Embed Size (px)
Citation preview

1
Identification of antiparkinsonian drugs in the 6-hydroxydopamine zebrafish model 1
Rita L. Vaz 1,2,†, Sara Sousa 1,†,*, Diana Chapela 1,3, Herma C. van der Linde 4, Rob 2
Willemsen 4, Ana D. Correia 3, &, Tiago F. Outeiro 5,6,7,8 and Nuno D. Afonso1* 3
4
1 TechnoPhage, SA, Av. Prof. Egas Moniz, 1649-028 Lisboa, Portugal 5
2 Faculdade de Medicina, Universidade de Lisboa, Lisboa, Portugal 6
3 Instituto de Medicina Molecular, Faculdade de Medicina, Universidade de Lisboa, Av. 7
Prof. Egas Moniz, 1649-028 Lisboa, Portugal 8
4 Department of Clinical Genetics, Erasmus MC, Rotterdam, the Netherlands. 9
5 Department of Experimental Neurodegeneration, Center for Nanoscale Microscopy and 10
Molecular Physiology of the Brain, Center for Biostructural Imaging of 11
Neurodegeneration, University Medical Center Göttingen, Göttingen, Germany 12
6 CEDOC, Chronic Diseases Research Centre, NOVA Medical School | Faculdade de 13
Ciências Médicas, Universidade NOVA de Lisboa, Campo dos Mártires da Pátria, 130, 14
1169-056 Lisboa, Portugal. 15
7 Max Planck Institute for Experimental Medicine, Göttingen, Germany 16
8 Institute of Neuroscience, Medical School, Newcastle University, Framlington Place, 17
Newcastle Upon Tyne, NE2 4HH, UK 18
19
† Equal contribution 20
& Current address: Battelle UK Limited, Chelmsford Business Park, Springfield, 21
Chelmsford CM2 5LB, United Kingdom 22
23
*Co-corresponding authors: 24
Dr. Nuno D. Afonso 25
TechnoPhage, SA, 26
Av. Prof. Egas Moniz 27
1649-028 Lisboa 28
Portugal 29
Telephone: +(351) 217999545 30
Email: [email protected] 31
32
Dr. Sara Sousa 33

2
TechnoPhage, SA, 34
Av. Prof. Egas Moniz 35
1649-028 Lisboa 36
Portugal 37
Telephone: +(351) 217999545 38
Email: [email protected] 39
40

3
Abstract 41
Parkinson’s disease (PD) is known as a movement disorder due to characteristic motor 42
features. Existing therapies for PD are only symptomatic, and their efficacy decreases as 43
disease progresses. Zebrafish, a vertebrate in which parkinsonism has been modelled, 44
offers unique features for the identification of molecules with antiparkinsonian properties. 45
Here, we developed a screening assay for the selection of neuroactive agents with 46
antiparkinsonian potential. First, we performed a pharmacological validation of the 47
phenotypes exhibited by the 6-hydroxydopamine zebrafish model, by testing the effects 48
of known antiparkinsonian agents. These drugs were also tested for disease-modifying 49
properties by whole mount immunohistochemistry to TH+ neurons and confocal 50
microscopy in the dopaminergic diencephalic cluster of zebrafish. Next, we optimized a 51
phenotypic screening using the 6-hydroxydopamine zebrafish model and tested 1600 52
FDA-approved bioactive drugs. We found that 6-hydroxydopamine-lesioned zebrafish 53
larvae exhibit bradykinetic and dyskinetic-like behaviours that are rescued by the 54
administration of levodopa, rasagiline, isradipine or amantadine. The rescue of 55
dopaminergic cell loss by isradipine was also verified, through the observation of a higher 56
number of TH+ neurons in 6-OHDA-lesioned zebrafish larvae treated with this compound 57
as compared to untreated lesioned larvae. The phenotypic screening enabled us to identify 58
several compounds previously positioned for PD, as well as, new molecules with potential 59
antiparkinsonian properties. Among these, we selected stavudine, tapentadol and 60
nabumetone as the most promising candidates. Our results demonstrate the functional 61
similarities of the motor impairments exhibited by 6-hydroxydopamine-lesioned 62
zebrafish with mammalian models of PD and with PD patients, and highlights novel 63
molecules with antiparkinsonian potential. 64
65
Keywords: Drug screening, Parkinson’s disease, repositioning, zebrafish. 66
67
List of Abbreviations 68
6-OHDA, 6-hydroxydopamine AIMs, abnormal involuntary movements 69
BBB, blood brain barrier BDNF, brain derived neurotrophic factor 70
dpf, days post fertilization EM, embryo medium 71

4
L-dopa, levodopa PBS, phosphate buffer saline 72
PD, Parkinson’s disease PFA, paraformaldehyde 73
TH, tyrosine hydroxylase TU, Tubingen 74
75

5
Introduction 76
Parkinson’s disease (PD) is the most common movement disorder, affecting around 5 77
million people worldwide (Dorsey et al., 2007). The characteristic motor features of this 78
neurodegenerative disorder include bradykinesia, resting tremor, postural instability and 79
muscular rigidity. While the causes of PD are still not completely understood, the 80
degeneration of dopaminergic neurons in the substantia nigra is a major pathological 81
feature of the disease (Hirsch et al., 2013). Current pharmacological therapies, such as 82
levodopa (L-dopa), dopamine agonists (apomorphine, bromocriptine, among others), 83
monoamine oxidase (MAO)-B inhibitors (rasagiline, selegiline and safinamide) and 84
catechol-o-methyl transferase inhibitors (entacapone, tolcapone, nitecapone and 85
opicapone), address the loss of dopamine by either replacing it or controlling its 86
metabolism (Oertel and Schulz, 2016). Although these drugs substantially improve 87
quality of life, they lose efficacy as the disease progresses, and give rise to L-dopa induced 88
motor fluctuations and dyskinesia. Amantadine, an NMDA antagonist, is the most 89
effective anti-dyskinetic drug, but the associated side effects limit its use (Vijayakumar 90
and Jankovic, 2016). Non-dopaminergic agents, including A2A-receptor antagonists and 91
modulators of glutamate receptors, as well as nicotine, caffeine and isradipine are 92
currently under clinical trials (Oertel and Schulz, 2016). Despite the number of new 93
candidate agents that have successfully displayed antiparkinsonian effects in preclinical 94
studies, there is a growing demand for the development of alternative drugs. 95
Zebrafish is particularly suitable for large-scale drugs screening due to its small size, 96
transparency and high permeability to compounds diluted in the surrounding media. In 97
addition, studies have explored the organization and function of the zebrafish 98
dopaminergic system and suggest its overall conserved when compared to mammalian 99
vertebrates (Godoy et al., 2015; Rico et al., 2011; Rink and Wullimann, 2002, 2001), 100
rendering zebrafish as a practical and economic alternative vertebrate model for testing 101
the effect of neuroactive compounds (Parng et al., 2006; Sun et al., 2012). The sensitivity 102
of zebrafish to specific neurotoxins known to induce dopaminergic cell loss in rodent 103
models of PD has also been well validated (Anichtchik et al., 2004; Babu et al., 2016; 104
Wang et al., 2017). Specifically, the exposure of zebrafish larvae to 6-hydroxydopamine 105
(6-OHDA) induces the typical phenotypic features of PD, namely death of dopaminergic 106
neurons and bradykinesia (Feng et al., 2014). Furthermore, 6-OHDA-lesioned zebrafish 107
respond to antiparkinsonian compounds, such as levodopa or rasagiline (Cronin and 108

6
Grealy, 2017; Feng et al., 2014). Therefore, this model constitutes an excellent tool for 109
evaluating novel therapeutic options, not only as an alternative but also to complement 110
studies in mammalian models (Chong et al., 2013; Flinn et al., 2008; Vaz et al., 2018; 111
Wang et al., 2011; Xi et al., 2011; Zhang et al., 2012, 2011). 112
Using the 6-OHDA zebrafish model, we developed a platform for the selection of novel 113
potential antiparkinsonian molecules. First, we evaluated the effects of established anti-114
parkinsonian agents, such as L-dopa, rasagiline and isradipine on two specific measures 115
of motor performance, total distance moved and burst swimming. These three compounds 116
were also tested for their disease-modifying properties as determined by dopaminergic 117
neuronal loss. We then evaluated a new parameter, immobile events, as a surrogate 118
marker for dyskinetic-like behaviour (Babu et al., 2016), using the previously described 119
drugs and amantadine. Finally, we performed a phenotypic screen of 1600 FDA approved 120
drugs and selected the most promising candidates based on their ability to rescue motor 121
impairments in 6-OHDA-lesioned zebrafish larvae and evaluated their potential for 122
repositioning. 123
124

7
Material and methods 125
Chemicals and reagents 126
6-hydroxydopamine hydrobromide (6-OHDA) stock solution was prepared in 0.2% (w/v) 127
ascorbic acid and tested at 250, 500, 600, 750 and 800 µM, (Supplementary fig. S1A and 128
B). Tapentadol hydrochloride was purchased from Thonson Technology Ltd. (Shanghai, 129
China). The library of bioactive, FDA approved drugs was purchased from Microsource 130
(Gaylordsville, CT, USA). Nabumetone (Cat# N6142), stavudine (Cat# Y0000408), 6-131
OHDA (Cat# 162957), levodopa methyl ester (L-dopa, Cat# D9628), rasagiline (Cat# 132
SML0124), isradipine (Cat# I6658), paraformaldehyde (Cat# P6148), bovine serum 133
albumin (Cat# A2153) and triton X-100 (Cat# X100) were purchased from Sigma-134
Aldrich (St Louis, MO, USA). Amantadine was used directly from the library of drugs. 135
DABCO (Cat# 803456) was obtained from Millipore (Darmstadt, Germany). Mouse anti-136
tyrosine hydroxylase (TH, Cat# 22941) antibody was purchased from ImmunoStar 137
(Hudson, WI, USA) and AlexaFluor 568 (Cat# A11004) secondary antibody was 138
obtained from ThermoFisher Scientific (Waltham, MA, USA). All other reagents were 139
purchased from AppliChem (Darmstadt, Germany). 140
141
Zebrafish maintenance 142
Animal procedures were performed in accordance with the European Community 143
guidelines (Directive 2010/63/EU), Portuguese law on animal care (DL 113/2013), and 144
approved by the Instituto de Medicina Molecular Internal Committee and the Portuguese 145
Animal Ethics Committee (Direcção Geral de Alimentação e Veterinária). Tubingen (TU) 146
wild-type zebrafish were obtained from ZIRC (University of Oregon, Eugene, USA), 147
maintained and bred in constant conditions, by following standard guidelines for fish care 148
and maintenance protocols (Westerfield, 2000). 149
150
Treatment protocol 151
The protocol optimized for exposure of zebrafish larvae to the compounds was the same 152
for testing therapeutic controls, screening the library and determining dose-response 153
curves (Fig. 1). All experimental procedures were conducted between 8AM and 8PM. 154

8
Investigators were blind to the experimental groups during the screening protocol. 155
Briefly, zebrafish larvae were allowed to develop in embryo medium (EM [5 mM NaCl, 156
0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, pH 7.4]) with 1µM methylene blue 157
until 4 days post fertilization (dpf). At this developmental stage, the zebrafish 158
catecholaminergic system is fully developed and the blood brain barrier (BBB) is already 159
functional (Du et al., 2016; Jeong et al., 2008; Kastenhuber et al., 2010). On the other 160
hand, 6-OHDA induces dopaminergic lesion in zebrafish larvae with up to 5 dpf, 161
suggesting the BBB permeability to this neurotoxin (Feng et al., 2014). 4 dpf zebrafish 162
larvae were arbitrarily distributed into 24-well plate (8 larvae/well) containing EM + 10 163
mM HEPES and treated with 750 µM of 6-OHDA, for 24h. The next day, compounds 164
were added to the medium at the established concentration and larvae (n= 8-16 larvae) 165
were incubated for 24h. To counteract zebrafish larvae neuroregenerative capability, 166
further 6-OHDA was added to each well simultaneously. For each experiment, healthy 167
larvae treated with 0.2% ascorbic acid (vehicle, n= 8-16 larvae) and untreated 6-OHDA-168
lesioned larvae (6-OHDA, n= 8-16 larvae) were used as controls. The sample size was 169
predetermined by non-statistical methods. Particularly, the exposure to 6-OHDA resulted 170
in a variable rate of survival dependent on zebrafish larvae fitness. For suitable statistical 171
analysis and in compliance with the 3Rs, this study was designed to ensure a minimum 172
of 5 zebrafish larvae per group and, given only experiments with more than 65% of 173
surviving animals were considered, minimum 8 zebrafish larvae were assigned to each 174
group. Behavioural assessment was performed at the end of the incubation with the 175
compounds. 176

9
177
Figure 1 – Schematic representation of the platform developed for the selection of 178
novel compounds with antiparkinsonian potential. The protocol of the phenotypic 179
screening starts with dopaminergic lesion by exposure of the zebrafish larvae to 6-OHDA. 180
The next day, the compounds are added to the medium and zebrafish larvae are incubated 181
for 24h. Finally, the behavioural analysis is conducted with a video tracking system and 182
drugs are selected for their capability to improve the motor performance of 6-OHDA-183
lesioned zebrafish larvae. 184
185
Behavioural analysis 186
Spontaneous locomotion was recorded using the DanioVision™ (Noldus Information 187
Technology, the Netherlands) automated tracking system for zebrafish larvae. Larvae 188
were allowed to swim freely in a 96-well plate (1 larva/well) with 1200 µl of EM + 10 189
mM HEPES and their swimming activity was tracked for 90 minutes, under 10 min light-190
dark cycles. 30 min acclimatization period was done followed by 60 min test. The 191
acquired track data was analysed using the Ethovision X.T. 10 software (Noldus, 192

10
Wageningen, Netherlands). Only swimming activity obtained in the dark periods (i.e. 193
under infrared light) were analysed (Esch et al., 2012). The parameters automatically 194
measured were total distance moved (mm), burst swimming (number of times larvae 195
reached velocities higher than 25 mm/s) and immobile events (mean time spent moving 196
less than 2 mm/s divided by number of events; protocol adapted from noldus technical 197
specifications (http://www.noldus.com/EthoVision-XT/Gathering-data; 198
http://www.noldus.com/animal-behavior-research/solutions/research-small-lab-199
animals/open-field-set)). To filter system noise, 0.2 mm was defined as the minimum 200
distance of movement. All values were normalized as a percentage of the mean of the 201
healthy control. 202
203
Dose-response curves 204
Dose-response curves were determined by adding to 6-OHDA-lesioned larvae different 205
doses of the compound under test (n= 8 larvae per condition). Dopaminergic lesion 206
induction, exposure to the compounds and behavioural analysis were conducted as 207
described above. Exceptionally, to depict L-dopa curve, larvae were treated 30 min prior 208
to the behavioural evaluation. The parameters were normalized as a percentage of the 209
mean of healthy control. Anti-TH immunostaining was performed after behavioural 210
evaluation to test the viability of dopaminergic neurons in zebrafish larvae treated with 211
an effective dose of the compound. 212
213
Screening of the library of FDA-approved compounds 214
To screen for compounds that can rescue 6-OHDA-induced motor impairments in 215
zebrafish, the treatment protocol and behavioural analysis were performed as described 216
above. 6-OHDA-lesioned larvae treated with 1250 µM of L-dopa were concomitantly 217
tested as positive control. L-dopa treated larvae were exposed to freshly prepared L-dopa, 218
30 min prior to behavioural evaluation. All experimental conditions (n= 8 larvae per 219
condition) were performed at 1% DMSO and with compounds at 25 µM, labelled with a 220
predefined code by an investigator not involved in the analysis, to ensure the blinding 221
procedure. Healthy larvae treated with vehicle and untreated 6-OHDA-lesioned larvae 222
were also exposed to 1% DMSO, a non-toxic concentration to zebrafish (Hallare et al., 223

11
2006). Compounds were considered as a positive hit, when there was a statistically 224
significant recovery of motor performance (total distance moved and burst swimming) in 225
the 3 independent experiments performed. 226
227
Evaluation of repositioning potential 228
After the first round of screenings, the feasibility of repositioning of the compounds that 229
rescued motor performance of 6-OHDA-lesioned zebrafish larvae was evaluated by an 230
investigator not involved in the screening experiments. Relevant data about the selected 231
molecules was searched in publicly available scientific databases and revised. All 232
compounds with active intellectual property, prior art for PD, prone to off-label use (e.g. 233
vitamins) and with safety concerns were excluded from further screenings. The 234
compounds that rescued motor performance in the three independent rounds of screenings 235
and feasible for repositioning were further investigated. Previous medical indications, 236
targets and BBBpermeability were evaluated. 237
238
Whole mount immunostaining and confocal microscopy 239
Whole mount immunostaining in zebrafish was performed as previously described (Wang 240
et al., 2011) with modifications. Zebrafish larvae were fixed overnight at 4 °C in 4% 241
paraformaldehyde (PFA). Larvae were then gradually dehydrated to methanol (Cat# 242
A3493) 100% and stored at −20 °C. For whole mount immunostaining, larvae were 243
gradually rehydrated to phosphate buffer saline (PBS). The tissue was then permeated in 244
100% acetone (Cat# 211007) for 15 min at -20°C, washed with 0.5% PBS-Triton X-100 245
and blocked in blocking solution (1% bovine serum albumin in PBS with 1% DMSO and 246
0.05% Triton X-100) for 2 hours at room temperature. Whole mount tissues were 247
incubated overnight at 4 ºC with anti-TH primary antibody (1:200 in blocking solution; 248
ImmunoStar Cat# 22941), washed in 0.1% PBS-Triton X-100, PBS and re-incubated 249
overnight at 4 ºC with AlexaFluor 568 secondary antibody (1:1000 in blocking solution; 250
ThermoFisher Scientific Cat# A-11004). After staining, larvae were washed in PBS and 251
flat-mounted on a fluorescent mounting medium with DABCO, under a stereoscope. Z-252
stack compositions of the dopaminergic diencephalic cluster were acquired in a confocal 253
microscope (Zeiss LSM 510 META, Carl Zeiss MicroImaging, Göttingen, Germany) 254
with 40x magnification. Dopaminergic cell content was assessed by counting the number 255
of TH+ cells from average intensity projections with Image J software (Schneider et al., 256

12
2012), as described by (Wang et al., 2011). The zebrafish dopaminergic diencephalic 257
cluster was outlined according to (Kastenhuber et al., 2010; Tay et al., 2011) (Figure 3D). 258
The number of TH+ neurons ranged between X and X in the 6-OHDA-lesioned zebrafish 259
larvae as compared to Y and Y in the healthy larvae. Results are expressed as a percentage 260
of the mean of TH+ cells in healthy controls. 261
262
Statistical analysis 263
Data analysis and graphical representation were performed using Prism 5 software 264
(GraphPad Software, Inc., San Diego, CA, USA). All values were normalized as 265
percentage of the mean of healthy control, because substantial variability was evidenced 266
in the behaviour of zebrafish larvae (previously described at this developmental stage 267
(Farrell et al., 2011). This variability was also observed in the number of TH+ neurons in 268
the dopaminergic diencephalic cluster, as well as, in the extent of the lesion induced by 269
different lots of 6-OHDA. This variability probably resulted from the instability of 6-270
OHDA, which is highly sensitive to light and easily oxidized. Values presented are mean 271
± s.e.m. of n animals. All statistical tests used were two-tailed and chosen according to 272
the distribution of the data. Mean comparisons between the different groups and 6-273
OHDA-lesioned untreated larvae were performed using one-way ANOVA with 274
Dunnett’s post-hoc test for experiments independently replicated or Kruskal-Wallis 275
ANOVA with Dunn’s post-hoc test for experiments performed once. Difference was 276
considered significant when P value < 0.05. 277
278

13
Results 279
L-dopa, rasagiline and isradipine rescue bradykinesia in zebrafish larvae 280
6-OHDA-lesioned zebrafish present bradykinetic-like behaviour that can be depicted 281
from quantification of the total distance moved (Feng et al., 2014). To further characterize 282
the motor impairments in this model, we calculated the number of times that larvae 283
reached velocities corresponding to escaping behaviour (>25 mm/s, burst swimming). 284
This parameter allows the evaluation of motor fitness. After 2 days of exposure to 6-285
OHDA, lesioned larvae exhibit a decrease of the burst swimming when compared to 286
healthy larvae, as well as, a reduction of the total distance moved (Fig. 2). We then 287
assessed whether, in this model, the motor impairments could be rescued by L-dopa (the 288
most effective treatment for PD to date, (Oertel and Schulz, 2016)), rasagiline (showed 289
disease-modifying properties in preclinical models of PD and induces motor 290
improvement in patients with PD (Oertel and Schulz, 2016)) and isradipine (in phase III 291
clinical trials as a disease-modifying agent for PD (Oertel and Schulz, 2016)). To 292
determine the effective doses, 6-OHDA-lesioned larvae were incubated with different 293
concentrations of each compound and the dose-response curves were outlined. First, we 294
tested a larger range of concentrations and determined the LD50 of each drug. The LD50 295
of L-dopa, rasagiline and isradipine on 6-OHDA-lesioned zebrafish larvae was above 296
5000, 150 and 40 µM, respectively. Then, we tested concentrations around the optimal 297
dose. As shown by the dose-response curve, L-dopa rescued the total distance moved at 298
125 and 1250 µM (Supplementary fig. S2A), but only at 125 µM there was rescue of the 299
burst swimming (Supplementary fig. S2B). Rasagiline was effective at lower 300
concentrations, 0.8 and 0.9 µM rescued the total distance moved and burst swimming 301
(Supplementary fig. S2C and D). Isradipine showed a wider range of effective doses, 0.04 302
µM of this drug rescued the total distance moved, but did not affect the burst swimming 303
(Supplementary fig. S2E). Additionally, several concentrations between 0.08 and 0.8 µM 304
rescued both parameters, with a peak effective dose at 0.5 µM (Supplementary fig. S2E 305
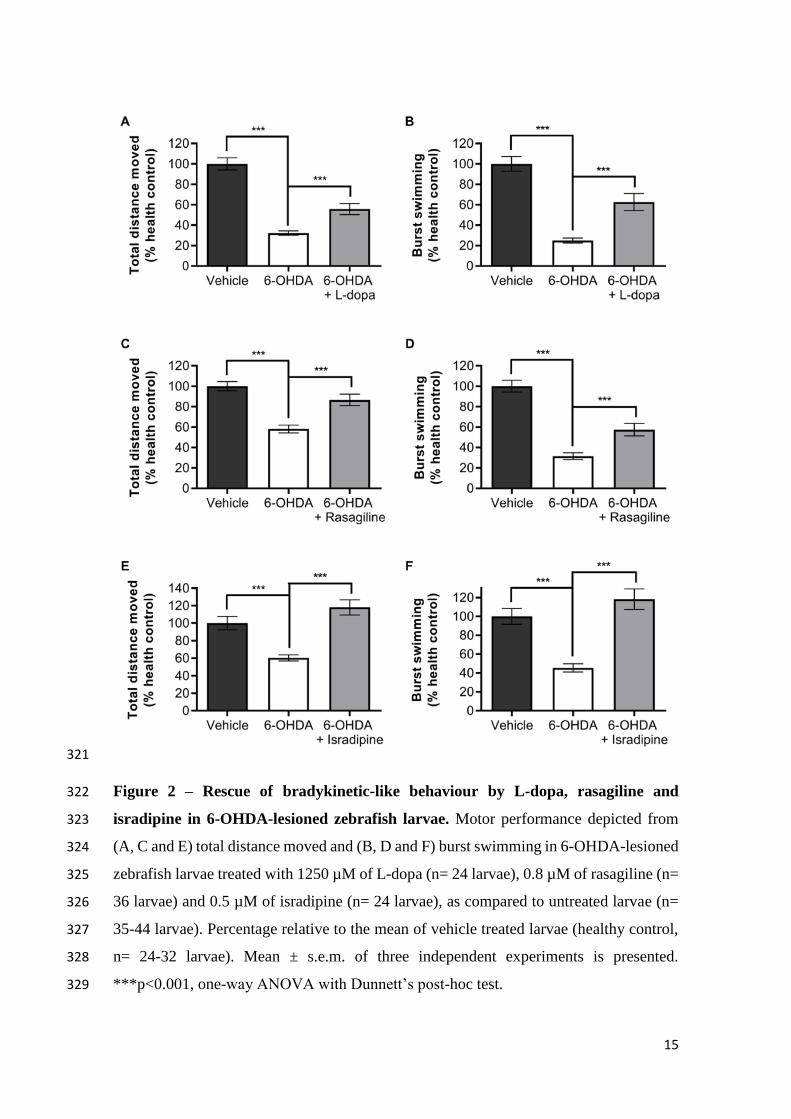
and F). To confirm the effective doses obtained from this data, three independent 306
experiments were performed with each of the compounds at two different concentrations, 307
125 and 1250 µM for L-dopa, 0.8 and 0.9 µM for rasagiline and 0.04 and 0.5 µM of 308
isradipine. 0.8 µM and 0.5 µM were confirmed to be the effective doses of rasagiline (Fig. 309
2C and D) and isradipine (Fig. 2E and F), respectively, as highlighted by the dose-310
response curves of each compound. In turn, while the dose of 125 µM was highlighted 311

14
by the dose-response curves of L-dopa, the dose of 1250 µM revealed more consistent 312
rescue of motor performance of 6-OHDA-lesioned zebrafish larvae in the three 313
independent experiments performed. The specificity of the motor effects induced by L-314
dopa, rasagiline and isradipine was then confirmed, as none of the compounds induced 315
behavioural changes in healthy zebrafish larvae (Supplementary fig. S3). This data shows 316
that the bradykinetic-like behaviour in 6-OHDA-lesioned zebrafish larvae can be 317
recovered by antiparkinsonian compounds, namely L-dopa, rasagiline and isradipine, and 318
that the effects observed are specific. 319
320
15
321
Figure 2 – Rescue of bradykinetic-like behaviour by L-dopa, rasagiline and 322
isradipine in 6-OHDA-lesioned zebrafish larvae. Motor performance depicted from 323
(A, C and E) total distance moved and (B, D and F) burst swimming in 6-OHDA-lesioned 324
zebrafish larvae treated with 1250 µM of L-dopa (n= 24 larvae), 0.8 µM of rasagiline (n= 325
36 larvae) and 0.5 µM of isradipine (n= 24 larvae), as compared to untreated larvae (n= 326
35-44 larvae). Percentage relative to the mean of vehicle treated larvae (healthy control, 327
n= 24-32 larvae). Mean ± s.e.m. of three independent experiments is presented. 328
***p<0.001, one-way ANOVA with Dunnett’s post-hoc test. 329

16
330
331
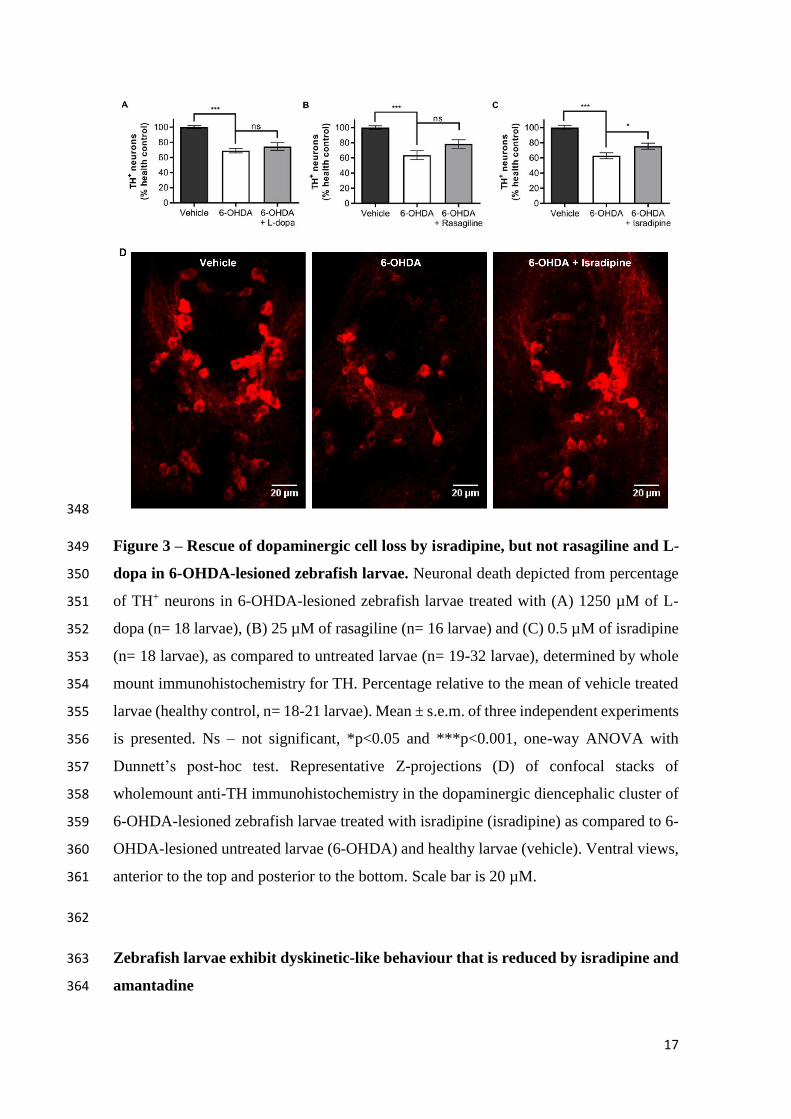
Isradipine rescues dopaminergic cell loss in zebrafish larvae 332
Previous studies demonstrated neuronal loss in the dopaminergic diencephalic cluster of 333
larvae lesioned with 6-OHDA (Feng et al., 2014). Therefore, we explored disease-334
modifying properties of L-dopa, rasagiline and isradipine in this model. Larvae were 335
treated with each compound at the effective dose and, after behavioural analysis, the 336
cellular content in the dopaminergic diencephalic cluster was determined by 337
immunohistochemistry against TH. 6-OHDA-lesioned larvae presented a reduction in the 338
number of dopaminergic cells, that was partially recovered by 0.5 µM of isradipine (Fig. 339
3C and D). On the contrary, neither L-dopa (Fig. 3A), nor rasagiline showed disease-340
modifying properties in this zebrafish model, at the optimal dose. Considering that, in 341
rats, rasagiline shows disease-modifying properties (Blandini et al., 2004), in addition to 342
the optimal dose, we also tested this compound at the concentration used in the phenotypic 343
screening, 25 µM, but no statistically significant cell recovery was observed (Fig. 3B). 344
This result indicates that the 6-OHDA-lesioned zebrafish larvae has a limited predictive 345
value to test disease-modifying agents for PD. 346
347
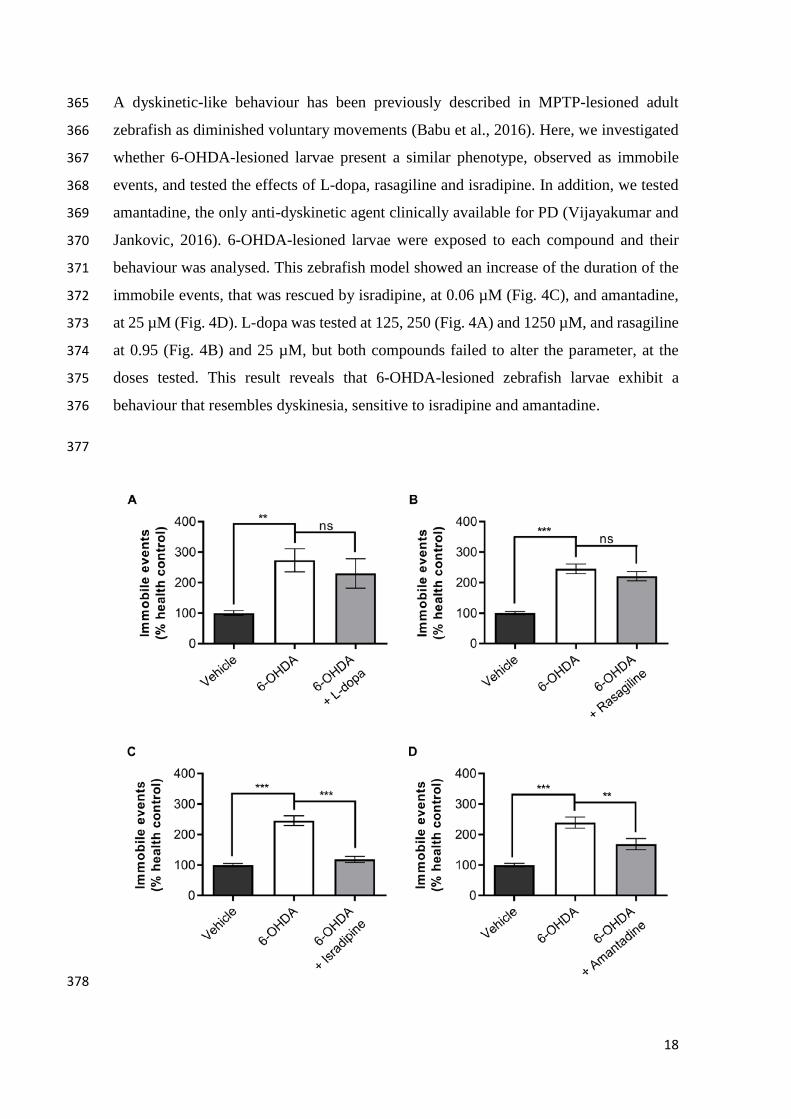
17
348
Figure 3 – Rescue of dopaminergic cell loss by isradipine, but not rasagiline and L-349
dopa in 6-OHDA-lesioned zebrafish larvae. Neuronal death depicted from percentage 350
of TH+ neurons in 6-OHDA-lesioned zebrafish larvae treated with (A) 1250 µM of L-351
dopa (n= 18 larvae), (B) 25 µM of rasagiline (n= 16 larvae) and (C) 0.5 µM of isradipine 352
(n= 18 larvae), as compared to untreated larvae (n= 19-32 larvae), determined by whole 353
mount immunohistochemistry for TH. Percentage relative to the mean of vehicle treated 354
larvae (healthy control, n= 18-21 larvae). Mean ± s.e.m. of three independent experiments 355
is presented. Ns – not significant, *p<0.05 and ***p<0.001, one-way ANOVA with 356
Dunnett’s post-hoc test. Representative Z-projections (D) of confocal stacks of 357
wholemount anti-TH immunohistochemistry in the dopaminergic diencephalic cluster of 358
6-OHDA-lesioned zebrafish larvae treated with isradipine (isradipine) as compared to 6-359
OHDA-lesioned untreated larvae (6-OHDA) and healthy larvae (vehicle). Ventral views, 360
anterior to the top and posterior to the bottom. Scale bar is 20 µM. 361
362
Zebrafish larvae exhibit dyskinetic-like behaviour that is reduced by isradipine and 363
amantadine 364
18
A dyskinetic-like behaviour has been previously described in MPTP-lesioned adult 365
zebrafish as diminished voluntary movements (Babu et al., 2016). Here, we investigated 366
whether 6-OHDA-lesioned larvae present a similar phenotype, observed as immobile 367
events, and tested the effects of L-dopa, rasagiline and isradipine. In addition, we tested 368
amantadine, the only anti-dyskinetic agent clinically available for PD (Vijayakumar and 369
Jankovic, 2016). 6-OHDA-lesioned larvae were exposed to each compound and their 370
behaviour was analysed. This zebrafish model showed an increase of the duration of the 371
immobile events, that was rescued by isradipine, at 0.06 µM (Fig. 4C), and amantadine, 372
at 25 µM (Fig. 4D). L-dopa was tested at 125, 250 (Fig. 4A) and 1250 µM, and rasagiline 373
at 0.95 (Fig. 4B) and 25 µM, but both compounds failed to alter the parameter, at the 374
doses tested. This result reveals that 6-OHDA-lesioned zebrafish larvae exhibit a 375
behaviour that resembles dyskinesia, sensitive to isradipine and amantadine. 376
377
378

19
Figure 4 – Rescue of dyskinetic-like behaviour by isradipine and amantadine, but 379
not L-dopa and rasagiline in 6-OHDA-lesioned zebrafish larvae. Dyskinetic-like 380
behaviour depicted from immobile events in 6-OHDA-lesioned zebrafish larvae treated 381
with (A) 250 µM of L-dopa (n= 30 larvae), (B) 0.95 µM of rasagiline (n= 31 larvae), (C) 382
0.06 µM of isradipine (n= 26 larvae) and (D) 25 µM of amantadine (n= 23 larvae), as 383
compared to untreated larvae (n= 44-49 larvae). Percentage relative to the mean of vehicle 384
treated larvae (healthy control, n= 23-31 larvae). Mean ± s.e.m. of three independent 385
experiments is presented. Ns – not significant, **p<0.01 and ***p<0.001, one-way 386
ANOVA with Dunnett’s post-hoc test. 387
388
A phenotypic-based screen identifies compounds with antiparkinsonian potential 389
To select candidate compounds capable of rescuing motor impairments in zebrafish 390
larvae lesioned with 6-OHDA, we developed a phenotypic assay that enabled the 391
screening of a library of 1600 FDA approved drugs (Fig. 1). The assay was three days 392
long and encompassed dopaminergic lesion with 6-OHDA (0h-24h), incubation with the 393
screening compounds (24h-48h) and behavioural evaluation (48h-72h). The locomotor 394
behaviour was tested by automatic measurement of total distance moved and burst 395
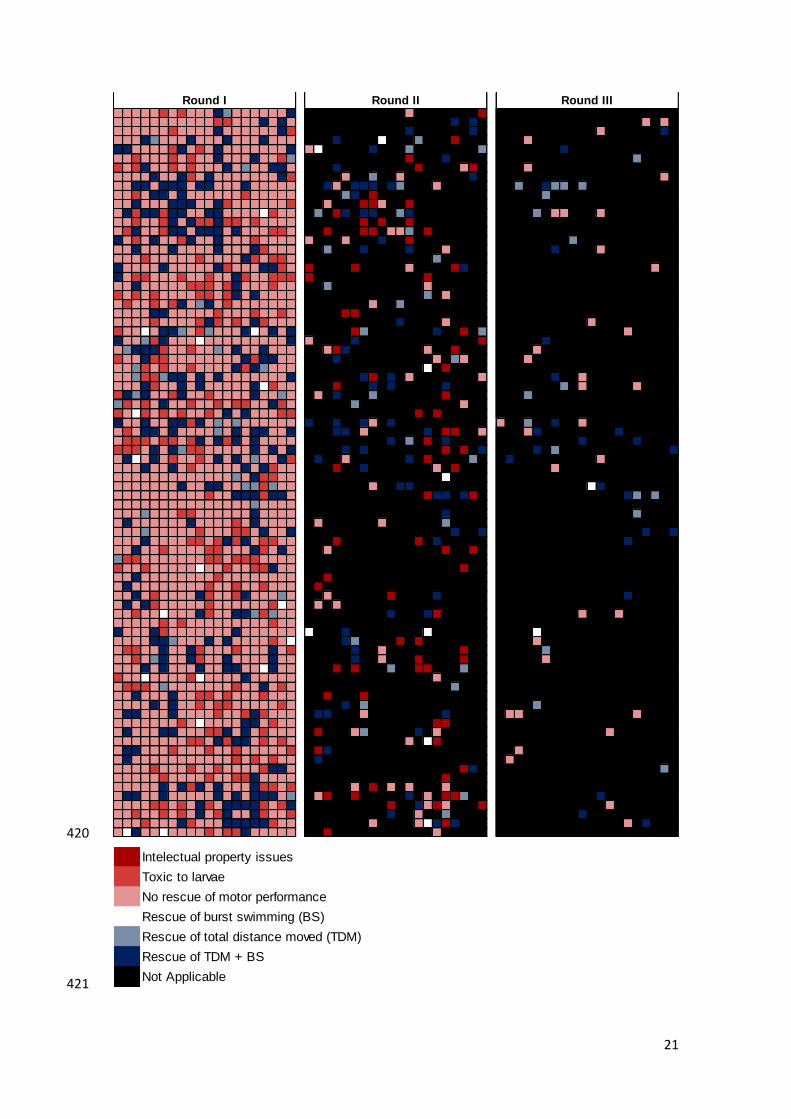
swimming, as previously described. From the 1600 compounds screened (Fig. 5), 258 396
(16%) rescued motor impairments in 6-OHDA-lesioned zebrafish larvae during the first 397
round of experiments performed (Table 1). These compounds were then evaluated for 398
repositioning and 83 (32%) were excluded from further analysis, based on pre-determined 399
exclusion criteria. From these 83 compounds, 26 (31%) had prior art for PD, 25 (30%) 400
had an active patent for other indications or were prone to off-label use, and 7 (8%) raised 401
safety concerns (Table 2). Nevertheless, these compounds were useful for validation of 402
the screen. This was the case, for example, of caffeine and carbinoxamine maleate, which 403
were blindly selected during the screen for rescuing motor impairments in 6-OHDA-404
lesioned zebrafish larvae. In contrast, bromocriptine mesylate and apomorphine 405
hydrochloride, two dopamine agonists used in the clinic, were also screened, but showed 406
no effect in the 6-OHDA-lesioned zebrafish larvae at the concentration tested. In the 407
second and third rounds of screenings, 78 (30%) and 23 (29%) compounds, respectively, 408
rescued motor impairments in 6-OHDA-lesioned zebrafish larvae (Table 1). After further 409
analysis, which included the evaluation of previous medical indications and targets and 410

20
BBB permeability, 3 drugs (13%), stavudine, tapentadol and nabumetone, showed to be 411
particularly promising for further experiments (Table 3). These three compounds had 412
minimally explored targets for the treatment of PD, high permeability to the BBB and no 413
further concerns from previous indications. The other 20 compounds had either no known 414
targets, low or non-described BBB permeability or concerns regarding safety, and were 415
discarded. Overall, the screening protocol developed was suitable for a quick selection of 416
neuroactive drugs with antiparkinsonian potential, but also presented limitations 417
concerning the activity of dopamine agonists. 418
419
21
420
421
Round I Round II Round III
-3 Intelectual property issues
-2 Toxic to larvae
-1 No rescue of motor performance
0 Rescue of burst swimming (BS)
1 Rescue of total distance moved (TDM)
2 Rescue of TDM + BS
Not Applicable

22
Figure 5 – Hit map representation of the screening of 1600 bioactive drugs. Each 422
square corresponds to a compound, identified as rescuing none of the parameters 423
measured (burst swimming, BS, and total distance moved, TDM), rescuing one of the two 424
parameters measured or rescuing both parameters measured in 6-OHDA-lesioned 425
zebrafish larvae. Only compounds that rescued both, BS and TDM, are represented in the 426
subsequent round of screenings. Compounds that presented toxic effects on zebrafish 427
larvae and intellectual property concerns, and therefore excluded from further screenings 428
are also represented. 429
430
431
Table 1 – Overview results of the screening of 1600 bioactive drugs. Number and 432
percentage of compounds with no effects, and with effects on one or both parameters 433
(burst swimming, BS, and total distance moved, TDM) measured during each round of 434
the screening assay, to depict motor performance in 6-OHDA-lesioned zebrafish larvae. 435
Number and percentage of compounds with toxic effects on zebrafish larvae and 436
intellectual property concerns also indicated. Percentage relative to the total of 437
compounds screened (1600) for round I and to the number of positive hits in the previous 438
round, for rounds II and III. 439
440
441
Table 2 – Intellectual property issues considered for exclusion of positive hits from 442
round I of phenotypic screenings. Number and percentage of compounds with prior art 443

23
for PD, active patent, prone to off-label use and safety concerns. Percentage relative to 444
the total of compounds selected for rescuing motor performance in 6-OHDA-lesioned 445
zebrafish larvae during round I of screenings. 446
447

24
448
Hits Targets BBB permeability
1 Human immunodeficiency virus type 1 protease inhibitor
Nuclear receptor subfamily 1 group I member 2 activator
Low
2 Tubulin beta-1 chain
Apoptosis regulator Bcl-2
Microtubule-associated protein 2 and 4
Microtubule-associated protein tau
Nuclear receptor subfamily 1 group I member 2
High
3 Not found High
4 Sodium channel protein type 5 subunit alpha antagonist High
5 Not found Not found
6 cAMP and cGMP-specific 3',5'-cyclic phosphodiesterases inhibitor
Adenosine deaminase inhibitor
Calcipressin-1
Alpha-1-acid glycoprotein 1
Low
7 Not found Not found
8 5-hydroxytryptamine receptors agonist
Alpha-2A adrenergic receptor agonist
High
9 Cross-linking/alkylation of DNA
Nuclear receptor subfamily 1 group I member 2
High
10 Arachidonate 5-lipoxygenase inhibitor
Prostaglandin G/H synthase 1 inhibitor
High
11 Prostaglandin G/H synthase 1 and 2 inhibitor High
12 Not found Not found
13 Reverse transcriptase/RNaseH inhibitor High
14 Mu-type opioid receptor agonist
Sodium-dependent noradrenaline transporter inhibitor
Kappa-type opioid receptor
Delta-type opioid receptor
5-hydroxytryptamine receptor 3A
Sodium-dependent serotonin transporter inhibitor
High
15 Alpha-1A, 1D and 1B adrenergic receptor antagonist Low
16 3 beta-hydroxysteroid dehydrogenase/Delta 5-->4-isomerase type 1 and 2 inhibitor
Estrogen receptor alpha and beta allosteric modulator
Low
17 Not found Not found
18 Not found Not found
19 Beta-1 and 2 adrenergic receptor antagonist
Alpha-1A, 1B, 1D, 2A-2C adrenergic receptor antagonist
NADH dehydrogenase [ubiquinone] 1 subunit C2 inhibitor
Vascular endothelial growth factor A
Natriuretic peptides B
Gap junction alpha-1 protein
Potassium voltage-gated channel subfamily H member 2 inhibitor
Vascular cell adhesion protein 1 inhibitor
E-selectin inhibitor
Hypoxia-inducible factor 1-alpha modulator
Inward rectifier potassium channel 4
Low
20 Penicillin binding protein 2a, 1A and 1B inhibitor
Peptidoglycan synthase FtsI inhibitor
D-alanyl-D-alanine carboxypeptidase DacA and DacC inhibitor
High
21 Muscarinic acetylcholine receptor M1 antagonist High
22 Not found Not found
23 Potassium voltage-gated channel subfamily H member 2 Not found

25
Table 3 – Overview of the 23 lead compounds of the screening assay. Targets and 449
BBB permeability of the 23 compounds selected for rescuing motor performance in 6-450
OHDA-lesioned zebrafish larvae during the screening assay. Data obtained from an initial 451
search at https://www.drugbank.ca/. 452
453
454
Three new compounds display antiparkinsonian activity in 6-OHDA-lesioned 455
zebrafish larvae 456
Stavudine is a nucleoside reverse transcriptase inhibitor, that stimulates the production of 457
brain derived neurotrophic factor (BDNF) (Renn et al., 2011), tapentadol is a µ-opioid 458
receptor agonist and norepinephrine reuptake inhibitor (Tzschentke et al., 2007) and 459
nabumetone is a cyclooxygenase enzyme inhibitor (Davies, 1997). These three 460
compounds were selected from the screening assay, for their antiparkinsonian properties 461
in 6-OHDA-lesioned zebrafish larvae (Supplementary fig. S4). Next, we determined the 462
effective dose of each of the purified compounds and evaluated their effect on dyskinetic-463
like behaviour. 6-OHDA-lesioned larvae were treated with different concentrations of 464
each compound and behavioural analysis was conducted. For both parameters, total 465
distance moved and burst swimming, we observed a significant recovery of larvae treated 466
with 50 µM of stavudine (Fig. 6A and B), 49 µM of tapentadol (Fig. 6D and E) and 0.9 467
µM of nabumetone (Fig. 6G and H). Stavudine and tapentadol also rescued dyskinetic-468
like behaviour at these doses, as opposed to nabumetone, in 6-OHDA-lesioned zebrafish 469
larvae (Fig. 6C, F and I). This data confirms that stavudine, tapentadol and nabumetone 470
are potential antiparkinsonian agents and supports further studies in mammalian models 471
of PD. 472
473

26
474
Figure 6 – Rescue of motor impairments by stavudine, tapentadol and nabumetone 475
in 6-OHDA-lesioned zebrafish larvae. Motor performance and dyskinetic-like 476
behaviour depicted from (A, D and G) total distance moved, (B, E and H) burst swimming 477
and (C, F and I) immobile events in 6-OHDA-lesioned zebrafish larvae treated with 50 478
µM of stavudine (n= 38-40 larvae), 49 µM of tapentadol (n= 32-46 larvae) and 0.9 µM 479
of nabumetone (n= 38-49 larvae), as compared to untreated larvae (n= 30-66 larvae). 480
Percentage relative to the mean of vehicle treated larvae (healthy control, n= 23-31 481
larvae). Mean ± s.e.m. of three independent experiments is presented. Ns – not significant, 482
and ***p<0.001, one-way ANOVA with Dunnett’s post-hoc test. 483
484

27
Discussion 485
Here, we developed a phenotypic screening assay for the selection of new drugs with 486
antiparkinsonian potential, using 6-OHDA-lesioned zebrafish larvae. This model exhibits 487
death of dopaminergic neurons, accompanied by a decrease of dopamine levels and by 488
motor impairments (Anichtchik et al., 2004; Feng et al., 2014; Vijayanathan et al., 2017). 489
These phenotypes can be rescued by antiparkinsonian compounds (L-dopa and rasagiline) 490
(Cronin and Grealy, 2017; Feng et al., 2014). After deeper characterization of this model, 491
we used it for screening of 1600 FDA approved drugs and identified 23 drugs with 492
antiparkinsonian potential. We selected stavudine, tapentadol and nabumetone due to 493
their effects on relevant targets for the treatment of PD. 494
In agreement with previous studies (Anichtchik et al., 2004; Cronin and Grealy, 2017; 495
Feng et al., 2014), we observed that zebrafish larvae exposed to 6-OHDA display 496
bradykinetic-like behaviour and loss of dopaminergic neurons. We also introduce the 497
burst swimming as a suitable parameter to assess motor performance. This parameter 498
measures the number of times that larvae reach velocities correspondent to escaping 499
behaviour (>25 mm/s). Burst swimming has been catalogued as a swimming behaviour 500
of zebrafish larvae, which exhibit locomotion at slow speed, sometimes followed by a 501
faster and more vigorous swimming, that is typically above 20 mm/s (Budick and 502
O’Malley, 2000; Kalueff et al., 2013). Importantly, we showed that burst swimming can 503
be rescued by L-dopa and rasagiline, as previously demonstrated for total distance moved 504
(Cronin and Grealy, 2017; Feng et al., 2014). 505
Since isradipine has been described as a disease-modifying agent for PD (Chan et al., 506
2007; Ilijic et al., 2011), we also explored the antiparkinsonian potential of this drug. We 507
report, for the first time, that isradipine reduces the loss of dopaminergic neurons and 508
rescues bradykinetic-like behaviour in 6-OHDA-lesioned zebrafish larvae. It has been 509
suggested that isradipine (calcium channel blocker) prevents the death of dopaminergic 510
cells through the reduction of calcium influx in neurons and consequently the decrease of 511
mitochondrial activity (Chan et al., 2007). Therefore, isradipine could have induced 512
changes in the mitochondrial activity of zebrafish larvae which decreased the oxidative 513
stress and cytotoxicity induced by 6-OHDA over dopaminergic neurons. In contrast, we 514
did not detect disease-modifying effect of rasagiline. This finding differs from previous 515
reports in zebrafish and mice (Blandini et al., 2004; Cronin and Grealy, 2017), likely due 516

28
to differences in the treatment protocols adopted. The time between dopaminergic lesion 517
and the exposure to rasagiline is a key factor that may dramatically influence the outcome 518
in the zebrafish model, where the regeneration capacity is greater in comparison to 519
mammals (Zupanc, 2008). In fact, rasagiline failed to demonstrate disease-modifying 520
properties in humans and isradipine is still under clinical evaluation (Oertel and Schulz, 521
2016; Olanow et al., 2009). 522
Previously, dyskinetic-like behaviour in MPTP-lesioned adult zebrafish was reported 523
(Babu et al., 2016). Now, we observed that 6-OHDA-lesioned zebrafish larvae exhibit a 524
similar behaviour, as measured by the average duration of immobile events (velocity <2 525
mm/s). While the evaluation of an hyperkinetic disease through complete lack of 526
movement might appear counter-intuitive, there is already evidence supporting this 527
approach in a zebrafish model of dystonia (Friedrich et al., 2012). On the other hand, 528
dyskinesia only occurs in PD patients under L-dopa or dopamine-agonist treatment. 529
Therefore, although the parameter we measured cannot be directly related with L-dopa-530
induced dyskinesia, it reports on non-induced dyskinesias, indicative of abnormal 531
plasticity of the motor circuitry. This could result from the dysregulation of different 532
neurotransmitters, sensitive to the impairment of the dopaminergic system, that are 533
essential for generating a normal swimming behaviour. Consistently, L-dopa and 534
rasagiline, two drugs that rely on the dopaminergic system, did not alter the average 535
duration of immobile events in 6-OHDA-lesioned zebrafish larvae. Accordingly, these 536
drugs do not have an anti-dyskinetic effect in humans (Pistacchi et al., 2014). In turn, one 537
would expect this parameter to worsen in larvae treated with L-dopa. However, this may 538
only be observed with chronic treatment, so it could not be assessed. Additionally, we 539
found that the drugs with non-dopaminergic targets, isradipine and amantadine, reduced 540
the average duration of immobile events. Amantadine is widely used in the clinic as an 541
anti-dyskinetic agent for PD (Vijayakumar and Jankovic, 2016). In the case of isradipine, 542
the pre-clinical evidence for anti-dyskinetic properties is limited (Rylander et al., 2009; 543
Schuster et al., 2009) and reports of this therapeutic indication in humans do not exist to 544
date. Whereas, amantadine is an NMDA antagonist and could have balanced the 545
dysregulation induced by the impaired dopaminergic system in zebrafish, isradipine does 546
not act in a specific neuronal circuitry. Therefore, the changes observed in the duration 547
of immobile events of zebrafish larvae treated with isradipine could have resulted from 548
the rescue of dopaminergic neurons and, consequently, from a lower dysregulation of this 549

29
and other neuronal systems. Additional studies will be necessary to further investigate the 550
predictive value of this parameter. As such, the phenotypic screening assay we developed 551
relied on the total distance moved and burst swimming parameters for the selection of 552
compounds with antiparkinsonian potential. 553
The phenotypic screening of 1600 FDA approved drugs, resulted in the identification of 554
26 drugs (31% of the drugs selected during round I) with prior art for PD, revealing the 555
sensitivity of the assay. In contrast, the screening set up failed to identify some dopamine 556
agonists used as monotherapy or in combination with L-dopa (Oertel and Schulz, 2016). 557
In zebrafish, eight proteins homologous to the human dopamine receptors have been 558
identified (Panula et al., 2010), but their affinity for dopamine agonists has not been 559
explored. Therefore, one possibility is that the dose tested during the screening was not 560
effective. Previously, it was reported that the non-selective dopamine agonist, 561
apomorphine, has a biphasic effect on the locomotor activity of zebrafish, that depends 562
on the dose applied (Irons et al., 2013). This dose-dependent effect has also been 563
evidenced in mice treated with dopamine receptor agonists (Lundblad et al., 2005). All 564
drugs were screened at a concentration of 25 µM, the highest concentration more 565
commonly used for drug screenings in zebrafish (Rennekamp and Peterson, 2015). 566
Importantly, only 16% of the screened drugs showed toxic effects in these conditions, 567
which did not compromise the screening. 568
Overall, the phenotypic screening identified 23 drugs (1.4% of the total drugs screened) 569
with antiparkinsonian potential in 6-OHDA-lesioned zebrafish larvae. Previous studies 570
allowed the analysis of targets, BBB permeability and clinical indications, and selection 571
of three drugs that were considered particularly interesting for further studies. Stavudine 572
is a reverse transcriptase inhibitor and previous reports have described the stimulation of 573
BDNF expression by this drug (Renn et al., 2011). Neurotrophic factors have proven their 574
efficacy in animal models of PD, but their inability to cross the BBB has limited their 575
application (Aron and Klein, 2011). In turn, tapentadol is a µ-opioid receptor agonist and 576
norepinephrine reuptake inhibitor (Tzschentke et al., 2007), two pathways long 577
implicated in PD (Espay et al., 2014; Samadi et al., 2006). Lastly, nabumetone is a 578
prostaglandin synthase inhibitor with anti-inflammatory properties (Davies, 1997). 579
Neuroinflammation is a key pathogenic mechanism of PD and anti-inflammatory agents 580
have also shown promising results in preclinical studies, although convincing clinical data 581
is still missing (Athauda and Foltynie, 2015). Interestingly, the three drugs rescued 582

30
bradykinetic-like behaviour, but nabumetone failed to alter the duration of immobile 583
events in 6-OHDA-lesioned zebrafish larvae. As discussed above, the target described for 584
nabumetone is not specific of a neuronal circuitry, while stavudine induces the expression 585
of BDNF and tapentadol acts on the opioid and noradrenergic systems. Further studies 586
need to be conducted to determine the exact mechanism of action behind the therapeutic 587
potential of these three compounds and to validate their antiparkinsonian efficacy in other 588
vertebrate models. Since the effects of stavudine, tapentadol and nabumetone in the motor 589
performance of healthy zebrafish larvae were not assessed, further studies should 590
determine the specificity of the therapeutic effects to dopaminergic disfunction. In 591
contrast to other screenings previously reported (Buckley et al., 2010; Parng et al., 2006; 592
Robertson et al., 2016; Sun et al., 2012), our selection of drugs was completely agnostic 593
to target or mechanism of action, which is a key distinction of the phenotypic screening, 594
as it can result in the identification of drugs with new mechanisms of action. 595
596

31
Conclusions 597
In general, our study provides further demonstration that the 6-OHDA-lesioned zebrafish 598
larvae exhibit bradykinetic-like behaviours that are sensitive to the motor improvement 599
of antiparkinsonian compounds. A dyskinetic-like behaviour was also observed, but 600
further investigations will be necessary in order to fully demonstrate the predictive value 601
of this parameter in zebrafish. Although we did not detect disease-modifying properties 602
for rasagiline or anti-bradykinetic properties for dopamine agonists, this might be due to 603
the screening protocol, and will require further investigation. Nevertheless, the 604
phenotypic screening we describe is a valid strategy for rapid selection of potential 605
antiparkinsonian agents. Importantly, it is simple, is based in objective and automated 606
parameters, and does not require invasive or stressful manipulation of the animals. The 607
screening led to the identification of three neuroactive drugs, stavudine, tapentadol and 608
nabumetone, and further studies regarding their mechanism of action could lead to the 609
discovery of targets or pathways relevant for PD mechanisms. 610

32
Acknowledgements 611
Funding: This study was sponsored by TechnoPhage S.A. and Eurostars program 612
(ES#5553) from EUREKA (a program run by the European Commission). Rita L. Vaz 613
was supported by a grant (SFRH/BD/78077/2011) from Fundação para a Ciência e 614
Tecnologia. Tiago F. Outeiro was supported by the DFG Center for Nanoscale 615
Microscopy and Molecular Physiology of the Brain (CNMPB). 616
617
Conflict of interest: S.S., D.C. and N.D.A. were employees of Technophage SA, at the 618
time of the study. The other authors declare no conflicts of interest. 619
620
Author contributions: S.S., R.W., T.F.O. and N.D.A. did study conception and design. 621
R.L.V., D.C. and S.S. performed experiments, did data acquisition and analysed the data. 622
All authors interpreted the data. R.L.V. drafted the paper. S.S. and N.D.A. did critical 623
revision. 624
625

33
References 626
Anichtchik, O. V., Kaslin, J., Peitsaro, N., Scheinin, M., Panula, P., 2004. 627
Neurochemical and behavioural changes in zebrafish Danio rerio after systemic 628
administration of 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6-629
tetrahydropyridine. J. Neurochem. 88, 443–453. doi:10.1046/j.1471-630
4159.2003.02190.x 631
Aron, L., Klein, R., 2011. Repairing the parkinsonian brain with neurotrophic factors. 632
Trends Neurosci. 34, 88–100. doi:10.1016/j.tins.2010.11.001 633
Athauda, D., Foltynie, T., 2015. The ongoing pursuit of neuroprotective therapies in 634
Parkinson disease. Nat. Rev. Neurol. 11, 25–40. doi:10.1038/nrneurol.2014.226 635
Babu, N.S., Murthy, C.L.N., Kakara, S., Sharma, R., Swamy, C.V.B., Idris, M.M., 636
2016. 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine induced Parkinson’s disease 637
in zebrafish. Proteomics 16, 1407–1420. doi:10.1002/pmic.201500291 638
Blandini, F., Armentero, M.T., Fancellu, R., Blaugrund, E., Nappi, G., 2004. 639
Neuroprotective effect of rasagiline in a rodent model of Parkinson’s disease. Exp. 640
Neurol. 187, 455–459. doi:10.1016/j.expneurol.2004.03.005 641
Buckley, C.E., Marguerie, A., Roach, A.G., Goldsmith, P., Fleming, A., Alderton, 642
W.K., Franklin, R.J.M., 2010. Drug reprofiling using zebrafish identifies novel 643
compounds with potential pro-myelination effects. Neuropharmacology 59, 149–644
159. doi:10.1016/j.neuropharm.2010.04.014 645
Budick, S.A., O’Malley, D.M., 2000. Locomotor repertoire of the larval zebrafish: 646
swimming, turning and prey capture. J. Exp. Biol. 203, 2565–2579. 647
doi:10.1242/jeb.01529 648
Chan, C.S., Guzman, J.N., Ilijic, E., Mercer, J.N., Rick, C., Tkatch, T., Meredith, G.E., 649
Surmeier, D.J., 2007. “Rejuvenation” protects neurons in mouse models of 650
Parkinson’s disease. Nature 447, 1081–1086. doi:10.1038/nature05865 651
Chong, C., Zhou, Z., Razmovski-Naumovski, V., Cui, G., Zhang, L., Sa, F., Hoi, P., 652
Chan, K., Lee, S.M., 2013. Danshensu protects against 6-hydroxydopamine-653
induced damage of PC12 cells in vitro and dopaminergic neurons in zebrafish. 654

34
Neurosci. Lett. 543, 121–125. doi:10.1016/j.neulet.2013.02.069 655
Cronin, A., Grealy, M., 2017. Neuroprotective and Neuro-restorative Effects of 656
Minocycline and Rasagiline in a Zebrafish 6-Hydroxydopamine Model of 657
Parkinson’s Disease. Neuroscience 367, 34–46. 658
doi:10.1016/j.neuroscience.2017.10.018 659
Davies, N.M., 1997. Clinical pharmacokinetics of nabumetone. The dawn of selective 660
cyclo-oxygenase-2 inhibition? Clin. Pharmacokinet. 33, 404–416. 661
Dorsey, E.R., Constantinescu, R., Thompson, J.P., Biglan, K.M., Holloway, R.G., 662
Kieburtz, K., Marshall, F.J., Ravina, B.M., Schifitto, G., Siderowf, A., Tanner, 663
C.M., 2007. Projected number of people with Parkinson disease in the most 664
populous nations, 2005 through 2030. Neurology 68, 384–386. 665
doi:10.1212/01.wnl.0000247740.47667.03 666
Du, Y., Guo, Q., Shan, M., Wu, Y., Huang, S., Zhao, H., 2016. Spatial and Temporal 667
Distribution of Dopaminergic Neurons during Development in Zebrafish 10, 1–7. 668
doi:10.3389/fnana.2016.00115 669
Esch, C. De, Linde, H. Van Der, Slieker, R., Willemsen, R., Wolterbeek, A., 670
Woutersen, R., Groot, D. De, 2012. Locomotor activity assay in zebrafish larvae: 671
Influence of age, strain and ethanol. Neurotoxicol. Teratol. 34, 425–433. 672
doi:10.1016/j.ntt.2012.03.002 673
Espay, A.J., Lewitt, P.A., Kaufmann, H., 2014. Norepinephrine deficiency in 674
Parkinson’s disease: The case for noradrenergic enhancement. Mov. Disord. 29, 675
1710–1719. doi:10.1002/mds.26048 676
Farrell, T.C., Cario, C.L., Milanese, C., Vogt, A., Jeong, J.-H., Burton, E.A., 2011. 677
Evaluation of spontaneous propulsive movement as a screening tool to detect 678
rescue of Parkinsonism phenotypes in zebrafish models. Neurobiol. Dis. 44, 9–18. 679
doi:10.1016/j.nbd.2011.05.016 680
Feng, C., Wen, Z., Huang, S., Hung, H., Chen, C., Yang, S., Chen, N., Wang, H., Hsiao, 681
C., Chen, W., 2014. Effects of 6-hydroxydopamine exposure on motor activity and 682
biochemical expression in zebrafish (Danio rerio) larvae. Zebrafish 11, 227–239. 683
doi:10.1089/zeb.2013.0950 684

35
Flinn, L., Bretaud, S., Lo, C., Ingham, P.W., Bandmann, O., 2008. Zebrafish as a new 685
animal model for movement disorders. J. Neurochem. 106, 1991–1997. 686
doi:10.1111/j.1471-4159.2008.05463.x 687
Friedrich, T., Lambert, A.M., Masino, M.A., Downes, G.B., 2012. Mutation of 688
zebrafish dihydrolipoamide branched-chain transacylase E2 results in motor 689
dysfunction and models maple syrup urine disease. Dis. Model. Mech. 5, 248–258. 690
doi:10.1242/dmm.008383 691
Godoy, R., Noble, S., Yoon, K., Anisman, H., Ekker, M., 2015. Chemogenetic ablation 692
of dopaminergic neurons leads to transient locomotor impairments in zebrafish 693
larvae. J. Neurochem. 135, 249–260. doi:10.1111/jnc.13214 694
Hallare, A., Nagel, K., Köhler, H.R., Triebskorn, R., 2006. Comparative embryotoxicity 695
and proteotoxicity of three carrier solvents to zebrafish (Danio rerio) embryos. 696
Ecotoxicol. Environ. Saf. 63, 378–388. doi:10.1016/j.ecoenv.2005.07.006 697
Hirsch, E.C., Jenner, P., Przedborski, S., 2013. Pathogenesis of Parkinson’s disease. 698
Mov. Disord. 28, 24–30. doi:10.1002/mds.25032 699
Ilijic, E., Guzman, J.N., Surmeier, D.J., 2011. The L-type channel antagonist isradipine 700
is neuroprotective in a mouse model of Parkinson’s disease. Neurobiol. Dis. 43, 701
364–371. doi:10.1097/MPG.0b013e3181a15ae8.Screening 702
Irons, T.D., Kelly, P.E., Hunter, D.L., MacPhail, R.C., Padilla, S., 2013. Acute 703
administration of dopaminergic drugs has differential effects on locomotion in 704
larval zebrafish. Pharmacol. Biochem. Behav. 103, 792–813. 705
doi:10.1016/j.pbb.2012.12.010 706
Jeong, J., Kwon, H., Ahn, J., Kang, D., Kwon, S., Park, J.A., Kim, K., 2008. Functional 707
and developmental analysis of the blood-brain barrier in zebrafish. Brain Res. Bull. 708
75, 619–628. doi:10.1016/j.brainresbull.2007.10.043 709
Kalueff, A. V, Gebhardt, M., Stewart, A.M., Cachat, J.M., Brimmer, M., Chawla, J.S., 710
Craddock, C., Kyzar, E.J., Roth, A., Landsman, S., Gaikwad, S., Robinson, K., 711
Baatrup, E., Tierney, K., Shamchuk, A., Norton, W., Miller, N., Nicolson, T., 712
Braubach, O., Gilman, C.P., Pittman, J., Rosemberg, D.B., Gerlai, R., Echevarria, 713
D., Lamb, E., Neuhauss, S.C.F., Weng, W., Bally-Cuif, L., Schneider, H., 2013. 714

36
Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 715
10, 70–86. doi:10.1089/zeb.2012.0861 716
Kastenhuber, E., Kratochwil, C.F., Ryu, S., Schweitzer, J., Driever, W., 2010. Genetic 717
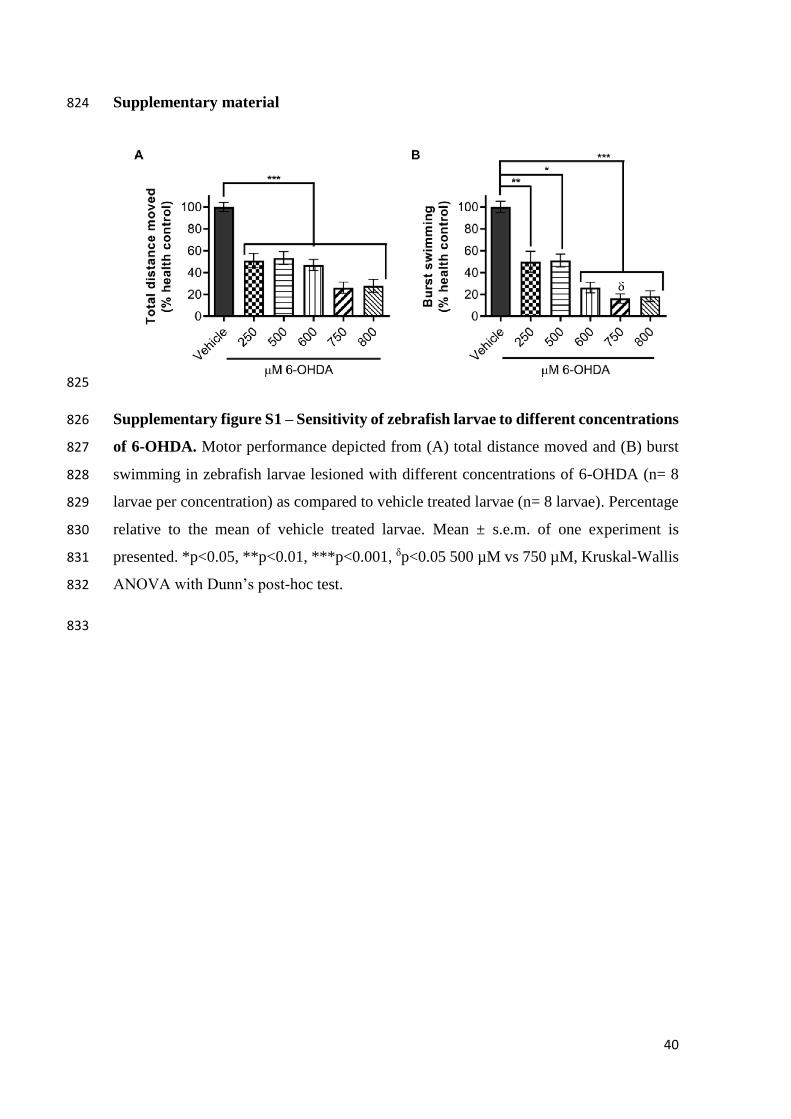
dissection of dopaminergic and noradrenergic contributions to catecholaminergic 718
tracts in early larval zebrafish. J. Comp. Neurol. 518, 439–458. 719
doi:10.1002/cne.22214 720
Lundblad, M., Usiello, A., Carta, M., Håkansson, K., Fisone, G., Cenci, M.A., 2005. 721
Pharmacological validation of a mouse model of L-DOPA-induced dyskinesia. 722
Exp. Neurol. 194, 66–75. doi:10.1016/j.expneurol.2005.02.002 723
Oertel, W., Schulz, J.B., 2016. Current and experimental treatments of Parkinson 724
disease: A guide for neuroscientists. J. Neurochem. 1–13. doi:10.1111/jnc.13750 725
Olanow, C.W., Rascol, O., Hauser, R., Feigin, P.D., Jankovic, J., Lang, A., Langston, 726
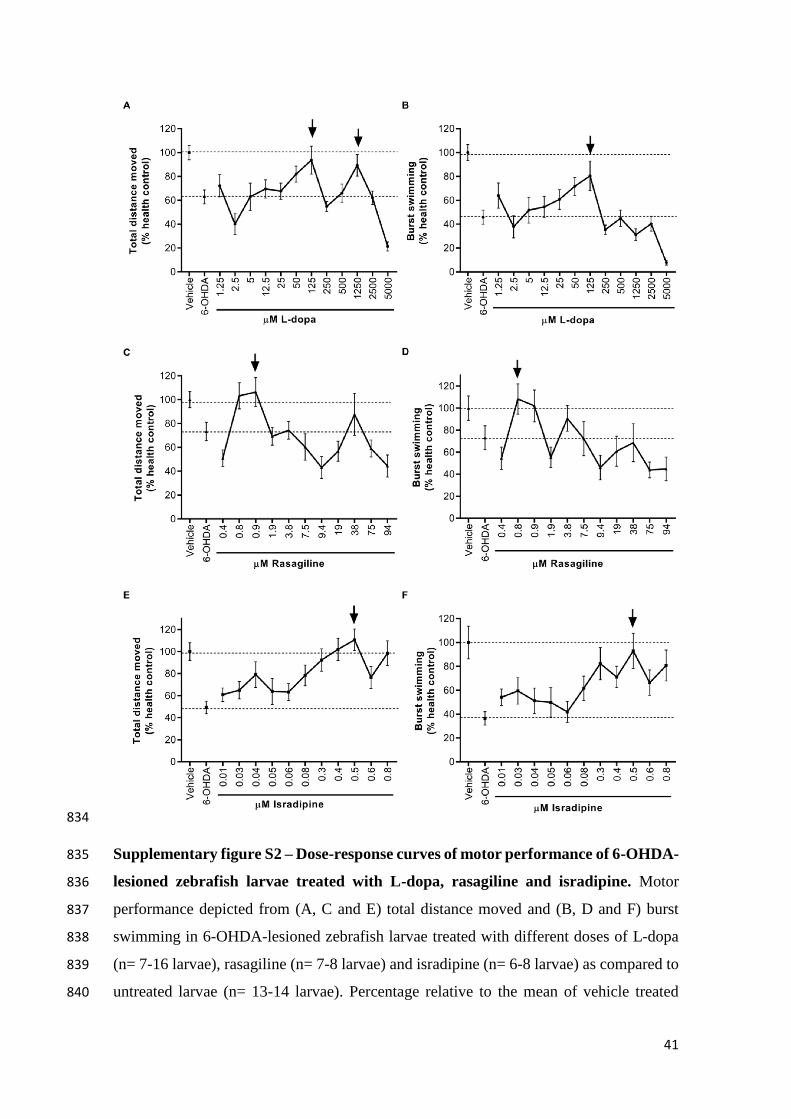
W., Melamed, E., Poewe, W., Stocchi, F., Tolosa, E., ADAGIO Study 727
Investigators, 2009. A Double-Blind, Delayed-Start Trial of Rasagiline in 728
Parkinson’s Disease. N. Engl. J. Med. 361, 1268–1278. 729
doi:10.1056/NEJMoa0809335 730
Panula, P., Chen, Y.C., Priyadarshini, M., Kudo, H., Semenova, S., Sundvik, M., 731
Sallinen, V., 2010. The comparative neuroanatomy and neurochemistry of 732
zebrafish CNS systems of relevance to human neuropsychiatric diseases. 733
Neurobiol. Dis. 40, 46–57. doi:10.1016/j.nbd.2010.05.010 734
Parng, C., Ton, C., Lin, Y.-X., Roy, N.M., McGrath, P., 2006. A zebrafish assay for 735
identifying neuroprotectants in vivo. Neurotoxicol. Teratol. 28, 509–516. 736
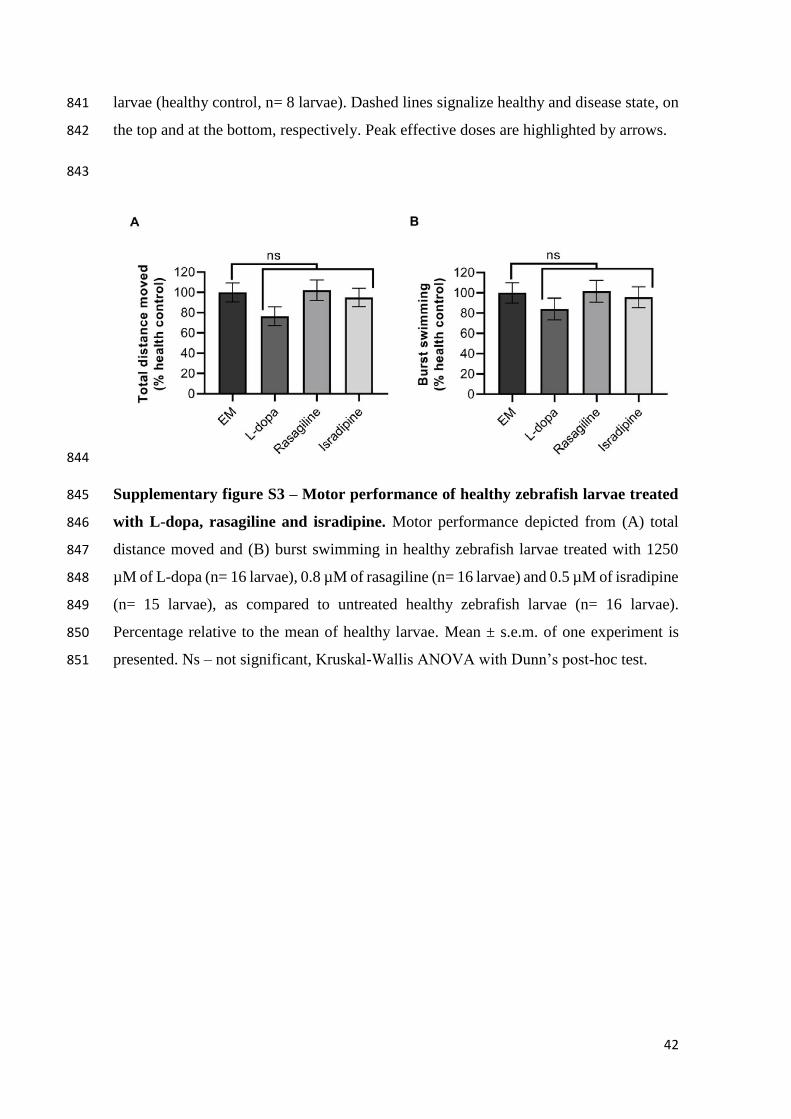
doi:10.1016/j.ntt.2006.04.003 737
Pistacchi, M., Martinello, F., Gioulis, M., Marsala, S.Z., 2014. Rasagiline and Rapid 738
Symptomatic Motor Effect in Parkinson’s Disease: Review of Literature. Neurol. 739
Ther. 3, 41–66. doi:10.1007/s40120-013-0014-1 740
Renn, C.L., Leitch, C.C., Lessans, S., Rhee, P., Mcguire, W.C., Smith, B.A., Traub, 741
R.J., Dorsey, S.G., 2011. Brain-derived neurotrophic factor modulates 742
antiretroviral-induced mechanical allodynia in the mouse. J. Neurosci. Res. 89, 743
1551–1565. doi:10.1002/jnr.22685 744

37
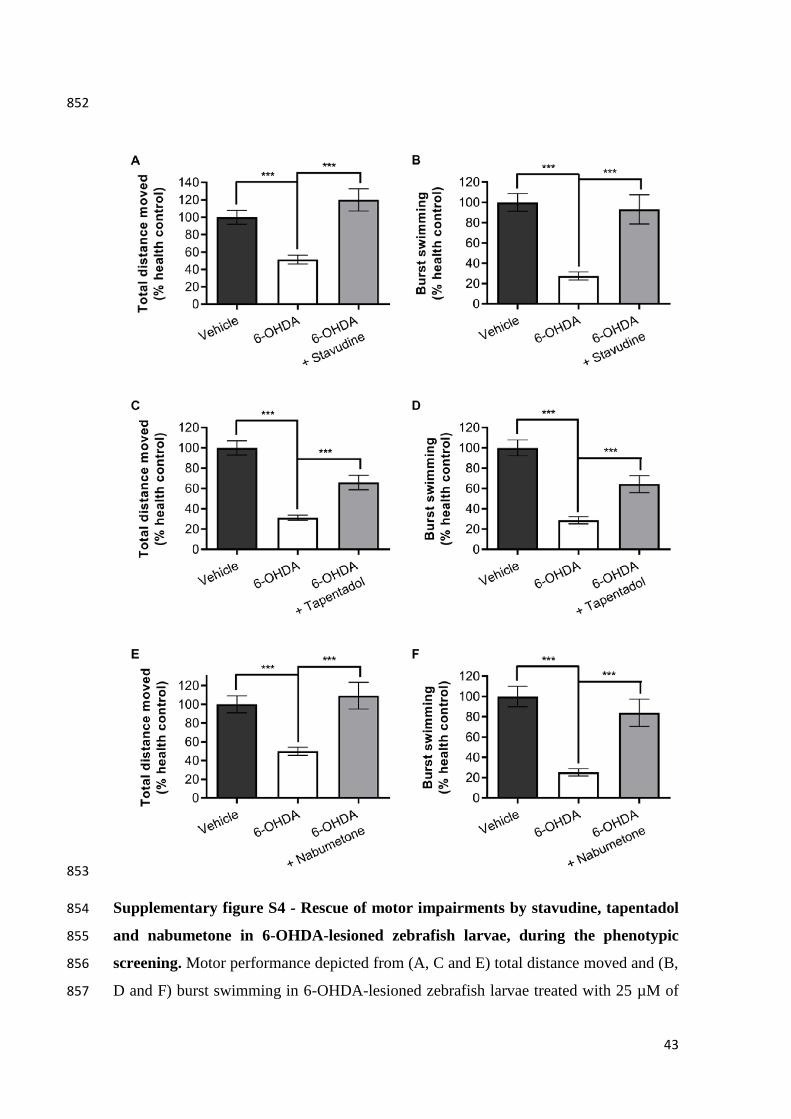
Rennekamp, A.J., Peterson, R.T., 2015. 15 Years of Zebrafish Chemical Screening. 745
Curr. Opin. Chem. Biol. 24, 58–70. doi:10.1016/j.cbpa.2014.10.025 746
Rico, E.P., Rosemberg, D.B., Seibt, K.J., Capiotti, K.M., Da Silva, R.S., Bonan, C.D., 747
2011. Zebrafish neurotransmitter systems as potential pharmacological and 748
toxicological targets. Neurotoxicol. Teratol. 33, 608–617. 749
doi:10.1016/j.ntt.2011.07.007 750
Rink, E., Wullimann, M.F., 2002. Connections of the ventral telencephalon and tyrosine 751
hydroxylase distribution in the zebrafish brain (Danio rerio) lead to identification 752
of an ascending dopaminergic system in a teleost. Brain Res. Bull. 57, 385–387. 753
doi:10.1016/S0361-9230(01)00696-7 754
Rink, E., Wullimann, M.F., 2001. The teleostean (zebrafish) dopaminergic system 755
ascending to the subpallium (striatum) is located in the basal diencephalon 756
(posterior tuberculum). Brain Res. 889, 316–330. doi:10.1016/S0006-757
8993(00)03174-7 758
Robertson, A.L., Ogryzko, N. V., Henry, K.M., Loynes, C.A., Foulkes, M.J., Meloni, 759
M.M., Wang, X., Ford, C., Jackson, M., Ingham, P.W., Wilson, H.L., Farrow, 760
S.N., Solari, R., Flower, R.J., Jones, S., Whyte, M.K.B., Renshaw, S.A., 2016. 761
Identification of benzopyrone as a common structural feature in compounds with 762
anti-inflammatory activity in a zebrafish phenotypic screen. Dis. Model. Mech. 9, 763
621–632. doi:10.1242/dmm.024935 764
Rylander, D., Recchia, A., Mela, F., Dekundy, A., Danysz, W., Cenci, M.A., 2009. 765
Pharmacological modulation of glutamate transmission in a rat model of L-DOPA-766
induced dyskinesia: effects on motor behavior and striatal nuclear signaling. J. 767
Pharmacol. Exp. Ther. 330, 227–235. doi:10.1124/jpet.108.150425 768
Samadi, P., Bédard, P.J., Rouillard, C., 2006. Opioids and motor complications in 769
Parkinson’s disease. Trends Pharmacol. Sci. 27, 512–517. 770
doi:10.1016/j.tips.2006.08.002 771
Schneider, C.A., Rasband, W.S., Eliceiri, K.W., 2012. NIH Image to ImageJ: 25 years 772
of image analysis. Nat. Methods 671–675. doi:10.1038/nmeth.2089 773
Schuster, S., Doudnikoff, E., Rylander, D., Berthet, A., Aubert, I., Ittrich, C., Bloch, B., 774

38
Cenci, M.A., Surmeier, D.J., Hengerer, B., Bezard, E., 2009. Antagonizing L-type 775
Ca2+ Channel Reduces Development of Abnormal Involuntary Movement in the 776
Rat Model of L-3,4-Dihydroxyphenylalanine-Induced Dyskinesia. Biol. Psychiatry 777
65, 518–526. doi:10.1016/j.biopsych.2008.09.008 778
Sun, Y., Dong, Z., Khodabakhsh, H., Chatterjee, S., Guo, S., 2012. Zebrafish chemical 779
screening reveals the impairment of dopaminergic neuronal survival by cardiac 780
glycosides. PLoS One 7, 1–11. doi:10.1371/journal.pone.0035645 781
Tay, T.L., Ronneberger, O., Ryu, S., Nitschke, R., Driever, W., 2011. Comprehensive 782
catecholaminergic projectome analysis reveals single-neuron integration of 783
zebrafish ascending and descending dopaminergic systems. Nat. Commun. 2, 171. 784
doi:10.1038/ncomms1171 785
Tzschentke, T.M., Christoph, T., Kögel, B., Schiene, K., Hennies, H., Englberger, W., 786
Haurand, M., Jahnel, U., Cremers, T.I.F.H., Friderichs, E., Vry, J. De, 2007. (-)-787
(1R,2R)-3-(3-Dimethylamino-1-ethyl-2-methyl-propyl)-phenol hydrochloride 788
(Tapentadol HCl): a Novel u-Opioid Receptor Agonist / Norepinephrine Reuptake 789
Inhibitor with Broad-Spectrum Analgesic Properties. J Pharmacol Exp Ther. 323, 790
265–276. doi:10.1124/jpet.107.126052.2002 791
Vaz, R.L., Outeiro, T.F., Ferreira, J.J., 2018. Zebrafish as an animal model for drug 792
discovery in Parkinson’s disease and other movement disorders: a systematic 793
review. Front. Neurol. 9, 1–23. doi:10.3389/FNEUR.2018.00347 794
Vijayakumar, D., Jankovic, J., 2016. Drug-Induced Dyskinesia, Part 1: Treatment of 795
Levodopa-Induced Dyskinesia. Drugs 76, 759–777. doi:10.1007/s40265-016-0566-796
3 797
Vijayanathan, Y., Lim, F.T., Lim, S.M., Long, C.M., Tan, M.P., Majeed, A.B.A., 798
Ramasamy, K., 2017. 6-OHDA-Lesioned Adult Zebrafish as a Useful Parkinson’s 799
Disease Model for Dopaminergic Neuroregeneration. Neurotox. Res. Epub ahead, 800
1–13. doi:10.1007/s12640-017-9778-x 801
Wang, M., Zhang, Z., Cheang, L.C., Lin, Z., Lee, S.M., 2011. Eriocaulon buergerianum 802
extract protects PC12 cells and neurons in zebrafish against 6-hydroxydopamine-803
induced damage. Chin. Med. 6, 1–10. doi:10.1186/1749-8546-6-16 804

39
Wang, Y., Liu, W., Yang, J., Wang, F., Sima, Y., Zhong, Z., Wang, H., Hu, L.-F., Liu, 805
C.-F., 2017. Parkinson’s disease-like motor and non-motor symptoms in rotenone-806
treated zebrafish. Neurotoxicology 58, 103–109. doi:10.1016/j.neuro.2016.11.006 807
Westerfield, M., 2000. The zebrafish book. A guide for the laboratory use of zebrafish 808
(Danio rerio)., 4th ed. Univ. of Oregon Press, Eugene. 809
Xi, Y., Noble, S., Ekker, M., 2011. Modeling neurodegeneration in zebrafish. Curr. 810
Neurol. Neurosci. Rep. 11, 274–282. doi:10.1007/s11910-011-0182-2 811
Zhang, Z., Cheang, L.C.V., Wang, M., Li, G., Chu, I.K., Lin, Z., Lee, S.M.Y., 2012. 812
Ethanolic extract of fructus alpinia oxyphylla protects against 6-hydroxydopamine-813
induced damage of PC12 cells in vitro and dopaminergic neurons in zebrafish. 814
Cell. Mol. Neurobiol. 32, 27–40. doi:10.1007/s10571-011-9731-0 815
Zhang, Z.J., Cheang, L.C.V., Wang, M.W., Lee, S.M., 2011. Quercetin exerts a 816
neuroprotective effect through inhibition of the iNOS/NO system and pro-817
inflammation gene expression in PC12 cells and in zebrafish. Int. J. Mol. Med. 27, 818
195–203. doi:10.3892/ijmm.2010.571 819
Zupanc, G.K.H., 2008. Adult neurogenesis and neuronal regeneration in the brain of 820
teleost fish. J. Physiol. Paris 102, 357–373. doi:10.1016/j.jphysparis.2008.10.007 821
822
823
40
Supplementary material 824
825
Supplementary figure S1 – Sensitivity of zebrafish larvae to different concentrations 826
of 6-OHDA. Motor performance depicted from (A) total distance moved and (B) burst 827
swimming in zebrafish larvae lesioned with different concentrations of 6-OHDA (n= 8 828
larvae per concentration) as compared to vehicle treated larvae (n= 8 larvae). Percentage 829
relative to the mean of vehicle treated larvae. Mean ± s.e.m. of one experiment is 830
presented. *p<0.05, **p<0.01, ***p<0.001, δp<0.05 500 µM vs 750 µM, Kruskal-Wallis 831
ANOVA with Dunn’s post-hoc test. 832
833
41
834
Supplementary figure S2 – Dose-response curves of motor performance of 6-OHDA-835
lesioned zebrafish larvae treated with L-dopa, rasagiline and isradipine. Motor 836
performance depicted from (A, C and E) total distance moved and (B, D and F) burst 837
swimming in 6-OHDA-lesioned zebrafish larvae treated with different doses of L-dopa 838
(n= 7-16 larvae), rasagiline (n= 7-8 larvae) and isradipine (n= 6-8 larvae) as compared to 839
untreated larvae (n= 13-14 larvae). Percentage relative to the mean of vehicle treated 840
42
larvae (healthy control, n= 8 larvae). Dashed lines signalize healthy and disease state, on 841
the top and at the bottom, respectively. Peak effective doses are highlighted by arrows. 842
843
844
Supplementary figure S3 – Motor performance of healthy zebrafish larvae treated 845
with L-dopa, rasagiline and isradipine. Motor performance depicted from (A) total 846
distance moved and (B) burst swimming in healthy zebrafish larvae treated with 1250 847
µM of L-dopa (n= 16 larvae), 0.8 µM of rasagiline (n= 16 larvae) and 0.5 µM of isradipine 848
(n= 15 larvae), as compared to untreated healthy zebrafish larvae (n= 16 larvae). 849
Percentage relative to the mean of healthy larvae. Mean ± s.e.m. of one experiment is 850
presented. Ns – not significant, Kruskal-Wallis ANOVA with Dunn’s post-hoc test. 851
43
852
853
Supplementary figure S4 - Rescue of motor impairments by stavudine, tapentadol 854
and nabumetone in 6-OHDA-lesioned zebrafish larvae, during the phenotypic 855
screening. Motor performance depicted from (A, C and E) total distance moved and (B, 856
D and F) burst swimming in 6-OHDA-lesioned zebrafish larvae treated with 25 µM of 857

44
stavudine (n= 19 larvae), tapentadol (n= 24 larvae) and nabumetone (n= 20 larvae), 858
picked from a library of FDA approved drugs, as compared to untreated larvae (n= 38-45 859
larvae). Percentage relative to the mean of vehicle treated larvae (healthy control, n= 23-860
24 larvae). Mean ± s.e.m. of three independent experiments is presented. ***p<0.001, 861
one-way ANOVA with Dunnett’s post-hoc test. 862
863





























