ethics committee approval. Disc biopsies wereWxed in 10% v/v neutral buVered formalin and embedded inparaYn wax. Positive control tissues were represented inhuman tissue microarrays (TMAs) that were obtained fromthe Cooperative Human Tissue Network (CHTN) of theNational Cancer Institute (NCI), the National Institutes ofHealth, Bethesda, MD (http://faculty.virginia.edu/chtn-tma/home.html). The TMAs contained formalin-Wxed paraYnembedded samples of 66 non-neoplastic adult tissuesincluding renal cortex, renal medulla and brain (includingthe following regions: cerebral cortex, cerebellar cortex,choroid plexus, hippocampus and meninges).
Tissue biopsy classiWcation
DeparaYnised 7 �m sections of disc tissue biopsies werestained with hematoxylin and eosin (H&E) and examined bylight microscopy. Tissues were graded histologically for fea-tures of degeneration using the grading system reported bySive et al. (2002) and divided into four groups based on thegrade of degeneration. The parameters used for grading werea loss of demarcation between NP and AF, loss of proteogly-cans from the NP, presence and extent of Wssures and cellcluster formation. Standard haematoxylin and eosin stainedsections were analysed independently by two independentobservers and given a score from 0 to 3 for each of theparameters. This gave a total score of between 0 and 12, with0–3 being non-degenerate, 4–6 being intermediate, 7–9 beingdegenerate and 10–12 being severely degenerate. Based onthis classiWcation there were ten samples in the non-degener-ate group, six in the intermediate, eight in the degenerate andseven samples in the severely degenerate group (Table 1).
Immunohistochemistry
Immunohistochemistry was used to determine the expressionof GLUT-1, GLUT-3 and GLUT-9 in the human IVD usinga previously described method on formalin-Wxed, paraYnembedded human cartilage tissues (Richardson et al. 2003).A rabbit polyclonal antibody to the C-terminus of rat GLUT-1 was obtained from Dr. S. Baldwin (University of Leeds,Leeds, UK). A rabbit polyclonal antibody to human GLUT-9
was kindly donated by J. F. Moley, and K. H. Moley (Wash-ington University, St Louis, MO, USA). These antibodieshave been used by numerous investigators in several keypapers (Davies et al. 1987 and 1990; Froehner et al. 1988;Kasanicki et al. 1987; Augustin et al. 2004) and we haverecently used them to conWrm that GLUT-1 and GLUT-9 arepresent in human (Richardson et al. 2003) and ovine (Moba-sheri et al. 2005) articular chondrocytes. A rabbit polyclonalantibody to human GLUT-3 was purchased from ChemiconInternational (AB 1345, Temecula, CA, USA). All antibodies
Table 1 Study population demographics
Details of grade of degeneration, age and gender of patients fromwhom the disc biopsies were obtained
Biopsy number
Grade of degeneration
Age (years)
Gender
Normal
1 0 41 Female
2 0 41 Female
3 0 15 Female
4 1 53 Male
5 1 53 Male
6 2 20 Female
7 2 40 Male
8 3 31 Male
9 3 57 Female
10 3 75 Male
Intermediate
11 5 55 Male
12 5 39 Male
13 5 28 Male
14 6 40 Male
15 6 35 Female
16 6 73 Female
Degenerate
17 7 55 Female
18 7 Unknown Female
19 8 33 Male
20 8 24 Female
21 9 46 Male
22 9 33 Female
23 9 64 Male
24 9 73 Female
Severely degenerate
25 10 32 Male
26 10 68 Male
27 10 45 Female
28 10 47 Male
29 11 45 Male
30 11 47 Male
31 12 39 Male
123
Histochem Cell Biol
were diluted 1:100 and Fast-Red TR/Naphthol AS-MX wasused as the detection step to give red positive staining.
Immunohistochemistry image analysis and statistical analysis
All slides were visualised using a Leica RMDB microscopeand images captured using a digital camera and BioquantNova image analysis system. Each area of the disc (NP,IAF and OAF) was analysed separately, and the number ofimmunopositive cells expressed as a percentage of the 200cells analysed for that area. Data was then plotted asmean § 2 standard errors to represent the 95% conWdenceintervals. As data was non-parametric, the Mann–WhitneyU test was used to compare the numbers of immunopositivecells in each region of degenerate discs compared to thesame region of normal discs. Wilcoxon paired sample testswere also used to compare the number of immunopositivecells in diVerent regions of the same disc, regardless ofgrade of degeneration (Le Maitre et al. 2005).
Results
Correlation of GLUT and HIF-1� mRNA expression in NP and AF tissues
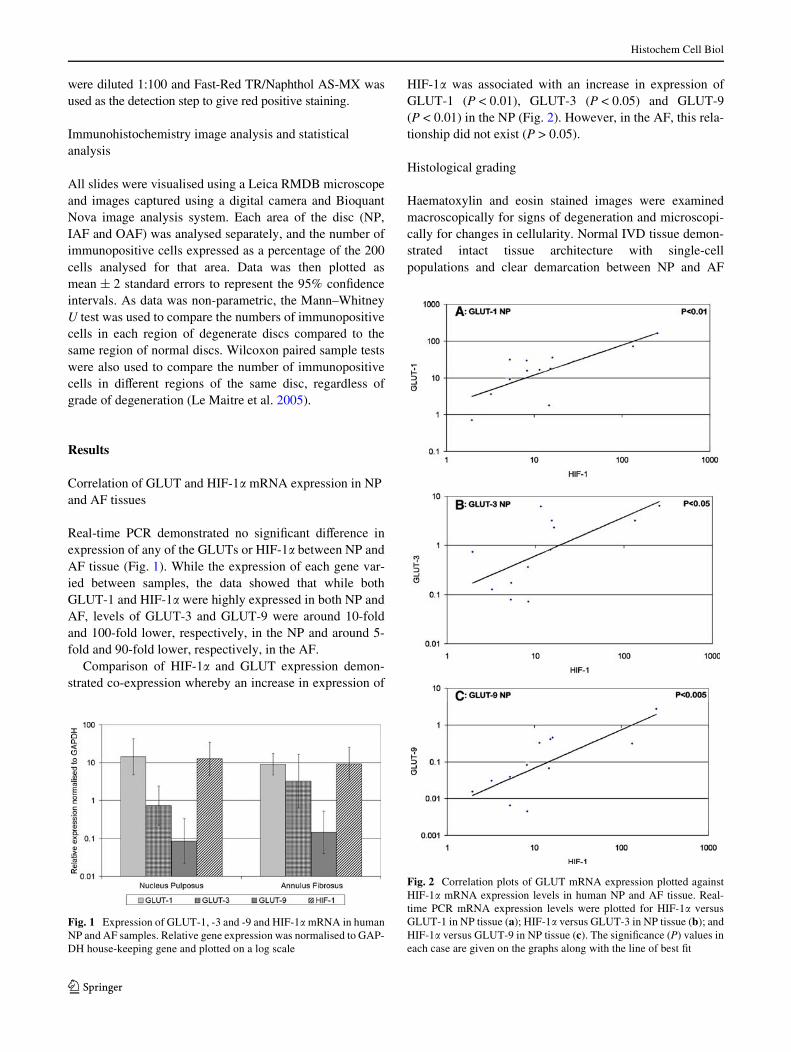
Real-time PCR demonstrated no signiWcant diVerence inexpression of any of the GLUTs or HIF-1� between NP andAF tissue (Fig. 1). While the expression of each gene var-ied between samples, the data showed that while bothGLUT-1 and HIF-1� were highly expressed in both NP andAF, levels of GLUT-3 and GLUT-9 were around 10-foldand 100-fold lower, respectively, in the NP and around 5-fold and 90-fold lower, respectively, in the AF.
Comparison of HIF-1� and GLUT expression demon-strated co-expression whereby an increase in expression of
HIF-1� was associated with an increase in expression ofGLUT-1 (P < 0.01), GLUT-3 (P < 0.05) and GLUT-9(P < 0.01) in the NP (Fig. 2). However, in the AF, this rela-tionship did not exist (P > 0.05).
Histological grading
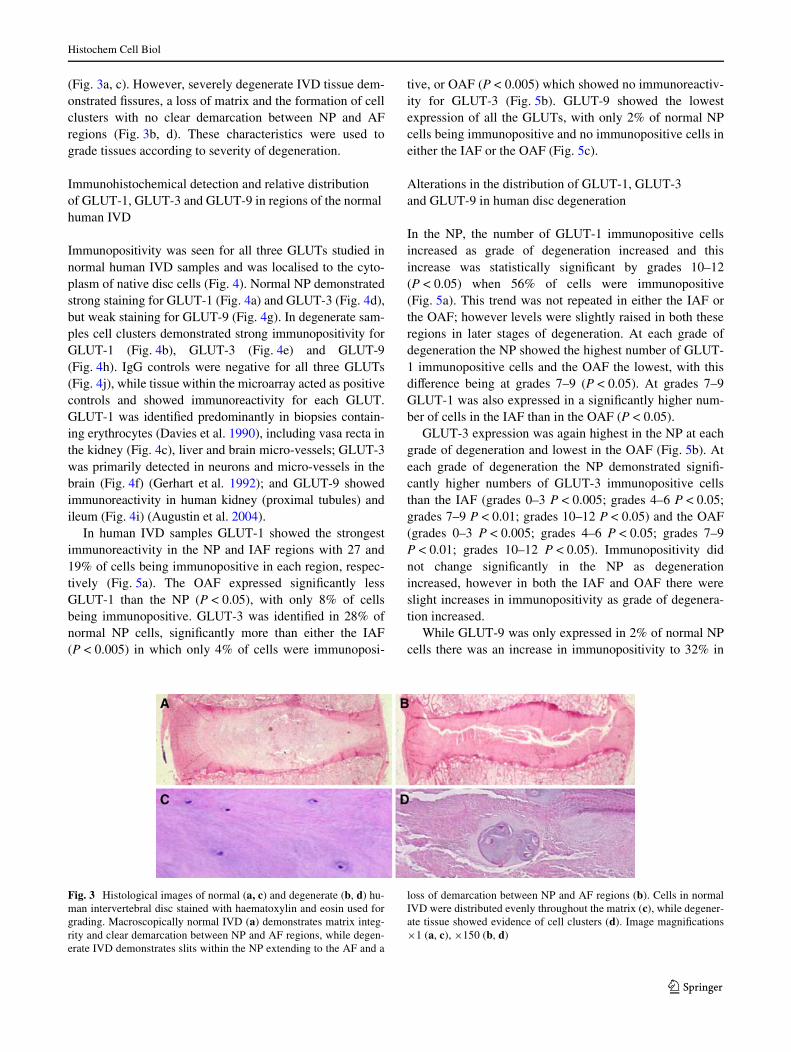
Haematoxylin and eosin stained images were examinedmacroscopically for signs of degeneration and microscopi-cally for changes in cellularity. Normal IVD tissue demon-strated intact tissue architecture with single-cellpopulations and clear demarcation between NP and AF
Fig. 1 Expression of GLUT-1, -3 and -9 and HIF-1� mRNA in humanNP and AF samples. Relative gene expression was normalised to GAP-DH house-keeping gene and plotted on a log scale
Fig. 2 Correlation plots of GLUT mRNA expression plotted againstHIF-1� mRNA expression levels in human NP and AF tissue. Real-time PCR mRNA expression levels were plotted for HIF-1� versusGLUT-1 in NP tissue (a); HIF-1� versus GLUT-3 in NP tissue (b); andHIF-1� versus GLUT-9 in NP tissue (c). The signiWcance (P) values ineach case are given on the graphs along with the line of best Wt
123
Histochem Cell Biol
(Fig. 3a, c). However, severely degenerate IVD tissue dem-onstrated Wssures, a loss of matrix and the formation of cellclusters with no clear demarcation between NP and AFregions (Fig. 3b, d). These characteristics were used tograde tissues according to severity of degeneration.
Immunohistochemical detection and relative distribution of GLUT-1, GLUT-3 and GLUT-9 in regions of the normal human IVD
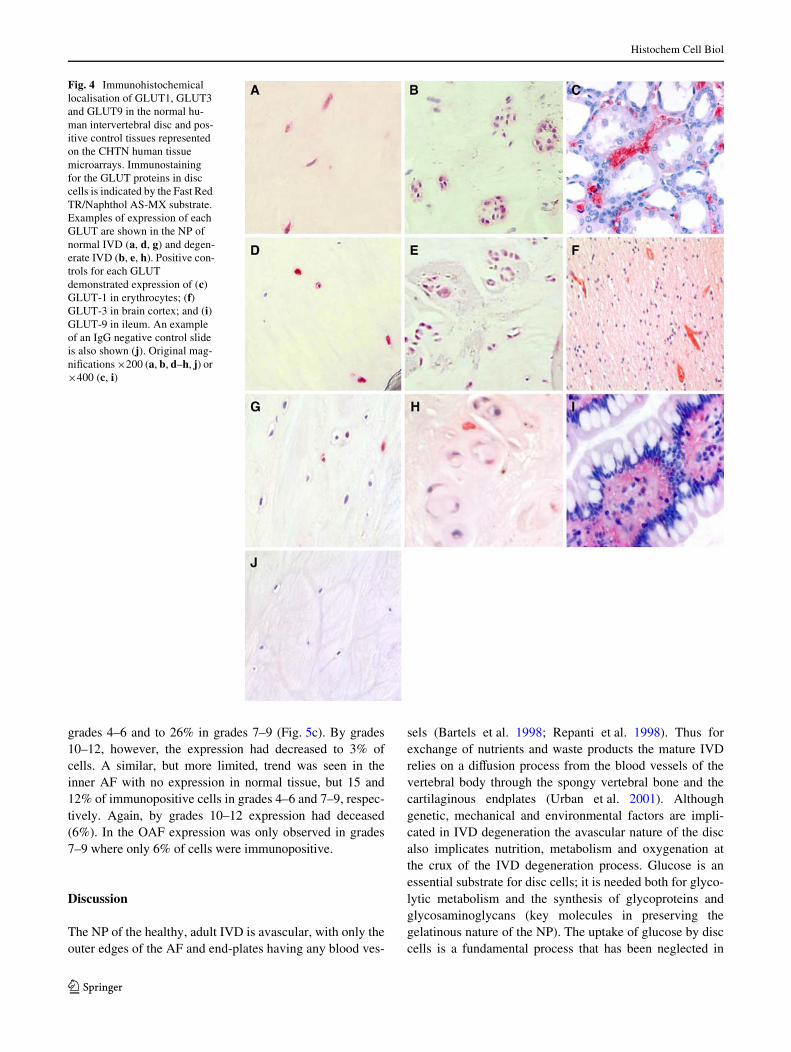
Immunopositivity was seen for all three GLUTs studied innormal human IVD samples and was localised to the cyto-plasm of native disc cells (Fig. 4). Normal NP demonstratedstrong staining for GLUT-1 (Fig. 4a) and GLUT-3 (Fig. 4d),but weak staining for GLUT-9 (Fig. 4g). In degenerate sam-ples cell clusters demonstrated strong immunopositivity forGLUT-1 (Fig. 4b), GLUT-3 (Fig. 4e) and GLUT-9(Fig. 4h). IgG controls were negative for all three GLUTs(Fig. 4j), while tissue within the microarray acted as positivecontrols and showed immunoreactivity for each GLUT.GLUT-1 was identiWed predominantly in biopsies contain-ing erythrocytes (Davies et al. 1990), including vasa recta inthe kidney (Fig. 4c), liver and brain micro-vessels; GLUT-3was primarily detected in neurons and micro-vessels in thebrain (Fig. 4f) (Gerhart et al. 1992); and GLUT-9 showedimmunoreactivity in human kidney (proximal tubules) andileum (Fig. 4i) (Augustin et al. 2004).
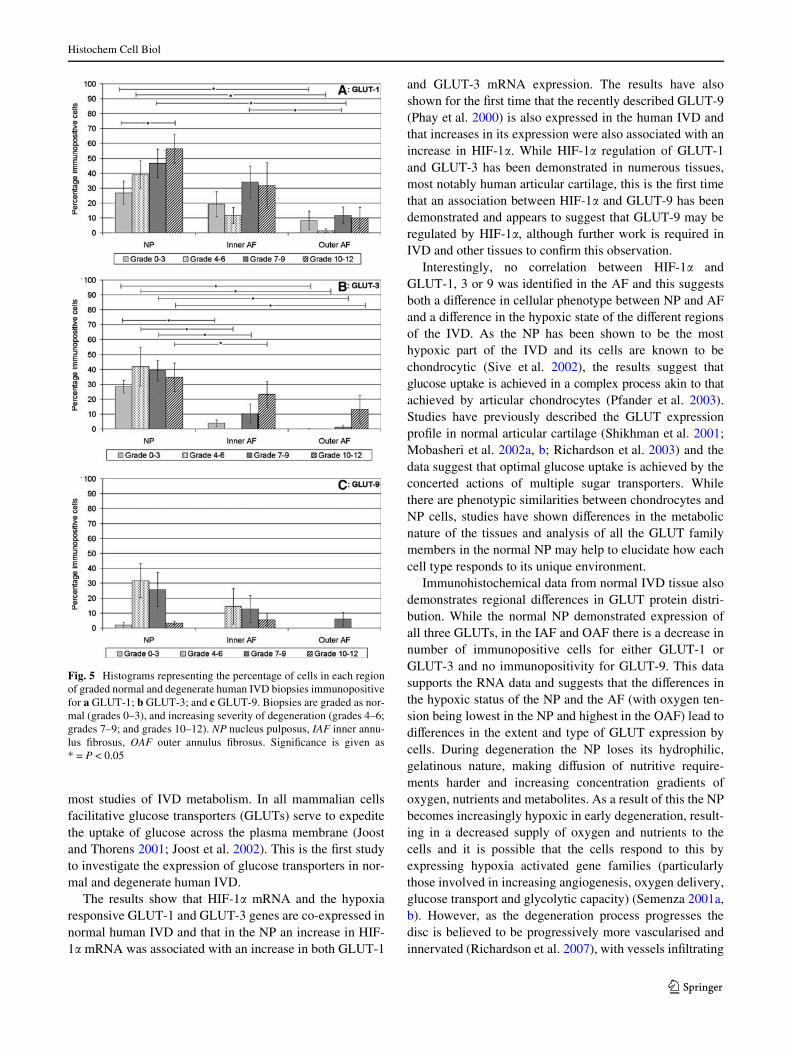
In human IVD samples GLUT-1 showed the strongestimmunoreactivity in the NP and IAF regions with 27 and19% of cells being immunopositive in each region, respec-tively (Fig. 5a). The OAF expressed signiWcantly lessGLUT-1 than the NP (P < 0.05), with only 8% of cellsbeing immunopositive. GLUT-3 was identiWed in 28% ofnormal NP cells, signiWcantly more than either the IAF(P < 0.005) in which only 4% of cells were immunoposi-
tive, or OAF (P < 0.005) which showed no immunoreactiv-ity for GLUT-3 (Fig. 5b). GLUT-9 showed the lowestexpression of all the GLUTs, with only 2% of normal NPcells being immunopositive and no immunopositive cells ineither the IAF or the OAF (Fig. 5c).
Alterations in the distribution of GLUT-1, GLUT-3 and GLUT-9 in human disc degeneration
In the NP, the number of GLUT-1 immunopositive cellsincreased as grade of degeneration increased and thisincrease was statistically signiWcant by grades 10–12(P < 0.05) when 56% of cells were immunopositive(Fig. 5a). This trend was not repeated in either the IAF orthe OAF; however levels were slightly raised in both theseregions in later stages of degeneration. At each grade ofdegeneration the NP showed the highest number of GLUT-1 immunopositive cells and the OAF the lowest, with thisdiVerence being at grades 7–9 (P < 0.05). At grades 7–9GLUT-1 was also expressed in a signiWcantly higher num-ber of cells in the IAF than in the OAF (P < 0.05).
GLUT-3 expression was again highest in the NP at eachgrade of degeneration and lowest in the OAF (Fig. 5b). Ateach grade of degeneration the NP demonstrated signiW-cantly higher numbers of GLUT-3 immunopositive cellsthan the IAF (grades 0–3 P < 0.005; grades 4–6 P < 0.05;grades 7–9 P < 0.01; grades 10–12 P < 0.05) and the OAF(grades 0–3 P < 0.005; grades 4–6 P < 0.05; grades 7–9P < 0.01; grades 10–12 P < 0.05). Immunopositivity didnot change signiWcantly in the NP as degenerationincreased, however in both the IAF and OAF there wereslight increases in immunopositivity as grade of degenera-tion increased.
While GLUT-9 was only expressed in 2% of normal NPcells there was an increase in immunopositivity to 32% in
Fig. 3 Histological images of normal (a, c) and degenerate (b, d) hu-man intervertebral disc stained with haematoxylin and eosin used forgrading. Macroscopically normal IVD (a) demonstrates matrix integ-rity and clear demarcation between NP and AF regions, while degen-erate IVD demonstrates slits within the NP extending to the AF and a
loss of demarcation between NP and AF regions (b). Cells in normalIVD were distributed evenly throughout the matrix (c), while degener-ate tissue showed evidence of cell clusters (d). Image magniWcations£1 (a, c), £150 (b, d)
123
Histochem Cell Biol
grades 4–6 and to 26% in grades 7–9 (Fig. 5c). By grades10–12, however, the expression had decreased to 3% ofcells. A similar, but more limited, trend was seen in theinner AF with no expression in normal tissue, but 15 and12% of immunopositive cells in grades 4–6 and 7–9, respec-tively. Again, by grades 10–12 expression had deceased(6%). In the OAF expression was only observed in grades7–9 where only 6% of cells were immunopositive.
Discussion
The NP of the healthy, adult IVD is avascular, with only theouter edges of the AF and end-plates having any blood ves-
sels (Bartels et al. 1998; Repanti et al. 1998). Thus forexchange of nutrients and waste products the mature IVDrelies on a diVusion process from the blood vessels of thevertebral body through the spongy vertebral bone and thecartilaginous endplates (Urban et al. 2001). Althoughgenetic, mechanical and environmental factors are impli-cated in IVD degeneration the avascular nature of the discalso implicates nutrition, metabolism and oxygenation atthe crux of the IVD degeneration process. Glucose is anessential substrate for disc cells; it is needed both for glyco-lytic metabolism and the synthesis of glycoproteins andglycosaminoglycans (key molecules in preserving thegelatinous nature of the NP). The uptake of glucose by disccells is a fundamental process that has been neglected in
Fig. 4 Immunohistochemical localisation of GLUT1, GLUT3 and GLUT9 in the normal hu-man intervertebral disc and pos-itive control tissues represented on the CHTN human tissue microarrays. Immunostaining for the GLUT proteins in disc cells is indicated by the Fast Red TR/Naphthol AS-MX substrate. Examples of expression of each GLUT are shown in the NP of normal IVD (a, d, g) and degen-erate IVD (b, e, h). Positive con-trols for each GLUT demonstrated expression of (c) GLUT-1 in erythrocytes; (f) GLUT-3 in brain cortex; and (i) GLUT-9 in ileum. An example of an IgG negative control slide is also shown (j). Original mag-niWcations £200 (a, b, d–h, j) or £400 (c, i)
123
Histochem Cell Biol
most studies of IVD metabolism. In all mammalian cellsfacilitative glucose transporters (GLUTs) serve to expeditethe uptake of glucose across the plasma membrane (Joostand Thorens 2001; Joost et al. 2002). This is the Wrst studyto investigate the expression of glucose transporters in nor-mal and degenerate human IVD.
The results show that HIF-1� mRNA and the hypoxiaresponsive GLUT-1 and GLUT-3 genes are co-expressed innormal human IVD and that in the NP an increase in HIF-1� mRNA was associated with an increase in both GLUT-1
and GLUT-3 mRNA expression. The results have alsoshown for the Wrst time that the recently described GLUT-9(Phay et al. 2000) is also expressed in the human IVD andthat increases in its expression were also associated with anincrease in HIF-1�. While HIF-1� regulation of GLUT-1and GLUT-3 has been demonstrated in numerous tissues,most notably human articular cartilage, this is the Wrst timethat an association between HIF-1� and GLUT-9 has beendemonstrated and appears to suggest that GLUT-9 may beregulated by HIF-1�, although further work is required inIVD and other tissues to conWrm this observation.
Interestingly, no correlation between HIF-1� andGLUT-1, 3 or 9 was identiWed in the AF and this suggestsboth a diVerence in cellular phenotype between NP and AFand a diVerence in the hypoxic state of the diVerent regionsof the IVD. As the NP has been shown to be the mosthypoxic part of the IVD and its cells are known to bechondrocytic (Sive et al. 2002), the results suggest thatglucose uptake is achieved in a complex process akin to thatachieved by articular chondrocytes (Pfander et al. 2003).Studies have previously described the GLUT expressionproWle in normal articular cartilage (Shikhman et al. 2001;Mobasheri et al. 2002a, b; Richardson et al. 2003) and thedata suggest that optimal glucose uptake is achieved by theconcerted actions of multiple sugar transporters. Whilethere are phenotypic similarities between chondrocytes andNP cells, studies have shown diVerences in the metabolicnature of the tissues and analysis of all the GLUT familymembers in the normal NP may help to elucidate how eachcell type responds to its unique environment.
Immunohistochemical data from normal IVD tissue alsodemonstrates regional diVerences in GLUT protein distri-bution. While the normal NP demonstrated expression ofall three GLUTs, in the IAF and OAF there is a decrease innumber of immunopositive cells for either GLUT-1 orGLUT-3 and no immunopositivity for GLUT-9. This datasupports the RNA data and suggests that the diVerences inthe hypoxic status of the NP and the AF (with oxygen ten-sion being lowest in the NP and highest in the OAF) lead todiVerences in the extent and type of GLUT expression bycells. During degeneration the NP loses its hydrophilic,gelatinous nature, making diVusion of nutritive require-ments harder and increasing concentration gradients ofoxygen, nutrients and metabolites. As a result of this the NPbecomes increasingly hypoxic in early degeneration, result-ing in a decreased supply of oxygen and nutrients to thecells and it is possible that the cells respond to this byexpressing hypoxia activated gene families (particularlythose involved in increasing angiogenesis, oxygen delivery,glucose transport and glycolytic capacity) (Semenza 2001a,b). However, as the degeneration process progresses thedisc is believed to be progressively more vascularised andinnervated (Richardson et al. 2007), with vessels inWltrating
Fig. 5 Histograms representing the percentage of cells in each regionof graded normal and degenerate human IVD biopsies immunopositivefor a GLUT-1; b GLUT-3; and c GLUT-9. Biopsies are graded as nor-mal (grades 0–3), and increasing severity of degeneration (grades 4–6;grades 7–9; and grades 10–12). NP nucleus pulposus, IAF inner annu-lus Wbrosus, OAF outer annulus Wbrosus. SigniWcance is given as* = P < 0.05
123

Histochem Cell Biol
Wrst the OAF, then progressing inwards and eventuallyinWltrating the NP. These changes are part of the metabolicand functional adaptation to the degenerated disc matrixand are intended to lead to an increased supply of oxygenand nutrients to the cells. These hypotheses are supportedby the immunohistochemical data from this study. In theNP, the most hypoxic region of the IVD, the number ofcells immunopositive for the three GLUTs studiedincreases in early grades of degeneration (grades 4–6 and7–9) as oxygen and glucose concentrations decrease. In thecase of GLUT-1, the most ubiquitous of the GLUTs, thisincrease continues to late stage degeneration (grades 10–12), but for GLUT-3 and particularly GLUT-9 the numberof immunopositive cells decreases from grades 7–9 togrades 10–12. These changes are likely to be a response,initially to the lack of available glucose within the extra-cellular matrix and later to a change in the physiology ofthe disc as new blood vessels inWltrate the NP.
The available literature suggests that GLUT-1 andGLUT-3 are regulated by hypoxia but there is no evidencefor hypoxic regulation of GLUT-9 (Vannucci et al. 1998;Badr et al. 1999; Zhang et al. 1999). Thus, it is reasonableto suggest that up-regulation of GLUT-1 and GLUT-3 inearly degeneration may be a metabolic adaptation to thehypoxic conditions. Many studies have show that GLUT-3is expressed in the brain and is important for glucose deliveryto neurons when glucose concentrations are low (i.e. inhypoxic and ischemic conditions) (Vannucci et al. 1998).Up-regulation of these proteins is likely to result from thecells attempt to increase their uptake of glucose, eitherbecause of decreased diVusion of glucose through thedegenerate matrix, increased cellular demands from glyco-lytic pathways or stimulation by catabolic, pro-inXamma-tory cytokines such as IL-1� (Mobasheri et al. 2002a, b).
In summary the results presented suggest that glucosetransport and metabolism are hypoxia regulated processesin the avascular regions of the human IVD and thatdegeneration aVects the expression of glucose transporters.
Acknowledgments We would like to thank Dr. Stephen Baldwin(Faculty of Biological Sciences, University of Leeds) and Dr. KelleMoley (Washington University School of Medicine, St Louis, MO,USA) for polyclonal antibodies to GLUT-1 and GLUT-9, respectively.We thank Dr. Christopher A. Moskaluk (Departments of Pathology,Biochemistry, and Molecular Genetics, University of Virginia HealthSystem, Charlottesville, VA, USA) and the staV of the CooperativeHuman Tissue Network of the National Cancer Institute (NationalInstitutes of Health, Bethesda, MD, USA) for access to CHTN tissuemicroarrays. Thanks also to Dr. Christine Le Maitre for assistance withthe real-time PCR. We gratefully acknowledge the support of the jointUK Research Councils’ Interdisciplinary Research Collaboration inTissue Engineering (Biotechnology and Biological Sciences ResearchCouncil, Engineering and Physical Sciences Research Council, andMedical Research Council).
ConXict of interest None of the authors declare any conXicts ofinterest for this work.
References
Augustin R, Carayannopoulos MO, Dowd LO, Phay JE, Moley JF,Moley KH (2004) IdentiWcation and characterization of humanglucose transporter-like protein-9 (GLUT9): alternative splicingalters traYcking. J Biol Chem 279(16):16229–16236
Badr GA, Zhang JZ, Tang J, Kern TS, Ismail-Beigi F (1999) Glut1 andglut3 expression, but not capillary density, is increased by cobaltchloride in rat cerebrum and retina. Brain Res Mol Brain Res64(1):24–33
Bartels EM, Fairbank JC, Winlove CP, Urban JP (1998) Oxygen andlactate concentrations measured in vivo in the intervertebral discsof patients with scoliosis and back pain. Spine 23(1):1–7
Bibby SR, Jones DA, Lee RB, Yu J, Urban JPG (2001) The pathophys-iology of the intervertebral disc. Joint Bone Spine 68(6):537–542
Crean JK, Roberts S, JaVray DC, Eisenstein SM, Duance VC (1997)Matrix metalloproteinases in the human intervertebral disc: rolein disc degeneration and scoliosis. Spine 22(24):2877–2884
Davies A, Meeran K, Cairns MT, Baldwin SA (1987) Peptide-speciWcantibodies as probes of the orientation of the glucose transporter inthe human erythrocyte membrane. J Biol Chem 262(19):9347–9352
Davies A, Ciardelli TL, Lienhard GE, Boyle JM, Whetton AD, Bald-win SA (1990) Site-speciWc antibodies as probes of the topologyand function of the human erythrocyte glucose transporter. Bio-chem J 266(3):799–808
Freemont AJ, Watkins A, Le MC, Jeziorska M, Hoyland JA (2002)Current understanding of cellular and molecular events in inter-vertebral disc degeneration: implications for therapy. J Pathol196(4):374–379
Froehner SC, Davies A, Baldwin SA, Lienhard GE (1988) The blood-nerve barrier is rich in glucose transporter. J Neurocytol17(2):173–178
Gerhart DZ, Broderius MA, Borson ND, Drewes LR (1992) Neuronsand microvessels express the brain glucose transporter proteinGLUT3. Proc Natl Acad Sci USA 89(2):733–737
Goupille P, Jayson MI, Valat JP, Freemont AJ (1998) Matrix metallo-proteinases: the clue to intervertebral disc degeneration? Spine23(14):1612–1626
Gruber HE, Hanley EN Jr (1998) Analysis of aging and degenerationof the human intervertebral disc. Comparison of surgical speci-mens with normal controls. Spine 23(7):751–757
Ha KY, Koh IJ, Kirpalani PA, Kim YY, Cho YK, Khang GS, Han CW(2006) The expression of hypoxia inducible factor-1alpha andapoptosis in herniated discs. Spine 31(12):1309–1313
Iatridis JC, Mente PL, Stokes IA, Aronsson DD, Alini M (1999) Com-pression-induced changes in intervertebral disc properties in a rattail model. Spine 24(10):996–1002
Ishihara H, Urban JP (1999) EVects of low oxygen concentrations andmetabolic inhibitors on proteoglycan and protein synthesis ratesin the intervertebral disc. J Orthop Res 17(6):829–835
Joost HG, Thorens B (2001) The extended GLUT-family of sugar/pol-yol transport facilitators: nomenclature, sequence characteristics,and potential function of its novel members (review). Mol MembrBiol 18(4):247–256
Joost HG, Bell GI, Best JD, Birnbaum MJ, Charron MJ, Chen YT, Do-ege H, James DE, Lodish HF, Moley KH, Moley JF, Mueckler M,Rogers S, Schurmann A, Seino S, Thorens B (2002) Nomencla-ture of the GLUT/SLC2A family of sugar/polyol transport facili-tators. Am J Physiol Endocrinol Metab 282(4):E974–E976
Kasanicki MA, Cairns MT, Davies A, Gardiner RM, Baldwin SA (1987)IdentiWcation and characterization of the glucose-transport proteinof the bovine blood/brain barrier. Biochem J 247(1):101–108
Le Maitre CL, Freemont AJ, Hoyland JA (2004) Localization of deg-radative enzymes and their inhibitors in the degenerate humanintervertebral disc. J Pathol 204(1):47–54
123

Histochem Cell Biol
Le Maitre CL, Freemont AJ, Hoyland JA (2005) The role of interleu-kin-1 in the pathogenesis of human intervertebral disc degenera-tion. Arthritis Res Ther 7(4):R732–R745
Le Maitre CL, Freemont AJ, Hoyland JA (2007) Accelerated cellularsenescence in degenerate intervertebral discs: a possible role inthe pathogenesis of intervertebral disc degeneration. Arthritis ResTher 9(3):R45
Lee RB, Urban JP (1997) Evidence for a negative Pasteur eVect inarticular cartilage. Biochem J 321(Pt 1):95–102
Lee RB, Urban JP (2002) Functional replacement of oxygen by otheroxidants in articular cartilage. Arthritis Rheum 46(12):3190–3200
Maroudas A, Stockwell RA, Nachemson A, Urban J (1975) Factorsinvolved in the nutrition of the human lumbar intervertebraldisc: cellularity and diVusion of glucose in vitro. J Anat 120(Pt 1):113–130
Mobasheri A, Vannucci SJ, Bondy CA, Carter SD, Innes JF, ArteagaMF, Trujillo E, Ferraz I, Shakibaei M, Martin-Vasallo P (2002a)Glucose transport and metabolism in chondrocytes: a key tounderstanding chondrogenesis, skeletal development and carti-lage degradation in osteoarthritis. Histol Histopathol 17(4):1239–1267
Mobasheri A, Neama G, Bell S, Richardson S, Carter SD (2002b) Hu-man articular chondrocytes express three facilitative glucosetransporter isoforms: GLUT1, GLUT3 and GLUT9. Cell Biol Int26(3):297–300
Mobasheri A, Dobson H, Mason SL, Cullingham F, Shakibaei M, Mo-ley JF, Moley KH (2005) Expression of the GLUT1 and GLUT9facilitative glucose transporters in embryonic chondroblasts andmature chondrocytes in ovine articular cartilage. Cell Biol Int29(4):249–260
Ohshima H, Urban JP (1992) The eVect of lactate and pH on proteo-glycan and protein synthesis rates in the intervertebral disc. Spine17(9):1079–1082
Pfander D, Cramer T, Schipani E, Johnson RS (2003) HIF-1alpha con-trols extracellular matrix synthesis by epiphyseal chondrocytes.J Cell Sci 116(Pt 9):1819–1826
Phay JE, Hussain HB, Moley JF (2000) Cloning and expression anal-ysis of a novel member of the facilitative glucose transporter fam-ily, SLC2A9 (GLUT9). Genomics 66(2):217–220
Pockert AJ, Le Maitre CL, Buttle DJ, Freemont AJ, Hoyland JA (2006)Gene expression of aggrecanolytic ADAMTS and TIMP-3 in thehuman intervertebral disc. Int J Exp Pathol 87:A19
Rajpurohit R, Risbud MV, Ducheyne P, Vresilovic EJ, Shapiro IM(2002) Phenotypic characteristics of the nucleus pulposus:expression of hypoxia inducing factor-1, glucose transporter-1and MMP-2. Cell Tissue Res 308(3):401–407
Repanti M, Korovessis PG, Stamatakis MV, Spastris P, Kosti P (1998)Evolution of disc degeneration in lumbar spine: a comparativehistological study between herniated and postmortem retrieveddisc specimens. J Spinal Disord 11(1):41–45
Richardson S, Neama G, Phillips T, Bell S, Carter SD, Moley KH, Mo-ley JF, Vannucci SJ, Mobasheri A (2003) Molecular characteriza-tion and partial cDNA cloning of facilitative glucose transporters
expressed in human articular chondrocytes; stimulation of 2-de-oxyglucose uptake by IGF-I and elevated MMP-2 secretion byglucose deprivation. Osteoarthritis Cartilage 11(2):92–101
Richardson SM, Walker RV, Parker S, Rhodes NP, Hunt JA, FreemontAJ, Hoyland JA (2006a) Intervertebral disc cell-mediated mesen-chymal stem cell diVerentiation. Stem Cells 24(3):707–716
Richardson SM, Curran JM, Chen R, Vaughan-Thomas A, Hunt JA,Freemont AJ, Hoyland JA (2006b) The diVerentiation of bonemarrow mesenchymal stem cells into chondrocyte-like cells onpoly-L-lactic acid (PLLA) scaVolds. Biomaterials 27(22):4069–4078
Richardson SM, Mobasheri A, Freemont AJ, Hoyland JA (2007) Inter-vertebral disc biology, degeneration and novel tissue engineeringand regenerative medicine therapies. Histol Histopathol22(9):1033–1041
Semenza GL, Roth PH, Fang HM, Wang GL (1994) Transcriptionalregulation of genes encoding glycolytic enzymes by hypoxia-inducible factor 1. J Biol Chem 269(38):23757–23763
Semenza GL, Jiang BH, Leung SW, Passantino R, Concordet JP,Maire P, Giallongo A (1996) Hypoxia response elements in thealdolase A, enolase 1, and lactate dehydrogenase A gene promot-ers contain essential binding sites for hypoxia-inducible factor 1.J Biol Chem 271(51):32529–32537
Semenza GL (2001a) Hypoxia-inducible factor 1: control of oxygenhomeostasis in health and disease. Pediatr Res 49(5):614–617
Semenza GL (2001b) Regulation of hypoxia-induced angiogenesis: achaperone escorts VEGF to the dance. J Clin Invest 108(1):39–40
Shikhman AR, Brinson DC, Valbracht J, Lotz MK (2001) Cytokineregulation of facilitated glucose transport in human articularchondrocytes. J Immunol 167(12):7001–7008
Shikhman AR, Brinson DC, Lotz MK (2004) Distinct pathways regu-late facilitated glucose transport in human articular chondrocytesduring anabolic and catabolic responses. Am J Physiol Endocri-nol Metab 286(6):E980–E985
Sive JI, Baird P, Jeziorsk M, Watkins A, Hoyland JA, Freemont AJ(2002) Expression of chondrocyte markers by cells of normal anddegenerate intervertebral discs. Mol Pathol 55(2):91–97
Thompson RE, Pearcy MJ, Downing KJ, Manthey BA, Parkinson IH,Fazzalari NL (2000) Disc lesions and the mechanics of the inter-vertebral joint complex. Spine 25(23):3026–3035
Urban MR, Fairbank JC, Etherington PJ, Loh FL, Winlove CP, UrbanJP (2001) Electrochemical measurement of transport into scoli-otic intervertebral discs in vivo using nitrous oxide as a tracer.Spine 26(8):984–990
Vannucci SJ, Reinhart R, Maher F, Bondy CA, Lee WH, Vannucci RC,Simpson IA (1998) Alterations in GLUT1 and GLUT3 glucosetransporter gene expression following unilateral hypoxia-ische-mia in the immature rat brain. Brain Res Dev Brain Res107(2):255–264
Yasuma T, Koh S, Okamura T, Yamauchi Y (1990) Histologicalchanges in aging lumbar intervertebral discs. Their role in protru-sions and prolapses. J Bone Joint Surg Am 72(2):220–229
Zhang JZ, Behrooz A, Ismail-Beigi F (1999) Regulation of glucosetransport by hypoxia. Am J Kidney Dis 34(1):189–202
123
Recommended
















