Embed Size (px)
Citation preview

Research Article
Geographical diversification of growth–defense strategiesin an invasive plant
Xiao‐Yun PAN* Xin JIA Dong‐Jing FU Bo LI(Coastal Ecosystems Research Station of Yangtze River Estuary, Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering,
Institute of Biodiversity Science, Fudan University, Shanghai 200433, China)
Abstract Interactions between herbivore pressure and resource availability may result in growth–defense trade‐offsin plants (resource availability hypothesis), which promote the potential for ecological diversification and haverecently been proposed as a new mechanism of plant invasion (resource–enemy release hypothesis). In the presentstudy, we tested this idea by comparing patterns of maximum growth rate and antiherbivore defense capacity againstAgasicles hygrophila (a specialist phytophagous insect on Alternanthera philoxeroides) among six morphs ofA. philoxeroides from both native (Argentina) and introduced (US and China) ranges. The results revealed thatherbivore resistance consistently exhibited a significant trade‐off against the growth rate of A. philoxeroides.Compared with native morphs in Argentina, introduced morphs generally exhibited a higher growth rate and lowerdefense. The results demonstrate that the combination of flea beetle pressure and resource availability facilitatesgeographical divergence of A. philoxeroides, and that release from natural specialists and increased resourceavailability may interact to contribute to its successful invasion.Key words Agasicles hygrophila, Alternanthera philoxeroides (alligator weed), growth–defense trade‐off, resourceavailability hypothesis.
Interactions between plants and herbivores haveprofound effects on both ecological and evolutionaryprocesses (Crawley, 1997; Pennings & Silliman, 2005;Fine et al., 2006; Agrawal, 2007) and have led to theevolution of a staggering diversity of plant defensestrategies (Crawley, 1997; Agrawal, 2007). The optimaldefensive investment depends not only on the intensityof herbivory, but also on resource availability in theenvironment.More benign physical environments favorplants with reduced allocation to resistance becausehigh resource availability increases the opportunity costof defensive investment (resource availability hypothe-sis; Coley et al., 1985). Empirical studies have generallysupported this hypothesis, although many exceptionsexist (for a review, see Crawley, 1997). However, thisinteractive effect between enemy pressure and resourceavailability has been largely unappreciated in invasionecology (but see Blumenthal, 2006; Zhang & Jiang,2006; Pan et al., 2012).
Recently, Blumenthal (2006) developed an inter-pretive framework suggesting that enemy release andincreased resource availability may act in concertto cause successful invasion among plant species
(resource–enemy release hypothesis, R‐ERH). To thebest of our knowledge, this potentially important hypo-thesis has not been tested formally (Pan et al., 2012).Although the R‐ERH (and the resource availabilityhypothesis) is largely concerned with variations ingrowth–defense strategies among plant species thatdiffer in inherent growth rates, a genetically basedvariation in growth–defense strategies within plantspecies (i.e. ecotypic differentiation) is theoreticallypredicted by the R‐ERH.
In the present study, our approach to testing thisidea arises out of geographical diversification ofgrowth–defense strategies in the invasive exotic plantAlternanthera philoxeroides (Mart.) Griseb. (Amaran-thaceae; alligator weed). We set out to screen sixgeographically isolated morphs for traits that aretheoretically correlated: (i) maximum relative growthrate (RGR); (ii) antiherbivore defense capacity againstAgasicles hygrophila (a specialist phytophagousinsect on A. philoxeroides); and (iii) distribution alonglatitude/resource gradients. Alternanthera philoxer-oides is a perennial herbaceous plant originating fromthe Parana River basin in Argentina that is now one ofthe most abundant and widespread plants in freshwaterecosystems, riparian zones, and floodplain wetlands inwarm regions (Pan et al., 2006a, 2007), where nutrientenrichment due to human waste disposal and intensive
Received: 17 September 2012 Accepted: 9 November 2012* Author for correspondence. E‐mail: [email protected]. Tel./Fax:86‐21‐65642178.
Journal of Systematics and Evolution 51 (3): 308–317 (2013) doi: 10.1111/j.1759-6831.2012.00239.x
© 2012 Institute of Botany, Chinese Academy of Sciences
agricultural practices typically occurs (Pan et al., 2006a;Verhoeven et al., 2006). Alternanthera philoxeroides ishighly polymorphic in its native range. The currenttaxonomic status of this species has been discussed bySosa et al. (2004). Most A. philoxeroides collections innative ranges can be assigned to two major morphs,namely var. acutifolia and var. obtusifolia, but anintermediate morph (var. lancifolia) has also beenreported (Mears, 1977; Jia et al., 2010). In addition toobvious variations in leaf morphology, natural pop-ulations of A. philoxeroides exhibit abundant variationsin stature, phenology, stomatal index, growth rate, andstem morphology (e.g. diameter, tissue density, and thespace occupied by the pith cavity; Pan et al.,2007, 2011). These diversifications may be attributableto the occurrence of different ecotypes along alatitudinal gradient (Jia et al., 2010). Previous studieshave suggested that a small stem diameter and highlycompact stem tissue play a pivotal role in preventingpupation of A. hygrophila (Coulson, 1977; Vogt et al.,1979; Stewart et al., 2000; Ma & Wang, 2004; Panet al., 2011). In addition, it is well known that plant traitssuch as stem tissue density and diameter are stronglycorrelated with RGR (Niklas, 1995). Thus, it isreasonable to speculate that the interactions betweenherbivore pressures of flea beetles and habitat produc-tivity may contribute to the geographical diversificationof this species. In the present study, we assume thatRGR is an appropriate proxy for plant fitness (Grime,1979).
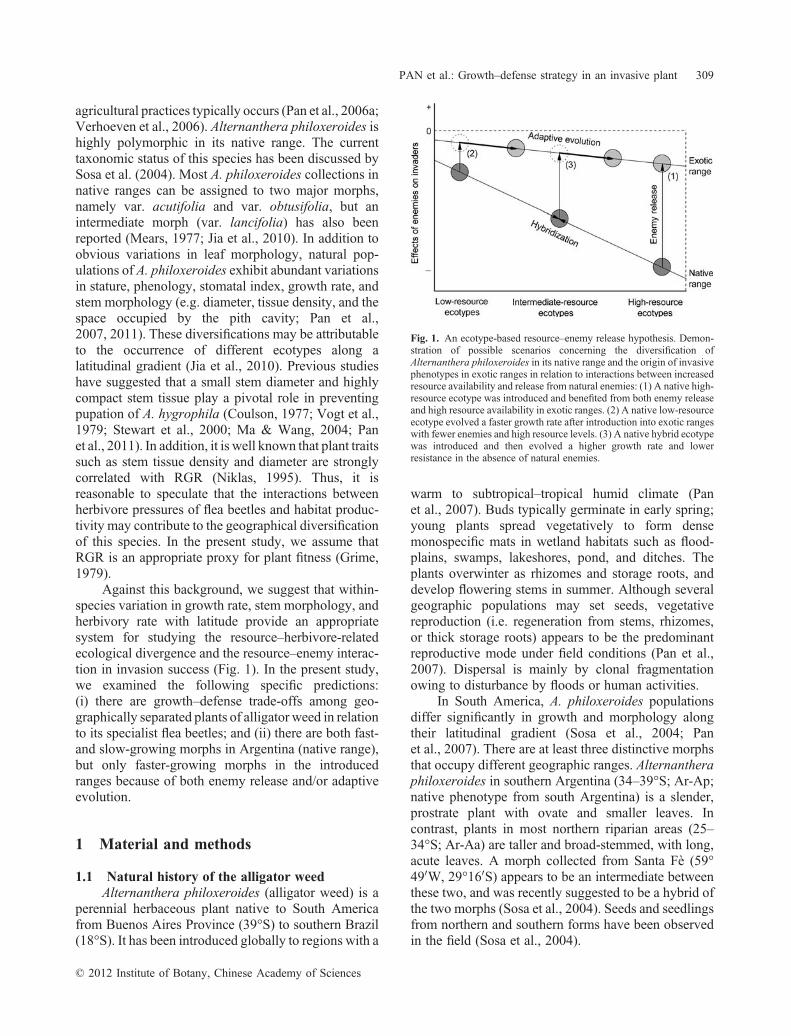
Against this background, we suggest that within‐species variation in growth rate, stem morphology, andherbivory rate with latitude provide an appropriatesystem for studying the resource–herbivore‐relatedecological divergence and the resource–enemy interac-tion in invasion success (Fig. 1). In the present study,we examined the following specific predictions:(i) there are growth–defense trade‐offs among geo-graphically separated plants of alligator weed in relationto its specialist flea beetles; and (ii) there are both fast‐and slow‐growing morphs in Argentina (native range),but only faster‐growing morphs in the introducedranges because of both enemy release and/or adaptiveevolution.
1 Material and methods
1.1 Natural history of the alligator weedAlternanthera philoxeroides (alligator weed) is a
perennial herbaceous plant native to South Americafrom Buenos Aires Province (39°S) to southern Brazil(18°S). It has been introduced globally to regions with a
warm to subtropical–tropical humid climate (Panet al., 2007). Buds typically germinate in early spring;young plants spread vegetatively to form densemonospecific mats in wetland habitats such as flood-plains, swamps, lakeshores, pond, and ditches. Theplants overwinter as rhizomes and storage roots, anddevelop flowering stems in summer. Although severalgeographic populations may set seeds, vegetativereproduction (i.e. regeneration from stems, rhizomes,or thick storage roots) appears to be the predominantreproductive mode under field conditions (Pan et al.,2007). Dispersal is mainly by clonal fragmentationowing to disturbance by floods or human activities.
In South America, A. philoxeroides populationsdiffer significantly in growth and morphology alongtheir latitudinal gradient (Sosa et al., 2004; Panet al., 2007). There are at least three distinctive morphsthat occupy different geographic ranges. Alternantheraphiloxeroides in southern Argentina (34–39°S; Ar‐Ap;native phenotype from south Argentina) is a slender,prostrate plant with ovate and smaller leaves. Incontrast, plants in most northern riparian areas (25–34°S; Ar‐Aa) are taller and broad‐stemmed, with long,acute leaves. A morph collected from Santa Fè (59°490W, 29°160S) appears to be an intermediate betweenthese two, and was recently suggested to be a hybrid ofthe two morphs (Sosa et al., 2004). Seeds and seedlingsfrom northern and southern forms have been observedin the field (Sosa et al., 2004).
Fig. 1. An ecotype‐based resource–enemy release hypothesis. Demon-stration of possible scenarios concerning the diversification ofAlternanthera philoxeroides in its native range and the origin of invasivephenotypes in exotic ranges in relation to interactions between increasedresource availability and release from natural enemies: (1) A native high‐resource ecotype was introduced and benefited from both enemy releaseand high resource availability in exotic ranges. (2) A native low‐resourceecotype evolved a faster growth rate after introduction into exotic rangeswith fewer enemies and high resource levels. (3) A native hybrid ecotypewas introduced and then evolved a higher growth rate and lowerresistance in the absence of natural enemies.
© 2012 Institute of Botany, Chinese Academy of Sciences
PAN et al.: Growth–defense strategy in an invasive plant 309

In the US, A. philoxeroides was first authenticallyrecorded in 1897 near Mobile, Alabama, although itmay have been found in Florida as early as 1894(Coulson, 1977). Within a few years, it was wellestablished in the wetland habitats from Virginia toFlorida in the east and westward to Texas, withpopulations also in California. It was recognized as athreat to waterways as early as 1901 (Coulson, 1977).As a consequence, alligator weed becomes the target ofthe first biocontrol program on aquatic plants estab-lished after 1960. Alternanthera philoxeroides has twomorphs in the US, one with slender stems and short,rounded leaves (USA‐NS; invasive narrow‐stemmedphenotype from the US) and the other with broad stemsand long, slender leaves (USA‐BS; invasive broad‐stemmed phenotype from the US; Kay & Haller, 1982;Wain et al., 1984).
The introduction of A. philoxeroides into Shang-hai, China, is believed to be incidental to the Sino‐Japanese war during the 1930s. It expanded its rangethrough cultivation practices and spread as a forage cropin southern China, where it had established itself as aprominent part of the Chinese weed flora by the1980s.Now, A. philoxeroides is one of the 12 most harmfulalien invasive species in China (for a review, see Panet al., 2007). Intensive studies have shown that there isvery little genetic variation among populations ofalligator weed in China (Xu et al., 2003; Yeet al., 2003). Thus, in the present study, we used plantscollected from Qingpu (121°060E, 31°090N) of Shang-hai as representative of the plants in China.
1.2 Clone collectionAll plant materials used in the present study
were clonally propagated. In December 2003 (inArgentina) and in September 2004 (in the US andChina), we collected ramets (generated from storageroots or rhizomes) from A. philoxeroides populations atsix sites (Table 1), namely San Ignacio (northernArgentina), Santa Fè (northern Buenos Aires,Argentina), Tandil (southern Buenos Aires, Argentina),
Baton Rouge (Louisiana, USA), Jackson (Mississippi,USA), and Shanghai (China). These sample sites weredesigned to cover the present distribution of differentmorphs of native and two major introduced populationsof alligator weed (Pan et al., 2007). We expectedthat the effects of geographic isolation on geneticallybased phenotypic differentiation would be more easilydetected in populations occurring at different sites on alarge spatial scale. At each site, we first selected three tofive separate (>10 km) populations in representativehabitats (e.g. roadsides, ponds, lakeshores, riparianzones, pastures etc.), which consisted largely ofherbaceous plant communities.Within each population,10–20 ramets were randomly collected. The distancesbetween the ramets were kept large enough to avoidsampling the same genet more than once. Aftersampling, the ramets were stored under moist con-ditions at approximately 8 °C. Because of the strictquarantine regulations between China and Argentina, itwas impossible for us to ship all the samples of thenative plants to China. Finally, five ramets wererandomly resampled from collections at each site andthen sent to Shanghai. All ramets were first planted inpots in the greenhouse at Fudan University untilJune 2006 to eliminate any possible environmentalmaterial effects and to produce plenty of ramets. Duringthis period, plants from different sites still differed inkey characters, such as stature, phenology, leaf angleand size, and stem morphology (Pan et al., 2006b). InJuly 2006, we established our common garden at theCoastal Ecosystems Research Station of Yangtze RiverEstuary (Chongming Island; 121°510E, 31°310N),Shanghai.
1.3 Agasicles hygrophilaAgasicles hygrophila Selman & Vogt (Coleoptera:
Chrysomelidae; alligator weed flea beetle) is a specialistphytophagous insect of alligator weed that has beenused as one of the most important biological controlagents of A. philoxeroides in North America (in 1964),Australia, New Zealand, and China (in 1986). It is
Table 1 Details of the sample collection sites for the six morphs of Alternanthera philoxeroides
Morphabbreviation
Site of clonecollection
Historicalpresence
Reproductivemode
Herbivore rateby flea beetles
Annual meantemperature (°C)/annual
precipitation (mm)
Geographicalcoordinates
Ar‐Ap Tandil, Argentina Native Seed/clonal No 13.8/860 37°110S, 59°040WAr‐As Santa Fè, Argentina Native Seed/clonal Seldom 18.8/950 29°170S, 59°490WAr‐Aa San Ignacio, Argentina Native Seed/clonal Frequent 22.5/1 300 27°250S, 55°530WUSA‐BS Baton Rouge, USA 1897 Seed clonal Frequent 19.5/1 600 29°060N, 90°080WUSA‐NS Jackson, USA 1897 Clonal Frequent 17.8/1 400 33°160N, 88°470WCH Shanghai, China 1930s Clonal Frequent 16.3/1 200 31°310 N, 121°510E
Ar‐Ap, native phenotype from south Argentina; Ar‐As, native phenotype from Santa Fè, Argentina; Ar‐Aa, native phenotype from north Argentina; USA‐BS, invasive broad‐stemmed phenotype from the US; USA‐NS, invasive narrow‐stemmed phenotype from the US; CH, invasive phenotype from China.
© 2012 Institute of Botany, Chinese Academy of Sciences
310 Journal of Systematics and Evolution Vol. 51 No. 3 2013

small, approximately 5 mm long, and the wing coversare striped with yellow and black. Agasicles hygrophiladeposits clusters of yellow eggs on the underside ofleaves. Newly hatched larvae are yellow, but theygradually darken to black as they mature. Both adultsand larvae feed on the leaves and other aerial portions ofalligator weed and can cause considerable damage(Maddox et al., 1971). Mature larvae chew into thehollow stems, where they pupate as adults approxi-mately 25 days after the eggs hatch (Maddoxet al., 1971). Little feeding occurs on terrestrial plants.Agasicles hygrophila is commonly multivoltine nearBuenos Aires, producing five generations per year, andprobably four to six generations in Florida and the lowerMississippi River Valley (Maddox et al., 1971) andseven to nine generations in south‐eastern coastalprovinces in China (Yang et al., 2002).
Agasicles hygrophila is specific to alligator weedand has not been reported on other host plants either inChina or the US, even after 20–40 years use (for areview, see Pan et al., 2007). It has been suggested thatcompletion ofA. hygrophila’s life cycle requires hollowstems for pupation, which may contribute, in part, to itsspecific adaptation to alligator weed (Coulson, 1977;Vogt et al., 1979; Stewart et al., 2000; Ma & Wang,2004). There is some evidence that A. hygrophilalocates its host plant because of the presence of feedingattractants and stimulants (Simons et al., 1968). The fleabeetles have a more restricted distribution than the hostalligator weed, not being found in hot tropical or coldregions despite the presence of the host plant (Coulson,1977). In the present study, specimens of A. hygrophilawere collected at the field station of the Center forManagement of Invasive Alien Species, Ministry ofAgriculture, China (CMIAS), at Wuhu City, AnhuiProvince, China, in July 2006. Flea beetles were raisedon alligator weed plants collected from Chongming,Shanghai.
1.4 Common garden experimentsWe first examined whether traits of alligator weed
morphs have any genetic differentiation. This questionwas addressed by common garden experiments of acompletely randomized design. We planted single stemcuttings (2‐ to 4‐cm stem segments with one node) foreach morph into pots containing a mixture of 50%sand and 50% vermiculite, and cultivated them in aglasshouse until they were approximately 2–5 cm high,depending on the morphs. Approximately 120 random-ly selected cuttings from each morph were transplantedinto 15‐cm diameter plastic pots for the common gardenexperiment and herbivory hierarchy experiment. Allplants were watered as needed and fertilized weekly
(half‐strength Hoagland solution, 50 mL per potper week) under conditions of 16:8 h light:dark,25 � 2 °C, 70%–100% relative humidity. The experi-ment lasted for 6 weeks.1.4.1 Maximum RGR of main stem length Themain stem length was measured every 3 days. The RGR(in mm/cm per day) for eachmorph was estimated fromthe slope of the regression of ln(plant stem length)versus time during the initial exponential phase ofgrowth.1.4.2 Leaf morphology Leaves were collectedfrom each plant of the six morphs and the length andwidth of three pairs of leaves were measured on eachplant. The ratio of leaf length:width was calculated.1.4.3 Stem morphology At final harvest, plantswere separated into leaves, main stems, roots, andoriginal stem cuttings. Tissues were dried at 65 °C for48 h and weighed. Specific stem length (SSL; the ratioof main stem length to its weight; in cm/g) wascalculated. The diameter (d) of the main stem from eachmorph was measured with calipers at the midpoints ofthe internodes between the third and fourth fullyexpanded leaf pairs. The mean tissue density (r) of themain stemwas determined from the ratio of stemweightto volume, the latter being determined by submerging astem in distilled water and recording the volume ofdisplaced water. In the case of alligator weed withhollow internodes, stems were cut into medianlongitudinal sections before being submerged in water.
There are two major sources of variability in thisexperiment, the first arising from the collection of rametmaterial and the second from differences among the sixmorphs. We used analysis of variance (ANOVA) andFisher’s least significant difference (LSD) test todetermine whether plants in the six morphs differedin terms of morphological and growth traits. Coef-ficients of variation (CV) were calculated. We used acylinder model (with a uniform cross‐sectional geome-try) to examine correlations between SSL and thetwo stem traits, namely tissue density (r) and stemdiameter (d):
SSL ¼ L=W ¼ L Vrð Þ ¼ 4= prd2� �
where L is length, W is weight, and V is stem volume.
1.5 Herbivory hierarchyWe used preference hierarchy (feeding and
oviposition) of flea beetles feeding on differentmorphs of alligator weed as the measure of herbivoryhierarchy. The preference hierarchy of A. hygrophilawas determined by multiple‐choice experiments(15 replicates). This type of experimentation gives
© 2012 Institute of Botany, Chinese Academy of Sciences
PAN et al.: Growth–defense strategy in an invasive plant 311

the same results as binary and no‐choice experiments(Fu et al., 2007), and is therefore considered to givereliable estimates of host preference. One pot of eachmorph was offered simultaneously to five female adultsduring a trial. All plants in the trial were of the same age(5 weeks), with leaf biomass sufficient to rear thebeetles. Pots were randomly positioned in a flight cage(50 � 75 � 75 cm). We observed beetles movingfreely among the six morph pots. The cages werekept at 25 � 2 °C and relative humidity of 75%–90%under a 14:10 h light–dark period. Each trial lastedfor 72 h, after which the number of eggs and leaf areaconsumed per genotype were recorded. We convertedthe leaf area consumed to leaf dry mass using thespecific leaf area, leaf area/leaf dry mass (SLA) ofeach alligator weed morph. We then estimated theherbivory intensity of flea beetles on the six morphsusing the number of eggs and the leaf mass consumedper day.
To test the prediction that stem diameter and tissuedensity of different morphs of alligator weed can affectthe pupation rate of flea beetle (five replicates), thepupation rates on six genotypes were determined byraising the eggs under a no‐choice experiment. Larvaewere drawn at random from eggs that were laid bymultiple females on each morph. Ten first instar larvae(<24 h old) were placed gently on apical leavesusing a paintbrush. One pot of each plant morph wasthen enclosed in a polythene bag with circular holes(0.4 mm diameter) and sealed around the base of thepot to prevent the larvae from escaping while theplants were allowed to transpire and facilitate nearfull light transmission. Pots were monitored daily forpupated and dead larvae. The positions of the larvalhost plants were randomized daily within the en-vironmental chamber to avoid position effects. After4 weeks, adult beetles emerged from pupation, withthe final number of adults that successfully emergedeasily determined by counting the eclosion holes onthe stems.
The results of the multiple‐choice experiments(feeding preference and oviposition preference) wereanalyzed using the Quade test (Quade, 1979). This non‐parametric test takes into account the lack of indepen-dence among the simultaneously offered food types(Roa, 1992). Offspring survival was analyzed usingmixed‐model logistic regression with logit link andbinomial errors. Pearson’s correlation analyses wereused to examine the relationships between meanpreference and mean performance on the differenthosts. Data that violated the ANOVA assumptions ofnormality and homogeneity of variance were ranktransformed.
2 Results
2.1 Phenotypic differences in growth andmorphological traits
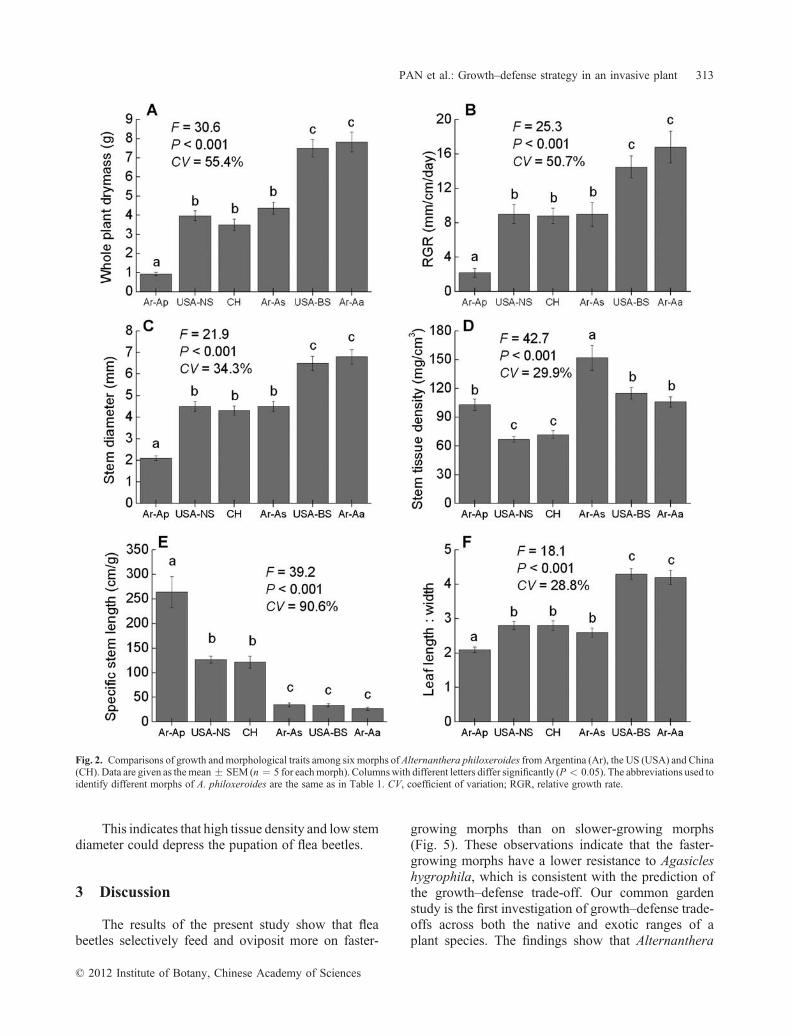
Both measures of growth and morphologicalcharacters differed considerably among the six morphsof alligator weed from wide geographical locations(CV 28.8%–90.6%; Fig. 2). First, the three nativemorphs differed significantly from each other (Fig. 1: A,B; Fig. 2). Native Ar‐Ap had the lowest growth rate(2.2 mm/cm per day), whole plant biomass, distinctslender stems, and small ovate leaves. Native Ar‐Aawas another extreme that grew fastest (16.8 mm/cm perday) and was distinguished by broad stems and longacute leaves. Except for having the highest stem density(Fig. 2: D), the hybrid morph Ar‐As (native phenotypefrom Santa Fè, Argentina) was intermediate between itstwo parents Ar‐Ap and Ar‐Aa. Second, the two USmorphs were clearly different, with USA‐BS exhibitinga higher growth rate (14.5 mm/cm per day), a broadstem, and long acute leaves, whereas USA‐NS had anintermediate growth rate (8.8 mm/cm per day) and stemand leaf morphology (Fig. 2). Third, the growth andmorphological traits of CH were all similar to those ofUSA‐NS (Fig. 2).
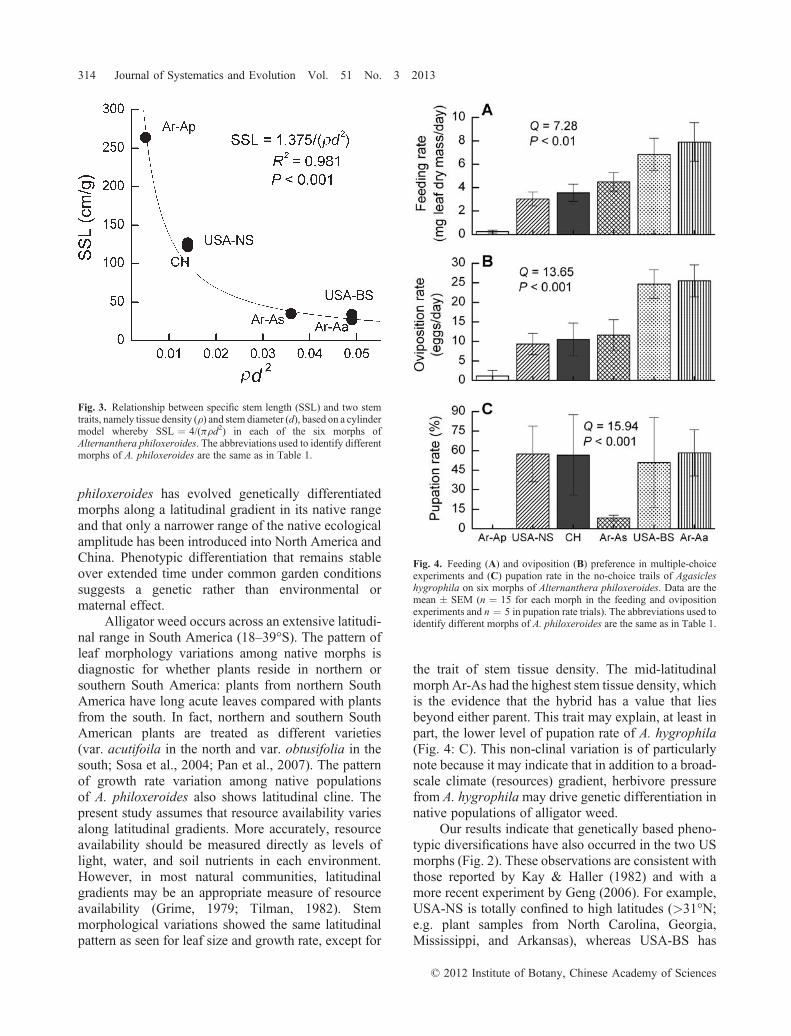
The highest variation was seen in SSL (CV ¼91%; Fig. 2: E). The relationship between SSL and theproduct of stem diameter and stem tissue density waswell fitted by a cylindrical model (Fig. 3). There was anegative correlation between SSL and stem growth rate(r ¼ � 0.879; P ¼ 0.021; n ¼ 10).
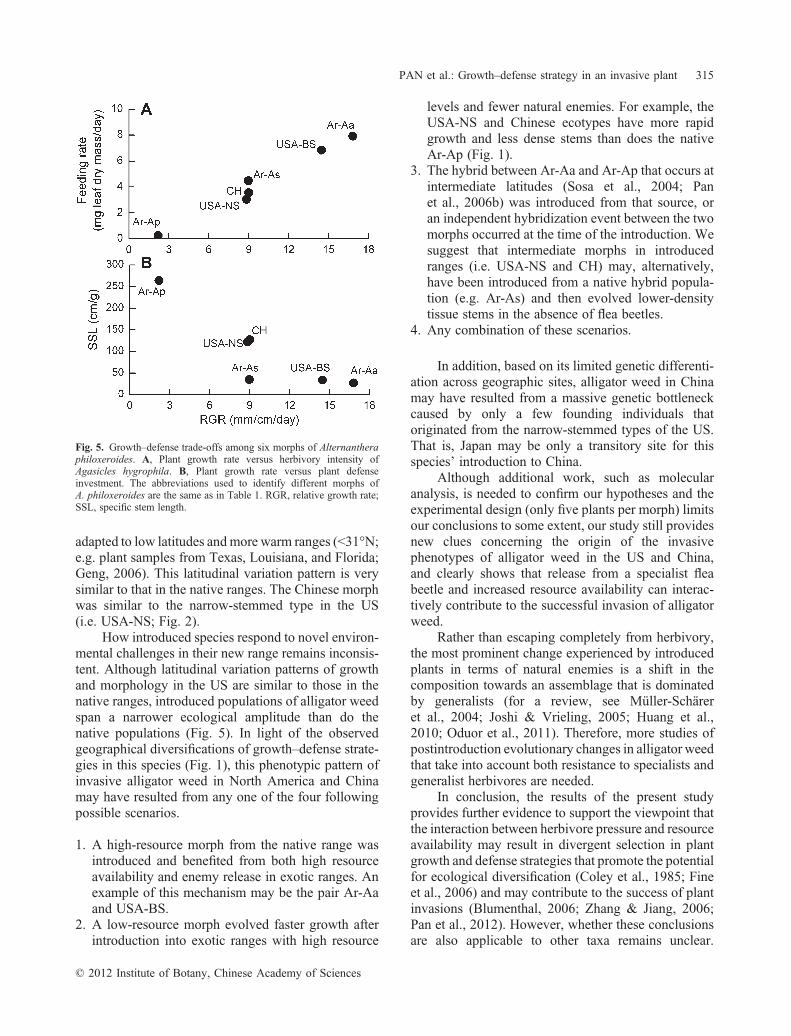
2.2 Herbivory hierarchyThe intensities of feeding and oviposition differed
significantly among the six morphs of alligator weed(Fig. 4: A, B), and were highly correlated with eachother (r ¼ 0.983; P < 0.001; n ¼ 10). The Ar‐Apmorph was the least preferred host morph, withpreference increasing in the order USA‐NSA, CH,USA‐BSA, and Ar‐Aa (Fig. 4: A, B). Both oviposition(r ¼ 0.989; P < 0.001) and feeding preference(r ¼ 0.986; P < 0.001; n ¼ 10) were significantlycorrelated with plant growth rate.
The pupation rate differed significantly betweenthe native Ar‐Ap (0%), Ar‐As (8%) and the other fourmorphs (51%–58%, Fig. 4: C). Regression analysisrevealed a linear relationship between pupation rate andboth stem tissue density and diameter (P ¼ 0.002;n ¼ 10) with the following equation:
Pupation rate ¼ 44:1� 668:6 stem tissue density
ðmg=cm3Þ þ 12:9 stem diameter ðmmÞ
© 2012 Institute of Botany, Chinese Academy of Sciences
312 Journal of Systematics and Evolution Vol. 51 No. 3 2013
This indicates that high tissue density and low stemdiameter could depress the pupation of flea beetles.
3 Discussion
The results of the present study show that fleabeetles selectively feed and oviposit more on faster‐
growing morphs than on slower‐growing morphs(Fig. 5). These observations indicate that the faster‐growing morphs have a lower resistance to Agasicleshygrophila, which is consistent with the prediction ofthe growth–defense trade‐off. Our common gardenstudy is the first investigation of growth–defense trade‐offs across both the native and exotic ranges of aplant species. The findings show that Alternanthera
Fig. 2. Comparisons of growth andmorphological traits among six morphs of Alternanthera philoxeroides fromArgentina (Ar), the US (USA) and China(CH). Data are given as themean � SEM (n ¼ 5 for eachmorph). Columnswith different letters differ significantly (P < 0.05). The abbreviations used toidentify different morphs of A. philoxeroides are the same as in Table 1. CV, coefficient of variation; RGR, relative growth rate.
© 2012 Institute of Botany, Chinese Academy of Sciences
PAN et al.: Growth–defense strategy in an invasive plant 313
philoxeroides has evolved genetically differentiatedmorphs along a latitudinal gradient in its native rangeand that only a narrower range of the native ecologicalamplitude has been introduced into North America andChina. Phenotypic differentiation that remains stableover extended time under common garden conditionssuggests a genetic rather than environmental ormaternal effect.
Alligator weed occurs across an extensive latitudi-nal range in South America (18–39°S). The pattern ofleaf morphology variations among native morphs isdiagnostic for whether plants reside in northern orsouthern South America: plants from northern SouthAmerica have long acute leaves compared with plantsfrom the south. In fact, northern and southern SouthAmerican plants are treated as different varieties(var. acutifoila in the north and var. obtusifolia in thesouth; Sosa et al., 2004; Pan et al., 2007). The patternof growth rate variation among native populationsof A. philoxeroides also shows latitudinal cline. Thepresent study assumes that resource availability variesalong latitudinal gradients. More accurately, resourceavailability should be measured directly as levels oflight, water, and soil nutrients in each environment.However, in most natural communities, latitudinalgradients may be an appropriate measure of resourceavailability (Grime, 1979; Tilman, 1982). Stemmorphological variations showed the same latitudinalpattern as seen for leaf size and growth rate, except for
the trait of stem tissue density. The mid‐latitudinalmorph Ar‐As had the highest stem tissue density, whichis the evidence that the hybrid has a value that liesbeyond either parent. This trait may explain, at least inpart, the lower level of pupation rate of A. hygrophila(Fig. 4: C). This non‐clinal variation is of particularlynote because it may indicate that in addition to a broad‐scale climate (resources) gradient, herbivore pressurefrom A. hygrophilamay drive genetic differentiation innative populations of alligator weed.
Our results indicate that genetically based pheno-typic diversifications have also occurred in the two USmorphs (Fig. 2). These observations are consistent withthose reported by Kay & Haller (1982) and with amore recent experiment by Geng (2006). For example,USA‐NS is totally confined to high latitudes (>31°N;e.g. plant samples from North Carolina, Georgia,Mississippi, and Arkansas), whereas USA‐BS has
Fig. 4. Feeding (A) and oviposition (B) preference in multiple‐choiceexperiments and (C) pupation rate in the no‐choice trails of Agasicleshygrophila on six morphs of Alternanthera philoxeroides. Data are themean � SEM (n ¼ 15 for each morph in the feeding and ovipositionexperiments and n ¼ 5 in pupation rate trials). The abbreviations used toidentify different morphs of A. philoxeroides are the same as in Table 1.
Fig. 3. Relationship between specific stem length (SSL) and two stemtraits, namely tissue density (r) and stem diameter (d), based on a cylindermodel whereby SSL ¼ 4/(prd2) in each of the six morphs ofAlternanthera philoxeroides. The abbreviations used to identify differentmorphs of A. philoxeroides are the same as in Table 1.
© 2012 Institute of Botany, Chinese Academy of Sciences
314 Journal of Systematics and Evolution Vol. 51 No. 3 2013
adapted to low latitudes and more warm ranges (<31°N;e.g. plant samples from Texas, Louisiana, and Florida;Geng, 2006). This latitudinal variation pattern is verysimilar to that in the native ranges. The Chinese morphwas similar to the narrow‐stemmed type in the US(i.e. USA‐NS; Fig. 2).
How introduced species respond to novel environ-mental challenges in their new range remains inconsis-tent. Although latitudinal variation patterns of growthand morphology in the US are similar to those in thenative ranges, introduced populations of alligator weedspan a narrower ecological amplitude than do thenative populations (Fig. 5). In light of the observedgeographical diversifications of growth–defense strate-gies in this species (Fig. 1), this phenotypic pattern ofinvasive alligator weed in North America and Chinamay have resulted from any one of the four followingpossible scenarios.
1. A high‐resource morph from the native range wasintroduced and benefited from both high resourceavailability and enemy release in exotic ranges. Anexample of this mechanism may be the pair Ar‐Aaand USA‐BS.
2. A low‐resource morph evolved faster growth afterintroduction into exotic ranges with high resource
levels and fewer natural enemies. For example, theUSA‐NS and Chinese ecotypes have more rapidgrowth and less dense stems than does the nativeAr‐Ap (Fig. 1).
3. The hybrid between Ar‐Aa and Ar‐Ap that occurs atintermediate latitudes (Sosa et al., 2004; Panet al., 2006b) was introduced from that source, oran independent hybridization event between the twomorphs occurred at the time of the introduction. Wesuggest that intermediate morphs in introducedranges (i.e. USA‐NS and CH) may, alternatively,have been introduced from a native hybrid popula-tion (e.g. Ar‐As) and then evolved lower‐densitytissue stems in the absence of flea beetles.
4. Any combination of these scenarios.
In addition, based on its limited genetic differenti-ation across geographic sites, alligator weed in Chinamay have resulted from a massive genetic bottleneckcaused by only a few founding individuals thatoriginated from the narrow‐stemmed types of the US.That is, Japan may be only a transitory site for thisspecies’ introduction to China.
Although additional work, such as molecularanalysis, is needed to confirm our hypotheses and theexperimental design (only five plants per morph) limitsour conclusions to some extent, our study still providesnew clues concerning the origin of the invasivephenotypes of alligator weed in the US and China,and clearly shows that release from a specialist fleabeetle and increased resource availability can interac-tively contribute to the successful invasion of alligatorweed.
Rather than escaping completely from herbivory,the most prominent change experienced by introducedplants in terms of natural enemies is a shift in thecomposition towards an assemblage that is dominatedby generalists (for a review, see Müller‐Schäreret al., 2004; Joshi & Vrieling, 2005; Huang et al.,2010; Oduor et al., 2011). Therefore, more studies ofpostintroduction evolutionary changes in alligator weedthat take into account both resistance to specialists andgeneralist herbivores are needed.
In conclusion, the results of the present studyprovides further evidence to support the viewpoint thatthe interaction between herbivore pressure and resourceavailability may result in divergent selection in plantgrowth and defense strategies that promote the potentialfor ecological diversification (Coley et al., 1985; Fineet al., 2006) and may contribute to the success of plantinvasions (Blumenthal, 2006; Zhang & Jiang, 2006;Pan et al., 2012). However, whether these conclusionsare also applicable to other taxa remains unclear.
Fig. 5. Growth–defense trade‐offs among six morphs of Alternantheraphiloxeroides. A, Plant growth rate versus herbivory intensity ofAgasicles hygrophila. B, Plant growth rate versus plant defenseinvestment. The abbreviations used to identify different morphs ofA. philoxeroides are the same as in Table 1. RGR, relative growth rate;SSL, specific stem length.
© 2012 Institute of Botany, Chinese Academy of Sciences
PAN et al.: Growth–defense strategy in an invasive plant 315

Increased resource availability in habitats and releasefrom natural enemies appear to be far more common inthe present human‐dominated world (Verhoevenet al., 2006); thus, an important and widespread rolefor resource–herbivory interactions in molding theadaptive evolution of invasive plants is plausible.
Acknowledgements The authors are grateful to DrsChen‐Yuan XU (CSIRO Entomology, Indooroopilly,Qld, Australia) and Yu‐Peng GENG (Yunnan Univer-sity, Kunming, China) for their comments on earlierversions of this manuscript. This research wassupported by the National Natural Science Foundationof China (Grant Nos. 31070369, 1131070417).
References
Agrawal AA. 2007. Macroevolution of plant defense strategies.Trends in Ecology and Evolution 22: 103–109.
Blumenthal DM. 2006. Interactions between resource availabili-ty and enemy release in plant invasion. Ecology Letters 9:887–895.
Coley PD, Bryant JP, Chapin FS. 1985. Resource availability andplant anti‐herbivore defense. Science 230: 895–899.
Coulson JR. 1977. Biological control of alligator weed, 1959–1972: A review and evaluation. Technical Bulletin No.1547. Washington, DC: U.S. Department of Agriculture,Agricultural Research Service.
Crawley MJ. 1997. Plant–herbivore dynamics. In: CrawleyMJ ed. Plant ecology. Oxford: Blackwell Science. 401–474.
Fine PVA, Miller ZJ, Mesones I, Irazuzta S, Appel HM, StevensMH, Saaksjarvi I, Schultz LC, Coley PD. 2006. Thegrowth–defense trade‐off and habitat specialization byplants in Amazonian forests. Ecology 87: S150–S162.
Fu DJ, Jia X, Yang XZ, Li B, Chen JK, Pan XY. 2007. Feedingpreference and growth adaptation of alligator weed fleabeetle (Agasicles hygrophila) in different ecotypes ofalligator weed (Alternanthera philoxeroides). Shanxi ForestScience and Technology 2: 21–25.
Geng YP. 2006. Adaptive strategies of invasive alligator weed,Alternanthera philoxeroides, in heterogeneous habitats.Ph.D. Dissertation. Shanghai: Fudan University.
Grime JP. 1979. Plant strategies and vegetation processes. NewYork: John Wiley & Sons.
HuangW, Siemann E,Wheeler GS, Zou JW, Carrillo J, Ding JQ.2010. Allocation to defence and growth are driven bydifferent responses to generalist and specialist herbivory inan invasive plant. Journal of Ecology 98: 57–167.
Jia X, Pan XY, Sosa A, Li B, Chen JK. 2010. Differentiationin growth and biomass allocation among three nativeAlternanthera philoxeroides varieties fromArgentina. PlantSpecies Biology 25: 85–92.
Joshi J, Vrieling K. 2005. The enemy release and EICAhypothesis revisited: Incorporating the fundamental differ-ence between specialist and generalist herbivores. EcologyLetters 8: 704–714.
Kay SH, Haller WT. 1982. Evidence for the existence of distinctalligator weed biotypes. Journal of Aquatic Plant Manage-ment 20: 37–41.
Ma RY, Wang R. 2004. Effect of morphological andphysiological variations in the ecotypes of alligatorweed,Alternanthera philoxerodies on the pupation rate of itsbiocontrol agent Agasicles hygrophila. Acta Phytoecolo-gica Sinica 28: 24–30.
Maddox DM, Andres LA, Hennessey RD, Blackburn RD,Spencer NR. 1971. Inescts to control alligator weed: Aninvader of aquatic ecosystems in the United States.BioScience 21: 985–991.
Mears JA. 1977. The nomenclature and type collections ofthe widespread taxa of Alternanthera (Amaranthaceae).Proceedings of the Natural Sciences of Philadelphia 129:1–2.
Müller‐Schärer H, Schaffner U, Steinger T. 2004. Evolution ininvasive plants: Implications for biological control. Trendsin Ecology and Evolution 19: 417–422.
Niklas K. 1995. Plant height and the properties of someherbaceous stems. Annals of Botany 75: 133–142.
Oduor AMO, Lankau RA, Strauss SY, Gómez JM. 2011.Introduced Brassica nigra populations exhibit greatergrowth and herbivore resistance but less tolerance thannative populations in the native range. New Phytologist191: 536–544.
Pan XY, Geng YP, Sosa AJ, Zhang WJ, Li B, Chen JK. 2007.Invasive Alternanthera philoxeroides: Biology, ecologyand management. Acta Phytotaxonomica Sinica 45: 884–900.
Pan XY, Geng YP, Zhang WJ, Li B, Chen JK. 2006a. Theinfluence of abiotic stress and phenotypic plasticity on thedistribution of invasiveAlternanthera philoxeroides along ariparian zone. Acta Oecollogica 30: 333–341.
Pan XY, Liang HZ, Sosa AJ, Geng YP, Li B, Chen JK.2006b. Morphological variation pattern of alligator weed(Alternanthera philoxeroides): From native to introducedregions. Biodiversity Science 14: 232–240.
Pan XY, Jia X, Chen JK, Li B. 2012. For or against: Theimportance of variation in growth rate for testing the EICAhypothesis. Biological Invasions 14: 1–8.
Pan XY, Jia X, Zeng J, Sosa A, Li B, Chen JK. 2011. Stem tissuemass density is linked to growth and resistance to a stem‐
boring insect in Alternanthera philoxeroides. Plant SpeciesBiology 26: 58–65.
Pennings SC, Silliman BR. 2005. Linking biogeography andcommunity ecology: Latitudinal variation in plant‐herbi-vore interaction strength. Ecology 86: 2310–2319.
Quade D. 1979. Using weighted rankings in the analysis ofcomplete blocks with additive block effects. Journal of theAmerican Statistical Association 74: 680–683.
Roa R. 1992. Design and analysis of multiple‐choice feedingpreference experiments. Oecologia 89: 509–515.
Simons JN, Silverstein RM, Bellas T. 1968. Tests of a feedingstimulant in alligator weed for Agasicles n. sp. Journal ofEconomic Entomology 61: 1448–1449.
Sosa AJ, Julien MH, Corda H. 2004. New research onAlternanthera philoxeroides (alligator weed) in its SouthAmerican native range. In: Cullen JM, Briese DT, KriticosDJ, Lonsdale WM, Morin L, Scott JK eds. Proceedings of
© 2012 Institute of Botany, Chinese Academy of Sciences
316 Journal of Systematics and Evolution Vol. 51 No. 3 2013

the XI international symposium on biological control ofweeds. Canberra, Australia: CSIRO Entomology. 180–185.
Stewart CA, Chapman RB, Frampton CMA. 2000. Growth ofalligator weed (Alternanthera philoxeroides) and popula-tion development of Agasicles hygrophila in northern NewZealand. Plant Protection Quarterly 15: 95–101.
TilmanD. 1982. Resource competition and community structure.Princeton: Princeton University Press.
Verhoeven JTA, Arheimer B, Yin CQ, Hefting MM. 2006.Regional and global concerns over wetlands and waterquality. Trends in Ecology and Evolution 21: 96–103.
Vogt GB,McGuire JU, Crushman AD. 1979. Probable evolutionand morphological variation in South American Disony-chine flea beatles (Coleoptera: Chrysomelidae) and theirAmaranthaceous hosts. Technical Bulletin No. 1593.Washington, DC: U.S. Department of Agriculture, Agri-cultural Research Service.
Wain RP, Haller WT, Martin DF. 1984. Genetic relationshipamong two forms of alligator weed. Journal of AquaticPlant Management 22: 104–105.
Xu CY, Zhang WJ, Fu CZ, Lu BR. 2003. Genetic diversity ofalligator weed in China byRAPD analysis. Biodiversity andConservation 12: 637–645.
Yang ZH, He CJ, Qian DM. 2002. A preliminary observation forpopulation dynamic of Agasicles hygrophila Selman &Vogt and its control effect on alligatorweed in the suburb ofShanghai. Acta Agriculturae Shanghai 18: 79–83.
Ye WH, Li J, Cao HL, Ge XJ. 2003. Genetic uniformityof Alternanthera philoxeroides in South China. WeedResearch 43: 297–302.
Zhang DY, Jiang XH. 2006. Interactive effects of habitatproductivity and herbivore pressure on the evolution of anti‐herbivore defense in invasive plant populations. Journal ofTheoretical Biology 242: 935–994.
© 2012 Institute of Botany, Chinese Academy of Sciences
PAN et al.: Growth–defense strategy in an invasive plant 317





























