
http://www.elsevier.com/locate/bba
Biochimica et Biophysica Ac
Review
Proangiogenic activity of beta-carotene is coupled with the
activation of endothelial cell chemotaxis
A. Dembinska-Kieca,*, A. Polusa, B. Kiec-Wilka, J. Grzybowskaa, M. Mikolajczyka, J. Hartwicha,
U. Raznya, K. Szumilasa, A. Banasa, M. Bodziocha, J. Stachurab, G. Dyduchb, P. Laidlerc,
J. Zagajewskic, T. Langmand, G. Schmitzd
aDepartment of Clinical Biochemistry, The Jagiellonian University Medical College, Kopernika 15a, 31-501 Krakow, PolandbDepartment of Pathology, The Jagiellonian University Medical College, Krakow, Poland
cDepartment of Medical Biochemistry, The Jagiellonian University Medical College, Krakow, PolanddDepartment of Clinical Chemistry, Laboratory Medicine and Transfusiology, University of Regensburg, Regensburg, Germany
Received 27 September 2004; received in revised form 5 November 2004; accepted 22 November 2004
Available online 28 December 2004
Abstract
Endothelial cells play an important role in angiogenesis (formation of new vessels from preexisting ones), which is essential for
organogenesis, tissue remodeling but also inflammatory response, carcinogenesis in all periods of our life. Beta-carotene (BC) in non-toxic
concentrations (up to 3 AM) had no detectable effect on HUVECs (human umbilical vein endothelial cells) proliferation or apoptosis,
despite significant changes of the expression patterns of pro- and anti-apoptotic genes. However beta-carotene did not change the
tubulogenic activity of HUVEC in the in vitro angiogenesis model, it potently accelerated the bFGF-induced development of
microcapillaries, as well as the migration of endothelial cells, in matrigel plug injected subcutaneously to mice. Potent activation of
endothelial cell migration in the in vitro model of chemotaxis was also observed. According to the microarray data, genes involved in cell/
cell and cell/matrix adhesion, matrix reorganization, activation of chemotaxis, the G-protein regulated intracellular signaling as well as
genes involved in the rapid remodeling of protein cytoskeleton were the most affected by BC in HUVEC. We conclude that beta-carotene in
the physiological concentration range stimulates early steps of angiogenesis by the activation of cellular migration as well as matrix
reorganization and decrease of cell adhesion.
D 2004 Published by Elsevier B.V.
Keywords: Beta-carotene; Angiogenesis; Endothelium; Microarray; Chemotaxis
1. Introduction
Angiogenesis, the formation of new blood vessels from
the preexisting ones, plays a central role in a number of
physiologic and pathologic events including vascular
remodeling of growing ischemic tissue, inflammation,
diabetic retinopathy, as well as in progression of solid
tumor growth and invasiveness [1,2].
The knowledge of the formation of new tubular
structures in angiogenesis is being continuously enriched
0925-4439/$ - see front matter D 2004 Published by Elsevier B.V.
doi:10.1016/j.bbadis.2004.11.017
* Corresponding author. Tel.: +48 12 421 40 06; fax: +48 12 421 40
73.
E-mail address: [email protected] (A. Dembinska-Kiec).
in new observations suggesting the influence of the new
regulators. A number of growth factors (such as VEGF,
bFGF, TGFb, PDGF, IGF and others), cell/matrix (integ-
rins) and cell/cell (VE-cadherins, catenins, endoglins,
ephrins, their receptors or Jagged/Notch pathway) inter-
actions, and environmental factors (such as shear stress,
nutrients, oxygen supply and others) regulate the most
important steps in angiogenesis, which include detachment,
proliferation, migration, homing and differentiation of the
vascular wall cells, mainly endothelial or their progenitors
[3,4].
Carotenoids are lipid soluble hydrocarbon pigments
(C40H56) containing 11 conjugated double bonds. The main
plant carotenoids detected in human blood are: beta-carotene
ta 1740 (2005) 222–239

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 223
(BC), alpha-carotene, gamma-carotene and lycopene. They
are isomers varying in the position of double bonds and the
presence of acyclic ring. Carotenoids present in human
tissues function as free radical scavengers, immunomodula-
tors, or some being the substrate for the synthesis of retinol
(Vitamin A) and retinoic acid (RA) are regulators of cell fate
and differentiation [5]. RA, derived from beta-carotene, is a
well known activator of nuclear receptors such as retinoid/
rexinoid receptors (RXR/RAR). RA in cooperation with fatty
acids and fatty acid metabolites regulates the activity of the
PPAR/RXR heterodimer (peroxisome proliferator-activated
receptors, PPARs) [6]. As regulators of transcription they
influence cell differentiation, maturation of tissues and body
organization in both fetal and adult periods of life [5–7].
Epidemiological studies suggested that the intake of
fruit and vegetables rich in carotenoids results in the
reduction of the cancer incidence, decreased risk of heart
diseases, and enhanced immunity due to its free radical
scavenging properties, anti-inflammatory and increased
activity of NK cells [8–11]. Surprisingly, some of the large
clinical studies, undertaken to prove the efficacy of beta-
carotene supplementation in the prevention of coronary
heart disease and cancer, showed that administration of
beta-carotene or vitamin A might increase the risk of lung
cancer, especially in smokers and patients afflicted with
asbestosis (Alpha-Tocopherol Beta-Carotene Cancer Pre-
vention Study (ATBC), [12], Beta-Carotene Retinol Effi-
cacy Trial (CARET) [13], Physician’s Health Study (PHS),
[14]).
Since 15% of beta-carotene ingested reach human target
cells in the non-metabolized form [15] and angiogenesis is
an important part of remodeling of an ischemic tissue as
well as solid tumor malignancy [1], this study was under-
taken to define the direct effects of beta-carotene on
endothelial cells in terms of angiogenic activity and
regulation of gene expression.
2. Methods
The HPLC grade beta-carotene (1 mg) filled with
nitrogen in vials made of dark glass were kindly provided
for the project by the Roche Vitamins AG, Kaiseraugust,
Switzerland.
2.1. The cell culture
Primary endothelial cells (HUVECs) were isolated from
human umbilical veins using collagenase digestion.
HUVECs were cultured in EBM (EGM Bullet Kit,
Clonetics) with supplements: hEGF (10 ng/ml), hydro-
cortisone (1 ng/ml), Bovine Brain Extract (12 Ag/ml),
antibiotics: gentamicin (50 Ag/ml) as well as amphoter-
icin-B (50 ng/ml), and 10% Fetal Bovine Serum (Clonetics).
The resulting cell lines were characterized by morphological
and immunohistochemical criteria such as desmin, alpha-
actin, vWF [16]. Experiments were performed on 70%
confluent cell cultures (up to 5th passage).
For cell culture, 4 mM beta-carotene (BC) stock solution
in tetrahydrofurane (THF) (Sigma) was further diluted in
ethanol (1:1 v/v THF/EtOH ratio) to obtain 2 mM
concentration of BC. The further dilutions were made with
the tissue culture medium (EBM). The final concentration of
THF/EtOH in the cell culture medium was 0.075%.
HUVECs were incubated with the solvent (0.075% THF/
EtOH) containing medium (as the control) and beta-carotene
(BC) (0.3–10 AM), arachidonic acid (AA) (3 AM) (Sigma),
or both BC (3 AM) and AA (3 AM) for 24 h at 37 8C in 5%
CO2 and 95% humidity (Jouan IG 150).
2.2. Beta-carotene uptake by HUVECs (measured by HPLC)
All solvents used for HPLC (THF, ethanol, tBME
(tetra-butylmethylether) (Sigma) and BHT (2,6-Di-tert-
butyl-p-cresol) (Sigma)) were analytical grade. THF and
ethanol were additionally filtered through the Aluminum
Oxide column (Sigma) before use. Following 24 h
incubation of HUVECs with the studied compounds, cells
were detached by trypsinization and washed three times
with Ca+2 and Mg+2 free PBS (centrifugation: 400�g for
10 min, at the room temperature). Cells were counted
using the Burker chamber and spinned down (400�g, 10
min, at room temperature). The resulted pellet was frozen
(at �80 8C) for further analysis. BC concentration in the
cells and BC stability in culture medium (lack of
spontaneous degradation products), were assessed with
the HPLC micromethod developed by Roche Vitamins AG
(Kaiseraugust, Switzerland). Briefly, for extraction, 200 Alof acetone (with 0.025% BHT f.c.) was added to a frozen
cell pellet, mixed for 1 min and dried in a vacuum
centrifuge under argon (30 min at 50 mbar). 200 Al of theBC extraction solvent (ethanol/THF/tBME 9:1:5, 0.025%
BHT f.c.) was applied to the dried pellet, vigorously mixed
and centrifuged (3 min, 8000�g at 4 8C). The supernatant
was used for HPLC.
For the measurement of BC content in the tissue culture
medium, 25 Al of the medium was mixed with 225 Al of theBC extraction solvent described above, mixed for 1 min and
centrifuged for 3 min at 8000�g at 4 8C. The HPLC sample
analysis was isocratically performed on a Vydac 218TP54
column C-18 (Roche) (4.6�250 mm) at a constant column
temperature 20 8C. The sample solvent consisted of ethanol/
THF/tBME 9:1:5, 0.025% BHT f.c. The mobile phase was:
acetonitryl/tert-butylmethylether/ammonium acetate (80
mM)/triethylamine (73:20:7:0.05) at a constant flow of
butylmethylether/1.5 ml/min. Shimadzu SCL-10AVP instru-
ment (Shimadzu, Kyoto, Japan) with the SPD-10AV
detector was set at 450 nm for the carotenoids estimation.
In this HPLC system, 12V-apocarotenal, 8V-apocarotenal, 4V-h-apocarotenal, all-trans-h-carotene and (Z)-h-carotenecould be detected. The amount of BC was expressed as
pmol of BC/106 cells.

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239224
2.3. Detection of cellular toxicity and apoptosis
The possible toxic effects of used compounds and solvents
on HUVECs were assessed by the cellular lactic dehydrogen-
ase (LDH) presence in the medium after 24-h incubation (Cy-
toTox 96 Non-Radioactive Cytotoxicity Assay, Promega).
For the estimation of apoptosis the ApoFluorR Green
Caspase Activity Assay (ICN Biomedicals Inc.) was used.
This test allows for the measurement of global cellular
caspase activity. Proapoptotic staurosporine (at 1 nM) was
used as the positive control for apoptosis. Following
incubation with the fluorescent dye (1 h) cells were
harvested, washed to remove unbound dye with the Ca+2
and Mg+2 free PBS and 1�106 cells were placed in black
microtiter plates (NUNC A/S, Roskilde). The fluorescence
was detected using a plate reader (Synergy HT, BIO-TEK)
and quantitated at 520 nm emission wavelength following
the excitation with the 490 nm light. Results are expressed
as the percentage of activity detected in the control
(incubated with the solvent only) HUVECs.
2.4. Assay of HUVEC proliferation (The bromo-
deoxyuridyne (BrdU) incorporation assay)
In order to determine the effect of BC and AA on the
HUVEC proliferation, the rate of DNA synthesis was
measured by the incorporation of the thymidine analog,
bromo-deoxyuridine (BrdU) to DNA [17]. HUVECs
(5�104 cells) were incubated with the compounds for 24
h, as described above, and additionally with BrdU for the
last 3 h. Following incubation, cells were fixed and stained
with the anti-BrdU kit (Roche) according to the manufac-
turer’s recommendations. HUVECs proliferation induced by
VEGF (Vascular Endothelial Growth Factor) (0.2 nM)
(Sigma) or bFGF (basic fibroblast growth factor) (0.5 nM)
(Sigma) for 24 h was served as the positive control. Results
are given as the percent of the control BrdU incorporation to
the HUVEC incubated in the medium with solvents only.
2.5. Migration of endothelial cells (The Boyden’s Chamber
assay)
HUVECs harvested from 3–4 passages were suspended
(105 cells/100 Al) in the EBM medium with 0.5% FBS and
seeded into BD Falconk FluoroBlokk Inserts (3 Am pore,
Becton Dickinson). The inserts containing HUVECs were
placed into a 24-well plate with 600 Al of the EBM medium
and 0.5% FBS, and incubated for 24 h at 37 8C. Media with
the addition of BC (3 AM) or AA (3 AM) as well as 1-
phospho-sphingosine (S1P) (500 nM) (Sigma) as the
positive control [18] were used as chemoattractant in the
lower chamber. After 24 h, HUVECs were stained with anti-
CD31 antibody conjugated with phycoerythrin (PE) (10 Ag/ml) (Becton Dickinson) for 30 min at 37 8C. Subsequently,cells in the inserts were washed with the Ca+2 and Mg+2 free
PBS and the fluorescence of the cells, which had migrated
through the pores to the bottom side of the inserts, was
measured with a fluorescence plate reader (Synergy HT,
BIO-TEK). The changes in intensity of fluorescence in a
sample with agents versus control (cells migrated against the
EBM with solvents only) corresponded to the amount of
migrating cells. The chemotactic activity of the endothelial
cells was expressed as the chemotaxis index (CHI) which
represents the ratio of migration stimulated by investigated
compound versus random migration of unstimulated
HUVECs in THF/EtOH control sample.
2.6. The three dimensional [3D] model of tubulogenesis in
matrigel in vitro [19]
HUVECs harvested from 3–4 passages were resuspended
in Matrigel (containing laminin, collagen IV, entactin,
heparan sulfate proteoglycans) (Becton Dickinson) to the
final concentration of 1�106 cells/ml on ice. The cell–
matrigel mixture (50 Al) was placed into the cell culture
dishes and incubated in a humidified CO2 incubator (Jouan)
at 37 8C for 30 min. Subsequently the EBM medium (100
Al) supplemented with solvent (control probe), or BC (3
AM), or AA (3 AM), or BC/AA mixture was applied on top
of the cells immersed in matrigel and was incubated in a
humidified CO2 incubator for 24 h at 37 8C. The number of
tubules formed in the presence of VEGF (0.2 nM) or bFGF
(0.5 nM) served as the positive control. The formation of
tubules by HUVECs suspended in matrigel was assessed
under the light microscope (at magnitude �10) and photo-
graphed. The lengths of the analyzed tubule-like structures
were calculated and expressed as the average sum of total
length of tubules visible under the light microscope.
2.7. Angiogenesis in vivo (the mouse model)
Protocol was accepted by the local University Ethic
Committee. All animal experiments were performed accord-
ing to Polish laws and approved by the Polish Animal
Inspectorate and Institutional Animal Care. Female Balb/c
mice (n=6) received sterile injections of 2�500 Al matrigel
(Becton Dickinson) s.c. (dorsally). The matrigel plug con-
tained solvent or BC (3 AM), bFGF (50 nM), or both BC and
bFGF at the given concentrations. Six days later the animals
were sacrificed by overdosing urethane anesthesia and the
matrigel plug were removed, fixed and immersed in paraffin.
Immunohistochemistry was performed using routine proto-
col. Primary anti-CD31 antibodies (anti-PECAM-1, BD
Pharmingen) at 1:300 dilution were used. The slides were
rehydrated and incubated in 3% peroxide solution for 10 min
to block endogenous peroxidase activity. The Streptavidin–
Biotin (BD Pharmingen) detection system was used. DAB
was used as chromogen. The slides were contra-stained with
Mayer hematoxylin (DAKO, Denmark). The amount of
capillaries was counted under the microscope in five different
fields in each of the three slices taken from different parts of
each plugs. The number of capillaries detected in slices from

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 225
the plugs containing the studied compounds was compared to
the plugs with solvents and expressed as the number of
vessels with or without the lumen. Additionally, the number
of separately detected endothelial PECAM-positive single
cells, not connected with capillaries, migrating to the matrigel
was calculated.
2.8. The protein synthesis (Western Blot)
Western-blot was used to detect the synthesis of
interleukin -8 (IL-8) by HUVECs incubated with inves-
tigated compounds.
The cells were rinsed with the Ca+2 and Mg+2 free PBS
and lysed in CelLyticTM-M Mammalian Cell Lysis/Extrac-
tion Reagent (Sigma) containing protease inhibitors (Protease
Inhibitor Cocktail Tablets complete TM, Roche). Protein
content was determined by Bradford’s method [20]. Proteins
were electrophoretically separated on an SDS-polyacryla-
mide gel under reducing conditions and transferred to PVDF
(polyvinylidine difluoride) membranes (BIORAD). Transfer
was performed in 100 V for 2 h. Blots, stained with 2%
Ponceau S (Sigma), in 3% trichloroacetic acid to visualize
proteins, were saturated with 1% blocking solution (Lumi-
LightPLUS Western Blotting Kit Mouse/Rabbit, Amersham
Pharmacia Biotech) and incubated for 1 hwithmouse anti-IL-
8 antibody (Mouse monoclonal anti-IL-8 antibody [B-2]).
Following washing in TBST (Lumi-LightPLUS Western
Blotting Kit Mouse/Rabbit), membranes were incubated for
30min, subsequently with anti-mouse Ig-POD Fab fragments
(Lumi-LightPLUS Western Blotting Kit Mouse/Rabbit,
Roche) or monoclonal anti-goat/sheep IgG Monoclonal
Anti-Goat/Sheep IgG Clone GT-34 peroxidase conjugated
(Sigma). Monoclonal antibodies against actin were used as a
control of protein loading (mouse monoclonal anti-actin
antibodies—Actin [C-2], Santa Cruz Biotechnology, Inc.).
Enhanced chemiluminescence, performed according to
manufacturer’s instructions (Amersham), was used to
demonstrate positive bands that were visualized after
exposure on a transparent medical X-ray film.
2.9. Isolation of total RNA
Following 24-h incubation with the studied compounds,
total RNA was isolated from HUVECs by the guanidine
thiocyanate–caesium chloride method [21] using Trizol
(Invitrogen Life Technologies) and was purified using the
SV total RNA Isolation System Kit (Promega). The quality
of RNA was confirmed by denaturing gel electrophoresis
and an analysis on the Agilent 2100 Bioanalyser (Agilent
Technologies).
2.10. Microarray affymetrix HG-U133A hybridization
The aim of the microarray experiments was to screen the
effects of beta-carotene (with or without arachidonic acid) on
gene expression patterns in HUVECs. For microarray
hybridization, RNAwas reverse transcribed into cDNA with
a primer containing the T7 promoter using Superscript II
(Invitrogen Life Technologies). cDNA was used as a
template in a biotin-labeled transcription reaction (Enzo
BioArray, Affymetrix). The resulting target cRNA was
purified on RNeasy columns (QIAGEN) and fragmented
for hybridization to Affymetrix HG-U133A GeneChips.
Hybridization was done overnight at 45 8C for 16 h in the
GeneChip Hybridization Oven 640 (Affymetrix). The
GeneChips were subsequently processed on the Affymetrix
GeneChip Fluidics Workstation 400 according to the
EukGE-WS2v4 protocol. The GeneChips were scanned with
the Hewlett Packard GeneArray Scanner, and the results
analyzed using Affymetrix Microarray Analysis Suit 4.0.
2.11. Analysis of the microarray data
Changes in relative gene expression were calculated
versus the control (THF/EtOH solvent). Only spots with
significant differences in signal intensity (more than 1.4-fold
and only when the P value was at least 0.05) were included
in the analysis.
In order to identify genes belonging to pathways regulated
by beta-carotene and/or arachidonic acid in HUVECs we
searched promotor sequences of the identified genes to find
similar transcription factors binding sites. The 4-kb upstream
sequences relative to the transcription start site were retrieved
from the database of the Transcriptional Start Sites DBTSS
(http://dbtss.hgc.jp/index.html). Searches of transcription
factor binding sites were done using the TRANSFAC
database at a default threshold setting of 90 (http://
molsun1.cbrc.aist.go.jp/research/db/TFSEARCH.html).
2.12. cDNA synthesis and quantitative real-time PCR
In order to confirm the regulation of the expression of
genes important for demonstration of the proangiogenic
activity of BC in HUVECs, which were identified by the
microarray experiments, the quantitative expression analysis
IL-8, CXCR4, VCAM-1, EGR-1, MAD1L1, BIRC5 and
MEOX2 was performed by real-time PCR using GAPDH as
the reference gene. For the cDNA synthesis 1 Ag of total RNAwas reverse transcribed at 42 8C for 50 min in a total volume
of 40 Al reaction buffer containing 5� First Strand Buffer,
DTT, oligo(dT) (Sigma), deoxy-NTPs (Promega), and 200
units of SUPERSCRIPT II reverse transcriptase (Invitrogen
Life Technologies). The reaction mixture was heated to 70 8Cfor 15 min and immediately chilled on ice. Subsequently,
cDNAwas subjected to real-time PCR in a reaction mixture
containing QuantiTect SYBR Green PCR (Qiagen) mix and
primers. The sequences of the primers used in this study are
presented in Table 1.The primers were designed to include an
intervening intron between the sense and antisense primers,
thereby eliminating the possibility of amplifying any genomic
DNA, and checked for specificity by BLAST searches. All
real-time PCR reactions were performed on the DNA Engine
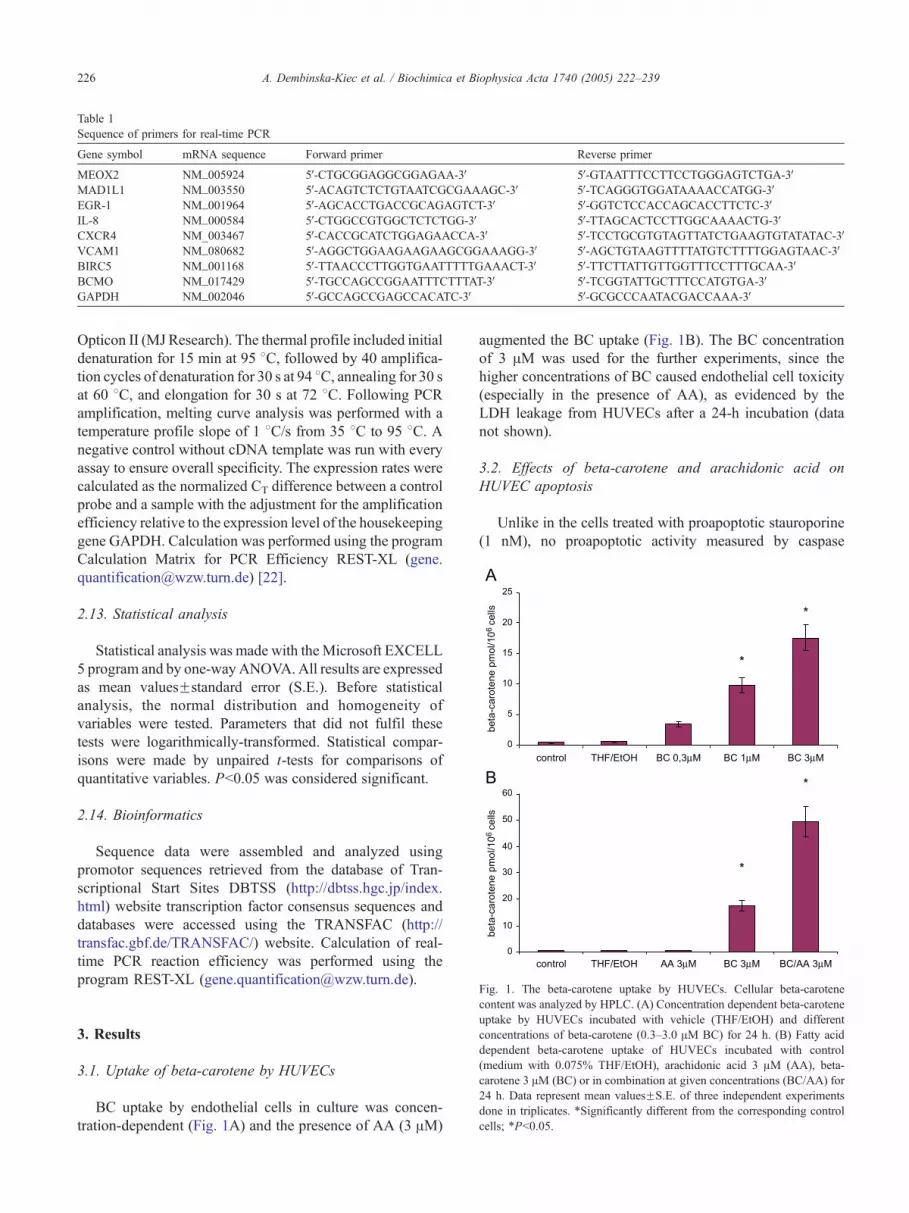
Table 1
Sequence of primers for real-time PCR
Gene symbol mRNA sequence Forward primer Reverse primer
MEOX2 NM_005924 5V-CTGCGGAGGCGGAGAA-3V 5V-GTAATTTCCTTCCTGGGAGTCTGA-3VMAD1L1 NM_003550 5V-ACAGTCTCTGTAATCGCGAAAGC-3V 5V-TCAGGGTGGATAAAACCATGG-3VEGR-1 NM_001964 5V-AGCACCTGACCGCAGAGTCT-3V 5V-GGTCTCCACCAGCACCTTCTC-3VIL-8 NM_000584 5V-CTGGCCGTGGCTCTCTGG-3V 5V-TTAGCACTCCTTGGCAAAACTG-3VCXCR4 NM_003467 5V-CACCGCATCTGGAGAACCA-3V 5V-TCCTGCGTGTAGTTATCTGAAGTGTATATAC-3VVCAM1 NM_080682 5V-AGGCTGGAAGAAGAAGCGGAAAGG-3V 5V-AGCTGTAAGTTTTATGTCTTTTGGAGTAAC-3VBIRC5 NM_001168 5V-TTAACCCTTGGTGAATTTTTGAAACT-3V 5V-TTCTTATTGTTGGTTTCCTTTGCAA-3VBCMO NM_017429 5V-TGCCAGCCGGAATTTCTTTAT-3V 5V-TCGGTATTGCTTTCCATGTGA-3VGAPDH NM_002046 5V-GCCAGCCGAGCCACATC-3V 5V-GCGCCCAATACGACCAAA-3V
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239226
Opticon II (MJ Research). The thermal profile included initial
denaturation for 15 min at 95 8C, followed by 40 amplifica-
tion cycles of denaturation for 30 s at 94 8C, annealing for 30 sat 60 8C, and elongation for 30 s at 72 8C. Following PCR
amplification, melting curve analysis was performed with a
temperature profile slope of 1 8C/s from 35 8C to 95 8C. Anegative control without cDNA template was run with every
assay to ensure overall specificity. The expression rates were
calculated as the normalized CT difference between a control
probe and a sample with the adjustment for the amplification
efficiency relative to the expression level of the housekeeping
gene GAPDH. Calculation was performed using the program
Calculation Matrix for PCR Efficiency REST-XL (gene.
[email protected]) [22].
2.13. Statistical analysis
Statistical analysis was made with the Microsoft EXCELL
5 program and by one-wayANOVA. All results are expressed
as mean valuesFstandard error (S.E.). Before statistical
analysis, the normal distribution and homogeneity of
variables were tested. Parameters that did not fulfil these
tests were logarithmically-transformed. Statistical compar-
isons were made by unpaired t-tests for comparisons of
quantitative variables. Pb0.05 was considered significant.
2.14. Bioinformatics
Sequence data were assembled and analyzed using
promotor sequences retrieved from the database of Tran-
scriptional Start Sites DBTSS (http://dbtss.hgc.jp/index.
html) website transcription factor consensus sequences and
databases were accessed using the TRANSFAC (http://
transfac.gbf.de/TRANSFAC/) website. Calculation of real-
time PCR reaction efficiency was performed using the
program REST-XL ([email protected]).
Fig. 1. The beta-carotene uptake by HUVECs. Cellular beta-carotenecontent was analyzed by HPLC. (A) Concentration dependent beta-carotene
uptake by HUVECs incubated with vehicle (THF/EtOH) and different
concentrations of beta-carotene (0.3–3.0 AM BC) for 24 h. (B) Fatty acid
dependent beta-carotene uptake of HUVECs incubated with control
(medium with 0.075% THF/EtOH), arachidonic acid 3 AM (AA), beta-
carotene 3 AM (BC) or in combination at given concentrations (BC/AA) for
24 h. Data represent mean valuesFS.E. of three independent experiments
done in triplicates. *Significantly different from the corresponding control
cells; *Pb0.05.
3. Results
3.1. Uptake of beta-carotene by HUVECs
BC uptake by endothelial cells in culture was concen-
tration-dependent (Fig. 1A) and the presence of AA (3 AM)
augmented the BC uptake (Fig. 1B). The BC concentration
of 3 AM was used for the further experiments, since the
higher concentrations of BC caused endothelial cell toxicity
(especially in the presence of AA), as evidenced by the
LDH leakage from HUVECs after a 24-h incubation (data
not shown).
3.2. Effects of beta-carotene and arachidonic acid on
HUVEC apoptosis
Unlike in the cells treated with proapoptotic stauroporine
(1 nM), no proapoptotic activity measured by caspase

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 227
activation was detected in HUVECs incubated for 24 h with
the BC or AA at 3 AM, as well as with the mixture of both
compounds (Fig. 2).
3.3. Effect of beta-carotene and arachidonic acid on
HUVEC proliferation
Unlike VEGF (0.2 nM) and bFGF (0.5 nM), which
stimulated cell proliferation, both BC as well as AA
used in the non-toxic concentrations did not influence
HUVEC proliferation measured by the BrdU incorporation
(Fig. 3).
3.4. Effect of beta-carotene and arachidonic acid on
HUVEC migration
BC (3 AM) four-fold increased the migration of HUVECs
(Fig. 4). Arachidonic acid alone did not stimulate HUVEC
migration, but the cell migration was further increased (six-
fold) when AA was used in combination with BC (Fig. 4).
Sphingosine-1-phosphate (S1P) the potent activator of
endothelial cell migration [18] was used as the positive
control to confirm HUVEC chemotactic potential in our
assay.
3.5. Effect of beta-carotene and arachidonic acid on
tubulogenesis in the 3D matrigel in vitro model
The 3D matrigel assay of tubulogenesis was used to
verify the angiogenic property of BC and AA in vitro [19].
Only the trace tubulogenic activity of HUVEC was detected
in cells cultured in matrigel covered with medium without
factors. No tubulogenic activity of BC and AA incubated
alone as well as together was observed in the above in vitro
model (Fig. 5A,B). The proangiogenic VEGF (0.2 nM) and
bFGF (0.5 nM) potently increased the number of tubules in
matrigel HUVEC suspension (Fig. 5A,B).
Fig. 2. Influence of beta-carotene and arachidonic acid on the HUVEC apoptosis.
caspase activity following a 24-h incubation of HUVEC with the studied facto
arachidonic acid (AA) 3 AM or both compounds (BC/AA). Data represent mean va
the percentage of activity detected in the control. *Significantly different from th
3.6. The proangiogenic activity of beta-carotene in the in
vivo mouse model
The presence of BC at 3 AM in the matrigel administered
subcutaneously significantly increased the density of capil-
lary network (with and without lumen) and the amount of
endothelial cells penetrating to the matrigel plug stimulated
by bFGF (Fig. 6).
3.7. The microarray analysis
Using the criteria described in Methods, we identified
838 genes, whose expression changed only in response to
the stimulation with BC (446 down-regulated, 393 up-
regulated). The expression of 644 genes was regulated by
AA (375 down-regulated, 269 up-regulated) and the
expression of 740 genes was regulated by beta carotene
and AA together (498 down-regulated, 242 up-regulated).
The analysis of microarray data, including genes regulated
(up or down) from 1.4-fold up to 10-fold, provided a list of
the selected significantly regulated genes coding for
proteins, which belong to the cellular pathways such as:
pathways contributing to proangiogenic activity (cell cycle,
adhesion, matrix remodeling, chemotaxis), apoptosis, recep-
tor-mediated signal transduction, as well as transcription
factors and regulators of protein synthesis (zinc finger
proteins, ribosomal proteins), xenobiotic metabolism,
inflammatory response (Table 2).
According to the recent data concerning the proangio-
genic activity of endothelial cells [1,23], special attention
was paid to the BC-regulated genes involved in the cell
cycle-proliferation. BC weakly up-regulated the key genes
coding for proteins participating in the regulation of a cell
cycle such as MCM5, MAD1L1 connected with G1/S check
point and polo-like kinase Plk1, NUCKS related to G2/M
check point. BC up-regulated such important inhibitors of
cell cycle as Wee1, PKMyt1 (Table 2) [24].
The rate of apoptosis was estimated by the measurement of global cellular
rs (control: medium with 0.075% THF/EtOH), beta-carotene (BC) 3 AM,
luesFS.E. of three independent experiments done in triplicates, expressed as
e corresponding control, *Pb0.05.

Fig. 3. Effects of beta-carotene and arachidonic acid on HUVEC proliferation. HUVEC cultured in EBM medium containing 10% FBS and antibiotics were
treated with medium with 0.075% THF/EtOH (control), beta-carotene (BC) 3 AM, arachidonic acid (AA) 3 AM or a combination of both (BC/AA) for 24 h
were labeled with BrdU for 3 h before the end of incubation. BrdU incorporation was determined by collorimetric immunoassay. The results are given as the
percent of the control proliferation. Data represent mean valuesFS.E. of three independent experiments performed in triplicates. Significantly different from the
corresponding control, *Pb0.05, **Pb0.005.
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239228
BC also weakly up-regulated the expression of apoptosis
inhibitors associated with the FASL pathway such as
CFLAR (Flip), BIRC5 (IAP) TRAF4 genes and down-
regulated the expression of apoptosis activators OPTN,
PAR1 and PAR4 [25]. The parallel up-regulation of the
expression of a number of pro- and anti-apoptotic members
of the Bcl-2 family was observed. BC up-regulated the
proapoptotic Bcl2L11/BAM, anti-apoptotic MCL-1 and
TEGT-BAX inhibitor (Table 2).
The group of BC-regulated genes, which code for
proteins participating in cell–cell interactions, such as
cadherins (CELSR1), catenins (CTNNA1L, CTNNB1),
and the leukocyte–endothelium adhesion mediating mole-
Fig. 4. Beta-carotene-induced chemotaxis of HUVEC. Cells were seeded onto up
prefilled with EBM medium with 0.5% FBS containing the chemoattractant b
sphingosine-1-phosphate (S1P) 500 nM. Cells treated with S1P were used as p
micropore inserts (5-Am pore size) to the lower surface of the membranes were
Chemotaxis index: the ratio of stimulated migration divided by that of basal, unst
valuesFS.E.; n=3 done in triplicates. Significantly different from the correspondi
cules (VCAM1, SELP, CD24) [26], were generally down-
regulated by both BC and AA (Table 2). On the contrary, the
expression of genes coding for proteins associated with cell-
extracellular matrix adhesion, such as integrins (ITGA6),
scavenger receptor SCARB1, was up-regulated by BC
(Table 2).
The expressions of genes encoding extracellular matrix
degrading enzymes and stimulators of chemotaxis, which
may regulate the matrix-degradation [27], receptor shading
[28] and cell migration, such as ADAMTS1, ADAMTS18,
MMP10, MMP12, MMP14, MMP24, were differentially
regulated. The down-regulation of different types of
extracellular matrix components collagens, fibrillin 1
per well membranes of Transwell plate inserts and then placed into wells
eta-carotene (BC) 3 AM or arachionic acid (AA) 3 AM or BC/AA and
ositive controls. After incubation for 24 h, cells that had migrated across
stained by anti-human CD31 antibody coupled with phycoerythrin (PE).
imulated migration of HUVEC in control medium sample. Values are mean
ng control, *Pb0.05, **Pb0.005.
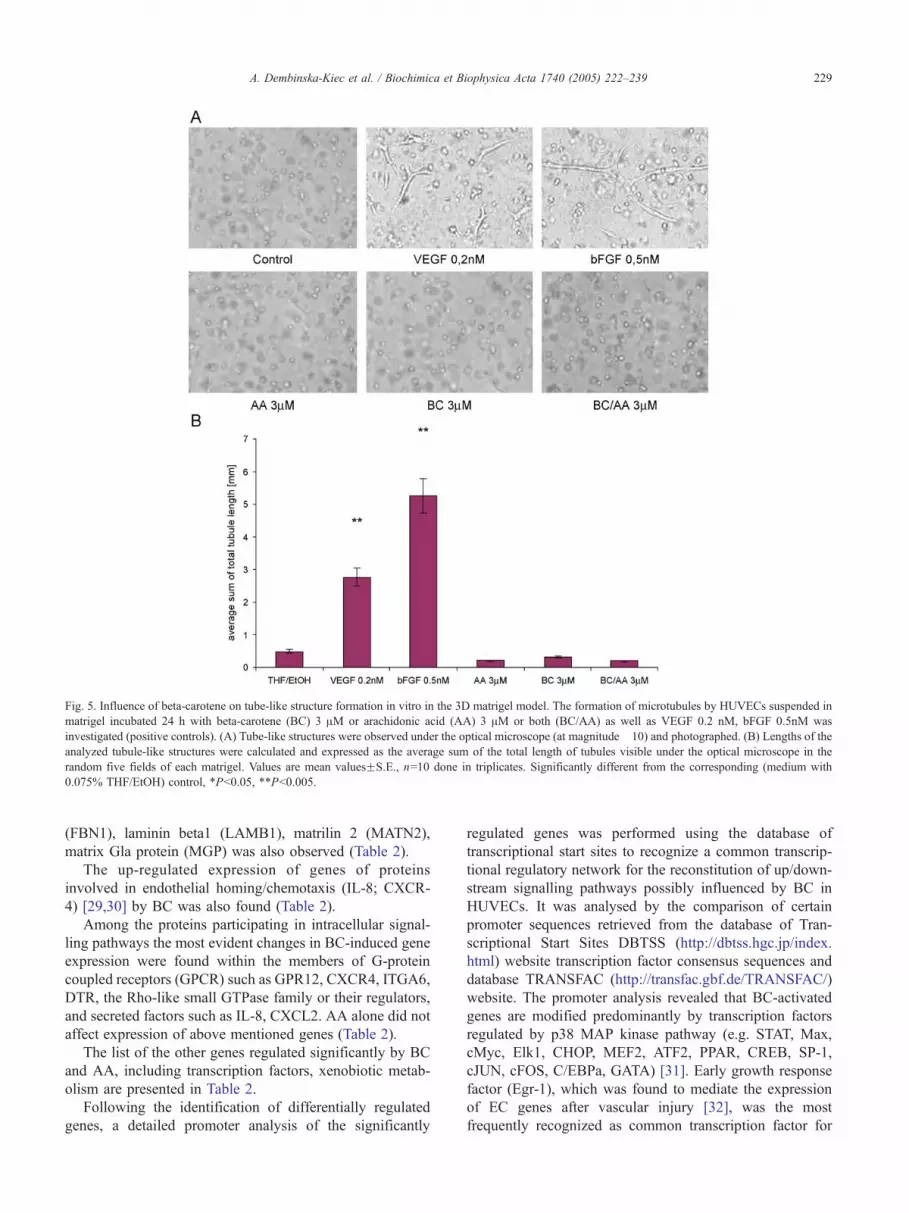
Fig. 5. Influence of beta-carotene on tube-like structure formation in vitro in the 3D matrigel model. The formation of microtubules by HUVECs suspended in
matrigel incubated 24 h with beta-carotene (BC) 3 AM or arachidonic acid (AA) 3 AM or both (BC/AA) as well as VEGF 0.2 nM, bFGF 0.5nM was
investigated (positive controls). (A) Tube-like structures were observed under the optical microscope (at magnitude �10) and photographed. (B) Lengths of the
analyzed tubule-like structures were calculated and expressed as the average sum of the total length of tubules visible under the optical microscope in the
random five fields of each matrigel. Values are mean valuesFS.E., n=10 done in triplicates. Significantly different from the corresponding (medium with
0.075% THF/EtOH) control, *Pb0.05, **Pb0.005.
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 229
(FBN1), laminin beta1 (LAMB1), matrilin 2 (MATN2),
matrix Gla protein (MGP) was also observed (Table 2).
The up-regulated expression of genes of proteins
involved in endothelial homing/chemotaxis (IL-8; CXCR-
4) [29,30] by BC was also found (Table 2).
Among the proteins participating in intracellular signal-
ling pathways the most evident changes in BC-induced gene
expression were found within the members of G-protein
coupled receptors (GPCR) such as GPR12, CXCR4, ITGA6,
DTR, the Rho-like small GTPase family or their regulators,
and secreted factors such as IL-8, CXCL2. AA alone did not
affect expression of above mentioned genes (Table 2).
The list of the other genes regulated significantly by BC
and AA, including transcription factors, xenobiotic metab-
olism are presented in Table 2.
Following the identification of differentially regulated
genes, a detailed promoter analysis of the significantly
regulated genes was performed using the database of
transcriptional start sites to recognize a common transcrip-
tional regulatory network for the reconstitution of up/down-
stream signalling pathways possibly influenced by BC in
HUVECs. It was analysed by the comparison of certain
promoter sequences retrieved from the database of Tran-
scriptional Start Sites DBTSS (http://dbtss.hgc.jp/index.
html) website transcription factor consensus sequences and
database TRANSFAC (http://transfac.gbf.de/TRANSFAC/)
website. The promoter analysis revealed that BC-activated
genes are modified predominantly by transcription factors
regulated by p38 MAP kinase pathway (e.g. STAT, Max,
cMyc, Elk1, CHOP, MEF2, ATF2, PPAR, CREB, SP-1,
cJUN, cFOS, C/EBPa, GATA) [31]. Early growth response
factor (Egr-1), which was found to mediate the expression
of EC genes after vascular injury [32], was the most
frequently recognized as common transcription factor for

Fig. 6. Effects of beta-carotene on angiogenesis in mouse matrigel model in
vivo. Angiogenesis was assayed in subcutaneously injected matrigel plugs
containing medium with 0.075% THF/EtOH (as control), bFGF 50 nM or
beta-carotene 3 AM (BC) or both isolated after 6 days. Angiogenesis was
quantified by counting. (A) The number of vessels without lumen. (B) The
number of vessels with lumen. (C) The number of endothelial CD31
(PECAM)-positive cells penetrating the matrigel plug, as average in five
different fields in each of the three slices taken from different regions
throughout the each matrigel plug. For immunostaining, sections were
incubated with mouse monoclonal anti-CD31 antibody. Values are mean
valuesFS.E., n=6 per group. *Significantly different from the correspond-
ing control, *Pb0.05.
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239230
BC-regulated genes or for described transcription factors
coactivator or target.
3.8. Changes in relative gene expression (real-time PCR)
In order to confirm the microarray results related to
the biological effects of BC in HUVECs, the expression
of selected genes was verified by the quantitative real-
time PCR.
The direction of beta-carotene expression regulation was
found similar to the microarray data with respect to the
factors, whose activity is related to proliferation (MAD1L1,
MEOX2) [24,33], chemotaxis (IL-8, CXCR-4, VCAM1),
[30,29,26], and inhibition of tubulogenesis (MEOX2) (Fig.
7). Additionally, the BC-induced difference in the EGR-1
gene expression was detected, consistent with the up-
regulation of DR1 and NAB1 (the inhibitors of the
transcription factor EGF-1, which is important for endothe-
lial cell proangiogenic activity) [32] observed in the
microarray experiments (Fig. 7).
The basal expression of h,h-carotene 15,15V-monoxyge-
nase (BCMO, EC 1.13.11.21) in HUVECs detected by real-
time PCR and also demonstrated the lack of BC-induced
changes was in agreement with the microarray assay (Fig. 8).
3.9. Western blotting analysis of IL-8 protein level
The activation of the protein expression of chemotactive
IL-8 by BC was confirmed by Western blot. Fig. 9A shows
a Western blot of total cell lysates probed with an antibody
to detect IL-8 protein. Densitometric analysis revealed that
incubation of HUVEC with BC, AA or BC with AA
increased IL-8 protein level (Fig. 9B).
4. Discussion
The main result of presented study is the original
demonstration of the proangiogenic activity of BC in vivo
and the involvement of BC-induced chemotactic activity of
endothelial cells in this process.
Beta-carotene, alpha-carotene, h-cryptoxanthin lutein
and lycopene belong to the well-characterized family of
carotenoids constituting approximately 90% of the total
plasma carotenoids in characterized humans in concentra-
tions ranging from undetectable to 3–10 Amol/l (dependent
upon diet or supplementation) [34]. Dietary lipids are
important for the absorption of carotenoids both in vivo
and in the cell culture system [35]. Also, in our in vitro
experimental model, AA increased the uptake of BC by
HUVECs, which was documented by HPLC (Fig. 1B). This
was not related to the increase of the basal expression of
h,h-carotene 15,15V-monoxygenase (BCMO) (the micro-
array experiments, confirmed later by RT-PCR). BCMO is
the key enzyme that cleaves beta-carotene centrally into two
molecules of retinal, the source of retinoic acid or retinol
(Vitamin A). Thus, the observed effects of BC may
be related to its direct free radical scavenger activity [36]
and/or possible activity of its metabolites (mainly retinoic
acid), generated locally by HUVECs.
The results of the microarray experiments indicate that
the degree of change in gene expression regulated by the
used concentration of BC in HUVECs is moderate. Also,
the expression of 95% of up- or down-regulated genes was
changed less than 2-fold. It is important to stress that non-
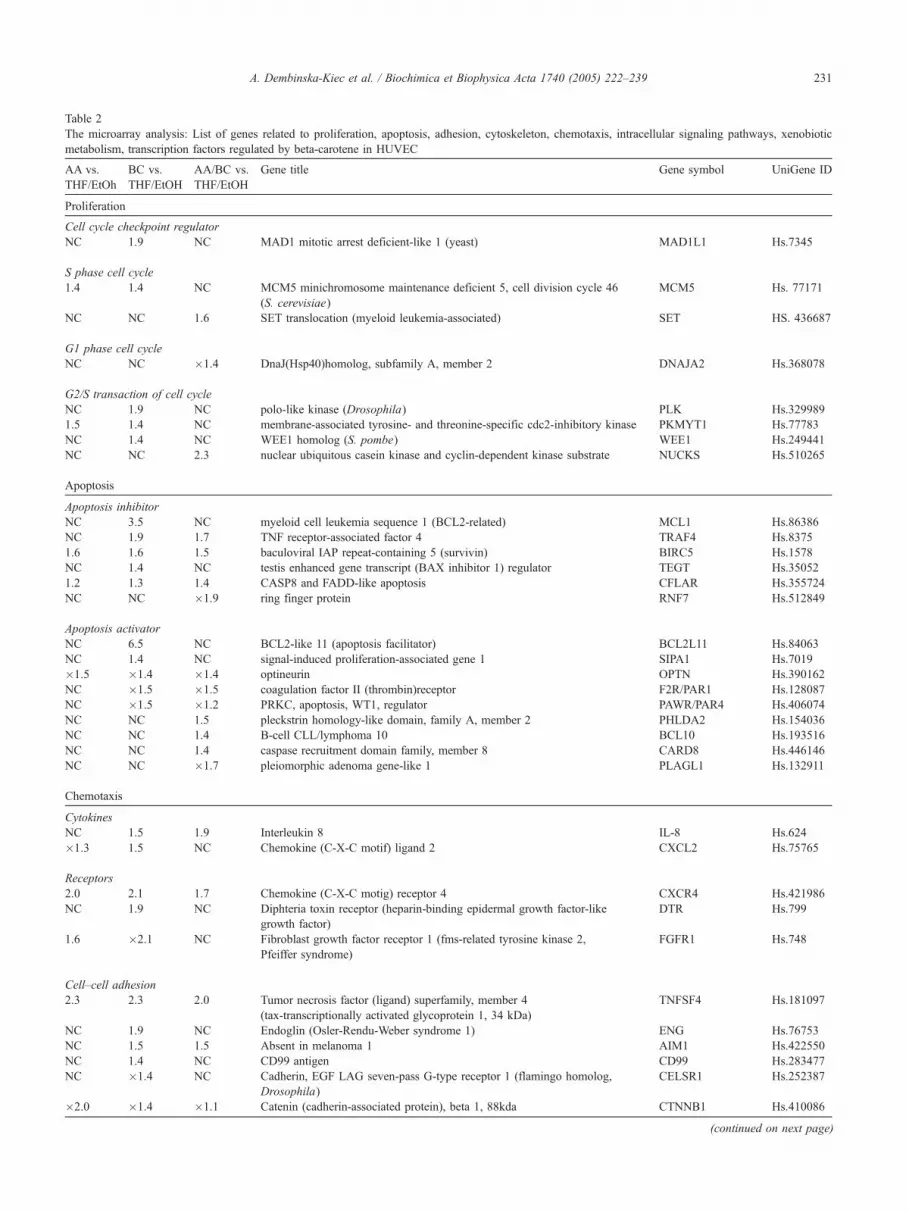
Table 2
The microarray analysis: List of genes related to proliferation, apoptosis, adhesion, cytoskeleton, chemotaxis, intracellular signaling pathways, xenobiotic
metabolism, transcription factors regulated by beta-carotene in HUVEC
AA vs.
THF/EtOh
BC vs.
THF/EtOH
AA/BC vs.
THF/EtOH
Gene title Gene symbol UniGene ID
Proliferation
Cell cycle checkpoint regulator
NC 1.9 NC MAD1 mitotic arrest deficient-like 1 (yeast) MAD1L1 Hs.7345
S phase cell cycle
1.4 1.4 NC MCM5 minichromosome maintenance deficient 5, cell division cycle 46
(S. cerevisiae)
MCM5 Hs. 77171
NC NC 1.6 SET translocation (myeloid leukemia-associated) SET HS. 436687
G1 phase cell cycle
NC NC �1.4 DnaJ(Hsp40)homolog, subfamily A, member 2 DNAJA2 Hs.368078
G2/S transaction of cell cycle
NC 1.9 NC polo-like kinase (Drosophila) PLK Hs.329989
1.5 1.4 NC membrane-associated tyrosine- and threonine-specific cdc2-inhibitory kinase PKMYT1 Hs.77783
NC 1.4 NC WEE1 homolog (S. pombe) WEE1 Hs.249441
NC NC 2.3 nuclear ubiquitous casein kinase and cyclin-dependent kinase substrate NUCKS Hs.510265
Apoptosis
Apoptosis inhibitor
NC 3.5 NC myeloid cell leukemia sequence 1 (BCL2-related) MCL1 Hs.86386
NC 1.9 1.7 TNF receptor-associated factor 4 TRAF4 Hs.8375
1.6 1.6 1.5 baculoviral IAP repeat-containing 5 (survivin) BIRC5 Hs.1578
NC 1.4 NC testis enhanced gene transcript (BAX inhibitor 1) regulator TEGT Hs.35052
1.2 1.3 1.4 CASP8 and FADD-like apoptosis CFLAR Hs.355724
NC NC �1.9 ring finger protein RNF7 Hs.512849
Apoptosis activator
NC 6.5 NC BCL2-like 11 (apoptosis facilitator) BCL2L11 Hs.84063
NC 1.4 NC signal-induced proliferation-associated gene 1 SIPA1 Hs.7019
�1.5 �1.4 �1.4 optineurin OPTN Hs.390162
NC �1.5 �1.5 coagulation factor II (thrombin)receptor F2R/PAR1 Hs.128087
NC �1.5 �1.2 PRKC, apoptosis, WT1, regulator PAWR/PAR4 Hs.406074
NC NC 1.5 pleckstrin homology-like domain, family A, member 2 PHLDA2 Hs.154036
NC NC 1.4 B-cell CLL/lymphoma 10 BCL10 Hs.193516
NC NC 1.4 caspase recruitment domain family, member 8 CARD8 Hs.446146
NC NC �1.7 pleiomorphic adenoma gene-like 1 PLAGL1 Hs.132911
Chemotaxis
Cytokines
NC 1.5 1.9 Interleukin 8 IL-8 Hs.624
�1.3 1.5 NC Chemokine (C-X-C motif) ligand 2 CXCL2 Hs.75765
Receptors
2.0 2.1 1.7 Chemokine (C-X-C motig) receptor 4 CXCR4 Hs.421986
NC 1.9 NC Diphteria toxin receptor (heparin-binding epidermal growth factor-like
growth factor)
DTR Hs.799
1.6 �2.1 NC Fibroblast growth factor receptor 1 (fms-related tyrosine kinase 2,
Pfeiffer syndrome)
FGFR1 Hs.748
Cell–cell adhesion
2.3 2.3 2.0 Tumor necrosis factor (ligand) superfamily, member 4
(tax-transcriptionally activated glycoprotein 1, 34 kDa)
TNFSF4 Hs.181097
NC 1.9 NC Endoglin (Osler-Rendu-Weber syndrome 1) ENG Hs.76753
NC 1.5 1.5 Absent in melanoma 1 AIM1 Hs.422550
NC 1.4 NC CD99 antigen CD99 Hs.283477
NC �1.4 NC Cadherin, EGF LAG seven-pass G-type receptor 1 (flamingo homolog,
Drosophila)
CELSR1 Hs.252387
�2.0 �1.4 �1.1 Catenin (cadherin-associated protein), beta 1, 88kda CTNNB1 Hs.410086
(continued on next page)
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 231

Table 2 (continued)
AA vs.
THF/EtOh
BC vs.
THF/EtOH
AA/BC vs.
THF/EtOH
Gene title Gene symbol UniGene ID
Chemotaxis
Cell–cell adhesion
NC �1.4 NC Sialophorin (gpl115, leukosialin,CD43) SPN Hs.461934
�2.1 �1.4 �1.7 Selectin P (granule membrane protein 140kDa,CD43) SELP Hs.73800
�2.6 �2.1 NC CD 24 antigen (small cell lung carcinoma cluster 4 antigen) CD24 Hs.375108
�2.6 �2.6 NC CD 24 antigen (small cell lung carcinoma cluster 4 antigen) CD24 Hs.375108
�10.6 �2.6 �5.3 Vascular cell adhesion molecule 1 VCAM1 Hs.109225
NC �5.7 1.6 EphB2 EPHB2 Hs.125124
NC NC �1.4 Activate leukocyte cell adhesion molecule ALCAM Hs.10247
NC NC 1.4 Carcinoembryonic antigen-related cell adhesion molecule 4 CEACAM4 Hs.12
NC NC �1.4 Catenin (cadherin-associated protein), alpha-like 1 CTNNAL1 Hs.58488
NC NC �1.4 Desmoplakin DSP Hs.349499
Cell–matrix adhesion
NC 1.9 NC Integrin, alpha ITGA6 Hs.212296
1.5 1.6 NC Scavenger receptor class B, member 1 SCARB1 Hs.130981
Exracellular matrix component
1.7 1.6 1.9 Collagen,type XIII, alpha 1 COL13A1 Hs.211933
1.7 1.6 NC Nidogen 2 (osteonidogen) NID2 Hs.147697
1.5 1.5 1.5 Fibrillin 2 (congenital contractural archnodactyly) FBN2 Hs.79432
�1.4 �1.4 �1.3 Collagen, type V, alpha 2 COL5A2 Hs.283393
�1.3 �1.4 �1.4 Fibrillin 1 (Marfan syndrome) FBN1 Hs.750
NC �1.4 NC Kallman syndrome 1 sequence KAL1 Hs.380850
�1.3 �1.4 �1.2 Laminin, beta 1 LAMB1 Hs.122645
�2.1 �1.9 �2.1 Sulfatase 1 SULF1 Hs.409602
NC �2.0 NC Exostoses (multiple) 1 EXT1 Hs.184161
�2.3 �2.8 �2.5 Collagen, type VIII, alpha COL8A1 Hs.114599
�3.2 �2.8 �3.5 Matrix Gla protein MGP Hs.365706
�4.0 �4.3 �3.7 Matrillin 2 MATN2 Hs.153647
�1.5 NC �1.6 Collagen, type III, alpha 1 (Ehlers-Danlos syndrome type IV, autosomal
dominant)
COL3A1 Hs.443625
Cellular proteases
NC 9.8 NC Membrane metallo-endopeptidase (neutral endopeptidase, enkephaline,
CALLA, CD10)
MME Hs.307734
NC 2.5 NC Matrix metalloproteinase 14 (membrane-inserted) MMP14 Hs.2399
1.3 1.9 1.6 Serine (or cysteine) proteinase inhibitor, clade B (ovalbumin), member 2 SERPINB2 Hs.75716
NC 1.7 NC Plasminigen activator, tissue PLAT Hs.274404
�1.7 �1.4 �1.7 A disintegrin-like and metalloprotease (reprolysin type) with
thrombosposin type 1 motif, 1
ADAMTS1 Hs.8230
�2.1 �2.3 �1.9 Matrix metalloprotainase 12 (macrophage elastase) MMP12 Hs.1695
NC NC 2.3 A disintegrin and metalloproteinase domain 18 ADAM18 Hs.127930
NC NC 1.4 Matrix metalloprotainase 24 (membrane-inserted) MMP24 Hs.212581
NC NC �1.4 Matrix metalloproteinase 10 (stromelysin 2) MMP10 Hs.2258
Cytoskeleton
NC 1.6 NC Intersectin 1 (SH3 Domain protein) ITSN1 Hs.66392
NC 1.6 NC Lamin B1 LMNB1 Hs.89497
NC 1.6 NC Tubulin, beta polypeptide TUBB Hs.512712
NC 1.5 1.3 Myosin IB MYO1B Hs.121576
NC 1.4 NC Actinin, alpha 4 ACTN4 Hs.443619
NC 1.4 NC Flotillin 2 FLOT2 Hs.18799
NC 1.4 NC Myosin IXB MYO9B Hs.159629
NC 1.4 NC Transgellin 2 TAGLN2 Hs.406504
NC �1.4 NC Transgellin TAGLN Hs.410977
�1.9 �1.7 �2.5 Myosin, heavy polypeptide 10, non-muscle MYH10 Hs.280311
NC NC 1.7 Caveolin 2 CAV2 Hs.139851
NC NC 1.4 Utophin (homologous to dystrophin) UTRN Hs.250607
NC NC �1.4 Kelch-like 3 (Drosophila) KLHL3 Hs.434434
NC NC �1.6 Filamin A,alpha (actin binding protein 280) FLNA Hs.195464
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239232

Table 2 (continued)
AA vs.
THF/EtOh
BC vs.
THF/EtOH
AA/BC vs.
THF/EtOH
Gene title Gene symbol UniGene ID
Receptors
G protein coupled receptor
2.0 2.1 1.7 Chemokine (C-X-C motif) receptor 4 CXCR4 Hs.421986
NC 2.0 NC G protein-coupled receptor 12 GPR12 Hs.123034
NC 1.9 NC Diphtheria toxin receptor (heparin-binding epidermal growth factor-like
growth factor)
DTR Hs.799
NC 1.9 NC Integrin, alpha 6 ITGA6 Hs.212296
NC �1.4 NC Cadherin, EGF LAG seven-pass G-type receptor 1 (flamingo homolog,
Drosophilia)
CELSR1 Hs.252387
NC �1.5 �1.5 Coagulation factor II (thrombin) receptor F2R/PAR1 Hs.128087
NC �1.5 �1.2 PRKC, apoptosis, WT1, regulator PAWR/PAR4 Hs.406074
NC NC 4.9 Somatostatin receptor 2 SSTR2 Hs.184841
NC NC 2.8 Pyrimidnergic receptor P2Y, G-protein coupled, 4 P2RY4 Hs.248157
NC NC 1.6 Dopamine receptor D5 DRD5 Hs.380681
NC NC 1.5 G protein-coupled receptor 27 GPR27 Hs.356084
�1.4 NC �1.7 Calcitonin receptor-like CALCRL Hs.152175
FGF receptor mediated signalling pathway
1.6 �2.1 NC Fibroblast growth factor receptor 1 (fms-related thyrosine kinase 2,Pfeiffer
syndrome)
FGFR1 Hs.748
Eph receptor mediated signalling pathway
NC NC 1.6 EphB2 EPHB2 Hs.125124
Interleukin receptor mediated signalling pathway
NC 1.5 1.9 Interleukin 8 IL-8 Hs.624
�1.3 1.5 NC Chemokine (C-X-C motif) ligand 2 CXCL2 Hs.75765
�1.5 �1.5 �1.3 Interleukin 1 receptor, type 1 IL1R1 Hs.82112
NC NC �1.4 Interleukin 10 receptor, beta IL1ORB Hs.418291
NC NC �1.5 Interleukin 13 receptor, alpha IL13RA1 Hs.285115
Intracellular signalling
Small GTPase mediated signal transdustion
NC 1.7 NC CAP, adenylate cyclase-associated protein, 2 (yeast) CAP2 Hs.296341
�1.3 1.5 NC Chemokine (C-X-C motif) ligand 2 CXCL2 Hs.75765
NC 1.5 1.9 Interleukin 8 IL-8 Hs.624
NC 1.4 NC Myosin IXB MYO9B Hs.159629
NC 1.4 NC Res-related C3 botulinum toxin substrate 2 (rho family, small GTP binding
protein Rac 2)
RAC2 Hs.301175
�1.9 �1.3 �1.4 ADP-ribisylation factor-like 4 ARL4 Hs.245540
NC �1.4 NC RAP2C, member of RAS oncogene family RAP2C Hs.225979
NC �1.6 NC Guanine nucleotide exchange factor for Rap 1 GFR Hs.449375
�1.5 �2.0 �1.7 Regulator of G-protein signalling 4 RGS4 Hs.386726
NC NC 1.4 Rho/Rac guanine nucleotide exchange factor (GEF) 2 ARHGEF2 Hs.337774
NC NC �1.4 RAN, member RAS oncogene family RAN Hs.10842
NC NC �1.7 Rap2 interacting protein x RIPX Hs.7972
Protein kinase cascade
NC �1.4 �1.9 Mitogen-activated protein kinase kinase kinase 7 MAP3K7 Hs.290346
�1.5 �1.9 �2.1 Transformer-2 alpha TRA2A Hs.445652
NC NC �1.4 Mitogen-activated protein kinase kinase kinase 5 MAP4K5 Hs.246970
NFjBNC 1.7 NC Toll-like receptor 4 TLR4 Hs.174312
JAK/STA
NC �1.5 �1.5 Coagulation factor II (thrombin) receptor F2R/PAR1 Hs.128087
JNK
NC NC �1.4 Mitogen-activated protein kinase kinase kinase kinase 5 MAP4K5 Hs.246970
(continued on next page)
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 233

Table 2 (continued)
AA vs.
THF/EtOh
BC vs.
THF/EtOH
AA/BC vs.
THF/EtOH
Gene title Gene symbol UniGene ID
Intracellular signalling
Phosphatidiloinositol pathway
NC 3.5 NC Myotubularin related protein 1 MTMR1 Hs.347187
NC �2.0 NC Dual adaptor of phosphotyrosine and 3-phosphoinositides DAPP1 Hs.62643
Nitric oxide
NC 1.4 NC Nitric oxide synthase 3 (endothelial cell) NOS3 Hs.446303
NC 1.6 1.3 NAD(P)H dehydrogenase, quinone 1 NQ01 Hs.406515
NC NC 1.5 GTP cyclohyrolase I feedback regulatory protein GCHFR Hs.245644
NC NC �1.6 Guanylate cyclase 1, soluble, alpha 3 GUCY1A3 Hs.433488
SH2/SH3
NC 1.7 NC SH3-domain binding protein 2 SH3BP2 Hs.167679
NC 1.4 NC 1 SHC1 Hs.433795
Transcription factors
NC 3.7 2.5 LIM homebox 3 LHX3 Hs.148427
NC 2.0 1.4 Cbp/p300-interacting transactivator, with Glu/Asp-rich carboxy-terminal
domain 2
CITED2 Hs.82071
NC 1.9 NC MAD1 mitotic arrest deficient-like 1 (yeast) MAD1L1 Hs.7345
2.0 1.9 1.3 Mesenchyme homeo box 2 (growth arrest-specific homeo box) MEOX2 Hs.77858
NC 1.7 NC General transcription factor IIIH, polypeptide 4 52 kDa GTF2H4 Hs.102910
NC 1.6 NC Forehead box M1 FOXM1 Hs.511941
NC 1.6 NC Jun B proto-oncogene JUNB Hs.400124
NC 1.6 2.5 Early growth response 1 EGR1 Hs.326035
NC 1.5 NC Heat shock transcription factor 1 HSF1 Hs.132625
NC 1.4 NC CAMP responsive element binding protein 1 CREB1 Hs.22315
NC 1.4 NC Down-regulator o transcription 1, TBP-binding (negative cofactor 2) DR1 Hs.348418
1.4 1.4 1.5 Hairy/enhancer-of-split related with YRPW motif 1 HEY1 Hs.234434
NC 1.4 NC Kruppel-like factor 2 (lung) KLF2 Hs.107740
1.3 1.4 NC NGFI-A binding protein 1 (EGR1 binding protein 1) NAB1 Hs.107474
NC 1.4 NC SRY (sex determining region Y)-box 13 SOX13 Hs.201671
NC 1.3 1.5 LIM domain only 4 LMO4 Hs.3844
1.4 1.3 1.4 Pituitary tumor-transforming 1 PTTG1 Hs.350966
NC 1.2 1.4 Hematopoietic cell-specific Lyn substrate HCLS1 Hs.14601
NC �1.3 �1.5 Zinc finger protein 36, C3H type-like 1 ZFP36L1 Hs.85155
NC �1.3 �1.4 Zinc finger protein 24 (KOX 17) ZNF24 Hs.173911
�1.6 �1.4 NC Basic transcription element binding protein 1 BTEB1 Hs.150557
NC �1.4 NC Nuclear receptor subfamily 2, group F, member 2 NR2F2/COU P-TFII Hs.347991
�1.4 �1.4 NC Ubinuclein 1 UBN1 Hs.21479
�1.5 �1.5 NC Delta sleep including peptide, immunoreactor DSIPI Hs.420569
�1.4 �1.5 �1.5 Nuclear receptor subfamily 1, group D, member 2 NR1D2/EAR-1R Hs.37288
�1.2 �1.5 NC Serologically defined colon cancer antigen 33 SDCCAG33 Hs.284217
1.1 �1.5 �1.3 Transcription factor 7 (T-cell specific, HMG-box) TCF7 Hs.169294
�1.4 �1.6 �1.5 Nuclear receptor subfamily 3, group C, member 1 (glucocorticoid receptor) NR3C1 Hs.512414
�1.6 �1.6 �1.3 OLF-1/EBF associated zinc finger gene OAZ Hs.137168
NC NC 1.6 Zinc finger protein 323 ZNF323 Hs.444116
NC NC 1.5 Interferon regulatory factor 7 IRF7 Hs.166120
NC NC 1.4 v-jun sarcoma virus 17 oncogene homolog (avian) JUN Hs.78465
NC NC 1.4 Pre-B-cell leukemia transcription factor 2 PBX2 Hs.93728
NC NC 1.4 Transcription factor 3 (E2A immunoglobin enhancer binding factors E12/E47) TCF3 Hs.371282
NC NC �1.4 v-ets erythroblastosis virus E26 oncogene homolog 2 (avian) ETS2 Hs.292477
NC NC �1.5 Friend leukemia virus integration 1 FLI1 Hs.257049
NC NC �1.5 Metastasis associated 1 MAT1 Hs.101448
Metabolism
Xenobiotic metabolism phase I
NC 1.6 1.3 NAD(P)H dehydrogenase, quinone 1 NQO1 Hs.406515
NC 1.4 NC Aldehyde dehydrogenase 6 family, member A1 ALDH6A1 Hs.293970
�1.3 NC �1.5 Aldehyde dehydrogenase 1 family, member A1 ALDH1A1 Hs.76392
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239234

Table 2 (continued)
AA vs.
THF/EtOh
BC vs.
THF/EtOH
AA/BC vs.
THF/EtOH
Gene title Gene symbol UniGene ID
Metabolism
Xenobiotic metabolism phase II
Sulfotransferase
�1.7 �2.0 �2.1 Sulfotransferase family, cytosolic, 1B, member 1 SULT1B1 Hs.129742
Transferase alkyl or aryl
NC NC 1.6 Glutathione S-transferase M5 GSTM5 Hs.75652
Glucuronosyltransferase
NC 1.1 1.4 Exostoses (multiple) 2 EXT2 Hs.75334
NC �2.0 NC Exostoses (multiple) 1 EXT1 Hs.184161
Epoxide hydrolase
�1.5 �1.2 �1.4 Leukotriene A4 hydrolase LTA4H Hs.81118
Changes in relative gene expression were calculated versus control (medium with 0.075%THF/EtOH), only genes with the significant change in expression
level higher than 1,4 are shown. NC_no change.
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 235
toxic, physiological concentrations of BC were used in this
study [7,35]. At the same time, these concentrations of BC,
mimicking those occurring physiologically, promoted endo-
thelial cell chemotaxis in vitro and applied subcutaneously
in matrigel significantly induced the proangiogenic effect of
bFGF in the in vivo mouse model.
Angiogenesis is the crucial event for the remodeling of
tissues of a growing body in embryonic and adult life,
female ovulatory cycle, wound healing, tissue ischemia and
inflammatory processes as well as tumor malignancy [1].
The induction of angiogenesis involves the activation of
endothelial cells from preexisting capillaries or mobilization
of the endothelial progenitor cells [23]. The proangiogenic
cytokines, such as VEGF, bFGF, PDGF, TGF-h, angiopoie-tin 1/2 and several others, activate specific receptors on EC,
inducing endothelial cell detachment, migration, prolifer-
Fig. 7. The expression of selected genes verified by the quantitative real-time PCR:
X-C motif) receptor 4), IL-8 (interleukin 8), VCAM-1 (vascular cell adhesion
(mesenchyme homeo box 2 (growth arrest-specific homeo box)), EGR-1 (early gro
apoptosis inhibitor). Data expressed as the relative gene expression ratio. The me
from the corresponding control, *Pb0.05.
ation, expression of specific matrix proteins and proteolytic
enzymes, and other factors, which are responsible for the
remodeling of matrix and the promotion of the outgrowth of
new capillaries [1,3]. Activated EC change their profiles of
gene expression, switching to proliferative, non-differenti-
ated status, investigated already by microarrays [37,38].
Several pathways, including retinoid signaling, have been
implicated in the development of the cardiovascular system
in the fetal period. Nutritional deficiency of retinoids, RXR/
RAR, or Raldh2 knock-out mice, is characterized by
multiple developmental malformations, including severe
cardiovascular defects and lack of ophthalmo-mesenteric
vessels with disrupted formation of extra-extraembryonic
vessels [39,40]. However, cellular mechanisms, thereby
carotenoid/retinoid participates in the assembly of mamma-
lian blood vessels, have not been defined.
influence of beta-carotene and arachidonic acid on CXCR4 (chemokine (C-
molecule 1), MAD1L1 (MAD1 mitotic arrest deficient-like 1), MEOX2
wth response 1) and BIRC5 (baculoviral IAP repeat-containing 5 (survivin)
an valuesFS.E., n=3 done in triplicates are shown; *Significantly different

Fig. 8. Evidence of beta-carotene 15,15V-monooxygenase 1 (BCMO) gene
expression in HUVEC. Gene expression was investigated using quantitative
real-time PCR method. PCR products were analyzed also by electrophoresis
carried out according to standard protocol.
Fig. 9. Influence of studied compounds on IL-8 protein expression. Western
blot analysis of protein content in the HUVECs treated 24 h with EBM
medium with 0.075% THF/EtOH (control), BC 3 AM or AA 3 AM or both
compounds. Beta-actin was used as a the control of the equally loading of
proteins. Values are meanFS.E., n=6 done in triplicates. *Pb0.05. (A)
Representative Western blot of IL-8 protein expression. (B) Quantitative
evaluation (densitometric analysis) of analyzed proteins, expressed as
percent of control.
A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239236
Using Raldh2 �/� mice, Lai et al. documented that
retinoic acid induced the expression of p21 and p27, the
Cip/Waf family of Cdk inhibitors controlling cell cycle
progression in EC. Retinoid treatment in normal EC had no
effect on the expression of cyclins: A, B, D3, E, or Cdc2 or
Cdk2 [41]. Using Western blot, those authors found that the
formation of the complex between cyclin D1 or D2 and
Cdk4 was significantly reduced, whereas the formation of
the complex between RA-induced p21 or p27 and Cdk4 as
well as cyclin D1 and D2 was greatly enhanced in the
presence of retinoic acid. The reduced Cdk/cyclin D
complex formation resulted in lower levels of phosphory-
lated retinoblastoma protein (Rb) in RA-treated EC. Thus,
there was no influence on the number of cells (prolifer-
ation), but there was a significant decrease in the proportion
of endothelial cells in phase S and an increase in the
proportion of cells in phase G1 with no evidence of
apoptosis in EC [41]. We have not observed any measurable
effect of BC on EC proliferation or apoptosis. Results from
our microarray experiments indicated that BC and AA
weakly up-regulated the key molecules participating in cell
cycle regulation, such as MCM-5 as well as polo-like kinase
Plk1, or NUCKS controlling the G2/M check point.
However, it was paralleled by the up-regulation of the
important inhibitors of G2/M cell cycle phase, as Wee1 and
PKMyt1. The expression of MAD1L1, participating in G1/S
cell cycle check point and inhibition of proliferation [24],
was also observed.
Although the analysis of the microarray data revealed a
strong (6.5-fold) up-regulation of apoptosis facilitator
Bcl2L11, a member of the Bcl2 family by BC, we did
not observe any change in the rate of apoptosis in
HUVECs measured by the caspase activity. The up-
regulation of some proapoptotic members of the Bcl-2
family, such as Bcl2L11/BAM and anti-apoptotic MCL-1,
TEGT-BAX inhibitor was noted in the microarray experi-
ments in BC treated HUVECs. It has been shown that
Bcl2L11 releases mitochondrial cytochrome c, but the
simultaneous increase of the MCL1 expression may have
attenuated the proapoptotic potential of Bcl2L11 [42]. The
BC and AA also up-regulated survivin (BIRC5), an anti-
apoptotic protein, which binds to pro-caspase-9, preventing
its recruitment to Apaf1 [43]. The observed change in the
expression of survivin in microarray was later confirmed
by the results of real-time PCR.
Thus, the proangiogenic activity of BC in HUVECs
seems to be related not to proliferation (or inhibition of
apoptosis) [41], but to the potent activation of chemotaxis
and changes in the expression of genes mediating cell
adhesion and matrix assembly [44,45]. The results obtained
by the microarray in our model strongly support this
hypothesis.
The microarray data demonstrated changes in the
expression of several genes coding for group of proteins
participating in cell–cell and cell–matrix adhesion, matrix
proteins and proteases which regulate cell/matrix interac-
tion. The list of the BC-responsive genes includes those
coding for proteins participating in: cell–cell adhesion
(VCAM1, SELP, CD24), cadherins (CELSR1), and catenins
(CTNNA1L, CTNNB1). Expression of those genes was
mainly down-regulated by both BC and AA. The down-
regulation of VCAM-1, ICAM and selectin E by carote-
noids, including BC, in human aortic endothelium stimu-
lated with Il-1h was suggested to be responsible for the
modulatory effect on inflammatory response [46] and may
reflect the anti-inflammatory, protective effect of BC on
endothelium.

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 237
The expression of the genes coding for proteins
associated with cell-extracellular matrix adhesion, such as
integrins (ITGA6) and SCARB1, was up-regulated by BC.
Members of the extracellular matrix degrading enzymes
involved in the regulation of the matrix degradation
products promoting cell migration, such MMP14, and the
tissue plasminogen activator gene (PLAT), were similarly
up-regulated, while MMP12 was down-regulated. The
integrin-mediated stimulation of chemotaxis by the metal-
loproteinase-mediated matrix degradation products is an
important step of the capillary network formation [44,
45,47]. The expression of desintegrin ADAMTS1, respon-
sible for receptor shadding [28], was also inhibited.
The BC-treated cells also showed the down-regulation
of genes coding for various extracellular matrix compo-
nents, such as collagens, fibrillin 1, laminin h1, matrilin 2
and matrix Gla protein. The importance of the expression
of cell surface proteins, which participate in cell/matrix
(e.g. integrins) and cell/cell (e.g. cadherins, catenins,
endoglin, ephrins and their receptors) interactions required
for the regulation of proliferation, migration and differ-
entiation during angiogenesis, has been well documented
[3]. Additionally BC up-regulates the expression of IL-8,
the potent activator of migration of EC [30], what was
confirmed in this study by both real-time PCR and Western
blot.
Remodeling of the extracellular matrix proteins by
enzymatic degradation and synthesis of proteoglycans
changes the extracellular matrix composition and regu-
lates the migration of endothelial cells. Migration and
cellular shape change are also associated with the stress
fiber formation and reorganization of the cellular cytos-
keletal proteins [48,49]. The contribution of nitric oxide
in the chemotactic activity and cytoskeleleton reorganiza-
tion of endothelial cells was also reported [50]. The
induction of the endothelial nitric oxide synthase (eNOS)
gene by BC in HUVEC was also observed in our
microarray data.
The G-protein binding receptors and regulators of Rho
GTPases were the second largest group of genes, whose
expression was significantly changed by BC. The expres-
sion of some of them was confirmed by real-time PCR. The
genes up-regulated by BC, such as CXCR4 and IL-8 (whose
induction was verified by real-time PCR), and others, such
as GPR12, were recently demonstrated to be activated by
sphingosine 1-phosphate (S1P)-Edg class receptor [51],
DTR-heparan-binding epidermal growth factor (EGF)-like
receptor, and the integrin alpha 6 (ITGA6) are known
regulators of HUVEC migration that activate Rho GTPases
[52,53]. Proangiogenic factors, such as stromal derived
factor (SDF-1), IL-8, or S1P, lead to the activation of Rho/
Rac/CDC42 small GTPases through the activation of G-
protein-coupled receptors and by interaction with adhesion-
mediated signaling pathway [18,29,30,53]. Rho GTPases
regulate cytoskeletal changes responsible for cell motility,
shape and contraction. Rho GTPases promote actin–myosin
interaction and contraction of the cell through the regulation
of phosphorylation of the myosin light chains (MLCs).
MLC phosphorylation occurs in the presence of specific
MLC kinases, such as MLCK Ca+2 dependent kinase, Rho
kinase (ROCK) and p21-activated kinase (PAK). Rho
kinases also inhibit dephosphorylation by the inhibition of
myosin phosphatase type I (PP1M) and increase MLC
phosphorylation [54,55]. The obtained microarray results
demonstrated that BC, through the regulation of expression
of Rho/Rac/Cdc42 pathway members, lead to the activation
of HUVEC migration.
Despite the evidence of their proangiogenic activity, BC
(and AA) do not seem to influence the differentiation of
HUVECs as reflected by the microarrays results and in vitro
3D model of tubulogenesis. Lai et al. reported that RA
deficiency had no effect on the differentiation of immature
endothelial cell as evidenced by the expression of VEGF,
Flk1-receptor, VE-cadherin or angiopoietin Tie2 receptor
[41]. We also did not observe any changes in the expression
of genes associated with HUVEC differentiation following
incubation with BC or AA.
It has been reported that cytokines (bFGF, VEGF)
stimulate formation of tube-like structures and endothelial
cell maturation in an in vitro matrigel model, and that
vascular repair after injury is associated with the increase of
the expression of early growth response transcription factor
(EGR-1) [32]. As suggested by the results of our microarray
experiments, the expression of the bFGF receptor (FGFR1)
was down-regulated by BC. The microarray data revealed
that BC induced two potent repressors of EGR-1: DR-1 and
NAB-1 [32]. Additionally, the expression of the tran-
scription factor MEOX2, which has an inhibitory effect on
the VEGF-stimulated tube-formation of HUVECs [33], was
up-regulated by BC and AA in our model. Moreover, the
highest expression activation by BC (9.8-fold) was observed
in gene coding for endopeptidase MME. The membrane-
bound metallopeptidase, which cleaves and degrades
angiogenic peptides, such as atrial natriuretic peptide,
endothelin, angiotensin I, substance P, and bradykinin,
was also shown to inhibit HUVEC differentiation [45].
These findings suggests the activation of migration and
proliferation of non-differentiated EC and may imply the
formation of non-mature and non-functional capillary net-
work in response to BC. Such a network is characteristic of
tumor vasculature [1].
In summary, we postulate that the originally observed
proangiogenic activity of BC is related to the activation of
chemotaxis of endothelial cells. The effect of BC on EC
gene expression as assessed with microarrays is moderate.
However, we were able to detect gene expression changes
that support the involvement of BC in the regulation of
synthesis of extracellular matrix and adhesion molecules
synthesis, resulting in a potent activation of Rho/Rac/Cdc42
GTPase signaling pathway, which in turn may promote
HUVEC migration and the formation of not a completely
matured capillary network.

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239238
Acknowledgements
This work was supported by the F5 EU DLARFID
project QLK1-CT-2001-00183 The Author would like to
thank Phd R. Goralczyk, Phd K. Wertz and Phd G. Riss
(DSM Nutritional Products, Human Nutrition and Health,
Carotenoid Section, Basel, Switzerland) for the kind supply
of HPLC-grade BC as well as for the training in the HPLC
methodology of measuring the levels of carotenoids.
References
[1] P. Carmeliet, Angiogenesis in health and disease, Nat. Med. 9 (2003)
653–660.
[2] Z.A. Khan, S. Chakrabarti, Growth factors in proliferative diabetic
retinopathy, Exp. Diabesity Res. 4 (2003) 287–301.
[3] G. Bazzoni, E. Dejana, M.G. Lampugnani, Endothelial adhesion
molecules in the development of the vascular tree: the garden of
forking paths, Curr. Opin. Cell Biol. 11 (1999) 573–581.
[4] J. Gerhart, 1998, Warkany lecture: signaling pathways in develop-
ment, Teratology 60 (1999) 226–239.
[5] D. Simoni, M. Tolomeo, Retinoids, apoptosis and cancer, Curr.
Pharm. Des. 7 (2001) 1823–1837.
[6] B.A. Stoll, Linkage between retinoid and fatty acid receptors:
implications for breast cancer prevention, Eur. J. Cancer Prev. 11
(2002) 319–325.
[7] S.A. Ross, P.J. McCaffery, U.C. Drager, L.M. De Luca, Retinoids in
embryonal development, Physiol. Rev. 80 (2000) 1021–1054.
[8] W. Stahl, N. Ale-Agha, M.C. Polidori, Non-antioxidant properties of
carotenoids, Biol. Chem. 383 (2002) 553–558.
[9] J.A. Fontana, A.K. Rishi, Classical and novel retinoids: their targets in
cancer therapy, Leukemia 16 (2002) 463–472.
[10] E. Dragnev, K.H. Rigas, J.R. Dmitrovsky, The retinoids and cancer
prevention mechanisms, Oncologist 5 (2000) 361–368.
[11] J.J.M. Castenmiller, C.E. West, Bioavailability and bioconversion of
carotenoids, Annu. Rev. Nutr. 18 (1998) 19–38.
[12] D. Albanes, O.P. Heinonen, P.R. Taylor, J. Virtamo, B.K. Edwards, M.
Rautalahti, A.M. Hartman, J. Palmgren, L.S. Freedman, J. Haapa-
koski, M.J. Barrett, P. Pietinen, N. Malila, E. Tala, K. Liippo, E.R.
Salomaa, J.A. Tangrea, L. Teppo, F.B. Askin, E. Taskinen, Y. Erozan,
P. Greenwald, J.K. Huttunen, Alpha-tocopherol and beta-carotene
supplements and lung cancer incidence in the alpha-tocopherol, beta-
carotene cancer prevention study: effects of baseline characteristics
and study compliance, J. Natl. Cancer Inst. 88 (1996) 1560–1570.
[13] G.S. Omenn, G. Goodman, M. Thornquist, J. Grizzle, L. Rosenstock,
S. Barnhart, J. Balmes, M.G. Cherniack, M.R. Cullen, A. Glass, The
beta-carotene and retinol efficacy trial (CARET) for chemoprevention
of lung cancer in high risk populations: smokers and asbestos-exposed
workers, Cancer Res. 54 (1994) 2038s–2043s.
[14] N.R. Cook, I.M. Le, J.E. Manson, J.E. Buring, C.H. Hennekens,
Effects of beta-carotene supplementation on cancer incidence by
baseline characteristics in the physicians’ health study (United States),
Cancer Causes Control 11 (2000) 617–626.
[15] D.S. Goodman, R. Blomstrand, B. Werner, H.S. Huang, T. Shiratori,
The intestinal absorption and metabolism of vitamin A and beta-
carotene in man, J. Clin. Invest. 45 (1966) 1615–1623.
[16] E.A. Jaffe, R.L. Nachman, C.G. Becker, C.R. Minick, Culture of
human endothelial cells derived from umbilical veins. Identification
by morphologic and immunologic criteria, J. Clin. Invest. 52 (1973)
2745–2756.
[17] G. Plickert, M. Kroiher, Proliferation kinetics and cell lineages can be
studied in whole mounts and macerates by means of BrdU/anti-BrdU
technique, Development 103 (1988) 791–794.
[18] F. Wang, J.R. Van Brocklyn, J.P. Hobson, S. Movafagh, Z.
Zukowska-Grojec, S. Milstien, S. Spiegel, Sphingosine 1-phosphate
stimulates cell migration through a G(i)-coupled cell surface receptor.
Potential involvement in angiogenesis, J. Biol. Chem. 274 (1999)
35343–35350.
[19] B. Vailhe, D. Vittet, J.J. Feige, In vitro models of vasculogenesis and
angiogenesis, Lab. Invest. 81 (2001) 439–452.
[20] M. Bradford, A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding, Anal. Biochem. 72 (1976) 248–254.
[21] P. Chomczynski, N. Sacchi, Single-step method of RNA isolation by
acid guanidinium thiocyanate-thiocyanatephenol-chloroform extrac-
tion, Anal. Biochem. 162 (1987) 156–159.
[22] M.W. Pfaffl, A new mathematical model for relative quantification in
real-time RT-PCR, Nucleic Acids Res. 29 (2001) 9–45.
[23] T. Asahara, H. Masuda, T. Takahashi, C. Kalka, C. Pastore, M. Silver,
M. Kearne, M. Magner, J.M. Isner, Bone marrow origin of endothelial
progenitor cells responsible for postnatal vasculogenesis in physio-
logical and pathological neovascularization, Circ. Res. 85 (1999)
221–228.
[24] R.N. Eisenman, Deconstructing myc, Genes Dev. 15 (2001)
2023–2030.
[25] D.R. Schultz, W.J. Harrington Jr., Apoptosis: programmed cell death
at a molecular level, Semin. Arthritis Rheum. 32 (2003) 345–369.
[26] B.R. Alevriadou, CAMs and Rho small GTPases: gatekeepers for
leukocyte transendothelial migration. Focus on bVCAM-1-medi-
ated Rac signaling controls endothelial cell–cell contacts and
leukocyte transmigrationQ, Am. J. Physiol., Cell Physiol. 285
(2003) C250–C252.
[27] M. Anghelina, A. Schmeisser, P. Krishnan, L. Moldovan, R.H.
Strasser, N.I. Moldovan, Migration of monocytes/macrophages in
vitro and in vivo is accompanied by MMP12-dependent tunnel
formation and by neovascularization, Cold Spring Harbor Symp.
Quant. Biol. 67 (2002) 209–215.
[28] B.L. Tang, ADAMTS: a novel family of extracellular matrix
proteases, Int. J. Biochem. Cell Biol. 33 (2001) 33–44.
[29] R. Henschler, A. Piiper, R. Bistrian, D. Mobest, SDF-1alpha-induced
intracellular calcium transient involves Rho GTPase signalling and is
required for migration of hematopoietic progenitor cells, Biochem.
Biophys. Res. Commun. 311 (2003) 1067–1071.
[30] I.U. Schraufstatter, K. Trieu, M. Zhao, D.M. Rose, R.A. Terkeltaub,
M. Burger, IL-8-mediated cell migration in endothelial cells depends
on cathepsin B activity and transactivation of the epidermal growth
factor receptor, J. Immunol. 171 (2003) 6714–6722.
[31] T.S. Lewis, P.S. Shapiro, N.G. Ahn, Signal transduction through MAP
kinase cascades, Adv. Cancer Res. 74 (1998) 49–139.
[32] R.G. Fahmy, C.R. Dass, L.Q. Sun, C.N. Chesterman, L.M.
Khachigian, Transcription factor Egr-1 supports FGF-dependent
angiogenesis during neovascularization and tumor growth, Nat.
Med. 9 (2003) 1026–1032.
[33] D.H. Gorski, A.J. Leal, Inhibition of endothelial cell activation by the
homeobox gene Gax, J. Surg. Res. 111 (2003) 91–99.
[34] P.D. Fraser, P.M. Bramley, The biosynthesis and nutritional uses of
carotenoids, Prog. Lipid Res. 43 (2004) 228–265.
[35] F.J. Schweigert, Metabolism of carotenoids in mammals, in: G.
Britton, S. Liaaen-Jensen, H. Pfandern (Eds.), Carotenoids, Biosyn-
thesis, vol. 3, Birkh7user Verlag, Basel, 1998, pp. 249–284.[36] D.V. Crabtree, A.J. Adler, Is beta-carotene an antioxidant? Med.
Hypotheses 48 (1997) 183–187.
[37] S.C. Bell, A. Avila, R. Salazar, K.J. Bayless, S. Kanagala, S.A.
Maxwell, G.E. Davis, Differential gene expression during capillary
morphogenesis in 3D collagen matrices: regulated expression of genes
involved in basement membrane matrix assembly, cell cycle
progression, cellular differentiation and G-protein signaling, J. Cell.
Sci. 114 (2001) 2755–2773.
[38] J. Kahn, F. Mahraban, G. Ingle, X. Xin, J.E. Bryant, G. Vehar, J.
Schoenfeld, C.J. Grimaldi, F. Peale, A. Draksharapu, D.A. Lewin,

A. Dembinska-Kiec et al. / Biochimica et Biophysica Acta 1740 (2005) 222–239 239
M.E. Gerritsen, Gene expression profiling in in vitro model of
angiogenesis, Am. J. Pathol. 156 (2000) 1887–1900.
[39] P. Kastner, N. Messaddeq, M. Mark, O. Wendiing, J.M. Grondona, S.
Ward, N. Ghyselinck, P. Chambon, Vitamin A deficiency and
mutations of RXR alpha, RXR beta and RAR alpha lead to the early
differentiation of embryonic ventricular cardiomyocytes, Develop-
ment 124 (1997) 4749–4758.
[40] M. Zile, Function of Vit A in vertebrate development, J. Nutr. 131
(2001) 705–708.
[41] L. Lai, B.L. Bohnsack, K. Niederreither, K.K. Hirschi, Retinoic
acid regulates endothelial cell proliferation during vasculogenesis,
Development 130 (2004) 6465–6474.
[42] R.D. Erwert, K.T. Eiting, J.C. Tupper, R.K. Winn, J.M. Harlan, D.D.
Bannerman, Shiga toxin induces decreased expression of the anti-
apoptotic protein Mcl-1 concomitant with the onset of endothelial
apoptosis, Microb. Pathog. 35 (2003) 87–93.
[43] B.B. Aggarwal, U. Bhardwaj, Y. Takada, Regulation of TRAIL-
induced apoptosis by ectopic expression of antiapoptotic factors,
Vitam. Horm. 67 (2004) 453–483.
[44] A. Collen, R. Hanemaaijer, F. Lupu, P.H. Quax, N. van Lent, J.
Grimbergen, E. Peters, P. Koolwijk, V.W. van Hinsbergh, Membrane-
type matrix metalloproteinase-mediated angiogenesis in a fibrin-
collagen matrix, Blood 101 (2003) 1810–1817.
[45] L.A. Cornelius, L.C. Nehring, E. Harding, M. Bolanowski, H.G.
Welgus, D.K. Kobayashi, R.A. Pierce, S.D. Shapiro, Matrix metal-
loproteinases generate angiostatin: effects on neovascularization, J.
Immunol. 161 (1998) 6845–6852.
[46] K.R. Martin, D. Wu, M. Meydani, The effects of carotenoids on the
expression of cell surface adhesion molecules and binding of
monocytes to human aortic endothelial cells, Atherosclerosis (2000)
265–274.
[47] J. Sottile, Regulation of angiogenesis by extracellular matrix,
Biochim. Biophys. Acta 1654 (2004) 13–22.
[48] C. Menager, J. Vassy, C. Doliger, Y. Legrand, A. Karniguian,
Subcellular localization of RhoA and ezrin at membrane ruffles of
human endothelial cells: differential role of collagen and fibronectin,
Exp. Cell Res. 249 (1999) 221–230.
[49] E.S. Wijelath, S. Rahman, J. Murray, Y. Patel, G. Savidge, M.
Sobel, Fibronectin promotes VEGF-VEGFinduced CD34 cell
differentiation into endothelial cells, J. Vasc. Surg. 39 (2004)
655–660.
[50] K. Kawasaki, R.S. Smith Jr., C.M. Hsieh, J. Sun, J. Chao, J.K. Liao,
Activation of the phosphatidylinositol 3-kinase/protein kinase Akt
pathway mediates nitric oxide-induced endothelial cell migration and
angiogenesis, Mol. Cell. Biol. 23 (2003) 5726–5737.
[51] Y. Takuva, Subtype-specific differential regulation of Rho family G
proteins and cell migration by the Edg family sphingosine-1-
phosphate receptors, Biochim. Biophys. Acta 1582 (2002) 112–120.
[52] A. Gohla, R. Harhammer, G. Schultz, The G-protein G13 but not
G12 mediates signaling from lysophosphatidic acid receptor via
epidermal growth factor receptor to Rho, J. Biol. Chem. 273 (1998)
4653–4659.
[53] J.W. Lee, R. Juliano, Mitogenic signal transduction by integrin- and
growth factor receptor-mediated pathways, Mol. Cells 17 (2004)
188–202.
[54] M.V. Hoang, M.C. Whelan, D.R. Senger, Rho activity critically and
selectively regulates endothelial cell organization during angiogenesis,
Proc. Natl. Acad. Sci. U. S. A. 101 (2004) 1874–1879.
[55] G.P. van Nieuw Amerongen, V.W. van Hinsbergh, Cytoskeletal
effects of rho-like small guanine nucleotide-binding proteins in
the vascular system, Arterioscler. Thromb. Vasc. Biol. 21 (2001)
300–311.
Recommended

















