
Molecular Microbiology (2005)
57
(5), 1210–1223 doi:10.1111/j.1365-2958.2005.04743.x
© 2005 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2005
? 2005
57
512101223
Original Article
Rhamnolipid-mediated biofilm detachmentB. R. Boles, M. Thoendel and P. K. Singh
Accepted 23 May, 2005. *For correspondence. E-mail [email protected]; Tel. (
+
1) 319 335 7608; Fax (
+
1) 319 335 7623.
Rhamnolipids mediate detachment of
Pseudomonas aeruginosa
from biofilms
Blaise R. Boles,
1
Matthew Thoendel
1,2
and Pradeep K. Singh
1,2
*
Departments of
1
Microbiology and
2
Internal Medicine, University of Iowa Roy J. and Lucille A. Carver College of Medicine, Iowa City, IA 52242, USA.
Summary
The process of detachment, through which bacteriause active mechanisms to leave biofilms and returnto the planktonic (free-living) state, is perhaps theleast understood aspect of the biofilm life cycle. Likeother stages of biofilm development, detachment is adynamic, regulated process, controlled by specificgenes, and induced by particular environmental cues.In previous work we discovered
Pseudomonasaeruginosa
variants that exhibit accelerated biofilmdetachment. These hyper-detaching variants arisespontaneously from biofilms at a high frequency, andthey exhibit robust detachment under different biofilmgrowth conditions. Here we show that these variantsdetach by a mechanism requiring the biosurfactantrhamnolipid and that this detachment mechanismrapidly restores antibiotic sensitivity to separatingbacteria. We also show that rhamnolipids can bringabout detachment in wild-type
P. aeruginosa
biofilms.These findings raise the possibility that this detach-ment mechanism may be useful as a treatment todisrupt established biofilms. Interestingly, the rham-nolipid-mediated detachment mechanism involvesthe formation of cavities within the centre of biofilmstructures. Our data suggest a model to explaindetachment that occurs via this pattern.
Introduction
Many bacterial species are capable of two general growthmodes. In planktonic growth, organisms exist as indepen-dent, free-living cells, usually suspended in a liquidmedium. Bacteria can also form biofilms in which the cellslive clustered together in matrix-encased groups (Coster-ton
et al
., 1999; Hoiby
et al
., 2001). Biofilms cause someof the most recalcitrant human infections (Costerton
et al
.,
1999; Parsek and Singh, 2003) and are the predominantgrowth mode for bacteria in the environment.
Biofilm growth provides important advantages to organ-isms. Biofilm bacteria manifest marked resistance tokilling by antimicrobial agents (Costerton
et al
., 1999;Stewart, 2002), increased synthesis of protective matrixmaterials (Costerton
et al
., 1999; Friedman and Kolter,2004) and in some cases, enhanced metabolic coopera-tion (Shapiro, 1998). Recent work also suggests that hor-izontal gene transfer and intercellular communication maybe facilitated in biofilms (Hausner and Wuertz, 1999;Parsek and Greenberg, 2000), and that biofilm growthincreases the genetic diversity of bacterial populations(Boles
et al
., 2004). All of these characteristics couldenhance the survival of bacterial communities in harshconditions.
While the benefits provided by biofilm growth areimpressive, these advantages come at some cost. Forexample, matrix production imposes a synthetic burdenon the bacteria and the nutrient gradients within biofilmscan limit growth and the cells’ ability to sustain defences,such as the production of antioxidants, and metabolicenzymes (Huang
et al
., 1998; Hassett
et al
., 1999). Per-haps the most significant risk for biofilm bacteria occurswhen local conditions deteriorate. This could occurbecause of nutrient depletion, the accumulation of wastes,the appearance of immune cells or antibiotics, or otherthreats. Biofilm bacteria have a reduced ability to evadestresses because they are physically confined by thematrix and their motility functions are repressed (Whiteley
et al
., 2001; Sauer
et al
., 2002).The costs associated with biofilm growth make it vital
that bacteria possess mechanisms to separate from bio-films and assume planktonic life. This process is referredto as detachment (some investigators use the term dis-persion). Unlike sloughing in which shear forces physicallydislodge bacteria (Picioreanu
et al
., 2001; Stoodley
et al
.,2002), in detachment, environmental conditions triggeractive mechanisms that bring about bacterial separation(Hunt
et al
., 2004; Sauer
et al
., 2004; Thormann
et al
.,2005). The fact that detachment can be triggered by sev-eral different cues could allow organisms to regulate theirmovement between the biofilm and planktonic growthstates as environmental conditions change.
In addition to its importance for biofilms in the environ-ment, detachment also plays an important role in infection

Rhamnolipid-mediated biofilm detachment
1211
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1210–1223
pathogenesis. For instance, organisms like
Vibrio chol-erae, Legionella pneumophila
and
Pseudomonas aerugi-nosa
are thought to live in soil or aquatic biofilms (Parsekand Singh, 2003). The process of detachment createsmobile bacteria (single cells or aggregates) that are capa-ble of causing infection. Detachment may also promotedissemination from an initial infection point to other sitesin the body. A well-characterized example is the devastat-ing embolic events of endocarditis caused by detachmentof the complex biofilm growing on heart valves (Parsekand Singh, 2003). In addition, many cases of hospital-acquired pneumonia are caused by bacteria detachedfrom biofilms that form in a patient’s endotracheal tube ororopharnyx (Bergmans
et al
., 1998; Adair
et al
., 1999;Feldman
et al
., 1999).The process of detachment is complex. Even within a
single bacterial species, multiple detachment patternshave been observed, suggesting that different detach-ment mechanisms may exist. In the pathogen
P. aerugi-nosa
, detachment can involve the discharge of individualbacteria (Stoodley
et al
., 2001), the piecemeal separationof cell clusters (Stoodley
et al
., 2001) and the massdetachment of whole colonies seen early in biofilm devel-opment (P. Singh and B. Boles, unpubl. obs.). Perhaps themost interesting pattern, often observed in aged biofilms(
~
10–12 days of growth in our system), involves the for-mation of internal cavities that eventually fracture torelease motile bacteria. This pattern has been termed‘central hollowing’ (Sauer
et al
., 2002; Hunt
et al
., 2004).Each of these detachment patterns may involve multiplesteps including degradation of the matrix, activation ofmotility and physiological changes that prepare cells forconditions outside the biofilm.
In addition to this mechanistic complexity, the timing andextent of biofilm detachment is likely to be under tight andlocal regulation. Under favourable conditions, most wild-type biofilms release a relatively small number of cellscontinually; they also sporadically undergo major detach-ment events after prolonged periods of growth. Thesemajor detachment events can be very heterogeneous interms of timing, the regions of the biofilm affected andperhaps, the mechanism of detachment employed. Thisheterogeneity poses challenges to the investigation ofdetachment mechanisms. One approach that has beenused to overcome this challenge involves exposing bio-films to major, sudden changes in environmental condi-tions in order to induce detachment. This approach hasled to greater understanding of the process by whichdetachment is triggered (Hunt
et al
., 2004; Sauer
et al
.,2004; Thormann
et al
., 2005).Here we took a different approach. In a previous study,
we found that variants with a hyper-detaching phenotypespontaneously arise at a high frequency after short-termgrowth of wild-type
P. aeruginosa
in biofilms (Boles
et al
.,
2004). The variant we chose to study (hereinafter called‘variant’) showed a normal pattern of initial biofilm forma-tion; however, after 2 days of growth, the biofilm rapidlydisperses (see Fig. 1 and Boles
et al
., 2004).We decided to study the variant’s detachment mecha-
nism because its detachment phenotype is robust; mostof the biofilm biomass simultaneously disperses after only2 days of growth and it detaches under different biofilmgrowth conditions (Boles
et al
., 2004). Furthermore,detachment of variant biofilms is spontaneous. This elim-inates the need for an exogenous disturbance to inducedetachment that could produce pleiotropic physiologicaleffects. It also allowed us to focus on an effector mecha-nism that brings about bacterial separation, rather thanthe triggering mechanisms that activate the detachmentprocess. Here we show that the variant detaches via thecentral hollowing pattern, and this detachment mecha-nism rapidly restores aspects of planktonic physiology todetaching cells. Our investigation of the variant’s detach-ment mechanism revealed that the bio-surfactant rhamno-lipid is required for the hyper-detachment phenotype.Importantly, using three independent methods, we foundthat rhamnolipids also produce detachment (via centralhollowing) in wild-type
P. aeruginosa
biofilms.
Methods
Strains, plasmids and growth conditions
The
P. aeruginosa
strains used in this work were derivedfrom the wild-type strain PA01 (obtained from B. Iglewski).The isolation of the hyper-detaching variants, which spon-taneously arise from wild-type PA01 biofilms, is describedin the study by Boles
et al
. (2004). For visualization byconfocal microscopy, pMRP9-1 (which expresses greenfluorescent protein) was transformed into appropriatestrains (Davies
et al
., 1998). For the variant–wild typecoculture experiments a red fluorescent protein (
rfp
)expressing plasmid (pMTRFP) was constructing by liga-tion of polymerase chain reaction (PCR)-amplified
rfp
from PRSETB (Campbell
et al
., 2002) into pUCP18. Inthis vector
rfp
expression is controlled by the
lac
promoterand has a
T7gene10
ribosome binding site. In someexperiments mucoid and non-mucoid
P. aeruginosa
iso-lates from cystic fibrosis patients (from the University ofIowa Cystic Fibrosis Clinic) were used.
A
rhlAB
insertion/deletion mutation was generated byinserting a gentamicin cassette into the
P. aeruginosarhlAB
genes digested with DraIII and MluI. The
rhlAB
mutation was moved on to the chromosome of PA01 andthe variant by homologous recombination. For comple-mentation studies,
rhlAB
including the
rhlA
promoter wasPCR-amplified from PA01 and cloned into pEX1.8 (Pear-son
et al
., 1997). A construct for the inducible expression

1212
B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1210–1223
of rhamnolipids was made by cloning
rhlAB
without itsnative promoter into pSW195 (obtained from D. Wozniak),which contains mini-CTX1 (Hoang
et al
., 2000; Boles
et al
., 2004) with an arabinose inducible promoter pBAD(Newman and Fuqua, 1999). This was then moved ontothe chromosome at the phage attachment site
attB
.Expression was induced with 0.5%
L
-arabinose. Trypti-case Soy Broth (Difco) was used as the growth mediumunless otherwise specified. Swarming motility plates con-sisted of 8 g l
-
1
nutrient broth, 5 g l
-
1
dextrose and 0.5%Bacto Agar (Difco). For orcinol assays, bacteria weregrown in a medium composed of 6 g l
-
1
Na
2
HPO
4
, 3 g l
-
1
KH
2
PO
4
, 0.5 g l
-
1
NaCl, 0.1 mM CaCl
2
, 2 mM MgSO
4
,0.2% glucose and 0.05% glutamate. Antibiotic concentra-tions (per millilitre) were: 300
m
g carbenicillin, 100
m
g gen-tamycin, 60
m
g tetracycline for
P. aeruginosa.
Biofilm experiments
Flow cell experiments and confocal microscopy were per-formed as previously described (Davies
et al
., 1998). Bio-films were grown in continuous culture flow cells similarto those described in the study by Davies
et al
. (1998).The size of the flow channel was 5
¥
35
¥
1 mm. An over-night culture diluted to 10
7
cells per millilitre in fresh biofilmmedium was used as the inoculum and flow was arrestedfor 45 min. Flow of biofilm medium was then initiated at a
rate of 170
m
l min
-
1
. Images were obtained using a Bio-Rad (Hercules, CA) scanning confocal microscope. Bio-films were grown in 1% TSB unless otherwise specified.To measure the proportion of each strain in the wild typeand variant coculture biofilms, either the wild type or thevariant was marked with tetracycline resistance by inser-tion of mini-CTX1 at the strains chromosomal phageattachment site. Just before detachment, the biofilm wasscraped from the growth surface, homogenized and platedon LB agar with and without tetracycline in order to enu-merate each strain in the biofilm. In some flow cell studies,3-day-old wild-type biofilms were exposed to 0.5 mg ml
-
1
of purified rhamnolipid (obtained from the Jeneil Biosur-factant, Saukville, WI) or 0.2% SDS (Sigma) by replacingthe biofilm medium with medium containing either of thesesurfactants. Viability of planktonic
P. aeruginosa
in thepresence of purified rhamnolipid was assayed by incubat-ing 10
6
stationary-phase cells in a range of rhamnolipidconcentrations in 1 ml of PBS for 6 h.
For antibiotic susceptibility testing of detached cells andintact and dispersed biofilms, biofilms were grown usingthe rotating disk biofilm reactor as previously described(Boles
et al
., 2004). To determine the antibiotic sensitivityof detached bacteria, the discs (with attached biofilm)were washed in PBS (to remove growth media), and incu-bated in 1 ml of PBS for 4 h at 25
∞
C. The overlying fluid(containing detached cells) was then moved to a new
Fig. 1.
Schematic of biofilm detachment by wild-type
P. aeruginosa
and the hyper-detaching variant (also see Boles
et al
., 2004). Some biofilm structures formed by the wild type detach via the central hollowing pattern after 10–12 days of growth (top). The variant exhibits a similar detachment pattern; however this strain detaches much more extensively and detachment occurs after only 2 days of biofilm development (bottom).
Day 2 Day 3 Day 4
Wild type
Variant
Day ~10-12
Day 2 Day 3 Day ~10-12Day 0
Day 0

Rhamnolipid-mediated biofilm detachment
1213
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1210–1223
container and incubated with tobramycin at indicated con-centrations for 4 h. The cell suspension was then homog-enized, and cell counts were determined by platecounting. To determine the antibiotic sensitivity of intactbiofilms, discs (with attached biofilm) were washed withPBS and immediately treated with tobramycin for 4 h. Thebiofilm was then homogenized to produce a cell suspen-sion, and cell counts were determined by plate counting.To determine the antibiotic sensitivity of mechanically dis-rupted biofilms, discs with attached biofilms were placedin 1 ml of PBS and homogenized for 30 s to disrupt thebiofilm (prior to antibiotic treatment). The tobramycin sen-sitivity of these dispersed bacteria was then determinedas described above.The rotating disk biofilm reactor (Boles
et al
., 2004) wasalso used to generate biofilms for the static detachmentassay. In this assay, discs (with attached biofilm) wereremoved from the reactor, washed in PBS and incubatedin 1 ml of PBS for 4 h. In some experiments 0.5 mg ml
-
1
of purified rhamnolipid was added to PBS during the 4 hdetachment period. The bacteria in the overlying fluid(detached cells) and bacteria that remained on the disk(biofilm cells) were counted as described above. Thedetachment fraction was calculated by dividing thenumber of detached cells by the total number ofcells (detached
+
biofilm cells). Detachment of the
P. aeruginosa
strain inducibly expressing rhamnolipidswas analysed by collecting 1 ml aliquots of the biofilmeffluent on ice (in triplicate) from flow cell biofilm reactorsat the indicated times and enumerating cell numbers ineach aliquot by plate counting.
Rhamnolipid assays
Several methods were used to assay rhamnolipid produc-tion. For rhamnolipid detection on plates, methylene blue-containing plates were prepared as previously describedin the study by Kohler
et al
. (2000). The orcinol assay(Koch
et al
., 1991) of ether-extracted culture supernatantswas used to measure rhamnolipids in liquid cultures. Cul-tures were grown for 48 h at 37
∞
C, and 0.5 ml of culturesupernatant was extracted twice with 1 ml of diethyl ether.The ether fractions were pooled and evaporated to dry-ness, and reconstituted in 0.5 ml H2O. Samples werediluted 1/10 in a solution containing 0.19% orcinol in 53%H2SO4. The sample was then placed in boiling water for30 min, cooled at room temperature for 15 min, and theabsorbance (A421) was measured. Rhamnolipid concen-trations were calculated by comparing the data with astandard curve generated from purified rhamnolipid.Rhamnolipid gene expression was measured using arhlA::gfp fusion construct by Y. Lequette and E. P. Green-berg (Lequette and Greenberg, 2005). To measurerhlA::gfp expression in liquid cultures, fluorescence was
measured with a microtitre plate fluormeter (Tecan). Theexcitation wavelength was 435 nm and the emissionlength was 535 nm. The relative level of rhlA::gfp expres-sion in nascent biofilms was determined using confocalmicroscopy. Images of 12 different 2-day-old variant andwild-type biofilms of similar size were acquired using iden-tical confocal microscope settings. A line profile of fluores-cence pixel intensity across each biofilm colony wasgenerated using Laser Sharp software (Bio-Rad, Her-cules, CA). These values were normalized for colonydimensions and averaged.
Results
Detachment rapidly produces physiologicalchanges in separating bacteria
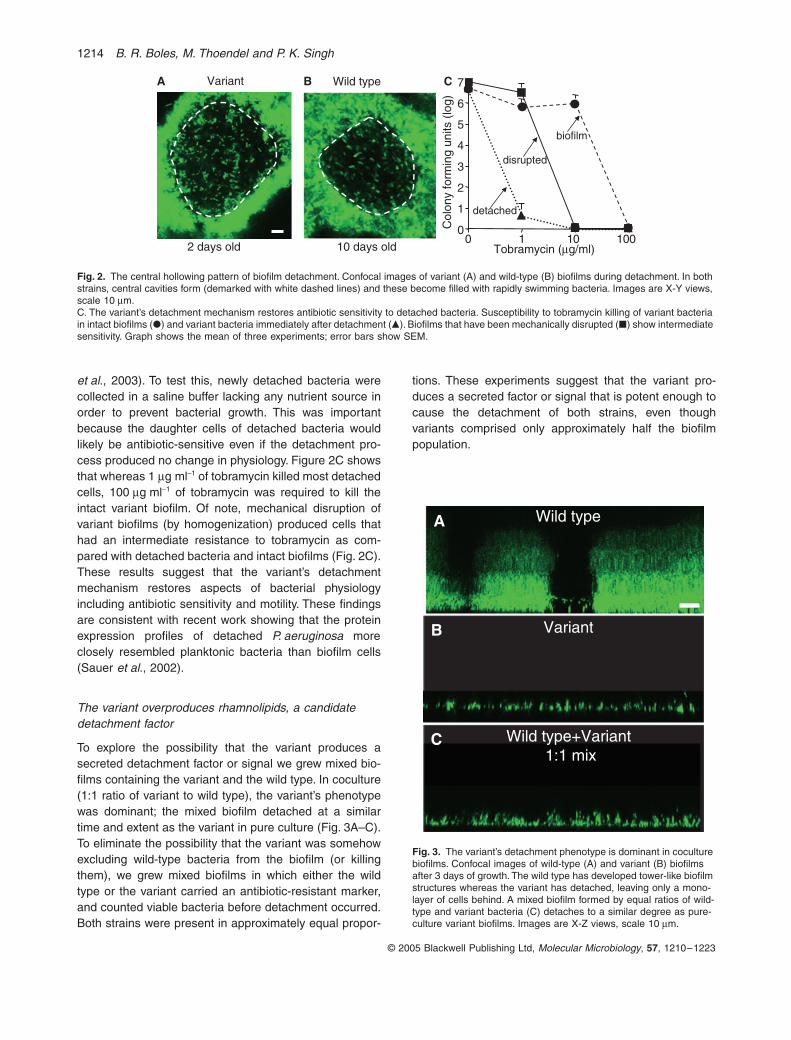
To begin our investigation of the variant’s detachmentmechanism, we performed time-lapse microscopy duringthe process of detachment (see Movie S1 in Supplemen-tary material). These images revealed that the variantreproducibly detached via a characteristic sequence ofevents. First, the biofilm began to slacken; individual bac-teria were seen moving to and fro within the previouslysolid structure. Soon after this, cavities formed in thecentre of biofilms and these became filled with rapidlyswimming bacteria (Fig. 2A). The cavities increased insize over time, and they eventually ruptured to releasemotile bacteria. Finally, the remaining biofilm biomassdetached from the growth surface leaving only a mono-layer of cells behind. In variant biofilms, the entire detach-ment process was completed in a short period of time (2–3 h), and it occurred nearly simultaneously in most all ofthe biofilm structures present within the reactor.
This pattern of detachment is very similar to the centralhollowing detachment pattern observed in aged wild-typebiofilms by several other groups and us (Sauer et al.,2002; Hunt et al., 2004) (Fig. 2B; see Movie S2 in Sup-plementary material). In the wild type, however, this typeof detachment occurs after much longer periods of growth(~10–12 days in our growth conditions versus 2 days forthe variant), and it affects only some areas of the biofilmat any given time. Nevertheless, the close resemblance ofthese detachment patterns suggests that a commonmechanism may be involved in both variant and wild-typebiofilms.
The presence of motile bacteria within the central cav-ities indicated that the variant’s detachment mechanismproduces some aspects of planktonic physiology in theseparating bacteria; previous work has shown that swim-ming motility is generally repressed during biofilm growth(Whiteley et al., 2001; Sauer et al., 2002). This led us toinvestigate whether detachment reversed another key bio-film phenotype, antibiotic resistance (Stewart, 2002; Mah
1214 B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
et al., 2003). To test this, newly detached bacteria werecollected in a saline buffer lacking any nutrient source inorder to prevent bacterial growth. This was importantbecause the daughter cells of detached bacteria wouldlikely be antibiotic-sensitive even if the detachment pro-cess produced no change in physiology. Figure 2C showsthat whereas 1 mg ml-1 of tobramycin killed most detachedcells, 100 mg ml-1 of tobramycin was required to kill theintact variant biofilm. Of note, mechanical disruption ofvariant biofilms (by homogenization) produced cells thathad an intermediate resistance to tobramycin as com-pared with detached bacteria and intact biofilms (Fig. 2C).These results suggest that the variant’s detachmentmechanism restores aspects of bacterial physiologyincluding antibiotic sensitivity and motility. These findingsare consistent with recent work showing that the proteinexpression profiles of detached P. aeruginosa moreclosely resembled planktonic bacteria than biofilm cells(Sauer et al., 2002).
The variant overproduces rhamnolipids, a candidate detachment factor
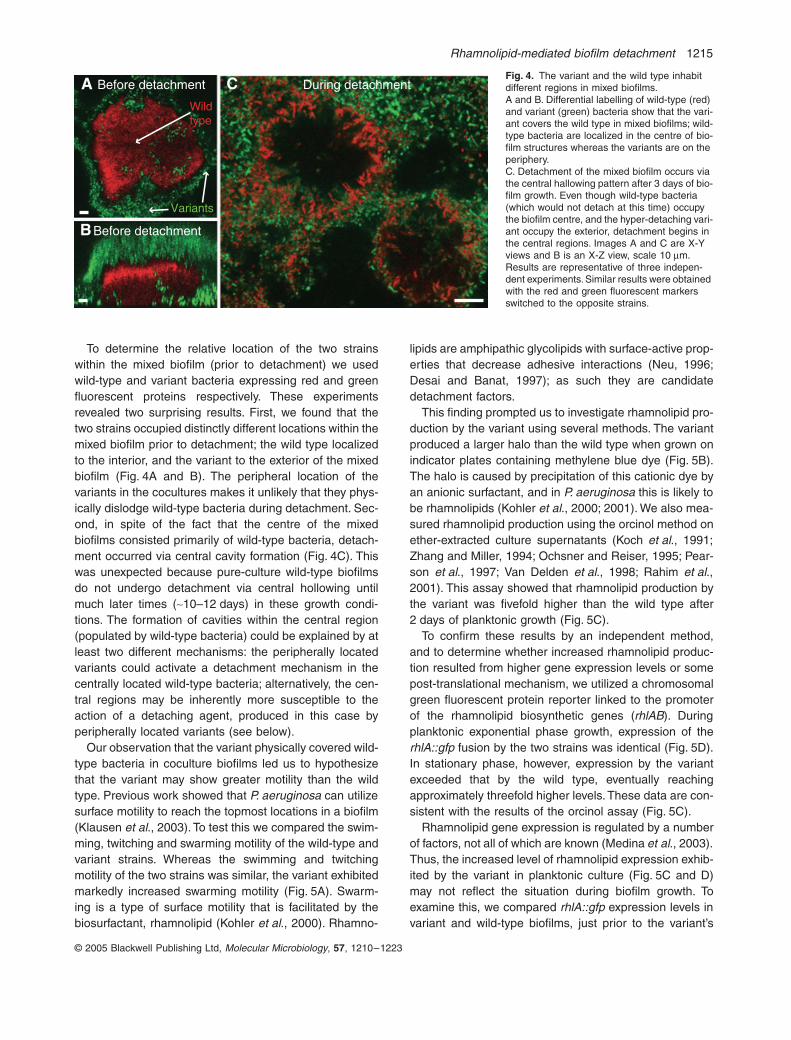
To explore the possibility that the variant produces asecreted detachment factor or signal we grew mixed bio-films containing the variant and the wild type. In coculture(1:1 ratio of variant to wild type), the variant’s phenotypewas dominant; the mixed biofilm detached at a similartime and extent as the variant in pure culture (Fig. 3A–C).To eliminate the possibility that the variant was somehowexcluding wild-type bacteria from the biofilm (or killingthem), we grew mixed biofilms in which either the wildtype or the variant carried an antibiotic-resistant marker,and counted viable bacteria before detachment occurred.Both strains were present in approximately equal propor-
tions. These experiments suggest that the variant pro-duces a secreted factor or signal that is potent enough tocause the detachment of both strains, even thoughvariants comprised only approximately half the biofilmpopulation.
Fig. 2. The central hollowing pattern of biofilm detachment. Confocal images of variant (A) and wild-type (B) biofilms during detachment. In both strains, central cavities form (demarked with white dashed lines) and these become filled with rapidly swimming bacteria. Images are X-Y views, scale 10 mm.C. The variant’s detachment mechanism restores antibiotic sensitivity to detached bacteria. Susceptibility to tobramycin killing of variant bacteria in intact biofilms (�) and variant bacteria immediately after detachment (�). Biofilms that have been mechanically disrupted (�) show intermediate sensitivity. Graph shows the mean of three experiments; error bars show SEM.
Tobramycin (mg/ml)10 days old
Wild type
2 days old0 1 10
0
1
2
3
4
5
6
7
biofilm
disrupted
Col
ony
form
ing
units
(lo
g)
C
detached
VariantA B
100
Fig. 3. The variant’s detachment phenotype is dominant in coculture biofilms. Confocal images of wild-type (A) and variant (B) biofilms after 3 days of growth. The wild type has developed tower-like biofilm structures whereas the variant has detached, leaving only a mono-layer of cells behind. A mixed biofilm formed by equal ratios of wild-type and variant bacteria (C) detaches to a similar degree as pure-culture variant biofilms. Images are X-Z views, scale 10 mm.
Wild typeA
VariantB
Wild type+Variant 1:1 mix
C
Rhamnolipid-mediated biofilm detachment 1215
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
To determine the relative location of the two strainswithin the mixed biofilm (prior to detachment) we usedwild-type and variant bacteria expressing red and greenfluorescent proteins respectively. These experimentsrevealed two surprising results. First, we found that thetwo strains occupied distinctly different locations within themixed biofilm prior to detachment; the wild type localizedto the interior, and the variant to the exterior of the mixedbiofilm (Fig. 4A and B). The peripheral location of thevariants in the cocultures makes it unlikely that they phys-ically dislodge wild-type bacteria during detachment. Sec-ond, in spite of the fact that the centre of the mixedbiofilms consisted primarily of wild-type bacteria, detach-ment occurred via central cavity formation (Fig. 4C). Thiswas unexpected because pure-culture wild-type biofilmsdo not undergo detachment via central hollowing untilmuch later times (~10–12 days) in these growth condi-tions. The formation of cavities within the central region(populated by wild-type bacteria) could be explained by atleast two different mechanisms: the peripherally locatedvariants could activate a detachment mechanism in thecentrally located wild-type bacteria; alternatively, the cen-tral regions may be inherently more susceptible to theaction of a detaching agent, produced in this case byperipherally located variants (see below).
Our observation that the variant physically covered wild-type bacteria in coculture biofilms led us to hypothesizethat the variant may show greater motility than the wildtype. Previous work showed that P. aeruginosa can utilizesurface motility to reach the topmost locations in a biofilm(Klausen et al., 2003). To test this we compared the swim-ming, twitching and swarming motility of the wild-type andvariant strains. Whereas the swimming and twitchingmotility of the two strains was similar, the variant exhibitedmarkedly increased swarming motility (Fig. 5A). Swarm-ing is a type of surface motility that is facilitated by thebiosurfactant, rhamnolipid (Kohler et al., 2000). Rhamno-
lipids are amphipathic glycolipids with surface-active prop-erties that decrease adhesive interactions (Neu, 1996;Desai and Banat, 1997); as such they are candidatedetachment factors.
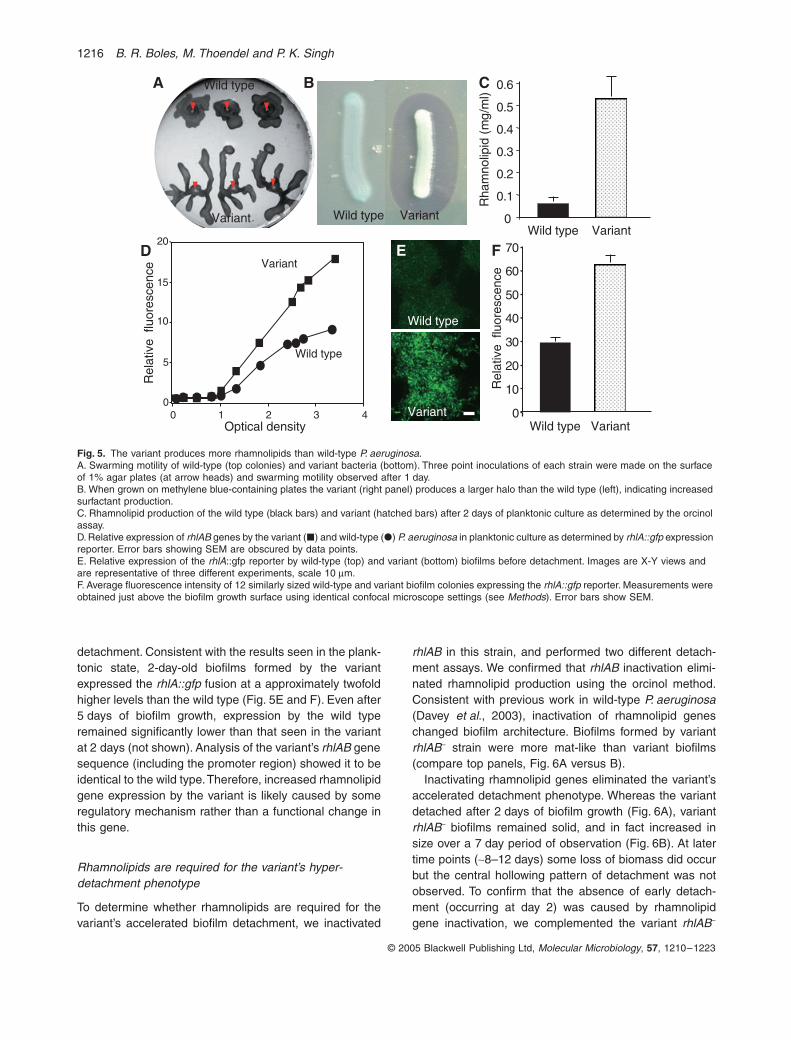
This finding prompted us to investigate rhamnolipid pro-duction by the variant using several methods. The variantproduced a larger halo than the wild type when grown onindicator plates containing methylene blue dye (Fig. 5B).The halo is caused by precipitation of this cationic dye byan anionic surfactant, and in P. aeruginosa this is likely tobe rhamnolipids (Kohler et al., 2000; 2001). We also mea-sured rhamnolipid production using the orcinol method onether-extracted culture supernatants (Koch et al., 1991;Zhang and Miller, 1994; Ochsner and Reiser, 1995; Pear-son et al., 1997; Van Delden et al., 1998; Rahim et al.,2001). This assay showed that rhamnolipid production bythe variant was fivefold higher than the wild type after2 days of planktonic growth (Fig. 5C).
To confirm these results by an independent method,and to determine whether increased rhamnolipid produc-tion resulted from higher gene expression levels or somepost-translational mechanism, we utilized a chromosomalgreen fluorescent protein reporter linked to the promoterof the rhamnolipid biosynthetic genes (rhlAB). Duringplanktonic exponential phase growth, expression of therhlA::gfp fusion by the two strains was identical (Fig. 5D).In stationary phase, however, expression by the variantexceeded that by the wild type, eventually reachingapproximately threefold higher levels. These data are con-sistent with the results of the orcinol assay (Fig. 5C).
Rhamnolipid gene expression is regulated by a numberof factors, not all of which are known (Medina et al., 2003).Thus, the increased level of rhamnolipid expression exhib-ited by the variant in planktonic culture (Fig. 5C and D)may not reflect the situation during biofilm growth. Toexamine this, we compared rhlA::gfp expression levels invariant and wild-type biofilms, just prior to the variant’s
Variants
Wild type
B
A C During detachmentBefore detachment
Before detachment
Fig. 4. The variant and the wild type inhabit different regions in mixed biofilms.A and B. Differential labelling of wild-type (red) and variant (green) bacteria show that the vari-ant covers the wild type in mixed biofilms; wild-type bacteria are localized in the centre of bio-film structures whereas the variants are on the periphery.C. Detachment of the mixed biofilm occurs via the central hallowing pattern after 3 days of bio-film growth. Even though wild-type bacteria (which would not detach at this time) occupy the biofilm centre, and the hyper-detaching vari-ant occupy the exterior, detachment begins in the central regions. Images A and C are X-Y views and B is an X-Z view, scale 10 mm. Results are representative of three indepen-dent experiments. Similar results were obtained with the red and green fluorescent markers switched to the opposite strains.
1216 B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
detachment. Consistent with the results seen in the plank-tonic state, 2-day-old biofilms formed by the variantexpressed the rhlA::gfp fusion at a approximately twofoldhigher levels than the wild type (Fig. 5E and F). Even after5 days of biofilm growth, expression by the wild typeremained significantly lower than that seen in the variantat 2 days (not shown). Analysis of the variant’s rhlAB genesequence (including the promoter region) showed it to beidentical to the wild type. Therefore, increased rhamnolipidgene expression by the variant is likely caused by someregulatory mechanism rather than a functional change inthis gene.
Rhamnolipids are required for the variant’s hyper-detachment phenotype
To determine whether rhamnolipids are required for thevariant’s accelerated biofilm detachment, we inactivated
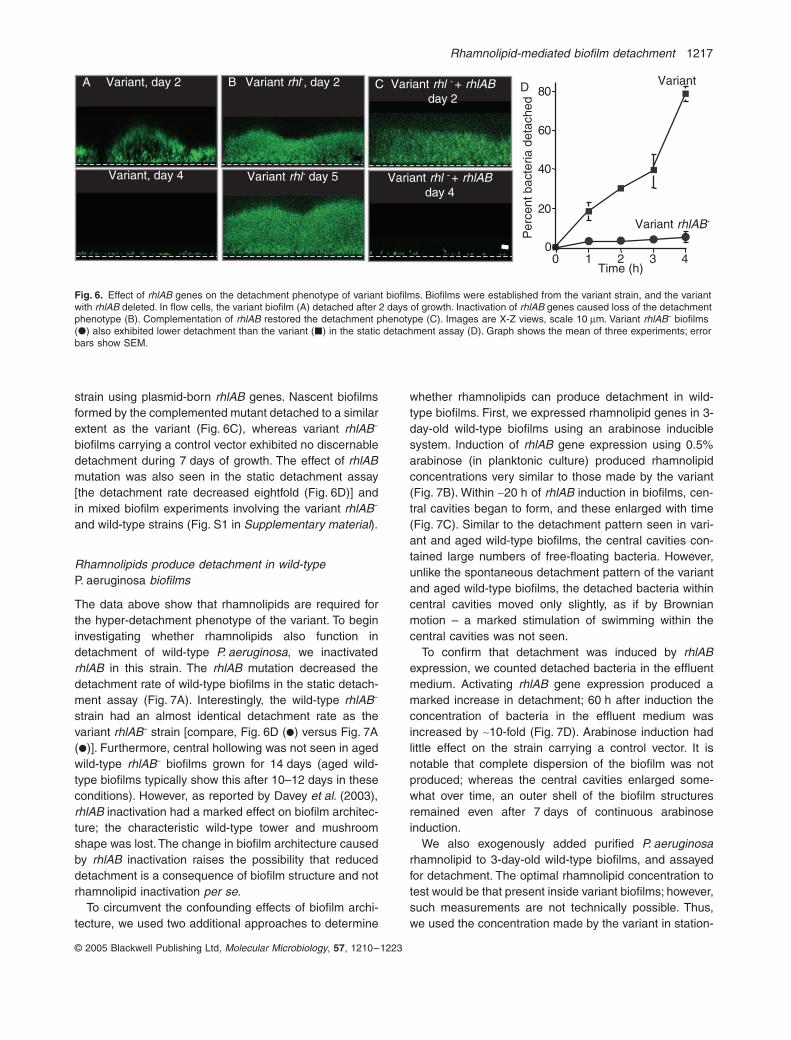
rhlAB in this strain, and performed two different detach-ment assays. We confirmed that rhlAB inactivation elimi-nated rhamnolipid production using the orcinol method.Consistent with previous work in wild-type P. aeruginosa(Davey et al., 2003), inactivation of rhamnolipid geneschanged biofilm architecture. Biofilms formed by variantrhlAB– strain were more mat-like than variant biofilms(compare top panels, Fig. 6A versus B).
Inactivating rhamnolipid genes eliminated the variant’saccelerated detachment phenotype. Whereas the variantdetached after 2 days of biofilm growth (Fig. 6A), variantrhlAB– biofilms remained solid, and in fact increased insize over a 7 day period of observation (Fig. 6B). At latertime points (~8–12 days) some loss of biomass did occurbut the central hollowing pattern of detachment was notobserved. To confirm that the absence of early detach-ment (occurring at day 2) was caused by rhamnolipidgene inactivation, we complemented the variant rhlAB–
Fig. 5. The variant produces more rhamnolipids than wild-type P. aeruginosa.A. Swarming motility of wild-type (top colonies) and variant bacteria (bottom). Three point inoculations of each strain were made on the surface of 1% agar plates (at arrow heads) and swarming motility observed after 1 day.B. When grown on methylene blue-containing plates the variant (right panel) produces a larger halo than the wild type (left), indicating increased surfactant production.C. Rhamnolipid production of the wild type (black bars) and variant (hatched bars) after 2 days of planktonic culture as determined by the orcinol assay.D. Relative expression of rhlAB genes by the variant (�) and wild-type (�) P. aeruginosa in planktonic culture as determined by rhlA::gfp expression reporter. Error bars showing SEM are obscured by data points.E. Relative expression of the rhlA::gfp reporter by wild-type (top) and variant (bottom) biofilms before detachment. Images are X-Y views and are representative of three different experiments, scale 10 mm.F. Average fluorescence intensity of 12 similarly sized wild-type and variant biofilm colonies expressing the rhlA::gfp reporter. Measurements were obtained just above the biofilm growth surface using identical confocal microscope settings (see Methods). Error bars show SEM.
Wild type
Variant
E
Rel
ativ
e flu
ores
cenc
e
VariantWild type0
10
20
30
40
50
60
70
0
5
10
15
20
0 1 2 3 4
Variant
Wild type
Rel
ativ
e flu
ores
cenc
e
Optical density
Rha
mno
lipid
(m
g/m
l)
C
D F
Wild type Variant 0
0.1
0.2
0.3
0.4
0.5
0.6
Wild type Variant
Wild type
Variant
A B
Rhamnolipid-mediated biofilm detachment 1217
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
strain using plasmid-born rhlAB genes. Nascent biofilmsformed by the complemented mutant detached to a similarextent as the variant (Fig. 6C), whereas variant rhlAB–
biofilms carrying a control vector exhibited no discernabledetachment during 7 days of growth. The effect of rhlABmutation was also seen in the static detachment assay[the detachment rate decreased eightfold (Fig. 6D)] andin mixed biofilm experiments involving the variant rhlAB–
and wild-type strains (Fig. S1 in Supplementary material).
Rhamnolipids produce detachment in wild-type P. aeruginosa biofilms
The data above show that rhamnolipids are required forthe hyper-detachment phenotype of the variant. To begininvestigating whether rhamnolipids also function indetachment of wild-type P. aeruginosa, we inactivatedrhlAB in this strain. The rhlAB mutation decreased thedetachment rate of wild-type biofilms in the static detach-ment assay (Fig. 7A). Interestingly, the wild-type rhlAB–
strain had an almost identical detachment rate as thevariant rhlAB– strain [compare, Fig. 6D (�) versus Fig. 7A(�)]. Furthermore, central hollowing was not seen in agedwild-type rhlAB– biofilms grown for 14 days (aged wild-type biofilms typically show this after 10–12 days in theseconditions). However, as reported by Davey et al. (2003),rhlAB inactivation had a marked effect on biofilm architec-ture; the characteristic wild-type tower and mushroomshape was lost. The change in biofilm architecture causedby rhlAB inactivation raises the possibility that reduceddetachment is a consequence of biofilm structure and notrhamnolipid inactivation per se.
To circumvent the confounding effects of biofilm archi-tecture, we used two additional approaches to determine
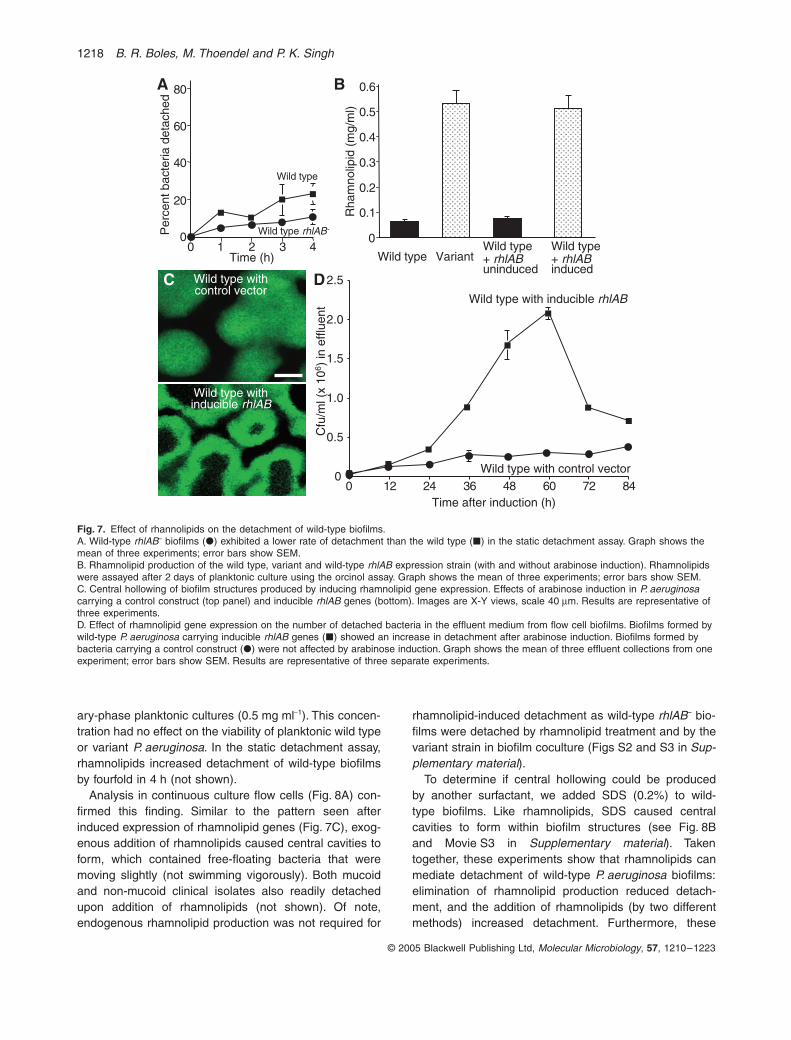
whether rhamnolipids can produce detachment in wild-type biofilms. First, we expressed rhamnolipid genes in 3-day-old wild-type biofilms using an arabinose induciblesystem. Induction of rhlAB gene expression using 0.5%arabinose (in planktonic culture) produced rhamnolipidconcentrations very similar to those made by the variant(Fig. 7B). Within ~20 h of rhlAB induction in biofilms, cen-tral cavities began to form, and these enlarged with time(Fig. 7C). Similar to the detachment pattern seen in vari-ant and aged wild-type biofilms, the central cavities con-tained large numbers of free-floating bacteria. However,unlike the spontaneous detachment pattern of the variantand aged wild-type biofilms, the detached bacteria withincentral cavities moved only slightly, as if by Brownianmotion – a marked stimulation of swimming within thecentral cavities was not seen.
To confirm that detachment was induced by rhlABexpression, we counted detached bacteria in the effluentmedium. Activating rhlAB gene expression produced amarked increase in detachment; 60 h after induction theconcentration of bacteria in the effluent medium wasincreased by ~10-fold (Fig. 7D). Arabinose induction hadlittle effect on the strain carrying a control vector. It isnotable that complete dispersion of the biofilm was notproduced; whereas the central cavities enlarged some-what over time, an outer shell of the biofilm structuresremained even after 7 days of continuous arabinoseinduction.
We also exogenously added purified P. aeruginosarhamnolipid to 3-day-old wild-type biofilms, and assayedfor detachment. The optimal rhamnolipid concentration totest would be that present inside variant biofilms; however,such measurements are not technically possible. Thus,we used the concentration made by the variant in station-
Fig. 6. Effect of rhlAB genes on the detachment phenotype of variant biofilms. Biofilms were established from the variant strain, and the variant with rhlAB deleted. In flow cells, the variant biofilm (A) detached after 2 days of growth. Inactivation of rhlAB genes caused loss of the detachment phenotype (B). Complementation of rhlAB restored the detachment phenotype (C). Images are X-Z views, scale 10 mm. Variant rhlAB– biofilms (�) also exhibited lower detachment than the variant (�) in the static detachment assay (D). Graph shows the mean of three experiments; error bars show SEM.
0
Variant
Variant rhlAB-
0 1 2 3 4Time (h)
Per
cent
bac
teria
det
ache
d
20
40
60
80DVariant rhl-, day 2Variant, day 2
Variant, day 4 Variant rhl- day 5
A B Variant rhl - + rhlABday 2
Variant rhl - + rhlABday 4
C
1218 B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
ary-phase planktonic cultures (0.5 mg ml-1). This concen-tration had no effect on the viability of planktonic wild typeor variant P. aeruginosa. In the static detachment assay,rhamnolipids increased detachment of wild-type biofilmsby fourfold in 4 h (not shown).
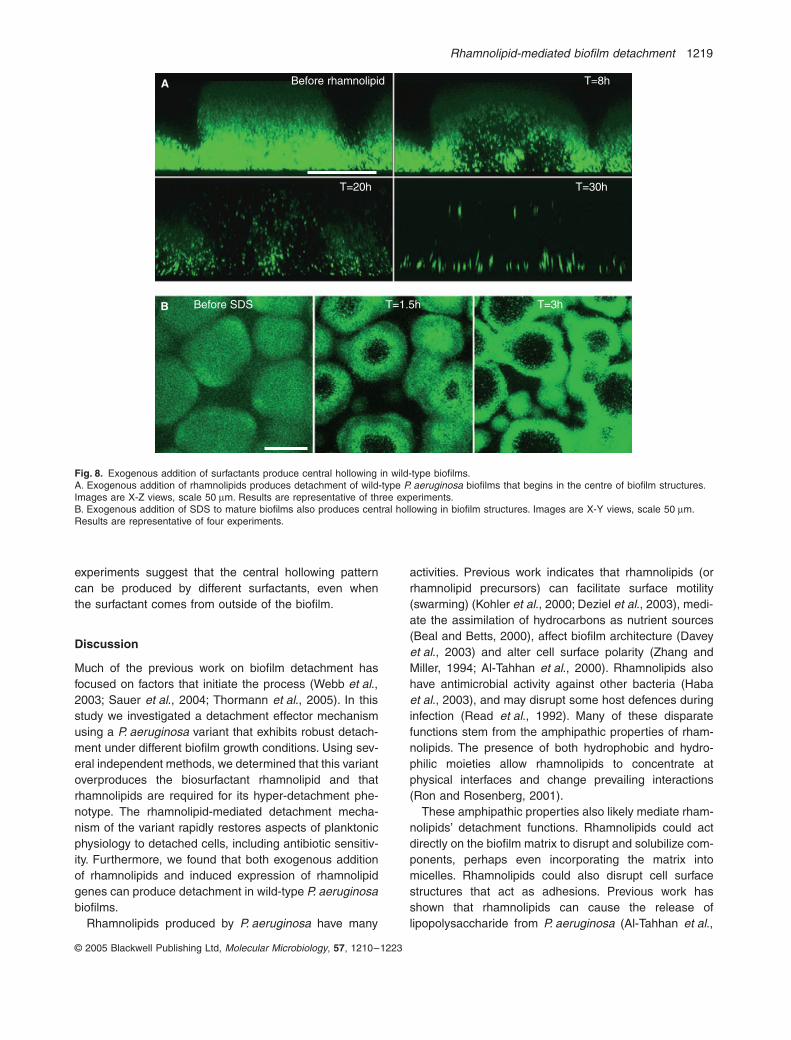
Analysis in continuous culture flow cells (Fig. 8A) con-firmed this finding. Similar to the pattern seen afterinduced expression of rhamnolipid genes (Fig. 7C), exog-enous addition of rhamnolipids caused central cavities toform, which contained free-floating bacteria that weremoving slightly (not swimming vigorously). Both mucoidand non-mucoid clinical isolates also readily detachedupon addition of rhamnolipids (not shown). Of note,endogenous rhamnolipid production was not required for
rhamnolipid-induced detachment as wild-type rhlAB– bio-films were detached by rhamnolipid treatment and by thevariant strain in biofilm coculture (Figs S2 and S3 in Sup-plementary material).
To determine if central hollowing could be producedby another surfactant, we added SDS (0.2%) to wild-type biofilms. Like rhamnolipids, SDS caused centralcavities to form within biofilm structures (see Fig. 8Band Movie S3 in Supplementary material). Takentogether, these experiments show that rhamnolipids canmediate detachment of wild-type P. aeruginosa biofilms:elimination of rhamnolipid production reduced detach-ment, and the addition of rhamnolipids (by two differentmethods) increased detachment. Furthermore, these
Fig. 7. Effect of rhannolipids on the detachment of wild-type biofilms.A. Wild-type rhlAB– biofilms (�) exhibited a lower rate of detachment than the wild type (�) in the static detachment assay. Graph shows the mean of three experiments; error bars show SEM.B. Rhamnolipid production of the wild type, variant and wild-type rhlAB expression strain (with and without arabinose induction). Rhamnolipids were assayed after 2 days of planktonic culture using the orcinol assay. Graph shows the mean of three experiments; error bars show SEM.C. Central hollowing of biofilm structures produced by inducing rhamnolipid gene expression. Effects of arabinose induction in P. aeruginosa carrying a control construct (top panel) and inducible rhlAB genes (bottom). Images are X-Y views, scale 40 mm. Results are representative of three experiments.D. Effect of rhamnolipid gene expression on the number of detached bacteria in the effluent medium from flow cell biofilms. Biofilms formed by wild-type P. aeruginosa carrying inducible rhlAB genes (�) showed an increase in detachment after arabinose induction. Biofilms formed by bacteria carrying a control construct (�) were not affected by arabinose induction. Graph shows the mean of three effluent collections from one experiment; error bars show SEM. Results are representative of three separate experiments.
Wild type
Wild type rhlAB-
0 1 2 3 4Time (h)
Per
cent
bac
teria
det
ache
d
20
40
60
80
Wild type Variant
Rha
mno
lipid
(m
g/m
l)
0
0.1
0.2
0.3
0.4
0.5
0.6
Wild type+ rhlABuninduced
Wild type+ rhlABinduced
Cfu
/ml (
x 10
6 ) in
effl
uent
0
0.5
1.0
1.5
2.0
2.5
0 12 24 36 48 60 72 84Time after induction (h)
A B
C D
Wild type with control vector
Wild type with inducible rhlAB
0
Wild type withcontrol vector
Wild type withinducible rhlAB
Rhamnolipid-mediated biofilm detachment 1219
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
experiments suggest that the central hollowing patterncan be produced by different surfactants, even whenthe surfactant comes from outside of the biofilm.
Discussion
Much of the previous work on biofilm detachment hasfocused on factors that initiate the process (Webb et al.,2003; Sauer et al., 2004; Thormann et al., 2005). In thisstudy we investigated a detachment effector mechanismusing a P. aeruginosa variant that exhibits robust detach-ment under different biofilm growth conditions. Using sev-eral independent methods, we determined that this variantoverproduces the biosurfactant rhamnolipid and thatrhamnolipids are required for its hyper-detachment phe-notype. The rhamnolipid-mediated detachment mecha-nism of the variant rapidly restores aspects of planktonicphysiology to detached cells, including antibiotic sensitiv-ity. Furthermore, we found that both exogenous additionof rhamnolipids and induced expression of rhamnolipidgenes can produce detachment in wild-type P. aeruginosabiofilms.
Rhamnolipids produced by P. aeruginosa have many
activities. Previous work indicates that rhamnolipids (orrhamnolipid precursors) can facilitate surface motility(swarming) (Kohler et al., 2000; Deziel et al., 2003), medi-ate the assimilation of hydrocarbons as nutrient sources(Beal and Betts, 2000), affect biofilm architecture (Daveyet al., 2003) and alter cell surface polarity (Zhang andMiller, 1994; Al-Tahhan et al., 2000). Rhamnolipids alsohave antimicrobial activity against other bacteria (Habaet al., 2003), and may disrupt some host defences duringinfection (Read et al., 1992). Many of these disparatefunctions stem from the amphipathic properties of rham-nolipids. The presence of both hydrophobic and hydro-philic moieties allow rhamnolipids to concentrate atphysical interfaces and change prevailing interactions(Ron and Rosenberg, 2001).
These amphipathic properties also likely mediate rham-nolipids’ detachment functions. Rhamnolipids could actdirectly on the biofilm matrix to disrupt and solubilize com-ponents, perhaps even incorporating the matrix intomicelles. Rhamnolipids could also disrupt cell surfacestructures that act as adhesions. Previous work hasshown that rhamnolipids can cause the release oflipopolysaccharide from P. aeruginosa (Al-Tahhan et al.,
Fig. 8. Exogenous addition of surfactants produce central hollowing in wild-type biofilms.A. Exogenous addition of rhamnolipids produces detachment of wild-type P. aeruginosa biofilms that begins in the centre of biofilm structures. Images are X-Z views, scale 50 mm. Results are representative of three experiments.B. Exogenous addition of SDS to mature biofilms also produces central hollowing in biofilm structures. Images are X-Y views, scale 50 mm. Results are representative of four experiments.
Before SDS T=1.5h T=3h
A Before rhamnolipid T=8h
T=20h T=30h
B

1220 B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
2000) and they may have similar effects on other surfaceappendages. If these structures act to tether bacteria toeach other or to the matrix, their disruption could inducedetachment. A similar mechanism may underlie rhamno-lipids’ action in maintaining the fluid channels (openareas) that surround biofilm structures (Davey et al.,2003). However in this case, rhamnolipids may act oncell–surface rather than cell–cell or cell–matrix interac-tions. The ability of rhamnolipids to promote swarmingmotility and alter cell surface charge could also help tomediate detachment. Of note, rhamnolipids have beenshown to disrupt Salmonella typhimurium biofilms, sug-gesting that they may have activity against a broad spec-trum of organisms (Mireles et al., 2001).
An interesting finding from our study was that the rham-nolipid-mediated detachment mechanism of the variantinvolved the central hollowing pattern that is also seen inaged wild-type biofilms. Detachment from the biofilm inte-rior has been observed in other organisms includingStaphlococcus aureus (Yarwood et al., 2004) and in theoral pathogen Actinobacillus actinomycetemcomitans(Kaplan et al., 2003a,b). In A. actinomycetemcomitansdetachment is mediated by the enzymatic action of a N-acetylglucosaminidase rather than a surfactant (Kaplanet al., 2003b), suggesting that the central hollowingdetachment pattern may be common across species,even when different detachment mechanisms areoperative.
What produces the central hollowing detachment pat-tern? Other investigators have proposed that central hol-lowing may occur because the signals that triggerdetachment are strongest in the interior of biofilm struc-tures (see model, Fig. 9A) (Allison et al., 1998; Hentzeret al., 2002; Hunt et al., 2004). This would result in pref-erential activation of detachment mechanisms in the bio-film centre. Two general types of triggers have beensuggested that could show this kind of centralized activity:the accumulation of a signalling molecule or waste prod-uct, or the depletion of one or multiple metabolic sub-strates (Hunt et al., 2004).
Interestingly, rhamnolipid expression can be regulatedby both of these putative triggers. The rhlAB genes areregulated by cell density-dependent quorum sensing, inwhich the level of expression is determined in part by theconcentration of asyl-homoserine lactone signalling mol-ecules present in the local environment (Ochsner andReiser, 1995). These (or other signals) could accumulatein the centre of biofilm structures. Rhamnolipids can alsobe induced by starvation of many nutrients includingnitrogen, phosphate, magnesium, calcium, potassium,sodium, iron and trace elements (Desai and Banat, 1997).As with signal accumulation, nutrient limitation may bemost severe in the biofilm interior. Furthermore, the mul-tiplicity of starvation signals could provide redundancy for
this important function. This could be of great advantageas nutrient limitation is a major threat to fixed, densebacterial populations.
While it is certainly possible that either starvation orsignal accumulation could induce rhamnolipid-mediateddetachment, our experiments suggest that central trigger-ing is not required to produce the central hollowing pat-tern. In all the experiments we conducted that involvedincreasing rhamnolipid levels in biofilms, the rhamnolipidsoriginated from outside of or uniformly throughout thebiofilm. In the case of the exogenous rhamnolipid additionexperiments (Fig. 7E), and the coculture experiments inwhich the variants localized to the outer biofilm regions(Figs 3 and 4), rhamnolipids likely originated from theperiphery. The system we used to induce rhamnolipidgene expression results in fairly uniform expressionthroughout the biofilm structures (P. K. Singh and B. R.Boles, unpubl. obs.; Lequette and Greenberg, 2005).
These findings suggest an alternative hypothesis toexplain central hollowing detachment that does not invokeregional triggering: biofilm structures may consist of dis-tinct subpopulations with differing susceptibility to detach-ing agents; the centre of biofilm structures are moresusceptible while the peripheral biofilm regions form amore durable shell that is relatively resistant (see model,Fig. 9B). The fact that the exogenous addition of bothrhamnolipids and SDS produced cavities in the centre ofbiofilm structures suggests that this region may be gen-erally more susceptible to various soluble agents thatproduce detachment. Increased susceptibility of internalpopulations could be caused by regional diversity in matrix
Fig. 9. Models to explain the central hallowing detachment pattern.A. Central triggering model. In this model, central detachment results from preferential triggering of detachment in the biofilm interior. The shaded region represents the intensity of a detachment trigger (such as starvation or signal accumulation). The graph depicts the relative concentration of the triggering signal along the biofilm cross-section. See text and the study by Hunt et al. (2004).B. Central susceptibility model. Central hollowing detachment could also be explained by the presence of distinct subpopulations with different susceptibility to detachment effector mechanisms. Physio-logical differences in the susceptible central regions (white area) result in preferential detachment irrespective of where the detach-ment mechanism is triggered.
a bCentral
triggeringmodel
Centralsusceptibility
model
Outer shell isresistant todetachment
Interior issusceptible todetachment
Intensity ofdetachment
signal
¨positionÆ

Rhamnolipid-mediated biofilm detachment 1221
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
or adhesin composition, or other physiological differences.The existence of populations with differing susceptibilitiesmay also explain how rhamnolipids act to maintain openareas (fluid channels) around biofilm structures withoutdisrupting the adjacent biofilms (Davey et al., 2003; Espi-nosa-Urgel, 2003). If bacteria in the durable outer shellsecrete moderate levels of rhamnolipids, this could detachbacteria in fluid channels without disrupting the biofilmitself. Of note, the two models are not mutually exclusive;some detachment mechanisms may involve both centralsusceptibility and central triggering.
Several of our findings lend support to the idea thatbiofilm detachment is under the control of a global regu-lator that might co-ordinate related detachment functions.First, whereas inactivation of rhamnolipid genes elimi-nated the detachment phenotype of the variant and rham-nolipids produced detachment in the wild type, neitherrhamnolipid addition nor induced expression of rhamno-lipids completely reproduced the pattern of detachmentseen in the variant or aged wild-type biofilms. Inducedexpression brought about incomplete detachment(Fig. 7C), and neither method produced rapidly swimmingbacteria within the central cavities (see Movies S1 and S2in Supplementary material). This finding raises thepossibility that the complete central hollowing patternmay require other co-ordinately expressed functions.Examples of such detachment cofactors could includemotility functions or matrix degrading enzymes such asexopolysaccharide lyases (Boyd and Chakrabarty, 1995).
Second, although the variant overexpressed rhamnolip-ids in all the conditions tested, we found no mutation inthe rhamnolipid gene sequence (including the promoterregion). This suggests that some regulatory mechanismis responsible for rhamnolipid overexpression in the vari-ant. Lastly, the variant’s detachment function rapidlyrestored some aspects of planktonic physiology to theseparating cells. This transition to planktonic physiologycould also be regulated and serve to prepare bacteria forconditions outside the biofilm.
Biofilms that exist in natural environments or withininfected patients are certain to be more complex thanthose studied here. These biofilms may differ physiologi-cally, contain multiple bacterial species and may incorpo-rate exogenous substances such as mucus and DNA froma host, or polysaccharides and proteins from the localenvironment. While it remains to be determined whetherrhamnolipids would be effective against such biofilms,inducing detachment is an attractive treatment strategyespecially given the difficulty in overcoming biofilm-induced resistance.
Acknowledgements
We thank P. Bontu, A. Turner, T. Moninger and M. Nevell for
technical assistance and M. J. Welsh, M. R. Parsek, T. L. Yahr,E. P. Greenberg and H. R. Wilson for helpful discussions.
References
Adair, C.G., Gorman, S.P., Feron, B.M., Byers, L.M., Jones,D.S., Goldsmith, C.E., et al. (1999) Implications of endot-racheal tube biofilm for ventilator-associated pneumonia.Intensive Care Med 25: 1072–1076.
Allison, D.G., Ruiz, B., SanJose, C., Jaspe, A., and Gilbert,P. (1998) Extracellular products as mediators of the forma-tion and detachment of Pseudomonas fluorescens bio-films. FEMS Microbiol Lett 167: 179–184.
Al-Tahhan, R.A., Sandrin, T.R., Bodour, A.A., and Maier,R.M. (2000) Rhamnolipid-induced removal of lipopolysac-charide from Pseudomonas aeruginosa: effect on cellsurface properties and interaction with hydrophobic sub-strates. Appl Environ Microbiol 66: 3262–3268.
Beal, R., and Betts, W.B. (2000) Role of rhamnolipid biosur-factants in the uptake and mineralization of hexadecane inPseudomonas aeruginosa. J Appl Microbiol 89: 158–168.
Bergmans, D.C., Bonten, M.J., Stobberingh, E.E., van Tiel,F.H., van der Geest, S., de Leeuw, P.W., and Gaillard, C.A.(1998) Colonization with Pseudomonas aeruginosa inpatients developing ventilator-associated pneumonia.Infect Control Hosp Epidemiol 19: 853–855.
Boles, B.R., Thoendel, M., and Singh, P.K. (2004) Self-gen-erated diversity produces ‘insurance effects’ in biofilm com-munities. Proc Natl Acad Sci USA 101: 16630–16635.
Boyd, A., and Chakrabarty, A.M. (1995) Pseudomonasaeruginosa biofilms: role of the alginate exopolysaccha-ride. J Ind Microbiol 15: 162–168.
Campbell, R.E., Tour, O., Palmer, A.E., Steinbach, P.A.,Baird, G.S., Zacharias, D.A., and Tsien, R.Y. (2002) Amonomeric red fluorescent protein. Proc Natl Acad SciUSA 99: 7877–7882.
Costerton, J.W., Stewart, P.S., and Greenberg, E.P. (1999)Bacterial biofilms: a common cause of persistent infec-tions. Science 284: 1318–1322.
Davey, M.E., Caiazza, N.C., and O’Toole, G.A. (2003) Rham-nolipid surfactant production affects biofilm architecture inPseudomonas aeruginosa PA01. J Bacteriol 185: 1027–1036.
Davies, D.G., Parsek, M.R., Pearson, J.P., Iglewski, B.H.,Costerton, J.W., and Greenberg, E.P. (1998) The involve-ment of cell-to-cell signals in the development of a bacterialbiofilm. Science 280: 295–298.
Desai, J.D., and Banat, I.M. (1997) Microbial production ofsurfactants and their commercial potential. Microbiol MolBiol Rev 61: 47–64.
Deziel, E., Lepine, F., Milot, S., and Villemur, R. (2003) rhlAis required for the production of a novel biosurfactant pro-moting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precur-sors of rhamnolipids. Microbiology 149: 2005–2013.
Espinosa-Urgel, M. (2003) Resident parking only: rhamnolip-ids maintain fluid channels in biofilms. J Bacteriol 185:699–700.
Feldman, C., Kassel, M., Cantrell, J., Kaka, S., Morar, R.,Goolam Mahomed, A., and Philips, J.I. (1999) The pres-ence and sequence of endotracheal tube colonization in

1222 B. R. Boles, M. Thoendel and P. K. Singh
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
patients undergoing mechanical ventilation. Eur Respir J13: 546–551.
Friedman, L., and Kolter, R. (2004) Two genetic loci producedistinct carbohydrate-rich structural components of thePseudomonas aeruginosa biofilm matrix. J Bacteriol 186:4457–4465.
Haba, E., Pinazo, A., Jauregui, O., Espuny, M.J., Infante,M.R., and Manresa, A. (2003) Physicochemical character-ization and antimicrobial properties of rhamnolipids pro-duced by Pseudomonas aeruginosa 47T2 NCBIM 40044.Biotechnol Bioeng 81: 316–322.
Hassett, D.J., Ma, J.F., Elkins, J.G., McDermott, T.R.,Ochsner, U.A., West, S.E., et al. (1999) Quorum sensingin Pseudomonas aeruginosa controls expression of cata-lase and superoxide dismutase genes and mediates bio-film susceptibility to hydrogen peroxide. Mol Microbiol 34:1082–1093.
Hausner, M., and Wuertz, S. (1999) High rates of conjugationin bacterial biofilms as determiend by quantitative in situanalysis. Appl Environ Microbiol 65: 3710–3713.
Hentzer, M., Riedel, K., Rasmussen, T.B., Heydorn, A.,Andersen, J.B., Parsek, M.R., et al. (2002) Inhibition ofquorum sensing in Pseudomonas aeruginosa biofilm bac-teria by a halogenated furanone compound. Microbiology148: 87–102.
Hoang, T.T., Kutchma, A.J., Becher, A., and Schweizer, H.P.(2000) Integration-proficient plasmids for Pseudomonasaeruginosa: site-specific integration and use for engineer-ing of reporter and expression strains. Plasmid 43: 59–72.
Hoiby, N., Krogh Johansen, H., Moser, C., Song, Z., Ciofu,O., and Kharazmi, A. (2001) Pseudomonas aeruginosaand the in vitro and in vivo biofilm mode of growth.Microbes Infect 3: 23–35.
Huang, C.T., Xu, K.D., McFeters, G.A., and Stewart, P.S.(1998) Spatial patterns of alkaline phosphatase expressionwithin bacterial colonies and biofilms in response to phos-phate starvation. Appl Environ Microbiol 64: 1526–1531.
Hunt, S.M., Werner, E.M., Huang, B., Hamilton, M.A., andStewart, P.S. (2004) Hypothesis for the role of nutrientstarvation in biofilm detachment. Appl Environ Microbiol70: 7418–7425.
Kaplan, J.B., Meyenhofer, M.F., and Fine, D.H. (2003a) Bio-film growth and detachment of Actinobacillus actinomyce-temcomitans. J Bacteriol 185: 1399–1404.
Kaplan, J.B., Ragunath, C., Ramasubbu, N., and Fine, D.H.(2003b) Detachment of Actinobacillus actinomycetemcom-itans biofilm cells by an endogenous beta-hexosaminidaseactivity. J Bacteriol 185: 4693–4698.
Klausen, M., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003) Involvement of bacterial migration in thedevelopment of complex multicellular structures inPseudomonas aeruginosa biofilms. Mol Microbiol 50: 61–68.
Koch, A.K., Kappeli, O., Fiechter, A., and Reiser, J. (1991)Hydrocarbon assimilation and biosurfactant production inPseudomonas aeruginosa mutants. J Bacteriol 173: 4212–4219.
Kohler, T., Curty, L.K., Barja, F., van Delden, C., andPechere, J.C. (2000) Swarming of Pseudomonas aerugi-nosa is dependent on cell-to-cell signaling and requiresflagella and pili. J Bacteriol 182: 5990–5996.
Kohler, T., Van Delden, C., Curty, L.K., Hamzehpour, M.M.,and Pechere, J.C. (2001) Overexpression of the MexEF-OprN multidrug efflux system affects cell-to-cell signalingin Pseudomonas aeruginosa. J Bacteriol 183: 5213–5222.
Lequette, Y., and Greenberg, E.P. (2005) Timing and local-ization of rhamnolipid synthesis gene expression inPseudomonas aeruginosa biofilms. J Bacteriol 187: 37–44.
Mah, T.F., Pitts, B., Pellock, B., Walker, G.C., Stewart, P.S.,and O’Toole, G.A. (2003) A genetic basis for Pseudomonasaeruginosa biofilm antibiotic resistance. Nature 426: 306–310.
Medina, G., Juarez, K., and Soberon-Chavev, G. (2003) ThePseudomonas aeruginosa rhlAB operon is not expressedduring the logarithmic phase of growth even in the pres-ence of its activator RhlR and the autoinducer N-butyryl-homoserine lactone. J Bacteriol 185: 377–380.
Mireles,J.R., II, Toguchi, A., and Harshey, R.M. (2001) Sal-monella enterica serovar typhimurium swarming mutantswith altered biofilm-forming abilities: surfactin inhibits bio-film formation. J Bacteriol 183: 5848–5854.
Neu, T.R. (1996) Significance of bacterial surface-activecompounds in interaction of bacteria with interfaces. Micro-biol Rev 60: 151–166.
Newman, J.R., and Fuqua, C. (1999) Broad-host-rangeexpression vectors that carry the L-arabinose-inducibleEcherichia coli araBAD promoter and the araC regulator.Gene 227: 197–203.
Ochsner, U.A., and Reiser, J. (1995) Autoinducer-mediatedregulation of rhamnolipid biosurfactant synthesis inPseudomonas aeruginosa. Proc Natl Acad Sci USA 92:6424–6428.
Parsek, M.R., and Greenberg, E.P. (2000) Acyl-homoserinelactone quorum sensing in Gram-negative bacteria: a sig-naling mechanism involved in associations with higherorganisms. Proc Natl Acad Sci USA 97: 8789–8793.
Parsek, M.R., and Singh, P.K. (2003) Bacterial biofilms: anemerging link to disease pathogenesis. Annu Rev Micro-biol 57: 677–701.
Pearson, J.P., Pesci, E.C., and Iglewski, B.H. (1997) Rolesof Pseduomonas aeruginosa las and rhl quorum-sensingsystems in control of elastase and rhamnolipid biosynthe-sis genes. J Bacteriol 179: 5756–5767.
Picioreanu, C., van Loosdrecht, M.C., and Heijnen, J.J.(2001) Two-dimensional model of biofilm detachmentcaused by internal stress from liquid flow. BiotechnolBioeng 72: 205–218.
Rahim, R., Ochsner, U.A., Olvera, C., Graninger, M., Mess-ner, P., Lam, J.S., and Soberon-Chavev, G. (2001) Cloningand functional characterization of the Pseudomonasaeruginosa rhlC gene that encodes rhamnosyltransferase2, an enzyme responsible for di-rhamnolipid biosynthesis.Mol Microbiol 40: 708–718.
Read, R.C., Roberts, P., Munro, N., Rutman, A., Hastie, A.,Shryock, T., et al. (1992) Effect of Pseudomonas aerugi-nosa rhamnolipids on mucociliary transport and ciliarybeating. J Appl Physiol 72: 2271–2277.
Ron, E.Z., and Rosenberg, E. (2001) Natural roles of biosur-factants. Environ Microbiol 3: 229–236.
Sauer, K., Camper, A.K., Ehrlich, G.D., Costerton, J.W., andDavies, D.G. (2002) Pseudomonas aeruginosa displays

Rhamnolipid-mediated biofilm detachment 1223
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1210–1223
multiple phenotypes during development as a biofilm. JBacteriol 184: 1140–1154.
Sauer, K., Cullen, M.C., Rickard, A.H., Zeef, L.A., Davies,D.G., and Gilbert, P. (2004) Characterization of nutrient-induced dispersion in Pseudomonas aeruginosa PAO1 bio-film. J Bacteriol 186: 7312–7326.
Shapiro, J.A. (1998) Thinking about bacterial populations asmulticellular organisms. Annu Rev Microbiol 52: 81–104.
Stewart, P.S. (2002) Mechanisms of antibiotic resistance inbacterial biofilms. Int J Med Microbiol 292: 107–113.
Stoodley, P., Wilson, S., Hall-Stoodley, L., Boyle, J.D., Lap-pin-Scott, H.M., and Costerton, J.W. (2001) Growth anddetachment of cell clusters from mature mixed-speciesbiofilms. Appl Environ Microbiol 67: 5608–5613.
Stoodley, P., Cargo, R., Rupp, C.J., Wilson, S., and Klapper,I. (2002) Biofilm material properties as related to shear-induced deformation and detachment phenomena. J IndMicrobiol Biotechnol 29: 361–367.
Thormann, K.M., Saville, R.M., Shukla, S., and Spormann,A.M. (2005) Induction of rapid detachment in Shewanellaoneidensis MR-1 biofilms. J Bacteriol 187: 1014–1021.
Van Delden, C., Pesci, E.C., Pearson, J.P., and Iglewski, B.H.(1998) Starvation selection restores elastase and rhamno-lipid production in a Pseudomonas aeruginosa quorum-sensing mutant. Infect Immun 66: 4499–4502.
Webb, J.S., Thompson, L.S., James, S., Charlton, T., Tolker-Nielsen, T., Koch, B., et al. (2003) Cell death in Pseudomo-nas aeruginosa biofilm development. J Bacteriol 185:4585–4592.
Whiteley, M., Bangera, M.G., Bumgarner, R.E., Parsek, M.R.,Teitzel, G.M., Lory, S., and Greenberg, E.P. (2001) Geneexpression in Pseudomonas aeruginosa biofilms. Nature413: 860–864.
Yarwood, J.M., Bartels, D.J., Volper, E.M., and Greenberg,E.P. (2004) Quorum sensing in Staphylococcus aureusbiofilms. J Bacteriol 186: 1838–1850.
Zhang, Y., and Miller, R.M. (1994) Effect of a Pseudomonasrhamnolipid biosurfactant on cell hydrophobicity and bio-degradation of octadecane. Appl Environ Microbiol 60:2101–2106.
Supplementary material
The following supplementary material is available for thisarticle online:Movie S1. Variant biofilms undergo detachment via the cen-tral hollowing pattern early in biofilm development.Movie S2. Wild-type biofilms undergo detachment via thecentral hollowing pattern late in biofilm development.Movie S3. The surfactant SDS produces central hollowing inbiofilms.Fig. S1. Inactivation of rhlAB genes eliminates the variantsability to produce detachment in mixed biofilms.Fig. S2. Exogenous addition of rhamnolipids producedetachment of the wild-type rhlAB– strain.Fig. S3. The variant causes detachment of wild-type rhlAB–
bacteria in mixed biofilms.
Recommended

















