Embed Size (px)
Citation preview

Journal club, 10 July 2013Hiromi MATSUMAE
中世ヨーロッパのハンセン病菌のゲノム解読
published in 13 June 2013
113年7月16日火曜日

development of ancient pathogen genomics
13 June 2013
Neanderthal genome, 2010
213年7月16日火曜日

development of ancient pathogen genomics
13 June 2013
Neanderthal genome, 2010
中世のペスト(黒死病)菌 ゲノム、in 2011
213年7月16日火曜日

Leprosy (ハンセン病)現代では抗生物質が有効なため、先進国ではほとんど患者がいないantibiotic is availableless infectious, most patients are infected in early childhood
prevalence in developing countries (Brazil, India, etc.)
historically, patients were the objects of discrimination in the world due to damaged skin
313年7月16日火曜日
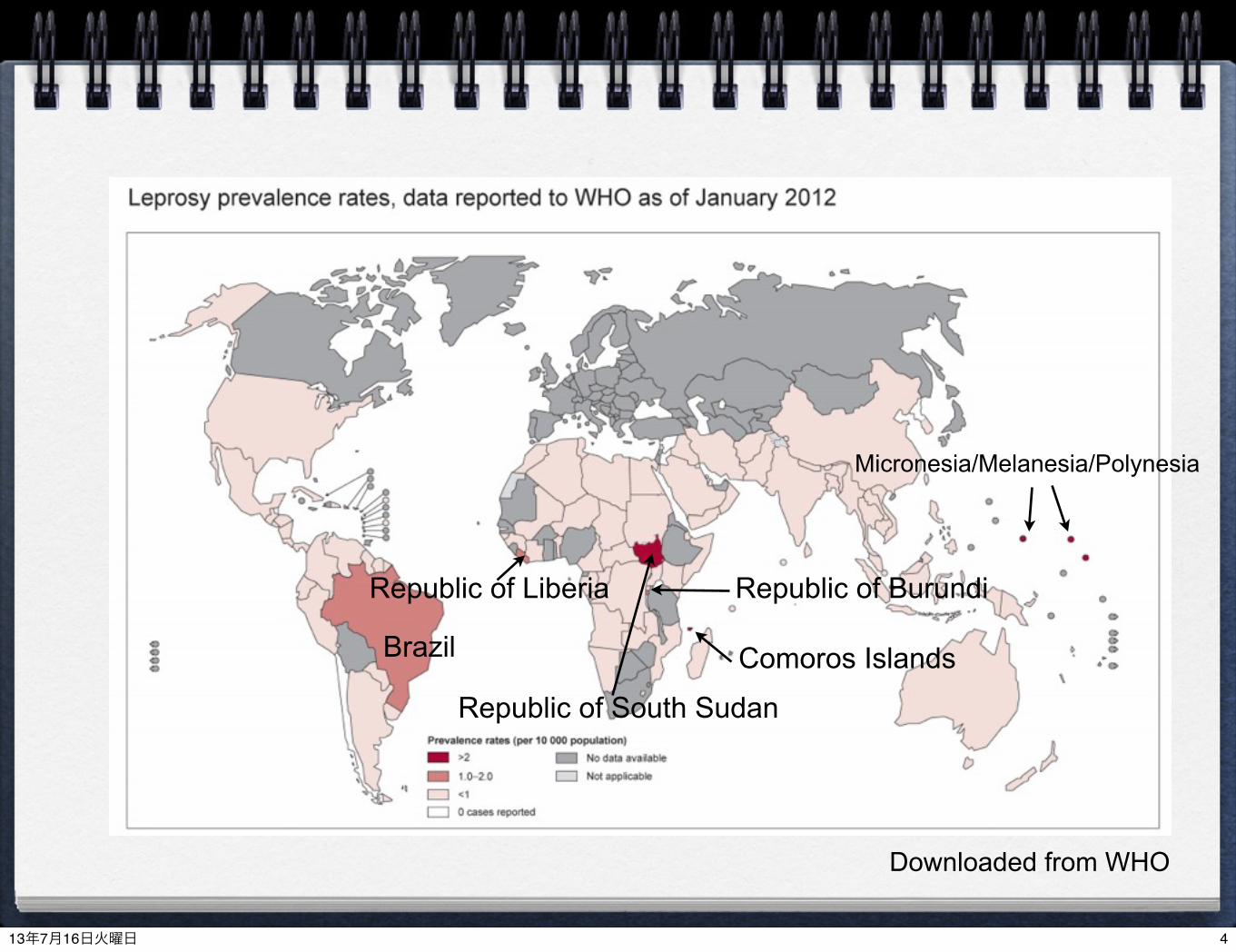
Brazil
Republic of South Sudan
Comoros Islands
Republic of BurundiRepublic of Liberia
Micronesia/Melanesia/Polynesia
Downloaded from WHO
413年7月16日火曜日
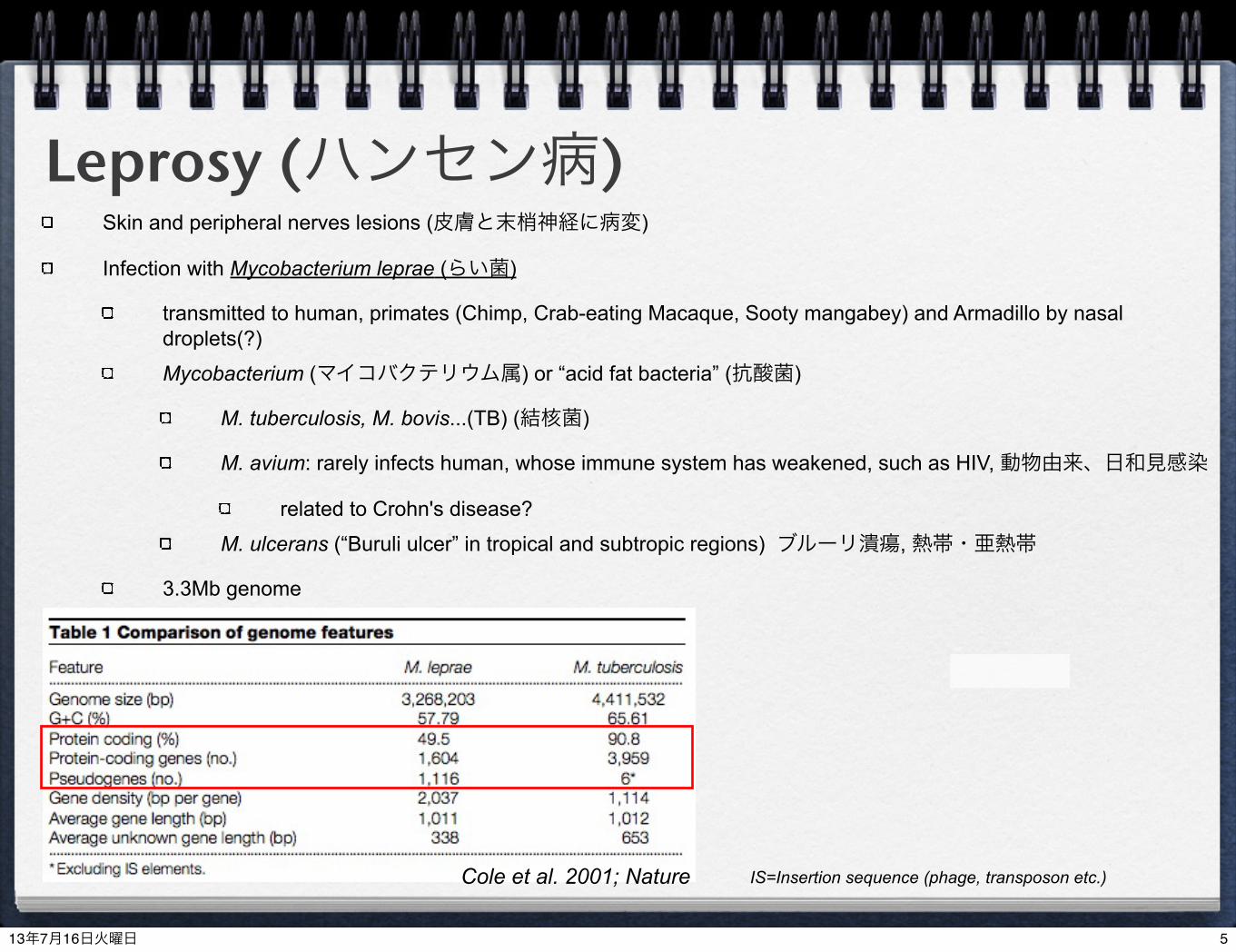
Leprosy (ハンセン病)Skin and peripheral nerves lesions (皮膚と末梢神経に病変)
Infection with Mycobacterium leprae (らい菌)
transmitted to human, primates (Chimp, Crab-eating Macaque, Sooty mangabey) and Armadillo by nasal droplets(?)
Mycobacterium (マイコバクテリウム属) or “acid fat bacteria” (抗酸菌)
M. tuberculosis, M. bovis...(TB) (結核菌)
M. avium: rarely infects human, whose immune system has weakened, such as HIV, 動物由来、日和見感染
related to Crohn's disease?
M. ulcerans (“Buruli ulcer” in tropical and subtropic regions) ブルーリ潰瘍, 熱帯・亜熱帯
3.3Mb genome
Cole et al. 2001; Nature IS=Insertion sequence (phage, transposon etc.)
513年7月16日火曜日
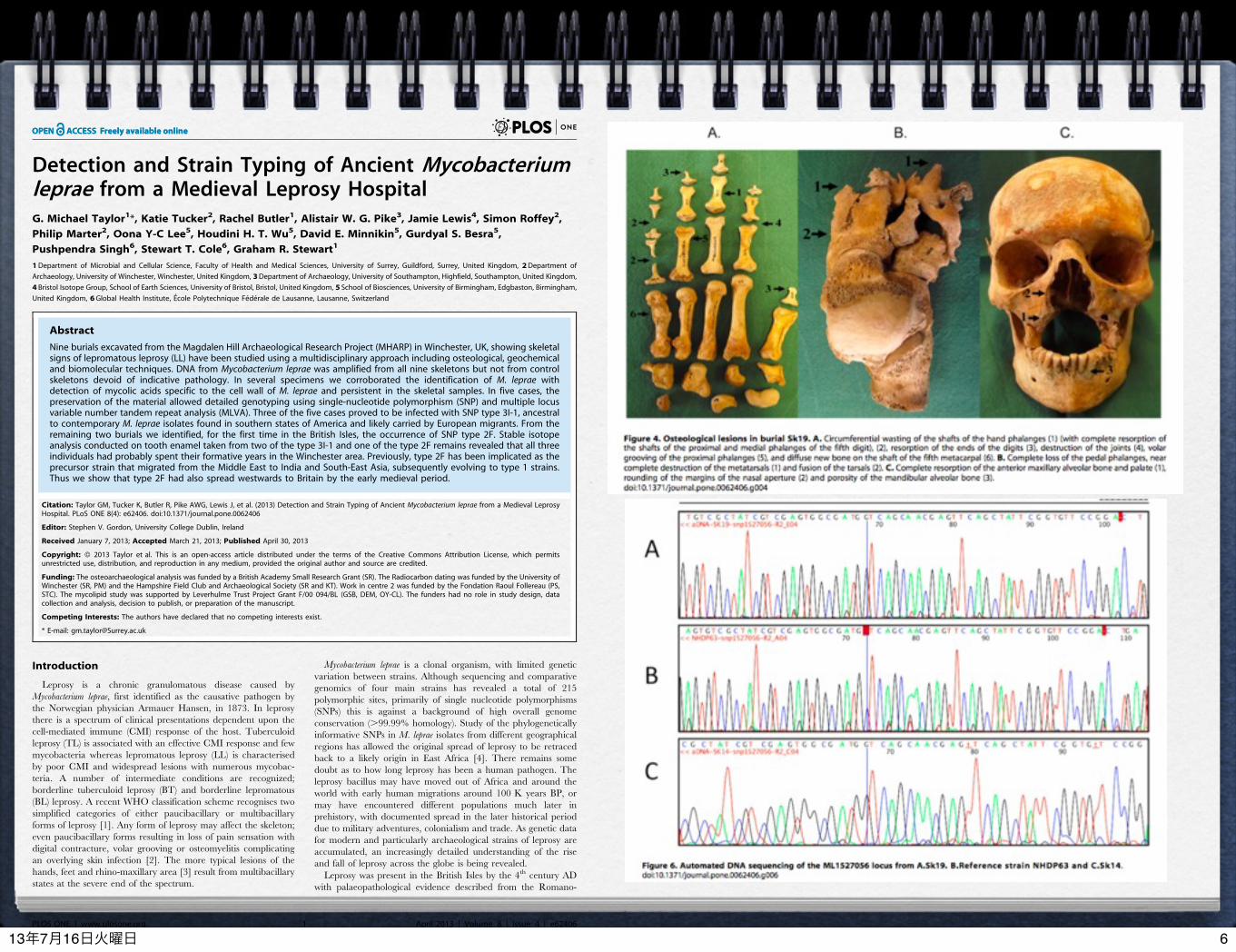
Detection and Strain Typing of Ancient Mycobacteriumleprae from a Medieval Leprosy HospitalG. Michael Taylor1*, Katie Tucker2, Rachel Butler1, Alistair W. G. Pike3, Jamie Lewis4, Simon Roffey2,
Philip Marter2, Oona Y-C Lee5, Houdini H. T. Wu5, David E. Minnikin5, Gurdyal S. Besra5,
Pushpendra Singh6, Stewart T. Cole6, Graham R. Stewart1
1Department of Microbial and Cellular Science, Faculty of Health and Medical Sciences, University of Surrey, Guildford, Surrey, United Kingdom, 2Department of
Archaeology, University of Winchester, Winchester, United Kingdom, 3Department of Archaeology, University of Southampton, Highfield, Southampton, United Kingdom,
4 Bristol Isotope Group, School of Earth Sciences, University of Bristol, Bristol, United Kingdom, 5 School of Biosciences, University of Birmingham, Edgbaston, Birmingham,
United Kingdom, 6Global Health Institute, Ecole Polytechnique Federale de Lausanne, Lausanne, Switzerland
Abstract
Nine burials excavated from the Magdalen Hill Archaeological Research Project (MHARP) in Winchester, UK, showing skeletalsigns of lepromatous leprosy (LL) have been studied using a multidisciplinary approach including osteological, geochemicaland biomolecular techniques. DNA from Mycobacterium leprae was amplified from all nine skeletons but not from controlskeletons devoid of indicative pathology. In several specimens we corroborated the identification of M. leprae withdetection of mycolic acids specific to the cell wall of M. leprae and persistent in the skeletal samples. In five cases, thepreservation of the material allowed detailed genotyping using single-nucleotide polymorphism (SNP) and multiple locusvariable number tandem repeat analysis (MLVA). Three of the five cases proved to be infected with SNP type 3I-1, ancestralto contemporary M. leprae isolates found in southern states of America and likely carried by European migrants. From theremaining two burials we identified, for the first time in the British Isles, the occurrence of SNP type 2F. Stable isotopeanalysis conducted on tooth enamel taken from two of the type 3I-1 and one of the type 2F remains revealed that all threeindividuals had probably spent their formative years in the Winchester area. Previously, type 2F has been implicated as theprecursor strain that migrated from the Middle East to India and South-East Asia, subsequently evolving to type 1 strains.Thus we show that type 2F had also spread westwards to Britain by the early medieval period.
Citation: Taylor GM, Tucker K, Butler R, Pike AWG, Lewis J, et al. (2013) Detection and Strain Typing of Ancient Mycobacterium leprae from a Medieval LeprosyHospital. PLoS ONE 8(4): e62406. doi:10.1371/journal.pone.0062406
Editor: Stephen V. Gordon, University College Dublin, Ireland
Received January 7, 2013; Accepted March 21, 2013; Published April 30, 2013
Copyright: ! 2013 Taylor et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: The osteoarchaeological analysis was funded by a British Academy Small Research Grant (SR). The Radiocarbon dating was funded by the University ofWinchester (SR, PM) and the Hampshire Field Club and Archaeological Society (SR and KT). Work in centre 2 was funded by the Fondation Raoul Follereau (PS,STC). The mycolipid study was supported by Leverhulme Trust Project Grant F/00 094/BL (GSB, DEM, OY-CL). The funders had no role in study design, datacollection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
Introduction
Leprosy is a chronic granulomatous disease caused byMycobacterium leprae, first identified as the causative pathogen bythe Norwegian physician Armauer Hansen, in 1873. In leprosythere is a spectrum of clinical presentations dependent upon thecell-mediated immune (CMI) response of the host. Tuberculoidleprosy (TL) is associated with an effective CMI response and fewmycobacteria whereas lepromatous leprosy (LL) is characterisedby poor CMI and widespread lesions with numerous mycobac-teria. A number of intermediate conditions are recognized;borderline tuberculoid leprosy (BT) and borderline lepromatous(BL) leprosy. A recent WHO classification scheme recognises twosimplified categories of either paucibacillary or multibacillaryforms of leprosy [1]. Any form of leprosy may affect the skeleton;even paucibacillary forms resulting in loss of pain sensation withdigital contracture, volar grooving or osteomyelitis complicatingan overlying skin infection [2]. The more typical lesions of thehands, feet and rhino-maxillary area [3] result from multibacillarystates at the severe end of the spectrum.
Mycobacterium leprae is a clonal organism, with limited geneticvariation between strains. Although sequencing and comparativegenomics of four main strains has revealed a total of 215polymorphic sites, primarily of single nucleotide polymorphisms(SNPs) this is against a background of high overall genomeconservation (.99.99% homology). Study of the phylogeneticallyinformative SNPs in M. leprae isolates from different geographicalregions has allowed the original spread of leprosy to be retracedback to a likely origin in East Africa [4]. There remains somedoubt as to how long leprosy has been a human pathogen. Theleprosy bacillus may have moved out of Africa and around theworld with early human migrations around 100 K years BP, ormay have encountered different populations much later inprehistory, with documented spread in the later historical perioddue to military adventures, colonialism and trade. As genetic datafor modern and particularly archaeological strains of leprosy areaccumulated, an increasingly detailed understanding of the riseand fall of leprosy across the globe is being revealed.Leprosy was present in the British Isles by the 4th century AD
with palaeopathological evidence described from the Romano-
PLOS ONE | www.plosone.org 1 April 2013 | Volume 8 | Issue 4 | e62406
613年7月16日火曜日
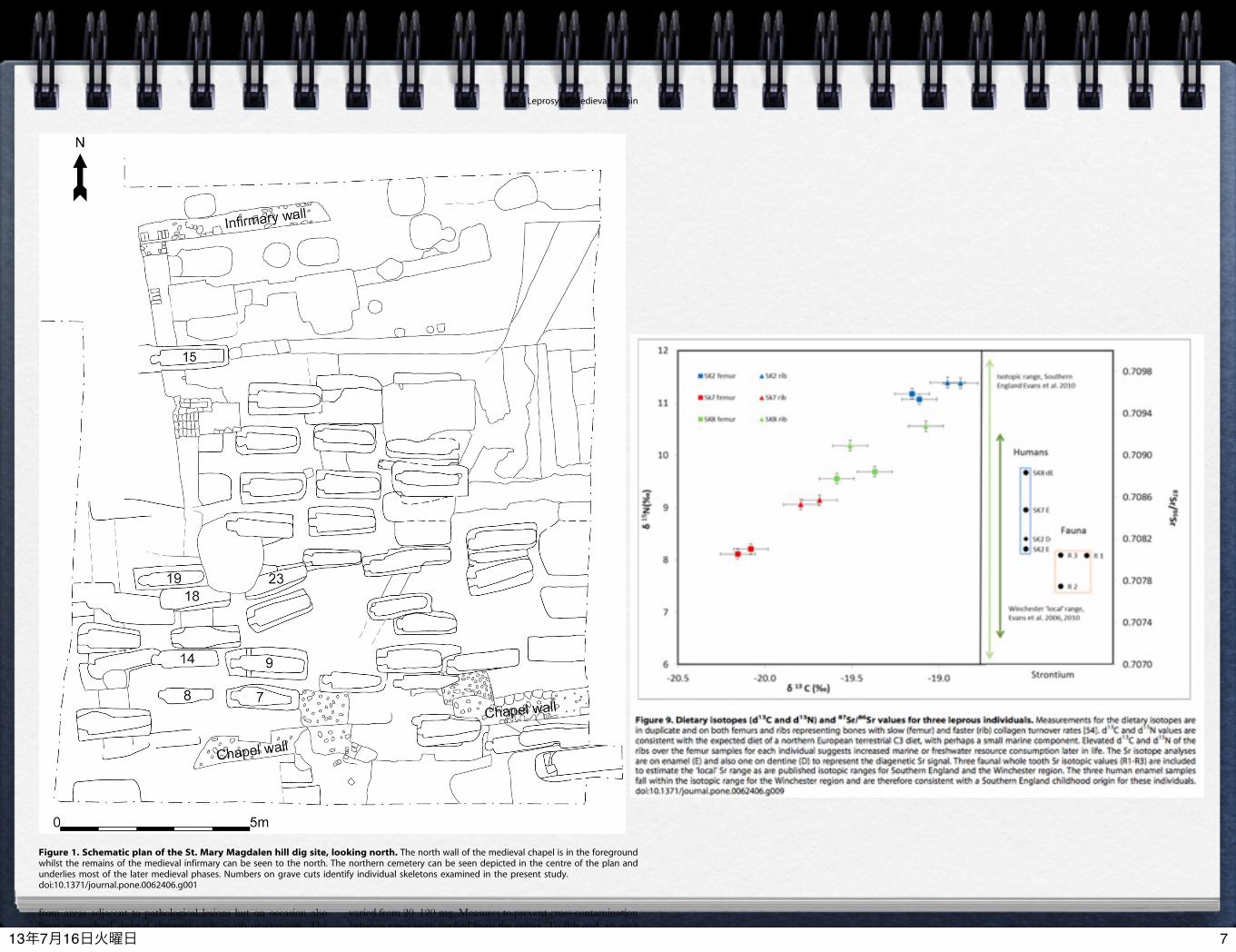
from areas adjacent to pathological lesions but on occasion alsofrom uninvolved skeletal elements such as rib or cranium. Theweights of bone fragments, roughly cuboidal (6–8 mm) in shape,
varied from 20–120 mg. Measures to prevent cross-contaminationbetween cases were applied from the outset. To this end, an areaof bench was set aside and cleaned before and between cases.
Figure 1. Schematic plan of the St. Mary Magdalen hill dig site, looking north. The north wall of the medieval chapel is in the foregroundwhilst the remains of the medieval infirmary can be seen to the north. The northern cemetery can be seen depicted in the centre of the plan andunderlies most of the later medieval phases. Numbers on grave cuts identify individual skeletons examined in the present study.doi:10.1371/journal.pone.0062406.g001
Leprosy in Medieval Britain
PLOS ONE | www.plosone.org 3 April 2013 | Volume 8 | Issue 4 | e62406
713年7月16日火曜日

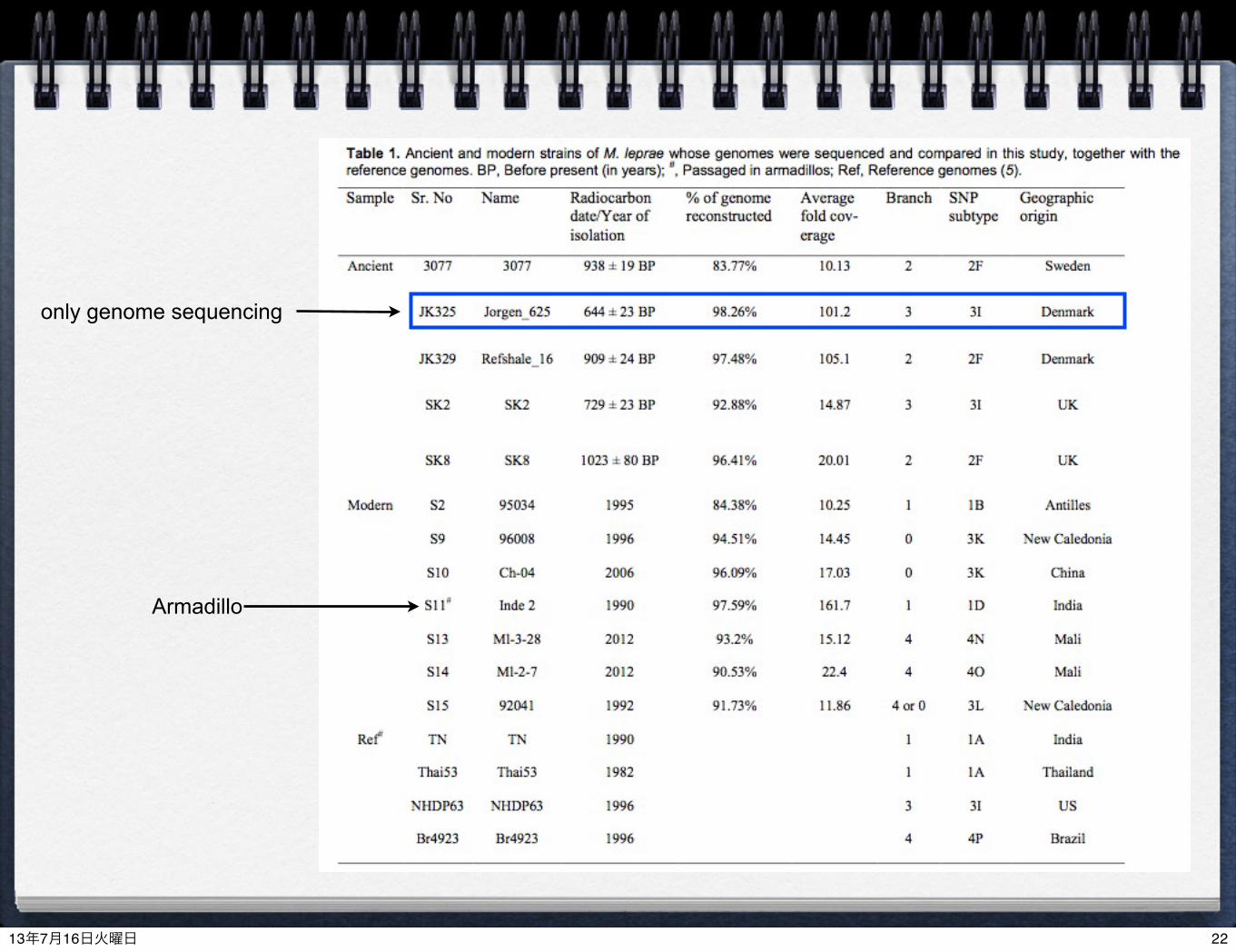
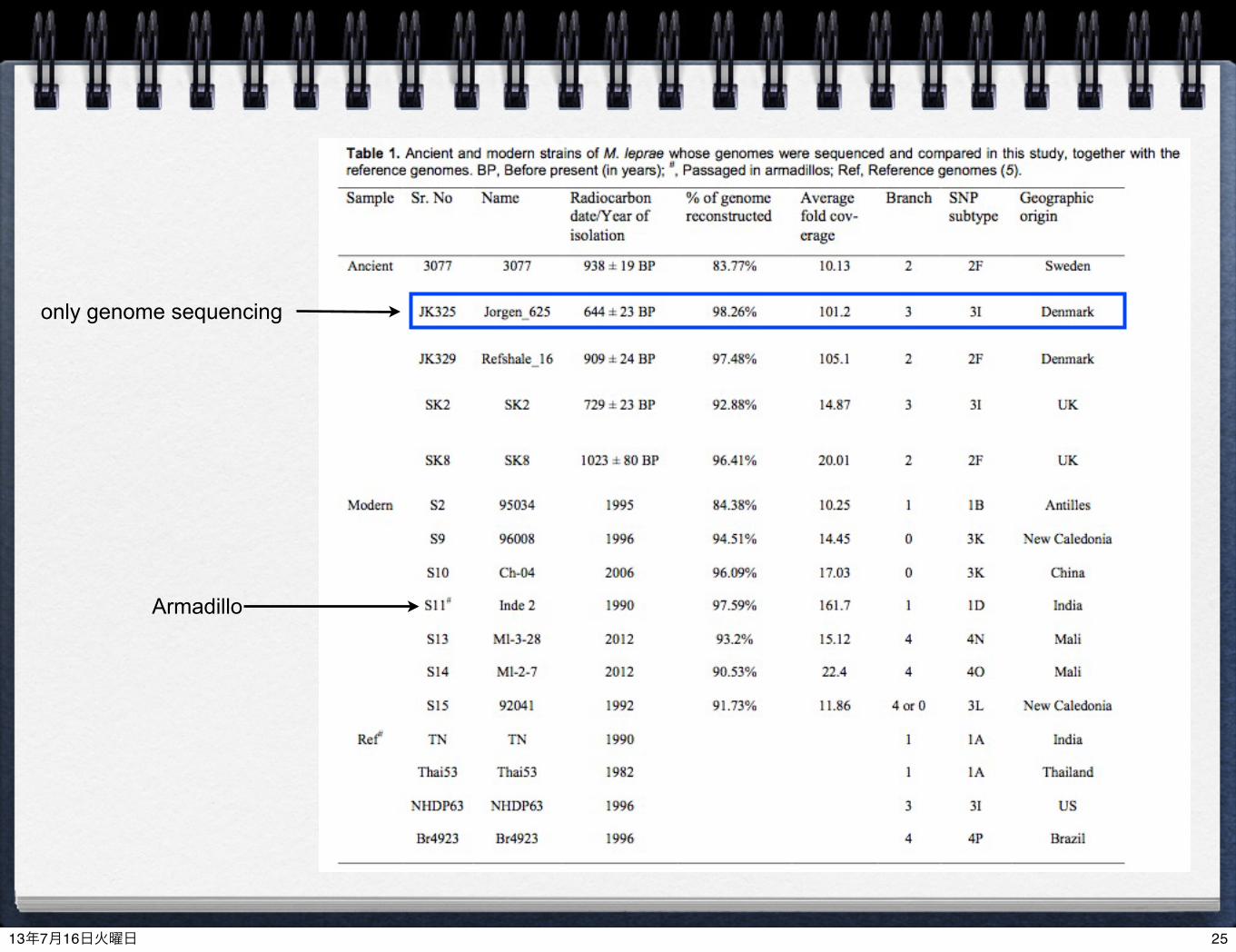
OverviewSamples:
Ancient: UK, Sweden and Denmark in the Middle Ages (11~17 Centuries)
Modern strains (1982~2012)
Methods: Illumina MiSeq/HiSeq/GAIIx (次世代シーケンサーを使った解析)
Two capture methods (target enrichment) for ancient DNA(1) Beads capture for screening(3) DNA array capture to cover whole genome of M. laprae
constructed in the Black Death project in 2011 (Bos, Schuenemann et al. 2011, Nature)(Schuenemann et al. 2011, PNAS)
Modern strains (S7, S9, S12 and S15)(2) Shotgun sequencing (re-sequencing or de novo assembly)
5 ancient strains (10th-14th century)12 modern strains (including 1 armadillo)
Comparative genomics (比較ゲノム解析)
(not sequenced) + a few published genomesPhylogenetic analysis (系統解析)
813年7月16日火曜日

27 samples from 24 skeletons (tooth and bones), with osteological lesions
(1) Screened for the M. leprae DNAby Bead Capture for three genomic loci
DNA extraction in five independent laboratories
(Stockholm, Surrey, Lausanne, Kiel, Tübingen)
NGS sequencing MiSeq (HiSeq) in Tübingen(2) shotgun sequencing (genome assembly or re-sequencing) for 5 ancient samples and 11 modern samples(3) Array-based enrichment for 5 samples showing low DNA yields
913年7月16日火曜日

27 samples from 24 skeletons (tooth and bones), with osteological lesions
(1) Screened for the M. leprae DNAby Bead Capture for three genomic loci
DNA extraction in five independent laboratories
(Stockholm, Surrey, Lausanne, Kiel, Tübingen)
NGS sequencing MiSeq (HiSeq) in Tübingen(2) shotgun sequencing (genome assembly or re-sequencing) for 5 ancient samples and 11 modern samples(3) Array-based enrichment for 5 samples showing low DNA yields
without target enrichment (DNA rich)
913年7月16日火曜日

Methods to identify SNPs
1013年7月16日火曜日
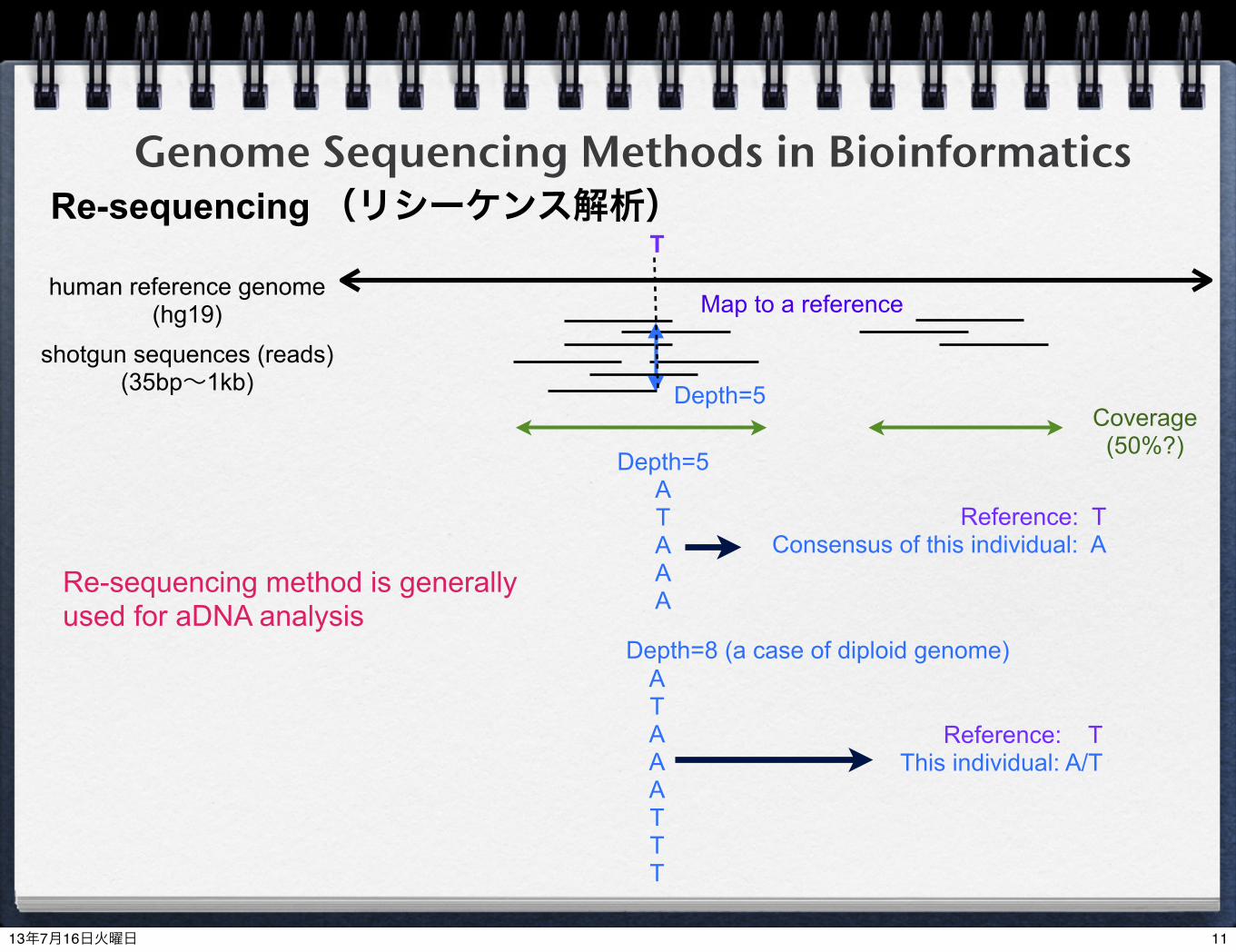
Genome Sequencing Methods in BioinformaticsRe-sequencing (リシーケンス解析)human reference genome
(hg19)shotgun sequences (reads)
(35bp~1kb)
Map to a reference
Depth=5
T
ATAAATTT
Depth=8 (a case of diploid genome)
Reference: TThis individual: A/T
ATAAA
Reference: TConsensus of this individual: A
Depth=5
Coverage(50%?)
Re-sequencing method is generally used for aDNA analysis
1113年7月16日火曜日
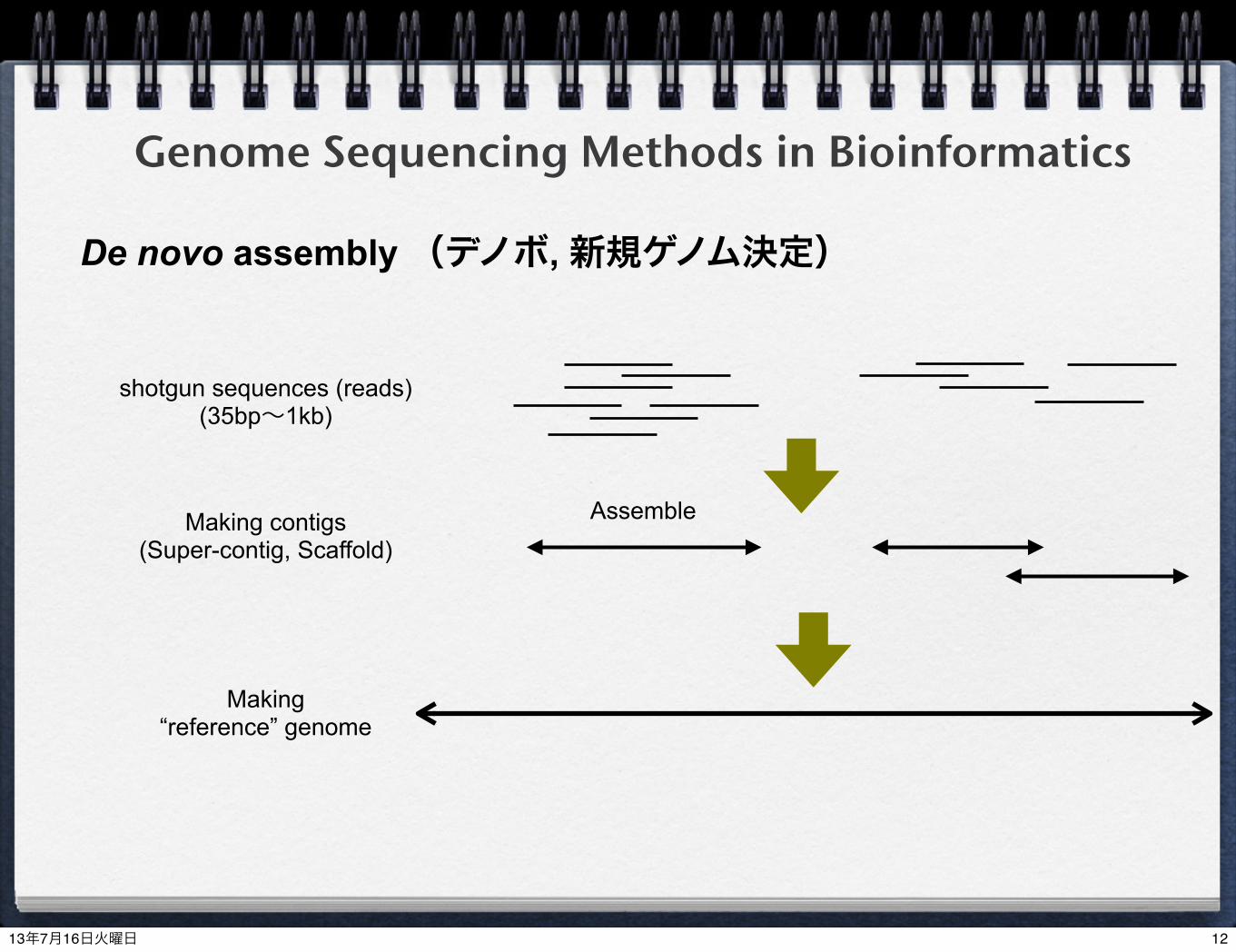
Genome Sequencing Methods in Bioinformatics
De novo assembly (デノボ, 新規ゲノム決定)
shotgun sequences (reads)(35bp~1kb)
Making “reference” genome
Making contigs(Super-contig, Scaffold)
Assemble
1213年7月16日火曜日
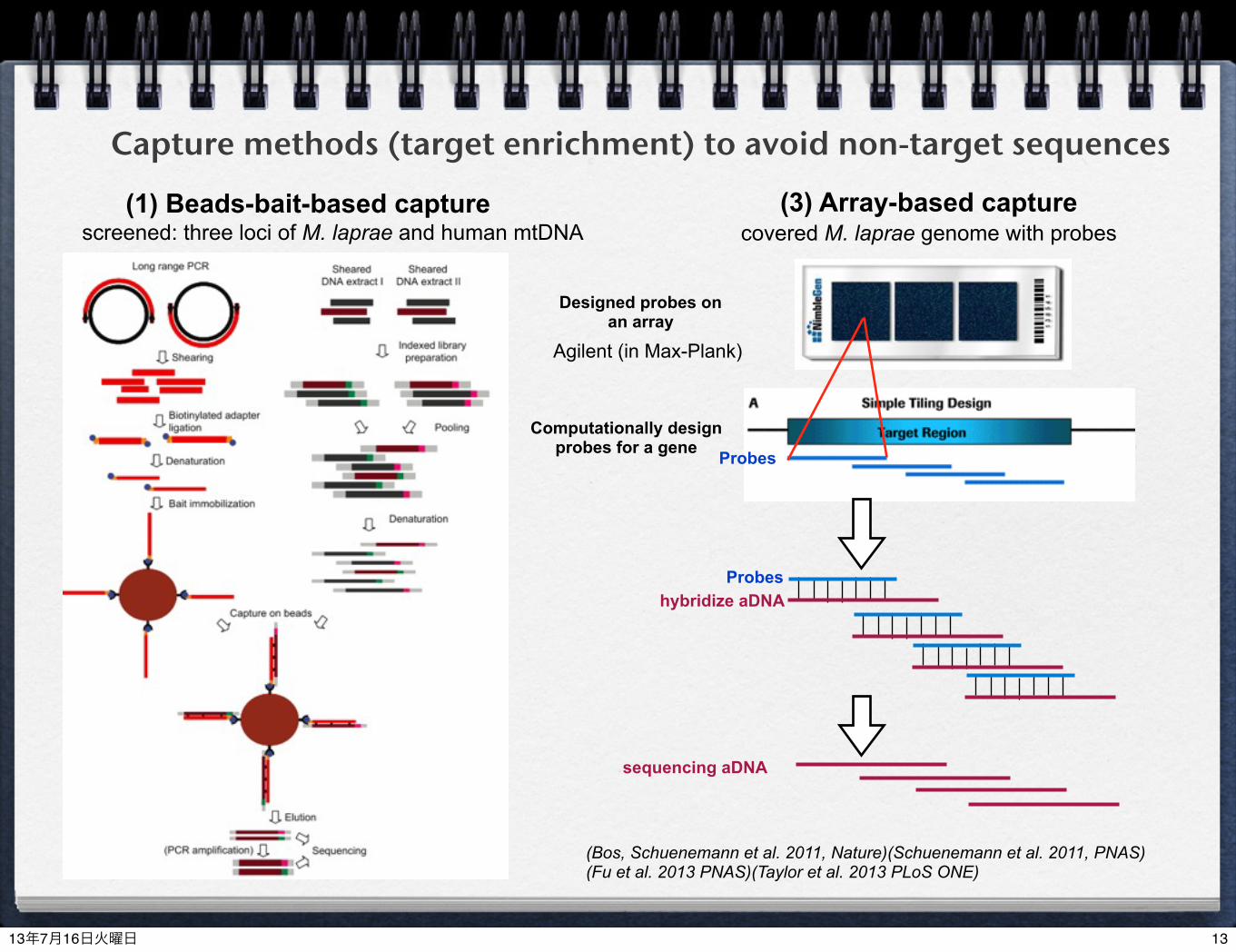
Capture methods (target enrichment) to avoid non-target sequences(1) Beads-bait-based capture (3) Array-based capture
Computationally design probes for a gene
Probeshybridize aDNA
Designed probes on an array
(Bos, Schuenemann et al. 2011, Nature)(Schuenemann et al. 2011, PNAS)(Fu et al. 2013 PNAS)(Taylor et al. 2013 PLoS ONE)
sequencing aDNA
Probes
screened: three loci of M. laprae and human mtDNA covered M. laprae genome with probes
Agilent (in Max-Plank)
1313年7月16日火曜日

27 samples from 24 skeletons (tooth and bones), with osteological lesions
(1) Screened for the M. leprae DNAby Bead Capture for three genomic loci
DNA extraction in five independent laboratories
(Stockholm, Surrey, Lausanne, Kiel, Tübingen)
NGS sequencing MiSeq (HiSeq) in Tübingen(2) shotgun sequencing (genome assembly or re-sequencing) for 5 ancient samples and 11 modern samples(3) Array-based enrichment for 5 samples showing low DNA yields
1413年7月16日火曜日
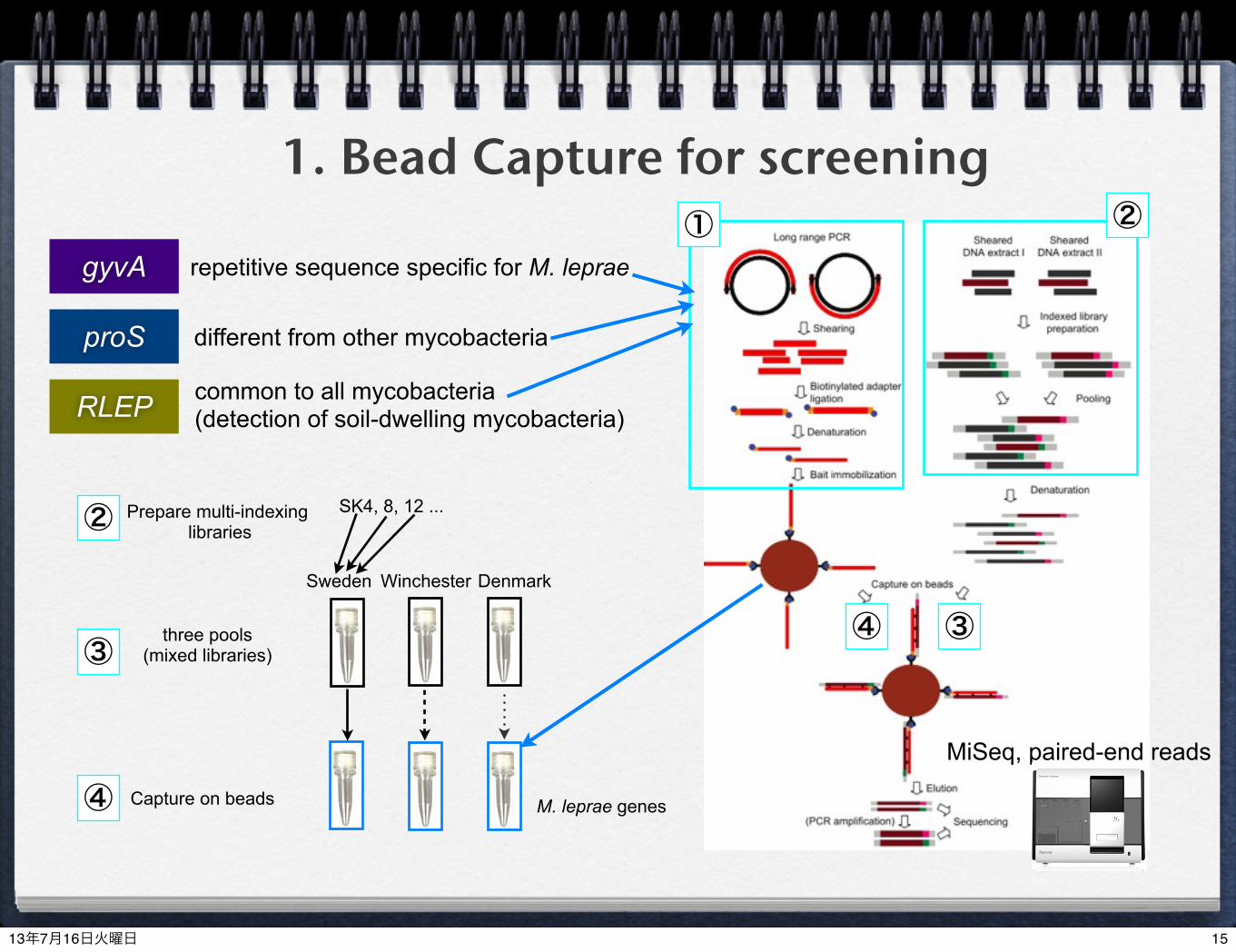
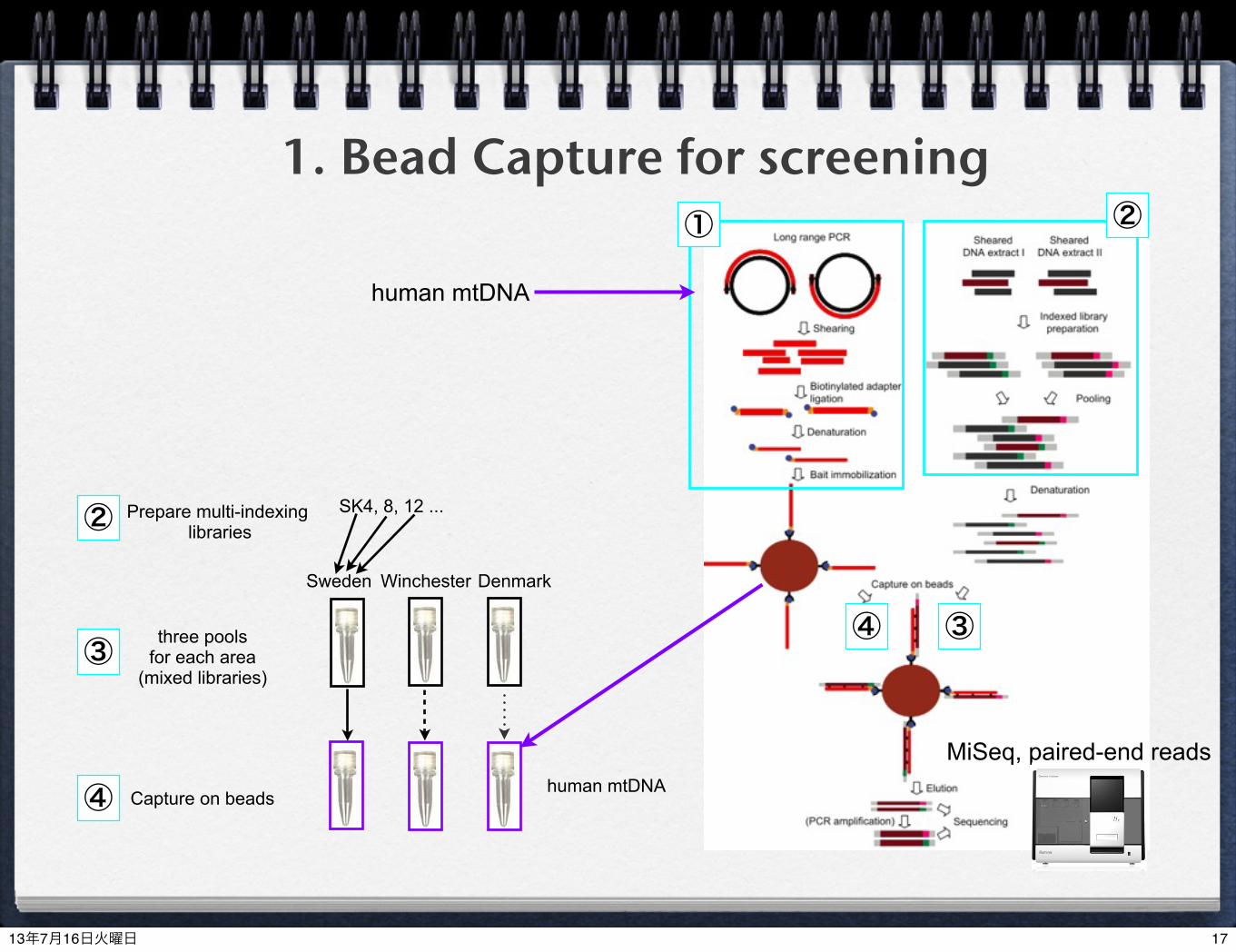
1. Bead Capture for screening
gyvA
RLEP
proS
repetitive sequence specific for M. leprae
different from other mycobacteria
common to all mycobacteria (detection of soil-dwelling mycobacteria)
MiSeq, paired-end reads
M. leprae genes
Sweden Winchester Denmark
Prepare multi-indexing libraries
three pools(mixed libraries)
Capture on beads
SK4, 8, 12 ...
②①
②
③
④
③④
1513年7月16日火曜日
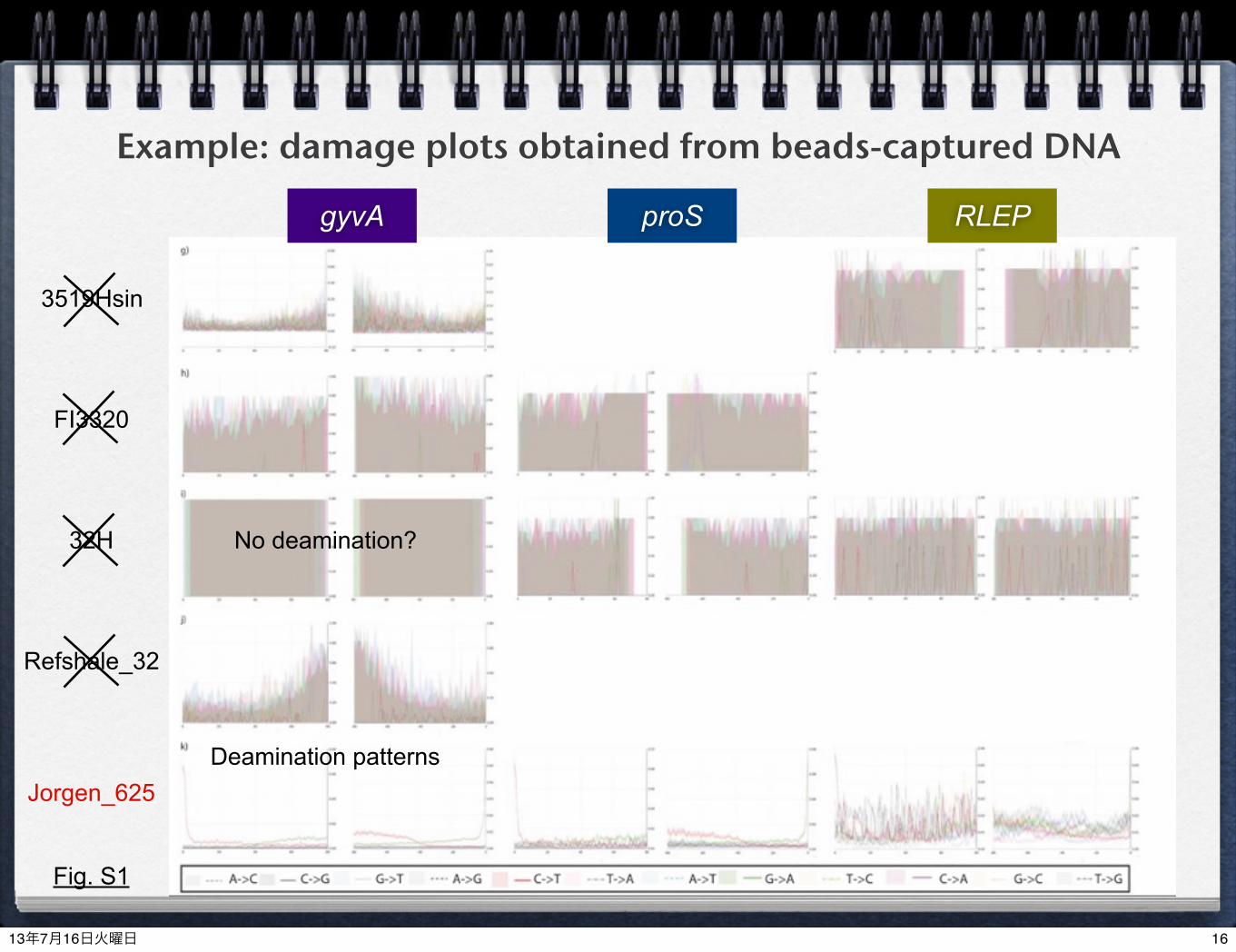
Example: damage plots obtained from beads-captured DNA
gyvA RLEPproS
3519Hsin
FI3320
32H
Refshale_32
Jorgen_625
Fig. S1
Deamination patterns
No deamination?
1613年7月16日火曜日
1. Bead Capture for screening
MiSeq, paired-end reads
Sweden Winchester Denmark
Prepare multi-indexing libraries
three pools for each area
(mixed libraries)
Capture on beads
SK4, 8, 12 ...
②①
②
③
④
③④
human mtDNA
human mtDNA
1713年7月16日火曜日
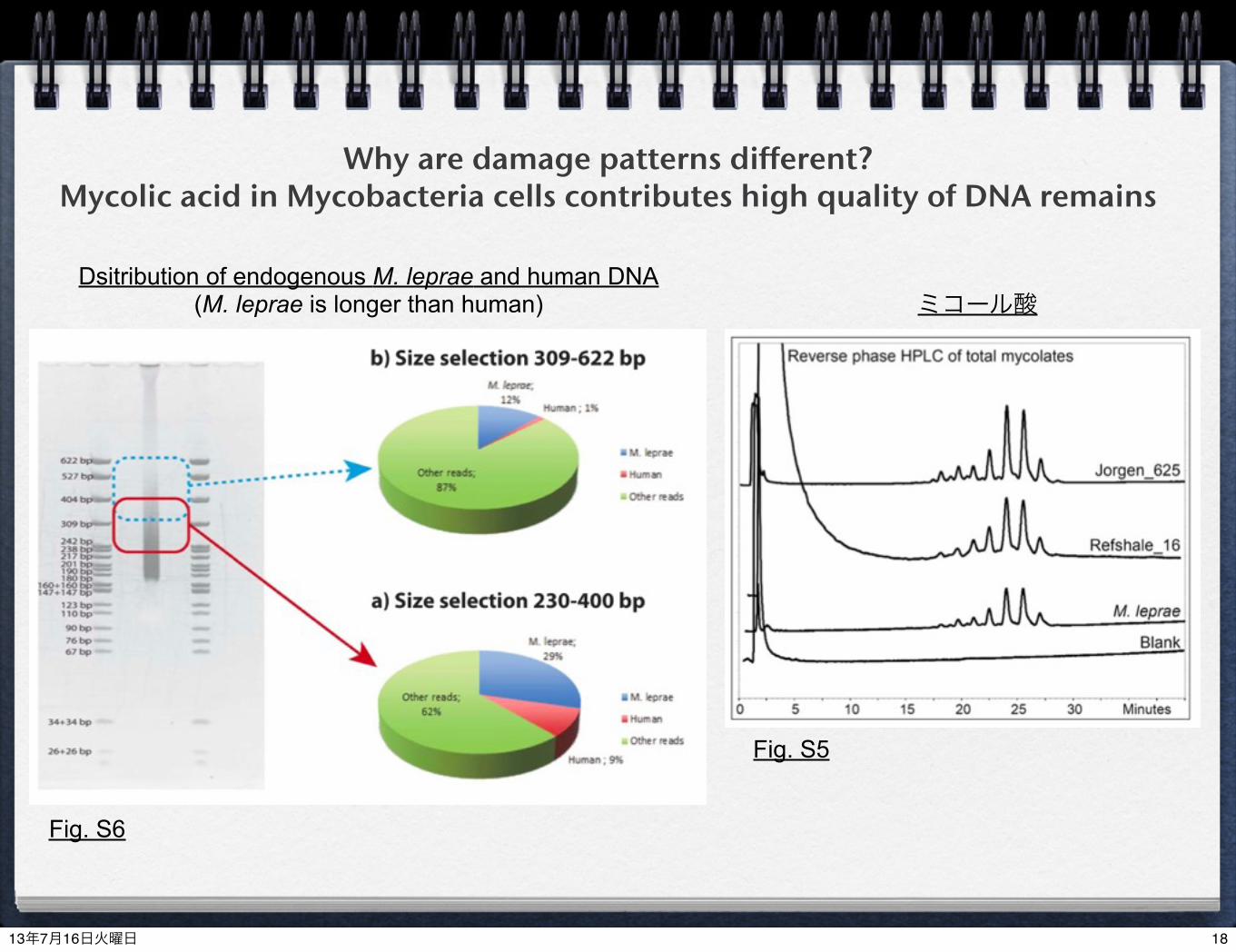
Why are damage patterns different?Mycolic acid in Mycobacteria cells contributes high quality of DNA remains
Fig. S6
Fig. S5
ミコール酸Dsitribution of endogenous M. leprae and human DNA
(M. leprae is longer than human)
1813年7月16日火曜日
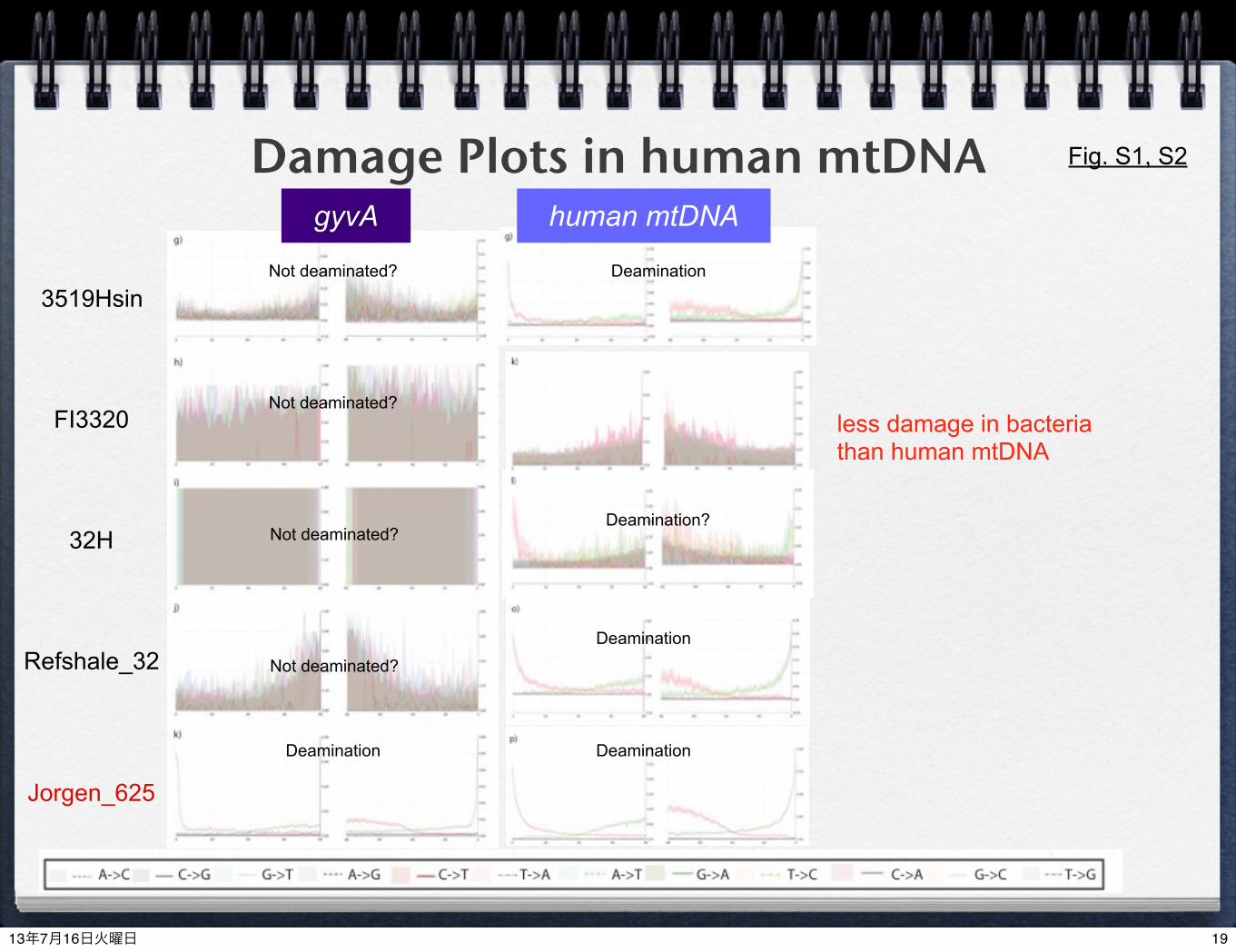
Damage Plots in human mtDNA
3519Hsin
FI3320
32H
Refshale_32
Jorgen_625
human mtDNAgyvA
Fig. S1, S2
Deamination
Deamination
DeaminationDeamination
Not deaminated?
Not deaminated?
Not deaminated?
Not deaminated?
Deamination?
less damage in bacteria than human mtDNA
1913年7月16日火曜日

5 samples passed their criteria (bead capture)10-14th century samples
2013年7月16日火曜日

27 samples from 24 skeletons (tooth and bones), with osteological lesions
(1) Screened for the M. leprae DNAby Bead Capture for three genomic loci
DNA extraction in five independent laboratories
(Stockholm, Surrey, Lausanne, Kiel, Tübingen)
NGS sequencing in Tübingen(2) shotgun sequencing (genome assembly or re-sequencing) for 5 ancient samples and 11 modern samples(3) Array-based enrichment for 5 samples showing low DNA yields
without target enrichment (DNA rich)
2113年7月16日火曜日
only genome sequencing
Armadillo
2213年7月16日火曜日
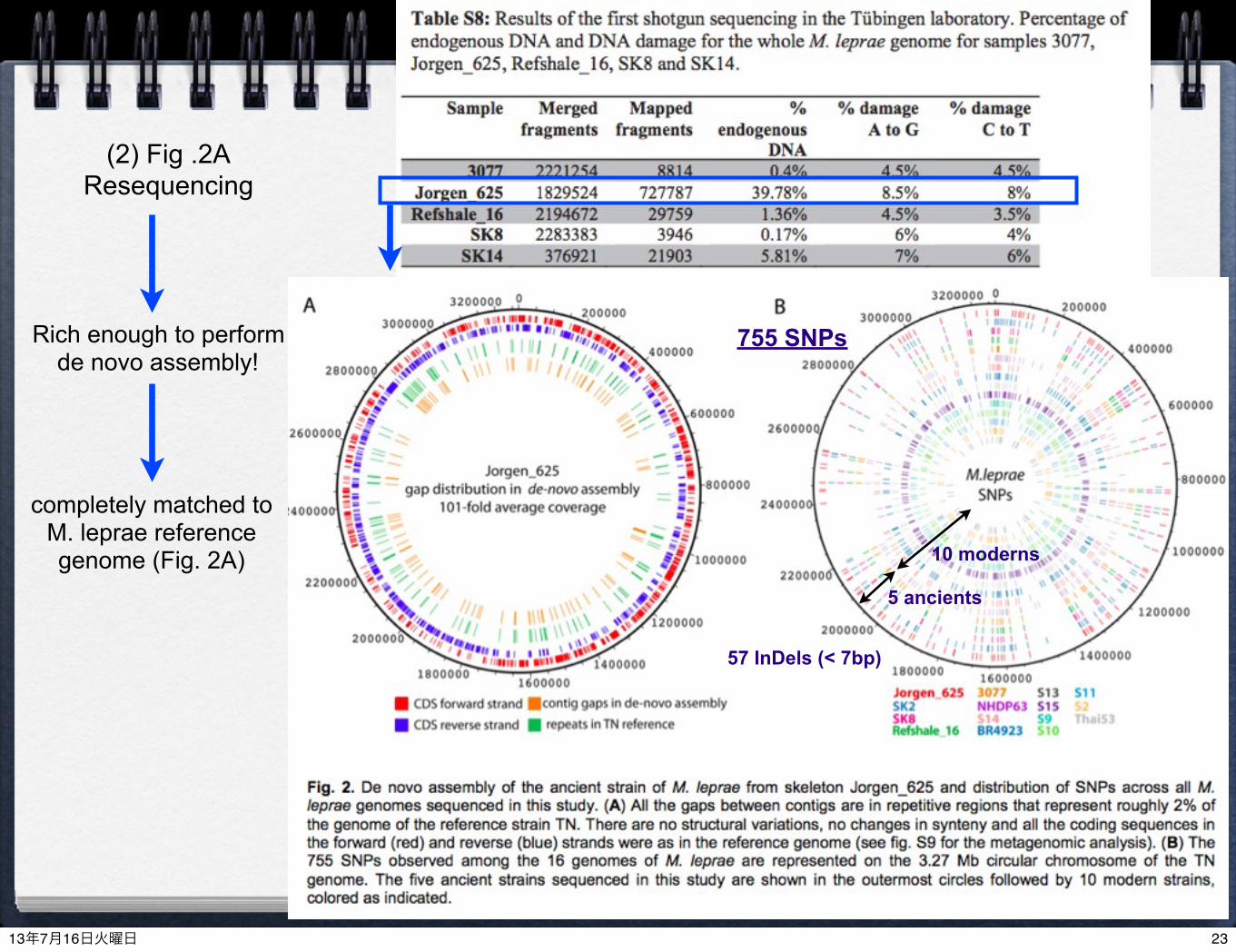
Rich enough to perform de novo assembly!
(2) Fig .2AResequencing
completely matched to M. leprae reference genome (Fig. 2A)
5 ancients
10 moderns
755 SNPs
57 InDels (< 7bp)
2313年7月16日火曜日
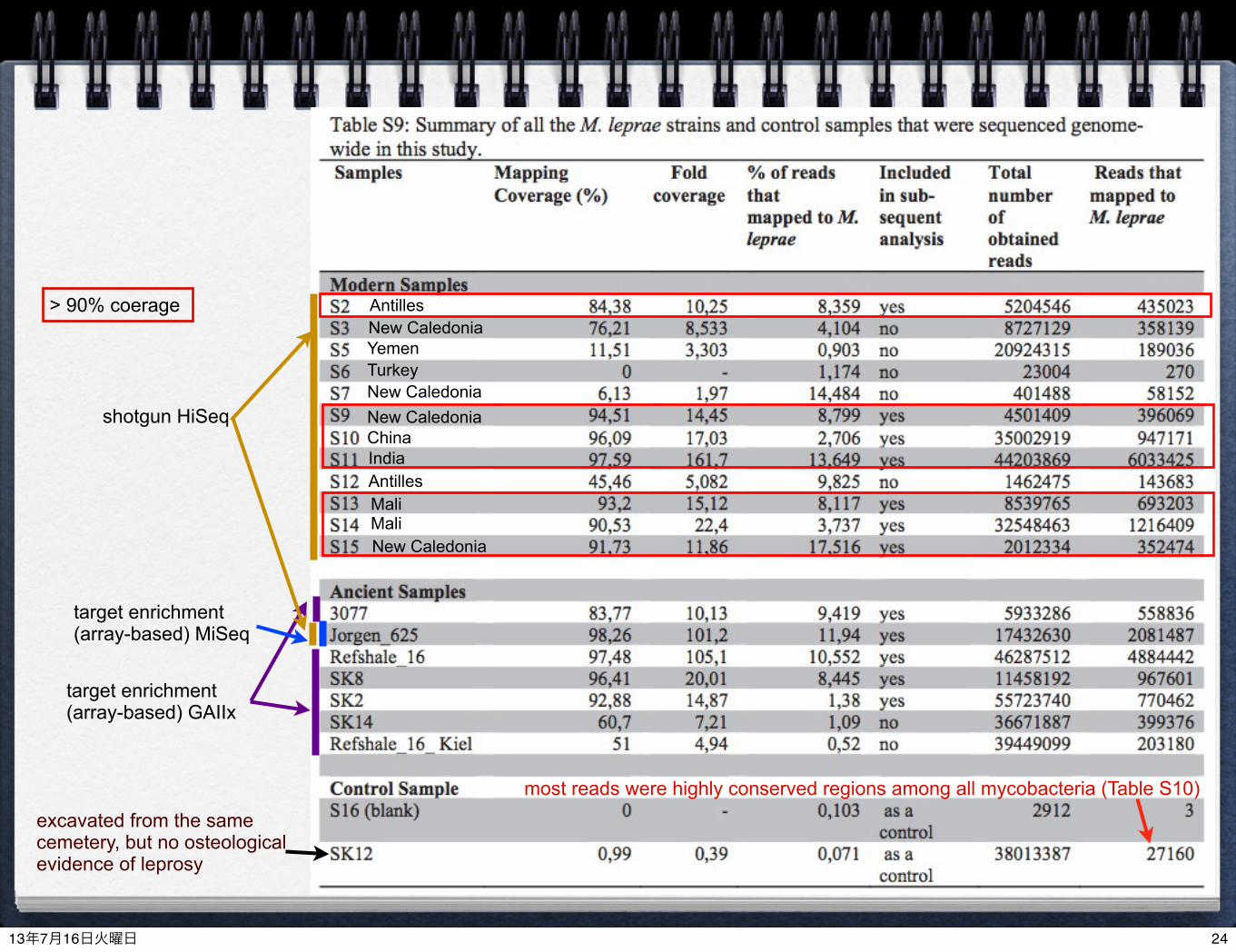
excavated from the same cemetery, but no osteological evidence of leprosy
target enrichment (array-based) GAIIx
shotgun HiSeq
target enrichment (array-based) MiSeq
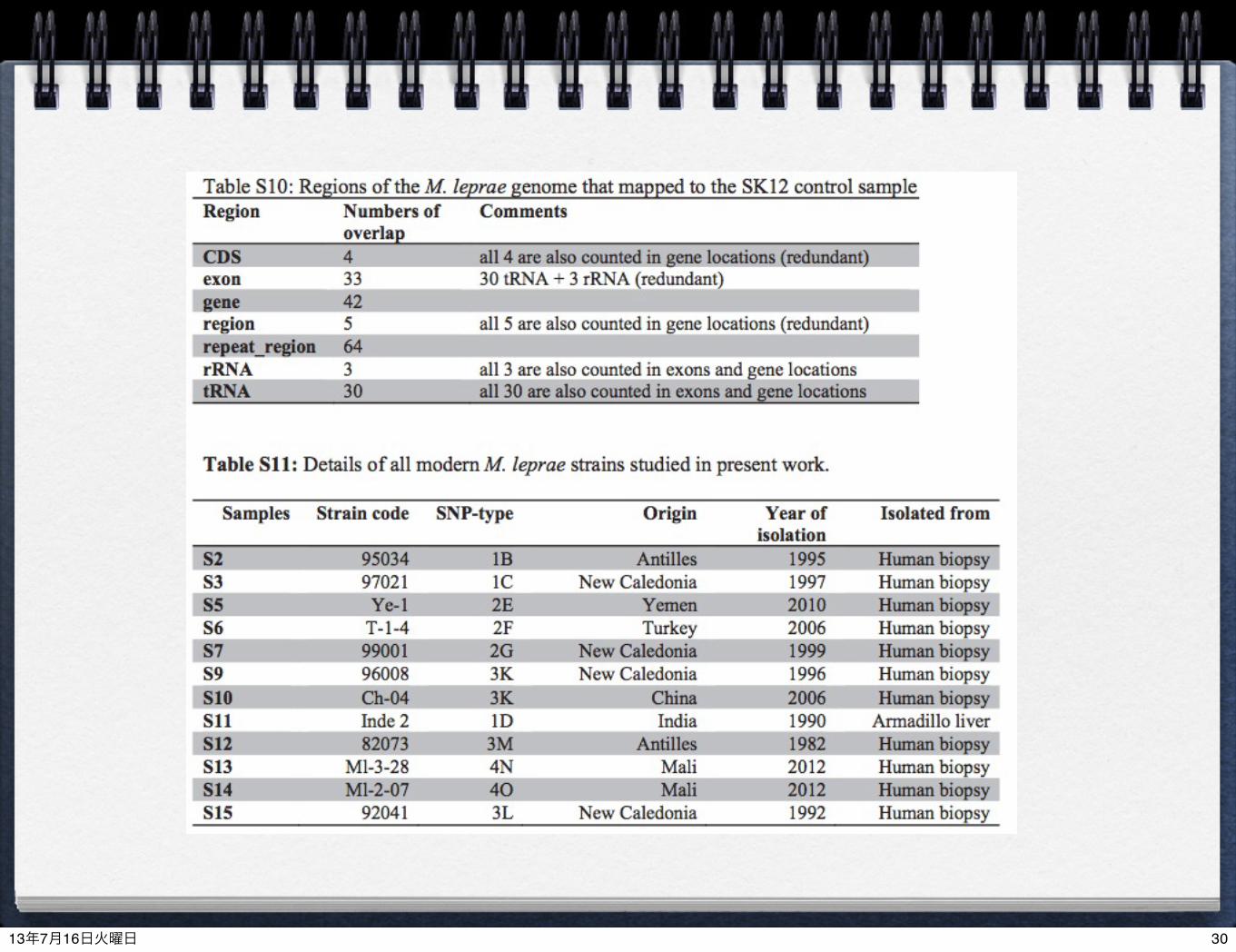
most reads were highly conserved regions among all mycobacteria (Table S10)
AntillesNew CaledoniaYemenTurkeyNew CaledoniaNew CaledoniaChinaIndia
MaliAntilles
MaliNew Caledonia
> 90% coerage
2413年7月16日火曜日
only genome sequencing
Armadillo
2513年7月16日火曜日
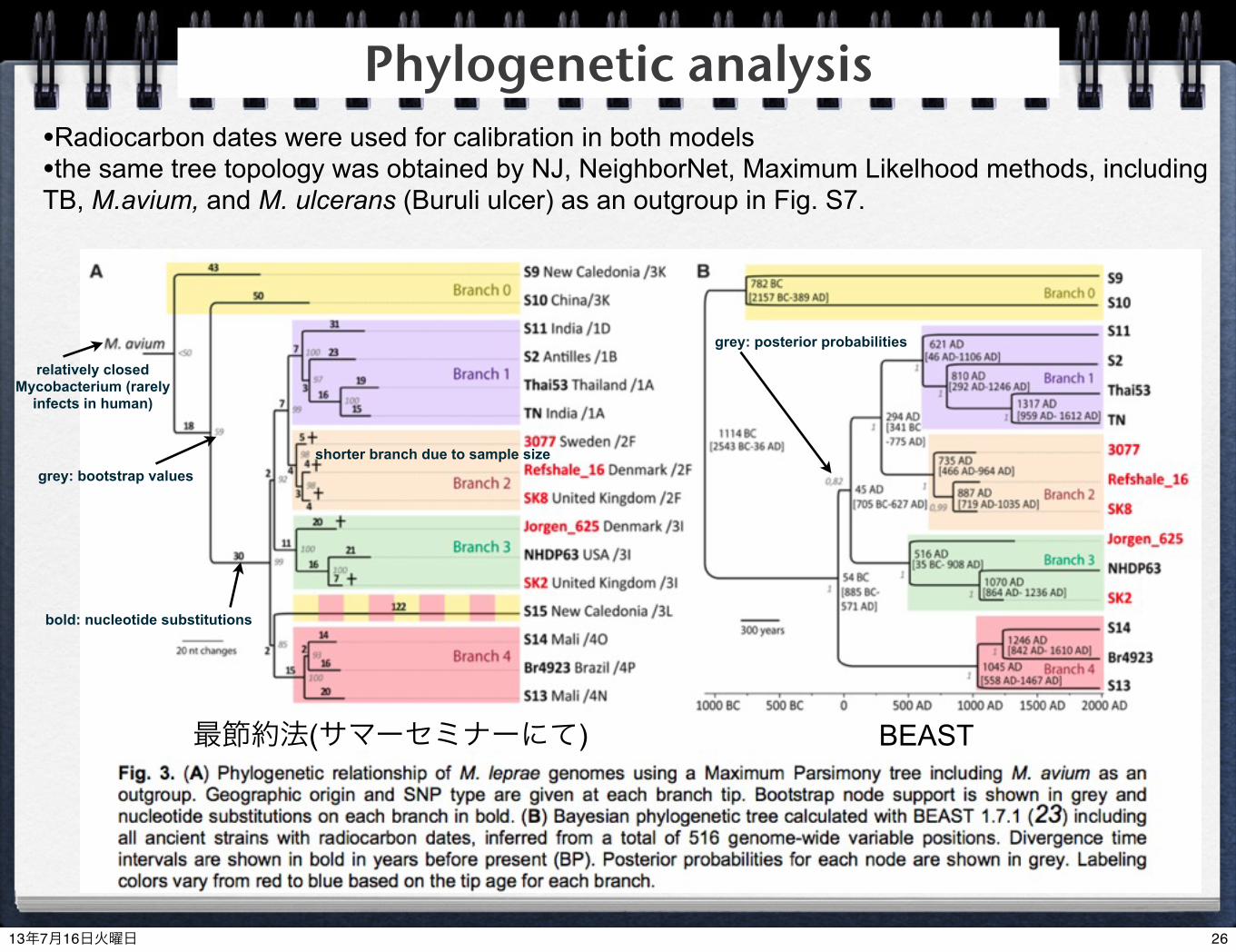
最節約法(サマーセミナーにて) BEAST
shorter branch due to sample sizegrey: bootstrap values
bold: nucleotide substitutions
grey: posterior probabilities
Phylogenetic analysis•Radiocarbon dates were used for calibration in both models•the same tree topology was obtained by NJ, NeighborNet, Maximum Likelhood methods, including TB, M.avium, and M. ulcerans (Buruli ulcer) as an outgroup in Fig. S7.
relatively closed Mycobacterium (rarely
infects in human)
2613年7月16日火曜日
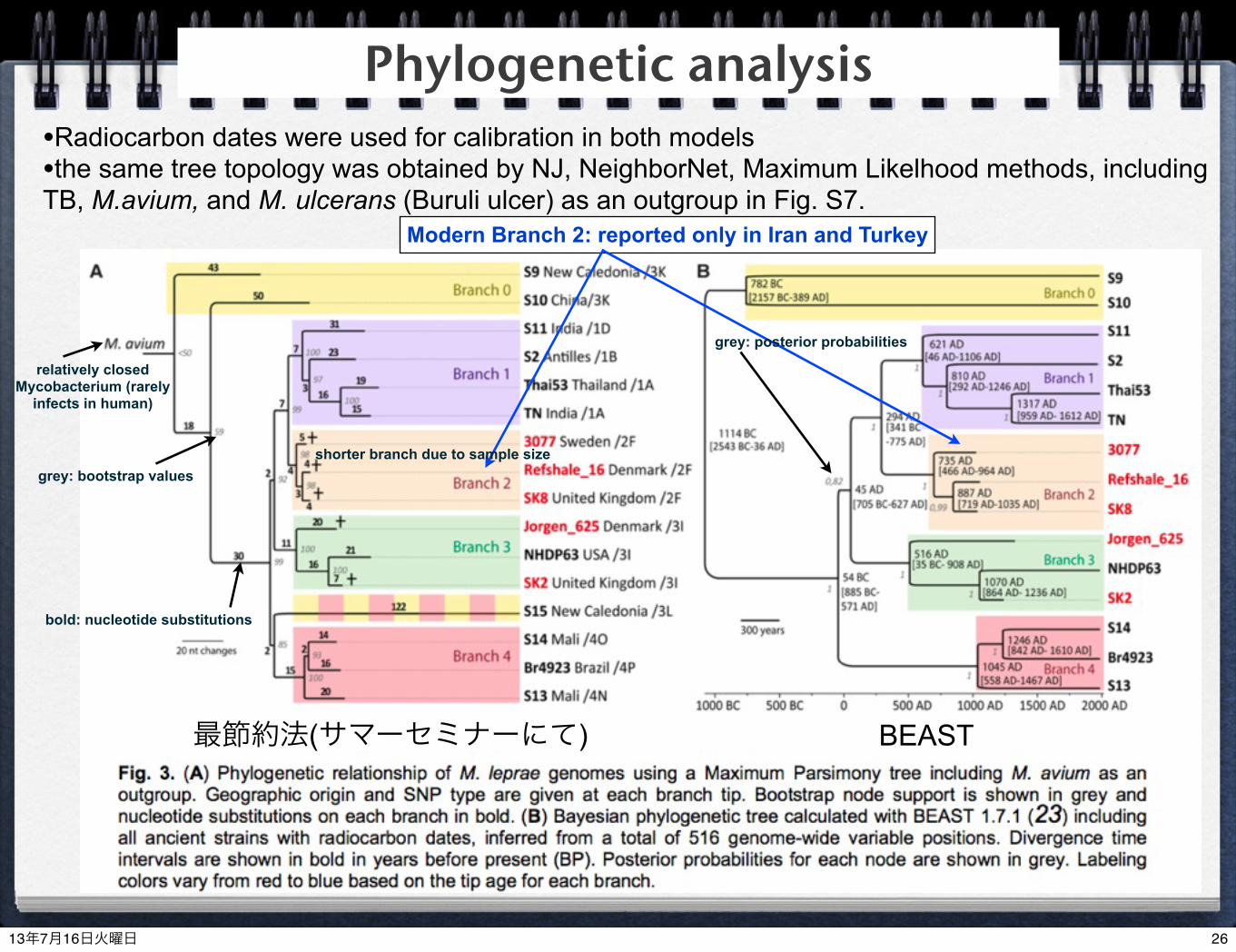
最節約法(サマーセミナーにて) BEAST
Modern Branch 2: reported only in Iran and Turkey
shorter branch due to sample sizegrey: bootstrap values
bold: nucleotide substitutions
grey: posterior probabilities
Phylogenetic analysis•Radiocarbon dates were used for calibration in both models•the same tree topology was obtained by NJ, NeighborNet, Maximum Likelhood methods, including TB, M.avium, and M. ulcerans (Buruli ulcer) as an outgroup in Fig. S7.
relatively closed Mycobacterium (rarely
infects in human)
2613年7月16日火曜日
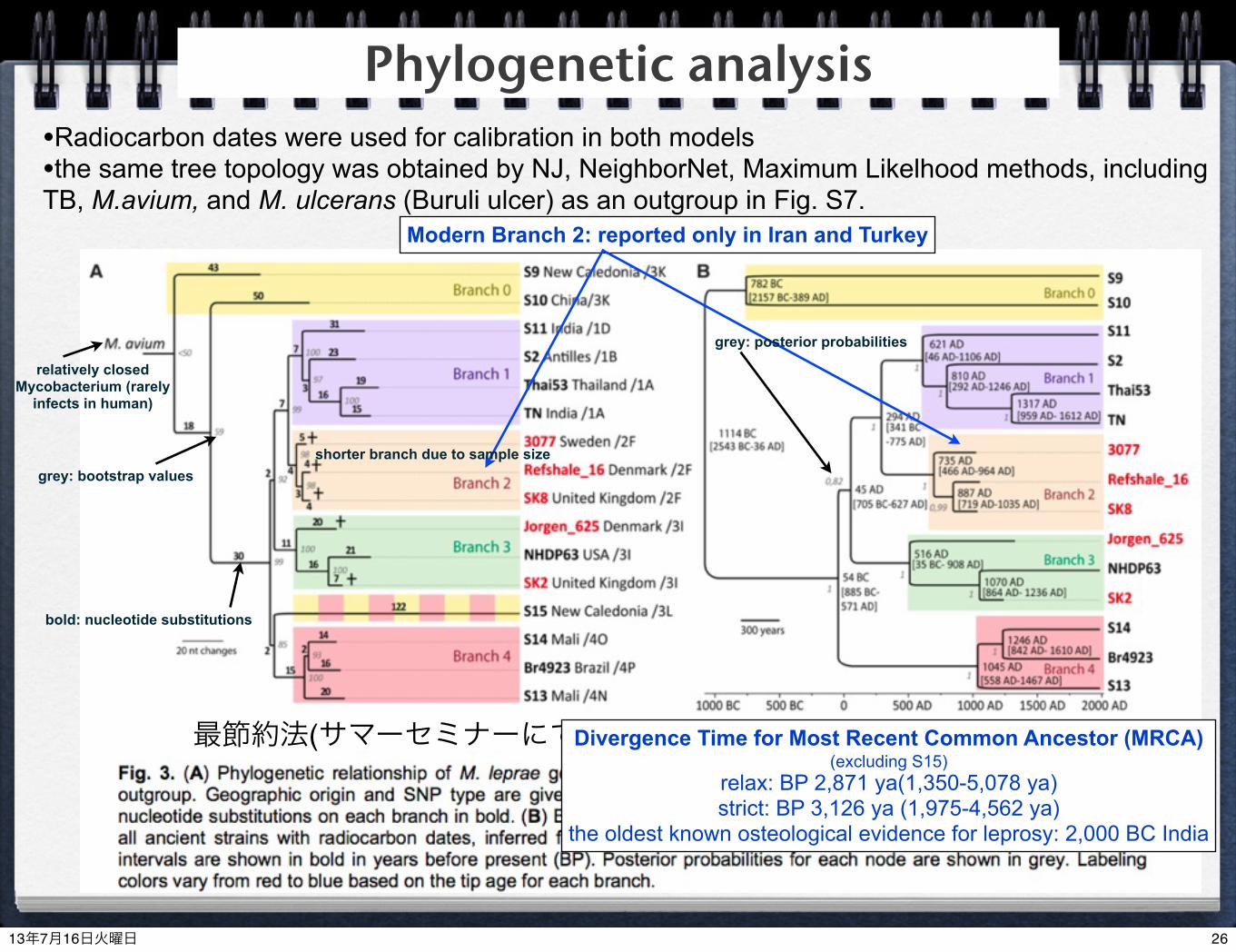
最節約法(サマーセミナーにて) BEAST
Modern Branch 2: reported only in Iran and Turkey
shorter branch due to sample size
Divergence Time for Most Recent Common Ancestor (MRCA) (excluding S15)
relax: BP 2,871 ya(1,350-5,078 ya)strict: BP 3,126 ya (1,975-4,562 ya)
the oldest known osteological evidence for leprosy: 2,000 BC India
grey: bootstrap values
bold: nucleotide substitutions
grey: posterior probabilities
Phylogenetic analysis•Radiocarbon dates were used for calibration in both models•the same tree topology was obtained by NJ, NeighborNet, Maximum Likelhood methods, including TB, M.avium, and M. ulcerans (Buruli ulcer) as an outgroup in Fig. S7.
relatively closed Mycobacterium (rarely
infects in human)
2613年7月16日火曜日
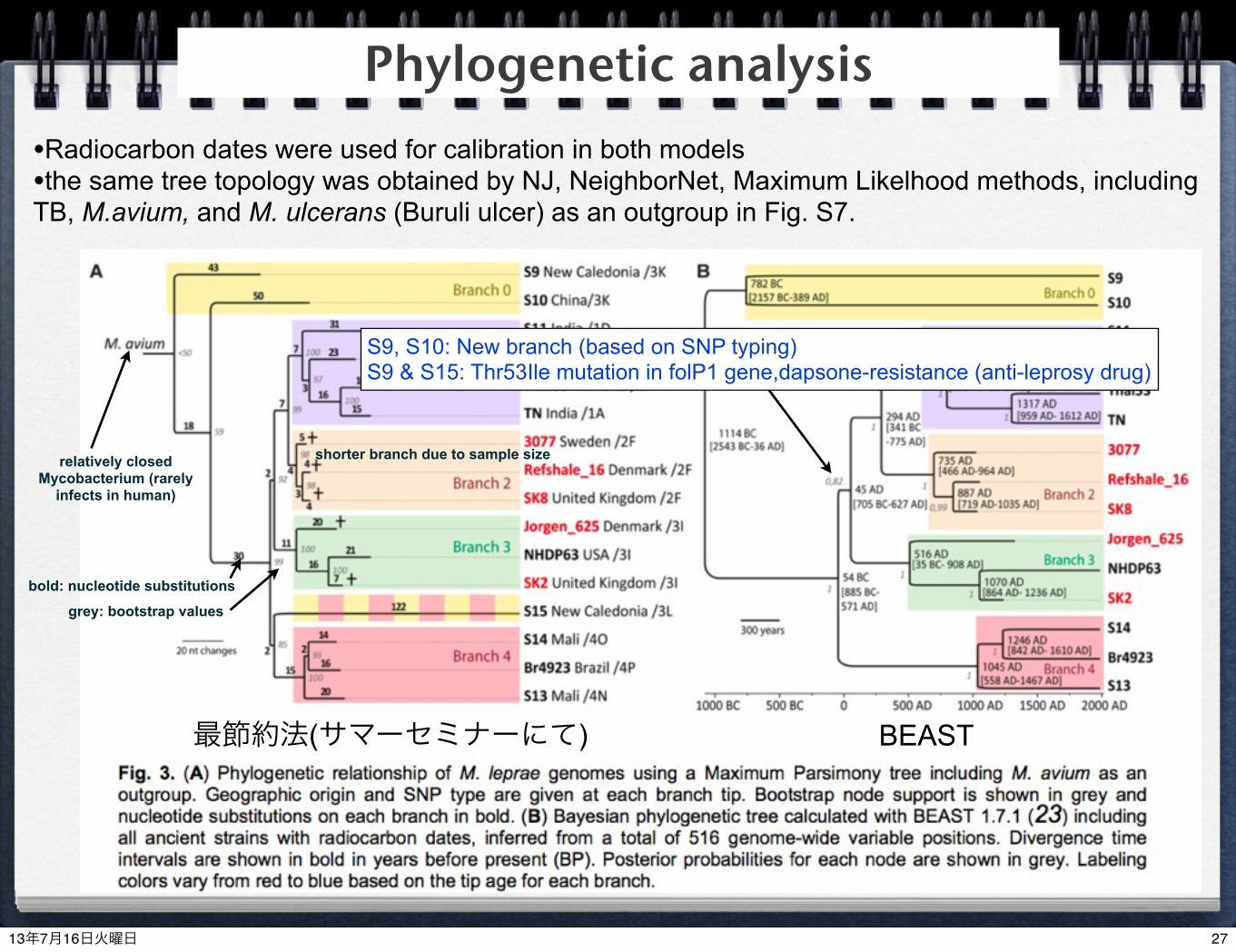
Phylogenetic analysis
最節約法(サマーセミナーにて) BEAST
•Radiocarbon dates were used for calibration in both models•the same tree topology was obtained by NJ, NeighborNet, Maximum Likelhood methods, including TB, M.avium, and M. ulcerans (Buruli ulcer) as an outgroup in Fig. S7.
shorter branch due to sample size
grey: bootstrap values
bold: nucleotide substitutions
grey: posterior probabilities
relatively closed Mycobacterium (rarely
infects in human)
S9, S10: New branch (based on SNP typing)S9 & S15: Thr53Ile mutation in folP1 gene,dapsone-resistance (anti-leprosy drug)
2713年7月16日火曜日

Summaryscreened 27 samples (24 skeletons with osteological evidence) by Bead-bait-capture method
damage patterns (deamination) in M. leprae were different from those in human mtDNAlonger reads were obtained from M. leprae than humanstrong lipid cells in Mycobacteria keep better quality of aDNA
five skeletons and modern strains were sequenced by Array-based capture and shotgun
one (Jorgen_625) included rich endogenous DNA, so that was enable to do de novo assembly755 SNPs among reference, 5 ancient and 10 modern strainsindicates large IN/DELs in 5 ancient strainsThree out of five strains in Europe in the Middle Ages belong to the same branch in modern Middle East (Branch2)Two ancient European strains grouped into Branch 3 with Modern USA
2813年7月16日火曜日
3013年7月16日火曜日





























